Как устроен и функционирует бактериальный жгутиковый двигатель. Каковы особенности его конструкции и принцип работы. Чем уникален этот биологический наномотор.
Что представляет собой бактериальный жгутиковый двигатель
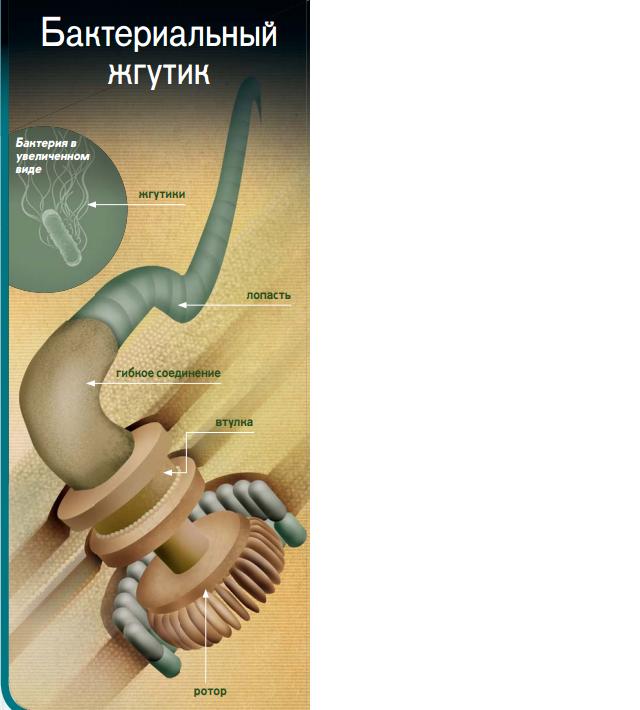
Бактериальный жгутиковый двигатель (BFM) — это сложный молекулярный комплекс, обеспечивающий движение многих бактерий. Он приводит во вращение длинные белковые нити — жгутики, которые проталкивают клетку в жидкой среде. BFM состоит примерно из 20 различных белков и имеет диаметр 40-45 нм. Этот наномотор способен вращаться с частотой до 40-50 тыс. оборотов в минуту, что значительно превышает показатели обычных двигателей.
Основные элементы конструкции бактериального мотора
В устройстве бактериального жгутикового двигателя можно выделить следующие ключевые компоненты:
- Ротор — подвижная часть, вращающаяся с высокой скоростью
- Статор — неподвижная часть, обеспечивающая вращение ротора
- Жгутик — длинная белковая нить, выполняющая роль гребного винта
- Крюк — гибкое соединение между ротором и жгутиком
- Система переключения направления вращения
Жгутик — уникальный гребной винт бактерий
Жгутик представляет собой полую спираль длиной до 10 мкм, что в 10 раз превышает размер самой бактериальной клетки. Он состоит из 11 параллельных нитей, образованных белками FliC. Жгутик соединяется с ротором через гибкий крюк из 120 молекул белка FlgE. Этот крюк выполняет функцию карданного шарнира, позволяя жгутику вращаться под углом к оси ротора для более эффективного движения.
Как происходит сборка и рост жгутика
Рост жгутика происходит на его дальнем конце, а не у основания. Белки жгутика проходят через центральный канал диаметром 2-3 нм и укладываются на своем месте под руководством молекул «кэпа» на кончике. Этот процесс напоминает работу системы секреции III типа, с помощью которой некоторые бактерии выделяют определенные белки. MS-кольцо на внутренней мембране клетки служит основой для сборки всей конструкции и обеспечивает транспорт белков наружу.
Ротор и статор — ключевые компоненты наномотора
Центральную часть ротора образует ось, соединенная с крюком и закрепленная на MS-кольце. Она вращается внутри поддерживающих втулок, зафиксированных в клеточной стенке. Вокруг оси располагается C-кольцо из белков FliG, FliM и FliY, которое взаимодействует со статором. Статор состоит из нескольких комплексов MotA/MotB, заякоренных во внутренней мембране клетки. Именно взаимодействие между C-кольцом и статорными комплексами обеспечивает вращение ротора.
Принцип работы бактериального жгутикового двигателя
Движущей силой BFM является поток протонов через статорные комплексы. Разница концентраций H+ снаружи и внутри клетки создает протонодвижущую силу. Протоны, проходя через канал в MotB, вызывают конформационные изменения в MotA. Это приводит к взаимодействию MotA с белками FliG на C-кольце, заставляя ротор вращаться. Таким образом, химическая энергия градиента протонов преобразуется в механическую энергию вращения.
Уникальные характеристики бактериального наномотора
Бактериальный жгутиковый двигатель обладает рядом впечатляющих характеристик:
- Высокая скорость вращения — до 40-50 тыс. об/мин
- Мгновенное торможение и реверс за четверть оборота
- КПД около 100% при низких скоростях
- Возможность функционировать при частичном отсутствии компонентов
- Самосборка и саморемонт
Эти свойства делают BFM уникальным биологическим наномотором, превосходящим по ряду параметров искусственные аналоги.
Эволюция бактериального жгутикового двигателя
Несмотря на сложность конструкции, бактериальный жгутиковый двигатель, вероятно, возник в ходе длительной эволюции. Исследователи выделяют несколько этапов его развития:
- Появление простой системы секреции белков
- Формирование жгутикоподобных выростов на поверхности клетки
- Возникновение вращательного движения
- Усложнение конструкции и оптимизация работы
Изучение промежуточных форм у современных бактерий подтверждает возможность постепенного формирования этого сложного молекулярного комплекса.
Перспективы изучения и применения бактериальных наномоторов
Исследование бактериальных жгутиковых двигателей представляет большой интерес для науки и технологий. Понимание принципов их работы может помочь в создании искусственных наномашин и молекулярных моторов. Потенциальные области применения включают:
- Разработку новых антибактериальных препаратов
- Создание микро- и нанороботов для медицины
- Конструирование молекулярных насосов и генераторов
- Развитие нанотехнологий и наноэлектроники
Таким образом, бактериальный жгутиковый двигатель остается одним из самых впечатляющих примеров молекулярных машин, созданных природой. Его изучение открывает широкие перспективы для развития науки и технологий.
Коллекторный двигатель постоянного тока: конструкция и принцип действия
Коллекторные электродвигатели довольно распространены в быту и на производстве. Они используются для привода различных механизмов, электроинструмента, в автомобилях. Отчасти популярность обусловлена простой регулировкой оборотов ротора, но есть и некоторые ограничения их применения и конечно же недостатки. Давайте разберемся что такое коллекторный двигатель постоянного тока (КДПТ), какие бывают разновидности данного вида электродвигателей и где они используются.
- Определение и устройство
- Принцип действия
- Виды КДПТ и схемы соединения обмоток
- Схема подключения и реверс
- Сфера применения
- Достоинства и недостатки
Определение и устройство
В справочниках и энциклопедиях приводят, такое определение:
«Коллекторным называется электродвигатель, у которого датчиком положения вала и переключателем обмоток является одно и то же устройство – коллектор. Такие двигатели могут работать либо только на постоянном токе, либо и на постоянном, и на переменном.»
Такие двигатели могут работать либо только на постоянном токе, либо и на постоянном, и на переменном.»
Коллекторный электродвигатель, как и любой другой, состоит из ротора и статора. В этом случае ротор – является якорем. Напомним, что якорем называется та часть электрической машины, которая потребляет основной ток, и в которой индуцируется электродвижущая сила.
Для чего нужен и как устроен коллектор? Коллектор расположен на валу (роторе), и представляет собой набор продольно расположенных пластин, изолированных от вала и друг от друга. Их называют ламелями. К ламелям подключаются отводы секций обмоток якоря (устройство якорной обмотки КДПТ вы видите на группе рисунков ниже), а точнее к каждой из них подключен конец предыдущей и начало следующей секции обмотки.
Ток к обмоткам подаётся через щетки. Щётки образуют скользящий контакт и во время вращения вала соприкасаются то с одной, то с другой ламелью. Таким образом происходит переключение обмоток якоря, для этого и нужен коллектор.
Щеточный узел состоит из кронштейна с щеткодержателями, непосредственно в них и устанавливаются графитовые или металлографитовые щетки. Для обеспечения хорошего контакта щетки прижимаются к коллектору пружинами.
На статоре устанавливаются постоянные магниты или электромагниты (обмотка возбуждения), которые создают магнитное поле статора. В литературе по электрическим машинам вместо слова «статор» чаще используют термины «магнитная система» или «индуктор». На рисунке ниже изображена конструкция ДПТ в разных проекциях. Теперь же давайте разберемся как работает коллекторный двигатель постоянного тока!
Принцип действия
Когда ток протекает через обмотку якоря, возникает магнитное поле, направление которого можно определить с помощью правила буравчика. Постоянное магнитное поле статора взаимодействует с полем якоря, и он начинает вращаться благодаря тому, что одноименные полюса отталкиваются, притягиваясь к разноимённым. Что отлично иллюстрирует рисунок ниже.
При переходе щеток на другие ламели ток начинает протекать в обратную сторону (если рассматривать приведенный выше пример), магнитные полюса меняются местами и процесс повторяется.
В современных коллекторных машинах не используется двухполюсная конструкция из-за неравномерности вращения, в момент переключения направления тока силы, действующие на якорь, будут минимальны. А если включить двигатель, вал которого остановился в этом «переходном» положении — он может и не начать вращаться совсем. Поэтому на коллекторе современного двигателя постоянного тока расположено значительно больше полюсов и секций обмоток, уложенных в пазах шихтованного сердечника, таким образом достигаются оптимальные плавность движения и момент на валу.
Принцип работы коллекторного двигателя простым языком для чайников раскрыт в следующем видеоролике, убедительно рекомендуем ознакомиться.
Виды КДПТ и схемы соединения обмоток
По способу возбуждения коллекторные двигатели постоянного тока различают двух типов:
- С постоянными магнитами (маломощные двигатели мощностью десятки и сотни Ватт).

- С электромагнитами (мощные машины, например, на грузоподъёмных механизмах и станках).
Различают такие типы КДПТ по способу соединения обмоток:
- Последовательного возбуждения (в старой отечественной литературе и от старых электриков можно услышать название «Сериесные», от англ. Serial). Здесь обмотка возбуждения подключена последовательно с обмоткой якоря. Высокий пусковой момент – преимущество такой схемы, а её недостаток – падение частоты вращения с увеличением нагрузки на валу (мягкая механическая характеристика), и то что двигатель идёт вразнос (неконтролируемый рост оборотов с последующим повреждением опорных подшипников и якоря) если работают на холостом ходу или с нагрузкой на валу в меньше 20-30% от номинальной.
- Параллельного (также называют «шунтовые»). Соответственно обмотка возбуждения подключена параллельно обмотке якоря. На низких оборотах на валу высокий момент и стабилен в относительно широком диапазоне оборотов, а с увеличением оборотов он уменьшается.

- Назависимого. Обмотки возбуждения и якоря питаются от разных источников. Такое решение позволяет точнее регулировать обороты вала. Особенности работы похожи на ДПТ с параллельным возбуждением.
- Смешанного. Часть обмотки возбуждения подключена параллельно, а часть последовательно с якорем. Совмещают достоинства последовательного и параллельного типов.
Условное графическое обозначение на схеме вы видите ниже.
В иностранной и современной отечественной литературе, а также на схемах можно встретить и другое представление УГО для КДПТ, как было приведено на предыдущем рисунке в виде круга с двумя квадратами, где круг обозначает якорь, а два квадрата – щетки.
Схема подключения и реверс
Схема соединения обмоток статора и ротора определяется при изготовлении, и, в зависимости от того, где применяется конкретный двигатель, нужно выбирать соответствующее решение. В определенных режимах работы (тормозной режим, например) схемы включения обмоток могут изменяться или вводиться дополнительные элементы.
В определенных режимах работы (тормозной режим, например) схемы включения обмоток могут изменяться или вводиться дополнительные элементы.
Включают маломощные коллекторные двигатели постоянного тока с помощью: полупроводниковых ключей (транзисторов), тумблеров или кнопок, специализированных микросхем-драйверов или с помощью маломощных реле. Крупные мощные машины подключаются к сети постоянного тока через двухполюсные контакторы.
Ниже вы видите реверсивную схему подключения двигателя постоянного тока к сети 220В. На практике, на производстве схема будет аналогичной, но диодного моста в ней не будет, поскольку все линии для подключения таких двигателей прокладываются от тяговых подстанций, где переменный ток выпрямляется.
Реверс осуществляется путем смены полярности на обмотке возбуждения или на якоре. Изменить полярность и там, и там нельзя, поскольку направление вращения вала не изменится, как это происходит с универсальными коллекторными двигателями при работе на переменном токе.
Для плавного пуска двигателя в цепь питания обмотки якоря или обмотки якоря и обмотки возбуждения (в зависимости от схемы их соединения) вводят регулировочное устройство, например, реостат, таким же образом регулируют и частоту вращения вала, но вместо реостата чаще используют набор постоянных резисторов, подключаемых с помощью набора контакторов.
В современных приложениях частота оборотов изменяется с помощью широтно-импульсной модуляции (ШИМ) и полупроводникового ключа, именно так это и сделано в аккумуляторном электроинструменте (шуруповёрт, например). КПД такого способа значительно выше.
Сфера применения
Коллекторные двигатели постоянного тока применяются повсеместно как в быту, так и в промышленных устройствах и механизмах, давайте кратко рассмотрим их область применения:
- В автомобилях используют 12В и 24В коллекторные ДПТ для привода щеток стеклоочистителей (дворников), в стеклоподъёмниках, для запуска двигателя (стартер — это коллекторный двигатель постоянного тока последовательного или смешанного возбуждения) и приводах другого назначения.

- В грузоподъёмных механизмах (краны, лифты и пр.) используются КДПТ, которые работают от сети постоянного тока с напряжением 220В или любым другим доступным напряжением.
- В детских игрушках и радиоуправляемых моделях малой мощности используются КДПТ с трёхполюсным ротором и постоянными магнитами на статоре.
- В ручном аккумуляторном электроинструменте — разнообразные дрели, болгарки, электроотвертки и т.д.
Отметим, что в современный дорогой электроинструмент устанавливают не коллекторные, а бесколлекторные электродвигатели.
Достоинства и недостатки
Разберем плюсы и минусы коллекторного двигателя постоянного тока. Преимущества:
- Соотношение размеров к мощности (массогабаритные показатели).
- Простота регулировки оборотов и реализации плавного пуска.
- Пусковой момент.
Недостатки у КДПТ следующие:
- Износ щеток. Высоконагруженные двигатели, которые регулярно эксплуатируются, требуют регулярного осмотра, замены щеток и обслуживания коллекторного узла.

- Коллектор изнашивается из-за трения щеток.
- Возможно искрение щеток, что ограничивает применение в опасных местах (тогда используют КДПТ взрывозащищенного исполнения).
- Из-за постоянного переключения обмоток этот тип двигателей постоянного тока вносит помехи и искажения в питающие цепи или электросеть, что приводит к сбоям и проблемам в работе других элементов схемы (особенно актуально для электронных схем).
- У ДПТ на постоянных магнитах магнитные силы со временем ослабевают (размагничиваются) и эффективность двигателя снижается.
Вот мы и рассмотрели, что такое коллекторный двигатель постоянного тока, как он устроен и какой у него принцип действия. Если остались вопросы, задавайте их в комментариях под статьей!
Материалы по теме:
- Что такое анод и катод
- Как работает магнитный пускатель
- Как понизить напряжение в сети
- Что такое асинхронный двигатель
Моторчик стеклоочистителя — ремонт и замена своими руками
При движении в условиях выпадения осадков, ветровое стекло покрывается каплями дождя или снегом, которые являются источником ограниченной видимости и ставят под риск управление автомобилем. Нередко возникает вопрос о работоспособности агрегата, предназначенного для очистки лобового стекла от частиц осадков. Постараемся подробно узнать, как проверить мотор стеклоочистителя и контролировать его рабочее состояние.
Нередко возникает вопрос о работоспособности агрегата, предназначенного для очистки лобового стекла от частиц осадков. Постараемся подробно узнать, как проверить мотор стеклоочистителя и контролировать его рабочее состояние.
Что такое моторчик стеклоочистителя?
Все легковые, грузовые и прочие транспортные средства уже с завода обязательно оснащаются механизмами для очистки стекол, теже щетки стеклоочистителя. В стандартном виде, он представляет собой два держателя с резинками на лобовом стекле и один держатель на заднем стекле (для автомобилей с кузовом «универсал» или «хэтчбэк»). Все стеклоочистители поддерживают три режима работы, в процессе которого они передвигаются быстро, медленно или в прерывистом темпе.
Все стеклоочистители снабжаются специальным электродвигателем с редукторной передачей крутящего момента. На валу двигателя крепится два механизма предназначенных для работы стеклоочистителей. Один поворачивает дворники до определенного положения на лобовом стекле, а второй отвечает за перемещение очистителей в исходное положение. Система рычагов поддерживает стеклоочистители на протяжении всей работы.
Система рычагов поддерживает стеклоочистители на протяжении всей работы.
По сути, электродвигатель вращается без остановок, а за перемещение дворников отвечает специальный механизм, который допускает их поворот и быстрый возврат в исходное положение. Этот механизм называется трапеция стеклоочистителя. Он состоит из комплекса рычагов, которые и перемещают дворники по лобовому стеклу.
Помимо стеклоочистителей, за нормальный обзор на фронтальную часть дороги отвечает и система омывания стекол. Он представляет собой бачок со специальной жидкостью, которая под действием установленного в бачке насоса, закачивается в систему и разбрызгивается на лобовом стекле. Крайне важно, чтобы эти механизмы были всегда в исправном и рабочем состоянии. Это связано с тем, что в запланированный день поездки погода может запросто испортиться и движение с неисправной системой стеклоочистителей может стать очень опасной.
Неисправности и ремонт стеклоочистителей
Причин, по которым стеклоочиститель может не работать достаточно много. Вначале, всегда проверяют состояние электродвигателя. Среди неисправностей можно отметить плохой контакт щеток и коллектора, межвитковые замыкания в обмотках статора или ротора или заклинивание ротора и статора в процессе износа опорных подшипников моторчика. Кроме того, неисправность может коснуться самих контактных соединений между двигателем и проводов электрического питания. Однако, если устранение данных неисправностей не привело к положительному результату, рекомендуем провести капитальный ремонт электрического двигателя.
Вначале, всегда проверяют состояние электродвигателя. Среди неисправностей можно отметить плохой контакт щеток и коллектора, межвитковые замыкания в обмотках статора или ротора или заклинивание ротора и статора в процессе износа опорных подшипников моторчика. Кроме того, неисправность может коснуться самих контактных соединений между двигателем и проводов электрического питания. Однако, если устранение данных неисправностей не привело к положительному результату, рекомендуем провести капитальный ремонт электрического двигателя.
В случаях, когда проблема кроется не в состоянии электродвигателя, то есть ряд других причин, по которым стеклоочиститель может не работать. Бывают случаи, когда причиной неисправности может стать заедание рабочих частей. Для предотвращения дальнейших поломок, срабатывает биметаллический предохранитель, который уберегает передаточный механизм моторчика от повреждений. Если предохранитель срабатывает, то дальнейшее использование стеклоочистителя невозможно. Проверьте состояние рабочих частей и при необходимости проведите соответствующий ремонт рычагов. После этого, замените использованный предохранитель.
Проверьте состояние рабочих частей и при необходимости проведите соответствующий ремонт рычагов. После этого, замените использованный предохранитель.
Если механизм стеклоочистителей работает только на одной скорости, то следует обратить особое внимание на состояние переключателя или специального резистора и при необходимости провести его замену. Нередки случаи, когда при работе, стеклоочистители выдают странные звуки при работе. Скрипы и стуки – все это может быть причиной неисправности шарнирного узла механизма. Если вы уверены, что подобные звуки никак не связаны с пылью под резинками или с покрытием стекла, то приобретите ремонтный комплект механизма стеклоочистителя и проведите замену проблемных шарниров.
Однако, если ремонт механизма не дал положительного результата, то рекомендуется провести замену мотора стеклоочистителя. Часто бывает такое, что подшипник двигателя начинает разбалтываться и издавать посторонние шумы при работе. При этом, он ставит под угрозу надежность стеклоочистителей.
Видео — Моторчик омывателя чиним своими руками
Как поменять моторчик стеклоочистителя?
Если это все же случилось, и моторчик перестал подавать какие-либо признаки жизни, наступил момент, когда его следует заменить. Многие водители пытаются отремонтировать электрическую машину самостоятельно. Делать это крайне не рекомендуется, если у вас нет соответствующих знаний и навыков электрика.
Порядок действий при замене неисправного моторчика
1. Выкрутите гайки, с помощью которых осуществляется рычагов щеток. После этого их демонтируют со специальных шлицов в виде конуса. Для облегчения рабочего процесса можно применять жидкость WD-40.
2. Вытащите пластмассовые втулки щеток и прокладки из резины.
3. Поднимите уплотнитель, который находится в небольшой прорези подкапотного пространства, и демонтируйте жгут проводов.
4. Открутите стеклоочиститель, который крепится посредством гаек и шайб. Затем открутите болты крепления моторчика и демонтируйте его.
Затем открутите болты крепления моторчика и демонтируйте его.
5. Установите новый электродвигатель и проведите сборку узлов в обратной последовательности.
В процессе внешнего осмотра необходимо убедиться, что все остальные составляющие находятся в исправном состоянии, и по возможности поменяйте все резиновые и пластиковые втулки. Как только все будет собрано – замена моторчика завершена.
Моторчик стеклоочистителя – самостоятельная диагностика и ремонт + видео » АвтоНоватор
При движении автомобиля в неблагоприятных погодных условиях большое значение имеет своевременная очистка ветрового стекла, и нередко возникает вопрос, как проверить моторчик стеклоочистителя. Основное предназначение агрегата заключается в избавлении ветрового проема автомобиля от влаги, снега и других атмосферных осадков, поэтому разберемся, как поддерживать этот механизм в рабочем состоянии.
Моторчик стеклоочистителя – что это и как работает?
Современные автомобили оснащаются двумя стеклоочистителями, которые для переднего стекла имеют двойную конструкцию (имеется две резинки на щетке для более эффективной работы), а для заднего – одинарный вариант. Большинство стеклоочистителей лобовика работают в трех основных режимах. Эти режимы представляют собой прерывистую, медленную и быструю работу.
Большинство стеклоочистителей лобовика работают в трех основных режимах. Эти режимы представляют собой прерывистую, медленную и быструю работу.
Стеклоочиститель лобовика включает в свою конструкцию электрический двигатель с редуктором, систему рычагов, щетки и механизм, автоматически возвращающий дворники авто в исходное положение. Одновременно с очистителем осуществляется работа омывателя ветрового стекла. Когда этот небольшой, но важный механизм перестает работать, не стоит медлить с его починкой, ведь погода редко бывает предсказуема.
Ремонт моторчика стеклоочистителя – как установить причину поломки?
Может быть несколько причин, почему не работает стеклоочиститель, рассмотрим часто встречающиеся ситуации чуть ниже. В первую очередь может отсутствовать контакт между коллектором и щетками. Если после разборки электродвигателя и зачистки контактов неисправность не устранилась, то в этом случае, скорее всего, необходим ремонт моторчика стеклоочистителя, но есть еще несколько вариантов, которые стоит проверить до кардинальных мер.
В случае, если заклинили рычаги или заел редуктор, может сработать биметаллический предохранитель. Тогда ваша система тоже не будет отзываться на попытки ее включить. Естественно, поправив положение указанных компонентов и заменив предохранитель, можно решить эту проблему. После устранения неисправности стеклоочиститель должен нормально работать.
Иногда этот механизм начинает функционировать только на одной скорости. В этом случае, скорее всего, неисправен переключатель или резистор. Щетки могут не устанавливаться в нижнее положение, для этого необходимо отрегулировать механизм остановки. Нередко бывают ситуации, когда при включении стеклоочистителя возникают посторонние звуки в виде скрипа или скрежета. Если это не связано с сухостью стекла или наличием под резинками щеток песка, то, видимо, барахлит что-то внутри, поэтому пора заглянуть поглубже.
Строение стеклоочистителя не сложное. На пальцы крепятся рычаги щеток, которые могут быть самыми первыми подозреваемыми источниками посторонних звуков. Как правило, они закисают во втулках кронштейна из-за нарушений защиты, когда внутрь попадает вода и грязь. Если же и эту проверку устройство прошло успешно, а звук сохранился, то необходим ремонт или замена моторчика стеклоочистителя.
Как правило, они закисают во втулках кронштейна из-за нарушений защиты, когда внутрь попадает вода и грязь. Если же и эту проверку устройство прошло успешно, а звук сохранился, то необходим ремонт или замена моторчика стеклоочистителя.
Для того чтобы снять моторчик, нужно обесточить систему электрооборудования, и только после этого его можно демонтировать.
Замена моторчика стеклоочистителя – как снять устройство?
Иногда моторчик заднего стеклоочистителя, также как и переднего, попросту выходит из строя. Вряд ли это будет частая поломка, но если она вас застала, ничего не поделаешь, придется разбираться. Для того чтобы произвести ремонт устройства, его нужно снять, а это довольно трудоемкий процесс. Для примера обычно приводятся классические ВАЗовские модели, наиболее полно демонстрирующие весь технический процесс. Итак, снятие моторчика проходит в несколько этапов:
- Откручиваются гайки, которыми крепятся рычаги щеток, после чего их можно снять с конусных шлицев привода.
 Если сделать это трудно, на шлицы может быть нанесена проникающая смазка.
Если сделать это трудно, на шлицы может быть нанесена проникающая смазка. - Приводы рычагов щеток демонтируются путем откручивания гаек крепления, а затем производится снятие пластмассовых втулок и резиновых прокладок.
- Колодка проводов в жгуте отсоединяется и вынимается путем приподнимания края уплотнителя, через прорезь в перегородке моторного отсека.
- Следующим этапом производится непосредственное снятие стеклоочистителя. Затем с него откручивается шайба и пружинный фиксатор, отсоединяется тяга.
- После откручивания трех болтов крепления можно достать и моторчик.
После визуального осмотра все негодные части следует заменить. Если восстановление невозможно, то следует приобрести новый моторчик. Сборка всей системы очистки стекол производится в обратной последовательности.
- Автор: Егор
- Распечатать
Оцените статью:
(15 голосов, среднее: 1. 9 из 5)
9 из 5)
Поделитесь с друзьями!
Adblock
detector
Как работает клеточный двигатель
Ротор, статор, тормоз и трансмиссия: как работает клеточный двигатель, изобретенный бактериями миллиарды лет до первого автомобиля, первого станка и первого человека.
Роман Фишман
С тех пор как десять лет назад в России случился нанотехнологический бум, мы с нетерпением ждем обещанной революции – «умных» приборов, материалов и механизмов, с ювелирной точностью собранных из отдельных молекул и даже атомов. К сожалению, до сих пор разработать, пустить в массовое производство и с успехом применять молекулярные наномашины удалось разве что самой природе. Один из самых поразительных примеров этому дают жгутики обыкновенных бактерий.
Их устройство столь совершенно, что сторонники некоторых псевдонаучных концепций часто упоминают жгутики в числе систем с «неупрощаемой сложностью»: дескать, этот механизм настолько сложен и внутренне взаимосвязан, что никак не мог возникнуть в ходе постепенной эволюции, по частям, а появился сразу в готовом виде, в результате «разумного замысла». Обоснованную критику подобных взглядов легко найти в Сети. Мы же лучше взглянем на устройство этой поразительной машины – пример высоких нанотехнологий от организмов, которые мы привыкли считать низшими.
Обоснованную критику подобных взглядов легко найти в Сети. Мы же лучше взглянем на устройство этой поразительной машины – пример высоких нанотехнологий от организмов, которые мы привыкли считать низшими.
youtube
Нажми и смотри
Двигатель
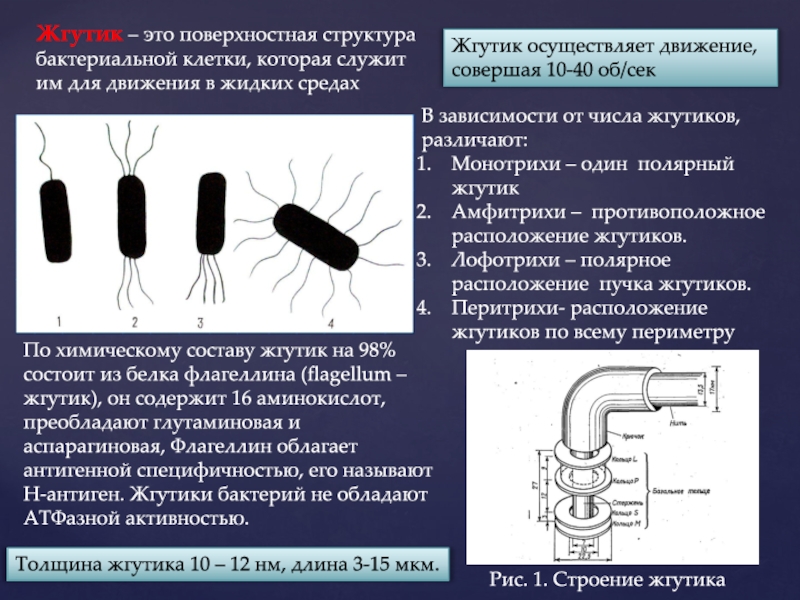
Большинство бактерий, способных к активному движению, используют для этого жгутики – длинные и упругие белковые нити, быстрое вращение которых проталкивает клетку в жидкости. Мощность этого молекулярного устройства поразительна, позволяя микробу преодолевать дистанцию до 100 длин собственного тела в секунду. Для сравнения, разогнавшийся гепард покрывает за секунду порядка 25 длин своего тела, – а ведь бактерии приходится двигаться в водной среде, весьма вязкой для крошечных микроорганизмов.
Жгутик приводится в движение огромным молекулярным комплексом, сложенным из примерно 20 различных белков – базальным телом, или «бактериальным жгутиковым двигателем» (BFM – Bacterial Flagellar Motor). Эта громадная (40-45 нм в диаметре) машина намертво вмонтирована в оболочку клетки, вращая свой ротор с частотой до 40-50 тыс. об./мин (для сравнения, двигатели болидов Формулы 1 совершают около 15 тыс. об./мин). При этом бактериальный «мотор» способен затормозить всего за четверть оборота – и тут же запуститься в противоположном направлении.
Эта громадная (40-45 нм в диаметре) машина намертво вмонтирована в оболочку клетки, вращая свой ротор с частотой до 40-50 тыс. об./мин (для сравнения, двигатели болидов Формулы 1 совершают около 15 тыс. об./мин). При этом бактериальный «мотор» способен затормозить всего за четверть оборота – и тут же запуститься в противоположном направлении.
В устройстве BFM действительно находится много общего с «настоящим» роторным двигателем. Помимо подвижной части – собственно, ротора, – здесь имеется и толкающий винт (жгутик), соединенный с ним посредством карданного шарнира (крюка). Он вращается, удерживаясь в полых втулках скольжения, и приводится в действие неподвижной частью – статором. Часть устройства играет роль трансмиссии, обеспечивая переключение «передней» (по часовой стрелке) и «задней» (скоростей). Впрочем, обо всем по порядку.
Гребной винт и кардан
Жгутик – самая длинная часть этого двигателя. У бактерий кишечной палочки, на примере которых изучено большинство деталей BFM, жгутик достигает 10 мкм (10000 нм) – вдесятеро больше самой клетки. Он представляет собой полую спираль, сложенную из 11 параллельных нитей – цепочек белков FliC – и соединяется с ротором через небольшой гибкий «крюк», образованный 120-ю копиями белка FlgE. Крюк выполняет роль карданного шарнира, позволяя жгутику вращаться под углом к оси самого ротора и эффективнее проталкивать клетку вперед.
Он представляет собой полую спираль, сложенную из 11 параллельных нитей – цепочек белков FliC – и соединяется с ротором через небольшой гибкий «крюк», образованный 120-ю копиями белка FlgE. Крюк выполняет роль карданного шарнира, позволяя жгутику вращаться под углом к оси самого ротора и эффективнее проталкивать клетку вперед.
Канал диаметром 2-3 нм, проходящий по центру жгутика и крюка, обеспечивает их рост. В отличие от наших волос, которые удлиняются у основания, «винт» бактериального двигателя наращивается на кончике. MS-кольцо из 26 белков FliF первым складывается и закрепляется на внутренней, основной мембране клетки, после чего начинает «выкачивать» наружу белки крюка, а затем и самого жгутика. Пройдя сквозь полость, эти белки укладываются на свое место, направляемые молекулами «кэпа» (белки FliD) – дальнего кончика жгутика.
Неудивительно, что MS-кольцо демонстрирует высокое сходство с другой белковой машиной – системой секреции III типа. С ее помощью некоторые бактерии выкачивают из клетки определенные белки, и рост жгутика действительно напоминает контролируемое выделение белков, которые складывают его детали одну за другой. Впрочем, синтез крюка и жгутика начинается позднее: при сборке бактериального двигателя следом после MS-кольца формируются его ротор и статор.
С ее помощью некоторые бактерии выкачивают из клетки определенные белки, и рост жгутика действительно напоминает контролируемое выделение белков, которые складывают его детали одну за другой. Впрочем, синтез крюка и жгутика начинается позднее: при сборке бактериального двигателя следом после MS-кольца формируются его ротор и статор.
Ротор и статор
Центральную деталь ротора образует ось, которая одним концом соединена с крюком, а другим крепится на MS-кольце. Она вращается внутри полых поддерживающих втулок, которые фиксируют ось в многочисленных оболочках бактериальной клетки. Во внутренней мембране роль втулки играет MS-кольцо, во внешней липосахаридной – L-кольцо, а в клеточной стенке, состоящей из пептидогликанов, – P-кольцо. С внутренней стороны клетки к MS-кольцу крепится большое С-кольцо, состоящее из более чем 150 отдельных белковых молекул. Именно на него передается усилие со статора.
Неподвижный статор состоит из 8-12 одинаковых элементов – комплексов из белков MotA и MotВ. MotВ намертво крепятся в клеточной стенке, а MotA связаны с C-кольцом и приводят его в движение. Каждый комплекс статора содержит два ионных канала, сквозь которые протоны движутся внутрь клетки, обеспечивая машину энергией. Прохождение протона меняет форму MotA, заставляя нижнюю часть молекулы выпрямляться, подобно пружине, и передавать усилие на белки FliG, из которых выложена верхняя часть С-кольца.
MotВ намертво крепятся в клеточной стенке, а MotA связаны с C-кольцом и приводят его в движение. Каждый комплекс статора содержит два ионных канала, сквозь которые протоны движутся внутрь клетки, обеспечивая машину энергией. Прохождение протона меняет форму MotA, заставляя нижнюю часть молекулы выпрямляться, подобно пружине, и передавать усилие на белки FliG, из которых выложена верхняя часть С-кольца.
При достаточном потоке протонов ротор начинает вращение, как в шаговом электродвигателе, – отдельными быстрыми перемещениями. В бактериальном моторе на каждый оборот приходится 26 таких шагов, по числу белков FliG, которые идут по периметру C-кольца. Стоит сказать, что за создание избытка протонов снаружи клетки отвечают другие молекулярные машины, занятые их активным перекачиванием через мембрану. А некоторые бактерии могут использовать в качестве источника энергии градиент других ионов – например, натрия.
Трансмиссия
Даже такой замечательный и сложный нанодвигатель был бы совершенно бесполезен, если бы его работу нельзя было контролировать. На самом деле, жгутик со всей его машинерией тесно интегрирован в другие системы бактериальной клетки, которые вместе обеспечивают реакции хемотаксиса – движение в сторону, где привлекательных веществ больше, или наоборот, меньше отталкивающих.
На самом деле, жгутик со всей его машинерией тесно интегрирован в другие системы бактериальной клетки, которые вместе обеспечивают реакции хемотаксиса – движение в сторону, где привлекательных веществ больше, или наоборот, меньше отталкивающих.
Роль датчиков при этом выполняют расположенные на клеточной мембране рецепторы – «метилируемые белки хемотаксиса». Разные виды этих белков реагируют на разные вещества, связываясь с ними и передавая сигнал клетке. Для этого рецепторы присоединяют фосфатную группу к сигнальным молекулам – например, у той же кишечной палочки такую функцию выполняют небольшие белки CheY. В зависимости от баланса свободных молекул CheY и фосфорилированных CheY‐P происходит переключение работы жгутика.
В состав С-кольца входят 34 молекулы белка FliM, с которыми связываются CheY‐P. Это заставляет их менять форму, а следом меняется и форма белков FliG, которые непосредственно связаны со статором и обеспечивают вращение. Один вариант FliG вызывает движение по часовой стрелке, другой – против, и чем больше фоссфорилированных CheY‐P связывает FliM, – тем сильнее «тянут» они двигатель вращаться по часовой стрелке.
Один вариант FliG вызывает движение по часовой стрелке, другой – против, и чем больше фоссфорилированных CheY‐P связывает FliM, – тем сильнее «тянут» они двигатель вращаться по часовой стрелке.
Тормоза
Последней по порядку – но не по значению – деталью бактериального двигателя является тормоз. В отличие от автомобильных колодок, которые намертво сжимают диск колеса, жгутику этого не требуется: вращаясь в чрезвычайно вязкой среде, он сам моментально остановится, как только перестанет получать усилие. Поэтому устройство тормоза BFM напоминает не тормоза автомобиля, а ту же трансмиссию: он лишает контакта ротор и статор, и двигатель перестает вращаться сам по себе.
Тормозной механизм BFM был изучен на примере обыкновенных Bacillus subtilis. Эти исключительно широко распространенные и безвредные почвенные бактерии способны переходить от подвижной планктонной фазы к фазе биопленки, в которой они образуют сложные стационарные микробные сообщества и не нуждаются в активном перемещении. При этом их жгутиковый двигатель отключается под действием «тормозных» белков EpsE.
При этом их жгутиковый двигатель отключается под действием «тормозных» белков EpsE.
Синтез EpsE начинается при переходе клетки к фазе биопленки. Эти белки связываются с FliG, которые входят в состав С-кольца двигателя. В результате форма FliG меняется, и они теряют контакт с белками статора MotA, – как колеса автомобиля, стоящего «на нейтралке». Движение жгутика прекращается вплоть до того момента, когда бактерии снова потребуется плавать, – и она снова запустит свой удивительный нанодвигатель.
особенности, преимущества и недостатки моторов
Идея роторного двигателя слишком заманчива: когда и конкурент весьма далек от идеала, кажется, что вот-вот преодолеем недостатки и получим не мотор, а само совершенство… Mazda находилась в плену этих иллюзий аж до 2012 года, когда была снята с производства последняя модель с роторным двигателем — RX-8.
История создания роторного двигателя
Второе имя роторного двигателя (РПД) — ванкель (этакий аналог дизеля). Именно Феликсу Ванкелю сегодня приписываются лавры изобретателя роторно-поршневого двигателя и даже рассказывается трогательная история о том, как Ванкель шел к поставленной цели тогда же, когда Гитлер шел к своей.
Именно Феликсу Ванкелю сегодня приписываются лавры изобретателя роторно-поршневого двигателя и даже рассказывается трогательная история о том, как Ванкель шел к поставленной цели тогда же, когда Гитлер шел к своей.
На самом деле все было чуточку иначе: талантливый инженер, Феликс Ванкель действительно трудился над разработкой нового, простого двигателя внутреннего сгорания, но это был другой двигатель, основанный на совместном вращении роторов.
После войны Ванкель был привлечен немецкой фирмой NSU, занимавшейся в основном выпуском мотоциклов, в одну из рабочих групп, трудившихся над созданием роторного двигателя под руководством Вальтера Фройде.
Вклад Ванкеля — это обширные исследования уплотнений вращающихся клапанов. Базовая схема и инженерная концепция принадлежат Фройде. Хотя у Ванкеля был патент на двойственное вращение.
Первый двигатель имел вращающуюся камеру и неподвижный ротор. Неудобство конструкции навело на мысль поменять схему местами.
Первый двигатель с вращающимся ротором начал работу в середине 1958 года. Он мало отличался от своего потомка наших дней — разве что свечи пришлось перенести на корпус.
Феликс Ванкель и его первый роторный двигатель
Вскоре фирма объявила о том, что ей удалось создать новый и очень перспективный двигатель. Почти сотня компаний, занимающихся производством автомобилей, закупила лицензии на выпуск этого мотора. Треть лицензий оказалась в Японии.
РПД в СССР
А вот Советский Союз лицензию не покупал вовсе. Разработки собственного роторного двигателя начались с того, что в Союз привезли и разобрали немецкий автомобиль Ro-80, производство которого NSU начала в 1967 году.
Через семь лет после этого на заводе ВАЗ появилось конструкторское бюро, разрабатывающее исключительно роторно-поршневые двигатели. Его трудами в 1976 году возник двигатель ВАЗ-311. Но первый блин получился комом, и его дорабатывали еще шесть лет.
Первый советский серийный автомобиль с роторным двигателем — это ВАЗ-21018, представленный в 1982 году. К сожалению, уже в опытной партии у всех машин вышли из строя моторы. Дорабатывали еще год, после чего появился ВАЗ-411 и ВАЗ 413, которые были взяты на вооружение силовыми ведомствами СССР. Там не особо переживали за расход топлива и малый ресурс мотора, зато нуждались в быстрых, мощных, но неприметных авто, способных угнаться за иномаркой.
ВАЗ с роторным двигателем (ГАИ)
РПД на Западе
На Западе роторный двигатель не произвел бума, а конец его разработкам в США и Европе положил топливный кризис 1973 года, когда цены на бензин резко взлетели, и покупатели машин стали прицениваться к моделям с экономным расходованием топлива.
Если учесть, что роторный двигатель съедал до 20 литров бензина на сотню км, продажи его во время кризиса упали до предела.
Единственной страной на Востоке, не утратившей веру, стала Япония. Но и там производители довольно быстро охладели к двигателю, который никак не желал совершенствоваться. И в конце концов там остался один стойкий оловянный солдатик — компания Mazda. В СССР топливный кризис не ощущался. Производство машин с РПД продолжалось и после распада Союза. ВАЗ прекратил заниматься РПД только в 2004 году. Mazda смирилась только в 2012.
Особенности роторного мотора
В основу конструкции положен ротор треугольной формы, каждая из граней которого имеет выпуклость (треугольник Рёло). Ротор вращается по планетарному типу вокруг центральной оси — статора. Вершины треугольника при этом описывают сложную кривую, именуемую эпитрохоидой. Форма этой кривой обуславливает форму капсулы, внутри которой вращается ротор.
У роторного мотора те же четыре такта рабочего цикла, что и у его конкурента — поршневого мотора.
Камеры образуются между гранями ротора и стенками капсулы, их форма — переменная серповидная, что является причиной некоторых существенных недостатков конструкции. Для изоляции камер друг от друга используются уплотнители — радиальные и торцевые пластины.
Для изоляции камер друг от друга используются уплотнители — радиальные и торцевые пластины.
Если сравнивать роторный ДВС с поршневым, то первым бросается в глаза то, что за один оборот ротора рабочий ход происходит три раза, а выходной вал при этом вращается в три раза быстрее, чем сам ротор.
У РПД отсутствует система газораспределения, что весьма упрощает его конструкцию. А высокая удельная мощность при малом размере и весе агрегата являются следствием отсутствия коленвала, шатунов и других сопряжений между камерами.
Достоинства и недостатки роторных двигателей
Преимущества
-
Роторный двигатель хорош тем, что состоит из куда меньшего числа деталей, чем его конкурент — процентов на 35-40.
-
Два двигателя одинаковой мощности — роторный и поршневый — будут сильно отличаться габаритами. Поршневый в два раза больше.
-
Роторный мотор не испытывает большой нагрузки на высоких оборотах даже в том случае, если на низкой передаче разгонять машину до скорости более 100 км/ч.

