Какова роль белка DSS1 в клетках человека. Как DSS1 взаимодействует с белком BRCA2. Каковы последствия подавления экспрессии DSS1 для стабильности BRCA2 и устойчивости клеток к повреждению ДНК. Может ли нарушение функции DSS1 быть связано с развитием рака молочной железы и яичников.
Открытие и структура белка DSS1
DSS1 (Deleted in Split hand/Split foot 1) представляет собой небольшой кислый белок массой около 70 аминокислот, который высококонсервативен в эволюции от дрожжей до человека. Этот белок был впервые обнаружен при изучении генетических причин редкого наследственного заболевания — синдрома расщепленной кисти/стопы.
Структурно DSS1 характеризуется следующими особенностями:
- Состоит преимущественно из отрицательно заряженных аминокислот (аспарагиновая и глутаминовая кислоты)
- Содержит неупорядоченные участки, что обеспечивает конформационную гибкость белка
- Имеет консервативный C-концевой домен, участвующий во взаимодействии с другими белками
Такая структура позволяет DSS1 выступать в роли белка-шаперона и участвовать в регуляции стабильности и функций других белков в клетке.

Взаимодействие DSS1 с белком BRCA2
Одним из ключевых партнеров DSS1 является белок BRCA2 — продукт гена-супрессора опухолей BRCA2, мутации в котором связаны с наследственными формами рака молочной железы и яичников. Исследования показали, что DSS1 напрямую связывается с ДНК-связывающим доменом BRCA2.
Основные особенности взаимодействия DSS1-BRCA2:
- DSS1 связывается с консервативным участком BRCA2, который необходим для его функции в репарации ДНК
- Практически весь клеточный пул BRCA2 находится в комплексе с DSS1
- Связывание DSS1 стабилизирует структуру BRCA2 и защищает его от деградации
Таким образом, DSS1 выступает в роли ко-шаперона BRCA2, поддерживая его нативную конформацию и функциональную активность в клетке.
Роль DSS1 в обеспечении стабильности белка BRCA2
Исследования показали, что DSS1 играет критическую роль в поддержании стабильности белка BRCA2 в клетках млекопитающих. При подавлении экспрессии DSS1 с помощью РНК-интерференции наблюдаются следующие эффекты:
- Резкое снижение уровня белка BRCA2 в клетке
- Усиление протеасомной деградации BRCA2
- Нарушение локализации BRCA2 в ядре
Важно отметить, что мутантные формы BRCA2, лишенные домена связывания DSS1, не подвергаются деградации при истощении DSS1. Это указывает на то, что стабилизирующий эффект DSS1 опосредован его прямым взаимодействием с BRCA2.

Влияние DSS1 на функции BRCA2 в репарации ДНК
BRCA2 играет ключевую роль в процессе гомологичной рекомбинации — одном из основных механизмов репарации двухцепочечных разрывов ДНК. Исследования показали, что DSS1 необходим для полноценного функционирования BRCA2 в этом процессе:
- DSS1 способствует правильной локализации BRCA2 в участках повреждения ДНК
- При истощении DSS1 нарушается формирование фокусов RAD51 — ключевого белка гомологичной рекомбинации
- Клетки с пониженным уровнем DSS1 проявляют повышенную чувствительность к ДНК-повреждающим агентам
Таким образом, DSS1 не только стабилизирует BRCA2, но и необходим для его функциональной активности в процессах репарации ДНК.
DSS1 и геномная нестабильность
Учитывая важную роль DSS1 в поддержании стабильности и функций BRCA2, нарушение экспрессии или функции DSS1 может приводить к геномной нестабильности — одному из ключевых признаков раковых клеток. Исследования показали:
- Клетки с пониженным уровнем DSS1 демонстрируют повышенную частоту хромосомных аберраций
- Дефицит DSS1 приводит к нарушению клеточного цикла и активации контрольных точек повреждения ДНК
- При истощении DSS1 наблюдается повышенный уровень спонтанных двухцепочечных разрывов ДНК
Эти данные указывают на то, что DSS1 играет важную роль в поддержании стабильности генома, в значительной степени через регуляцию функций BRCA2.

