Как хлорохин взаимодействует с ионами цинка в клетках. Какие механизмы лежат в основе его противоопухолевого действия. Каковы перспективы применения хлорохина в комбинированной терапии рака. Может ли хлорохин усиливать действие других противоопухолевых препаратов. Какие побочные эффекты возможны при использовании хлорохина как ионофора цинка.
Механизм действия хлорохина как ионофора цинка
Хлорохин, широко известный противомалярийный препарат, в последние годы привлек внимание исследователей как потенциальное противоопухолевое средство. Одним из ключевых механизмов его действия является способность выступать в роли ионофора цинка. Это означает, что хлорохин облегчает проникновение ионов цинка через клеточные мембраны, повышая его внутриклеточную концентрацию.
Как именно происходит этот процесс? Хлорохин, будучи слабым основанием, накапливается в кислых компартментах клетки, таких как лизосомы и эндосомы. Там он связывается с ионами цинка, образуя липофильные комплексы, способные проникать через мембраны. В результате концентрация свободного цинка в цитоплазме и других органеллах повышается, что запускает ряд биологических эффектов.

Влияние хлорохина на метаболизм опухолевых клеток
Повышение концентрации цинка в опухолевых клетках под действием хлорохина приводит к серьезным нарушениям их метаболизма. Какие основные эффекты наблюдаются?
- Ингибирование гликолиза — основного источника энергии для быстро делящихся раковых клеток
- Нарушение функции митохондрий и снижение синтеза АТФ
- Индукция окислительного стресса из-за избытка свободных ионов цинка
- Ингибирование ключевых ферментов и сигнальных путей, необходимых для роста опухоли
- Активация процессов клеточной гибели — апоптоза и аутофагии
Таким образом, хлорохин, выступая как ионофор цинка, способен атаковать сразу несколько «ахиллесовых пят» раковых клеток, что делает его перспективным противоопухолевым агентом.
Комбинированная терапия с использованием хлорохина
Особый интерес представляет возможность использования хлорохина в комбинации с другими противоопухолевыми препаратами. Какие преимущества дает такой подход?
- Усиление цитотоксического действия химиопрепаратов за счет повышения их внутриклеточной концентрации
- Преодоление лекарственной устойчивости опухолей
- Сенсибилизация опухолевых клеток к лучевой терапии
- Снижение дозировок стандартных препаратов при сохранении эффективности
- Уменьшение побочных эффектов терапии
Исследования показывают, что хлорохин способен значительно усиливать действие таких препаратов как доксорубицин, цисплатин, гемцитабин и других. Это открывает новые возможности для повышения эффективности противоопухолевой терапии.

Влияние хлорохина на микроокружение опухоли
Помимо прямого воздействия на опухолевые клетки, хлорохин как ионофор цинка способен модулировать микроокружение опухоли. Каковы основные эффекты?
- Ингибирование ангиогенеза — формирования новых кровеносных сосудов в опухоли
- Подавление инвазии и метастазирования раковых клеток
- Модуляция иммунного ответа против опухоли
- Снижение гипоксии в опухолевой ткани
- Нормализация pH в микроокружении опухоли
Эти эффекты дополняют прямое цитотоксическое действие хлорохина и могут способствовать повышению эффективности противоопухолевой терапии в целом.
Перспективные направления исследований хлорохина в онкологии
Учитывая многообещающие результаты доклинических и ранних клинических исследований, какие направления дальнейшего изучения хлорохина как противоопухолевого агента представляются наиболее перспективными?
- Поиск оптимальных комбинаций хлорохина с другими препаратами для различных типов опухолей
- Разработка новых аналогов хлорохина с улучшенными фармакологическими свойствами
- Изучение механизмов резистентности опухолей к хлорохину и способов их преодоления
- Исследование возможностей применения хлорохина для профилактики рака
- Разработка методов адресной доставки хлорохина к опухолевым клеткам
Эти направления исследований могут привести к созданию новых эффективных стратегий противоопухолевой терапии на основе хлорохина как ионофора цинка.
Побочные эффекты и меры предосторожности при использовании хлорохина
Несмотря на многообещающие результаты, применение хлорохина в онкологии требует осторожности. Какие основные побочные эффекты и риски следует учитывать?
- Кардиотоксичность — нарушения сердечного ритма, удлинение интервала QT
- Ретинопатия при длительном применении высоких доз
- Гематологические нарушения — анемия, тромбоцитопения
- Гастроинтестинальные расстройства
- Нейротоксичность — судороги, психические нарушения
Для минимизации рисков необходим тщательный мониторинг состояния пациентов, получающих хлорохин. Важно также учитывать возможные взаимодействия хлорохина с другими лекарственными препаратами.
Роль цинка в противоопухолевом действии хлорохина
Хотя способность хлорохина выступать в роли ионофора цинка считается ключевым механизмом его противоопухолевого действия, остается ряд вопросов. Как именно повышение концентрации цинка влияет на опухолевые клетки?
- Цинк является кофактором более 300 ферментов и участвует в регуляции множества клеточных процессов
- Избыток свободного цинка может нарушать функцию митохондрий и индуцировать окислительный стресс
- Цинк способен модулировать активность ключевых сигнальных путей, контролирующих рост и выживание клеток
- Ионы цинка могут взаимодействовать с ДНК, влияя на экспрессию генов
- Цинк играет важную роль в регуляции иммунного ответа против опухоли
Дальнейшее изучение этих механизмов может помочь в разработке более эффективных стратегий применения хлорохина в онкологии.

Схемы на все случаи жизни » Ионофон на NE555
Добрый день, уважаемые радиолюбители. Сегодня я хотел бы предложить Вам схему простого ионофона. Сейчас этой тематике посвящено множество сайтов, форумов и тем… Вот и я решил приложить к этому свою руку… Не судите строго, это первый мой опыт конструирования подобной техники.
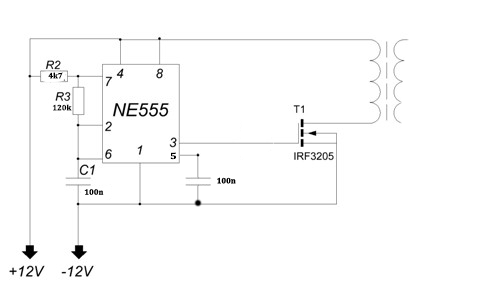
Схема электрическая принципиальная показана на рисунке ниже. Итак, рассмотрим схему предложенного ионофона подробнее.
Сигнал с линейного выхода какого-либо аудиоустройства поступает через фильтр-пробку С1-R1 на вторичную обмотку (На ту, при нормальном включении трансформатора на которой выходное напряжение равно 12 вольт) трансформатора ТВК-110. Данный трансформатор выполняет 2 роли: 1 – осуществляет гальваническую развязку Вашего аудиоустройства и ионофона; 2 – усиливает входной аудио сигнал до нужного уровня.