-
Автомобиль, на котором стоит роторный двигатель, проще уравновесить, что дает повышенную устойчивость машины на дороге.
-
Даже самые легкие из транспортных средств не страдают от вибрации, потому что РПД вибрирует куда меньше, чем «поршневик». Это происходит в силу большей сбалансированности РПД.
Недостатки
-
Главным недостатком роторного двигателя автомобилисты назвали бы его малый ресурс, который является прямым следствием его конструкции. Уплотнители изнашиваются крайне быстро, так как их рабочий угол постоянно меняется.
-
Мотор испытывает перепады температур через каждый такт, что также способствует износу материала. Добавьте к этому давление, которое оказывается на трущиеся поверхности, что лечится только впрыскиванием масла непосредственно в коллектор.

-
Износ уплотнителей становится причиной утечки между камерами, перепады давления между которыми слишком велики. Из-за этого КПД двигателя падает, а вред экологии растет.
-
Серповидная форма камер не способствует полноте сгорания топлива, а скорость вращения ротора и малая длина рабочего хода — причина выталкивания еще слишком горячих, не до конца сгоревших газов на выхлоп. Помимо продуктов сгорания бензина там еще присутствует масло, что в совокупности делает выхлоп весьма токсическим. Поршневый — приносит меньше вреда экологии.
-
Непомерные аппетиты двигателя на бензин уже упоминались, а масло он «жрет» до 1 литр на 1000 км. Причем стоит раз забыть про масло и можно попасть на крупный ремонт, если не замену двигателя.
-
Высокая стоимость — из-за того, что для изготовления мотора нужно высокоточное оборудование и очень качественные материалы.

Как видите, недостатков у роторного двигателя полно, но и поршневый мотор несовершенен, поэтому состязание между ними не прекращалось так долго. Закончилось ли оно навсегда? Время покажет.
Рассказываем как устроен и работает роторный двигатель
Мотор-редуктор и мотор-редукторы | SEW-EURODRIVE
Наша модульная система мотор-редукторов ориентируется на многообразие ваших сфер применения. Выберите для своего привода идеальный вариант из мотор-редукторов стандартного исполнения, для сервопривода, с вариатором, из нержавеющей стали или взрывозащищенных.
Что такое мотор-редуктор?
Мотор-редуктор Мотор-редукторМотор-редуктор – это единый компактный узел, состоящий из редуктора и двигателя. В электроприводной технике, изготавливаемой компанией SEW-EURODRIVE, двигатель всегда электрический. Идея „агрегата из двигателя и редуктора“ восходит к патенту конструктора и предпринимателя Альберта Обермозера из г. Брухзаль от 1928 года: он изобрел так называемый „двигатель с промежуточной передачей“.
Идея „агрегата из двигателя и редуктора“ восходит к патенту конструктора и предпринимателя Альберта Обермозера из г. Брухзаль от 1928 года: он изобрел так называемый „двигатель с промежуточной передачей“.
С тех пор мотор-редукторы постоянно совершенствовались, были изобретены новые типы редукторов. Двигатели постоянного тока утратили свое значение, поэтому сегодня редукторы чаще всего комбинируются с двигателями переменного тока или с серводвигателями.
Как работает мотор-редуктор?
Главным компонентом мотор-редуктора является редуктор с его ступенями – парами зубчатых колес. Они передают усилие двигателя от входной стороны к выходной. Таким образом, редуктор работает как преобразователь вращающего момента и частоты вращения.
В большинстве случаев применения редуктор замедляет скорость вращения двигателя, а вращающий момент при этом становится значительно больше, чем у электродвигателя без редуктора. Поэтому от конструкции редуктора зависит, будет ли мотор-редуктор использоваться для малых, средних или тяжелых нагрузок, для коротких или долгих периодов включенного состояния.
В зависимости от того, уменьшает или увеличивает редуктор частоту вращения двигателя (т. е. частоту вращения на входе), говорят о понижающем или повышающем редукторе. Мерой этого служит передаточное отношение i между значениями частоты вращения на входе и выходе редуктора.
Еще одним важным параметром мотор-редуктора является максимальный вращающий момент на выходном валу. Он указывается в ньютон-метрах (Нм) и является мерой усилия мотор-редуктора и нагрузки, которую он может привести в движение этим усилием.
Какие типы мотор-редукторов существуют?
Тип мотор-редуктора определяется прежде всего направлением передачи усилия в редукторе. При этом различают три основных варианта конструкции: редуктор с параллельными валами, угловой редуктор и планетарный редуктор.
Где применяются мотор-редукторы?
Возможности применения мотор-редукторов чрезвычайно разнообразны. Без мотор-редукторов остановились бы целые отрасли экономики по всему миру. Так, в промышленном производстве они приводят в движение бесчисленные конвейерные линии, поднимают и опускают грузы и перемещают самые разные товары в различных системах транспортировки из пункта А в пункт Б.
Так, в промышленном производстве они приводят в движение бесчисленные конвейерные линии, поднимают и опускают грузы и перемещают самые разные товары в различных системах транспортировки из пункта А в пункт Б.
Вот лишь малая доля возможных применений:
В автомобилестроении мотор-редукторы можно встретить на каждом этапе производства от штамповки кузовных деталей до окончательной сборки. А в производстве безалкогольных напитков они перемещают бутылки, упаковки и ящики, а также применяются при розливе напитков или сортировке пустой тары. Вся внутренняя логистика производственных предприятий полностью зависит от приводов, будь то складирование, сортировка или выдача товара.
Также и в аэропортах без мотор-редукторов ничего бы уже не двигалось, и пассажиры напрасно ждали бы своего багажа в зоне выдачи.
Манипуляторы и роботы, для которых очень важна высокая динамика и точность движений, были бы немыслимы без мотор-редукторов для сервопривода.
И последнее, но не менее важное: совсем не было бы некоторых аттракционов в индустрии развлечений, и мы, наверное, не знали бы, как захватывает дух на американских горках.
Наверх
‘ data-close-others=»true» data-title=»Обозначение типа » data-max-width=»90%» >1
R37 DRE90L4:
- R = R.. series helical gear unit (two and three stages)
- 37 = gear unit size 37
- DRE = asynchronous DRE.. series AC motor (efficiency class IE2)
- 90 = motor size 90
- L = long length
- 4 = 4-pole
‘ data-close-others=»true» data-title=»Заводской номер » data-max-width=»90%» >2
The gearmotor’s serial number is used, for example, to order appropriate replacement parts.
‘ data-close-others=»true» data-title=»Номинальная частота » data-max-width=»90%» >3
The mains frequency to which the gearmotor can be connected.
‘ data-close-others=»true» data-title=»Скорость мотора/редуктора » data-max-width=»90%» >4
Ratio between the motor’s rated speed and the speed at the gear unit’s output shaft in rpm (revolutions per minute), depending on the nominal frequency applied (here 50 Hz).
- Motor speed 1430 revolutions per minute
- Gear unit output speed 141 revolutions per minute
‘ data-close-others=»true» data-title=»Номинальное напряжение» data-max-width=»90%» >5
Permitted voltage range in which the gearmotor can be operated:
- Lower value: Max. voltage to which one phase (winding) of the installed motor can be subjected (here 220-242 V)
- Higher value: Max. voltage the motor’s outer conductor can accommodate (here 380-420 V)
- These values are valid for the nominal frequency applied (here 50 Hz)
‘ data-close-others=»true» data-title=»Номинальная мощность » data-max-width=»90%» >6
Rated power and operating mode:
- Rated power in kW (here 1.5 kW)
- Here operating mode S1: Continuous operation with a constant load
‘ data-close-others=»true» data-title=»Номинальный ток » data-max-width=»90%» >7
Permitted current range in which the gearmotor can be operated:
- Higher value (here 6.
 00 A): Maximum current to which one phase (winding) of the installed motor can be subjected (corresponds to maximum voltage of 230 V)
00 A): Maximum current to which one phase (winding) of the installed motor can be subjected (corresponds to maximum voltage of 230 V) - Lower value (here 3.45 A): Maximum current the motor’s outer conductor can accommodate (corresponds to maximum voltage of 400 V)
- These values are valid for the nominal frequency applied (here 50 Hz)
‘ data-close-others=»true» data-title=»cos φ» data-max-width=»90%» >8
Phase shift angle with sinusoidal currents and voltages (AC motors)
‘ data-close-others=»true» data-title=»КПД » data-max-width=»90%» >9
Indicates how energy efficient the gearmotor is. When operating at 50 Hz, this gearmotor has an efficiency of 84% and is in line with IE2.
‘ data-close-others=»true» data-title=»Номинальная частота » data-max-width=»90%» >10
The mains frequency to which the gearmotor can be connected (here 60 Hz).
‘ data-close-others=»true» data-title=»Скорость мотора/редуктора » data-max-width=»90%» >11
Ratio between the motor’s rated speed and the speed at the gear unit’s output shaft in rpm (revolutions per minute), depending on the nominal frequency applied (here 60 Hz).
- Motor speed 1745 revolutions per minute
- Gear unit output speed 173 revolutions per minute
‘ data-close-others=»true» data-title=»Номинальное напряжение » data-max-width=»90%» >12
Permitted voltage range in which the gearmotor can be operated:
- Lower value: Max. voltage to which one phase (winding) of the installed motor can be subjected (here 254-277 V)
- Higher value: Max. voltage the motor’s outer conductor can accommodate (here 440-480 V)
- These values are valid for the nominal frequency applied (here 60 Hz)
‘ data-close-others=»true» data-title=»Номинальная мощность » data-max-width=»90%» >13
Rated power and operating mode:
- Rated power in kW (here 1.
 5 kW)
5 kW) - Here operating mode S1: Continuous operation with a constant load
‘ data-close-others=»true» data-title=»Номинальный ток » data-max-width=»90%» >14
Permitted current range in which the gearmotor can be operated:
- Higher value (here 4.95 A): Maximum current to which one phase (winding) of the installed motor can be subjected (corresponds to maximum voltage of 254-277 V)
- Lower value (here 2.85 A): Maximum current the motor’s outer conductor can accommodate (corresponds to maximum voltage of 440-480 V)
- These values are valid for the nominal frequency applied (here 60 Hz)
‘ data-close-others=»true» data-title=»cos φ» data-max-width=»90%» >15
Phase shift angle with sinusoidal currents and voltages (AC motors)
‘ data-close-others=»true» data-title=»КПД» data-max-width=»90%» >16
Indicates how energy efficient the gearmotor is. When operating at 60 Hz, this gearmotor has an efficiency of 85.5% and is in line with IE2.
When operating at 60 Hz, this gearmotor has an efficiency of 85.5% and is in line with IE2.
‘ data-close-others=»true» data-title=»Класс изоляции » data-max-width=»90%» >17
The thermal class or insulating material classification indicates the maximum temperature to which the insulation can be subjected at the rated power. In other words, the material used for the gearmotor’s insulating system can withstand temperatures up to the one indicated.
According to the nameplate shown here, the gearmotor complies with insulating material classification B and is designed for a max. temperature of up to 130°C.
‘ data-close-others=»true» data-title=»Коэффициент перегрузки » data-max-width=»90%» >18
The motor’s permitted overload factor in line with NEMA Section 12.51. Indicates how much above the indicated rated power the motor can be loaded without being damaged.
‘ data-close-others=»true» data-title=»Передаточное число » data-max-width=»90%» >19
The factor by which e.g. the speed changes between the gear unit’s output and input sides.
i = 10.11: 1011 revolutions per minute on the gear unit would be converted into a speed of 100 revolutions per minute
‘ data-close-others=»true» data-title=»Максимальный крутящий момент » data-max-width=»90%» >20
«Nm 101/83» indicates the maximum output torque – 101 Nm with 50 Hz operation and 83 Nm with 60 Hz operation.
‘ data-close-others=»true» data-title=»Монтажная позиция » data-max-width=»90%» >21
The spatial orientation in the room/system for which the gearmotor is designed. Depending on the mounting position, a different lubricant fill quantity (oil volume) and possibly an oil expansion tank may be required.
‘ data-close-others=»true» data-title=»Объем масла » data-max-width=»90%» >22
«CLP HC-460-NSF-h2 Lebmi.Öl/0,30 l» as displayed here means:
- CLP HC-460 – fully synthetic lubricant with a viscosity of 460 mm²/s (40°C)
- NSF-h2: Classification for foodstuff applications – used when contact with food cannot be ruled out if damage occurs
- Fill quantity 0.30 liters
‘ data-close-others=»true» data-title=»Масса » data-max-width=»90%» >23
This gearmotor’s weight (here 29.108 kg)
‘ data-close-others=»true» data-title=»Количество фаз/стандарт » data-max-width=»90%» >24
«3~IEC60034» has the following meaning:
- «3~»: 3-phase motor
- «IEC60034»: International standard IEC 60034 is the underlying rating and performance standard
‘ data-close-others=»true» data-title=»Степень защиты » data-max-width=»90%» >25
Degree of protection IP 54 has the following meaning:
- First number (5): Complete protection against contact, protection against internal dust accumulation
- Second number (4): Protected against spraying water
‘ data-close-others=»true» data-title=»» data-max-width=»90%» >26
Ключ
-
1
Обозначение типа
-
2
Заводской номер
-
3
Номинальная частота
-
4
Скорость мотора/редуктора
-
5
Номинальное напряжение
-
6
Номинальная мощность
-
7
Номинальный ток
-
8
cos φ
-
9
КПД
-
10
Номинальная частота
-
11
Скорость мотора/редуктора
-
12
Номинальное напряжение
-
13
Номинальная мощность
-
14
Номинальный ток
-
15
cos φ
-
16
КПД
-
17
Класс изоляции
-
18
Коэффициент перегрузки
-
19
Передаточное число
-
20
Максимальный крутящий момент
-
21
Монтажная позиция
-
22
Объем масла
-
23
Масса
-
24
Количество фаз/стандарт
-
25
Степень защиты
-
26
Наверх
Наверх
Мотор-редукторы из модульной системы SEW-EURODRIVE
Как и сферы применения наших мотор-редукторов, столь же разнообразны и широки возможности их комбинирования. Благодаря разработанной в SEW-EURODRIVE универсальной модульной системе наши клиенты могут использовать миллионы вариантов и найти индивидуальное техническое решение для любых задач. При этом цель модульной системы – суметь из минимального числа компонентов составить максимальное многообразие конечных продуктов.
Благодаря разработанной в SEW-EURODRIVE универсальной модульной системе наши клиенты могут использовать миллионы вариантов и найти индивидуальное техническое решение для любых задач. При этом цель модульной системы – суметь из минимального числа компонентов составить максимальное многообразие конечных продуктов.
Мотор-редукторы SEW-EURODRIVE делятся на следующие категории: стандартные мотор-редукторы, мотор-редукторы для сервопривода, мотор-редукторы для троллейного привода, мотор-редукторы с вариатором, мотор-редукторы из нержавеющей стали и взрывозащищенные мотор-редукторы.
Стандартные мотор-редукторы:
Стандартные мотор-редукторыСтандартные мотор-редукторы отличаются разнообразием конструкций, оптимальной градацией множества типоразмеров и самыми разными исполнениями. Это делает их незаменимыми и надежными приводами, особенно в сфере производства и логистики. В зависимости от количества типоразмеров редукторов возможны вращающие моменты до 50 000 Нм.
Мотор-редукторы для сервопривода:
Сила, динамика и точность. Это основные особенности мотор-редукторов для сервопривода. Наша модульная система и в этом случае является ключом к широким возможностям комбинирования и позволяет реализовать в этом сегменте самые разнообразные конфигурации из редукторов и двигателей. Поскольку для любой задачи можно подобрать идеальный вариант мотор-редуктора.
Какой бы ни была конфигурация сервопривода из наших планетарных редукторов PF.. или цилиндрических редукторов BF.. в сочетании с синхронными серводвигателями CMP, асинхронными серводвигателями типа DRL.. или с асинхронными двигателями DR..: Всякий раз специальная согласованность двигателя и редуктора дает вам именно те характеристики привода, которые идеально подходят к вашей системе и ее задачам.
Наши редукторы стандартной категории тоже позволяют вам создавать разнообразные комбинации с нашими серводвигателями, чтобы вполне индивидуально компоновать и оптимизировать свою приводную систему.
Мотор-редукторы с вариатором:
Для таких систем, где частота вращения привода должна регулироваться плавно, применяются наши механические мотор-редукторы с вариатором. Такие требования характерны, например, для простых ленточных конвейеров или мешалок, скорость которых должна постоянно адаптироваться к различным производственным процессам. При этом скорость регулируется бесступенчато с помощью либо маховичка, либо устройства дистанционного регулирования.
Мотор-редукторы из нержавеющей стали:
Если привод применяется в гигиенических зонах с высокими требованиями к чистоте, мотор-редуктор должен выдерживать воздействие химикатов и влаги. Для этих целей разработаны наши мотор-редукторы из нержавеющей стали, устойчивые к воздействию кислот и щелочей. Кроме того, их оптимизированная для очистки поверхность и отсутствие крыльчатки на дают грязи скапливаться в углублениях. Что же касается мощности, то никаких компромиссов от вас не потребуется. Будь то цилиндрический мотор-редуктор из нержавеющей стали RES.. или конический мотор-редуктор из нержавеющей стали KES..: Эти мотор-редукторы особенно прочны, долговечны и просты в обслуживании, а с коническим редуктором еще и очень компактны.
Будь то цилиндрический мотор-редуктор из нержавеющей стали RES.. или конический мотор-редуктор из нержавеющей стали KES..: Эти мотор-редукторы особенно прочны, долговечны и просты в обслуживании, а с коническим редуктором еще и очень компактны.
Взрывозащищенные мотор-редукторы:
Большинство наших стандартных и сервоприводных мотор-редукторов при соблюдении местных нормативов доступны по всему миру как взрывозащищенные мотор-редукторы. Это мощные и безопасные приводы, которые обеспечивают вам необходимую высокую производительность даже во взрывоопасных средах с воздушно-газовыми или воздушно-пылевыми смесями.
Наверх
Motor Cortex (раздел 3, глава 3) Neuroscience Online: электронный учебник по нейронаукам | Кафедра нейробиологии и анатомии
3.1 Введение
В предыдущих главах обсуждались нижние уровни моторной иерархии (спинной мозг и ствол головного мозга), которые участвуют в низкоуровневой обработке по принципу «гайки и болты», контролирующей деятельность отдельные мышцы. Отдельные альфа-мотонейроны контролируют силу, действующую на конкретную мышцу, а спинномозговые цепи могут контролировать сложное и сложное поведение, такое как ходьба и рефлекторные действия. Однако типы движений, контролируемые этими цепями, не инициируются сознательно. Произвольные движения требуют участия третьего и четвертого уровней иерархии: моторной коры и ассоциативной коры. Эти области коры головного мозга планируют произвольные действия, координируют последовательность движений, принимают решения о надлежащих поведенческих стратегиях и выборе, оценивают уместность конкретного действия с учетом текущего поведенческого или экологического контекста и передают команды соответствующим группам нижних двигательных нейронов. для выполнения желаемых действий.
Отдельные альфа-мотонейроны контролируют силу, действующую на конкретную мышцу, а спинномозговые цепи могут контролировать сложное и сложное поведение, такое как ходьба и рефлекторные действия. Однако типы движений, контролируемые этими цепями, не инициируются сознательно. Произвольные движения требуют участия третьего и четвертого уровней иерархии: моторной коры и ассоциативной коры. Эти области коры головного мозга планируют произвольные действия, координируют последовательность движений, принимают решения о надлежащих поведенческих стратегиях и выборе, оценивают уместность конкретного действия с учетом текущего поведенческого или экологического контекста и передают команды соответствующим группам нижних двигательных нейронов. для выполнения желаемых действий.
3.2 Моторная кора Включает первичную моторную кору, премоторную кору и дополнительную моторную область
| Рисунок 3.1 |
 Первичная моторная кора расположена непосредственно перед центральной бороздой.
Первичная моторная кора расположена непосредственно перед центральной бороздой. моторная кора включает три различные области лобной доли, расположенные непосредственно перед центральной бороздой . Этими областями являются первичная моторная кора (зона Бродмана 4), премоторная кора и дополнительная моторная зона (рис. 3.1). Электрическая стимуляция этих областей вызывает движения определенных частей тела. Первичная моторная кора, или M1 , расположена на прецентральной извилине и на передней парацентральной дольке на медиальной поверхности головного мозга. Из трех областей моторной коры для стимуляции первичной моторной коры требуется наименьшее количество электрического тока, чтобы вызвать движение. Низкие уровни кратковременной стимуляции обычно вызывают простые движения отдельных частей тела. Стимуляция премоторной коры или дополнительной моторной области требует более высоких уровней тока для вызова движений и часто приводит к более сложным движениям, чем стимуляция первичной моторной коры. Стимуляция в течение более длительных периодов времени (500 мс) у обезьян приводит к движению определенной части тела в стереотипную позу или положение, независимо от исходной точки этой части тела (рис. 3.2). Таким образом, премоторная кора и дополнительные двигательные области представляют собой области более высокого уровня, которые кодируют сложные паттерны двигательной активности и выбирают соответствующие двигательные планы для достижения желаемых конечных результатов.
Стимуляция премоторной коры или дополнительной моторной области требует более высоких уровней тока для вызова движений и часто приводит к более сложным движениям, чем стимуляция первичной моторной коры. Стимуляция в течение более длительных периодов времени (500 мс) у обезьян приводит к движению определенной части тела в стереотипную позу или положение, независимо от исходной точки этой части тела (рис. 3.2). Таким образом, премоторная кора и дополнительные двигательные области представляют собой области более высокого уровня, которые кодируют сложные паттерны двигательной активности и выбирают соответствующие двигательные планы для достижения желаемых конечных результатов.
| Рис. 3.2 |
 Стимуляция второго участка (нажмите СТИМУЛИРОВАТЬ 2) заставляет обезьяну подносить руку ко рту и открывать рот независимо от исходного положения руки, как если бы она подносила ко рту кусок пищи (Graziano et al. , 2002).
Стимуляция второго участка (нажмите СТИМУЛИРОВАТЬ 2) заставляет обезьяну подносить руку ко рту и открывать рот независимо от исходного положения руки, как если бы она подносила ко рту кусок пищи (Graziano et al. , 2002). Как и соматосенсорная кора постцентральной извилины, первичная моторная кора имеет соматотопическую организацию (рис. 3.3). Стимуляция передней парацентральной дольки вызывает движения контралатеральной ножки. По мере того как стимулирующий электрод перемещается по прецентральной извилине от дорсомедиального к вентролатеральному, движения вызываются постепенно от туловища, руки, кисти и лица (наиболее латерально). Репрезентации частей тела, совершающих точные, деликатные движения, таких как руки и лицо, непропорционально велики по сравнению с репрезентациями частей тела, совершающих лишь грубые, грубые движения, таких как туловище или ноги. Премоторная кора и дополнительная моторная область также содержат соматотопические карты.
| Рисунок 3. |
 3
3 Можно предположить, что моторная кора « гомункул » возникает из-за того, что нейроны, контролирующие отдельные мышцы, сгруппированы в коре. То есть все нейроны, контролирующие двуглавую мышцу, могут располагаться вместе, и все нейроны, контролирующие трехглавую мышцу, могут быть сгруппированы рядом, а нейроны, контролирующие камбаловидную мышцу, могут быть сгруппированы в отдаленной области. Однако электрофизиологические записи показали, что это не так. Движения отдельных мышц коррелируют с активностью распространенных отделов первичной моторной коры. Точно так же стимуляция небольших областей первичной моторной коры вызывает движения, требующие активности многочисленных мышц. Таким образом, первичная моторная кора гомункула не представляет активность отдельных мышц. Скорее, оно, по-видимому, представляет собой движения отдельных частей тела, которые часто требуют скоординированной деятельности больших групп мышц по всему телу.
3.3 Корковые афференты и эфференты
Моторная кора оказывает влияние на мышцы различными нисходящими путями (рис. 3.4). На некоторые нисходящие пути, рассмотренные в предыдущей главе, может влиять продукция моторной коры. Таким образом, в дополнение к прямой корковой иннервации альфа-мотонейронов через корково-спинномозговой путь следующие корковые эфферентные пути влияют на оставшиеся нисходящие пути:
- корково-рубральный путь позволяет коре модулировать руброспинальный путь0016
- корково-спинномозговой путь позволяет коре головного мозга модулировать покрышечно-спинномозговой путь
- кортико-ретикулярный тракт позволяет коре модулировать ретикулоспинальные тракты
| Рис. 3.4 |

Кора также может влиять на обработку боковых петель моторной иерархии. Корково-стриарный тракт иннервирует хвостатое ядро и скорлупу базальных ганглиев. Корково-мостовой и корково-оливарный пути иннервируют важные входы в мозжечок. Наконец, области коры могут влиять на другие области коры непосредственно через корково-кортикальные пути и косвенно через кортико-таламические пути (рис. 3.5). Большинство этих путей являются двунаправленными. Таким образом, моторная кора получает информацию от других областей коры прямо или косвенно через таламус, а также от мозжечка и базальных ганглиев, всегда через таламус.
| Рисунок 3.5 |
 Наведите курсор на каждое поле справа, чтобы выделить входы (синий) и выходы (красный) каждого региона.
Наведите курсор на каждое поле справа, чтобы выделить входы (синий) и выходы (красный) каждого региона. 3.4 Цитоархитектура моторной коры
Как и все части неокортекса, первичная моторная кора состоит из шести слоев (рис. 3.6). В отличие от первичных сенсорных областей, первичная моторная кора представляет собой агранулярную кору; то есть он не имеет упакованного клетками зернистого слоя (слой 4). Вместо этого наиболее характерным слоем первичной моторной коры является ее нисходящий выходной слой (слой 5), который содержит гигантские клетки Беца. Эти пирамидные клетки и другие проекционные нейроны первичной моторной коры составляют около 30% волокон корково-спинномозгового тракта. Остальные волокна исходят из премоторной коры и дополнительной моторной области (~30%), соматосенсорной коры (~30%) и задней теменной коры (~10%).
| Рисунок 3. |
 6
6 3.5 Кодирование движения моторной корой
Первичная моторная кора
Как обсуждалось выше, первичная моторная кора обычно не контролирует отдельные мышцы напрямую, а, скорее, контролирует отдельные движения или последовательности движений, которые требуют активности нескольких групп мышц. Альфа-мотонейроны спинного мозга, в свою очередь, кодируют силу сокращения групп мышечных волокон, используя код скорости и принцип размера. Таким образом, в соответствии с концепцией иерархической организации двигательной системы информация, представляемая моторной корой, представляет собой более высокий уровень абстракции, чем информация, представляемая спинальными мотонейронами.
Что кодируют нейроны первичной моторной коры? Подсказки пришли из записи активности этих нейронов, когда экспериментальные животные выполняют различные двигательные задачи. В общем, первичная моторная кора кодирует параметры, определяющие отдельные движения или простые последовательности движений.
- Нейроны первичной моторной коры возбуждаются за 5-100 мс до начала движения. Таким образом, вместо возбуждения в результате мышечной активности эти нейроны участвуют в передаче двигательных команд альфа-мотонейронам, которые в конечном итоге вызывают сокращение соответствующих мышц.
- Первичная моторная кора кодирует силу движения. Сила, необходимая для подъема руки из одного места в другое, намного больше, если человек держит шар для боулинга, чем если он держит воздушный шар. Многие нейроны первичной моторной коры кодируют силу, необходимую для совершения такого движения (рис. 3.7). Обратите внимание на различие между силой движения и мышечной силой.
 В то время как меньшинство первичных нейронов моторной коры кодирует индивидуальную мышечную силу, большее число кодирует количество силы, необходимой для конкретного движения, независимо от того, какие отдельные мышцы используются. Альфа-мотонейроны, в свою очередь, переводят команды нейронов моторной коры и контролируют количество силы, генерируемой отдельными мышцами для выполнения этого движения, в соответствии с принципами кода скорости и принципа размера.
В то время как меньшинство первичных нейронов моторной коры кодирует индивидуальную мышечную силу, большее число кодирует количество силы, необходимой для конкретного движения, независимо от того, какие отдельные мышцы используются. Альфа-мотонейроны, в свою очередь, переводят команды нейронов моторной коры и контролируют количество силы, генерируемой отдельными мышцами для выполнения этого движения, в соответствии с принципами кода скорости и принципа размера. Рисунки 3.7A, 3.7B и 3.7C
Моторная кора кодирует силу, необходимую для совершения движения. (Эвартс 1968)Рисунок 3.7А. Когда нагрузка невелика, двигательный нейрон в первичной моторной коре, контролирующий разгибание запястья, срабатывает, когда запястье разгибается. Двигательный нейрон, контролирующий сгибание запястья, не изменяет свою низкую скорость активности. Обратите внимание, что двигательный нейрон разгибания начинает запускать спайки до начала движения.

текст идет сюдаРисунок 3.7Б. Когда на левый шкив помещается груз весом 5 фунтов, необходимо приложить большее усилие, чтобы сначала удерживать вес неподвижно, а затем поднять его. Расширенный двигательный нейрон в первичной моторной коре возбуждается сильнее, чтобы произвести большую силу.
текст идет сюдаРисунок 3.7C. Когда на правый шкив помещается груз весом 5 фунтов, нагрузка приходится на сгибатели. Таким образом, первичные нейроны моторной коры для сгибания активируются, чтобы поддерживать стабильный вес. Когда запястье разгибается, нейроны успокаиваются, так как сила движения создается самим весом. (Обратите внимание, что моторная кора кодирует силу движения, такого как разгибание запястья или более сложные многосуставные движения.
 Сила отдельных мышц кодируется альфа-мотонейронами в спинном мозге и стволе головного мозга.)
Сила отдельных мышц кодируется альфа-мотонейронами в спинном мозге и стволе головного мозга.)
текст идет сюда - Первичная моторная кора кодирует направление движения. Многие нейроны первичной моторной коры избирательны в отношении определенного направления движения. Например, одна клетка может сильно возбуждаться при движении руки влево, тогда как при движении руки вправо она будет подавляться (рис. 3.8).
Рисунок 3.8
Направленная настройка нейронов моторной коры. Ячейка срабатывает максимально при движении руки в направлениях 135º или 180º, умеренно при движении руки в направлении 9.0º и 225º и молчит, когда рука движется в противоположных направлениях (0º, 45º, 270º и 315º) (Georgopoulos et al., 1982).
- Первичная моторная кора кодирует степень движения. Возбуждение некоторых нейронов коррелирует с расстоянием движения.
 Обезьяну обучали перемещать руку в разные целевые точки, которые различались по направлению и расстоянию от центра. Возбуждение многих нейронов коррелировало с направлением движения (как в пункте 3), тогда как возбуждение других нейронов коррелировало с расстоянием движения. Интересно, что некоторые нейроны коррелировали с взаимодействием определенного расстояния и направления; то есть они были связаны с определенной целевой позицией.
Обезьяну обучали перемещать руку в разные целевые точки, которые различались по направлению и расстоянию от центра. Возбуждение многих нейронов коррелировало с направлением движения (как в пункте 3), тогда как возбуждение других нейронов коррелировало с расстоянием движения. Интересно, что некоторые нейроны коррелировали с взаимодействием определенного расстояния и направления; то есть они были связаны с определенной целевой позицией. - Нейроны первичной моторной коры кодируют скорость движения. Почти все целенаправленные движения следуют типичной колоколообразной кривой зависимости скорости от расстояния (рис. 3.9). Например, когда рука перемещает предмет, такой как кофейная чашка, из одного места в другое (цель), рука ускоряется в течение первой половины движения, достигает максимальной скорости примерно на полпути к цели, а затем замедляется, пока не достигнет цели. достигает цели. Скорость возбуждения некоторых первичных нейронов моторной коры у обезьян коррелирует с этим колоколообразным профилем скорости, демонстрируя, что информация о скорости движения содержится в последовательности импульсов этих нейронов.

| Рисунок 3.9 |
Премоторная кора
Премоторная кора посылает аксоны в первичную моторную кору, а также непосредственно в спинной мозг. Он выполняет более сложную обработку, связанную с задачей, чем первичная моторная кора. Стимуляция премоторных областей у обезьяны при высоком уровне тока вызывает более сложные позы, чем стимуляция первичной моторной коры. Премоторная кора, по-видимому, участвует в выборе подходящих моторных планов для произвольных движений, тогда как первичная моторная кора участвует в выполнении этих произвольных движений.
- Нейроны премоторной коры сигнализируют о подготовке к движению. Обезьян обучали совершать определенные движения в ответ на визуальный сигнал с переменной задержкой между началом сигнала и началом движения (рис. 3.10). Записи премоторной коры показали, что многие нейроны избирательно возбуждаются в интервале задержки, за много секунд до начала движения.
 Конкретный нейрон сработает, когда обезьяна готовится сделать движение, например, влево, но будет молчать, когда обезьяна готовится сделать движение вправо. Таким образом, возбуждение этого типа нейронов не вызывает само движение, а, по-видимому, участвует в подготовке обезьяны к правильному движению при подаче сигнала «Вперед». Этот тип нейрона называется 9.0023 двигательный нейрон , поскольку он срабатывает, когда обезьяна готовится или готовится совершить движение.
Конкретный нейрон сработает, когда обезьяна готовится сделать движение, например, влево, но будет молчать, когда обезьяна готовится сделать движение вправо. Таким образом, возбуждение этого типа нейронов не вызывает само движение, а, по-видимому, участвует в подготовке обезьяны к правильному движению при подаче сигнала «Вперед». Этот тип нейрона называется 9.0023 двигательный нейрон , поскольку он срабатывает, когда обезьяна готовится или готовится совершить движение. Рисунок 3.10
Нейроны премоторной коры сигнализируют о подготовке к движению. Обезьяну учат готовиться к движению вправо или влево в зависимости от команды-сигнала, но задерживать движение до тех пор, пока не будет дан сигнал «Движение» (Weinrich and Wise, 1982). Некоторые нейроны срабатывают выборочно, когда животное готовится сделать движение вправо (ячейка Play Prepare right). Другие нейроны будут активироваться выборочно, когда животное готовится сделать движение влево (играть в «Подготовить левую ячейку»). Обратите внимание, что ячейки срабатывают в промежутке между инструкциями Prepare и Move, но не срабатывают во время самого движения.
Обратите внимание, что ячейки срабатывают в промежутке между инструкциями Prepare и Move, но не срабатывают во время самого движения.
- Нейроны премоторной коры сигнализируют о различных сенсорных аспектах, связанных с определенными двигательными актами. Некоторые премоторные нейроны возбуждаются, когда животное выполняет определенное действие, например, ломает арахис (рис. 3.11). Интересно, что один и тот же нейрон активируется избирательно, когда животное видит, как другая обезьяна или человек ломают арахис. Он также выборочно срабатывает на звук разбивающейся скорлупы арахиса, даже без какой-либо зрительной или двигательной активности. Эти нейроны называются «зеркальные» нейроны , потому что они реагируют не только на конкретное действие обезьяны, но и на вид (или звук) другой особи, выполняющей то же действие. (Интересное видео PBS о зеркальных нейронах см. на http://www.pbs.org/wgbh/nova/sciencenow/3204/01.
 html.)
html.) Рис. 3.11
Зеркальный нейрон в премоторной коре возбуждается в ответ на действие обезьяны, а также на восприятие обезьяной человека, выполняющего то же действие (Kohler et al., 2002).
- Премоторная кора чувствительна к поведенческому контексту конкретного движения. Премоторная кора головного мозга людей была визуализирована с помощью функциональной МРТ, когда они наблюдали видеозапись руки, сжимающей чашку (рис. 3.12). В одном случае чаша была полной и окружена полными тарелками с едой; подразумевалось, что человек схватил чашку, чтобы выпить. В другом состоянии чашка была пуста и окружена грязной посудой; подразумевалось, что человек схватил чашку, чтобы убрать со стола. В этом эксперименте премоторная кора была более активной, когда испытуемые смотрели первое видео, чем второе, хотя движения были одинаковыми. Таким образом, нейроны премоторной коры чувствительны к предполагаемым намерениям движения, а не только к самому движению, как следует из поведенческого контекста, в котором произошло движение.