Потенциальная роль нарушений DSS1 в канцерогенезе
Учитывая важность DSS1 для стабильности и функций BRCA2, нарушения в гене DSS1 или регуляции его экспрессии могут потенциально приводить к развитию рака, особенно рака молочной железы и яичников. Возможные механизмы включают:
- Мутации в гене DSS1, нарушающие его взаимодействие с BRCA2
- Эпигенетическое подавление экспрессии DSS1
- Нарушение пост-трансляционных модификаций DSS1, влияющих на его функции
Интересно, что повышенная экспрессия DSS1 также ассоциирована с неблагоприятным прогнозом при некоторых типах рака молочной железы. Это указывает на то, что точная регуляция уровня DSS1 критически важна для нормального функционирования клеток.
Терапевтический потенциал DSS1 при раке
Понимание роли DSS1 в регуляции функций BRCA2 открывает новые возможности для терапии рака:
- Стабилизация комплекса DSS1-BRCA2 может повысить эффективность ДНК-повреждающих противоопухолевых препаратов
- Нарушение взаимодействия DSS1-BRCA2 в раковых клетках может сенсибилизировать их к PARP-ингибиторам
- Модуляция уровня DSS1 может быть использована для регуляции активности BRCA2 в терапевтических целях
Однако для реализации этого потенциала необходимы дальнейшие исследования механизмов регуляции DSS1 и его взаимодействия с BRCA2 в нормальных и раковых клетках.

Заключение
Исследования последних лет показали, что небольшой белок DSS1 играет критически важную роль в обеспечении стабильности и функциональной активности BRCA2 — ключевого белка репарации ДНК и супрессора опухолей. DSS1 не только защищает BRCA2 от деградации, но и необходим для его правильной локализации и функционирования в процессах репарации повреждений ДНК.
Нарушение функций DSS1 приводит к геномной нестабильности и повышенной чувствительности клеток к повреждающим воздействиям, что может способствовать развитию рака. Дальнейшее изучение механизмов регуляции DSS1 и его взаимодействия с BRCA2 может открыть новые возможности для диагностики и терапии рака, особенно наследственных форм рака молочной железы и яичников.
Добро пожаловать на сайт АЕК
| Абонентская цифровая сигнализация DSS1 |
|---|