С первичной обмотки трансформатора (Той, на которую при нормальном включении подаётся 220 вольт), усиленный сигнал, через фильтр-пробку C2-R2 и диод VD3 поступает на вход генератора сигналов, реализованного на микросхеме DD1. Частота генератора определяется цепочкой R3-R4-C3 и при указанных на схеме номиналах составляет чуть-более 30 килогерц. Вход 2 микросхемы NE555 является запускающим т.е. при отсутствии на нём напряжения на выходе оно так же будет отсутствовать. Диод VD3 выделяет положительные полуволны сигнала. Таким образом при подаче аудио сигнала генератор запускается от каждого положительного полупериода сигнала и на выходе 3 микросхемы мы получаем пачки сигнала частотой 30 килогерц, следующие с частотой входного сигнала (По факту промодулированный сигнал). Данный сигнал, через резистор R5 поступает на вход составного транзистораVT1-VT2. Его нагрузкой является первичная обмотка трансформатора Tr2. Со вторичной обмотки данного трансформатора и снимается выходное высокое напряжение. Диод VD1 защищает схему от обратного напряжения, VD2 защищает схему от бросков напряжения по питанию.
Частота генератора определяется цепочкой R3-R4-C3 и при указанных на схеме номиналах составляет чуть-более 30 килогерц. Вход 2 микросхемы NE555 является запускающим т.е. при отсутствии на нём напряжения на выходе оно так же будет отсутствовать. Диод VD3 выделяет положительные полуволны сигнала. Таким образом при подаче аудио сигнала генератор запускается от каждого положительного полупериода сигнала и на выходе 3 микросхемы мы получаем пачки сигнала частотой 30 килогерц, следующие с частотой входного сигнала (По факту промодулированный сигнал). Данный сигнал, через резистор R5 поступает на вход составного транзистораVT1-VT2. Его нагрузкой является первичная обмотка трансформатора Tr2. Со вторичной обмотки данного трансформатора и снимается выходное высокое напряжение. Диод VD1 защищает схему от обратного напряжения, VD2 защищает схему от бросков напряжения по питанию.
Основным достоинством схемы является её простота. Схема не требует наладки и начинает работать сразу после включения.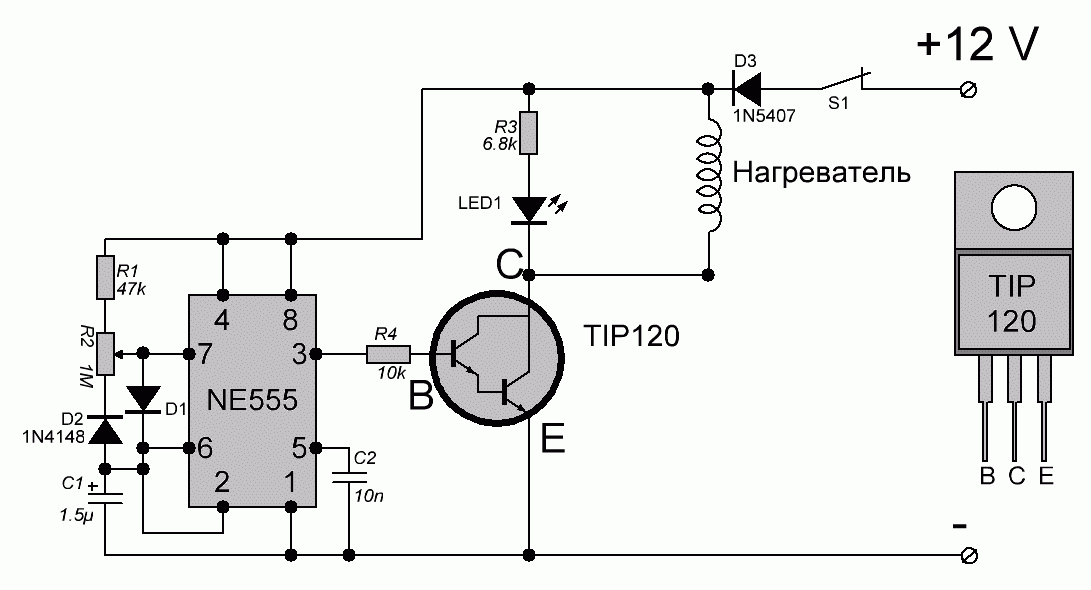 Но у схемы есть и небольшой недостаток, она потребляет в работе значительный импульсный ток, до 10 ампер.
Но у схемы есть и небольшой недостаток, она потребляет в работе значительный импульсный ток, до 10 ампер.
Все используемые детали указаны на схеме. Стоит только обратить внимание на изготовление трансформатора Tr2. Трансформатор Tr2 — это переделанный строчный трансформатор от старого лампового телевизора. Для его переделки снимаем первичную обмотку и мотаем свою. Первичная обмотка содержит 20 витков провода ПЭЛ-1.5. Вторичная обмотка (высоковольтная, залитая пластмассой) остается штатной, после чего трансформатор собирается. При сборке между половинок сердечника следует сделать зазор около 1 мм из тонкого гетинакса или стеклотекстолита. Транзисторы VT1 и VT2 следует установить на теплоотвод.
В подборках фото и видео ниже наглядно показана работа ионофона. Т.к. камера у меня не профессиональная, то голос и музыку слышно плохо, глушит треск горящей искры. При прослушивании на самом деле звук намного чище и отчётливее!!! На одном из видео видно что к выходному трансформатору подключён конденсатор, в конечной схеме он отсутствует т. к. особой роли на качество звучания не сыграл, хотя его присутствие несколько повышает громкость звучания отдельных композиций…
к. особой роли на качество звучания не сыграл, хотя его присутствие несколько повышает громкость звучания отдельных композиций…
Ну вот, на этом вроде бы на сегодня всё. До новых встреч. С уважением, Андрей Савченко.
P.S. Обновление от 29.03.2020:
Итак, данная конструкция собиралась мной летом 2013-го года и являлась первым опытом в применении таймера NE555 для ШИМ-управления (и подобных целей) на её основе. Я считаю в целом данную конструкцию НЕУДАЧНОЙ т.к. в самой конструкции и описании есть фактические ошибки. Данные ошибки возникли ввиду того, что я воспользовался источником с ошибочным описанием алгоритма работы данного таймера.
Во-первых, для данной конструкции справедлива рекомендация, данная для выходного каскада высоковольтного генератора на NE555.
Во-вторых, при подобной реализации управления пачек сигнала на выходе в реальности нет, но проконтролировать на тот момент я этого не смог т.к. у меня отсутствовал осциллограф.