Рисунок 3.12
Активность премоторной коры различает одно и то же движение на основе поведенческого контекста движения (Iacoboni et al., 2005). Когда испытуемый видел, как рука движется, чтобы взять чашку, чтобы выпить (ИГРА вверху), активность премоторной коры была выше, чем когда испытуемый видел руку, двигающуюся, чтобы взять чашку, чтобы убрать со стола после еды (ИГРА внизу). . Обратите внимание, что сила активности в коре (обозначаемая яркостью активированной области коры) больше в верхней, чем в нижней анимации.
- Премоторная кора сигнализирует о правильных и неправильных действиях. Людей испытуемых изучали в эксперименте с фМРТ, поскольку они наблюдали видеоклипы различных правильных и неправильных двигательных актов. Правильным действием считалось то, в котором движение и связанный с ним объект были выполнены правильно, например, установка времени на часах.
 Объектной ошибкой считалась ошибка, при которой действие было правильным, но объект был неправильным, например, полировка коричневой обуви черным кремом для обуви. Ошибка движения — это ошибка, при которой объект был правильным, но движение было неправильным, например, попытка положить монету в копилку, когда монета была ориентирована перпендикулярно прорези для монет. В этом эксперименте премоторная кора активировалась билатерально при пробах правильных действий и пробах ошибок движений; при пробах на предметную ошибку преимущественно активировалась только премоторная кора левого полушария.
Объектной ошибкой считалась ошибка, при которой действие было правильным, но объект был неправильным, например, полировка коричневой обуви черным кремом для обуви. Ошибка движения — это ошибка, при которой объект был правильным, но движение было неправильным, например, попытка положить монету в копилку, когда монета была ориентирована перпендикулярно прорези для монет. В этом эксперименте премоторная кора активировалась билатерально при пробах правильных действий и пробах ошибок движений; при пробах на предметную ошибку преимущественно активировалась только премоторная кора левого полушария.
Дополнительная моторная зона
Дополнительная моторная зона (SMA) участвует в программировании сложных последовательностей движений и координации двусторонних движений. В то время как премоторная кора, по-видимому, участвует в выборе моторных программ на основе визуальных стимулов или абстрактных ассоциаций, дополнительная моторная область, по-видимому, участвует в выборе движений на основе запомненной последовательности движений.
- СМА реагирует на последовательность движений и на мысленное повторение последовательности движений (рис. 3.13). Активность мозга измерялась с помощью ПЭТ-сканера, в то время как испытуемые выполняли простые и сложные последовательности движений. Когда движения были простыми, такими как повторяющееся движение одной цифры, первичная моторная кора и первичная соматосенсорная кора активировались в контралатеральном полушарии. Когда испытуемого просили выполнить сложную последовательность движений пальцев, SMA активировалась билатерально, в дополнение к активации контралатеральной первичной моторной и соматосенсорной коры. Наконец, когда испытуемого попросили оставаться неподвижным, но мысленно репетировать сложную последовательность действий, SMA все еще был активен, хотя первичные моторные и соматосенсорные области коры молчали. Таким образом, SMA, по-видимому, участвует в билатеральных движениях и в мысленной репетиции этих движений.
Рисунок 3.
 13
13
Исследование позитронно-эмиссионной томографии (ПЭТ) простых и сложных движений пальцев (Roland et al., 1980). SMA активируется двусторонне, когда субъекты выполняют сложные движения, и даже когда они только воображают, что выполняют движения.
- SMA участвует в преобразовании кинематической информации в динамическую. Движения могут быть определены с точки зрения динамика (величина силы, необходимая для совершения движения) и кинематика (расстояние и углы, определяющие конкретное движение в пространстве). Многие планы движений представлены в кинематических терминах (например, Переместите руку влево). Однако двигательная система должна в конечном итоге преобразовать это в представление, основанное на динамике, чтобы проинструктировать соответствующие мышцы сокращаться с соответствующей силой. Записи на обезьянах показали, что во время подготовительной задержки перед тем, как обезьяна совершает заданное движение, некоторые нейроны SMA меняют свои активирующие корреляты с кинематического представления на представление, основанное на динамике, что позволяет предположить, что SMA играет жизненно важную роль в этой трансформации.

Ассоциативная кора
Четвертым уровнем моторной иерархии является ассоциативная кора , в частности префронтальная кора и задняя теменная кора (рис. 3.14). Эти области мозга не являются двигательными областями в строгом смысле слова. Их активность не коррелирует точно с отдельными двигательными актами, а стимуляция этих областей не приводит к двигательной активности. Однако эти области необходимы для обеспечения того, чтобы движения были адаптивными к потребностям организма и соответствовали поведенческому контексту.
| Рисунок 3.14 |
- Задняя теменная кора участвует в обеспечении точного направления движений на объекты во внешнем пространстве. Эта область участвует в обработке пространственных отношений объектов в мире и в построении представления внешнего пространства, которое не зависит от положения глаз или тела наблюдателя.
 Такие представления обеспечивают стабильное восприятие мира, не зависящее от ориентации зрителя, а также представление желаемых траекторий в пространстве, не зависящих от положения тела. Повреждение задней теменной коры может привести к ряду апраксии , то есть невозможность совершать сложные координированные движения. Например, пациент с конструктивной апраксией не может воспроизвести конфигурацию набора блоков в правильной последовательности, даже если пациент может ловко маневрировать каждым блоком по отдельности.
Такие представления обеспечивают стабильное восприятие мира, не зависящее от ориентации зрителя, а также представление желаемых траекторий в пространстве, не зависящих от положения тела. Повреждение задней теменной коры может привести к ряду апраксии , то есть невозможность совершать сложные координированные движения. Например, пациент с конструктивной апраксией не может воспроизвести конфигурацию набора блоков в правильной последовательности, даже если пациент может ловко маневрировать каждым блоком по отдельности.
- Префронтальная кора участвует в выборе подходящих действий для определенного поведенческого контекста. Он также участвует в оценке последствий определенного образа действий. У пациентов с повреждением префронтальной коры возникают проблемы с исполнительной обработкой. Они принимают неадекватные поведенческие решения и часто не могут предвидеть вероятные последствия своих действий. Они проявляют импульсивное поведение, часто демонстрируя неспособность отсрочить мгновенное удовлетворение ради более крупного вознаграждения в долгосрочной перспективе.

Проверьте свои знания
- Вопрос 1
- А
- Б
- С
- Д
- Е
Клетки Беца наиболее многочисленны в слое…
А. IV соматосенсорной коры.
B. V соматосенсорной коры.
C. IV моторной коры.
Д. В моторной коры.
E. III моторной коры.
Клетки Беца наиболее многочисленны в слое…
А. IV соматосенсорной коры. Этот ответ НЕВЕРНЫЙ.
Клетки Беца не находятся в соматосенсорной коре.
B. V соматосенсорной коры.
C. IV моторной коры.
Д. В моторной коры.
E. III моторной коры.
Клетки Беца наиболее многочисленны в слое. ..
..
А. IV соматосенсорной коры.
B. V соматосенсорной коры. Этот ответ НЕВЕРНЫЙ.
Клетки Беца не находятся в соматосенсорной коре.
C. IV моторной коры.
Д. В моторной коры.
E. III моторной коры.
Клетки Беца наиболее многочисленны в слое…
А. IV соматосенсорной коры.
B. V соматосенсорной коры.
C. IV моторной коры. Этот ответ НЕВЕРНЫЙ.
Клетки Беца не находятся в слое IV.
Д. В моторной коры.
E. III моторной коры.
Клетки Беца наиболее многочисленны в слое…
А. IV соматосенсорной коры.
B. V соматосенсорной коры.
C. IV моторной коры.
Д. В моторной коры. Этот ответ ПРАВИЛЬНЫЙ!
E.
III моторной коры.
Клетки Беца наиболее многочисленны в слое…
8]А. IV соматосенсорной коры.
B. V соматосенсорной коры.
C. IV моторной коры.
Д. В моторной коры.
E. III моторной коры. Этот ответ НЕВЕРНЫЙ.
Клетки Беца не находятся в слое III.
вопросА Б С Д Е Корково-спинномозговой нейрон в первичной моторной коре может выполнять все следующие действия, КРОМЕ:
A. Проекция на множественные пулы двигательных нейронов в спинном мозге.
B. Принять участие в инициации движения.
C. Код силы отдельных мышц.
D. Код направления движения.
E.
Код степени перемещения.
Корково-спинномозговой нейрон в первичной моторной коре может выполнять все следующие действия, КРОМЕ:
A. Проекция на множественные пулы двигательных нейронов в спинном мозге. Этот ответ НЕВЕРНЫЙ.
Это ИСТИННОЕ утверждение. Многие различные группы мышц находятся под влиянием активности отдельных нейронов в моторной коре.
B. Принять участие в инициации движения.
C. Код силы отдельных мышц.
D. Код направления движения.
E. Код степени перемещения.
Корково-спинномозговой нейрон в первичной моторной коре может выполнять все следующие действия, КРОМЕ:
A. Проекция на множественные пулы двигательных нейронов в спинном мозге.
B. Принять участие в инициации движения. Этот ответ НЕВЕРНЫЙ.
Это ИСТИННОЕ утверждение.

C. Код силы отдельных мышц.
D. Код направления движения.
E. Код степени перемещения.
Корково-спинномозговой нейрон в первичной моторной коре может выполнять все следующие действия, КРОМЕ:
A. Проекция на множественные пулы двигательных нейронов в спинном мозге.
B. Принять участие в инициации движения.
C. Код силы отдельных мышц. Этот ответ ПРАВИЛЬНЫЙ!
Это ЛОЖНОЕ утверждение. Нейроны моторной коры кодируют силу отдельных движений, а не отдельных мышц. Нижние двигательные нейроны (альфа-мотонейроны) кодируют силу отдельных мышц.
D. Код направления движения.
E. Код степени перемещения.
Корково-спинномозговой нейрон в первичной моторной коре может выполнять все следующие действия, КРОМЕ:
A.
Проекция на множественные пулы двигательных нейронов в спинном мозге.
B. Принять участие в инициации движения.
C. Код силы отдельных мышц.
D. Код направления движения. Этот ответ НЕВЕРНЫЙ.
Это ИСТИННОЕ утверждение.
E. Код степени перемещения.
Корково-спинномозговой нейрон в первичной моторной коре может выполнять все следующие действия, КРОМЕ:
A. Проекция на множественные пулы двигательных нейронов в спинном мозге.
B. Принять участие в инициации движения.
C. Код силы отдельных мышц.
D. Код направления движения.
E. Код степени перемещения. Этот ответ НЕВЕРНЫЙ.
Это ИСТИННОЕ утверждение.
Физиология, моторная кортикальная — StatePears
Derek W.
Yip; Форшинг Луи.
Информация об авторе
Последнее обновление: 20 сентября 2021 г.
Введение
Основная функция моторной коры — генерировать сигналы, управляющие движением тела. Это часть лобной доли, расположенная впереди центральной борозды. Он состоит из первичной моторной коры, премоторной коры и дополнительной моторной области. Не все участки моторной коры имеют зернистый клеточный слой. Первичная моторная кора, расположенная в области Бродмана 4, посылает большую часть электрических импульсов, исходящих из моторной коры. Эти волокна напрямую синапсируют с двигательными нейронами спинного мозга. Стоит отметить, что корково-спинномозговые волокна отходят как от лобной, так и от теменной коры. Перед первичной моторной корой премоторная кора расположена непосредственно перед первичной моторной корой в области Бродмана 6. Ее функция заключается в подготовке к движению, особенно проксимальной мускулатуры. Дополнительная моторная зона находится на медиальной поверхности продольной щели, непосредственно перед представительством ног в первичной моторной коре.
Предлагаемые функции включают стабилизацию положения тела и координацию, хотя и не до конца поняты.[2]
Существуют важные отличия корковых проекций от корковых двигательных нейронов. Волокна, исходящие из первичной моторной коры (область 4), оканчиваются в основном в спинном мозге, образуя синапсы непосредственно на двигательных нейронах. Ростральные лобные двигательные области не заканчиваются непосредственно в спинном мозге. Они посылают волокна, которые заканчиваются в различных областях ствола мозга. Поэтому они контролируют движения косвенно через ствол мозга, например, через покрышечно-спинномозговой и ретикулоспинальный тракты. Также более понятно, что они будут больше контролировать проксимальные и осевые мышцы и движения.
Корковые афференты к лобной моторной коре возникают из трех источников: теменной соматосенсорной коры, префронтальной коры и поясной коры. Теменно-лобные связи представляют собой различные сенсомоторные цепи. Поясная кора посылает волокна в премоторные и дополнительные моторные области для лимбической информации, мотивации и принятия решений.
Наконец, префронтальная кора посылает волокна в моторную кору, выполняющую важные функции планирования и выполнения движений.
Вопросы, вызывающие озабоченность
Слабость и паралич, возникающие в результате дисфункции моторной коры, могут значительно снизить качество жизни пациентов. К счастью, некоторые факторы риска инсульта, такие как гипертония, курение, диабет, плохое питание, высокий уровень холестерина и отсутствие физической активности, можно смягчить. Крайне важно, чтобы медицинские работники консультировали пациентов по поводу этих факторов, чтобы предотвратить серьезные неврологические нарушения.[3]
Сотовый
На клеточном уровне нейрон отвечает за связь в моторной коре. Нейрон состоит из трех основных частей: дендритов, сомы и аксона — других нейронов, которые получают сигналы от дендритов. Затем дендрит передает этот сигнал телу клетки или соме. Сома содержит ядро и аксонный холмик. Ядро отвечает за создание всех белков и мембран, поддерживающих жизнь нейрона.
Холм аксона — это место, где протенциональное действие передается на аксон. Затем аксон доставляет потенциал действия к окончаниям аксона, которые образуют синапс с дендритами других нейронов. В моторной коре есть сомы, которые генерируют потенциалы действия вплоть до спинного мозга; это называется верхним моторным нейроном. Верхний мотонейрон связывается с нижним мотонейроном через глутамат.[4] Моторная кора имеет только рудиментарный слой IV, который функционирует для приема таламокортикальных проекций. Поэтому эти области также называют агранулярной корой без входа от таламуса.
Развитие
После трех недель развития нервная система делится на три первичных мозговых пузырька: передний мозг (прозэнцефалон), средний мозг (мезэнцефалон) и задний мозг (ромбэнцефалон). Затем нервная система становится стадией с пятью пузырьками, при этом передний мозг разделяется на полушария головного мозга, таламус и гипоталамус. Вслед за формированием различных областей головного мозга происходит пролиферация нейронов и миграция этих нейронов из перивентрикулярной зоны в кору наизнанку к ее окончательной 6-слойной корковой цитоархитектуре.
Верхние моторные нейроны происходят из слоев 5 и 6 полушарий головного мозга, в частности, из первичной моторной коры.[5]
Задействованные системы органов
Скелетно-мышечная система полагается на двигательные пути, чтобы управлять ими. В нервно-мышечном соединении нижний мотонейрон синапсирует с мышечным волокном. После того, как потенциал действия достигает окончания аксона нижнего двигательного нейрона, потенциал-зависимые кальциевые каналы открываются для увеличения концентрации внутриклеточных ионов кальция, что позволяет пузырькам ацетилхолина сливаться с мембраной и высвобождаться в синаптическую щель, вызывая мышечные сокращения.
Функция
Основной функцией моторной коры является планирование и создание электрических импульсов, вызывающих произвольные мышечные сокращения. Нервы, отходящие от моторной коры, могут быть как черепными, так и спинномозговыми. Черепные нервы исходят непосредственно из ствола головного мозга, тогда как спинномозговые нервы выходят из спинного мозга.
Есть несколько черепных нервов (ЧН), которые выполняют двигательную функцию, в том числе ЧН III (глазодвигательный нерв), ЧН IV (блоковый нерв), ЧН V (тройничный нерв), ЧН VI (отводящий нерв), ЧН VII (лицевой нерв), CN IX (языкоглоточный нерв), CN X (блуждающий нерв), CN XI (добавочный нерв) и CN XII (подъязычный нерв). CN III, CN IV и CN VI участвуют в движении глаз. CN V иннервирует жевательные мышцы. CN VII обеспечивает движение лицевых мышц. CN IX иннервирует шилоглоточную мышцу, которая обеспечивает движение глотки и гортани. CN X имеет много функций, которые играют роль в речи и глотании. CN XI участвует в пожимании плечами и поворотах головы с иннервацией трапециевидной и грудино-ключично-сосцевидной мышц соответственно. CN XII иннервирует большинство мышц языка.[6]
Дополнительная область двигателя
Прежде всего, дополнительная моторная область проецируется на моторную кору. По-видимому, он участвует в основном в программировании последовательностей движений.
Любые поражения этой области у обезьян вызывают неловкость при выполнении сложных действий с бимануальной координацией.
Когда люди считают про себя, не говоря ни слова, моторная кора находится в состоянии покоя, но когда они считают число вслух, при этом увеличивается кровоток в моторной коре и дополнительной моторной области. Кроме того, дополнительная моторная область, как и моторная кора, участвует в произвольных движениях, когда выполняемые движения сложны и требуют планирования. Кровоток увеличивается независимо от того, выполняется запланированное движение или нет. Увеличение кровотока происходит независимо от того, выполняется ли движение контралатеральной или ипсилатеральной рукой.[7]
Механизм
Начиная с первичной моторной коры, верхний двигательный нейрон спускается и проходит через ипсилатеральную заднюю ножку внутренней капсулы. Большинство волокон перекрещиваются в пирамидном перекресте в каудальном отделе продолговатого мозга. Отсюда верхний мотонейрон находится на контралатеральной стороне своего происхождения.
Верхний мотонейрон образует синапсы с нижним мотонейроном в передних рогах спинного мозга на соответствующем уровне. Нижний мотонейрон выходит из спинного мозга и соединяется с мышцей [8].
Сопутствующее тестирование
Тестирование интактного двигательного кортикального пути может включать тестирование глубокого сухожильного рефлекса. Бицепс (С5, С6), плечелучевую мышцу (С6), трицепс (С7), надколенник (L4) и ахиллово сухожилие (S1) можно исследовать для оценки целостности нервных корешков. При растяжении мышечное веретено посылает сигнал по нервному волокну 1-альфа. Волокно входит в позвоночник через задний корешок и образует синапсы с нижним мотонейроном и интернейроном. Нижний двигательный нейрон вызывает сокращение мышцы-агониста, а интернейрон вызывает расслабление мышцы-антагониста.]
Патофизиология
Столбняк – это состояние, вызываемое экзотоксином тетаноспазмином из Clostridium tetani. Столбняк вызывает болезненные сокращения мышц, особенно челюсти и шеи.
Тетаноспазмин ингибирует высвобождение глицина и ГАМК, которые являются тормозными нейротрансмиттерами. Без торможения скелетных мышц происходит устойчивое сокращение.
Инсульт – это когда часть мозга не получает достаточного количества крови. В результате симптомы инсульта могут проявляться в виде дефицита двигательной функции, если части моторной коры поражены инфарктом. Однако это часто сопровождается другими симптомами, такими как дефицит речи или зрения. Возможен чисто моторный инсульт, который является симптомом лакунарного инсульта, вызывающего инфаркт задней ножки внутренней капсулы. Этот инфаркт может проявляться гемипарезом, то есть слабостью одной стороны тела.[10]
Клиническое значение
Повреждение моторной коры приводит к клиническому состоянию, называемому синдромом верхнего двигательного нейрона. Он также может возникать в результате повреждения аксонов, отходящих от моторной коры, т. е. корково-спинномозгового пути. Клинически наблюдается слабость без значительной атрофии мышц, спастичность (повышенный мышечный тонус с ригидностью складного ножа), гиперрефлексия и положительный симптом Бабинского.
Симптом Бабинского является нормальным рефлексом у младенцев, но ненормальным у взрослых. Это происходит, когда большой палец движется вверх после сильного удара по подошве.[11] Ригидность или спастичность ножа с зажимом обычно проявляется начальным сопротивлением движению, за которым следует быстрое снижение сопротивления. Поражения верхних двигательных нейронов проявляются характерной слабостью, которая также типична для меньшей слабости, поражающей антигравитационные мышцы. Эта картина порождает привычный вид после инсульта с поражением моторной коры. Пациент будет иметь согнутую позу в пораженной верхней конечности и вытянутую позу в нижней конечности. Пациент будет ходить типичной гемипаретической походкой. Поражения нижних двигательных нейронов возникают в результате повреждения двигательных нейронов спинного мозга и обычно проявляются слабостью, атрофией, фасцикуляциями, гипорефлексией, снижением тонуса и вялыми параличами.
Контрольные вопросы
Доступ к бесплатным вопросам с несколькими вариантами ответов по этой теме.

Комментарий к этой статье.
Рисунок
Моторная кора. Изображение предоставлено S Bhimji MD
Ссылки
- 1.
Meier JD, Aflalo TN, Kastner S, Graziano MS. Сложная организация первичной моторной коры человека: исследование фМРТ с высоким разрешением. J Нейрофизиол. 2008 г., октябрь; 100 (4): 1800-12. [Бесплатная статья PMC: PMC2576195] [PubMed: 18684903]
- 2.
He SQ, Dum RP, Strick PL. Топографическая организация корково-спинномозговых проекций от лобной доли: двигательные зоны на медиальной поверхности полушария. Дж. Нейроски. 1995 г., май; 15 (5, часть 1): 3284–306. [Бесплатная статья PMC: PMC6578253] [PubMed: 7538558]
- 3.
de Brouwer EJM, Kockelkoren R, De Vis JB, Dankbaar JW, Velthuis BK, Takx RA, De Jonghe A, Emmelot-Vonk MH, Koek Vonk MH HL, de Jong PA., Голландские исследователи острого инсульта (DUST). Распространенность и сосудистые факторы риска кальцификации базальных ганглиев у пациентов с риском цереброваскулярных заболеваний.
J Нейрорадиол. 2020 сен;47(5):337-342. [В паблике: 31034898]
- 4.
де Карвалью М. Патофизиологическое значение фасцикуляций в ранней диагностике БАС. Боковой амиотроф склеры Другое заболевание двигательных нейронов. 2000 март; 1 Приложение 1: S43-6. [PubMed: 11464925]
- 5.
Стайлз Дж., Джерниган ТЛ. Основы развития мозга. Neuropsychol Rev. 2010 Dec; 20(4):327-48. [Бесплатная статья PMC: PMC2989000] [PubMed: 21042938]
- 6.
Davis MC, Griessenauer CJ, Bosmia AN, Tubbs RS, Shoja MM. Именование черепных нервов: исторический обзор. Клин Анат. 2014 янв; 27(1):14-9. [PubMed: 24323823]
- 7.
Шибата С., Ямао Ю., Куниеда Т., Инано Р., Накаэ Т., Нисида С., Инада Т., Такахаши Ю., Кикучи Т., Аракава Ю., Ёсида К., Мацумото Р., Икеда А., Мима Т., Миямото С. Интраоперационное электрофизиологическое картирование медиальных лобных моторных зон и функциональные результаты. Мировой нейрохирург. 2020 июнь; 138: e389-e404.
[PubMed: 32145417]
- 8.
Лимонный РН. Нисходящие пути в моторной регуляции. Annu Rev Neurosci. 2008;31:195-218. [В паблике: 18558853]
- 9.
Hofstoetter US, Danner SM, Freundl B, Binder H, Mayr W, Rattay F, Minassian K. Периодическая модуляция повторяющихся моносинаптических рефлексов пояснично-крестцового отдела спинного мозга человека. J Нейрофизиол. 2015 июль; 114 (1): 400-10. [Бесплатная статья PMC: PMC4507962] [PubMed: 25
Хан А., Каснер С.Е., Линн М.Дж., Чимовиц М.И., Симптоматическое внутричерепное заболевание варфарин-аспирин (WASID) Исследователи. Факторы риска и исход у пациентов с симптоматическим внутричерепным стенозом, проявляющимся лакунарным инсультом. Инсульт. 2012 май; 43(5):1230-3. [Бесплатная статья PMC: PMC3336041] [PubMed: 22363054]
Ачарья А.Б., Джамиль Р.Т., Дьюи Дж.Дж. StatPearls [Интернет]. Издательство StatPearls; Остров сокровищ (Флорида): 1 августа 2021 г. Рефлекс Бабинского. [PubMed: 30085551]
Рефлекс Бабинского. [PubMed: 30085551]
Физиология, моторная кора — PubMed
Сохранить цитату в файл
Формат: Резюме (текст) PubMedPMIDAbstract (текст) CSV
Добавить в коллекции
- Создать новую коллекцию
- Добавить в существующую коллекцию
Назовите свою коллекцию:
Имя должно содержать менее 100 символов
Выберите коллекцию:
Не удалось загрузить вашу коллекцию из-за ошибки
Повторите попытку
Добавить в мою библиографию
- Моя библиография
Не удалось загрузить делегатов из-за ошибки
Повторите попытку
Ваш сохраненный поиск
Название сохраненного поиска:
Условия поиска:
Тестовые условия поиска
Эл. адрес:
(изменить)
адрес:
(изменить)
Который день? Первое воскресеньеПервый понедельникПервый вторникПервая средаПервый четвергПервая пятницаПервая субботаПервый деньПервый будний день
Который день? воскресеньепонедельниквторниксредачетвергпятницасуббота
Формат отчета: SummarySummary (text)AbstractAbstract (text)PubMed
Отправить максимум: 1 шт. 5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
Отправить, даже если нет новых результатов
Необязательный текст в электронном письме:
Создайте файл для внешнего программного обеспечения для управления цитированием
Полнотекстовые ссылки
StatPearls [Интернет]
Книжная полка NCBIПолнотекстовые ссылки
Книга
Дерек В. Йип
1
, Форшинг Луи
2
Йип
1
, Форшинг Луи
2
Источник: StatPearls [Интернет]. Остров сокровищ (Флорида): StatPearls Publishing; 2022 9 января0003
.
Принадлежности
Принадлежности
- 1 Университет Северного штата Калифорния
- 2 CA Northstate Uni, Медицинский колледж
- PMID: 311
- Идентификатор книжной полки: НБК542188
Бесплатные книги и документы
Книга
Дерек В. Йип и др.
Йип и др.
Бесплатные книги и документы
Источник: StatPearls [Интернет]. Остров сокровищ (Флорида): StatPearls Publishing; 2022 янв.
.
Авторы
Дерек В. Йип 1 , Форшинг Луи 2
Принадлежности
- 1 Университет Северного штата Калифорния
- 2 CA Northstate Uni, Медицинский колледж
- PMID: 311
- Идентификатор книжной полки: НБК542188
Выдержка
Основная функция моторной коры — генерировать сигналы, управляющие движением тела. Это часть лобной доли, расположенная впереди центральной борозды. Она состоит из первичной моторной коры, премоторной коры и дополнительной моторной области. Не все участки моторной коры имеют зернистый клеточный слой. Первичная моторная кора, расположенная в области Бродмана 4, посылает большую часть электрических импульсов, исходящих из моторной коры. Эти волокна напрямую синапсируют с двигательными нейронами спинного мозга. Стоит отметить, что корково-спинномозговые волокна отходят как от лобной, так и от теменной коры. Перед первичной моторной корой премоторная кора расположена непосредственно перед первичной моторной корой в области Бродмана 6. Ее функция заключается в подготовке к движению, особенно проксимальной мускулатуры. Дополнительная моторная зона находится на медиальной поверхности продольной щели, непосредственно перед представительством ног в первичной моторной коре. Хотя это не совсем понятно, предлагаемые функции включают стабилизацию положения тела и координацию.
Это часть лобной доли, расположенная впереди центральной борозды. Она состоит из первичной моторной коры, премоторной коры и дополнительной моторной области. Не все участки моторной коры имеют зернистый клеточный слой. Первичная моторная кора, расположенная в области Бродмана 4, посылает большую часть электрических импульсов, исходящих из моторной коры. Эти волокна напрямую синапсируют с двигательными нейронами спинного мозга. Стоит отметить, что корково-спинномозговые волокна отходят как от лобной, так и от теменной коры. Перед первичной моторной корой премоторная кора расположена непосредственно перед первичной моторной корой в области Бродмана 6. Ее функция заключается в подготовке к движению, особенно проксимальной мускулатуры. Дополнительная моторная зона находится на медиальной поверхности продольной щели, непосредственно перед представительством ног в первичной моторной коре. Хотя это не совсем понятно, предлагаемые функции включают стабилизацию положения тела и координацию.
Существуют важные отличия корковых проекций от корковых мотонейронов. Волокна, исходящие из первичной моторной коры (область 4), оканчиваются в основном в спинном мозге, образуя синапсы непосредственно на двигательных нейронах. Ростральные лобные двигательные области не заканчиваются непосредственно в спинном мозге. Они посылают волокна, которые заканчиваются в различных областях ствола мозга. Поэтому они контролируют движения косвенно через ствол мозга, например, через покрышечно-спинномозговой и ретикулоспинальный тракты. Также более понятно, что они будут больше контролировать проксимальные и осевые мышцы и движения.
Волокна, исходящие из первичной моторной коры (область 4), оканчиваются в основном в спинном мозге, образуя синапсы непосредственно на двигательных нейронах. Ростральные лобные двигательные области не заканчиваются непосредственно в спинном мозге. Они посылают волокна, которые заканчиваются в различных областях ствола мозга. Поэтому они контролируют движения косвенно через ствол мозга, например, через покрышечно-спинномозговой и ретикулоспинальный тракты. Также более понятно, что они будут больше контролировать проксимальные и осевые мышцы и движения.
Корковые афференты к лобной моторной коре возникают из трех источников: теменной соматосенсорной коры, префронтальной коры и поясной коры. Теменно-лобные связи представляют собой различные сенсомоторные цепи. Поясная кора посылает волокна в премоторные и дополнительные моторные области для лимбической информации, мотивации и принятия решений. Наконец, префронтальная кора посылает волокна в моторную кору, выполняющую важные функции планирования и выполнения движений.
Авторское право © 2022, StatPearls Publishing LLC.
Разделы
- Введение
- Проблемы, вызывающие озабоченность
- Сотовый
- Разработка
- Задействованные системы органов
- Функция
- Механизм
- Связанное тестирование
- Патофизиология
- Клиническое значение
- Обзорные вопросы
- использованная литература
Похожие статьи
-
Происхождение корково-спинномозговых проекций из премоторных областей лобной доли.