Данный материал содержит следующие 5 лекций:
Лекция 1: Архитектура протоколов абонентской сигнализации
Лекция 2: Протокол доступа к каналу D (LAPD-Q.921)
Лекция 3: Формат сообщений сигнализации DSS1
Лекция 4: Протокол абонентской сигнализации Q.931
Лекция 5: Процедуры управления вызовом
Лекция 1: АРХИТЕКТУРА ПРОТОКОЛОВ АБОНЕНТСКОЙ СИГНАЛИЗАЦИИ
Абонентская сигнализация обеспечивает понимание между устройством пользователя и сетью.
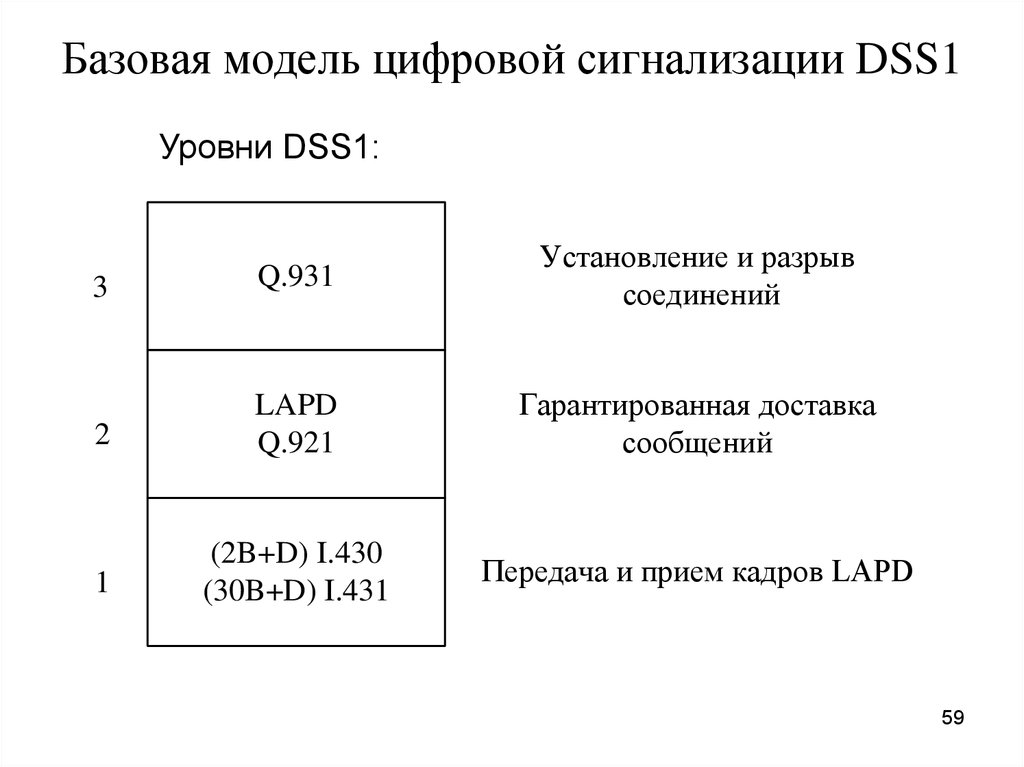
Протоколы абонентской сигнализации структурированы в соответствии с семиуровневой моделью взаимодействия открытых систем (ЭМВОС – OSI).
Сигнализация «пользователь-сеть» реализуется в пределах трех нижних уровней ЭМВОС.
- Уровень передачи бит (физический уровень, уровень 1) обеспечивает синхронизируемую сетью передачу информации по каналам одновременно в обоих направлениях. В случае основного абонентского окончания уровень передачи бит дополнительно делает возможным систематическую активизацию и деактивизацию и регулирует одновременный доступ нескольких оконечных устройств к совместно используемому D-каналу.
Детальные спецификации функций физического уровня приводятся в рек. МСЭ-Т I.430 (BRI), I.431 (PRI).
- Уровень защиты D-канала (уровень канала данных, уровень 2) обеспечивает защищенную от ошибок передачу информации сигнализации для уровня коммутации и, возможно, пакетированных данных, передаваемых в D-канале, в обоих направлениях между сетью и оконечным устройством.
Детальные спецификации функций этого уровня приводятся в рек.![]() МСЭ-Т Q.921 (протокол LAP-D).
МСЭ-Т Q.921 (протокол LAP-D).
- На уровне маршрутизации (сетевой уровень, уровень 3) заканчивается сигнализация «пользователь-сеть». Детальные спецификации протокола третьего уровня приводятся в рек. МСЭ-Т Q.931
- Протоколы более «высоких» уровней реализуются в терминалах пользователей. Сеть передает соответствующую информацию «прозрачно», т.е. без обработки.
Архитектура (стек) протоколов сигнализации DSS1
В таблицах 1, 2, 3 приведены функции протоколов физического (таблица 1), канального (таблица 2) и сетевого (таблица 3) уровней, поддерживаемых в доступе ЦСИО.