При работе подобная схема работает в реальности следующим образом: при подаче на вход запуска (вход 2) импульса низкого уровня (т.е. при приходе отрицательной полуволны входного сигнала) таймер запускается и на выходе устанавливается напряжение высокого уровня на время, которое определяется внешней RC-цепочкой. Такая ситуация будет происходить при условии, если длительность входного импульса меньше времени заряда конденсатора RC-цепи. Если же входной импульс по длительности все-таки длиннее, то выход микросхемы будет оставаться в состоянии высокого уровня до тех пор, пока на входе не установится опять высокий уровень т.е. не придёт положительная полуволна сигнала.
Таким образом, в реальности, на выходе формируются отдельные выходные импульсы при превышении/понижении порога встроенного компаратора сигнала. Ни пачек импульсов, ни ШИМ-модуляции в такт амплитуде входного сигнала нет. Фактически это достаточно грубый пик-детектор (и если уж быть совсем точным даже не пик-детектор, а детектор перехода через пороговый уровень встроенного компаратора сигнала). Отсюда такой посредственный результат. При построении ионофона на таймере NE555 модуляцию необходимо осуществлять, например, по 5-му выводу. Модернизированный вариант со всеми необходимыми доработками описан в статье Вторая жизнь ионофона на NE555.
Схема ионофона — поющей дуги
-
08.10.2010 Электронная техника
-
Очередной увлекательный ознакомительный проект, который связан с высоким напряжением. Исходя из этого устраивайтесь эргономичнее. На протяжении ролика канала Aka Kasyan соберем одну весьма познавательную конструкцию — “Ионофон”, либо поющую приятелю.
 Быть может, многим это наименование ни о чем не говорит, но такие игрушки достаточно популярны среди начинающих радиолюбителей. Радиодетали дешево возможно приобрести в этом китайском магазине.
Быть может, многим это наименование ни о чем не говорит, но такие игрушки достаточно популярны среди начинающих радиолюбителей. Радиодетали дешево возможно приобрести в этом китайском магазине.Плагин на Google Хром для экономии в нём: 7 процентов с приобретений возвращается вам.
Мы привыкли к тому то что звук обязан воспроизводиться с громкоговорителя либо динамика. Ионофон же разрешит воспроизводить тот же звук, применяя ионизированный поток либо же высоковольтную дугу.
И прямо на данный момент вы услышите, как звучит музыка высоковольтной дуги.В действительности конструкция таковой занимательной игрушки достаточно несложна. Складывается из генератора в виде микросхемы NE55, силовой части в лице n-канального высоковольтного трансформатора и полевого транзистора от строчной развертки советского телевизора.
Начнем с генератора. Микросхема NE55 трудится в качестве генератора прямоугольных импульсов с возможностью аудио модуляций. Имеется возможность регулировки частоты в пределах 6-48 КГц посредством подстроечного резистора на плате.
 Звуковой сигнал подается на пятой вывод микросхемы либо вывод контроля через разделительный конденсатор. Разрешает это руководить длительностью выходных импульсов.
Звуковой сигнал подается на пятой вывод микросхемы либо вывод контроля через разделительный конденсатор. Разрешает это руководить длительностью выходных импульсов.Выход микросхемы нагружен полевым транзистором, раскачивающим высоковольтный трансформатор.
Схема ионофона
Полевой транзистор в обязательном порядке устанавливается на радиатор. Подойдут с током от 20 А и с расчетным напряжением выше 40 вольт. Нужно забрать полевики на 100 вольт.
Применять возможно дешёвые IRFZ44, 46, 48 либо им подобные, возможно и высоковольтные.
А сейчас перейдем к строчному трансформатору, подходящему для поющей дуги. В данном варианте схемы Ионофона это ТВС 110 ПЦ 15. В первую очередь на свободную часть сердечника обмотаем 12 витков изолированного миллиметрового проводам.
В принципе диаметр провода возможно от 0,7 до 1,5-2 мм. Позже необходимо отыскать выход высоковольтной обмотки. В большинстве случаев, это обмотка с самым громадным сопротивлением. Мультиметр в этом деле окажет помощь.
 Или возможно скачать документацию в сети, где все детально продемонстрировано. При подобных строчных трансформаторов один из выводов высоковольтной обмотки находится прямо на катушке — второй снизу.
Или возможно скачать документацию в сети, где все детально продемонстрировано. При подобных строчных трансформаторов один из выводов высоковольтной обмотки находится прямо на катушке — второй снизу.Выяснить его весьма легко, потому, что к нему подключен провод в изоляции.
Потом подключаем два изолированных провода к контактам высоковольтной обмотки. Протяженность отрезков 15-20 сантиметров.Поющая дуга
Позже собираем плату Ионофона и подключаем к ней строчный трансформатор. В качестве источника питания сойдет любой блок питания с напряжением от 5 вольт, с током от 2 А. Для самая эффективной работы схемы нужен источник питания с напряжением 10-12 вольт и с токам 2 А и выше. Возможно питать поющую дугу и от аккумуляторная батарей.
Звуковой сигнал подается от любого плеера, планшета либо сотового телефона, применяя штатной разъем для наушников.
Ну а сейчас и наслаждаемся зрелищем. Дуга не просто звучит, но и меняет форму. Это особенно заметно при низких частотах.
Собирается на здоровье, но помните о мерах безопасности! Еще схема для начинающих мастеров электроники.
Случайные записи:
- Вот что можно сделать из старой микроволновки
- Обзор nikon keymission 360
Ионофон(Поющая дуга) на NE555
Похожие статьи, которые вам понравятся:
-
Трансформатор тесла своими руками — простейшая схема
Первый в данной публикации видеоролик канала E-Station — самый простой и доступный для сборки вариант трансформатора тесла, что не сложно будет собрать…
-
Схема зарядного устройства для литиевых li-ion аккумуляторов
В этом видеоуроке, что выложил на своем канале блогер Ака Касьян, вы сможете ознакомиться со схемой зарядного устройства, которая превосходно подойдет…
-
Схема и описание блока питания без трансформатора на 5 вольт, 40-50 ма
Многие радиолюбители не вычисляют блоки питания без трансформаторов.
 Но не обращая внимания на это, они употребляются достаточно деятельно. В частности,…
Но не обращая внимания на это, они употребляются достаточно деятельно. В частности,… -
Схема музыкальной катушки тесла
В этом видео уроке канала youtube «Alpha Mods» будем собирать мелкий поющий качер из покупного китайского набора, продается в этом китайском магазине….