Дум РП, Стрик ПЛ. Дум РП и др. Дж. Нейроски. 1991 март; 11 (3): 667-89. doi: 10.1523/JNEUROSCI.11-03-00667.1991. Дж. Нейроски. 1991. PMID: 1705965 Бесплатная статья ЧВК.
-
Сравнение ипсилатеральных корковых проекций с дорсальными и вентральными подразделениями премоторной коры макаки.
Гош С., Гаттера Р. Гош С. и др. Соматосенс Мот Рез. 1995;12(3-4):359-78. дои: 10.3109/089
50 - 68. Соматосенс Мот Рез. 1995. PMID: 8834308
Топографическая организация корково-спинномозговых проекций от лобной доли: двигательные зоны на медиальной поверхности полушария.
He SQ, Dum RP, Strick PL. He SQ и др. Дж. Нейроски. 1995 г., май; 15 (5, часть 1): 3284–306. doi: 10.1523/JNEUROSCI.
 15-05-03284.1995.
Дж. Нейроски. 1995.
PMID: 7538558
Бесплатная статья ЧВК.
15-05-03284.1995.
Дж. Нейроски. 1995.
PMID: 7538558
Бесплатная статья ЧВК. Моторные зоны в лобной доле примата.
Дум РП, Стрик ПЛ. Дум РП и др. Физиол Поведение. 2002 декабрь; 77 (4-5): 677-82. doi: 10.1016/s0031-9384(02)00929-0. Физиол Поведение. 2002. PMID: 12527018 Обзор.
Моторные зоны на медиальной стенке полушария.
Strick PL, Dum RP, Picard N. Стрик П.Л. и др. Novartis обнаружила Symp. 1998;218:64-75; обсуждение 75-80, 104-8. doi: 10.1002/9780470515563.ch5. Novartis обнаружила Symp. 1998. PMID: 9949816 Обзор.
Посмотреть все похожие статьи
использованная литература
-
-
Мейер Д.
 Д., Афлало Т.Н., Кастнер С., Грациано М.С. Сложная организация первичной моторной коры человека: исследование фМРТ с высоким разрешением. J Нейрофизиол. 2008 г., октябрь; 100 (4): 1800-12.
—
ЧВК
—
пабмед
Д., Афлало Т.Н., Кастнер С., Грациано М.С. Сложная организация первичной моторной коры человека: исследование фМРТ с высоким разрешением. J Нейрофизиол. 2008 г., октябрь; 100 (4): 1800-12.
—
ЧВК
—
пабмед
-
Мейер Д.
-
- He SQ, Дум RP, Стрик PL. Топографическая организация корково-спинномозговых проекций от лобной доли: двигательные зоны на медиальной поверхности полушария. Дж. Нейроски. 1995 г., май; 15 (5, часть 1): 3284–306. — ЧВК — пабмед
-
-
de Brouwer EJM, Kockelkoren R, De Vis JB, Dankbaar JW, Velthuis BK, Takx RA, De Jonghe A, Emmelot-Vonk MH, Koek HL, de Jong PA, Голландские исследователи острого инсульта (DUST) Распространенность и сосудистые факторы риска кальцификаты базальных ганглиев у пациентов с риском цереброваскулярных заболеваний.
 J Нейрорадиол. 2020 сен;47(5):337-342.
—
пабмед
J Нейрорадиол. 2020 сен;47(5):337-342.
—
пабмед
-
de Brouwer EJM, Kockelkoren R, De Vis JB, Dankbaar JW, Velthuis BK, Takx RA, De Jonghe A, Emmelot-Vonk MH, Koek HL, de Jong PA, Голландские исследователи острого инсульта (DUST) Распространенность и сосудистые факторы риска кальцификаты базальных ганглиев у пациентов с риском цереброваскулярных заболеваний.
-
- де Карвалью М. Патофизиологическое значение фасцикуляций в ранней диагностике БАС. Боковой амиотроф склеры Другое заболевание двигательных нейронов. 2000 март; 1 Приложение 1: S43-6. — пабмед
-
- Стайлз Дж., Джерниган Т.Л. Основы развития мозга. Neuropsychol Rev. 2010 Dec; 20(4):327-48. — ЧВК — пабмед
Типы публикаций
Полнотекстовые ссылки
StatPearls [Интернет]
Книжная полка NCBIПроцитируйте
Формат: ААД АПА МДА НЛМ
Отправить по номеру
Сигналы моторной коры для каждой руки смешиваются между полушариями и нейронами, но разделяются в ответе популяции
Реферат
Моторная кора (M1) имеет латеральные выходы, однако нейроны могут быть активны во время движений любой руки.
 Какова природа и роль активности двух полушарий? Мы регистрировали мышцы и нейроны с двух сторон, в то время как обезьяны вращались каждой рукой. Большинство нейронов были активны во время движения любой руки. Ответы сильно зависели от руки, что дает две возможности. Во-первых, сигналы на уровне популяции могут различаться в зависимости от используемой руки. Во-вторых, одни и те же сигналы на уровне популяции могут присутствовать, но по-разному распределяться по нейронам. Данные подтвердили эту вторую гипотезу. Мышечная активность была точно предсказана по активности ипсилатерального или контралатерального полушария. В более общем плане нам не удалось найти сигналы, характерные только для контралатерального полушария. Но если сигналы распределяются между полушариями, как они избегают воздействия не на ту руку? Мы обнаружили, что активность, связанная с каждой рукой, занимает отдельное подпространство, что позволяет декодерам мышечной активности естественным образом игнорировать сигналы, относящиеся к другой руке.
Какова природа и роль активности двух полушарий? Мы регистрировали мышцы и нейроны с двух сторон, в то время как обезьяны вращались каждой рукой. Большинство нейронов были активны во время движения любой руки. Ответы сильно зависели от руки, что дает две возможности. Во-первых, сигналы на уровне популяции могут различаться в зависимости от используемой руки. Во-вторых, одни и те же сигналы на уровне популяции могут присутствовать, но по-разному распределяться по нейронам. Данные подтвердили эту вторую гипотезу. Мышечная активность была точно предсказана по активности ипсилатерального или контралатерального полушария. В более общем плане нам не удалось найти сигналы, характерные только для контралатерального полушария. Но если сигналы распределяются между полушариями, как они избегают воздействия не на ту руку? Мы обнаружили, что активность, связанная с каждой рукой, занимает отдельное подпространство, что позволяет декодерам мышечной активности естественным образом игнорировать сигналы, относящиеся к другой руке.
https://doi.org/10.7554/eLife.46159.001
Введение
Выходы моторной коры (M1) латерализованы: большинство проекций позвоночника влияют на контралатеральную мускулатуру. Таким образом, поражения M1 вызывают контралатеральный моторный дефицит (Liu and Rouiller, 1999; Murata et al., 2008; Passingham et al., 1983; Vilensky and Gilman, 2002). Точно так же электрическая микростимуляция активирует контралатеральную мускулатуру (Kwan et al., 1978; Sessle and Wiesendanger, 19).82). Степень латерализации вычислений внутри M1 остается менее ясной. Мозолистое тело соединяет M1 между полушариями, создавая потенциал для широкого сотрудничества (Gould et al., 1986; Jenny, 1979; Jones and Wise, 1977). Мозолисто-опосредованные взаимодействия легко выявляются с помощью протоколов парных импульсов TMS и могут включать чистое облегчение или подавление (Ferbert et al., 1992; Hanajima et al., 2001; Meyer et al., 1995). Очевидной ролью межполушарной коммуникации является координация бимануальных движений (Donchin et al.
 , 19).98; Хакен и др., 1985; Келсо, 1984; Кермади и др., 1998). Тем не менее, есть свидетельства того, что одноручные движения также включают обмен информацией между полушариями.
, 19).98; Хакен и др., 1985; Келсо, 1984; Кермади и др., 1998). Тем не менее, есть свидетельства того, что одноручные движения также включают обмен информацией между полушариями. Большинство физиологических исследований одноручных движений были сосредоточены на активности, противоположной подвижной конечности, на том основании, что контралатеральная активность является наиболее функционально значимой и, вероятно, наиболее распространенной. Тем не менее, ипсилатеральная активность может быть устойчивой. Во время движений пальцев активность ипсилатеральных одиночных нейронов умеренная, но присутствует (Aizawa et al., 19).90; Мацунами и Хамада, 1981 год; Tanji et al., 1988), а поведение можно расшифровать по активации BOLD в ипсилатеральной моторной коре (Berlot et al., 2019; Diedrichsen et al., 2013). Сообщалось о значительной ипсилатеральной активности во время движений плеча, таких как вытягивание еды из ящика (Kermadi et al., 1998; Kazennikov et al., 1999) или выполнение движений от центра кнаружи (Cisek et al.
 , 2003; Дончин и др., 2002; Гангули и др., 2009; Стейнберг и др., 2002).
, 2003; Дончин и др., 2002; Гангули и др., 2009; Стейнберг и др., 2002). Хотя наличие ипсилатеральной активности установлено, природа этой активности менее ясна. Немногие исследования напрямую сравнивали паттерны нейронной реакции, когда одно и то же движение выполняется одной рукой по сравнению с другой. В премоторных областях реакции периода задержки могут кодировать информацию о предстоящем достижении (Cisek et al., 2003) или захвате (Michaels and Scherberger, 2018) независимо от того, какая рука впоследствии будет двигаться, что позволяет предположить, что подготовительная деятельность в значительной степени не зависит от конечностей. Однако активность во время движения в большей степени зависела от конечностей как для премоторной коры, так и для M1 (Cisek et al., 2003). Стейнберг и др. (2002) сообщили об аналогичной направленной настройке одного нейрона в M1, независимо от того, какая рука двигалась, но также обнаружили доказательства зависимого от конечностей кодирования направления на популяционном уровне.
 Дончин и др. (1998) обнаружили, что активность во время бимануальных движений отличается от таковой во время мономануальных движений, но не анализировали, различается ли настройка реакции между контралатеральными и ипсилатеральными движениями. Таким образом, остается неясным, в какой степени паттерн ответов М1 — на уровне отдельных нейронов или на уровне популяции — зависит от используемой конечности.
Дончин и др. (1998) обнаружили, что активность во время бимануальных движений отличается от таковой во время мономануальных движений, но не анализировали, различается ли настройка реакции между контралатеральными и ипсилатеральными движениями. Таким образом, остается неясным, в какой степени паттерн ответов М1 — на уровне отдельных нейронов или на уровне популяции — зависит от используемой конечности. Если ответы не зависят от конечностей, то отношения между полушариями неизбежно просты: оба содержат одну и ту же информацию, закодированную одинаковым образом. Напротив, ответы, сильно зависящие от конечностей, вызовут дополнительные вопросы. Являются ли сигналы «более низкого уровня» (например, описывающие мышечную активность) более распространенными в контралатеральном полушарии? В более общем плане, какие сигналы являются общими для полушарий? Как устроена деятельность, при которой двигается только одна рука, когда оба полушария активны?
Мы исследовали эти вопросы с помощью новой задачи «езда на велосипеде», выполняемой либо левой, либо правой рукой.
 Мы регистрировали нейронную активность обоих полушарий одновременно. В отдельных сеансах мы регистрировали двустороннюю мышечную активность. Отдельные нейроны реагировали надежно независимо от того, какая рука выполняла задачу. Тем не менее, ответы сильно зависели от конечностей: для данного нейрона паттерны ответов, относящиеся к двум рукам, были по существу не связаны. Тем не менее, мы не нашли четких доказательств того, что два полушария содержат разные типы информации. Например, мышечная активность может быть одинаково хорошо расшифрована по контралатеральной или ипсилатеральной нейронной активности. В более широком смысле любой сигнал, сильно присутствующий в одном полушарии, присутствовал (и столь же силен) в другом. Таким образом, активность в данном полушарии содержит одинаковую информацию при движении одной руки по сравнению с другой, но эта информация очень по-разному распределяется по отдельным нейронам. Может показаться, что это приводит к парадоксу: как M1 может быть устойчиво активным, не управляя контралатеральным плечом? Решение появилось, когда мы исследовали популяционную активность: сигналы, специфичные для рук, были разделены на ортогональные измерения, что позволило простому декодеру естественным образом разделить сигналы, относящиеся к двум рукам.
Мы регистрировали нейронную активность обоих полушарий одновременно. В отдельных сеансах мы регистрировали двустороннюю мышечную активность. Отдельные нейроны реагировали надежно независимо от того, какая рука выполняла задачу. Тем не менее, ответы сильно зависели от конечностей: для данного нейрона паттерны ответов, относящиеся к двум рукам, были по существу не связаны. Тем не менее, мы не нашли четких доказательств того, что два полушария содержат разные типы информации. Например, мышечная активность может быть одинаково хорошо расшифрована по контралатеральной или ипсилатеральной нейронной активности. В более широком смысле любой сигнал, сильно присутствующий в одном полушарии, присутствовал (и столь же силен) в другом. Таким образом, активность в данном полушарии содержит одинаковую информацию при движении одной руки по сравнению с другой, но эта информация очень по-разному распределяется по отдельным нейронам. Может показаться, что это приводит к парадоксу: как M1 может быть устойчиво активным, не управляя контралатеральным плечом? Решение появилось, когда мы исследовали популяционную активность: сигналы, специфичные для рук, были разделены на ортогональные измерения, что позволило простому декодеру естественным образом разделить сигналы, относящиеся к двум рукам.
Результаты
Терминология
Мы используем следующую терминологию. Для нейронов в данном полушарии мы называем контралатеральную руку «ведомой рукой» (что отражает сильные связи с контралатеральным спинным мозгом). Мы называем ипсилатеральную руку «неуправляемой рукой». Таким образом, для нейрона, зарегистрированного в правом полушарии, левая рука является ведомой рукой, а правая рука — неуправляемой рукой. Для мышц ведомая рука — это просто рука, на которую действует мышца. Мы используем термин «ведущая кора» для обозначения полушария, контралатерального исполнительной руке. Мы используем термин «неуправляющая кора» для обозначения полушария, ипсилатерального по отношению к работающей руке.
Поведение
Двух обезьян (E и F) обучали езде на велосипеде, которую можно было выполнять любой рукой (рис. 1А). Левая и правая руки сжимали педали. Обезьяны выполняли блоки попыток левой и правой руки. Вращение правильной педали приводило к движению в виртуальной среде.
 Для успеха требовалось, чтобы неработающая рука оставалась неподвижной. В каждом испытании обезьяны циклически переходили от одной цели к другой, расположенной в семи циклах от нее (рис. 1В). Мишени располагались таким образом, чтобы цикл начинался и заканчивался либо в верхней части цикла («верхнее начало»), либо в нижней части цикла («нижнее начало»).
Для успеха требовалось, чтобы неработающая рука оставалась неподвижной. В каждом испытании обезьяны циклически переходили от одной цели к другой, расположенной в семи циклах от нее (рис. 1В). Мишени располагались таким образом, чтобы цикл начинался и заканчивался либо в верхней части цикла («верхнее начало»), либо в нижней части цикла («нижнее начало»). Поведение.
( A ) Схема задания. Вращение одной из двух педалей приводило к прохождению через виртуальную среду. Другая педаль должна была оставаться неподвижной. Эта схема упрощает физическую установку. В частности, в педалях использовалась ручка, которая обеспечивала постоянное положение рук, и скоба, сводившая к минимуму движение запястья. ( Б ) Поведение на двух примерах испытаний. После сигнала движения обезьяна сделала семь кругов одной рукой, удерживая другую руку неподвижно.
https://doi.org/10.7554/eLife.46159.002 Красный след: правое вертикальное положение. Синий след: левое вертикальное положение. ( C ) Структура задачи. Блоки правосторонних, левосторонних и бимануальных состояний предъявлялись в псевдослучайном порядке. В каждом блоке из 20 испытаний испытания были представлены в подблоках по пять испытаний. Для каждой комбинации направления езды на велосипеде и стартовой позиции был один подблок. ( D ) Распределение скорости езды на велосипеде для работающих и неработающих рук. Распределения по испытаниям. В каждом испытании средняя угловая скорость вычислялась после определения величины скорости в каждый момент времени. Красный: правая рука. Синий: левая рука. Точками показаны медианы распределения. Данные для обезьяны E. ( E ) То же, что и D, но для обезьяны F. ( F ) Для обезьяны E: расположение трепанационных отверстий, из которых были сделаны записи, наложенные на анатомическую карту, полученную с помощью МРТ. ( G ) То же, что и F, для Monkey F.
Красный след: правое вертикальное положение. Синий след: левое вертикальное положение. ( C ) Структура задачи. Блоки правосторонних, левосторонних и бимануальных состояний предъявлялись в псевдослучайном порядке. В каждом блоке из 20 испытаний испытания были представлены в подблоках по пять испытаний. Для каждой комбинации направления езды на велосипеде и стартовой позиции был один подблок. ( D ) Распределение скорости езды на велосипеде для работающих и неработающих рук. Распределения по испытаниям. В каждом испытании средняя угловая скорость вычислялась после определения величины скорости в каждый момент времени. Красный: правая рука. Синий: левая рука. Точками показаны медианы распределения. Данные для обезьяны E. ( E ) То же, что и D, но для обезьяны F. ( F ) Для обезьяны E: расположение трепанационных отверстий, из которых были сделаны записи, наложенные на анатомическую карту, полученную с помощью МРТ. ( G ) То же, что и F, для Monkey F.
В каждом блоке из 20 попыток каждая комбинация исходного положения и направления движения выполнялась в пяти последовательных попытках (рис. 1С). Обезьяны выполняли в среднем 29 и 21 испытание на каждое условие в день (обезьяны E и F соответственно). Обезьяны двигались быстро, со средней угловой скоростью 2,2 цикла/с и 1,8 цикла/с (обезьяны E и F; рис. 1D, E). Напротив, неэффективная рука двигалась очень мало (крайние левые распределения на рис. 1D, E). Средняя угловая скорость для неработающей руки составила 0,0016 цикла/с и 0,024 цикла/с (обезьяны E и F).
Нервные и мышечные реакции
Мы проанализировали частоту срабатывания 1150 единиц, зарегистрированных в обоих полушариях (рис. 1F–G). Большинство записей были сделаны из собственно M1, а некоторые из прилегающей каудальной области PMd.
 Усредненная по пробам частота срабатывания была рассчитана путем фильтрации последовательностей импульсов в одной попытке для получения непрерывной частоты, а затем корректировки временной базы для временного выравнивания поведения между испытаниями (рис. 2). Нейронные ответы обычно были ритмичными (рис. 3E–H) и могли быть почти синусоидальными (рис. 3E) или могли содержать дополнительную высокочастотную структуру (рис. 3G). Для сравнения мы записали активность основных мышц обеих рук (всего 48 записей). Временные характеристики ответов отдельных мышц (рис. 3A-D) во многом напоминали ответы отдельных нейронов. Однако мышцы и нейроны сильно различались по степени, в которой ответы были ограничены движениями одной руки.
Усредненная по пробам частота срабатывания была рассчитана путем фильтрации последовательностей импульсов в одной попытке для получения непрерывной частоты, а затем корректировки временной базы для временного выравнивания поведения между испытаниями (рис. 2). Нейронные ответы обычно были ритмичными (рис. 3E–H) и могли быть почти синусоидальными (рис. 3E) или могли содержать дополнительную высокочастотную структуру (рис. 3G). Для сравнения мы записали активность основных мышц обеих рук (всего 48 записей). Временные характеристики ответов отдельных мышц (рис. 3A-D) во многом напоминали ответы отдельных нейронов. Однако мышцы и нейроны сильно различались по степени, в которой ответы были ограничены движениями одной руки. Двухэтапная пробная процедура выравнивания.
( A ) Вертикальное положение рук во всех испытаниях (в рамках одного сеанса) для примера.
https://doi.org/10.7554/eLife.461590,003 Данные выровнены по первому полупериоду движения. Испытания окрашены в зеленый или фиолетовый цвет в зависимости от средней скорости езды на велосипеде для этого испытания. ( B ) Растровая диаграмма времени выброса для примера нейрона для тех же испытаний, что и в A. Испытания упорядочены по средней скорости цикла и выровнены, как в A. ( C ) Следы положения рук после второго шага выравнивания: корректировка временной базы каждого испытания таким образом, чтобы езда на велосипеде в течение средних шести циклов соответствовала типичной скорости вращения педалей 2 Гц. ( D ) Количество всплесков после второго шага выравнивания. ( E ) Средняя скорострельность, рассчитанная после второго шага выравнивания. Черный: средняя скорострельность. Серая заливка: стандартная ошибка в испытаниях. Метка внизу справа дает идентификацию нейрона.
Данные выровнены по первому полупериоду движения. Испытания окрашены в зеленый или фиолетовый цвет в зависимости от средней скорости езды на велосипеде для этого испытания. ( B ) Растровая диаграмма времени выброса для примера нейрона для тех же испытаний, что и в A. Испытания упорядочены по средней скорости цикла и выровнены, как в A. ( C ) Следы положения рук после второго шага выравнивания: корректировка временной базы каждого испытания таким образом, чтобы езда на велосипеде в течение средних шести циклов соответствовала типичной скорости вращения педалей 2 Гц. ( D ) Количество всплесков после второго шага выравнивания. ( E ) Средняя скорострельность, рассчитанная после второго шага выравнивания. Черный: средняя скорострельность. Серая заливка: стандартная ошибка в испытаниях. Метка внизу справа дает идентификацию нейрона. Активность в зависимости от времени для четырех примеров мышц (
AD ) и четырех примеров нейронов ( E–H ).
Каждая панель показывает активность для одного условия, выполненного либо с ведомым рычагом (синий), либо с неведомым рычагом (красный). Каждая трасса отображает активность, усредненную по пробам, с боковыми огибающими (иногда едва видимыми), показывающими стандартные ошибки. Серые прямоугольники показывают, когда педаль двигалась, с делениями, разделяющими каждый цикл. Все примеры взяты из Monkey E. Примеры нейронных записей были взяты из одиночной изоляции для всех фигур (анализ популяции включает одиночную и множественную изоляцию). Калибровочные столбцы составляют 10 импульсов в секунду. Для мышц вертикальный масштаб произволен, но сохраняется в пределах каждой панели.
https://doi.org/10.7554/eLife.46159.004Мышцы демонстрировали сильную активность только тогда, когда их ведомая рука выполняла задачу (рис.
 3A-D). Например, левая передняя дельтовидная мышца (рис. 3А) была активна, когда левая рука выполняла задание ( синий ), но не когда правая рука выполняла задание ( красный ). Хотя это и ожидалось, это прямое подтверждение важно из-за возможности того, что мышцы могли быть активны таким образом, чтобы педаль не двигалась (например, совместное сокращение). Такая активность потенциально могла быть существенной, что усложнило бы интерпретацию нейронной активности. Несколько мышц проявляли слабую активность, когда задача выполнялась не ведущей рукой (рис. 3А, С). Однако обычно это происходило только в конце движения, что согласуется с напряжением для обеспечения устойчивости при остановке.
3A-D). Например, левая передняя дельтовидная мышца (рис. 3А) была активна, когда левая рука выполняла задание ( синий ), но не когда правая рука выполняла задание ( красный ). Хотя это и ожидалось, это прямое подтверждение важно из-за возможности того, что мышцы могли быть активны таким образом, чтобы педаль не двигалась (например, совместное сокращение). Такая активность потенциально могла быть существенной, что усложнило бы интерпретацию нейронной активности. Несколько мышц проявляли слабую активность, когда задача выполнялась не ведущей рукой (рис. 3А, С). Однако обычно это происходило только в конце движения, что согласуется с напряжением для обеспечения устойчивости при остановке. В отличие от мышц, нейроны, как правило, были активны на протяжении всего движения, независимо от того, выполнялась ли задача ведомой (контралатеральной) или не ведущей (ипсилатеральной) рукой. Несколько нейронов были активны только при езде на велосипеде с ведомой рукой (рис. 3E), а в редких случаях нейрон был активен только при езде на велосипеде с не ведущей рукой (рис.
 3H). Однако большинство нейронов были активны в обеих ситуациях (рис. 3F, G). Нейронные реакции могут сильно различаться при езде на велосипеде с ведомой и не ведущей рукой. Паттерны нейронных реакций могут меняться как по фазе (рис. 3F), так и по структуре (рис. 3G) в зависимости от того, какая рука выполняла задачу.
3H). Однако большинство нейронов были активны в обеих ситуациях (рис. 3F, G). Нейронные реакции могут сильно различаться при езде на велосипеде с ведомой и не ведущей рукой. Паттерны нейронных реакций могут меняться как по фазе (рис. 3F), так и по структуре (рис. 3G) в зависимости от того, какая рука выполняла задачу. Нейроны активны во время движений любой руки
Для количественной оценки предпочтения руки отдельными единицами мы сравнили модуляцию скорости стрельбы, когда задача выполнялась с ведомой и не ведущей рукой. Модуляцию оценивали как стандартное отклонение скорости стрельбы в зависимости от времени и условий. Средняя модуляция вычислялась один раз для всех условий, в которых ведомая рука выполняла задачу, и еще раз для всех условий, когда задачу выполняла неведомая рука. Мы анализировали только средние циклы движения (исключая первый цикл и два последних цикла). Сосредоточение внимания на этом «устойчивом состоянии» нервной реакции помогает интерпретации, потому что неработающие мышцы рук имели минимальную устойчивую модуляцию.
 Мы рассчитали «индекс предпочтения руки»: разницу в модуляции для ведомой и не ведомой руки, деленную на сумму. Этот индекс колеблется от -1 до 1, причем крайние значения указывают на полное предпочтение неведущего и ведомого рычагов соответственно. Нулевой индекс предпочтения руки указывает на то, что нейрон одинаково реагировал независимо от используемой руки.
Мы рассчитали «индекс предпочтения руки»: разницу в модуляции для ведомой и не ведомой руки, деленную на сумму. Этот индекс колеблется от -1 до 1, причем крайние значения указывают на полное предпочтение неведущего и ведомого рычагов соответственно. Нулевой индекс предпочтения руки указывает на то, что нейрон одинаково реагировал независимо от используемой руки. Чтобы установить базовый уровень для сравнения, мы рассчитали индекс предпочтения руки для каждой мышцы. Индексы предпочтения рук, как правило, были высокими для мышц (рис. 4А), подтверждая, что мышцы были активны в первую очередь, когда задача выполнялась ведомой рукой. Несколько мышц показали слабую активацию независимо от используемой руки, что привело к более низким показателям. Тем не менее, большинство мышц реагировали хорошо во время езды на велосипеде и были гораздо более активны, когда задача выполнялась ведомой рукой. Медианный индекс предпочтения руки был близок к единице (E: 0,86; F: 0,9).8; Рисунок 4A, 91 105 синих точек (91 106) и модальный отклик возник при единице.

Мышечные реакции латерализованы, а нервные — нет.
( A ) Гистограммы индекса предпочтения руки для всех зарегистрированных мышц. Заштрихованные области указывают на предпочтение не ведущего рычага (красный) и ведомого рычага (синий). Синяя точка указывает на медиану. ( B ) Гистограммы индекса предпочтения руки для всех зарегистрированных нейронов. ( C ) Гистограммы, обобщающие для отдельных нейронов сходство ответов при выполнении задачи с ведомой и не ведомой рукой. Для каждого нейрона мы рассчитали корреляцию между этими двумя ответами. Черные гистограммы отображают распределение таких корреляций по всем нейронам. Оранжевая кривая: контроль, демонстрирующий, что высокие корреляции наблюдаются, как и ожидалось, при сравнении откликов в условиях верхнего и нижнего начала. Зеленый след: распределение корреляций между парами разных единиц.
https://doi.org/10.7554/eLife.46159.005 Это распределение иллюстрирует диапазон случайных корреляций, наблюдаемых, если нет тенденции к сходству ответов, за исключением того, что они получены из одного и того же общего набора ответов.
Это распределение иллюстрирует диапазон случайных корреляций, наблюдаемых, если нет тенденции к сходству ответов, за исключением того, что они получены из одного и того же общего набора ответов. Напротив, нейроны редко имели индексы предпочтения рук, близкие к единице (рис. 4В). Вместо этого центр распределения был лишь немного выше нуля (медиана = 0,07 и 0,31 для Monkey E/F). Таким образом, нервные реакции с гораздо большей вероятностью, чем мышечные, будут одинаковыми по величине независимо от используемой руки. Кроме того, многие нейроны имели индексы предпочтения руки <0, что указывает на более сильную модуляцию, когда неуправляемая рука выполняла задачу (обезьяна E: 201/533 нейрона; обезьяна F: 107/617 нейронов). Результаты были практически идентичными (медианные индексы предпочтения руки 0,06 и 0,31), если анализ был ограничен более задними записями (исключая влияние любых нейронов, зарегистрированных из PMd или из пограничной области между M1 и PMd).

Таким образом, нейроны могут быть достаточно активны, даже когда задача выполняется их не ведущей рукой. Могут ли такие реакции быть связаны с небольшими движениями ведомой руки? Это объяснение маловероятно априори. Как описано выше, движения неработающей руки были небольшими (рис. 1D, E), а соответствующая мышечная активность была слабой (рис. 4A). В принципе, нервные реакции, связанные со слабой мышечной активностью, могут усиливаться за счет нормализации или какой-либо другой нелинейности. Однако такое увеличение должно быть очень сильным. Чтобы соответствовать медианным индексам нейронных предпочтений руки, мышечная активность в неработающей руке должна быть увеличена в 12 раз (обезьяна E) и в 52 раза (обезьяна F). Кроме того, увеличение не может объяснить тот факт, что нейроны обычно имеют отрицательные индексы предпочтения руки, в то время как мышцы редко (обезьяна E) или никогда (обезьяна F).
Мы провели дополнительный контроль, чтобы узнать, действительно ли ответы нейронов, когда их неведомая рука выполняла задачу, отражали небольшие движения их ведомой руки.
 Такие движения различались в разных испытаниях (рис. 5А), что позволило нам разделить испытания на те, в которых движения превышали медиану (очень скромное движение, красный ) и ниже медианы (почти стационарное, черный ). Средняя частота срабатывания была очень похожа в этих двух случаях, как показано для одного примера нейрона (рис. 5B). Различия были небольшими и были значимыми только в некоторые моменты ( черных точек ) р<0,05 при 1000 повторных выборках, материалы и методы). Во всех нейронах 5% точек данных показали значительные различия (рис. 5C, D). Это равно проценту, ожидаемому случайно, и, таким образом, согласуется с отсутствием надежного влияния небольших движений. Применение того же анализа к (слабой) мышечной активности в неработающей руке выявило значительные различия при удвоенной частоте шансов (10% точек данных) в целом и с пиком в четыре раза выше вероятности ближе к концу движения, когда мышечная активность стала более заметной (21% точек данных).
Такие движения различались в разных испытаниях (рис. 5А), что позволило нам разделить испытания на те, в которых движения превышали медиану (очень скромное движение, красный ) и ниже медианы (почти стационарное, черный ). Средняя частота срабатывания была очень похожа в этих двух случаях, как показано для одного примера нейрона (рис. 5B). Различия были небольшими и были значимыми только в некоторые моменты ( черных точек ) р<0,05 при 1000 повторных выборках, материалы и методы). Во всех нейронах 5% точек данных показали значительные различия (рис. 5C, D). Это равно проценту, ожидаемому случайно, и, таким образом, согласуется с отсутствием надежного влияния небольших движений. Применение того же анализа к (слабой) мышечной активности в неработающей руке выявило значительные различия при удвоенной частоте шансов (10% точек данных) в целом и с пиком в четыре раза выше вероятности ближе к концу движения, когда мышечная активность стала более заметной (21% точек данных).
Небольшие движения неработающей руки не могут объяснить модуляцию нейронной активности в неуправляемой коре.
( A ) В анализе использовалось распределение (между испытаниями) средней угловой скорости неработающей руки. Это распределение показано для одного состояния, зарегистрированного в 1 день. Испытания были разделены на те, в которых средняя скорость была меньше (серый) или больше (красный) медианы (вертикальная пунктирная линия). ( B ) Частота срабатывания одного примера нейрона для этих двух групп: испытания со скоростями меньше (черный) и выше (красный) медианы. Конверты показывают стандартные ошибки среднего. Черные точки вверху обозначают время, когда эти два показателя значительно различались (p<0,05). Условные обозначения, как на рисунке 3. ( C ) Процент нейронов (черная кривая), показывающих статистически значимую разницу ( p<0,05) в частоте возбуждения между испытаниями со скоростями меньше и выше медианы.
https://doi.org/10.7554/eLife.46159.006 Различия возникали так часто, как и ожидалось, случайно (красная линия на уровне 5%). Серый прямоугольник обозначает время движения. Каждая отметка обозначает цикл. Данные относятся к обезьяне Е. Анализ основан на 426 единицах. ( D ) То же, что и C, но для Monkey F. Анализ основан на 479 единицах.
Различия возникали так часто, как и ожидалось, случайно (красная линия на уровне 5%). Серый прямоугольник обозначает время движения. Каждая отметка обозначает цикл. Данные относятся к обезьяне Е. Анализ основан на 426 единицах. ( D ) То же, что и C, но для Monkey F. Анализ основан на 479 единицах. Таким образом, мышцы молчали или в лучшем случае были слабо активны, когда задача выполнялась не ведущей рукой. Эта слабая мышечная активность была статистически связана с небольшими движениями ведомой (не работающей) руки. Напротив, большинство нейронов демонстрировали устойчивые ответы, когда задача выполнялась их неуправляемой рукой. Эти ответы не были статистически связаны с небольшими движениями их ведомой (не работающей) руки. Предыдущие исследования показали, что нейроны могут быть активны, когда задача выполняется не ведущей рукой, хотя и в разной степени (Cisek et al.
 , 2003; Donchin et al., 19).98; Кермади и др., 1998 г.; Стейнберг и др., 2002; Танджи и др., 1988). Настоящие результаты повторяют обнаружение слабо латеральных ответов в моторной коре и в значительной степени исключают возможные объяснения, основанные на небольших остаточных движениях ведомой руки.
, 2003; Donchin et al., 19).98; Кермади и др., 1998 г.; Стейнберг и др., 2002; Танджи и др., 1988). Настоящие результаты повторяют обнаружение слабо латеральных ответов в моторной коре и в значительной степени исключают возможные объяснения, основанные на небольших остаточных движениях ведомой руки. Паттерны нейронных реакций зависят от конечностей
Одним из правдоподобных объяснений слабо латеральных ответов является то, что нейронная активность кодирует высокоуровневые, независимые от конечностей особенности движения. Например, активность может кодировать скорость руки, цель движения или какую-то другую величину, независимо от того, какая конечность движется. Подготовительная активность в более передней ростральной премоторной коре может проявлять в значительной степени независимые от конечностей ответы (Cisek et al., 2003). Может ли это быть верно и для моторной коры во время движения? Две руки выполняли очень похожие движения в нашей задаче. Таким образом, независимость конечностей должна отражаться сходными нейронными реакциями независимо от работающей руки.
 Вместо этого нейронные реакции сильно зависели от конечностей. Ответы часто различались по фазе (рис. 3F) и/или структуре (рис. 3G) в зависимости от того, какая рука выполняла задачу.
Вместо этого нейронные реакции сильно зависели от конечностей. Ответы часто различались по фазе (рис. 3F) и/или структуре (рис. 3G) в зависимости от того, какая рука выполняла задачу. Чтобы обеспечить количественную оценку, для каждого нейрона мы рассчитали корреляцию между паттернами скорости возбуждения, когда ведомая и не ведомая рука выполняла задачу (рис. 4C). При анализе учитывались только средние циклы (2-5). Это гарантировало, что высокие корреляции указывают на схожие модели ответов, а не просто на частоту стрельбы, которая неспецифически возрастает во время движения. В среднем корреляция была близка к нулю (средняя корреляция: 0,16 и 0,08 для Monkey E и F). Сильно коррелированные ответы были в меньшинстве: только 18/533 (E) и 9/617 (F) нейронов имели корреляции >= 0,75. Таким образом, для данного нейрона было удивительно мало связи между реакциями, когда задача выполнялась с управляемой и неуправляемой рукой.
Для сравнения мы рассчитали распределение корреляций ответов между случайными парами нейронов.
 Это дает распределение корреляций, ожидаемое случайно, если ответы связаны только в том смысле, что они исходят из одного и того же общего набора ответов. Эмпирические распределения ( черных ) были значительно смещены вправо по сравнению с этими «перетасованными» ( зелеными ) распределениями (p<10−3 для обеих обезьян, начальной загрузки, материалов и методов). Тем не менее, хотя этот сдвиг был значительным, он был очень скромным: эмпирические распределения были сосредоточены гораздо ближе к нулю, чем к единице. Таким образом, корреляция между ответами двух плеч для данного нейрона была лишь немного больше, чем корреляция между двумя произвольно выбранными нейронами.
Это дает распределение корреляций, ожидаемое случайно, если ответы связаны только в том смысле, что они исходят из одного и того же общего набора ответов. Эмпирические распределения ( черных ) были значительно смещены вправо по сравнению с этими «перетасованными» ( зелеными ) распределениями (p<10−3 для обеих обезьян, начальной загрузки, материалов и методов). Тем не менее, хотя этот сдвиг был значительным, он был очень скромным: эмпирические распределения были сосредоточены гораздо ближе к нулю, чем к единице. Таким образом, корреляция между ответами двух плеч для данного нейрона была лишь немного больше, чем корреляция между двумя произвольно выбранными нейронами. Могут ли корреляции казаться искусственно заниженными, если ответы слабые или шумные? Хотя ошибка выборки неизбежно снижает корреляции, она вряд ли является причиной наблюдаемых нами низких корреляций. Езда на велосипеде вызывала особенно сильные нейронные реакции с соответственно небольшими стандартными ошибками средней частоты возбуждения (рис.
 3E-H, конверты показывают SEM). Мы также решили эту проблему, вычислив для каждого нейрона корреляцию между частотой срабатывания для условий верхнего старта и нижнего старта. Поведение этих двух условий было очень схожим во время средних циклов (после фазы выравнивания), поэтому корреляции должны быть высокими. Это действительно было так (рис. 4C, 9).1105 оранжевый , медианы 0,72 и 0,77), подтверждая, что ошибка выборки не препятствует возможности измерения высоких корреляций.
3E-H, конверты показывают SEM). Мы также решили эту проблему, вычислив для каждого нейрона корреляцию между частотой срабатывания для условий верхнего старта и нижнего старта. Поведение этих двух условий было очень схожим во время средних циклов (после фазы выравнивания), поэтому корреляции должны быть высокими. Это действительно было так (рис. 4C, 9).1105 оранжевый , медианы 0,72 и 0,77), подтверждая, что ошибка выборки не препятствует возможности измерения высоких корреляций. Эти результаты исключают гипотезу преимущественно независимых от конечностей реакций. Отдельные нейроны показывали очень разные ответы в зависимости от того, какая рука выполняла задачу. Ответы были почти такими же разными, как если бы не было никакой связи между паттернами активности, связанными с двумя руками.
Нейрон-нейронные корреляции зависят от конечностей
Рассмотрим два нейрона, которые имеют схожий паттерн ответа, когда задача выполняется с ведомой рукой (на рис. 6A-D показаны четыре такие пары примеров).
 Что происходит, когда задача выполняется с неведомой рукой? Из приведенного выше анализа мы знаем, что характер реакции каждого нейрона, вероятно, изменится. Происходит ли это скоординировано, так что два нейрона остаются коррелированными друг с другом? Это согласуется с идеей о том, что нейроны со связанными ответами «кодируют» связанные признаки и продолжают делать это в новых контекстах. На самом деле это свойство редко наблюдалось. Иногда мы наблюдали нейроны, которые сильно коррелировали, когда ведомая рука выполняла задачу, и оставались сильно коррелированными, когда задача выполнялась не ведущей рукой (рис. 6А). Тем не менее, корреляции также часто инвертировались (рис. 6B), исчезали сильные корреляции (рис. 6C) или нейроны претерпевали очень разные изменения в величине ответа (рис. 6D).
Что происходит, когда задача выполняется с неведомой рукой? Из приведенного выше анализа мы знаем, что характер реакции каждого нейрона, вероятно, изменится. Происходит ли это скоординировано, так что два нейрона остаются коррелированными друг с другом? Это согласуется с идеей о том, что нейроны со связанными ответами «кодируют» связанные признаки и продолжают делать это в новых контекстах. На самом деле это свойство редко наблюдалось. Иногда мы наблюдали нейроны, которые сильно коррелировали, когда ведомая рука выполняла задачу, и оставались сильно коррелированными, когда задача выполнялась не ведущей рукой (рис. 6А). Тем не менее, корреляции также часто инвертировались (рис. 6B), исчезали сильные корреляции (рис. 6C) или нейроны претерпевали очень разные изменения в величине ответа (рис. 6D). Корреляции между нейронами зависят от того, какое плечо используется.
( A ) Средняя частота срабатывания двух нейронов (зеленая и оранжевая кривые) при одном условии, выполненном с ведомой рукой (верхняя пара дорожек) и неведомой рукой (нижняя пара дорожек).
https://doi.org/10.7554/eLife.461590,007 Ответы показаны для средних циклов, которые составляют основу анализа ниже. В этом примере частота возбуждения двух нейронов сильно коррелировала, когда ведомая рука выполняла задачу, и оставалась такой, когда задача выполнялась не ведущей рукой. Затенение показывает стандартные ошибки среднего. ( B–D ) Ответы трех других пар нейронов. Все пары показывают коррелированные ответы, когда задача выполнялась с ведомой рукой. Корреляции исчезают или даже инвертируются при выполнении задачи неведомой рукой. ( E ) Попарные корреляции между всеми единицами, зарегистрированными в левом полушарии обезьяны E. Каждая панель отображает матрицу корреляции для одного состояния, указанного вверху. Порядок единиц был основан на данных на левой панели и сохраняется на всех панелях. ( F ) то же, но для Monkey F. ( G ) Диаграммы рассеяния парных корреляций. Каждая точка соответствует паре единиц для данного условия и отображает корреляцию скорострельности при использовании неприводной руки по сравнению с таковой при использовании ведомой руки.
Ответы показаны для средних циклов, которые составляют основу анализа ниже. В этом примере частота возбуждения двух нейронов сильно коррелировала, когда ведомая рука выполняла задачу, и оставалась такой, когда задача выполнялась не ведущей рукой. Затенение показывает стандартные ошибки среднего. ( B–D ) Ответы трех других пар нейронов. Все пары показывают коррелированные ответы, когда задача выполнялась с ведомой рукой. Корреляции исчезают или даже инвертируются при выполнении задачи неведомой рукой. ( E ) Попарные корреляции между всеми единицами, зарегистрированными в левом полушарии обезьяны E. Каждая панель отображает матрицу корреляции для одного состояния, указанного вверху. Порядок единиц был основан на данных на левой панели и сохраняется на всех панелях. ( F ) то же, но для Monkey F. ( G ) Диаграммы рассеяния парных корреляций. Каждая точка соответствует паре единиц для данного условия и отображает корреляцию скорострельности при использовании неприводной руки по сравнению с таковой при использовании ведомой руки. Для облегчения визуализации показаны случайно выбранные 1% точек данных. ( H ) График плотности парных корреляций для Обезьяны E (слева) и Обезьяны F (справа).
Для облегчения визуализации показаны случайно выбранные 1% точек данных. ( H ) График плотности парных корреляций для Обезьяны E (слева) и Обезьяны F (справа). Мы рассчитали матрицы корреляции для оценки таких эффектов среди населения. Чтобы облегчить визуализацию, мы приказали нейронам сгруппировать ответы, которые были похожи, когда задача выполнялась с ведомой рукой, что привело к блочной структуре (рис. 6E-F, слева ). Мы спросили, остается ли эта корреляционная структура аналогичной, когда задача выполнялась не ведущей рукой. Обратите внимание, что корреляционная матрица может оставаться идентичной, даже если каждый нейрон меняет свой ответ, пока коррелированные нейроны остаются коррелированными. Вместо этого корреляционная структура резко изменилась. В результате первоначальный порядок больше не группирует нейроны с похожими свойствами ответа (9).
 1105, правый столбец на рис. 6E-F).
1105, правый столбец на рис. 6E-F). Это изменение в структуре корреляции произошло не из-за того, что корреляции были в значительной степени ложными, как это могло бы произойти, если бы предполагаемая частота стрельбы была зашумлена. Чтобы исследовать эту возможность, мы оценили структуру корреляции между условиями, при которых езда на велосипеде начинается в верхней и нижней частях цикла. Структура корреляции была очень похожей в этих двух случаях (сравните 91 105 средних 91 106 и 91 105 левых столбцов 91 106 на рис. 6E–F). Этот вывод также показывает, что не всякое изменение задачи приводит к изменению корреляционной структуры; изменение исходного положения оказало относительно небольшое влияние, в то время как изменение рабочей руки оказало существенное влияние.
Каждая матрица на рис. 6E–F соответствует заданному условию (начальное положение и направление цикла). Мы хотели обобщить во всех таких условиях степень, в которой корреляции сохраняются или не сохраняются, когда задача выполняется одной рукой по сравнению с другой.
 Для этого для каждого состояния и каждой пары нейронов мы нанесли на график их корреляцию частоты возбуждения, когда неуправляемая рука выполняла задачу, по сравнению с их корреляцией, когда задача выполнялась управляемой рукой (для облегчения визуализации диаграммы рассеяния на рис. 6G показывают случайный 1% точек данных; графики плотности на рисунке 6H охватывают все точки данных). При сохранении корреляций получится диагональная структура. Фактически, коррелированные нейроны оставались коррелированными, а антикоррелированные нейроны — антикоррелированными. Метакорреляция составила 0,1 и 0,05 (обезьяны E и F). Таким образом, если два нейрона реагировали одинаково, когда их ведомая рука выполняла задачу, это мало что говорило о том, будут ли эти нейроны реагировать одинаково, когда их не ведущая рука выполняла задачу.
Для этого для каждого состояния и каждой пары нейронов мы нанесли на график их корреляцию частоты возбуждения, когда неуправляемая рука выполняла задачу, по сравнению с их корреляцией, когда задача выполнялась управляемой рукой (для облегчения визуализации диаграммы рассеяния на рис. 6G показывают случайный 1% точек данных; графики плотности на рисунке 6H охватывают все точки данных). При сохранении корреляций получится диагональная структура. Фактически, коррелированные нейроны оставались коррелированными, а антикоррелированные нейроны — антикоррелированными. Метакорреляция составила 0,1 и 0,05 (обезьяны E и F). Таким образом, если два нейрона реагировали одинаково, когда их ведомая рука выполняла задачу, это мало что говорило о том, будут ли эти нейроны реагировать одинаково, когда их не ведущая рука выполняла задачу. Активность населения изоморфна по полушариям
Приведенные выше результаты показывают, что реакции отдельных нейронов и корреляции между нейронами сильно различаются в зависимости от того, какая рука выполняет задание.
 Одно из возможных объяснений состоит в том, что присутствуют очень разные сигналы. Возможно, мышечные сигналы преобладают при использовании ведомой руки, в то время как более абстрактные сигналы становятся преобладающими при использовании не ведущей руки. Альтернативное объяснение заключается в том, что многие из одних и тех же сигналов присутствуют, но по-разному отражаются на уровне отдельных нейронов.
Одно из возможных объяснений состоит в том, что присутствуют очень разные сигналы. Возможно, мышечные сигналы преобладают при использовании ведомой руки, в то время как более абстрактные сигналы становятся преобладающими при использовании не ведущей руки. Альтернативное объяснение заключается в том, что многие из одних и тех же сигналов присутствуют, но по-разному отражаются на уровне отдельных нейронов. Сначала мы спросили, присутствуют ли мышечные сигналы в обоих полушариях. Мы обучили регуляризованный линейный декодер прогнозировать активность мышц рабочей руки на основе нейронной активности (рис. 7В). Мы сравнили прогнозы, основанные на активности управляющей коры (контралатеральной по отношению к работающей руке) и неуправляющей коры (ипсилатеральной по отношению к работающей руке). Производительность декодера была количественно определена путем тестирования обобщения для состояния удерживания с повторением этой процедуры для каждого состояния.
Доминирующие сигналы очень похожи в ведущей и не ведущей коре.