Таблица 1 – Протоколы физического уровня
Функции |
Описание (Рекомендации I. |
|
Конфигурация проводки |
Соединение между TE и NT |
Линейный код |
AMI (инверсия) |
Структура цикла |
Синхронизация битов, октетов, циклов |
Управление конфликтами в канале D |
Управление доступом к каналу D |
Идентификация каналов |
Идентификация каналов В и D |
Техническая эксплуатация |
Действия по ТЭ доступа к абоненту и его установкам |
Электрические характеристики |
Реализация стыка в соединениях с пассивной шиной (см. раздел “Эталонные конфигурации доступа к ресурсам У-ЦСИО”.) |
Физические характеристики |
Распределение контактов розетки для TE1 |
 430, I.412)
430, I.412)
Таблица 2 – Протоколы канального уровня
Функции |
Описание (Рекомендация Q. |
Исключение ложных флагов |
Обеспечение информационной прозрачности при переносе через сеть |
Идентификация цикла |
Опознавание и проверка на достоверность всех 48-битовых циклов |
Установка режима передачи |
Посылка вызова в сеть для инициализации услуги |
Обнаружение ошибок |
Обнаружение ошибок в принимаемом сообщении и в логике обмена с сетью |
Программное управление |
Обеспечение непрерывной последовательности кадров в соединении |
Восстановление |
Исправление ошибок средствами канального уровня (обнаружение ошибок в принимаемых кадрах и их исправление с использованием решающей обратной связи) и информирование сетевого протокола об ошибках, не поддающихся исправлению средствами в звене. |
Возможности вещания |
Предоставление звеньев данных вещания, которые могут быть идентифицированы с помощью глобального (группового) идентификатора TEI. |
 921)
921)
Таблица 3 – Протоколы сетевого уровня
Функции |
Описание (Рекомендация Q.931) |
Сообщения идентификации и обработки |
Опознавание и проверка правильности форматов сообщений |
Указатель (метка) вызова CRV |
Идентификация запроса вызова в интерфейсе “пользователь-сеть” |
Информация для обмена с сетью |
Спецификация типов сообщений (в фазах установления, передачи, разъединения) |
Сообщения управления вызовом |
Формирование обязательных сообщений для процедур управления основным вызовом |
|
Цифровой абонентский доступ
Основной предпосылкой создания сети ISDN (Integrated Service Digital Network) является организация цифрового абонентского доступа. В настоящее время, в ряде стран существуют различные протоколы сигнализации на абонентском уровне. Это связано с тем, что разработка их проводилась различными фирмами при отсутствии единого стандарта. В 1990 году были разработаны стандарты, регламентирующие систему сигнализации на абонентском участке ISDN. Эта система сигнализации была названа EDSS1 (Европейская цифровая система сигнализации №1), которая является единственной системой сигнализации, используемой на сети общего пользования России на абонентском участке сети ISDN. Данная система сигнализации применяется как для базового, так и для первичного доступа. Цифровой абонентский доступ Цифровой абонентский доступ характеризуется физическими параметрами интерфейса «пользователь — сеть» и абонентской сигнализацией.
Основная конфигурация абонентского доступа приведена на рис.1. Несколько абонентских устройств (TE — terminal equipment) подключаются к одному устройству сетевого окочания (NT — network terminator), при этом оконечные устройства могут быть либо однотипными, либо разнотипными (устройства для передачи текста, данных, речи). Сетевое окончание предназначено для подключения абонентской установки к абонентской линии и для обеспечения совместного использования одной абонентской линии несколькими абонентскими установками. В связи с этим блок NT разделяется на два блока — NT1 и NT2.
Между функциональными блоками определяются контрольные точки (интерфейсы обмена информации): точка S, точка T и точка U (в протоколе EDSS1 эти точки обозначаются как S0, T, Uk0 соответственно). В этих точках образован стандартизованный физический стык. Абонентская сигнализация
Задачей абонентской сигнализации является обеспечение взаимодействия между оборудованием и сетью (сигнализация «пользователь — сеть»).
|
 ::. Абонентская сигнализация DSS1
::. Абонентская сигнализация DSS1 

 На сети ISDN допускается также в ограниченном объеме обмен информацией между двумя устройствами пользователя по D-каналу (сигнализация «пользователь — пользователь»).
На сети ISDN допускается также в ограниченном объеме обмен информацией между двумя устройствами пользователя по D-каналу (сигнализация «пользователь — пользователь»). Третьим уровнем заканчивается сигнализация «пользователь — сеть».
Третьим уровнем заканчивается сигнализация «пользователь — сеть».
DSS1 требуется для стабильности BRCA2
-
Band V, Sager R . (1989). Proc Natl Acad Sci USA 86 : 1249–1253.
-
Скучный CC, Squires TS, Tong T . (1991). CA Cancer J Clin 41 : 19–36.
-
Броди Л.С., Бизекер Б.Б. (1998). Медицина (Балтимор) 77 : 208–226.
-
Chen CF, Chen PL, Zhong Q, Sharp ZD, Lee WH . (1999). J Биол Хим 274 : 32931–32935.