Tags: высоковольтной обмотки, дуга, ионофона, схема
Хлорохин – ионофор цинка
1. Крафтс К., Хемпельманн Э., Скорска-Стания А. (2012) От метиленового синего к хлорохину: краткий обзор разработки противомалярийной терапии. Паразитол Рез 111: 1–6. [PubMed] [Google Scholar]
2. Lamoureux F, Zoubeidi A (2013)Двойное ингибирование аутофагии и пути AKT при раке простаты. аутофагия 9: 1119–1120. [Бесплатная статья PMC] [PubMed] [Google Scholar]
3.
Элдридж Х.Б., Дениттис А., Духадавей Дж.Б., Черник М., Мец Р. и соавт. (2013) Параллельная лучевая терапия всего мозга и краткосрочный курс хлорохина у пациентов с метастазами в головной мозг: экспериментальное исследование.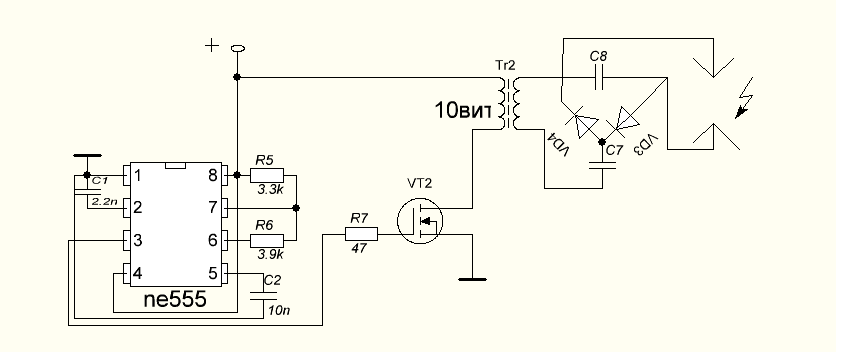 J Радиат Онкол
2. [Бесплатная статья PMC] [PubMed] [Google Scholar]
J Радиат Онкол
2. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Голдберг С.Б., Супко Дж.Г., Нил Дж.В., Музыканский А., Дигумарти С. и соавт. (2012)Исследование фазы I эрлотиниба и гидроксихлорохина при распространенном немелкоклеточном раке легкого. Дж. Торак Онкол 7: 1602–1608. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Рохас-Пуэнтес Л.Л., Гонсалес-Пинедо М., Крисматт А., Ортега-Гомес А., Гамбо-Виньоль С. и др. (2013) Фаза II рандомизированного двойного слепого плацебо-контролируемого исследования облучения всего мозга с сопутствующим хлорохином при метастазах в головной мозг. Радиат Онкол 8: 209. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Цзоу Ю., Линг Ю.Х., Сирони Дж., Шварц Э.Л., Перес-Солер Р. и соавт. (2013) Ингибитор аутофагии хлорохин преодолевает врожденную устойчивость клеток немелкоклеточного рака легкого EGFR дикого типа к эрлотинибу. Дж. Торак Онкол 8: 693–702. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Selvakumaran M, Amaravadi RK, Vasilevskaya IA, O’Dwyer PJ (2013)Ингибирование аутофагии повышает чувствительность клеток рака толстой кишки к антиангиогенной и цитотоксической терапии. Клин Рак Рез
19: 2995–3007. [PubMed] [Google Scholar]
Selvakumaran M, Amaravadi RK, Vasilevskaya IA, O’Dwyer PJ (2013)Ингибирование аутофагии повышает чувствительность клеток рака толстой кишки к антиангиогенной и цитотоксической терапии. Клин Рак Рез
19: 2995–3007. [PubMed] [Google Scholar]
8. Зинн Р.Л., Гарднер Э.Е., Добромильская И., Мерфи С., Маркионни Л. и соавт. (2013)Комбинированное лечение ABT-737 и хлорохином на доклинических моделях мелкоклеточного рака легкого. Мол Рак 12: 16. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Maycotte P, Aryal S, Cummings CT, Thorburn J, Morgan MJ, et al. (2012)Хлорохин повышает чувствительность клеток рака молочной железы к химиотерапии независимо от аутофагии. аутофагия 8: 200–212. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Kimura T, Takabatake Y, Takahashi A, Isaka Y (2013)Хлорохин в терапии рака: обоюдоострый меч аутофагии. Рак Рез 73: 3–7. [PubMed] [Google Scholar]
11.
Бристоль М.Л., Эмери С.М., Майкотт П., Торберн А., Чакрадео С. и др. (2013) Ингибирование аутофагии для химиосенсибилизации и радиосенсибилизации при раке: подтверждают ли доклинические данные эту терапевтическую стратегию? J Pharmacol Exp Ther
344: 544–552. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Соломон В.Р., Ли Х. (2009 г.)) Хлорохин и его аналоги: новое обещание старого препарата для эффективной и безопасной терапии рака. Евр Дж Фармакол 625: 220–233. [PubMed] [Google Scholar]
13. Мизусима Н., Йошимори Т., Левин Б. (2010) Методы исследования аутофагии млекопитающих. Клетка 140: 313–326. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Ламуре Ф., Томас С., Крафтер С., Кумано М., Чжан Ф. и др. (2013)Блокированная аутофагия с использованием лизосомотропных агентов повышает чувствительность резистентных клеток опухоли предстательной железы к новому ингибитору Akt AZD5363. Клин Рак Рез 19: 833–844. [PubMed] [Google Scholar]
15. Yu H, Zhou Y, Lind SE, Ding WQ (2009) Клиохинол нацеливает цинк на лизосомы раковых клеток человека. Биохим Дж 417: 133–139. [PubMed] [Google Scholar]
16.