( A ) Пример паттернов мышечной активности (черный) и прогнозы мышечной активности на основе линейного декодирования нейронной активности в управляющей (синий) и неуправляемой (красный) коре. Примеры показаны для одной мышцы от каждой обезьяны. В обоих случаях данные взяты из тестового условия и иллюстрируют эффективность обобщения. ( B ) Карикатура, иллюстрирующая подход к анализу для сравнения производительности декодирования мышц от управляющей и неуправляемой коры. Активность исполнительных мышц руки (синяя кривая) является результатом нисходящих связей (черные стрелки) от нейронов в управляющей коре (синяя сеть). Таким образом, должно быть возможно предсказать (синяя стрелка), что мышечная активность по нейронной активности регистрируется из управляющей коры. Наличие или отсутствие мышечных сигналов в неуправляемой коре (красная сеть) оценивали, задавая вопрос, насколько хорошо такая активность предсказывает (красная стрелка) активность исполнительных мышц руки.
https://doi.org/10.7554/eLife.461590,008 ( C ) Эффективность прогнозирования для сравнения, показанного на B. Данные относятся к обезьяне E. Каждая линия соответствует одному поведенческому состоянию и показывает процентную дисперсию (активности мышечной популяции), предсказанную декодированием на основе активности нейронов коры головного и неуправляющего секторов. . ( D ) Как и в C, для Обезьяны F. ( E ) Карикатура, иллюстрирующая анализ вопроса о том, присутствуют ли сигналы, передаваемые управляющей корой, в неуправляемой коре. Когда задача выполняется данной рукой, нейронная активность в соответствующей управляющей коре предсказывается либо по активности других нейронов либо в управляющей коре (синяя стрелка), либо в неуправляющей коре (красная стрелка). ( F ) Эффективность прогнозирования для сравнения, показанного на E. Данные относятся к Monkey E. Каждая цветовая кривая показывает среднюю производительность обобщения по восьми поведенческим условиям для данного ранга прогноза с использованием цветов радуги от красного (10 измерений) до фиолетового (один измерение).
( C ) Эффективность прогнозирования для сравнения, показанного на B. Данные относятся к обезьяне E. Каждая линия соответствует одному поведенческому состоянию и показывает процентную дисперсию (активности мышечной популяции), предсказанную декодированием на основе активности нейронов коры головного и неуправляющего секторов. . ( D ) Как и в C, для Обезьяны F. ( E ) Карикатура, иллюстрирующая анализ вопроса о том, присутствуют ли сигналы, передаваемые управляющей корой, в неуправляемой коре. Когда задача выполняется данной рукой, нейронная активность в соответствующей управляющей коре предсказывается либо по активности других нейронов либо в управляющей коре (синяя стрелка), либо в неуправляющей коре (красная стрелка). ( F ) Эффективность прогнозирования для сравнения, показанного на E. Данные относятся к Monkey E. Каждая цветовая кривая показывает среднюю производительность обобщения по восьми поведенческим условиям для данного ранга прогноза с использованием цветов радуги от красного (10 измерений) до фиолетового (один измерение). В каждой строке сравнивается производительность при прогнозировании ведущей активности коры на основе ведущей активности коры (слева) и без вождения (справа). ( G ) Как на панели F, для данных от Monkey F.
В каждой строке сравнивается производительность при прогнозировании ведущей активности коры на основе ведущей активности коры (слева) и без вождения (справа). ( G ) Как на панели F, для данных от Monkey F. Мышечная активность была точно предсказана по активности как управляющей коры (рис. 7A, синий ), так и активности неуправляемой коры ( красный ). Во всех условиях обобщение R2 было высоким как для управляющей, так и для неуправляющей коры (рис. 7C, D, эффективность обобщения, рассчитанная для всех мышц). Производительность обобщения была ниже для неуправляемой коры, но это было небольшим эффектом и было значимым только для одной обезьяны (p = 0,15 и p = 0,016 для обезьян E и F, двусторонний знаковый ранговый критерий Уилкоксона для восьми условий). Таким образом, мы не увидели доказательств того, что сигналы, связанные с мышечной активностью, отсутствовали в неуправляемой коре.
Могут ли другие сигналы ограничиваться управляющей корой? Вместо того, чтобы пытаться вывести конкретные сигналы-кандидаты, мы разработали метод для общего решения этого вопроса. Мы оценили, насколько хорошо можно предсказать активность управляющей коры по доминантным сигналам от неуправляющей коры. Для этого мы спроецировали неуправляющую активность коры на ее главные основные компоненты, а затем регрессировали управляющую активность коры относительно этих проекций (рис. 7Е). Для сравнения мы подсчитали, насколько хорошо активность управляющей коры может предсказывать сама себя, путем регрессии активности каждой единицы управляющей коры относительно проекций основных компонентов, полученных из всех других единиц управляющей коры, методом исключения одного.
Производительность обобщения зависела от количества основных компонентов (рис. 7F, G) и была выше для большего количества компонентов (указывая на то, что переоснащение не было серьезной проблемой). Ключевой вопрос заключается в том, имеет ли значение для данного количества компонентов, объясняется ли активность управляющей коры проекциями управляющей коры или проекциями неуправляющей коры.
Это действительно было так, но это был очень скромный эффект. Например, при использовании 10 основных компонентов обобщение R2 составило 82% и 85% (обезьяны E и F) при регрессии по проекциям ведущей коры по сравнению с 81% и 81% при регрессии по проекциям не ведущей коры. Это различие было статистически достоверным (p<0,01 для обеих обезьян, тест рандомизации со 100 повторными выборками, материалы и методы), но было намного меньше, чем ожидалось, если бы были сигналы, которые сильно присутствовали в управляющей коре, но были слабыми или отсутствовали в неуправляющей коре. (Рисунок 7 Приложение 1). Таким образом, доминирующие сигналы, присутствующие в верхней части основных компонентов, почти идентичны между управляющей и неуправляющей корой. Предостережение заключается в том, что меньшие сигналы могут быть совершенно другими, что было бы трудно различить, учитывая нашу относительно простую задачу. Активность охватывает небольшое количество измерений при циклировании в устойчивом состоянии (на пять основных компонентов приходится ~ 80 % дисперсии).
Вполне возможно, что различия в меньших сигналах, переносимых другими измерениями, могут быть обнаружены при дополнительной сложности задачи (например, при езде на велосипеде с разной скоростью или при разных нагрузках). Запутывание нейронных траекторий низкое для обоих полушарий
Недавно мы описали основные различия между активностью популяции M1 и активностью нижестоящих мышц (Russo et al., 2018). Нейронная популяционная активность, но не мышечная популяционная активность, позволяет избежать «запутывания траекторий». Запутывание траекторий определяется как появление сходных состояний популяций с очень разными производными. Запутанность траекторий становится высокой, если траектория популяции пересекает саму себя или если траектория для одного состояния проходит рядом с траекторией для другого условия, двигаясь в другом направлении. Рекуррентные сети, генерирующие шаблоны, устойчивы к шуму только в том случае, если запутывание траекторий низкое, что позволяет объяснить, почему в M1 наблюдалось низкое запутывание траекторий.
Неизвестно, участвует ли неуправляющая кора (через мозолистые связи) в формировании паттернов. Примечательно, что область коры головного мозга может быть активной во время езды на велосипеде, но при этом иметь сильное запутывание траектории; это верно для проприоцептивной первичной соматосенсорной коры (Russo et al., 2018). Мы рассчитали индекс запутывания (Russo et al., 2018) для каждой временной точки в середине цикла во всех условиях. Мышцы часто демонстрировали сильное запутывание траектории, что выявлялось длинным правым хвостом в кумулятивных распределениях (рис. 8A, B, черных линий ). Ведущая кора демонстрировала стабильно низкую запутанность траекторий: кумулятивное распределение ( синий ) рано стабилизировалось. Это повторяет предыдущие результаты: запутывание траекторий для движущей коры намного ниже, чем для популяции мышц, расположенных ниже по течению. Примечательно, что это верно даже при том, что реакции отдельных нейронов и отдельных мышц внешне похожи.
Запутывание траекторий одинаково для управляющей и неуправляющей коры.
( A ) Кумулятивное распределение запутывания траекторий для управляющей коры (синий), неуправляемой коры (красный) и мышечной активности (черный). Распределения были составлены по временным точкам и условиям. Данные для Monkey E. ( B ) Как в A, для Monkey F.
https://doi.org/10.7554/eLife.46159.010Запутанность траектории также была низкой для неуправляемой коры ( красный ). У обезьяны E запутывание было немного выше в коре головного мозга, не управляющей автомобилем, по сравнению с ведущей (468 ± 201 по сравнению с 420 ± 153; среднее значение ± стандартное отклонение), в то время как для обезьяны F все было наоборот (374 ± 105 по сравнению с 430 ± 146).
Таким образом, запутывание траекторий одинаково мало для обеих кор, с небольшими и непостоянными различиями. Важно отметить, что как для управляющей, так и для неуправляющей коры запутывание нервных траекторий было намного ниже, чем запутывание мышечных траекторий. В среднем 2296 ± 1766 для Обезьяны E и 4392 ± 2950 для Обезьяны F. Наличие одинаково низкой запутанности в коре головного мозга, отвечающей за вождение, по сравнению с корой, не отвечающей за вождение, согласуется с приведенным выше выводом о сходстве основных сигналов. Таким образом, между полушариями мало различий ни в основных сигналах, ни в организации популяционных траекторий. Примечательно, что такое сходство не является неизбежным при сравнении ответов населения; мышечные траектории имели другую и гораздо более запутанную организацию.
Нервная активность занимает разные измерения при движениях разных рук
Приведенные выше результаты подтверждают, что во время движения данной руки нейроны в обоих полушариях активны и передают одинаковые сигналы.
В самом деле, по реакции данного нейрона редко можно сказать, в каком полушарии находится этот нейрон. С одной стороны, такой широкий обмен информацией имеет смысл, учитывая тесную взаимосвязь двух полушарий. С другой стороны, нисходящее управление, обеспечиваемое каждым полушарием, должно по-прежнему иметь возможность разделять сигналы, относящиеся к соответствующему плечу, и избегать загрязнения сигналами, относящимися к другому плечу. Структурирован ли ответ населения таким образом, чтобы способствовать такому разделению? Можно ли добиться разделения, основываясь исключительно на свойствах самих ответов, не зная полушарной идентичности каждого нейрона? При ответе на эти вопросы мы черпали вдохновение из недавней работы, в которой предполагалось, что некоторые нейронные измерения в моторной коре являются «мышечно-потенциальными» — активность в этих измерениях производит выходные данные, которые будут влиять на мышцы, — в то время как другие измерения являются «мышечными нулевыми» — активность в эти размеры не имеют прямого исходящего влияния на мышечную активность (Druckmann and Chklovskiy, 2012; Kaufman et al.
, 2014). Наличие выходных нулевых измерений является естественным (и, как правило, неизбежным) в рекуррентных сетях, генерирующих шаблоны. Мы задались вопросом, может ли этот принцип применяться к настоящему делу. Мы рассматривали все зарегистрированные нейроны в обоих полушариях как единую популяцию. Мы спросили, разделены ли сигналы, связанные с движением каждой руки, таким образом, чтобы сигналы, относящиеся к одной руке, естественным образом не воздействовали на другую руку. Мы применили анализ основных компонентов (PCA) к ответу популяции (усредненная по пробам скорость стрельбы каждой единицы) в течение средних циклов каждого поведенческого состояния. PCA применяли отдельно для условий, когда задача выполнялась левой, а не правой рукой. Например, при езде на велосипеде вперед, начиная сверху, PCA применялся один раз при езде на велосипеде левой рукой (получая подпространство «левой руки») и еще раз при езде на велосипеде правой рукой (получая подпространство «правой руки»).
Важно отметить, что в обоих случаях PCA учитывал ответы одной и той же объединенной популяции нейронов (все зарегистрированные нейроны в обоих полушариях). Ключевой вопрос заключался в том, в какой степени левостороннее подпространство, построенное без привязки к деятельности при выполнении задания правой рукой, улавливало или не улавливало эту деятельность (и, наоборот, для праворукого подпространства). Активность в состоянии удержания использовалась для проверки того, насколько хорошо подпространства левой и правой руки фиксируют ответы, когда каждая рука выполняет задание. Например, на рис. 9А используется подпространство левой руки, основанное на реакциях, когда левая рука выполняла определенное условие (цикл вперед, начиная сверху). В этом же пространстве зафиксирована устойчивая реакция, когда левая рука выполняла условие удержания (91 105, синяя траектория 91 106; циклическое движение вперед, начиная снизу), но не когда правая рука выполняла то же самое состояние удержания (9).
1105 красный траектория). Мы количественно оценили этот эффект, рассчитав кумулятивную дисперсию, объясняемую пространством левого плеча, как функцию количества используемых основных компонентов. Объясненная кумулятивная дисперсия быстро росла (как и ожидалось) при рассмотрении условий удерживания, выполняемых левой рукой (рис. 9B, 91 105 синих 91 106 кривых), но не при рассмотрении условий удерживания, выполняемых правой рукой (91 105 красных 91 106 кривых). Действия, связанные с левой и правой рукой, лежат в ортогональных подпространствах.
( A ) Проекция активности населения на первые два измерения левого подпространства. Подпространство левой руки было найдено на основе данных одного условия (вперед, начало сверху), выполняемого левой рукой. Это же подпространство зафиксировало ответы во время другого условия (вперед, снизу-старт), выполняемого левой рукой (синяя траектория), но не для условия (вперед, сверху-старт), выполняемого правой рукой (красная траектория).
https://doi.org/10.7554/eLife.46159.011 При анализе учитываются данные средних циклов и все зарегистрированные нейроны обоих полушарий. Данные для обезьяны E. ( B ) Кумулятивная процентная дисперсия, объясненная подпространством левой руки, для ответов, зарегистрированных, когда левая (синяя) или правая (красная) руки выполняли задание. Объясненная дисперсия всегда вычислялась для исключенных условий (условия, которые не использовались для нахождения пробела). Анализ проводился с каждым условием, служащим в качестве условия обучения (используемого для нахождения пространства) один раз. Таких условий было четыре (два направления по двум начальным позициям), что давало четыре (в значительной степени перекрывающихся) трассы каждого цвета. Данные для обезьяны E. ( C ) Величина вклада в левое подпространство единиц правого (ведущего) и левого (не ведущего) полушарий. Сдвиг вправо указывал бы на больший вклад этого полушария. Медианы распределения обозначены красными и синими точками. Распределение было основано на абсолютном значении весов, содержащихся в пяти верхних ПК (после чего полученная процентная дисперсия в значительной степени стабилизировалась). Гистограммы объединяются для всех поведенческих состояний, поэтому каждый нейрон вносит 4 состояния x 5 ПК = 20 весовых коэффициентов. ( D–F ) Как и в A–C, для подпространства «правая рука». Обратите внимание, что проецируемые данные такие же, как и на панели А, отличается только подпространство. ( G–L ) Как в A-F, для Monkey F. Аналогичные результаты наблюдались для правой руки (рис. 9D, E) и для другой обезьяны (рис. 9G, H, J, K). Всегда было так, что относительно небольшая дисперсия регистрировалась, когда активность для правильных условий выполнения проецировалась на пространство левой руки, и наоборот. В среднем по всем таким условиям пять основных компонентов объясняли только 7 ± 1% (обезьяна E) и 2 ± 0,5% (обезьяна F) дисперсии ответа (среднее значение ± стандартное значение, рассчитанное для восьми условий).
Это контрастировало со значительной дисперсией, зафиксированной в условиях вытянутой руки, выполненной той же рукой, на которой тренировался свободный ход: 65 ± 3% (обезьяна E) и 69± 2% (обезьяна F). Таким образом, нейронные реакции, связанные с двумя руками, занимают почти ортогональные подпространства. Измерения, которые надежно фиксируют активность, когда одна рука выполняет задачу, не продолжают делать это, когда другая рука выполняет задачу. Это было верно как при анализе единой популяции (рис. 9), так и при анализе каждого полушария по отдельности (рис. 9 — приложение к рисунку 1,2).
Ортогональные подпространства могут возникать тривиально, если нейроны делятся на две группы: нейроны, которые реагируют только тогда, когда правая рука выполняет задание, и нейроны, которые реагируют, только когда задание выполняет левая рука. Однако, как показано выше (рис. 4В), большинство нейронов реагировало, когда задача выполнялась либо левой, либо правой рукой, и лишь с умеренной тенденцией к более сильной реакции при использовании контралатеральной руки.
В соответствии с этим, в обоих подпространствах участвовали нейроны обоих полушарий (рис. 9).C, I, F, L) с несколько большим вкладом от управляющей коры (например, правое полушарие для подпространства левой руки), как и ожидалось, с учетом немного большей реакции отдельных нейронов в управляющей коре. Нейронные траектории в одиночных испытаниях
Верно ли также, что однократное действие избегает подпространства, связанного с неработающей рукой? Предположительно, это подпространство должно быть занято хотя бы слабо при единичных испытаниях; неэффективная рука иногда двигается (хотя бы очень незначительно). Ключевой вопрос заключается в том, является ли занятие более чем скромным, что возможно, если усредненные данные испытаний нерепрезентативны по сравнению с тем, что происходит в отдельных испытаниях. Чтобы задать вопрос, являются ли репрезентативными усредненные по пробам частоты срабатывания, мы сначала рассмотрели вариабельность между испытаниями на уровне отдельных нейронов.
Как правило, исследования показывают, что фактор Фано (отношение дисперсии количества импульсов к среднему значению) в коре приблизительно равен единице, когда животное полностью вовлечено в задачу и последовательно выполняет ее (Churchland et al., 2010b). Наличие дополнительной изменчивости «сетевого уровня» от испытания к испытанию увеличивает коэффициент Фано выше единицы (Churchland and Abbott, 2012; Litwin-Kumar and Doiron, 2012; Shadlen and Newsome, 19).98). Во время езды на велосипеде фактор Фано был меньше или равен единице (0,99 и 0,69 для обезьян E и F, усредненные по условиям), что свидетельствует о значительной изменчивости на уровне сети. Затем мы изучили вариабельность отдельных испытаний на уровне популяции. Мы использовали недавно разработанный метод: латентный факторный анализ с помощью динамических систем (LFADS) (Pandarinath et al., 2018). LFADS использует скрытую нелинейную динамику для получения частоты возбуждения в одной попытке, которая объясняет наблюдаемые всплески.
Не гарантируется, что LFAD идентифицирует все формы изменчивости между испытаниями, но было продемонстрировано, что он успешно выводит значимые вариации активности в одном испытании при различных обстоятельствах. Мы использовали LFADS, чтобы определить частоту возбуждения в одной попытке, используя сеансы, в которых одновременно регистрировалось разумное количество нейронов (20–41). Затем оценочные частоты стрельбы в одной попытке были проецированы на подпространства левой и правой руки (рассчитаны на основе усредненной частоты испытаний). Траектории одной попытки имели умеренную вариабельность, но, тем не менее, в пространстве левой руки (рис. 10A, B, E, F) наблюдалась значительная дисперсия условий выполнения левой рукой (9).1105 синий ), но не для правой руки ( красный ). Аналогичные результаты были получены для правого подпространства (рис. 10C, D, G, H). Таким образом, разделение сигналов, связанных с левой и правой рукой, на разные подпространства присутствовало на уровне одной попытки. Разделение, возможно, было немного слабее (например, на рисунке 10C синяя кривая имеет незначительную дисперсию), чем при анализе усредненных по испытанию показателей, но это ожидается при анализе меньшего количества нейронов (рисунок 10 — дополнение к рисунку 1). Активность левой и правой руки в отдельных испытаниях.
( A ) Аналогичен анализу, показанному на рисунке 9A, но с прогнозированием показателей однократной стрельбы, полученным с использованием LFADS. Каждая трасса представляет собой проекцию ответа популяции в течение одного цикла одного испытания (показаны как верхние, так и нижние начальные условия). Данные относятся к одному сеансу, выполненному обезьяной Е. ( B ) Кумулятивная доля дисперсии, объясненная подпространством левой руки, для однократных ответов, зарегистрированных, когда левая (синяя) или правая (красная) руки выполняли задание.
https://doi.org/10.7554/eLife.46159.014 Каждая сессия записи вносит четыре дорожки: по одной для каждого направления вращения педалей и исполняющей руки. Темная кривая показывает среднее значение. Доля дисперсии была нормализована, чтобы всегда быть единицей для 35 ПК (что соответствует максимальному размеру популяции во всех включенных наборах данных для этой обезьяны), чтобы можно было сравнивать сеансы с разным количеством изоляций. ( C ) Тот же пример испытаний, показанный на A, спроецированный на первые два измерения подпространства правой руки. ( D ) Как в B, для кумулятивной процентной дисперсии, объясненной правым подпространством. ( E–H ) Как и в AD, для Monkey F. Линейные декодеры естественным образом разделяют сигналы, относящиеся к двум плечам
Разделение активности на ортогональные подпространства может позволить нисходящему управлению одной рукой естественным образом игнорировать сигналы, относящиеся к другой руке.
Чтобы проверить правдоподобность этой гипотезы, мы обучили линейные декодеры прогнозировать мышечную активность для данной руки на основе активности всей нервной популяции в обоих полушариях. Декодер обучался, используя только те условия, в которых эта рука выполняла задачу. Например, декодер был обучен предсказывать мышечную активность правой руки, в то время как правая рука выполняла задачу. В этой ситуации декодеры работали хорошо, прогнозируя медиану 91% (Обезьяна E) и 93% (Обезьяна F) дисперсии в условиях ожидания. Примеры прогнозируемой мышечной активности (фиг. 11A,G, оранжевых дорожек вверху ) показаны для одной мышцы у каждой обезьяны. Декодеры использовали активность в обоих полушариях (рис. 11E, K), как и ожидалось, учитывая, что нейроны в обоих полушариях активны и предсказывают мышечную активность (как описано выше). Декодеры мышечной активности игнорируют активность, связанную с «неправильной» рукой.
( A ) Фактическая (черный цвет) и прогнозируемая активность длинной головки трицепса, зарегистрированная с правой руки обезьяны E. Верхние следы: активность, когда правая рука выполняла задание. Нижние следы: активность во время выполнения задачи левой рукой. Оранжевые следы: прогнозы декодера, обученного только для условий выполнения правой руки (декодер «только для обучения»). Зеленые трассы: прогнозы декодера, обученного на подмножестве условий выполнения как левой, так и правой руки (декодер «обучения обоих»). Как верхний, так и нижний ответы получены из состояния ожидания, на котором декодеры не были обучены. ( B ) Фактическая и прогнозируемая модуляция мышечной активности, когда задача выполняется с помощью руки, содержащей эту мышцу (перф.), или другой руки (нерабочая). Каждая тонкая линия отображает модуляцию для одной мышцы в одном состоянии (например, дельтовидная мышца для велотренажера вперед с верхним стартом).
https://doi.org/10.7554/eLife.46159.016 Толстые линии показывают медианы. Три подпанели соответствуют эмпирической мышечной активности, декодеру, выполняющему только тренировки (обучались только условия, когда задача выполнялась рукой, содержащей мышцу), и декодеру обучения обоих (обучались в условиях обоих типов). Модуляцию всегда измеряли для прогнозирования мышечной активности в условиях удержания, для которых декодеры должны были обобщать. Данные для обезьяны E. ( C ) Те же данные, что и в B, но в виде графика распределения. На каждой гистограмме показано распределение эмпирической (черный) или прогнозируемой (оранжевый и зеленый) мышечной активности по всем мышцам/состояниям. Распределения наносятся отдельно, когда расшифрованная мышца находилась в работающей (вверху) или неработающей руке (внизу), чтобы облегчить сравнение в каждой ситуации. ( D ) Распределение индексов предпочтения руки для нейронов (красный цвет), мышц (серый цвет), прогнозы декодера, обученного только на работающих условиях руки (оранжевый), и прогнозы декодера, обученного как на работающей, так и на неработающей руке условия (зеленый). Данные относятся к обезьяне E. Красная и серая гистограммы немного отличаются от гистограмм на рис. 4A–B, поскольку они рассчитаны для каждого условия, учитывая, что настоящий анализ фокусируется на производительности обобщения для исключенных условий. ( E ) Распределение значений веса декодера (вклад каждого нейрона в декодирование мышечной активности) для нейронов ведущей коры (синие) и не ведущей коры (красные). Веса предназначены только для декодеров, выполняющих поезда. Точками показаны медианы распределения. ( F ) Как в E, для обучения обоих декодеров. ( G–L ) Как в A-F, для Monkey F. Ключевой вопрос касается того, что происходит, когда декодеры просят обобщить условия, при которых другая рука выполняла задачу. Например, может ли декодер, обученный предсказывать активность правого трицепса, когда задание выполняет правая рука, успешно предсказывать небольшую модуляцию трицепса, когда задание выполняет левая рука? Это строгий тест на обобщение как потому, что он включает условия, сильно отличающиеся от условий обучения, так и потому, что нейроны, на которых основано декодирование, активны независимо от используемой руки.
Декодеры точно обобщили и правильно предсказали небольшую модуляцию мышечной активности, когда рука не использовалась для выполнения задачи (рис. 11A, G, оранжевых кривых на нижних подпанелях ). Чтобы обеспечить количественную оценку, мы рассчитали степень модуляции (стандартное отклонение активности с течением времени) фактической и декодированной мышечной активности. Действительная мышечная модуляция снижалась (как и ожидалось), когда соответствующая рука менялась с рабочей на неработающую (рис. 11B,H, , черный 9).1106). Декодированная мышечная модуляция претерпела аналогичное снижение ( оранжевый ). Снижение расшифрованной мышечной активности было значительным для обеих обезьян (обезьяна E: p = 1,8 × 10–17, n = 96; обезьяна F: p = 1,8 × 10–17, n = 96; критерий знакового ранга Вилкоксона для мышц и условия).
Мы дополнительно задокументировали этот эффект, вычислив распределения (рис. 11C,I) фактической ( серый ) и прогнозируемой ( оранжевый ) мышечной модуляции.
Модуляция была сильной, когда соответствующая рука (рука, содержащая рассматриваемую мышцу) была работающей рукой (9).1105 верхние подпанели ). Модуляция была намного слабее, когда эта рука была неработающей ( нижние подпанели ). Примечательно, что это произошло для декодированной мышечной активности, даже несмотря на то, что декодер не был обучен этим неработающим условиям. Способность декодера игнорировать активность, связанную с «неправильной» рукой, наследуется от ортогональности подпространств правой и левой руки. При обучении с использованием условий правой руки декодеры естественным образом используют измерения в подпространстве правой руки. Поскольку эти измерения почти не заняты, когда левая рука выполняет задачу, декодирование показывает минимальную модуляцию. Благодаря этим свойствам декодеры естественным образом производят прогнозируемую мышечную активность с положительными индексами предпочтения рук (рис. 11D, J, 9).1105 оранжевых гистограмм ). Эти распределения сдвинуты вправо по сравнению с распределениями нейронной активности, на которых основывалось декодирование ( красная гистограмма) .
Таким образом, структура активности населения гарантирует, что декодер, обученный извлекать активность, относящуюся к одному плечу, будет естественным образом игнорировать активность, связанную с другим плечом. Тем не менее, в то время как декодеры, как правило, естественным образом игнорировали активность, связанную с «неправильной» рукой, эта функция была несовершенной: небольшое количество остаточной модуляции все еще присутствовало. Рисунок 11A, G нижняя подпанель ). В целом прогнозируемая модуляция ЭМГ в неработающей руке (рис. 11C,I, распределение 91 105 оранжевых 91 106 в нижней части панели) была значительно выше, чем истинная модуляция ЭМГ (обезьяна E: p = 0,0013; обезьяна F: p < 0,001; знак Вилкоксона). ранговый тест по 96 мышцам и состояниям). Это привело к тому, что индексы предпочтения рук для декодера были меньше, чем у мышц (рис. 11D,J, распределения оранжевых лежат слева от распределений черных ). Является ли это фундаментальным ограничением линейного декодера или просто отражает сложность обобщения на совсем другую ситуацию?
Чтобы решить вторую возможность, мы обучили декодеры, используя как условия выполнения левой рукой, так и условия выполнения правой руки (при проверке обобщения с использованием исключенных условий каждого типа).
Эта стратегия «обучение обоих» привела к дальнейшему улучшению способности игнорировать нейронную активность, связанную с «неправильной» рукой (рис. 11, 91–105 зеленых кривых и распределений 91–106). Например, зеленые распределения на рисунке 11C,I сдвинуты влево по сравнению с оранжевыми распределениями. (p=0,0013 и p<0,001 для двух обезьян, знаковый ранговый критерий Уилкоксона для 96 мышц и состояний). Тем не менее, расшифрованная мышечная активность все еще была модулирована немного больше, чем наблюдалось эмпирически (обезьяна E: p = 0,01, обезьяна F: p<0,001). В результате оба декодера для обучения предсказывали мышечную активность с индексами предпочтения руки намного выше, чем у нейронной активности, но не такими высокими, как у самих мышц (рис. 11D, J). Эта оставшаяся модуляция декодирования, вероятно, отражает использование линейного приближения к нелинейной функции, связывающей нейронную и мышечную активность, и/или неполную выборку нейронной популяции. Приведенные выше результаты показывают, что даже простые линейные декодеры могут в значительной степени игнорировать активность, связанную с неправильной рукой.
Эту способность можно улучшить, непосредственно обучая этому декодеры. Тем не менее, даже без такой подготовки декодеры успешно обобщают и предсказывают небольшую мышечную активность, когда соответствующая рука не выполняет задачу. Таким образом, нет никакого парадокса в наличии сильной нервной активности в обоих полушариях, несмотря на отсутствие мышечной активности в неработающей руке. Обсуждение
Нейронные сигналы, связанные с движениями правой и левой руки, были смешаны между полушариями; когда двигалась одна рука, модулировались нейроны обоих полушарий. Сигналы также были смешаны между нейронами; большинство нейронов реагировало, когда и их ведомая, и неведомая рука выполняли задачу. Отдельные нейроны реагировали очень по-разному в зависимости от того, какая рука двигалась. Однако на уровне населения оба полушария содержали одинаковую информацию. Удивительно, но мы не обнаружили сигналов, которые сильно присутствовали в управляющей коре, но отсутствовали в неуправляющей коре.
Это было верно даже для мышечных сигналов, которые можно было одинаково хорошо декодировать в любом полушарии. Несмотря на это смешение, сигналы, соответствующие двум плечам, были естественным образом разделены на уровне нейронной популяции. Активность, связанная с двумя рукавами, занимала почти ортогональные подпространства. В результате даже простые линейные декодеры могли считывать команды для одной руки, игнорируя команды для другой руки. Действительно, декодеры естественным образом игнорировали действия, связанные с «неправильной» рукой, даже не будучи обучены этому.
Разделение информации по измерениям является общей чертой активности коры головного мозга
Наши результаты вносят вклад во все более широкий набор исследований, сообщающих о том, что нейронная активность, связанная с различными вычислениями или параметрами задач, часто разделяется по нейронным измерениям, а не на уровне областей мозга или отдельных нейронов (Druckmann and Chklovskiy, 2012; Machens et al.
., 2010; Mante et al., 2013; Raposo et al., 2014). Например, во время достижения размеры, передающие подготовительную активность, ортогональны измерениям, передающим деятельность, связанную с движением в целом (Elsayed et al., 2016), и особенно сигналы, связанные с мышцами (Kaufman et al., 2014). Таким образом, предлагается, чтобы подготовительные размеры были «нулевыми мышцами» — без чистого драйва к нижестоящим мышцам. Нулевые размеры мышц также ожидаются во время движения; они важны для сетевых моделей формирования охвата (Churchland et al., 2012; Sussillo et al., 2015) и для гипотезы о том, что популяционная активность структурирована таким образом, чтобы избежать запутывания траекторий (Russo et al., 2018). Наши результаты согласуются с гипотезой о том, что нервная активность, связанная с правой рукой, падает в размерах, которые являются нулевыми по отношению к левой руке, и наоборот. Эта гипотеза делает предположение, которое можно проверить с помощью одновременных ЭМГ и крупномасштабных нейронных записей: небольшие сокращения в неработающей руке должны коррелировать с небольшими отклонениями в соответствующем подпространстве. Хотя мы еще не знаем механизмов, с помощью которых устанавливается ортогональность различных сигналов, следует отметить, что два случайно ориентированных измерения практически гарантированно почти ортогональны в многомерном пространстве. Таким образом, если механизм обучения гарантирует, что нейроны имеют случайные предпочтения для двух ветвей (случайная «смешанная» избирательность), ортогональность может быть достигнута без явного принуждения.
Учитывая, что разделение сигналов на основе подпространства, по-видимому, является обычной нейронной стратегией, стоит подчеркнуть два момента, касающихся интерпретации. Во-первых, веса, определяющие каждое подпространство, не следует рассматривать как отображение непосредственно на анатомию. Например, нейрон может нести мышечно-подобные сигналы и, таким образом, вносить свой вклад в «мышечно-потенциальное» подпространство, даже если он не является выходным нейроном. Такой нейрон может управлять выходными нейронами, может передавать сигналы, связанные с мышцами, в области, отличные от спинного мозга, или может просто участвовать в вычислениях, которые генерируют сигналы, связанные с мышцами.
В настоящем исследовании нейрон может вносить вклад в подпространство «правой руки» независимо от того, в каком полушарии он находится. И наоборот, выходной нейрон может содержать вклады как от мышечно-потенциальных, так и от мышечно-нулевых измерений. Это естественно происходит в сетевых моделях и возможно до тех пор, пока немышечные сигналы усредняются по выходным нейронам (Kaufman et al., 2014). Например, было показано, что некоторые корково-спинномозговые нейроны имеют «зеркальные» ответы на наблюдаемые действия, несмотря на отсутствие соответствующей мышечной активности (Красков и др., 2014). Во-вторых, ортогональность подпространств не означает, что релевантные сигналы не взаимодействуют — обычно предполагается, что они взаимодействуют. Предполагается, что подготовительная деятельность задает динамику, связанную с движением (Afshar et al., 2011; Churchland et al., 2006; Churchland et al., 2010a), а динамика периода движения предлагается охватывать параметры отсутствия мышц и мышечного потенциала.
(Черчленд и др., 2012; Черчленд и Шеной, 2007). Смешивание сигналов внутри данного нейрона открывает возможности для таких взаимодействий. В случае двух рук возможность взаимодействия может иметь отношение к бимануальной координации. Сравнение с предыдущими исследованиями латерализации M1
Наш вывод о том, что отдельные нейроны часто реагируют во время движений любой руки, в целом согласуется с предыдущими исследованиями записи на приматах. В большинстве этих исследований описывается смешение ответов правой и левой руки в активности отдельных нейронов M1 (Cisek et al., 2003; Donchin et al., 2002; Donchin et al., 1998; Kermadi et al., 1998). Стейнберг и др., 2002). Однако в некоторых исследованиях, в которых использовались небольшие движения пальцев, сообщалось о гораздо меньшем проценте ипсилатеральных нервных реакций (Aizawa et al., 19).90; Tanji et al., 1988), предполагая большую латерализацию для вычислений, связанных с руками, по сравнению с вычислениями, связанными с руками.
В соответствии с этой интерпретацией область рук M1 имеет меньше мозолистых соединений и меньше ипсилатеральных выступов (Jenny, 1979; Jones and Wise, 1977; Rouiller et al., 1994). Однако эту интерпретацию следует считать предварительной; Исследования фМРТ обнаруживают значительную двустороннюю настройку M1 во время движения пальцев (Berlot et al., 2019; Diedrichsen et al., 2013; Verstynen et al., 2005). Реакции отдельных нейронов сильно зависят от конечностей
Два предыдущих исследования явно сравнивали свойства нервной реакции во время одноручных движений каждой руки. Цисек и др. (2003) обнаружили, что нейроны в M1 имеют предпочтительные направления, зависящие от конечностей, однако Steinberg et al. (2002) сообщили о сохранении предпочтительных направлений. В то же время Steinberg и соавт. обнаружили, что направления досягаемости левой и правой руки могут быть независимо декодированы отдельными пулами нейронов, что предполагает некоторую степень зависимости от конечностей.
Настоящие результаты показывают, что нервные реакции сильно зависят от конечностей. В отличие от предыдущих исследований, наши сравнения свойств ответов касались их временной структуры, а не предпочтительного направления как такового, и использовали совсем другую задачу. Тем не менее, наши ответы согласуются как с Cisek et al. (2003) и Heming et al. (2019), которые обнаружили, что предпочтительные направления в ответ на нагрузку были независимыми для двух конечностей. Кроме того, хотя мономануальные сравнения не были центральной темой Donchin et al. (2002 г.); Дончин и др. (1998), примеры единиц, показанные в этом исследовании, не подтверждают гипотезу о предпочтительном направлении, одинаковом для всех конечностей. Таким образом, оказывается, что ответы M1 сильно зависят от конечностей в различных контекстах.