-
Chen PL, Chen CF, Chen Y, Xiao J, Sharp ZD, Lee WH . (1998). Proc Natl Acad Sci USA 95 : 5287–5292.
-
Коннор Ф., Бертуисл Д., Ми П.Дж., Росс Г.М., Свифт С., Григорьева Е. и др. . (1997). Nat Genet 17 : 423–430.
-
Краковер М.
 А., Шерер С.В., Ромменс Дж.М., Хуэй К.С., Пуркай П., Содер С. и др. . (1996). Хум Мол Жене 5 : 571–579.
А., Шерер С.В., Ромменс Дж.М., Хуэй К.С., Пуркай П., Содер С. и др. . (1996). Хум Мол Жене 5 : 571–579. -
Daniels MJ, Wang Y, Lee M, Venkitaraman AR . (2004). Наука 306 : 876–879.
-
Дэвис А.А., Массон Дж.Ю., Макилрайт М.Дж., Стасиак А.З., Стасиак А., Венкитараман А.Р. и др. . (2001). Мол Ячейка 7 : 273–282.
-
Димри Г.П., Итахана К., Акоста М., Кампизи Дж. (2000). Mol Cell Biol 20 : 273–285.
-
Голдман Л.А., Кутроне Э.К., Котенко С.В., Краузе К.Д., Лангер Дж.А. (1996). Биотехнологии 21 : 1013–1015.
-
Гудмундсдоттир К., Лорд С.Дж., Витт Э., Тутт А.Н., Эшворт А. . (2004). EMBO Rep 5 : 989–993 [E-pub, 10 сентября 2004 г.].
-
Howlett NG, Taniguchi T, Olson S, Cox B, Waisfisz Q, De Die-Smulders C et al . (2002). Science 297 : 606–609 [E-pub 2002, 13 июня].

-
Хьюз-Дэвис Л., Хантсман Д., Руас М., Фукс Ф., Бай Дж., Чин С.Ф. и др. . (2003). Сотовый 115 : 523–535.
-
Исоно Э., Саэки Ю., Йокосава Х., Тох-э А. . (2004). J Biol Chem 279 : 27168–27176 [E-pub 2004, 21 апреля].
-
Джантти Дж., Лахденранта Дж., Олкконен В.М., Содерлунд Х., Керанен С. . (1999). Proc Natl Acad Sci USA 96 : 909–914.
-
Койич М., Ян Х., Коструб С.Ф., Павлетич Н.П., Холломан В.К. (2003). Мол Ячейка 12 : 1043–1049.
-
Краакман-ван дер Цвет М., Оверкамп В.Дж., ван Ланге Р.Е., Эссерс Дж., ван Дайн-Годхарт А., Виггерс И. и др. . (2002). Mol Cell Biol 22 : 669–679.
-
Марстон Н.Дж., Ричардс В.Дж., Хьюз Д., Бертуистл Д., Маршалл С.Дж., Эшворт А. . (1999). Mol Cell Biol 19 : 4633–4642.
-
Милнер Дж., Пондер Б., Хьюз-Дэвис Л., Зельтманн М.
 , Кузаридес Т. . (1997). Природа 386 : 772–773.
, Кузаридес Т. . (1997). Природа 386 : 772–773. -
Mizuta R, LaSalle JM, Cheng HL, Shinohara A, Ogawa H, Copeland N и др. . (1997). Proc Natl Acad Sci USA 94 : 6927–6932.
-
Мойнахан М.Э., Пирс А.Дж., Джасин М. . (2001). Мол Ячейка 7 : 263–272.
-
Patel KJ, Yu VP, Lee H, Corcoran A, Thistlethwaite FC, Evans MJ и др. . (1998). Мол Ячейка 1 : 347–357.
-
Рахман Н., Стрэттон М.Р. (1998). Annu Rev Genet 32 : 95–121.
-
Рач С.Б., Гао К., Шринивасан С., Вазер Д.Е., Бэнд В. . (2001). Radiat Res 155 : 143–150.
-
Шаран С.К., Моримацу М., Альбрехт У., Лим Д.С., Регель Э., Динь С. и др. . (1997). Природа 386 : 804–810.