Zheng J, Zhang XX, Yu H, Taggart JE, Ding WQ (2012)Цинк в цитотоксических концентрациях влияет на посттранскрипционные события экспрессии генов в раковых клетках. Клеточный Физиол Биохим
29: 181–188. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Клеточный Физиол Биохим
29: 181–188. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Ding WQ, Lind SE (2009) Металлические ионофоры — развивающийся класс противоопухолевых препаратов. МЮБМБ Жизнь 61: 1013–1018. [PubMed] [Академия Google]
18. Liuzzi JP, Yoo C (2013) Роль цинка в регуляции аутофагии при воздействии этанола на клетки гепатомы человека. Биол Трейс Элем Рез. [PubMed] [Google Scholar]
19. Блэки М.А. (2013) Металл, содержащий хлорохинолины: не только противомалярийная эффективность наугад, но и твердая наука. Mini Rev Med Chem 13: 597–606. [PubMed] [Google Scholar]
20. Ding WQ, Liu B, Vaught JL, Yamauchi H, Lind SE (2005)Противораковая активность антибиотика клиохинола. Рак Рез 65: 3389–3395. [PubMed] [Google Scholar]
21. Ding WQ, Liu B, Vaught JL, Palmiter RD, Lind SE (2006) Клиохинол и докозагексаеновая кислота действуют синергетически, убивая опухолевые клетки. Мол Рак Тер 5: 1864–1872 гг. [PubMed] [Google Scholar]
22. Canzoniero LM, Manzerra P, Sheline CT, Choi DW (2003)Проникающие в мембрану хелаторы могут ослаблять Zn2+-индуцированную гибель кортикальных нейронов. нейрофармакология
45: 420–428. [PubMed] [Google Scholar]
Canzoniero LM, Manzerra P, Sheline CT, Choi DW (2003)Проникающие в мембрану хелаторы могут ослаблять Zn2+-индуцированную гибель кортикальных нейронов. нейрофармакология
45: 420–428. [PubMed] [Google Scholar]
23. Ding WQ, Yu HJ, Lind SE (2008)Соединения, связывающие цинк, вызывают гибель раковых клеток с помощью различных способов действия. Рак Летт 271: 251–259. [PubMed] [Google Scholar]
24. Li M, Khambu B, Zhang H, Kang JH, Chen X и др. (2013)Подавление функции лизосом индуцирует аутофагию посредством подавления обратной связи активности MTORC1. Дж. Биол. Хим. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Мерсер Э., Рекедал Л., Гарг Р., Лу Б., Массаротти Э.М. и др. (2012)Гидроксихлорохин улучшает чувствительность к инсулину у людей с ожирением, не страдающих диабетом. Артрит Рез Тер 14: 135 р. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26.
Соломон Д.Х., Массаротти Э., Гарг Р., Лю Дж., Каннинг С. и др. (2011) Связь между болезнь-модифицирующими противоревматическими препаратами и риском диабета у пациентов с ревматоидным артритом и псориазом. Джама
305: 2525–2531. [PubMed] [Google Scholar]
Джама
305: 2525–2531. [PubMed] [Google Scholar]
27. Qin Y, Thomas D, Fontaine CP, Colvin RA (2008)Механизмы оттока Zn2+ в культивируемых корковых нейронах. Джей Нейрохим 107: 1304–1313. [PubMed] [Google Scholar]
28. Aiba I, West AK, Sheline CT, Shuttleworth CW (2013) Внутриклеточный диализ нарушает динамику Zn2+ и позволяет селективно обнаруживать приток Zn2+ в препаратах срезов головного мозга. Джей Нейрохим 125: 822–831. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Baum L, Ng A (2004)Взаимодействие куркумина с медью и железом предполагает один возможный механизм действия на животных моделях болезни Альцгеймера. Дж. Альцгеймерс Дис 6: 367–377, обсуждение 443–369. [PubMed] [Google Scholar]
30.
Ричи К.В., Буш А.И., Маккиннон А., Макфарлейн С., Маствик М. и др. (2003)Аттенюация белков металлов с помощью йодохлоргидроксихина (клиохинола), направленная на отложение бета-амилоида и токсичность при болезни Альцгеймера: пилотная фаза 2 клинических испытаний. Арка Нейрол
60: 1685–1691. [PubMed] [Google Scholar]
Арка Нейрол
60: 1685–1691. [PubMed] [Google Scholar]
31. Bincoletto C, Bechara A, Pereira GJ, Santos CP, Antunes F, et al. (2013)Взаимодействие между апоптозом и аутофагией, сложная головоломка: новые взгляды на противоопухолевую химиотерапию. Химия Биол Взаимодействие 206: 279–288. [PubMed] [Google Scholar]
CCCP | КАС № 555-60-2 | Протонофор и разобщитель
Связанный номер CAS: 555-60-2
Синоним: CCCP; мезоксалонитрил 3-хлорфенилгидразон;
ИЮПАК/химическое название: Карбонилцианид 3-хлорфенилгидразон
Ключ ИнЧи: UGTJLJZQQFGTJD-UHFFFAOYSA-N
Код ИнЧи: InChI=1S/C9H5ClN4/c10-7-2-1-3-8(4-7)13-14-9(5-11)6-12/h2-4,13H
СМАЙЛЫ Код: N#C/C(C#N)=N/NC1=CC=CC(Cl)=C1
Вид: Твердый порошок
Чистота: >98% (или см. сертификат анализа)
Состояние доставки:
Транспортируется при температуре окружающей среды как неопасный химикат.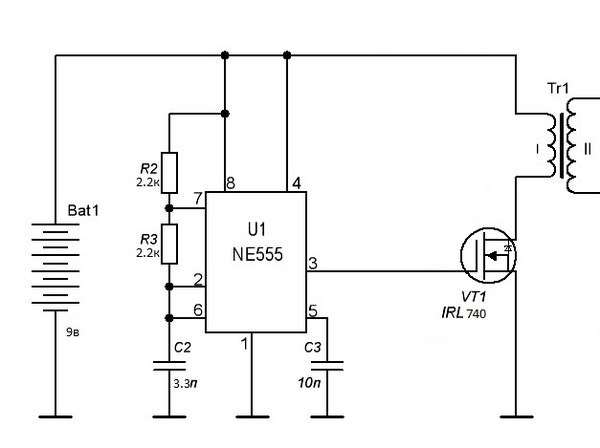 Этот продукт достаточно стабилен в течение нескольких недель при обычной доставке и времени, проведенном на таможне.
Этот продукт достаточно стабилен в течение нескольких недель при обычной доставке и времени, проведенном на таможне.
Условия хранения: Сухой, темный и при температуре от 0 до 4 C на короткое время (от дней до недель) или от -20 C на длительный срок (от месяцев до лет).
Растворимость: Растворим в ДМСО
Срок годности: >3 лет при правильном хранении
Препаративная форма: Этот препарат может быть приготовлен в ДМСО.
Хранение стокового раствора: 0–4 C на короткий срок (от дней до недель) или -20 C на длительный срок (месяцы).