По оценке фМРТ отдельные воксели в моторной коре сохранили «предпочтение пальцев» (мизинец, средний или большой палец) на конечностях (Diedrichsen et al., 2013).
Это может показаться противоречащим нынешним результатам, когда ответы не были сохранены. Вероятное решение состоит в том, что соседние нейроны имеют ответы, связанные с аналогичным набором суставов для обеих рук, но без сохранения настройки на особенности движения. Например, нейрон может реагировать при сгибании правого мизинца и разгибании левого мизинца. Возможные причины ипсилатеральной моторной активности коры
Существует несколько причин, по которым моторная кора может быть активна, когда неведущая рука выполняет задачу. Прямой возможностью является то, что моторная кора использует абстрактное независимое от конечностей представление движения. Однако эта гипотеза маловероятна, учитывая сильно зависящий от конечностей характер ответов. В качестве альтернативы две коры могут обрабатывать разную, но дополняющую информацию. Эта гипотеза также маловероятна; мы не обнаружили сильных сигналов, которые присутствовали в управляющей коре, но отсутствовали в неуправляющей коре.
Также возможно, что активность ипсилатеральнее подвижного плеча может быть связана с неперекрещенными нисходящими связями (Kuypers, 1981; Rosenzweig et al., 2009). Например, активность правой моторной коры могла бы управлять через непересекающиеся связи мышечной активностью правой руки, когда эта рука выполняет задачу. Тем не менее, в предыдущих исследованиях было обнаружено мало доказательств надежной связи между M1 и активацией ипсилатеральных мышц. Внутрикорковая микростимуляция легко вызывает реакцию контралатеральной мышцы (Kwan et al., 19).78; Sessle and Wiesendanger, 1982), но очень редко вызывает реакцию ипсилатеральных мышц (Aizawa et al., 1990). Внутриклеточные записи мотонейронов не выявляют моносинаптических вызванных потенциалов от стимуляции ипсилатерального корково-спинномозгового тракта, а эффекты ЭМГ, запускаемые спайками, присутствуют только для контралатеральных мышц (Soteropoulos et al., 2011). По этим причинам мы подозреваем, что непересекающиеся проекции вряд ли могут быть основной причиной активности неуправляющей моторной коры.
Другая возможность состоит в том, что активность в неуправляемой коре является результатом эфферентной копии сигналов, генерируемых и используемых управляющей корой. Эфферентная копия может помочь в координации между конечностями. Хотя в данной задаче в этом не было необходимости (одна рука просто оставалась неподвижной), возможно, сигналы эфферентного копирования передаются по умолчанию и игнорируются, если они не нужны. Наши результаты согласуются с этой возможностью и утверждают, что если она верна, эфферентная копия должна быть вполне полной. То есть управляющая кора должна передавать большую часть генерируемых ею сигналов, а не (например) только выходные сигналы.
Независимо от роли в онлайн-координации, эфферентная копия может позволить неуправляющей коре «наблюдать» за вычислениями в управляющей коре, способствуя межручному обмену навыками (Westler et al., 2014). Если это так, то ответы в неуправляемой коре можно рассматривать почти так же, как зеркальные ответы.
Зеркальные нейроны обнаружены во многих двигательных областях (Fogassi et al., 2005; Rizzolatti et al., 1996), включая первичную моторную кору (Dushanova and Donoghue, 2010; Vigneswaran et al., 2013). Даже идентифицированные нейроны пирамидного пути могут проявлять зеркальные свойства (Красков и др., 2014). Эти результаты доказывают, что общим свойством нейронов в двигательных областях является реакция во время движений, в которых они не участвуют напрямую, возможно, в целях обучения. Последняя возможность заключается в том, что вычисления моторной коры распределены по обоим полушариям (Li et al., 2016). В крайнем случае, нейроны неуправляющей коры можно просто рассматривать так же, как мы рассматриваем большинство нейронов управляющей коры; они могут участвовать в вычислениях, даже если они представляют собой один или несколько синапсов корково-спинномозговых нейронов, которые будут передавать выходные данные. Эта гипотеза привлекательна, потому что она может объяснить тот факт, что все большие сигналы распределяются между полушариями.
В более общем смысле, если случайно выбранный нейрон из неведущей коры имеет ответы, которые почти неотличимы от нейрона, выбранного из движущей коры, возможно, нашим исходным предположением должно быть то, что они участвуют в одних и тех же вычислениях. Остается неясным, можно ли согласовать эту гипотезу с тем фактом, что инактивация моторной коры главным образом затрагивает контралатеральные конечности (Glees and Cole, 19).50; Лю и Руйе, 1999 г.; Пассингем и др., 1983). Если оба полушария участвуют в управлении обоими руками, можно было бы ожидать более двустороннего дефицита. Возможное решение состоит в том, что сеть достаточно надежна, чтобы она могла функционировать, когда многие нейроны неактивны, пока выходные нейроны все еще могут передавать необходимые команды. Ипсилатеральные сигналы руки в других областях мозга
Области коры, подкорковые области и спинной мозг участвуют в управлении ловкими движениями. Действительно, другие исследования, сравнивающие контралатеральные и ипсилатеральные движения, показали, что не только M1, но и дорсальная премоторная кора (Cisek et al.
, 2003; Kermadi et al., 2000; Tanji et al., 1988), вентральная премоторная кора (Michaels, Scherberger, 2018), дополнительная моторная зона (Donchin et al., 2002; Грибова и др., 2002; Казенников и др., 1999; Kermadi et al., 2000; Kermadi et al. , 1998; Tanji et al., 1988), поясная моторная зона (Kermadi et al., 2000) и задняя теменная кора (Kermadi et al., 2000) содержат нейроны, которые реагируют на движения ипсилатеральной руки. Более того, в стволе и спинном мозге есть цепи, которые специально поддерживают генерацию скоординированных ритмичных движений, таких как передвижение (Duysens and Van de Crommert, 19).98). Другие области мозга, такие как передняя интратеменная область, кодируют параметры движения в значительной степени независимо от конечностей (Michaels and Scherberger, 2018), что позволяет предположить, что связь моторных и зрительно-моторных областей с ипсилатеральными движениями может варьироваться в зависимости от их роли в расчет двигателя. В целом движение генерируется широкой взаимосвязанной сетью отделов головного и спинного мозга. Мы сосредоточились на M1, потому что это корковая область, которая, основываясь на результатах анатомии и микростимуляции, скорее всего, имеет латерализованное представление движения. Тем не менее, даже в M1 нам не удалось найти доказательств сильно латерализованной активности. Материалы и методы
Поведение
Запросить подробный протоколВсе процедуры для животных были одобрены Институциональным комитетом по уходу и использованию животных Колумбийского университета. Данные были собраны у двух самцов обезьян ( Macaca mulatta ), когда они выполняли задание на велосипеде за вознаграждение соком. Эксперименты контролировались, а данные собирались под контролем компьютера (целевая машина в реальном времени: Speedgoat, Liebfeld, Швейцария). Во время выполнения задания каждая обезьяна сидела в специальном кресле для приматов, а голова фиксировалась с помощью хирургического имплантата. Видеомонитор отображал виртуальную среду, в которой перемещалась обезьяна.
Каждой рукой обезьяна сжимала специальную педаль. Руки были слегка зафиксированы тканевой лентой, чтобы держать их в постоянном положении на педалях. Сама педаль также была разработана для обеспечения постоянного положения рук и включала ручку и скобу, которые уменьшали движение запястья. Педали вращали коленчатый вал, прикрепленный к двигателю (Applied Motion Products, Watsonville, CA). Вращающийся энкодер внутри двигателя сообщал о положении с точностью 1/8000 цикла. Информация об угловом положении и его производных использовалась системой реального времени для подачи силовых команд, которые создавали виртуальную массу и вязкость. Это придало педали естественное ощущение и обеспечило достаточное сопротивление, чтобы обеспечить надежные и повторяемые модели мышечной активности. Обезьяны крутили педаль, чтобы контролировать свое положение в виртуальной среде (рис. 1А). Угловое положение педали было сопоставлено непосредственно с линейным положением в виртуальном мире. Задача заключалась в том, чтобы останавливаться на мишенях (неподвижные белые квадраты на земле), чтобы получить награду в виде сока.
В каждом испытании обезьяна продвигалась от начальной цели к конечной. Соотношение между положением в мире и положением педали было установлено таким образом, что цели находились на расстоянии ровно семи циклов друг от друга. Обезьяны выполняли блоки из двадцати последовательных испытаний. Для данного блока одна рука была «работающей рукой», а другая — «неработающей рукой». Мировое положение определялось педалью, которой манипулировала рабочая рука. Другая педаль должна была оставаться в окне узкого положения в нижней части цикла (см. ниже). После завершения блока следующий блок сигнализировался 5-секундным нежным «жужжанием», подаваемым на одну педаль (посредством двигателя), указывающим на новую исполнительную руку. Блоки были представлены в рандомизированном порядке (рис. 1C). Обезьяны также выполняли блоки, в которых обе руки вращались вместе (бимануальный вариант задачи), которые не анализируются в этом исследовании. В начале каждого испытания первоначальная цель появлялась на один-два цикла перед обезьяной.
Обезьяна подошла к этой цели и «приобрела» ее, остановившись на ней. Окно приемлемости составляло ± 0,15 цикла для Обезьяны E и ± 0,01 цикла для Обезьяны F. Удержание цели также требовало оставаться ниже жесткого порога скорости: 0,01 цикла / с для Обезьяны E и 0,0125 цикла / с для Обезьяны F. После удержания цели на 1000-2000 мс (обезьяна E) или 600-1000 мс (обезьяна F) начальная цель исчезала, а конечная цель появлялась на семь циклов впереди текущей позиции. Затем обезьяна направилась к этой последней цели. Чтобы стимулировать езду на велосипеде с быстрой и постоянной скоростью, мы ввели минимальный порог скорости и максимальную продолжительность езды на велосипеде. Как только конечная цель была достигнута, обезьяна оставалась внутри цели в течение 1000–1600 мс, чтобы получить вознаграждение в виде сока. Порог скорости был одинаковым для начальной и конечной целей. Для Monkey F, когда педаль была остановлена в пределах цели, двигатель создавал небольшие стабилизирующие силы, минимизируя мышечную активацию, необходимую для удержания педали в неподвижном состоянии. Во время проведения испытания педаль, связанная с неработающей рукой, должна была оставаться в пределах окна (±0,05 и ±0,07 циклов для обезьян E и F), центрированного в нижней части цикла. Движение за пределы этого окна привело к провалу испытания, за которым последовал короткий тайм-аут. Обезьяны заняли стереотипное положение внутри навязанного окна и мало отошли от этого положения. Для неэффективной руки диапазон положений, исследованных в рамках испытания, составил в среднем 0,006 цикла (обезьяна E) и 0,029 цикла (обезьяна F).
В каждом блоке из 20 испытаний было четыре поведенческих состояния (рис. 1С). Каждое условие использовало одно из двух начальных положений педали (начальное верхнее и нижнее начальное) и одно из двух направлений вращения (вперед и назад). В условиях старта сверху начальная цель располагалась таким образом, чтобы положение педали, необходимое для достижения этой цели, находилось в самом начале цикла. В условиях старта снизу необходимое положение педали было в самом низу.
В обоих случаях конечная цель находилась ровно в семи циклах и, таким образом, была достигнута при одном и том же положении педали. В условиях езды на велосипеде вперед движение вперед в виртуальной среде производилось за счет движения вперед, когда рука отходила от тела в верхней части цикла (движение, аналогичное движению стопы при вращении педалей на велосипеде). В условиях обратного цикла отображение было обратным. Направление движения определялось цветом ландшафта в виртуальной среде: зеленый — вперед, оранжевый — назад. Каждая из четырех комбинаций исходного положения и направления езды выполнялась в подблоке из пяти попыток. Порядок субблоков был идентичен для каждого блока (рис. 1C). Обезьяны были хорошо обучены этому заданию еще до начала экспериментов, поэтому их результаты оставались неизменными в течение нескольких дней. Для обезьяны E данные были собраны в течение 19 дней записи нейронов и 8 дней записи мышц. Для обезьяны F данные были собраны в течение 20 дней записи нейронов и 6 дней записи мышц.
Хирургические процедуры и нейронная запись
Запросить подробный протоколобезьяны были подвергнуты анестезии и имплантированы головные уборы в стерильных условиях. А 19Цилиндр диаметром мм (Crist Instruments) помещали над первичной моторной корой каждого полушария под контролем структурной МРТ, выполненной до операции. Череп остался неповрежденным под цилиндром, покрытым тонким слоем стоматологического акрила. Перед регистрацией обезьян подвергали анестезии и вручную просверливали трепанационное отверстие диаметром 3,5 мм через зубной акрил и череп, оставляя твердую мозговую оболочку нетронутой. В ходе эксперимента трепанационные отверстия были открыты в разных местах (рис. 1F, G). Записи велись с разных глубин. Примечательно, что проникновения, которые вошли в передний M1 / задний PMd, могли давать записи в борозде M1. После записи трепанационные отверстия были закрыты стоматологическим акрилом, что позволило черепу зажить. После завершения нашего эксперимента Обезьяне Е сделали операцию по имплантации записывающего массива в левое полушарие в рамках другого исследования.
Во время этой операции мы подтвердили, что расположение фрезевых отверстий совпало с ожидаемыми ориентирами борозды. После вскрытия каждого трепанационного отверстия мы сначала регистрировали нейронную активность с помощью обычных одиночных электродов (компания Frederick Haer). Это позволило провести несколько дней внутрикортикальной микростимуляции (ICMS) и пальпации мышц, чтобы подтвердить, что записи были в области руки M1. Для левого полушария обезьяны F (рис. 1G) минимальные пороги ICMS в заднем и переднем трепанационных отверстиях составляли 10 мкА и 20 мкА соответственно. Для правого полушария минимальные пороги ICMS в двух задних трепанационных отверстиях составляли 10 мкА (медиально) и 20 мкА (латерально). Эти пороги согласуются с проникновениями, лежащими в первичной моторной коре проксимальных отделов рук, в соответствии с физическим расположением этих четырех трепанационных отверстий вблизи центральной борозды. Для более переднего трепанационного отверстия минимальный порог ICMS составлял 40 мкА, что соответствовало либо задней дорсальной премоторной коре (PMd), либо, возможно, передней первичной моторной коре.
Четвертое, более заднее трепанационное отверстие из правого полушария не было зарегистрировано из-за отсутствия ответа ICMS до 150 мкА; предполагалось, что это трепанационное отверстие находится над S1. Для левого полушария обезьяны E (рис. 1F) минимальные пороги ICMS в заднем и переднем трепанационных отверстиях составляли 30 мкА и 75 мкА соответственно. Эти пороги согласуются с первичной моторной корой и PMd, соответственно, в соответствии с их расположением. Для правого полушария обезьяны E минимальные пороги ICMS в заднем и переднем трепанационных отверстиях составляли 20 и 25 мкА. Эти пороги согласуются с первичной моторной корой, хотя более переднее трепанационное отверстие, вероятно, также содержало некоторые записи от PMd (учитывая его расположение).
Таким образом, пороги и анатомические локализации указывают на то, что большинство записей было сделано из первичной моторной коры. (Обратите внимание, что места на рис. 1F, G показывают поверхностные ориентиры — проникновения, которые вошли в PMd, могут попасть в первичную моторную кору ниже).
Однако меньшая часть записей почти наверняка была сделана из премоторной коры. Таким образом, для обеих обезьян ключевые анализы были повторены с использованием данных только по задним фрезевым отверстиям. Результаты были практически идентичными, подтверждая, что свойства, о которых мы сообщаем, присущи первичной моторной коре, а не возникают в результате контаминации записями из премоторной коры. Мы регистрировали нейронную активность с помощью 24-канальных V-зондов (Monkey E) или 32-канальных S-зондов (Monkey F) (Plexon Inc, Даллас, Техас). Мы опускали по одному зонду в каждое полушарие каждый день, удаляя зонд в конце этого сеанса. Зонды перемещали в разные места внутри каждого трепанационного отверстия в каждый день записи. Нейронные сигналы обрабатывались и записывались с использованием цифрового концентратора и 128-канального процессора нейронных сигналов (Blackrock Microsystems, Солт-Лейк-Сити, Юта). Пороговые пересечения для каждого канала регистрировались и сортировались в автономном режиме (Plexon Offline Sorter).
Изоляцию единиц оценивали на основе разделения сигналов в пространстве PCA, гистограмм интервалов между спайками и стабильности сигналов в течение сеанса. Анализы рассматривают стабильные, хорошо изолированные единичные и множественные выделения. Многоэлементная изоляция состояла из идентифицируемых спайков (то есть не просто пересечений порога) от двух (а иногда и более) нейронов, которые нельзя было с уверенностью отличить друг от друга. Все примеры скорострельности, показанные на рисунках, относятся к одиночным установкам. Мы зарегистрировали 263 единицы в левом полушарии и 270 единиц в правом полушарии для обезьяны Е (8-47 в день), и 338 единиц в левом полушарии и 279 единиц.ед. в правом полушарии у обезьяны F (12–64 в сутки). Записи ЭМГ
Запросить подробный протоколВ отдельные дни, вперемежку с нейронными записями, регистрировались внутримышечные ЭМГ-сигналы. Мы записывали данные по двуглавой мышце плеча (длинная и короткая головки), трехглавой мышце плеча (медиальная, длинная и латеральная головки), дельтовидной мышце (передняя, латеральная и задняя головки), широчайшей мышце спины, грудной, трапециевидной и плечелучевой мышцам.
Пары электродов с крючками вставляли примерно на 1 см в брюшную полость мышцы в начале каждого сеанса и удаляли в конце. На каждом сеансе производили от 1 до 3 записей ЭМГ на руку. Напряжения на электродах усиливали, фильтровали полосой пропускания (10–500 Гц) и оцифровывали с частотой 1000 Гц (Monkey E) или 30 000 Гц (Monkey F). Записи далее не рассматривались, если они содержали значительный артефакт движения или слабые сигналы. В автономном режиме записи ЭМГ подвергались фильтрации верхних частот на частоте 40 Гц, выпрямлению и сглаживанию с помощью гауссовой кривой 25 мс. Это дало измерение интенсивности в зависимости от времени, которое затем усреднялось по испытаниям. Пробное усреднение ЭМГ использовало ту же процедуру выравнивания, которая использовалась при вычислении пробной усредненной частоты срабатывания (см. ниже). Усредненная скорострельность
Запросить подробный протоколВремя спайков каждого нейрона в каждом испытании было преобразовано в частоту возбуждения путем свертки спайков с гауссовой кривой 25 мс.
Чтобы получить усредненную по испытаниям скорость стрельбы, мы сначала выровняли все испытания по общему времени: моменту, когда был завершен первый полупериод. Это прекрасно выровненное поведение между испытаниями в начале каждого испытания. Однако, поскольку испытания длились несколько секунд, небольшие различия в скорости езды на велосипеде могли накапливаться и вызывать значительные расхождения в поведении между испытаниями (рис. 2А). Возникающее в результате несоответствие показателей однократной попытки стрельбы (рис. 2B) приведет к нерепрезентативной средней частоте стрельбы (та же проблема повлияет на средние значения мышечной активности и кинематических переменных). Таким образом, временное выравнивание применялось к средним шести циклам, начиная с полупериода после начала движения (т. е. для испытаний с верхним стартом, в момент, когда педаль достигла низа) до полуцикла до окончания движения. Выравнивание выполнено в два этапа. Во-первых, мы временно преобразовали скорострельность в базовую позицию, а не во временную.
Например, рассмотрим rtrial(t), частоту срабатывания нейрона как функцию времени. Мы построили rtrial'(θ), где θ — угловое положение педали (от 0 до 12π, так как имеется шесть средних циклов). Поскольку все испытания теперь находятся в одной и той же области, их можно усреднить, чтобы получить r′¯(θ). Затем мы преобразовали обратно во временную область, чтобы получить r¯(t). Хотя в разных испытаниях и условиях наблюдалась небольшая вариабельность, обезьяны работали с частотой, близкой к 2 Гц. Таким образом, мы использовали приближение, согласно которому каждый цикл длился 500 мс, и линейно отображали позиции 0-12π на времена 0-3000 мс (6 циклов по 500 мс на цикл). Это сопоставление гарантирует, что заданное время всегда соответствует одному и тому же положению, помогая сравнивать разные условия и объединять нейроны разных сеансов в единую популяцию. Недостаток этого простого сопоставления в том, что поведение становится идеализированным; то есть вертикальное положение руки становится идеальной синусоидой после выравнивания. На практике этот недостаток был допустим; вертикальное положение рук уже во время средних циклов было близко к идеальной синусоиде. Эта процедура очень незначительно изменила временную базу отдельных испытаний, но сохранила надлежащее согласование между испытаниями (рис. 2D) и произвела усредненные по испытаниям оценки частоты стрельбы (рис. 2E), которые репрезентативны для того, что произошло в одиночных испытаниях. На рисунке 2D показаны времена всплесков, чтобы проиллюстрировать полезность выравнивания. Тем не менее, мы подчеркиваем, что временное выравнивание было выполнено после преобразования пиков в однократную частоту срабатывания с помощью свертки с гауссианой. Таким образом, выравнивание никогда не меняло скорость стрельбы, а только время, когда эта скорость была достигнута. Выравнивание применялось только при расчете средних проб, а не в случае анализа отдельных проб.
Обратите внимание, что невозможно выполнить выравнивание на основе изменения положения руки во времени, если положение не меняется.
Хотя мы могли бы оценить самый первый момент начала движения педали и последний момент ее движения, было более надежно и удобно просто начинать и заканчивать на первом и последнем полупериодах. (Поскольку первый и последний полупериоды длились всего несколько сотен миллисекунд, испытания в это время оставались в разумных пределах). В большинстве анализов скорострельности использовались средние четыре цикла (2-5), за исключением первого цикла и последних двух циклов. Этот анализ был сосредоточен на стационарной реакции, а не на реакциях, связанных с запуском, остановкой или удержанием. Это помогло интерпретации двумя способами. Во-первых, мышечная активность в неработающей руке была особенно слабой во время средних циклов (напротив, умеренная активность иногда наблюдалась при остановке). Сосредоточение внимания на средних циклах, таким образом, в значительной степени обходит стороной опасения, что активность ипсилатеральнее работающей руки связана с мышечной активностью неработающей руки.
Во-вторых, мы хотели сфокусировать ключевой анализ на ритмическом паттерне модуляции скорости стрельбы, а не на общих изменениях чистой скорости стрельбы при движении по сравнению с отсутствием движения. Например, при прогнозировании мышечной активности на основе нейронной активности относительно «легко» зафиксировать обычно повышенный уровень активности во время движения, что приводит к высоким значениям R2, даже если прогнозы не отражают модели активности цикл за циклом. Мы хотели избежать этого и считать прогнозы успешными только в том случае, если они учитывают аспекты ритмической реакции. Анализ отдельных нейронов
Запросить подробный протоколАнализы были выполнены на основе усредненной по результатам испытаний частоты возбуждения, рассчитанной, как описано выше, и учитывали средние четыре цикла (циклы 2–5) изменяющейся во времени реакции. Мы хотели сравнить для каждого нейрона силу модуляции при выполнении задачи управляемой и неуправляемой рукой.
Под модуляцией мы подразумеваем степень, в которой частота срабатывания нейрона варьировалась в пределах циклов, между циклами и/или между условиями (вперед или назад, а также начало сверху или начало снизу). Для каждого нейрона мы составили единый вектор скорости возбуждения, rdriven, объединив векторы частоты возбуждения в четырех условиях, когда ведомая рука выполняла задачу. Таким образом, rdriven имел размер ct, где c — количество условий, а t — количество раз в течение четырех средних циклов одного состояния. Мы определили Modulationdriven как стандартное отклонение rdriven, которое отражает степень, в которой средняя скорость срабатывания варьируется во времени и в зависимости от условий. Неуправляемая модуляция была рассчитана аналогичным образом на основе rнеуправляемой (объединенной скорости срабатывания в четырех условиях, когда неуправляемая рука выполняла задачу). Чтобы оценить, в какой степени нейрон сильнее модулируется при выполнении задачи управляемой рукой по сравнению с неуправляемой, мы рассчитали индекс предпочтения руки:
(1) Modulationdriven-Modulationnon-drivenModulationdriven+Modulationnon-driven
Этот индекс предпочтения плеча равен нулю, если нейрон одинаково модулируется независимо от используемого плеча, приближается к единице, если модуляция намного больше при использовании ведомого плеча, и приближается к отрицательному значению, если модуляция намного больше при использовании неприводной руки.
Чтобы сравнить паттерны ответов при выполнении задания с управляемой и неуправляемой рукой, для каждого нейрона мы рассчитали корреляцию Пирсона между rуправляемой и rнеуправляемой рукой. rdriven и rnon-driven были определены, как указано выше, за одним исключением. Мы хотели, чтобы настоящий анализ сосредоточился на моделях скорости стрельбы цикл за циклом во время каждого поведенческого состояния, а не на глобальных изменениях скорости стрельбы в зависимости от условий. Таким образом, мы центрировали среднее значение частоты срабатывания для каждого условия (так, чтобы среднее значение было равно нулю) до конкатенации. Это гарантировало, что паттерны скорости стрельбы не могли казаться одинаковыми, например, из-за сильных, но в остальном не связанных ответов во время циклического движения вперед для двух рук.
Чтобы определить, была ли корреляция между rdriven и rnon-driven больше, чем ожидалось случайно, мы выполнили случайное управление. Для каждого нейрона в нашей популяции мы заменили его rуправляемый и rнеуправляемый на rуправляемый от одного случайно выбранного нейрона и rнеуправляемый от другого случайно выбранного нейрона (с заменой).
Затем мы вычислили средний (по повторной выборке населения) коэффициент корреляции Пирсона между новыми r-управляемыми и r-неуправляемыми значениями. Мы повторили эту процедуру 1000 раз. Средняя корреляция истинной совокупности считалась значимой, если она превышала 95% этих оценок с повторной выборкой. Чтобы определить ожидаемое распределение корреляций, если нейронная активность остается одинаковой в двух наборах условий, мы сравнили rtop и rbottom для каждого нейрона. rtop был построен путем объединения скоростей стрельбы для четырех условий (вперед и назад, для обеих рук) с расположением цели в верхней части цикла. rbottom был построен аналогично для четырех условий с целевым расположением в нижней части цикла. Поскольку в испытаниях с началом сверху и с началом снизу положение рук сдвинуто по фазе на 180 градусов, мы использовали циклы 2–5 при построении rtop и циклы 1,5–4,5 при построении rbottom. Для каждого нейрона мы вычислили коэффициент корреляции Пирсона между rtop и rbottom.
Поскольку поведение было очень похожим для условий начала сверху и снизу (за исключением разницы фаз), ожидается, что эти корреляции будут близки к единице, если только не будет ослаблена ошибка выборки в оценках скорострельности. Нормализация
Запросить подробный протоколПоскольку абсолютные напряжения кривых ЭМГ в значительной степени произвольны, шкала мышечной активности может быть совершенно разной для разных мышц. Таким образом, реакция каждой мышцы была нормализована по ее диапазону. Нейронные ответы оставляли ненормализованными для анализа отдельных нейронов. Для анализа на уровне популяции ответы были нормализованы, чтобы предотвратить чрезмерное смещение результатов в сторону свойств нескольких высокочастотных нейронов. Например, анализ основных компонентов (PCA) стремится зафиксировать максимальную дисперсию, и нейрон с модуляцией частоты возбуждения 100 импульсов в секунду будет вносить в 25 раз большую дисперсию, чем нейрон с модуляцией 20 импульсов в секунду.
Уменьшение этого несоответствия побуждает PCA суммировать ответ всех нейронов. Мы нормализовали скорость возбуждения каждого нейрона, используя (2) FRsoftnorm=FRrangeFR+5
Добавление 5 к знаменателю дает «мягкую» нормализацию и гарантирует, что мы не преувеличиваем активность очень низкочастотных нейронов. Мы использовали это значение ранее (Lara et al., 2018; Russo et al., 2018), поскольку оно обеспечивает разумный баланс между фокусировкой анализа на всех нейронах и позволяет нейронам с высокой частотой импульсов вносить несколько больший вклад, чем нейронам с очень низкой частотой. нейроны.
Скорострельное воздействие малых движений нерабочей руки
Запросить подробный протоколМы хотели контролировать возможность того, что нейронные реакции в неведущем полушарии (ипсилатеральном по отношению к работающей руке) могут быть связаны с небольшими движениями (контралатеральной) недействующей руки. Для каждого испытания мы рассчитывали среднюю (абсолютную) скорость неработающей руки.
Для каждого условия мы разделили испытания на те, в которых скорость была больше или меньше медианы. Мы не применяли этот анализ, если было менее восьми испытаний для этого состояния для этого нейрона (что иногда происходило, если нейрон был хорошо изолирован только в течение части сеанса записи). После деления мы повторно вычислили среднюю частоту стрельбы для каждого из двух наборов испытаний, получив одну частоту стрельбы, когда неэффективная рука двигалась умеренно, и другую, когда она была практически неподвижна. Для каждой временной точки мы спрашивали, отличались ли эти две скорости срабатывания больше, чем ожидалось, учитывая ошибку выборки. Это было достигнуто с помощью теста рандомизации, в котором испытания были разделены на две группы случайным образом, а не на основе скорости. Мы провели 1000 таких случайных делений. Различия считались значимыми, если они были больше, чем для 95% случайных делений.
Сравнение прогнозов мышечной активности между полушариями
Запросить подробный протоколЧтобы предсказать мышечную активность по нейронной активности, мы использовали регрессию с частичными наименьшими квадратами (PLS) (plsregress в MATLAB).
, мы получим матрицу весов ранга r, которая прогнозирует мышечную активность непосредственно на основе нейронной активности. Все предсказания касались средних циклов (2-5) движения. Мы центрировали все данные так, чтобы средняя нервная и мышечная активность была нулевой для каждого условия (гарантируя, что успешное предсказание включало в себя захват структуры скорости стрельбы, а не просто общее среднее значение). Мы выбрали одно поведенческое условие (например, старт сверху, вперед, игра правой рукой) в качестве тестового условия. Мы устанавливаем условие тренировки, чтобы оно было соответствующим условием с противоположным стартовым положением педали (например, старт снизу, вперед, выполнение правой рукой). Мы запустили регрессию PLS для условия обучения, чтобы найти матрицу ранга-r B. Чтобы выбрать оптимальный ранг, мы выбрали один цикл из нашего тестового условия, чтобы он служил проверочными данными. Мы оценили эффективность этого цикла проверки и выбрали ранг r* и соответствующую весовую матрицу B*, которые генерировали максимальную проверку R2.
Мы оценили эффективность предсказания (обобщение) на оставшихся циклах тестового условия. Эта процедура повторялась для каждого состояния и полушария. Эффективность обобщения была определена количественно на основе объясненной процентной дисперсии генеральной совокупности: (3) R2=1-Y-Ypredf2Yf2
где Ypred=XB* и ∙f указывает норму Фробениуса.
Сравнение моделей активности в разных полушариях
Запросить подробный протоколЧтобы оценить, отсутствуют ли основные сигналы от управляющей коры в неуправляющей коре, мы проанализировали, насколько хорошо можно предсказать активность управляющей коры по активности неуправляющей коры. Для этого анализа мы использовали регрессию основных компонентов, которая регрессирует управляющую активность коры по сравнению с верхними ПК-проекциями неуправляемой активности коры. Это позволяет нам определить как наличие, так и относительную величину сигналов в каждом полушарии. Этот анализ проводился на средних циклах (2-5) движения.
Регрессия PLS дала аналогичное качество прогноза (данные не показаны). Однако регрессия PLS специально разработана так, чтобы быть устойчивой к различиям в величинах сигналов между предиктором и целевым набором данных, поэтому она сделала сравнение величин сигналов между полушариями менее простым. 9, мы получаем матрицу весов ранга r, которая предсказывает управляющую активность коры непосредственно из неуправляющей активности коры. Мы рассчитали B отдельно для значений r от 1 до 10. Чтобы определить, насколько хорошо B предсказывает активность коры головного мозга, мы количественно оценили популяцию R2 (уравнение 3) при тестовом условии: поведение с тем же направлением вращения педалей и работающей рукой, но противоположное исходное и конечное положение в качестве условия тренировки. Для сравнения мы также подсчитали, насколько хорошо активность управляющей коры может быть предсказана другими единицами управляющей коры. Мы повторили описанные выше шаги, за исключением того, что X определили как все единицы, кроме единицы p из управляющей коры, и Y как активность единицы p.
Таким образом, веса регрессии определяются для каждой единицы отдельно, но по-прежнему ограничиваются извлечением только из верхних сигналов r ПК, рассчитанных по всем другим единицам. Эффективность обобщения рассчитывалась с использованием совокупности R2 (уравнение 3) в условиях теста. Чтобы оценить значимость, мы выполнили повторную выборку, чтобы оценить величину ожидаемой разницы в R2, если бы не было основного разделения между полушариями. Мы случайным образом переназначили каждый нейрон как «нейрон левого полушария» или «нейрон правого полушария». «управляющая кора», чтобы предсказать себя, как указано выше. Этот тест перетасовки был выполнен 100 раз. Разница в R2 между истинным прогнозом вождение→вождение и отсутствие вождения→вождение была определена как значительная, если она превышала 9.5/100 этих передискретизированных различий.
Чтобы подтвердить, что приведенного выше анализа действительно достаточно для выявления различий между двумя полушариями, мы провели два контроля.
– матрица ПК с отсутствующим первым столбцом. Этот анализ подтвердил, что, когда сигнал отсутствует в неуправляемой коре, предсказание управляющей активности коры из неуправляющей активности коры страдает (рис. 7 — дополнение к рисунку 1А). Мы выполнили соответствующий контроль, в котором мы изменили размер сигналов в популяции неуправляющей коры. Для этого мы спроецировали X в его пространство PC, в результате чего XPC=XW. Мы разделили первые два столбца XPC на два и умножили столбцы три и четыре на два, получив X~PC. Затем мы реконструировали активность каждого нейрона: X~=X~PCWt и проанализировали X~, как и раньше. Этот анализ подтвердил, что предсказание страдает, если размеры сигналов не совпадают между двумя полушариями (рис. 7 — дополнение к рисунку 1B). Ортогональность левого и правого подпространств
Запросить подробный протоколМы использовали PCA, чтобы найти нейронные измерения, занятые, когда задача выполняется с данной рукой (например, левой рукой).
Затем мы спросили, насколько дисперсия объясняется этими размерами, когда задача выполняется с использованием другой руки (например, правой руки). В этих анализах использовалась усредненная по пробам частота стрельбы (как описано в этом разделе), и они были повторены с использованием оценок единичной пробной частоты стрельбы (см. раздел, описывающий LFADS ниже). При анализе учитывались три условия: условие обучения, условие проверки «одной руки» и условие проверки «противоположной руки». Условие тренировки и условие теста с одной и той же рукой имели одну и ту же руку и направление вращения, но имели разные начальные положения. Условия тренировки и условия теста с противоположной рукой были идентичными, но использовались разные руки. Анализ проводился восемь раз для каждой обезьяны, при этом каждое условие использовалось в качестве условия обучения ровно один раз. Примеры проекций показаны для пар тестовых условий. Количественные сводки (например, объясненная дисперсия) усредняются по восьми соответствующим условиям испытаний.
Для данного условия обучения мы применили PCA к матрице t×n частот срабатывания, где t – количество временных точек в течение четырех средних циклов движения, а n – количество нейронов. Это дало матрицу n × n ПК. Для каждого из двух тестовых условий мы проецировали соответствующую матрицу t × n частот стрельбы на эти ПК и оценивали дисперсию, объясняемую каждым ПК.
Насколько два измеренных подпространства близки к ортогональным, зависит от количества нейронов в популяциях выборки. Небольшие выборки смещают оценки от ортогональности. Поэтому мы исследовали, как наши оценки перекрытия подпространств масштабируются с количеством нейронов (рис. 10 — дополнение к рисунку 1). Эти результаты подтвердили, что большое количество нейронов (> 200 на полушарие) важно для точной оценки перекрытия подпространств.
Коэффициент Фано
Запросить подробный протоколМы использовали фактор Фано как грубую меру того, являются ли усредненные по пробам частоты срабатывания репрезентативными для индивидуальных пробных ответов.