-
Шаран С.К., Пайл А., Коппола В., Бабус Дж., Сваминатан С., Бенедикт Дж. и др. . (2004). Разработка 131 : 131–142.

-
Шин С., Верма И.М. (2003). Proc Natl Acad Sci USA 100 : 7201–7206 [E-pub 2003, 19 мая].
-
Соне Т., Саэки Ю., Тох-э А., Йокосава Х. (2004). J Biol Chem 279 : 28807–28816 [E-pub 2004, 26 апреля].
-
Тавтигян С.В., Симард Дж., Ромменс Дж., Коуч Ф., Шаттак-Эйденс Д., Нойхаузен С. и др. . (1996). Nat Genet 12 : 333–337.
-
Tian XX, Rai D, Li J, Zou C, Bai Y, Wazer D и др. . (2005). Рак Res 65 : 4747–4753.
-
Тутт А., Габриэль А., Бертуистл Д., Коннор Ф., Патерсон Х., Пикок Дж. и др. . (1999). Curr Biol 9 : 1107–1110.
-
Ван С.К., Макино К., Су Л.К., Пао А.И., Ким Дж.С., Хунг М.С. (2001). Рак Res 61 : 2838–2842.
-
Ван С.К., Шао Р., Пао А.И., Чжан С., Хунг М.С., Су Л.К. (2002). Cancer Res 62 : 1311–1314.
-
Вэй С.
 Дж., Тремпус К.С., Кэннон Р.Э., Бортнер К.Д., Теннант Р.В. (2003). J Biol Chem 278 : 1758–1768 [E-pub 2002, 4 ноября].
Дж., Тремпус К.С., Кэннон Р.Э., Бортнер К.Д., Теннант Р.В. (2003). J Biol Chem 278 : 1758–1768 [E-pub 2002, 4 ноября]. -
Вонг А.К., Перо Р., Ормонде П.А., Тавтигян С.В., Бартел П.Л. (1997). J Biol Chem 272 : 31941–31944.
-
Вустер Р., Бигнелл Г., Ланкастер Дж., Свифт С., Сил С., Мэнджион Дж. и др. . (1995). Природа 378 : 789–792.
-
Вустер Р., Нойхаузен С.Л., Мангион Дж., Квирк Ю., Форд Д., Коллинз Н. и др. . (1994). Наука 265 : 2088–2090.
-
Ся Ф., Тагиан Д.Г., ДеФранк Дж.С., Цзэн З.К., Виллерс Х., Илиакис Г. и др. . (2001). Proc Natl Acad Sci USA 98 : 8644–8649.
-
Ян Х., Джеффри П.Д., Миллер Дж., Киннукан Э., Сан И., Тома Н.Х. и др. . (2002). Наука 297 : 1837–1848.
Ссылки на скачивание
DSS1 требуется для стабильности BRCA2
Сохранить цитату в файл
Формат: Резюме (текст)PubMedPMIDAbstract (текст)CSV
Добавить в коллекции
- Создать новую коллекцию
- Добавить в существующую коллекцию
Назовите свою коллекцию:
Имя должно содержать менее 100 символов
Выберите коллекцию:
Не удалось загрузить вашу коллекцию из-за ошибки
Повторите попытку
Добавить в мою библиографию
- Моя библиография
Не удалось загрузить делегатов из-за ошибки
Повторите попытку
Ваш сохраненный поиск
Название сохраненного поиска:
Условия поиска:
Тестовые условия поиска
Электронная почта: (изменить)
Который день? Первое воскресеньеПервый понедельникПервый вторникПервая средаПервый четвергПервая пятницаПервая субботаПервый деньПервый рабочий день
Который день? ВоскресеньеПонедельникВторникСредаЧетвергПятницаСуббота
Формат отчета: SummarySummary (text)AbstractAbstract (text)PubMed
Отправить максимум:
1 шт. 5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
Отправить, даже если нет новых результатов
Необязательный текст в электронном письме:
Создайте файл для внешнего программного обеспечения для управления цитированием
. 23 февраля 2006 г.; 25 (8): 1186-94.
doi: 10.1038/sj.onc.1209153.