Тарифный код ТН ВЭД: 2934.99.9001
Данные продукта:
Данные продукта
Инструкция:
Посмотреть инструкцию по обращению
| Биологическая мишень: | CCCP представляет собой разобщитель окислительного фосфорилирования (OXPHOS).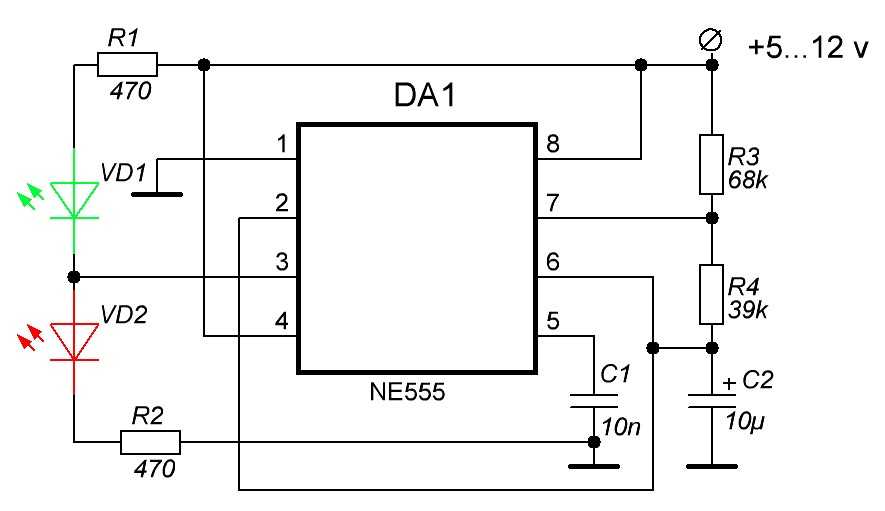 CCCP индуцирует активацию PINK1, что приводит к фосфорилированию Parkin Ser65. CCCP индуцирует активацию PINK1, что приводит к фосфорилированию Parkin Ser65. |
| Активность in vitro: | CCCP ингибирует продукцию IFN-β, индуцированную различными типами активаторов пути STING. CCCP подавляет фосфорилирование STING, TBK1 и IRF3, нарушая связь STING и TBK1. CCCP ингибирует активацию STING и его нижестоящих сигнальных молекул, TBK1 и IRF3, но не транслокацию STING в околоядерную область. CCCP нарушает взаимодействие между STING и TBK1 и одновременно запускает деление митохондрий. Важно отметить, что нокаут ключевого регулятора деления митохондрий Drp1 восстанавливал активность STING, указывая на то, что CCCP понижает модуляцию пути STING посредством DRP1-опосредованной фрагментации митохондрий. Протонофор CCCP, нарушающий мембранный потенциал, подавляет сигнальный путь STING, запускаемый DMXAA. CCCP резко подавляет продукцию IFN-β в обработанных DMXAA клетках RAW264.7 и MEF.
Ссылка: Biochem Biophys Res Commun. 2017 4 ноября; 493(1):737-743. https://linkinghub.elsevier.com/retrieve/pii/S0006-291X(17)31704-7 https://linkinghub.elsevier.com/retrieve/pii/S0006-291X(17)31704-7
|
| Активность in vivo: | Сигналы (99m) Tc-MIBI были проанализированы в сердцах крыс Sprague-Dawley (SD), перфузированных карбонилцианидом м-хлорфенилгидразоном (CCCP), митохондриальным разобщителем, который, как известно, снижает потенциал митохондриальной мембраны. Сигналы (99m)Tc-MIBI можно использовать для обнаружения изменений потенциала митохондриальной мембраны с чувствительностью, сравнимой с чувствительностью, полученной с помощью двухфотонной лазерной микроскопии с катионным зондом тетраметилродамин этилового эфира (TMRE). (9Сигналы 9m)Tc-MIBI также измеряли в сердцах крыс SD, которым вводили CCCP (4 мг/кг внутрибрюшинно) или носитель. Сигналы (99m)Tc-MIBI уменьшились в сердцах крыс, которым вводили CCCP, и одновременно уменьшилось содержание АТФ, измеренное с помощью (31)P магнитно-резонансной спектроскопии. Затем (99m)Tc-MIBI вводили чувствительным к соли крысам Dahl, получавшим диету с высоким содержанием соли, что приводит к гипертонии и сердечной недостаточности.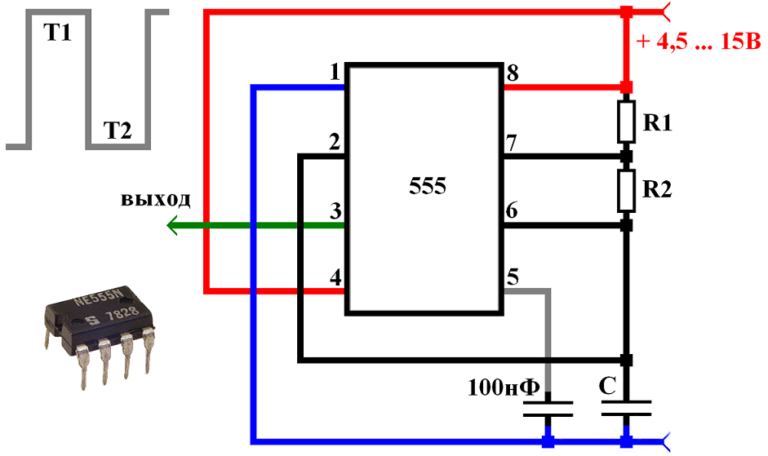 Сигнал (99m)Tc-MIBI на массу ткани сердца обратно коррелировал с массой сердца, сердечной функцией и экспрессией предсердного натрийуретического фактора, маркера сердечной недостаточности, и положительно коррелировал с накоплением меченого аналога жирной кислоты. (9Сигнал 9m)Tc-MIBI на массу ткани печени был ниже, чем на массу ткани сердца.
Ссылка: PLoS One. 16 января 2015 г .; 10 (1): e0117091. https://www.ncbi.nlm.nih.gov/pmc/articles/pmid/25594546/ Сигнал (99m)Tc-MIBI на массу ткани сердца обратно коррелировал с массой сердца, сердечной функцией и экспрессией предсердного натрийуретического фактора, маркера сердечной недостаточности, и положительно коррелировал с накоплением меченого аналога жирной кислоты. (9Сигнал 9m)Tc-MIBI на массу ткани печени был ниже, чем на массу ткани сердца.
Ссылка: PLoS One. 16 января 2015 г .; 10 (1): e0117091. https://www.ncbi.nlm.nih.gov/pmc/articles/pmid/25594546/
|
Данные о растворимости
| Растворитель | Макс. конц. мг/мл | Макс. конц. мМ | |
|---|---|---|---|
| Растворимость | |||
| ДМСО | 41,0 | 200,37 | |
| Этанол | 21,0 | 102,63 |
Подготовка маточных растворов
Следующие данные основаны на
товар
молекулярный вес
204.617
Удельная молекулярная масса партии может варьироваться от партии к партии из-за степени гидратации, которая будет влиять на объемы растворителя, необходимые для приготовления исходных растворов.