Фактор Фано определяется как отношение дисперсии количества спайков (по испытаниям) к среднему количеству спайков, при этом подсчеты производятся во временном окне фиксированной продолжительности. Мы рассчитали фактор для всех единичных выделений, по крайней мере, с десятью испытаниями на состояние. Чтобы подсчитать количество всплесков, мы сопоставили все испытания для определенного состояния с самой серединой цикла (время, когда положение руки пересекло 3,5 цикла, на полпути между началом и концом испытания). В каждом испытании пики подсчитывали в двух окнах по 250 мс: одно до и одно после времени выравнивания. Для каждого окна (рассматриваемого отдельно) мы рассчитали среднее значение и дисперсию количества спайков. Мы не изменяли временную основу отдельных испытаний. Модифицированная временная база усложняет интерпретацию изменчивости всплесков и была ненужной, учитывая относительно короткие промежутки времени, рассматриваемые здесь (поведение дрейфовало немногим более ± 250 мс). Чтобы вычислить один фактор Фано для каждого набора данных, мы рассмотрели как окна, так и все нейроны и условия, и использовали регрессию для оценки взаимосвязи между дисперсией количества спайков и средним значением (Churchland et al. , 2010a). LFADS
Запросить подробный протоколЧтобы визуализировать однократные нейронные траектории, мы использовали недавно разработанный метод машинного обучения: анализ скрытых факторов с помощью динамических систем (LFADS) (Pandarinath et al., 2018). LFADS обучает вариационный автоматический кодировщик генерировать частоту возбуждения нейронов в одной попытке с помощью динамики, созданной рекуррентной нейронной сетью. Этот подход использует одновременные нейронные записи, чтобы вывести скрытую динамику, которая генерирует нейронную активность, и использует эту динамику, чтобы генерировать очищенные от шума оценки частоты срабатывания одной попытки для каждого нейрона. Это позволило нам повторить анализ подпространств левой и правой руки (описанных выше для ответов, усредненных по пробам) на оценках частоты стрельбы в одной попытке. Мы использовали LFADS Run Manager (написанный Daniel J. O’Shea: https://lfads.github.io/lfads-run-manager/).
Преимущество LFADS заключается в том, что он может использовать данные, записанные между сеансами.
Скрытые траектории в одном испытании ограничиваются как спайками в этом испытании, так и динамикой, предполагаемой между испытаниями и сеансами. В настоящем исследовании все анализы однократных траекторий выполняются в рамках одного сеанса, но включение нескольких сеансов помогает уменьшить шум траекторий за счет лучшего ограничения динамической модели. Сеансы выбирались по двум критериям. Во-первых, сеанс должен был включать как минимум двадцать единиц (для обоих полушарий), все из которых оставались изолированными на протяжении всего сеанса (в отличие от усредненных данных, где мы могли исключить испытания, если изоляция стала плохой). Во-вторых, мы проанализировали только сеансы, в которых выборка свойств ответа не была чрезмерно «однобокой», при одном или нескольких состояниях было мало реагирующих нейронов. Это очень маловероятно в больших наборах данных, но может происходить в отдельных сеансах и усложняет сравнение дисперсии, полученной для разных условий. Поэтому мы потребовали, чтобы дисперсия нейронной популяции в условиях наибольшей дисперсии поведения не превышала более чем в три раза дисперсию нейронной популяции в условиях наименьшей дисперсии: (4) ∑nσn,C+2≤3∑nσn,C-2
, где σn,C+2 обозначает дисперсию нейрона n при условии с максимальной суммированной дисперсией, а σn,C-2 обозначает дисперсию нейрона n при условии с минимальной суммированной дисперсией.
Для Monkey E этим критериям соответствовало в общей сложности 8 наборов данных с диапазоном от 21 до 35 одновременно зарегистрированных единиц. Для Monkey F этим критериям соответствовало в общей сложности 11 наборов данных с 20-41 одновременно зарегистрированной единицей. LFADS работает при подсчете спайков, в бинах по 1 мс, в окне одинакового размера для каждой попытки. Вход представляет собой трехмерный тензор размера, умноженный на количество нейронов по числу испытаний. При каждом испытании LFADS определяет начальное состояние и скрытую траекторию, которая развивается в соответствии с изученной динамикой. Чтобы избежать сложности модели LFADS, мы не позволили ей также вывести изменяющиеся во времени входные данные. Это ограничивало модель, чтобы фиксировать все различия между испытаниями, используя разные начальные состояния. По этой причине важно, чтобы испытания не были слишком продолжительными. Для длительных испытаний вариации между испытаниями, которые происходят поздно, как правило, не фиксируются, если их нельзя объяснить различиями в начальном состоянии.
Мы специально хотели зафиксировать как можно большую изменчивость от испытания к испытанию. Чтобы предоставить LFADS наилучшую возможность для этого, мы определили «испытание» LFADS как единый цикл поведения. Таким образом, каждое поведенческое испытание было разделено на четыре испытания LFADS (по одному на каждый цикл от 2 до 5). Каждый из них охватывал окно продолжительностью 500 мс, центр которого находился в момент, когда рука достигла верхнего положения педали (обратите внимание, что в разных испытаниях педаль может перемещаться чуть больше или меньше одного цикла в течение этого окна). Мы обучили сшитый многодневный автоэнкодер LFADS со следующими параметрами. Размер бина: 5 мс; размер партии: 32; факторные размеры: 16; размеры генератора: 64; размеры энкодера: 64; остановка скорости обучения: 0,001. Другие параметры были оставлены со значениями по умолчанию, предоставленными LFADS Run Manager. Анализ подпространств левой и правой руки в отдельных испытаниях проводился параллельно исходному анализу, основанному на средних значениях испытаний.
Показатели частоты стрельбы, полученные на основе LFADS, были проецированы на подпространства левой и правой руки. Как и в исходном анализе, подпространства были найдены на основе условий обучения, а спроецированные данные были получены из двух тестовых условий (одна и та же рука, но другое начальное положение или соответствующее условие для другой руки). Мы вычислили захваченную дисперсию в каждом подпространстве для каждого условия теста. Несмотря на то, что LFADS использовал данные между сеансами, описанные выше этапы анализа выполнялись независимо для каждого сеанса. Подпространства были найдены на основе внутрисеансовых пробных средних, чтобы ответить на ключевой вопрос о том, разделяют ли подпространства, выведенные из пробных средних, ответы одиночных проб так же, как они делают ответы, усредненные пробами. Аналогичные результаты были получены, если подпространства были найдены на основе одиночных испытаний. Запутывание траекторий
Запросить подробный протоколМы оценили запутывание траектории, как описано Russo et al.
(2018). Чтобы провести параллель с другими анализами популяционной активности, запутывание траекторий было рассчитано для траекторий во время средних циклов. Усредненная по пробам нейронная активность была снижена до восьми верхних измерений с помощью PCA. Затем мы рассчитали запутывание, Qt, в каждый момент времени: (5) Q(t)=maxt′‖x˙t−x˙t′‖2‖xt−xt′‖2+ε
, где xt – состояние нервной системы в момент t, x˙t – временная производная состояния нервной системы. , ∙ – это евклидова норма, а ε – небольшая константа, которая предотвращает деление на ноль (здесь установлено значение 10 % от общей нейронной дисперсии по восьми верхним измерениям). И t, и t’ индексируются во времени в нескольких условиях. Таким образом, Qt становится большим, если состояние нейрона в момент времени t близко к состоянию нейрона в другое время или при других условиях, но эти два состояния имеют очень разные производные. В настоящем исследовании запутывание оценивалось отдельно для каждого полушария.
Запутывание также оценивали отдельно для ведомых и неведомых плеч. Таким образом, t и t’ индексируются во все времена в рамках четырех условий (например, четырех условий, когда ведомая рука выполняет задачу). Кумулятивные распределения Qt были объединены по полушариям, чтобы создать одно распределение для «управляющей коры», а другое — для «неуправляющей коры». Приведенный выше анализ также применялся к мышечным популяциям. Запутывание вычислялось для каждой руки (используя только условия, при которых эта рука выполняла задачу), а затем распределения были объединены между двумя руками, чтобы создать единое распределение для сравнения с нейронными данными.
Способность декодеров ЭМГ игнорировать действия, связанные с неработающей рукой
Запросить подробный протоколМы оценили, может ли линейный нейронный декодер предсказать мышечную активность в данной руке как тогда, когда эта рука выполняет задачу (и необходимо декодировать надежную ЭМГ), так и когда другая рука выполняет задачу (и должна быть декодирована почти нулевая ЭМГ).
. Как и в нашем предыдущем анализе прогнозирования ЭМГ, мы использовали регрессию PLS (описанную выше) и сосредоточились на средних четырех циклах. Для этого анализа мы предсказали активность ЭМГ, используя все нейроны, независимо от полушария. Сначала мы оценили эффективность обобщения с помощью декодера «движущийся поезд», который был обучен только условиям, в которых соответствующая рука выполняла задачу, а затем его попросили обобщить условия, в которых другая рука выполняла задачу. Это потенциально сложная форма обобщения, поскольку декодер должен предсказать активность ЭМГ в ситуации (рука не двигается), сильно отличающейся от ситуации, в которой он обучался (рука двигается). Мы также рассчитали производительность обобщения декодера «обучение обоих», обученного с использованием набора условий, включающих соответствующую руку, выполняющую и не выполняющую задачу. Обобщение заключалось в исключении условий каждого типа.
Для декодера движения поезда мы использовали следующее разделение условий обучения, проверки и тестирования: Обучение : Направление 1, Начальное положение 1, Движение руки.
Валидация : Направление 1, Начальное положение 2, Движение рычага (один цикл). Тестирование : Направление 1, Начальное положение 2, Движение другой руки. Для декодера поезд-оба мы использовали следующее разделение обучения, проверки и тестирования: Обучение : Направление 1, Начальное положение 1, Движение руки и Направление 1, Начальное положение 1, Движение другой руки. Проверка : Направление 1, начальное положение 2, движение рычага (один цикл). Тестирование : Направление 1, Начальное положение 2, Движение другой руки. Доступность данных
Усредненные по испытаниям нейронные данные и данные ЭМГ, а также данные одного испытания, обработанные LFADS, были загружены в Open Science Framework (OSF), и к ним можно получить доступ по следующей ссылке: https://osf.io/859st/.
Были сгенерированы следующие наборы данных
-
- Эймс К
- Черчленд М
(2019) Open Science Framework
Данные для: Сигналы моторной коры для каждой руки смешиваются между полушариями и нейронами, но разделяются в ответе популяции.
https://osf.io/859st/
Каталожные номера
-
- Афшар А
- Сантанам Г
- Ю БМ
- Рю СИ
- Сахани М
- Шеной КВ
(2011) Однократные нейронные корреляты подготовки к движению руки
Нейрон 71 : 555–564.
https://doi.org/10.1016/j.neuron.2011.05.047
- пабмед
- Google ученый
-
- Айзава Х
- Мусиаке H
- Инасе М
- Танджи Дж
(1990) Выходная зона первичной моторной коры обезьяны, специализирующаяся на билатеральных движениях рук
Экспериментальные исследования мозга 82 :219–221.
https://doi.org/10.1007/BF00230856
- пабмед
- Google ученый
-
- Берло Е
- Причард Г
- О’Рейли J
- Эджаз N
- Дидрихсен J
(2019) Ипсилатеральные представления пальцев в сенсомоторной коре управляются активными процессами движения, а не пассивным сенсорным входом
https://doi.org/10.1152/jn.00439.2018
- пабмед
- Google ученый
-
- Черчленд ММ
- Афшар А
- Шеной КВ
(2006) Центральный источник двигательной изменчивости
Нейрон 52 :1085–1096.
https://doi.org/10.1016/j.neuron.2006.10.034
- пабмед
- Google ученый
-
- Черчленд ММ
- Каннингем JP
- Кауфман МТ
- Рю СИ
- Шеной КВ
(2010а) Корковая подготовительная деятельность: представление о движении или первый винтик динамической машины?
Нейрон 68 :387–400.
https://doi.org/10.1016/j.neuron.2010.09.015
- пабмед
- Google ученый
-
- Черчленд ММ
- Ю БМ
- Каннингем JP
- Сугрю LP
- Коэн MR
- Коррадо GS
- Ньюсом WT
- Кларк AM
- Хоссейни П
- Скотт ББ
- Брэдли, округ Колумбия,
- Смит Массачусетс
- Кон А
- Мовшон ЮА
- Армстронг КМ
- Мур Т
- Чан SW
- Снайдер LH
- Лисбергер СГ
- Прибе, штат Нью-Джерси,
- Финн IM
- Ферстер Д
- Рю Си
- Сантанам Г
- Сахани М
- Шеной КВ
(2010б) Возникновение стимула подавляет нервную изменчивость: широко распространенный кортикальный феномен
Nature Neuroscience 13 :369–378.
https://doi.org/10.1038/nn.2501
- пабмед
- Google ученый
-
- Черчленд ММ
- Каннингем JP
- Кауфман МТ
- Фостер JD
- Нуюджукян П
- Рю СИ
- Шеной КВ
(2012) Динамика нейронной популяции при достижении
Природа 487 :51–56.
https://doi.org/10.1038/nature11129
- пабмед
- Google ученый
-
- Черчленд ММ
- Эббот ЛФ
(2012) Два уровня нейронной изменчивости
Nature Neuroscience 15 :1472–1474.
https://doi.org/10.1038/nn.3247
- пабмед
- Google ученый
-
- Черчленд ММ
- Шеной КВ
(2007) Временная сложность и гетерогенность активности отдельных нейронов в премоторной и моторной коре
Журнал нейрофизиологии 97 :4235–4257.
https://doi.org/10.1152/jn.00095.2007
- пабмед
- Google ученый
-
- Чисек П
- Краммонд DJ
- Каласка JF
(2003) Нейронная активность в первичной моторной и дорсальной премоторной коре при выполнении задач контралатеральной рукой по сравнению с ипсилатеральной
Journal of Neurophysiology 89 :922–942.
https://doi.org/10.1152/jn.00607.2002
- пабмед
- Google ученый
-
- Дидрихсен J
- Вистлер Т
- Кракауэр JW
(2013) Два отдельных ипсилатеральных корковых представления для индивидуальных движений пальцев
Кора головного мозга 23 :1362–1377.
https://doi.org/10.1093/cercor/bhs120
- пабмед
- Google ученый
-
- Дончин О
- Грибова А
- Стейнберг О
- Бергман Х
- Ваадия E
(1998) Первичная моторная кора участвует в бимануальной координации
Природа 395 :274–278.
https://doi.org/10.1038/26220
- пабмед
- Google ученый
-
- Дончин О
- Грибова А
- Стейнберг О
- Митц AR
- Бергман H
- Ваадия E
(2002) Единичная активность, связанная с бимануальными движениями рук в первичной и дополнительной моторной коре
Journal of Neurophysiology 88 :3498–3517.
https://doi.org/10.1152/jn.00335.2001
- пабмед
- Google ученый
-
- Дракманн S
- Чкловский ДБ
(2012) Нейронные цепи, лежащие в основе постоянных репрезентаций, несмотря на меняющуюся во времени активность
Современная биология 22 :2095–2103.
https://doi.org/10.1016/j.cub.2012.08.058
- пабмед
- Google ученый
-
- Душанова Ж
- Донохью Дж.
(2010) Нейроны первичной моторной коры, задействованные во время наблюдения за действием
European Journal of Neuroscience 31 :386–398.
https://doi.org/10.1111/j.1460-9568.2009.07067.x
- пабмед
- Google ученый
-
- Дуйсенс J
- Ван де Кроммер HW
(1998) Нейронный контроль передвижения; Центральный генератор паттернов от кошек к людям
Походка и осанка 7 :131–141.
https://doi.org/10.1016/S0966-6362(97)00042-8
- пабмед
- Google ученый
-
- Эльсайед ГФ
- Лара AH
- Кауфман МТ
- Черчленд ММ
- Каннингем JP
(2016) Реорганизация между подготовительными и двигательными популяционными реакциями в моторной коре
Nature Communications 7 :13239.
https://doi.org/10.1038/ncomms13239
- пабмед
- Google ученый
-
- Ферберт А
- Приори А
- Ротвелл JC
- День BL
- Колебэтч JG
- Марсден CD
(1992) Межполушарное торможение моторной коры человека
Журнал физиологии 453 : 525–546.
https://doi.org/10.1113/jphysiol.1992.sp019243
- пабмед
- Google ученый
-
- Фогасси L
- Феррари ПФ
- Гезиерих Б
- Роззи S
- Черси F
- Риццолатти G
(2005) Теменная доля: от организации действия к пониманию намерения
Наука 308 :662–667.
https://doi.org/10.1126/science.1106138
- пабмед
- Google ученый
-
- Гангули К
- Секундо L
- Ранаде Г
- Орсборн А
- Чанг ЭФ
-
Димитров Д. Ф.
- Уоллис JD
- Барбаро Н.М.
- Рыцарь РТ
- Кармена JM
(2009) Корковое представление ипсилатеральных движений рук у обезьяны и человека
Journal of Neuroscience 29 :12948–12956.
https://doi.org/10.1523/JNEUROSCI.2471-09.2009
- пабмед
- Google ученый
-
- Глис П
- Коул Дж
(1950) Восстановление квалифицированных двигательных функций после небольших повторных поражений моторной коры у макак
Journal of Neurophysiology 13 :137–148.
https://doi.org/10.1152/jn.1950.13.2.137
- Google ученый
-
- Гулд HJ
- Кьюсик CG
- Мост ТП
- Каас JH
(1986) Взаимосвязь связей мозолистого тела с картами электрической стимуляции моторных, дополнительных моторных и лобных полей глаза у сов-обезьян
Журнал сравнительной неврологии 247 : 297–325.
https://doi.org/10.1002/cne.
0303 0111- пабмед
- Google ученый
- Грибова А
- Дончин О
- Бергман H
- Ваадия E
- Кардосо Де Оливейра S
(2002) Синхронизация бимануальных движений человека и приматов в зависимости от активности нейронов в первичной моторной коре и дополнительной моторной области
Experimental Brain Research 146 :322–335.
https://doi.org/10.1007/s00221-002-1174-x
- пабмед
- Google ученый
- Хакен Х
- Келсо JAS
- Бунц H
(1985) Теоретическая модель фазовых переходов в движениях рук человека
Биологическая кибернетика 51 :347–356.
https://doi.org/10.1007/BF00336922
- Google ученый
- Ханадзима Р
- Угава Y
- Мачии К
- Мотидзуки H
- Терао Y
- Эномото H
- Фурубаяси Т
- Шио Y
- Уэсуги Х
- Канадзава I
(2001) Межполушарное облегчение моторной области рук у людей
Журнал физиологии 531 : 849–859.
https://doi.org/10.1111/j.1469-7793.2001.0849h.x
- пабмед
- Google ученый
Препринт
- Хеминг Е
- Крест К
- Такей Т
- Кук диджей
- Скотт СХ
(2019) Держите руки врозь: независимые представления ипсилатеральных и контралатеральных передних конечностей в первичной моторной коре
биоRxiv.
https://doi.org/10.1101/587378
- Google ученый
- Дженни AB
(1979) Комиссуральные проекции кортикальной двигательной области руки у обезьян
Журнал сравнительной неврологии 188 : 137–145.
https://doi.org/10.1002/cne.
- пабмед
- Google ученый
-
- Джонс ЭГ
- Мудрый СП
(1977) Размер, ламинарное и столбчатое распределение эфферентных клеток в сенсорно-моторной коре обезьян
The Journal of Comparative Neurology 175 :391–437.
https://doi.org/10.1002/cne.0403
- пабмед
- Google ученый
-
- Кауфман МТ
- Черчленд ММ
- Рю СИ
- Шеной КВ
(2014) Корковая активность в нулевом пространстве: подготовка без движения
Nature Neuroscience 17 :440–448.
https://doi.org/10.1038/nn.3643
- пабмед
- Google ученый
-
- Казенников О
- Хайленд Б
- Корбоз М
- Бабалян А
- Руйе EM
- Визендангер М
(1999) Нейронная активность дополнительных и первичных двигательных областей у обезьян и ее связь с бимануальными и мономануальными последовательностями движений
Неврология 89 :661–674.
https://doi.org/10.1016/S0306-4522(98)00348-0
- пабмед
- Google ученый
-
- Келсо JA
(1984) Фазовые переходы и критическое поведение при бимануальной координации человека
Американский журнал физиологии – регуляторная, интегративная и сравнительная физиология 246 :R1000–R1004.
https://doi.org/10.1152/ajpregu.1984.246.6.R1000
- Google ученый
-
- Кермади I
- Лю Ю
- Темпини А
- Кальциати E
- Руйе EM
(1998) Нейронная активность в дополнительной моторной области приматов и первичной моторной коре в связи с пространственно-временной бимануальной координацией
Соматосенсорные и двигательные исследования 15 :287–308.
https://doi.org/10.1080/089
870709 - пабмед
- Google ученый
- Кермади I
- Лю Ю
- Руйе EM
(2000) Вовлекаются ли бимануальные двигательные действия в дорсальную премоторную (PMd), поясную (CMA) и заднюю теменную (PPC) кору? сравнение с первичными и дополнительными двигательными областями коры
Соматосенсорные и двигательные исследования 17 :255–271.
https://doi.org/10.1080/089
- пабмед
- Google ученый
- Красков А
- Филипп Р
- Вальдерт С
- Виньесваран G
- Куалло ММ
- Лимон РН
(2014) Корково-спинномозговые зеркальные нейроны
Philosophical Transactions of the Royal Society B: Biological Sciences 369 :20130174.
https://doi.org/10.1098/rstb.2013.0174
- Google ученый
Книга
- Кайперс HG
(1981)
Анатомия нисходящих путей Справочник по физиологии, Нервная система, двигательный контроль, 2
Интернет-библиотека Wiley.
- Google ученый
- Кван ХК
- Маккей Вашингтон
- Мерфи JT
- Вонг YC
(1978) Пространственная организация прецентральной коры бодрствующих приматов. II. мощность двигателя
Журнал нейрофизиологии 41 :1120–1131.
https://doi.org/10.1152/jn.1978.41.5.1120
- пабмед
- Google ученый
- Лара AH
- Эльсайед ГФ
- Зимник AJ
- Каннингем JP
- Черчленд ММ
(2018) Сохранение подготовительных нейронных событий в моторной коре обезьян независимо от того, как инициируется движение
eLife 7 :e31826.
https://doi.org/10.7554/eLife.31826
- Google ученый
- Ли Н
- Дайе К
- Свобода К
- Дракманн S
(2016) Надежная динамика нейронов в премоторной коре во время планирования движений
Природа 532 :459–464.
https://doi.org/10.1038/nature17643
- пабмед
- Google ученый
- Литвин-Кумар А
- Дуарон Б
(2012) Медленная динамика и высокая изменчивость в сбалансированных корковых сетях с кластерными связями
Nature Neuroscience 15 :1498–1505.
https://doi.org/10.1038/nn.3220
- Google ученый
- Лю Ю
- Руйе EM
(1999) Механизмы восстановления ловкости при одностороннем поражении сенсомоторной коры у взрослых обезьян
Экспериментальные исследования мозга 128 :149–159.
https://doi.org/10.1007/s002210050830
- пабмед
- Google ученый
- Махенс СК
- Ромо Р
- Броды CD
(2010) Функциональное, но не анатомическое разделение «что» и «когда» в префронтальной коре
Journal of Neuroscience 30 :350–360.
https://doi.org/10.1523/JNEUROSCI.3276-09.2010
- пабмед
- Google ученый
- Манте В
- Суссильо D
- Шеной КВ
- Ньюсом WT
(2013) Контекстно-зависимые вычисления с помощью рекуррентной динамики в префронтальной коре
Природа 503 : 78–84.
https://doi.org/10.1038/nature12742
- пабмед
- Google ученый
- Мацунами К
- Хамада I
(1981) Характеристики ипсилатерального связанного с движением нейрона в моторной коре обезьяны
Brain Research 204 :29–42.
https://doi.org/10.1016/0006-8993(81)
-1
- пабмед
- Google ученый
- Мейер БУ
- Рерихт S
- Графин фон Айнзидель H
- Круггель F
- Вайндл А
(1995) Тормозные и возбуждающие межполушарные передачи между моторными областями коры у здоровых людей и пациентов с аномалиями мозолистого тела
Головной мозг 118 ( Pt 2 :429–440.
https://doi.org/10.1093/brain/118.2.429
- пабмед
- Google ученый
- Майклс JA
- Шербергер H
(2018) Популяционное кодирование информации, связанной с хваткой и латеральностью, в лобно-теменной сети макак
Scientific Reports 8 :1710.
https://doi.org/10.1038/s41598-018-20051-7
- Google ученый
- Мурата Y
- Хиго N
- Оиси Т
- Ямасита А
- Мацуда К
- Хаяши М
- Яманэ С
(2008) Влияние двигательной тренировки на восстановление ловкости рук после поражения первичной моторной коры у макак-обезьян
Журнал нейрофизиологии 99 :773–786.
https://doi.org/10.1152/jn.01001.2007
- Google ученый
- Пандаринат C
- О’Ши DJ
- Коллинз Дж
- Юзефович Р
- Ставиский SD
- Као JC
- Траутманн EM
- Кауфман МТ
- Рю СИ
- Хохберг LR
- Хендерсон JM
- Шеной КВ
- Эббот ЛФ
- Суссильо Д
(2018) Вывод динамики нейронной популяции в одном испытании с использованием последовательных автокодировщиков
Nature Methods 15 :805–815.
https://doi.org/10.1038/s41592-018-0109-9
- пабмед
- Google ученый
- Пассингхэм RE
- Перри VH
- Уилкинсон F
(1983) Долгосрочные последствия удаления сенсомоторной коры у детенышей и взрослых макак-резусов
Мозг 106 (Pt 3 :675–705.
https://doi.org/10.1093/brain/106.3.675
- пабмед
- Google ученый
- Рапозо Д
- Кауфман МТ
- Черчленд AK
(2014) Свободная от категорий нейронная популяция поддерживает изменяющиеся потребности в процессе принятия решений
Nature Neuroscience 17 :1784–1792.
https://doi.org/10.1038/nn.3865
- пабмед
- Google ученый
- Риццолатти G
- Фадига Л
- Галлезе В
- Фогасси L
(1996) Премоторная кора и распознавание двигательных действий
Когнитивные исследования мозга 3 :131–141.
https://doi.org/10.1016/0926-6410(95)00038-0
- пабмед
- Google ученый
- Розенцвейг ES
- Брок JH
- Калбертсон MD
- Лу П
- Мосеенко Р
- Эджертон VR
-
Хэвтон Л. А.
- Тушинский MH
(2009) Обширный перекрест позвоночника и двусторонняя терминация шейных корково-спинномозговых проекций у макак-резусов
The Journal of Comparative Neurology 513 :151–163.
https://doi.org/10.1002/cne.21940
- пабмед
- Google ученый
- Руйе EM
- Бабалян А
- Казенников О
- Море В
- Ю ХН
- Визендангер М
(1994) Транскаллозальные соединения дистальных представительств передних конечностей первичной и дополнительной двигательных областей коры у макак
Experimental Brain Research 102 :227–243.
https://doi.org/10.1007/BF00227511
- пабмед
- Google ученый
- Руссо АА
- Биттнер SR
- Перкинс СМ
- Сили JS
- Лондон BM
- Лара AH
- Мири А
- Маршалл, штат Нью-Джерси,
- Кон А
- Джесселл ТМ
- Эббот ЛФ
- Каннингем JP
- Черчленд ММ
(2018) Моторная кора встраивает мышечные команды в реакцию распутанной популяции
Нейрон 97 :953–966.
https://doi.org/10.1016/j.neuron.2018.01.004
- Google ученый
- Сессле БЖ
- Визендангер М
(1982) Структурно-функциональное определение моторной коры у обезьяны ( Macaca fascicularis )
Журнал физиологии 323 :245–265.
https://doi.org/10.1113/jphysiol.1982.sp014071
- пабмед
- Google ученый
- Шадлен MN
- Ньюсом WT
(1998) Переменный разряд корковых нейронов: значение для связности, вычислений и кодирования информации
Журнал неврологии 18 :3870–3896.
https://doi.org/10.1523/JNEUROSCI.18-10-03870.1998
- пабмед
- Google ученый
- Сотеропулос DS
- Эджли С.А.
- Бейкер СН
(2011) Отсутствие доказательств прямого корково-спинномозгового вклада в контроль ипсилатеральной передней конечности у обезьян
Journal of Neuroscience 31 :11208–11219.
https://doi.org/10.1523/JNEUROSCI.0257-11.2011
- Google ученый
- Стейнберг О
- Дончин О
- Грибова А
- Кардоса-де-Оливейра S
- Бергман H
- Ваадия E
(2002) Популяции нейронов в первичной моторной коре кодируют бимануальные движения рук
European Journal of Neuroscience 15 : 1371–1380.
https://doi.org/10.1046/j.1460-9568.2002.01968.x
- пабмед
- Google ученый
- Суссильо D
- Черчленд ММ
- Кауфман МТ
- Шеной КВ
(2015) Нейронная сеть, которая находит натуралистическое решение для производства мышечной активности
Nature Neuroscience 18 :1025–1033.
https://doi.org/10.1038/nn.4042
- пабмед
- Google ученый
- Танджи Дж
- Окано К
- Сато KC
(1988) Нейронная активность в двигательных областях коры головного мозга, связанная с ипсилатеральными, контралатеральными и двусторонними движениями пальцев обезьяны
Journal of Neurophysiology 60 :325–343.
https://doi.org/10.1152/jn.1988.60.1.325
- пабмед
- Google ученый
- Верстинен Т
- Дидрихсен J
- Альберт N
- Апарисио П
- Иври РБ
(2005) Активность ипсилатеральной моторной коры во время одноручных движений руки связана со сложностью задачи
Журнал нейрофизиологии 93 :1209–1222.
https://doi.org/10.1152/jn.00720.2004
- Google ученый
- Виньесваран G
- Филипп Р
- Лимон РН
- Красков А
(2013) Корково-спинномозговые зеркальные нейроны M1 и их роль в подавлении движения во время наблюдения за действием
Current Biology 23 :236–243.
https://doi.org/10.1016/j.cub.2012.12.006
- пабмед
- Google ученый
- Виленский ЖК
- Гилман С
(2002) Поражения прецентральной извилины у нечеловеческих приматов: библиография Pre-Medline
International Journal of Primatology 23 : 1319–1333.
https://doi.org/10.1023/A:1021135122571
- Google ученый
- Вистлер Т
- Уотерс-Метенье S
- Дидрихсен J
(2014) Эффекторно-независимые представления моторных последовательностей существуют во внешней и внутренней системах отсчета
The Journal of Neuroscience 34 :5054–5064.
https://doi.org/10.1523/JNEUROSCI.5363-13.2014
- пабмед
- Google ученый
Письмо-решение
Благодарим вас за отправку статьи «Сигналы моторной коры для каждой руки смешиваются между полушариями и нейронами, но разделяются в популяционном ответе» на рассмотрение eLife . Ваша статья была проверена тремя рецензентами, в том числе Йорном Дидрихсеном в качестве редактора-рецензента и рецензентом № 1, а оценка проводилась под контролем Джошуа Голда в качестве старшего редактора. Следующее лицо, участвовавшее в рассмотрении вашей заявки, согласилось раскрыть свою личность: Кристиан К. Махенс (Рецензент №3).
Рецензенты обсудили обзоры друг с другом, и редактор-рецензент подготовил это решение, чтобы помочь вам подготовить исправленное представление.
Резюме:
В статье Эймса и Черчленда представлен важный и свежий взгляд на характеристики нейронной активности в моторной коре во время ипсилатеральных движений рук.
Во многих отношениях результаты подтверждают исследование Donchin et al., 1998, проведенное 20 лет назад, показывающее высокую распространенность ипсилатерально настроенной активности и отсутствие тесной связи между настройкой на контралатеральное и ипсилатеральное направления движения. Новый анализ, однако, показывает ортогональность популяционной активности для контра- и ипсилатеральной руки, что создает важные ограничения в понимании функции широко распространенной ипсилатеральной активности. Существенные исправления:
Рецензенты всесторонне обсудили вопросы, поднятые в отдельных обзорах, и пришли к соглашению по следующим основным вопросам, которые следует рассмотреть в пересмотре.
1) В то время как эти результаты вполне убедительны, объяснение, почему это двустороннее представительство необходимо (референсная копия для поддержки моторного планирования?), менее ясно. Например, движения пальцев часто требуют повышенных усилий по координации между обеими руками, например, для бимануальных движений рук.
Это кажется противоречащим точке зрения авторов о том, что дистальные (пальцевые) движения менее представлены билатерально, чем проксимальные (достигающие) движения. Дополнительная гипотеза, которую, возможно, следует рассмотреть в ходе обсуждения, заключается в том, что неуправляющая кора просто «наблюдает» за активностью управляющего полушария, но не вносит причинного вклада в движение управляющей руки. Как и при наблюдении за действием, нейронная активность будет происходить в нуль-пространстве мышц (Красков и др., 2014). Возможная функция такой активности (если таковая имеется) может заключаться в том, что она помогает закладывать сети в ипсилатеральном полушарии, которые затем лежат в основе межпальцевой передачи двигательных навыков (Wiestler et al., 2014). 2) Как вы могли подтвердить, что записи были на самом деле с M1, а не, например, с PMd? Вы искали низкие пороги ICMS? Это важно, поскольку двусторонние представительства в PMd могут сильно отличаться от M1.
3) Похоже, что все анализы проводились на усредненных данных.
Мы считаем, что крайне важно, чтобы авторы показали, по-прежнему ли ортогональные подпространства надежно разделяют активность популяции в отдельных испытаниях. Необходимо, чтобы однократная активность также попадала в нуль-пространство для контралатеральной руки, чтобы она могла избежать генерации контралатеральной активности. Хотя ваш анализ показывает, что в среднем активность попадает в нулевое пространство, мы считаем, что было бы информативно увидеть распространение проекции. Это можно сделать, спроецировав одиночные испытания на ортогональные подпространства и показав, что информация там есть, или применив методы уменьшения размерности с одним испытанием. (В этом контексте было бы интересно узнать, можно ли предсказать однократную мышечную активность в неведомой руке по отклонениям от ортогональных подпространств. Насколько я понял ограничения данных, это не будет проверить в текущем наборе данных, так как мышечная активность и мозговая активность регистрировались отдельно.
Я не прошу авторов делать это, но хотел бы отметить, что это может значительно усилить гипотезу авторов.) 4 ) Как отмечалось в Обсуждении, активность в одном полушарии M1 мало влияет на мышечную активность в неведомой руке. Тем не менее, уменьшение размерности и прогнозный анализ на рисунках 9и 10 учитывают полную популяцию в разных полушариях. Чтобы ответить на вопрос, почему популяционная активность в одном полушарии не активирует неуправляемую руку, анализ уменьшения размерности и прогнозирования в идеале должен выполняться на одном полушарии. Эффект может быть слабее (из-за меньшего количества данных), но он все равно должен быть. В качестве альтернативы авторы могли бы продемонстрировать, что веса декодирования (или веса, характеризующие подпространства) должным образом разделяют информацию между двумя полушариями.
5) Даже внутри одного полушария дополнительная проблема заключается в том, что может существовать смесь типов клеток, некоторые из которых отвечают за прямой двигательный контроль (например, небольшая доля клеток с индексом предпочтения, близким к 1), а некоторые отвечают за для общего расчета, скажем.
Предыдущая работа на мышах ALM показала, что такое разделение может существовать со связанными анатомическими различиями в предпочтениях левых/правых (Li et al., 2015, Nature 519 51-56). Хотелось бы, чтобы авторы (как минимум) выводили некоторую информацию о весах декодеров или подпространств (см. также пункт 2). 6) Авторы используют PLS, чтобы найти низкоразмерное линейное отображение между активностью популяции X и активностью популяции Y, и оценили его в параметрах обобщения, насколько хорошо можно предсказать новые данные. Основные результаты заключаются в том, что мышечная активность может быть почти одинаково хорошо предсказана по контралатеральному и ипсилатеральному M1, и что едва ли есть какие-либо различия в предсказании левого M1 по левому M1 и по правому M1. Основное ограничение этого анализа заключается в том, что он может показать, что два кода совокупности занимают общее линейное подпространство, но не показывает, что два кода совокупности структурно одинаковы (или имеют одинаковую репрезентативную геометрию — Дидрихсен и Кригескорте, 2017).
. Например, рассмотрим две популяции, изображенные на Рисунке 1. У обеих есть нейронное измерение, которое кодирует вертикальное положение руки, и нейронное измерение, которое кодирует горизонтальное положение руки (конечно, нейронные измерения не точно отражают конкретные физические особенности). переменные, но это не главное). Это означает, что вы можете найти двумерное отображение, которое будет предсказывать популяционную активность в регионе А по региону В, вероятно, почти так же хорошо, как вы могли бы предсказать регион А по региону А. Кроме того, специфическая мышечная активность, которая представляет собой линейную комбинацию положение можно одинаково хорошо (или плохо) считывать из активности населения. Однако это затемняет тот факт, что область А придает чрезмерное значение вертикальному измерению движения, тогда как область В делает упор на кодирование горизонтального измерения. Два разных кода нейронной популяции, которые занимают одно и то же функциональное подпространство, но имеют разную репрезентативную геометрию.
https://doi.org/10.7554/eLife.46159.021
Эта проблема особенно распространена, поскольку лежащее в ее основе поведение относительно простое. В то время как направление велосипедного движения разделяет положение, скорость и мышечную активность, две исходные точки, вероятно, очень мало делают для добавления новых измерений, которые функциональное подпространство должно кодировать в мозгу. Таким образом, пока оба полушария занимают это относительно ограниченное подпространство нейронной активности, прогнозирование модели PLS будет достаточно хорошим. То есть два очень разных кода совокупности могут выглядеть очень похожими, если в ходе эксперимента не будут диссоциированы критические измерения.
Мы считаем, что авторы должны, по крайней мере, признать эти два недостатка своего регрессионного подхода.
https://doi.org/10.7554/eLife.46159.026
Ответ автора
Основные версии:
Рецензенты всесторонне обсудили вопросы, поднятые в отдельных обзорах, и пришли к соглашению по следующим основным вопросам, которые следует рассмотреть при пересмотре.
1) Хотя эти выводы вполне убедительны, объяснение, почему необходимо такое двустороннее представительство (референсная копия для поддержки моторного планирования?), менее ясно. Например, движения пальцев часто требуют повышенных усилий по координации между обеими руками, например, для бимануальных движений рук. Это кажется противоречащим точке зрения авторов о том, что дистальные (пальцевые) движения менее представлены билатерально, чем проксимальные (достигающие) движения. Дополнительная гипотеза, которую, возможно, следует рассмотреть в ходе обсуждения, заключается в том, что неуправляющая кора просто «наблюдает» за активностью управляющего полушария, но не вносит причинного вклада в движение управляющей руки.
Как и при наблюдении за действием, нейронная активность будет происходить в нуль-пространстве мышц (Красков и др., 2014). Возможная функция такой активности (если таковая имеется) может заключаться в том, что она помогает закладывать сети в ипсилатеральном полушарии, которые затем лежат в основе межпальцевой передачи двигательных навыков (Wiestler et al., 2014). Мы согласны со всеми вышеперечисленными пунктами. Во-первых, действительно неясно, зачем нужно двустороннее представительство, и остается множество разумных возможностей. Во-вторых, мы согласны с тем, что нет четкой причины, по которой реакции, связанные с пальцами/руками, должны быть более латерализованными (можно было бы ожидать как раз обратного, как отмечают рецензенты).
Кроме того, неясно, действительно ли реакции, связанные с пальцами/руками, более латерализованы. Результаты фМРТ показывают двусторонние ответы, связанные с пальцами. Наконец, мы согласны с тем, что возможной целью активности в неуправляющей коре является «наблюдение» за управляющей активностью коры с целью межмануальной передачи навыка.
Мы внесли следующие изменения в рукопись. 1) Мы пересмотрели соответствующий раздел Обсуждения (Возможные причины ипсилатеральной двигательной активности коры), чтобы учесть вышеуказанные пункты. В частности, теперь мы повышаем возможность наблюдения за межручным переносом навыков:
«Независимо от роли в онлайн-координации, эфферентная копия может позволить неуправляющей коре «наблюдать» за вычислениями в управляющей коре, помогая межручная передача навыков (Weastler et al., 2014). […] Эти результаты доказывают, что общим свойством нейронов в двигательных областях является реакция во время движений, в которых они не участвуют напрямую, возможно, в целях обучения».
2) Мы пересмотрели параграф в Обсуждении, где мы обсуждаем степень латерализации, обнаруженную в различных исследованиях. Теперь мы подчеркиваем отсутствие ясности относительно того, являются ли ответы цифр более латерализованными.
«По оценке с помощью фМРТ, отдельные воксели в моторной коре сохранили «предпочтение пальцев» (мизинец, средний или большой палец) на конечностях (Diedrichsen et al.
, 2013). […] Например, нейрон может реагировать при сгибании правого мизинца и разгибании левого мизинца». 2) Как вы могли подтвердить, что записи действительно были с M1, а не, например, с PMd? Вы искали низкие пороги ICMS? Это важно, поскольку двусторонние представительства в PMd могут сильно отличаться от M1.