Дж Ли 1 , C Zou, Y Bai, D E Wazer, V Band, Q Gao
принадлежность
- 1 Отделение биологии рака, Медицинский факультет, Эванстонский Северо-Западный научно-исследовательский институт здравоохранения, Северо-Западный университет, Медицинская школа им. Файнберга, Эванстон, Иллинойс 60201, США.
- PMID: 16205630
-
DOI:
10.
 1038/sj.onc.1209153
1038/sj.onc.1209153
Дж. Ли и соавт. Онкоген. .
. 2006 23 февраля; 25 (8): 1186-94.
дои: 10.1038/sj.onc.1209153.
Авторы
Дж Ли 1 , C Цзоу, Y Bai, D E Wazer, V Band, Q Gao
принадлежность
- 1 Отделение биологии рака, Медицинский факультет, Эванстонский Северо-Западный научно-исследовательский институт здравоохранения, Северо-Западный университет, Медицинская школа им. Файнберга, Эванстон, Иллинойс 60201, США.
- PMID: 16205630
-
DOI:
10.
 1038/sj.onc.1209153
1038/sj.onc.1209153
Абстрактный
DSS1 представляет собой эволюционно консервативный кислый белок, который связывается с BRCA2. Однако изучение функции DSS1 в клетках млекопитающих затруднено, потому что эндогенный DSS1 не обнаруживается вестерн-блоттингом. Здесь мы разработали модифицированный протокол вестерн-блоттинга, который обнаруживает эндогенный белок DSS1, и использовали его для изучения функции DSS1 и его взаимодействия с BRCA2 в клетках млекопитающих. Мы обнаружили, что практически все BRCA2 в клеточных линиях человека связаны с DSS1. Важно отметить, что мы обнаружили, что нокдаун DSS1 с помощью РНК-интерференции в клеточных линиях человека приводит к резкой потере белка BRCA2, в основном из-за его усиленной деградации. Более того, истощение DSS1 не влияет на стабильность мутанта BRCA2, лишенного домена связывания DSS1. В частности, как и истощение BRCA2, истощение DSS1 также приводило к гиперчувствительности к повреждению ДНК. Эти результаты показали, что стабильность белка BRCA2 в клетках млекопитающих зависит от присутствия DSS1. Таким образом, делеция или мутация DSS1 или подавление его экспрессии другими механизмами являются потенциальными причинными механизмами рака молочной железы и яичников человека. Такие механизмы могут иметь отношение как к спорадическому, так и к обычному раку молочной железы, при котором мутации BRCA1 и BRCA2 отсутствуют.
Эти результаты показали, что стабильность белка BRCA2 в клетках млекопитающих зависит от присутствия DSS1. Таким образом, делеция или мутация DSS1 или подавление его экспрессии другими механизмами являются потенциальными причинными механизмами рака молочной железы и яичников человека. Такие механизмы могут иметь отношение как к спорадическому, так и к обычному раку молочной железы, при котором мутации BRCA1 и BRCA2 отсутствуют.
Онкоген (2006) 25, 1186-1194. doi:10.1038/sj.onc.1209153; опубликовано в Интернете 3 октября 2005 г.
Похожие статьи
-
DSS1 необходим для формирования фокуса RAD51 и стабильности генома в клетках млекопитающих.
Гудмундсдоттир К., Лорд С.Дж., Витт Э., Тутт А.Н., Эшворт А. Гудмундсдоттир К. и соавт. EMBO Rep. 2004 Oct;5(10):989-93. doi: 10.1038/sj.embor.7400255. Epub 2004, 10 сентября. Представитель EMBO, 2004 г.
 PMID: 15359272
Бесплатная статья ЧВК.
PMID: 15359272
Бесплатная статья ЧВК. -
Рак молочной железы с высокой экспрессией DSS1, который потенциально поддерживает стабильность BRCA2, имеет плохой прогноз в отношении безрецидивной выживаемости.
Резано А., Кувахара К., Ямамото-Ибусуки М., Китабатакэ М., Мултия П., Фимсен С., Суда Т., Тоне С., Ямамото Ю., Ивасе Х., Сакагути Н. Резано А. и др. БМК Рак. 2013 1 декабря; 13:562. дои: 10.1186/1471-2407-13-562. БМК Рак. 2013. PMID: 24289229 Бесплатная статья ЧВК.