| Концентрация / объем растворителя / масса | 1 мг | 5 мг | 10 мг |
|---|---|---|---|
| 1 мМ | 1,15 мл | 5,76 мл | 11,51 мл |
| 5 мМ | 0,23 мл | 1,15 мл | 2,3 мл |
| 10 мМ | 0,12 мл | 0,58 мл | 1,15 мл |
| 50 мМ | 0,02 мл | 0,12 мл | 0,23 мл |
| Протокол in vitro: | 1. Квон Д., Пак Э., Сесаки Х., Канг С.Дж. Карбонилцианид 3-хлорфенилгидразон (CCCP) подавляет STING-опосредованный путь распознавания ДНК, вызывая деление митохондрий. Biochem Biophys Res Commun. 2017 4 ноября; 493 (1): 737-743. doi: 10.1016/j.bbrc.2017.08.121. Epub 2017, 30 августа. PMID: 28859978. |
| Протокол in vivo: | 1. Кавамото А., Като Т., Шиои Т., Окуда Дж., Кавасима Т., Тамаки Й., Ниидзума С.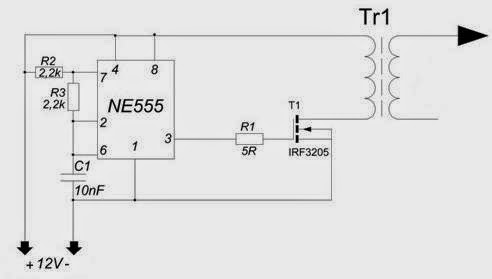 , Танада Й., Такемура Г., Нарадзаки М., Мацуда Т., Кимура Т. Измерение технеция-9Сигналы 9m sestamibi у крыс, которым вводили митохондриальный разобщитель, и в крысиной модели сердечной недостаточности. ПЛОС Один. 16 января 2015 г .; 10 (1): e0117091. doi: 10.1371/journal.pone.0117091. PMID: 25594546; PMCID: PMC4297193.
2. Синха Д., Панди С., Сингх Р., Тивари В., Сад К., Тандон В. Синергическая эффективность комбинации бисбензимидазола и карбонилцианида 3-хлорфенилгидразона против бактериальных штаммов MDR. Научный представитель 2017 г., 17 марта; 7:44419. дои: 10.1038/srep44419. PMID: 28303897; PMCID: PMC5355889. , Танада Й., Такемура Г., Нарадзаки М., Мацуда Т., Кимура Т. Измерение технеция-9Сигналы 9m sestamibi у крыс, которым вводили митохондриальный разобщитель, и в крысиной модели сердечной недостаточности. ПЛОС Один. 16 января 2015 г .; 10 (1): e0117091. doi: 10.1371/journal.pone.0117091. PMID: 25594546; PMCID: PMC4297193.
2. Синха Д., Панди С., Сингх Р., Тивари В., Сад К., Тандон В. Синергическая эффективность комбинации бисбензимидазола и карбонилцианида 3-хлорфенилгидразона против бактериальных штаммов MDR. Научный представитель 2017 г., 17 марта; 7:44419. дои: 10.1038/srep44419. PMID: 28303897; PMCID: PMC5355889.
|
1: Yamaguchi A, Ishikawa H, Furuoka M, Yokozeki M, Matsuda N, Tanimura S, Takeda K. Расщепленный PGAM5 высвобождается из митохондрий в зависимости от опосредованного протеасомами разрыва внешней митохондриальной мембраны во время митофагии. Дж Биохим. 2019 1 января; 165 (1): 19-25. дои: 10.1093/jb/mvy077. PubMed PMID: 30247576.
2: Minanimo-Muta E, Kato T, Shioi T, Tanada Y, Kimura T. Сердечные эффекты острого введения протонофора на крысиной модели. Дж Фарм Фармакол. 2018 Сен;70(9): 1209-1215. дои: 10.1111/jphp.12956. Epub 2018 Jun 25. PubMed PMID: 29943452.
Сердечные эффекты острого введения протонофора на крысиной модели. Дж Фарм Фармакол. 2018 Сен;70(9): 1209-1215. дои: 10.1111/jphp.12956. Epub 2018 Jun 25. PubMed PMID: 29943452.
3: Wauman J, Tavernier J. Внутриклеточный домен рецептора лептина предотвращает митохондриальную деполяризацию и митофагию. Biochim Biophys Acta Mol Cell Res. 2018 сен; 1865 (9): 1312-1325. doi: 10.1016/j.bbamcr.2018.06.009. Epub 2018 Jun 19. PubMed PMID: 29932990.
4: Zhang X, Zhang Y, Wang F, Wang C, Chen L, Liu H, Lu H, Wen H, Zhou T. Механизмы устойчивости к нитрофурантоину и эпидемиологические характеристики среди Клинические изоляты кишечной палочки. Противомикробные агенты Int J. 2018 авг; 52(2):226-232. doi: 10.1016/j.ijantimicag.2018.04.021. Epub 2018 9 мая. PubMed PMID: 29753133.
5: Stanford KR, Taylor-Clark TE. Индуцированная митохондриальной модуляцией активация субпопуляций сенсорных нейронов блуждающего нерва антимицином А, но не CCCP или ротеноном, коррелирует с продукцией митохондриального супероксида.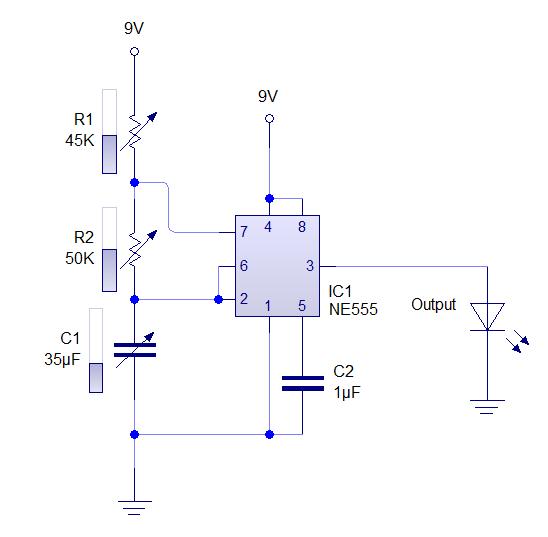 ПЛОС Один. 7 мая 2018 г .; 13 (5): e0197106. doi: 10.1371/journal.pone.0197106. eCollection 2018. PubMed PMID: 29734380; Центральный PMCID в PubMed: PMC5937758.
ПЛОС Один. 7 мая 2018 г .; 13 (5): e0197106. doi: 10.1371/journal.pone.0197106. eCollection 2018. PubMed PMID: 29734380; Центральный PMCID в PubMed: PMC5937758.
6: Ван Ю.Х., Ли С.Н., Чен С., Чжан Дж., Ван Г.К. Обнаружение гена floR и активного механизма оттока Escherichia coli в Нинся, Китай. Микроб Патог. 2018 Апрель; 117: 310-314. doi: 10.1016/j.micpath.2018.02.042. Epub 2018, 23 февраля. PubMed PMID: 29481973.