Это хороший вопрос, особенно потому, что у одной обезьяны (E) некоторые записи почти наверняка были из каудальной части PMd (рядом с M1). Даже у обезьяны F несколько записей находились вблизи «пограничной зоны» (четкой границы, конечно, нет). Важно отметить, что мы нашли одни и те же свойства независимо от места записи. Теперь об этом прямо говорится в рукописи.
«Результаты были практически идентичными (медианные индексы предпочтения руки 0,06 и 0,31), если анализ был ограничен более задними записями (исключая влияние любых нейронов, зарегистрированных из PMd или из пограничной области между M1 и PMd)». [Для справки, при анализе всех записей индексы были 0,07 и 0,31.
] Мы показываем на изображении ответа автора 1 соответствующий рисунок (рис. 4), воспроизведенный с помощью этого ограниченного анализа. На изображении ответа автора 2 мы показываем ключевой анализ (ортогональность подпространств) после ограничения задними заусенцами. Результаты не изменились. При желании их можно добавить в качестве дополнительных цифр.
Мы также добавили в рукопись сведения о локализации записей. Это включает в себя новые подпанели на рисунке 1, показывающие расположение трепанационных отверстий (имея в виду, что многие проходы проходили значительно ниже показанных элементов поверхности и в борозды). Мы также сообщаем о пороговых значениях ICMS, которые обычно были низкими (10-40 ед., что согласуется с M1 проксимального отдела руки), за исключением одного переднего трепанационного отверстия у обезьяны E (75 ед., что согласуется с PMd, как отмечено выше). Добавленный текст находится в подразделе «Хирургические процедуры и нейронная запись».
Вариант рисунка 4: нейронные ответы не сильно латерализованы (ограничены нейронами из задних трепанационных отверстий).
( A ) Гистограммы индекса предпочтения руки для всех зарегистрированных нейронов из задних фрезевых отверстий. ( C ) Гистограммы, обобщающие для отдельных нейронов сходство ответов при выполнении задачи с ведомой и не ведомой рукой. Для каждого нейрона мы рассчитали корреляцию между этими двумя ответами. 9Гистограммы 1105 Black отображают распределение таких корреляций по всем нейронам. Оранжевая кривая : контроль, демонстрирующий, что высокие корреляции наблюдаются, как и ожидалось, при сравнении откликов в условиях верхнего и нижнего начала. Зеленая кривая : ожидаемое распределение, если нет особой связи между ответами, соответствующими двум плечам. Это было вычислено как корреляция между каждой несовпадающей парой нейронов.
https://doi.org/10.7554/eLife.461590,022Вариант рисунка 9: Действия, связанные с левой и правой рукой, лежат в ортогональных подпространствах.
( A ) Проекция активности популяции во время примерного условия, выполняемого левой рукой ( синий ) и того же поведения, выполняемого правой рукой ( красный ), при этом ПК были обнаружены только с использованием данных во время движения левой руки. Нейронная популяция включает все зарегистрированные нейроны обоих полушарий, ограниченные задними трепанационными отверстиями. Анализ рассматривает данные средних циклов. ПК были найдены с использованием одного поведенческого состояния, а показанные данные относятся к состояниям удерживания, спроецированным в пространство ПК: либо для условия с тем же направлением вращения педалей и рукой, но противоположным исходным положением, либо для состояния с тем же исходным положением и направление вращения педалей, но с другой стороны. Данные для обезьяны E. ( B ) Кумулятивная процентная дисперсия, объясненная в условиях, выполняемых левой рукой ( синий ) и условиях, выполняемых правой рукой ( красный ), с PC, найденными с использованием только данных во время движения левой руки.
https://doi. Нейронная популяция включает все зарегистрированные нейроны обоих полушарий, ограниченные задними трепанационными отверстиями. ПК были найдены с использованием одного поведенческого условия, а объясненная дисперсия оценивалась в условиях задержки: либо одно и то же направление вращения педалей и работающая рука, но противоположное исходное положение, либо одно и то же исходное положение и направление вращения педалей, но противоположная рабочая рука. Каждая трасса представляет собой отдельное условие обобщения (всего четыре на руку). Данные для Monkey E. ( C ) Распределение абсолютного значения весов нейронов на первые пять ПК «левого плеча». Синие следы: подразделения в правом полушарии (которые управляют левой рукой). Красный: юниты в левом полушарии (правая рука). Гистограммы объединяются по всем поведенческим состояниям, поэтому каждый нейрон вносит 4 состояния x 5 ПК = 20 весов. (G-L) Как в A-F, для Monkey F. org/10.7554/eLife.46159.023
3) Похоже, что все анализы проводились на основе усредненных данных. Мы считаем, что крайне важно, чтобы авторы показали, по-прежнему ли ортогональные подпространства надежно разделяют активность популяции в отдельных испытаниях. Необходимо, чтобы однократная активность также попадала в нуль-пространство для контралатеральной руки, чтобы она могла избежать генерации контралатеральной активности. Хотя ваш анализ показывает, что в среднем активность попадает в нулевое пространство, мы считаем, что было бы информативно увидеть распространение проекции. Это можно сделать, спроецировав одиночные испытания на ортогональные подпространства и показав, что информация там есть, или применив методы уменьшения размерности с одним испытанием.
Это хорошее предложение. Немного сложно анализировать отдельные испытания, потому что мы одновременно регистрировали лишь небольшое количество нейронов: 8–47 (обезьяна E) и 12–64 (обезьяна F).
Это является следствием желания использовать v-зонды (а не массивы Юта) для доступа к центральной борозде и обеспечения гибкой записи из ряда мест (что дает более высокое общее количество нейронов, которое может быть достигнуто с массивами Юта). Тем не менее, в лучшие дни количество зарегистрированных нейронов приближалось к количеству хороших выделений на массиве Юты, что позволяло проводить анализ с одной пробой. Теперь мы включаем анализ отдельных испытаний (с использованием LFADS), чтобы выяснить, действительно ли средние значения испытаний являются репрезентативными. Мы также решаем этот вопрос, используя фактор Фано, чтобы спросить, есть ли доказательства «дополнительной» изменчивости, помимо той, что ожидается от грубой статистики Пуассона. Эти анализы описаны в новом разделе (Нейронные траектории в одиночных испытаниях) и на новом рисунке (рис. 10). Результаты показывают, что отдельные испытания ведут себя аналогично средним значениям испытаний; деятельность, связанная с двумя руками, в значительной степени разделена на разные подпространства.
В соответствии с этим фактор Фано ниже единицы. (В этом контексте было бы интересно узнать, можно ли предсказать мышечную активность в одиночной попытке в неведомой руке по отклонениям от ортогональных подпространств. Насколько я понял ограничения данных, это невозможно проверить. в текущем наборе данных, так как мышечная активность и мозговая активность регистрировались отдельно.Я не прошу авторов делать это, но хотел бы отметить, что это может значительно усилить гипотезу авторов.)
Это действительно было бы полезно знать. Мы предполагаем, что рецензент прав, и что небольшие однократные активации мышц неработающей руки связаны с небольшими экскурсиями в подпространство «неправильной» руки. Для обнаружения этого, конечно, потребуются одновременные записи мышц и нейронов (которых мы не делали) и, вероятно, также записи с гораздо большим числом нейронов (учитывая, что соответствующие отклонения, вероятно, будут небольшими). Тем не менее, это ясное предсказание, которое можно будет проверить в будущем, и мы согласны с тем, что его следует сформулировать.
Теперь делаем так: «Наши результаты согласуются с гипотезой о том, что нервная активность, связанная с правой рукой, падает в размерах, которые являются нулевыми по отношению к левой руке, и наоборот. Эта гипотеза делает предположение, которое можно проверить с помощью одновременных ЭМГ и крупномасштабных нейронных записей: небольшие сокращения в неработающей руке должны коррелировать с небольшими отклонениями в соответствующем подпространстве».
4) Как отмечалось в Обсуждении, активность в одном полушарии М1 мало влияет на мышечную активность в неведомой руке. Тем не менее, уменьшение размерности и прогнозный анализ на рисунках 9и 10 учитывают полную популяцию в разных полушариях. Чтобы ответить на вопрос, почему популяционная активность в одном полушарии не активирует неуправляемую руку, анализ уменьшения размерности и прогнозирования в идеале должен выполняться на одном полушарии. Эффект может быть слабее (из-за меньшего количества данных), но он все равно должен быть.
В качестве альтернативы авторы могли бы продемонстрировать, что веса декодирования (или веса, характеризующие подпространства) должным образом разделяют информацию между двумя полушариями. Мы согласны с тем, что важно показать, что то, что справедливо для всего населения, справедливо и для каждого полушария. Это действительно так, и теперь это продемонстрировано четырьмя дополнительными рисунками. Мы добавили два дополнительных рисунка, которые воспроизводят анализ на рисунке 9 для каждого полушария в отдельности (рисунок 9 — дополнение к рисунку 1, 2). Мы также добавили два дополнительных рисунка, воспроизводящих результаты, показанные на рисунке 11 (ранее рисунок 10) для каждого полушария в отдельности (рисунок 11 — рисунок дополняет 1, 2). Эффекты по-прежнему очень сильны. Эти дополнительные цифры также позволяют оценить дополнительный момент: способность игнорировать активность, связанную с «неправильной рукой», облегчается тем фактом, что не только подпространства ортогональны, но и активность для «правильной» руки немного выше.
Таким образом, разделение по подпространствам особенно четко для наиболее релевантного подпространства (например, подпространство левого плеча для правой коры, как показано в верхнем ряду на рис. 9).— приложение к рисунку 1). Нам нравятся эти дополнительные анализы, но мы сохранили анализ в основных цифрах, ориентированный на все население как по практическим, так и по концептуальным причинам. На практике это позволяет использовать меньше субпанелей (например, на рисунке 9 может быть 12 субпанелей, а не 24). Концептуально, на этом этапе мы хотим перевести разговор в сторону размышлений о моторной коре как о полной популяции. Результаты на более ранних рисунках показывают, что обе стороны реагируют во время движения обеих рук и содержат аналогичные сигналы. Анатомически обе стороны сильно взаимосвязаны. Таким образом, становится разумным спросить: «Если все релевантные сигналы являются общими для большой двусторонней популяции, как могут быть разделены сигналы, специфичные для рук».
две руки могут быть разделены без необходимости знать, с какой стороны был записан каждый нейрон. Мы изменили начало соответствующего раздела, чтобы отразить эту мотивацию. Мы также согласны с тем, что к соответствующим цифрам (сейчас 9 и 11) стоит добавить веса подпространства/декодирования. Это было сделано. Как и ожидалось, оба полушария вносят вклад в оба пространства/декодирования (с ожидаемым небольшим смещением управляющей коры).
5) Даже внутри одного полушария дополнительная проблема заключается в том, что может существовать смесь типов клеток, некоторые из которых отвечают за прямой двигательный контроль (например, небольшая доля клеток с индексом предпочтения, близким к 1), а некоторые отвечают за общий расчет, скажем. Предыдущая работа с ALM на мышах показала, что такое разделение может существовать со связанными с ним анатомическими различиями в предпочтениях левых и правых (Li et al., 2015, Nature 519).51-56). Хотелось бы, чтобы авторы (как минимум) выводили некоторую информацию о весах декодеров или подпространств (см.
также пункт 2). Мы согласны с тем, что интересно рассмотреть, могут ли быть подгруппы, которые более точно соответствуют конкретным параметрам вычисления. Теперь мы показываем распределения проекционных весов для PCA на рис. 9 и весов декодирования EMG на рис. нейроны руки). Существует несколько непрерывное распределение. Тем не менее, мы не хотели бы полностью исключать эту гипотезу. Например, кажется вероятным, что кортико-мотонейроны могут быть более «чистыми» и, таким образом, занимать хвост распределения.
6) Авторы используют PLS, чтобы найти низкоразмерное линейное отображение между активностью популяции X и активностью популяции Y, и оценили его в параметрах обобщения, насколько хорошо можно предсказывать новые данные. Основные результаты заключаются в том, что мышечная активность может быть почти одинаково хорошо предсказана по контралатеральному и ипсилатеральному M1, и что едва ли есть какие-либо различия в предсказании левого M1 по левому M1 и по правому M1.
Основное ограничение этого анализа заключается в том, что он может показать, что два кода совокупности занимают общее линейное подпространство, но не показывает, что два кода совокупности структурно одинаковы (или имеют одинаковую репрезентативную геометрию — Дидрихсен и Кригескорте, 2017). . Например, рассмотрим две популяции, изображенные на Рисунке 1. Обе имеют нейронное измерение, которое кодирует вертикальное положение руки, и нейронное измерение, которое кодирует горизонтальное положение руки (конечно, нейронные измерения не точно отражают конкретное положение руки). физические переменные, но речь здесь не об этом). Это означает, что вы можете найти двумерное отображение, которое будет предсказывать популяционную активность в регионе А по региону В, вероятно, почти так же хорошо, как вы могли бы предсказать регион А по региону А. Кроме того, специфическая мышечная активность, которая представляет собой линейную комбинацию положение можно одинаково хорошо (или плохо) считывать из активности населения.
Однако это затемняет тот факт, что область А придает чрезмерное значение вертикальному измерению движения, тогда как область В делает упор на кодирование горизонтального измерения. Эта проблема особенно распространена, поскольку лежащее в ее основе поведение относительно простое. В то время как направление велосипедного движения разделяет положение, скорость и мышечную активность, две исходные точки, вероятно, очень мало делают для добавления новых измерений, которые функциональное подпространство должно кодировать в мозгу. Таким образом, пока оба полушария занимают это относительно ограниченное подпространство нейронной активности, прогнозирование модели PLS будет достаточно хорошим. То есть два очень разных кода совокупности могут выглядеть очень похожими, если в ходе эксперимента не будут диссоциированы критические измерения.
Мы считаем, что авторы должны, по крайней мере, признать эти два недостатка своего регрессионного подхода.
Отличные очки.
Мы согласны с тем, что подход PLS имеет два заметных ограничения. Во-первых, PLS не различает два ответа популяции, которые «растянуты» друг относительно друга (т. е. имеют одни и те же сигналы, но в совершенно разной степени). Во-вторых, простая задача может дать ограниченную возможность «видеть» разные измерения. Например, более сложная задача может вызвать 50-мерную реакцию, при этом первые десять измерений будут общими для ведущего и не ведущего полушарий, а остальные будут совершенно разными. Таким образом, мы должны быть осторожны, подчеркивая, что наш вывод состоит просто в том, что сигналы, переносимые доминирующими измерениями, очень похожи. Мы внесли следующие изменения: 1) Мы изменили наш анализ, чтобы устранить первое ограничение. Теперь мы используем PCA-регрессию, а не PLS-регрессию. Чтобы понять, почему PCA-регрессия устраняет это ограничение, рассмотрим сценарий типа, описанного рецензентами: сигналы X1 и Χ2 присутствуют в обоих полушариях, но X1 выделяется в ведущей коре, а Χ2 — в не ведущей коре.
Если мы применим PCA к популяции управляющей коры и спроецируем на первый PC, мы восстановим X1. Когда мы регрессируем активность ведущей коры относительно X1, она фиксирует значительную дисперсию (скажем, 60%). Если мы применим PCA к популяции неведущей коры и спроецируем на первый PC, мы восстановим Χ2. Когда мы регрессируем популяцию управляющей коры относительно Χ2, она будет охватывать гораздо меньшую дисперсию (скажем, 30%). Таким образом, регрессия PCA, в отличие от регрессии PLS, выявит несоответствие размеров сигналов. Поэтому теперь мы используем регрессию PCA для этого анализа (другие анализы все еще используют регрессию PLS). Мы также подтверждаем (Рисунок 7 — дополнение к рисунку 1), что этот анализ действительно выявил бы различия, если бы они были. Соответствующие панели рисунков (рис. 7E-G) были обновлены, как и соответствующий текст. (Мы также продолжаем отмечать, для полноты картины, что регрессия PLS дала аналогичные результаты.) 2) Мы добавили следующий текст, чтобы явно признать второе ограничение:
«Таким образом, доминирующие сигналы — те, что присутствуют в верхней горстке главных компонентов — почти идентичны между управляющей и неуправляющей корой.
Предостережение заключается в том, что меньшие сигналы могут быть совершенно другими, что было бы трудно различить, учитывая нашу относительно простую задачу. Активность охватывает скромное количество измерений при циклировании в устойчивом состоянии (на 5 основных компонентов приходится ~ 80% дисперсии). Вполне возможно, что различия в меньших сигналах, переносимых другими измерениями, могут быть обнаружены при дополнительной сложности задачи (например, при езде на велосипеде на разных скоростях или при разных нагрузках)». https://doi.org/10.7554/eLife.46159.027
Статья и информация об авторе
Сведения об авторе
-
Кэтрин Кора Эймс
- Факультет неврологии, Колумбийский университет, Нью-Йорк, США
- Институт Цукермана, Колумбийский университет, Нью-Йорк, США
- Центр статистики сознания Гроссмана, Колумбийский университет, Нью-Йорк, США
- Центр теоретической неврологии Колумбийского университета, Нью-Йорк, США
Взнос
Концептуализация, обработка данных, программное обеспечение, формальный анализ, исследование, визуализация, методология, написание — первоначальный проект, написание — просмотр и редактированиеДля корреспонденции
kca2120@columbia. edu
Конкурирующие интересы
Конкурирующие интересы не заявлены «Этот ORCID iD идентифицирует автора этой статьи:» 0000-0003-0657-0162 -
Марк М. Черчленд
- Департамент неврологии, Колумбийский университет, Нью-Йорк, США
- Институт Цукермана, Колумбийский университет, Нью-Йорк, США
- Центр статистики сознания Гроссмана, Колумбийский университет, Нью-Йорк, США
- Институт изучения мозга Кавли, Колумбийский университет, Нью-Йорк, США
Взнос
Концептуализация, надзор, получение финансирования, методология, администрирование проекта, написание — обзор и редактированиеКонкурирующие интересы
Конкурирующие интересы не заявлены «Этот ORCID iD идентифицирует автора этой статьи:» 0000-0001-9123-6526
Финансирование
Национальный институт неврологических расстройств и инсульта (1DP2NS083037)
- Марк М Черчленд
Фонд Саймонса (SCGB#325233)
- Марк М Черчленд
Фонд Макнайта (P30 EY019007)
- Марк М Черчленд
Товарищество Клингенштейна-Саймонса
- Марк М Черчленд
Программа стипендий Серла
- Марк М Черчленд
Национальные институты здравоохранения (CRCNS R01 NS100066)
- Марк М Черчленд
Национальный институт неврологических расстройств и инсульта (1U19NS104649)
- Марк М Черчленд
Фонд Саймонса (SCGB#542957)
- Марк М Черчленд
Центр статистики сознания Гроссмана
- Марк М Черчленд
Фонд Саймонса (постдокторская стипендия SCGB)
- Кэтрин Кора Эймс
Спонсоры не участвовали в разработке исследования, сборе и интерпретации данных или принятии решения о представлении работы для публикации.
Благодарности
Благодарим Ю. Павлову за квалифицированный уход за животными. Руссо предоставил код для расчета запутывания траектории. Эта работа была поддержана NINDS 1DP2NS083037, NIH CRCNS R01NS100066, NINDS 1U19NS104649, Фондом Саймонса (SCGB#325233 и SCGB#542957), Центром статистики сознания Гроссмана, Фондом Макнайта, P30 EY019007, Товариществом Клингенштейна-Саймонса, и Программа ученых Серла. KCA финансировалось сотрудничеством Фонда Саймона в рамках стипендии Global Brain Post-doctor.
Этика
Эксперименты на животных: это исследование было проведено в соответствии с рекомендациями, изложенными в Руководстве по уходу и использованию лабораторных животных Национального института здравоохранения. Все протоколы животных были одобрены Институциональным комитетом по уходу и использованию животных Колумбийского университета (номер протокола AC-AAAQ7409).
Старший редактор
- Джошуа I Голд, Пенсильванский университет, США
Рецензирующий редактор
- Йорн Дидрихсен, Университет Западного Онтарио, Канада
рецензентов
- Йорн Дидрихсен, Университет Западного Онтарио, Канада
- Кристиан К. Маченс, Центр неизвестного Шампалимо, Португалия
История публикаций
- Получено: 16 февраля 2019 г.
- Принято: 24 сентября 2019 г.
- Версия записи опубликована: 9 октября, 2019 (версия 1)
Авторское право
© 2019, Ames and Churchland
Эта статья распространяется в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование и распространение при условии указания оригинального автора и источника.
Показатели
Число цитирований статей, полученное путем опроса самых высоких значений из следующих источников: Scopus, Crossref, PubMed Central.
Ссылки для скачивания
Список ссылок, состоящий из двух частей, для загрузки статьи или частей статьи в различных форматах.
Открытые цитаты (ссылки для открытия цитат из этой статьи в различных онлайн-сервисах управления ссылками)
- Менделей
- ЧитатьКуб»>
Процитировать эту статью (ссылки для загрузки цитат из этой статьи в форматах, совместимых с различными инструментами управления ссылками)
- Кэтрин Кора Эймс
- Марк М Черчленд
(2019)
Сигналы моторной коры для каждой руки смешиваются между полушариями и нейронами, но разделяются в ответе популяции
eLife 8 :e46159.
https://doi.org/10.7554/eLife.46159
- Скачать БибТекс
- Скачать .RIS
Категории и теги
- Исследовательская статья
- Неврология
- моторная кора
- движение руки
- двуручный
- государственный космический
Исследовательский организм
- Макак-резус
МОЗГ ОТ ВЕРХА К НИЗУ
Финансирование этого сайта осуществляется такими читателями, как вы. Making a Voluntary Movement
Бродманн Районы
В течение всего срока службы различные детали мозга, которые контролируют движения тела, могут пострадать от различных причинами, такими как травмы головы, инсульты и дегенеративные заболевания головного мозга.
Один типичный
Например, что происходит, когда одна сторона моторной коры полностью разрушена.
по инсульту. Из-за скрещенных
контроль, этот центр обычно контролирует произвольные движения на
противоположная сторона тела. Когда этот центр разрушен, дальнейшее движение невозможно;
эта сторона тела становится полностью парализованной. МОТОРНАЯ КОРА движения контролируются мозгом. Одна из областей мозга, наиболее вовлеченная в Эти произвольные движения контролируются моторной корой.
моторная кора расположена в задней части лобной доли, непосредственно перед центральная борозда (борозда), отделяющая лобную долю от теменной доли. Моторная кора разделена на две основные области: область 4 и область 6. Область 4 также
известный как первичный двигатель кора , образует тонкую полосу вдоль
центральная борозда. Зона 6 находится непосредственно перед Зоной 4. Зона 6 шире.
и является
далее подразделяется на две самостоятельные подобласти. Кому осуществлять целенаправленные движения, ваши моторная кора должна сначала получать различного рода информацию от различных доли головного мозга: информация о положении тела в пространстве, от теменная доля; о цели, которая должна быть достигнута, и соответствующей стратегии для достигая его, от переднего отдела лобной доли; о воспоминаниях о прошлые стратегии из височной доли; и так далее.
В 1870 году Хитциг и Фрич электрически стимулировали различные участки моторной коры собаки. Они заметили, что в зависимости какую часть коры они возбуждали, другая часть тела сокращалась.
Затем они обнаружили, что если разрушить этот же небольшой участок коры, то
соответствующая часть тела была парализована. Вот как это было обнаружено
что каждая часть тела имеет определенную область первичной моторной коры
который управляет его движением. Но что примечательного в этом моторе » карта» состоит в том, что определенные части тела – те, которые могут тончайшие движениязанимают гораздо больше места, чем другие. Эти части тела показаны больше, чем другие на иллюстрации здесь.
./../../../d_img/d_vl_right.gif»>
The putamen and the caudate nucleus проходят миелинизированные аксоны внутренней капсулы. Эти пучки белое вещество образует полосы, отличающие их от серого вещества ядер которые они пересекают. Вот почему эта группа нервных структур, вместе взятых часто упоминается как полосатое тело, или полосатое тело.
Точно так же форма скорлупы и бледного шара напоминает форму линза, поэтому эти два ядра вместе известны как чечевицеобразный ядро .
Обучение Как возбудить любопытство
БАЗАЛЬНЫЕ ГАНГЛИИ
Как их название предполагает, что базальные ганглии состоят набора нервных структур, спрятанных глубоко внутри головного мозга. Основной базальный ганглиями являются хвостатое ядро, скорлупа и бледный шар.
Эти ганглии или скопления нервных клеток тесно взаимосвязаны. Они также получают информацию из нескольких разных регионов. коры головного мозга. Как только базальные ганглии обработали эту информацию, они возвращают его в моторную кору через таламус.
Один вероятных функций этой петли, которая действует совместно с другим, задействующим мозжечок, заключается в выборе и вызывают хорошо скоординированные произвольные движения.
Эта роль базальные ганглии в инициировании и регуляции двигательных команд становятся ясно выраженными у людей с повреждением базальных ганглиев, например у пациентов с болезнью Паркинсона болезнь. Эти пациенты испытывают трудности с началом движений, которые у них есть. запланированные, а также дрожь и медлительность, как только они начинают их.
The cerebellum is composed of a количество долей и долек которые, как и извилины в мозговых коры, значительно увеличивают площадь поверхности коры мозжечка.
Этот
большая площадь поверхности серого вещества обеспечивает очень высокую плотность мозжечка
нейронов настолько высока, что мозжечок, на долю которого приходится лишь около 10%
от общего объема мозга, составляет более 50% его нейронов!
Анатомическое расположение мозжечка помогает нам лучше понять его функции. Лежит параллельно к двум основным нервным путям: один, который несет сенсорные сообщения в часть мозг, который их анализирует, и другой, который выходит из коры и спускается к мышцам, чтобы заставить их сокращаться.
Таким образом, мозжечок получает копия всей информации, поступающей от органов чувств к органам чувств. кору и всю информацию, которая передается из моторной коры в спинной мозг. Мозжечок также получает информацию из многих других областей мозга. кору больших полушарий и подкорковые отделы головного мозга.
THE CEREBELLUM
For даже такой простой жест, как прикосновение к кончику носа, недостаточно, чтобы ваш мозг просто приказал мышцам рук и рук сокращаться. Чтобы различные сегменты кисти и предплечья работали плавно, вам нужен внутренние «часы», которые могут точно регулировать последовательность и продолжительность элементарных движений каждого из этих сегментов. Эти часы — мозжечок. Как это часто бывает в нейробиологии, чтобы точно понять, что делает мозжечок, мы можем наблюдать пациентов, у которых часть этой структуры разрушена (например, опухолью или инсультом).
Когда эти больные пытаются схватить предмет, их руки начинают двигаться с опозданием, выдвигаются вперед.
неустойчиво и либо останавливаются, не достигнув цели, либо, часто, ускоряются
мимо него. Что касается осанки, то для людей с поврежденным мозжечком характерно
проявляют проблемы с балансом, подобные тем, которые обнаруживаются у людей, которые находятся в состоянии алкогольного опьянения. Фактически,
неуклюжесть, сопровождающая чрезмерное употребление алкоголя
непосредственно связано с его депрессивным влиянием на деятельность мозжечка.
У здорового человека мозжечок сначала получает информацию о предполагаемом движении от сенсорных и моторных коры. Затем он посылает информацию обратно в моторную кору о требуемом направление, сила и продолжительность этого движения. петля, включающая мозжечок, работает в дополнение к петля, включающая базальные ганглии, чтобы регулировать детали моторного контроля. Еще одна метафора резюмирует роль вашего мозжечка. довольно хорошо: он действует как авиадиспетчер, который собирает невероятные количество информации в каждый момент, в том числе (чтобы вернуться к нашему исходному примеру) положение вашей руки, вашей руки и вашего носа, скорость их движений, и последствия потенциальных препятствий на их пути, так что ваш палец может добиться «мягкой посадки» на кончик носа.
./../../../d_img/d_vl_left.gif»>
Производство механизма организовано в различные уровни управления . На высшем уровне кора контролирует произвольные движения. Это все движения, которые требуют координации и точность адаптации к конкретным ситуациям на основе предоставленной информации по ощущениям.
На самом базовом уровне движение контролируется только спинной мозг, без помощи головного мозга. Нейроны спинного мозга таким образом взять на себя ответственность за рефлекс движения, а также ритмические движения при ходьбе.
Между этими двумя уровнями есть все другие виды движений.
Например,
как и движения, связанные с ходьбой, движения, связанные с дыханием,
автоматический компонент, но также может быть изменен по желанию (например, если вы
хотите, вы можете задерживать дыхание, точно так же, как вы можете бегать вместо ходьбы). Последовательность активации для моторных областей Основная функция мозг должен производить действия, которые в первую очередь являются движениями. Несколько разных регионов коры головного мозга участвуют в управлении движениями тела.
Эти регионы организованы в иерархию, как экипаж корабля. На древняя галера, например, капитан определял место плавания путем оценки различных факторов, которые могут сделать такую поездку стоящей. затем его лейтенанты рассчитали направление, в котором должен был двигаться корабль, чтобы достичь этого пункта назначения в зависимости от погодных условий.
Наконец лейтенанты передали
отдавали приказы экипажу на веслах, которые использовали свои мускулы, чтобы двигать лодку.
корабль в нужном направлении. Точно так же в человеческом мозгу планирование для любого заданного движения делается в основном в передней части лобной доля. Эта часть коры получает информацию о текущем состоянии человека. позицию из нескольких других частей. Затем, подобно капитану корабля, команды, в Зону 6. Зона 6 действует как лейтенанты корабля. Это решает какой набор мышц нужно сократить для достижения необходимого движения, затем отдает соответствующие распоряжения «гребцам» – основной двигатель коры, также известной как область 4. Эта область , в свою очередь, активирует определенные мышцы или группы мышц через двигательные нейроны спинного мозга. Даже для такого простого движения, как поднятие стакан воды, вряд ли можно себе представить попытку сознательно указать последовательность, сила, амплитуда и скорость сокращений каждой задействованной мышцы.
А также
тем не менее, если мы здоровы, мы все делаем такие движения все время, даже не задумываясь
из них. Решение взять стакан воды сопровождается повышенная электрическая активность в лобной области коры. Нейроны в лобной коре затем посылают импульсы вниз по их аксонам, чтобы активировать моторную самой коры. Используя информацию, поступающую от зрительной коры, двигательная кора головного мозга планирует идеальный путь, по которому рука дотянется до стакана. Мотор Затем кора обращается к другим частям мозга, таким как центральная серые ядра и мозжечок, которые помогают инициировать и координировать активация мышц по очереди.
Аксоны нейронов первичная моторная кора опускается вплоть до спинного мозга, где происходит последняя передача информации к двигательным нейронам спинного мозга. Эти нейроны связаны напрямую к мышцам и заставляют их сокращаться. Наконец, заключая контракт и тем самым натягивая кости руки и кисти, мышцы выполняют движение, которое позволяет поднять стекло.
Кроме того, чтобы убедиться, что все эти движения быстрые, точные и скоординированные, нервная система должна постоянно получать сенсорную информацию из внешнего мира и использовать эту информацию для регулировать и корректировать траекторию движения руки. Нервная система достигает этих приспособлений главным образом с помощью мозжечка, который получает информацию о положениях в пространстве суставов и тела от проприорецепторы.
Презентации | Кредиты | Контакт | Копилефт ОЦЕНКА МОТОРНОЙ ФУНКЦИИ | LHSC
EDUBRIEFS в CCTC
Оценка двигательной функции
Как контролируется двигательная функция?
Произвольное движение требует передачи сообщения от двигательной полосы коры головного мозга (верхний мотонейрон) к соответствующей мышце на противоположной стороне тела.
Таким образом, повреждение коры головного мозга вызывает снижение мышечной функции контралатерально (на стороне тела, противоположной повреждению головного мозга). В то время как способность приказать мышце двигаться является функцией коры головного мозга, способность сделать движение плавным и скоординированным требует мозжечка. Шаткие или нескоординированные движения могут быть результатом дисфункции мозжечка. Мозжечок контролирует плавность движения на той же стороне (сообщение о движении левой ноги исходит из правой коры головного мозга, но координируется и сглаживается левым полушарием мозжечка). Как оценивается двигательная функция?
- Если двигательная функция не нарушена, мышцы можно двигать по команде. Симметричное движение и сила являются одним из наиболее важных результатов оценки. Снижение двигательной функции может возникнуть в результате повреждения коры головного мозга, двигательных путей, периферических нервов или мышц. В то время как для движения мышцы по команде требуется определенный уровень функционирования, для преодоления гравитации требуется повышенная иннервация и мышечная сила. Еще большая сила требуется, чтобы преодолеть сопротивление экзаменатора.
- Оценка двигательной функции у пациентов, способных подчиняться командам, может быть проведена следующим образом (справа по сравнению с левой):
5 = нормальная сила (нормальная сила, способность поддерживать мышечное сокращение при сопротивлении исследователя)
4 = легкая слабость (слабая или кратковременная способность преодолевать сопротивление исследователя)
3 = способность поддерживать конечность при сопротивлении, но неспособность преодолевать сопротивление исследователя
2 = может двигать конечностью, но не может подняться против силы тяжести
1 = мерцает, но движения нет
0 = нет движения
Как можно быстро оценить двигательную функцию пациента в сознании?
- В то время как интенсивная оценка может быть выполнена для каждой группы мышц, быстрый способ определить двигательную слабость — это оценка дрейфа конечностей.
Сила верхних конечностей
Человек в сознании
Попросите пациента вытянуть руки горизонтально ладонями вверх с закрытыми глазами. Если есть слабость в верхней конечности, пораженная сторона «дрейфует» или пронируется в течение 30 секунд.Сила нижних конечностей
Человек в сознании
Когда пациент лежит на спине, согните колени на 30 градусов. Если есть слабость в нижних конечностях, пораженная нога будет дрейфовать вниз в течение 30 секунд.
- Когда пациент лежит на спине, согните оба колена и поддерживайте одну из рук исследователя. Оставьте одну пятку на кровати. Вытяните другую ногу в колене и дайте ей мягко опуститься на кровать. Сравните скорость падения для обеих ног.
Как можно быстро оценить двигательную функцию пациента, находящегося без сознания?
- Сила верхних конечностей
Человек без сознанияПоднимите обе руки пациента вместе.
Защищая конечности от травм, отпустите обе руки вместе. Парализованная рука упадет быстрее.
- Сила нижних конечностей
Человек без сознания
- Положение пациента на спине. Согните колени, поставив обе ноги на кровать. Отпустите колени одновременно. Парализованная нога упадет в вытянутое положение, а бедро будет вращаться наружу. Нормальная нога остается согнутой в течение нескольких секунд и постепенно принимает прежнее положение.
- Какие другие наблюдения можно сделать для оценки двигательной функции у пациента без сознания?
- Наблюдайте за пациентом, когда он делает спонтанные движения. Обратите внимание на симметричность движений. Если человек не отвечает на команду, но делает целенаправленные движения, такие как дергание за линии или трубы, реакция называется 9.1105 локализация. Это адекватная реакция, требующая функциональных двигательных путей.
- Если спонтанных движений не наблюдается, проведите стимуляцию центральной боли . Центральную боль можно проверить, растирая грудину, сдавливая ткань в подмышечной впадине, сжимая трапециевидную мышцу в области шеи и плеча или прикладывая супраорбитальное давление (избегайте при наличии переломов лица). Меняйте технику, чтобы избежать синяков или травм тканей. Если получен ответ на центральную боль, периферическая стимуляция не нужен. Периферическая боль может вызывать спинальный рефлекс и, следовательно, не является эффективным тестом функции верхнего двигательного нейрона.
- Изъятие может быть описано наличием нормального сгибания в ответ на боль. Оттягивание без сгибания запястья — это один из способов отличить нормальное сгибание от ненормального сгибания. Жесткое сгибание считается ненормальным сгибанием. Жесткое расширение упоминается как ненормальное расширение . Отсутствие движения или тонуса называется вялым параличом.
- Какие другие обследования оценивают двигательную функцию?
- В дополнение к оценке силы, описанной выше, следует осмотреть и пропальпировать мышцы. Осмотрите на предмет асимметричного движения или неправильного вращения конечности. Пальпируйте мышцу на предмет уменьшения (вялости) или увеличения (спастичности). Снижение тонуса может свидетельствовать о раннем повреждении верхнего мотонейрона или периферического нерва. Повышенный тонус связан с повреждением верхних двигательных нейронов.
-
Бренда Морган, клинический педагог
19 ноября 1999 г. - Обновлено: 15 января 2019 г.
Ссылки:
Бейтс, Б. (1983). Руководство по физическому осмотру. (3-е издание). Филадельфия: Липпинкотт. стр. 411.
Прайс, М., и Деврум, Х. (1985). Краткое и простое руководство по неврологической оценке. Журнал нейронауки сестринского дела.
Похожие записи
 .
.