-
Взаимодействие между продуктом гена предрасположенности к раку молочной железы BRCA2 и DSS1, белком, функционально консервативным от дрожжей до млекопитающих.
Marston NJ, Richards WJ, Hughes D, Bertwistle D, Marshall CJ, Ashworth A. Марстон, штат Нью-Джерси, и др.
 Мол Селл Биол. 1999 июль; 19 (7): 4633-42. doi: 10.1128/MCB.19.7.4633.
Мол Селл Биол. 1999.
PMID: 10373512
Бесплатная статья ЧВК.
Мол Селл Биол. 1999 июль; 19 (7): 4633-42. doi: 10.1128/MCB.19.7.4633.
Мол Селл Биол. 1999.
PMID: 10373512
Бесплатная статья ЧВК. -
Роль BRCA1 и BRCA2 и ассоциированных белков в поддержании стабильности генома.
Гудмундсдоттир К., Эшворт А. Гудмундсдоттир К. и соавт. Онкоген. 2006 г., 25 сентября; 25 (43): 5864-74. doi: 10.1038/sj.onc.1209874. Онкоген. 2006. PMID: 16998501 Обзор.
-
Всесторонний анализ гена BRCA2: основное внимание уделяется механистическим аспектам его функций, спектру вредных мутаций и терапевтическим стратегиям, нацеленным на опухоли с дефицитом BRCA2.
Шайлани А., Каур Р.П., Мунши А. Шайлани А. и др. Мед Онкол. 2018 31 января; 35 (3): 18. doi: 10.1007/s12032-018-1085-8.
 Мед Онкол. 2018.
PMID: 29387975
Обзор.
Мед Онкол. 2018.
PMID: 29387975
Обзор.
Посмотреть все похожие статьи
Цитируется
-
Патогенный вариант R3052W BRCA2 нарушает гомологически направленную репарацию из-за невозможности локализации в ядре.
Хименес-Сайнс Дж., Криштофяк А., Гарбарино Дж., Роджерс Ф., Дженсен Р.Б. Хименес-Сайнс Дж. и соавт. Фронт Жене. 2022 30 мая; 13:884210. doi: 10.3389/fgene.2022.884210. Электронная коллекция 2022. Фронт Жене. 2022. PMID: 35711920 Бесплатная статья ЧВК.
-
Взаимодействие BRCA2-DSS1 необходимо для рекрутирования RAD51 при индуцированных репликацией и мейотических двухцепочечных разрывах ДНК.
Мишра А.П., Хартфорд С.А., Саху С., Кларманн К., Читтела Р.
 К., Бисвас К., Чон А.Б., Мартин Б.К., Беркетт С., Саутон Э., Рейд С., Олбо М.Е., Карим Б., Тессаролло Л., Келлер Дж.Р., Шаран С.К.
Мишра А.П. и др.
Нац коммун. 2022 1 апреля; 13 (1): 1751. дои: 10.1038/s41467-022-29409-й.
Нац коммун. 2022.
PMID: 35365640
Бесплатная статья ЧВК.
К., Бисвас К., Чон А.Б., Мартин Б.К., Беркетт С., Саутон Э., Рейд С., Олбо М.Е., Карим Б., Тессаролло Л., Келлер Дж.Р., Шаран С.К.
Мишра А.П. и др.
Нац коммун. 2022 1 апреля; 13 (1): 1751. дои: 10.1038/s41467-022-29409-й.
Нац коммун. 2022.
PMID: 35365640
Бесплатная статья ЧВК. -
OB-кратные семьи хранителей генома: универсальная тема, построенная из небольшого строительного блока β-ствола.
Бьянко Пр. Бьянко пиар. Фронт Мол Биоски. 2022, 11 февраля; 9:784451. doi: 10.3389/fmolb.2022.784451. Электронная коллекция 2022. Фронт Мол Биоски. 2022. PMID: 35223988 Бесплатная статья ЧВК. Обзор.
-
Интегрированный геномный анализ протеасомных изменений у 11 057 пациентов с 33 типами рака: клинически значимые результаты в рамках 3P-медицины.