7: Setz C, Benischke AS, Pinho Ferreira Bento AC, Brand Y, Levano S, Paech F, Leitmeyer K, Bodmer D. Индукция митофагии в слуховой клеточной линии HEI-OC1 и активация Atg12/LC3 пути в кортиевом органе. Услышьте рез. 2018 Апрель; 361: 52-65. doi: 10.1016/j.heares.2018.01.003. Epub 2018 Jan 11. PubMed PMID: 29352609.
8: Kane MS, Paris A, Codron P, Cassereau J, Procaccio V, Lenaers G, Reynier P, Chevrollier A. Текущее механистическое понимание реакции выживания клеток, вызванной CCCP. Биохим Фармакол. 2018 Февраль; 148: 100-110. doi: 10.1016/j.bcp.2017. 12.018. Epub 2017 22 декабря. Обзор. PubMed PMID: 29277693.
12.018. Epub 2017 22 декабря. Обзор. PubMed PMID: 29277693.
9: Park YS, Choi SE, Koh HC. PGAM5 регулирует митофагию, опосредованную PINK1/Parkin, посредством DRP1 при митохондриальной дисфункции, индуцированной CCCP. Токсикол Летт. 2018 1 марта; 284: 120-128. doi: 10.1016/j.toxlet.2017.12.004. Epub 2017, 11 декабря. PubMed PMID: 29241732.
10: Lombardo T, Folgar MG, Salaverry L, Rey-Roldan E, Alvarez EM, Carreras MC, Kornblihtt L, Blanco GA. Регулируемая клеточная гибель лимфомных клеток после постепенного повреждения митохондрий по-разному зависит от лекарств, воздействующих на реакцию клеток на стресс. Основной Клин Фармакол Токсикол. 2018 май;122(5):489-500. doi: 10.1111/bcpt.12945. Epub 2018, 3 января. PubMed PMID: 29205851.
11: Шимасаки К., Ватанабэ-Такахаши М., Умеда М., Фунамото С., Сайто Ю., Ногучи Н., Кумагаи К., Ханада К., Цукахара Ф., Мару Ю., Сибата Н., Наито M, Nishikawa K. Домен гомологии плекстрина p210 BCR-ABL взаимодействует с кардиолипином, регулируя его митохондриальную транслокацию и последующую митофагию.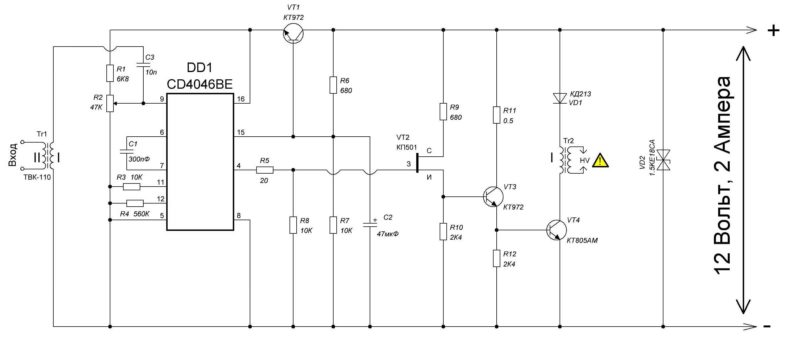 Клетки генов. 2018 янв; 23(1):22-34. doi: 10.1111/gtc.12544. Epub 2017 Dec 5. PubMed PMID: 29205725.
Клетки генов. 2018 янв; 23(1):22-34. doi: 10.1111/gtc.12544. Epub 2017 Dec 5. PubMed PMID: 29205725.
12: Kansaku K, Takeo S, Itami N, Kin A, Shirasuna K, Kuwayama T, Iwata H. Материнское старение влияет на устойчивость ооцитов к митохондриальным индуцированным карбонилцианид-м-хлорфенилгидразоном дисфункции у коров. ПЛОС Один. 2017 28 ноября; 12 (11): e0188099. doi: 10.1371/journal.pone.0188099. eCollection 2017. PubMed PMID: 29182624; Центральный PMCID в PubMed: PMC5705080.
13: член парламента Ярви, Бертон А.Дж. Аденилатный контроль способствует термической акклиматизации тонкокорневого дыхания клена сахарного в экспериментально прогретой почве. Окружающая среда растительной клетки. 2018 март; 41 (3): 504-516. doi: 10.1111/pce.13098. Epub 2017 Dec 11. PubMed PMID: 29124767.
14: Liang P, Rosas-Lemus M, Patel D, Fang X, Tuz K, Juárez O. Динамическая энергетическая зависимость Chlamydia trachomatis от метаболизма клетки-хозяина во время внутриклеточного роста: роль натриевая энергетика в хламидийной генерации АТФ.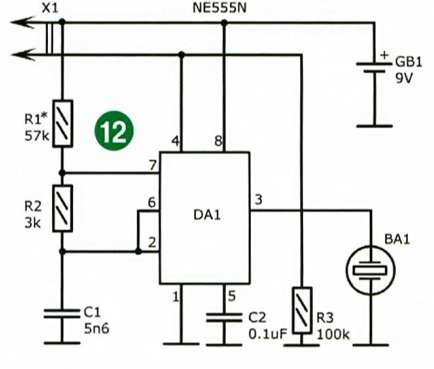 Дж. Биол. Хим. 2018 12 января; 293(2):510-522. doi: 10.1074/jbc.M117.797209. Epub 2017, 9 ноября. PubMed PMID: 29123027; Центральный PMCID в PubMed: PMC5767857.
Дж. Биол. Хим. 2018 12 января; 293(2):510-522. doi: 10.1074/jbc.M117.797209. Epub 2017, 9 ноября. PubMed PMID: 29123027; Центральный PMCID в PubMed: PMC5767857.
15: Хайлова Л.С., Рокитская Т.И., Котова Е.А., Антоненко Ю.Н. Влияние цианида на деполяризацию митохондриальной мембраны, вызванную разобщителями. Биохимия (Москва). 2017 Октябрь;82(10):1140-1146. дои: 10.1134/S0006297917100066. PubMed PMID: 29037134.
16: Xiao B, Deng X, Lim GGY, Xie S, Zhou ZD, Lim KL, Tan EK. Супероксид управляет развитием паркин/PINK1-зависимой митофагии после перемещения паркина в митохондрии. Клеточная смерть Дис. 2017 12 октября; 8 (10): e3097. doi: 10.1038/cddis.2017.463. Опечатка в: Cell Death Dis. 2018 19 июля; 9 (8): 794. PubMed PMID: 298; Центральный PMCID в PubMed: PMC5680585.
17: Polo M, Alegre F, Moragrega AB, Gibellini L, Marti-Rodrigo A, Blas-Garcia A, Esplugues JV, Apostolova N. Протеаза Lon: новый белок митохондриального матрикса во взаимосвязи между лекарственной дисфункцией митохондрий и стресс эндоплазматического ретикулума.