Что такое минископ и для чего он используется. Какие модели минископов существуют. Как применяются минископы при таможенном контроле. Какие преимущества и недостатки имеют разные модели минископов.
Что такое минископ и его основные характеристики
Минископ — это компактный портативный микроскоп, предназначенный для оперативного увеличения и детального рассмотрения мелких объектов. Основные характеристики минископов:
- Увеличение: как правило, от 20х до 100х
- Компактные размеры: длина 10-15 см, диаметр 1-2 см
- Легкий вес: 30-100 г
- Возможность работы одной рукой
- Наличие подсветки у некоторых моделей
Минископы широко применяются в криминалистике, таможенном контроле, нумизматике и других областях, где требуется быстрый осмотр мелких деталей.
Сравнение популярных моделей минископов
Рассмотрим характеристики нескольких распространенных моделей минископов:
Минископ модели 1171
- Увеличение: 30х
- Линейное поле зрения: 5 мм
- Размеры: длина 125 мм, диаметр 15 мм
- Отсутствует встроенная подсветка
- Перевернутое изображение
Микроскоп FF-393
- Увеличение: 30х
- Размеры: 140х48х22 мм
- Вес: 30 г
- Встроенная подсветка, работающая до 3 часов
- Возможность работы при естественном освещении
- Фокусировочное кольцо
Минископ Steiner Miniscope 8×22
- Увеличение: 8х
- Диаметр объектива: 22 мм
- Размеры: 51x49x46 мм
- Вес: 80 г
- Возможность складывания
- Минимальная дистанция фокусировки: 4 м
Применение минископов в таможенном контроле
Минископы активно используются сотрудниками таможни для проверки подлинности документов и атрибутов таможенного обеспечения. Основные задачи применения минископов в таможенном деле:

- Проверка защитных элементов документов (микропечать, микроузоры)
- Выявление следов подчистки и травления в документах
- Контроль целостности пломб и печатей
- Осмотр маркировки товаров
- Проверка подлинности акцизных и специальных марок
Компактность минископов позволяет сотрудникам таможни всегда иметь их при себе и оперативно проводить проверку при возникновении подозрений.
Преимущества и недостатки разных моделей минископов
У каждой модели минископа есть свои сильные и слабые стороны:
Минископ модели 1171
Преимущества:
- Компактность
- Простота конструкции
Недостатки:
- Отсутствие подсветки
- Перевернутое изображение
- Сложность использования при осмотре пломб
Микроскоп FF-393
Преимущества:
- Наличие подсветки
- Возможность фокусировки
- Удобство использования
Недостатки:
- Больший размер по сравнению с моделью 1171
Минископ Steiner Miniscope 8×22
Преимущества:
- Возможность складывания
- Широкий обзор
- Высокое качество оптики
Недостатки:
- Меньшее увеличение
- Большая минимальная дистанция фокусировки
Советы по выбору минископа для таможенного контроля
При выборе минископа для использования в таможенном деле следует учитывать несколько факторов:

- Увеличение: оптимально 30-60х для проверки документов
- Наличие подсветки: желательно для работы в условиях недостаточного освещения
- Компактность: прибор должен легко помещаться в кармане
- Простота использования: возможность работы одной рукой
- Качество оптики: четкое изображение без искажений
Исходя из этих критериев, для таможенного контроля больше всего подходят модели типа FF-393, сочетающие достаточное увеличение, подсветку и удобство использования.
Дополнительное оборудование для проверки документов в таможне
Помимо минископов, для проверки подлинности документов в таможенных органах используются и другие технические средства:
- Ультрафиолетовые осветители для выявления люминесцирующих защитных элементов
- Лупы с подсветкой для общего осмотра документов
- Приборы для проверки машиносчитываемой зоны паспортов
- Видеоспектральные компараторы для сложных исследований
- Портативные детекторы для проверки акцизных марок
Комплексное применение различных технических средств позволяет повысить эффективность выявления поддельных документов при таможенном контроле.

Перспективы развития портативных микроскопов для таможенных нужд
Развитие технологий открывает новые возможности для совершенствования минископов и других портативных средств контроля документов:
- Интеграция с мобильными устройствами для передачи изображений
- Применение технологий машинного зрения для автоматического выявления признаков подделки
- Использование спектральных методов анализа материалов документов
- Создание многофункциональных устройств, сочетающих функции микроскопа, УФ-детектора и других приборов
- Разработка специализированного программного обеспечения для анализа изображений
Внедрение таких инноваций позволит вывести оперативный контроль документов на таможне на новый уровень эффективности и надежности.
Сравнение двух минископов, Переделываем стандартное айфоновское крепление под своей телефон. Special Microscope 60X Currency Detecting with LED Microscope for iPhone 4G 4S HMM-84759
Всем привет:) решил сделать сравнение двух микроскопов (как их гордо называются китайцы) а по факту минископов, заодно и попробуем приделать его к камере своего телефона. Обзор получился излишние переполненный фото и текстом, уж простите, но и так ужимал как мог.
Был у меня один микроскоп, вот такой www.tinydeal.com/60x-100x-zoom-pocket-illuminated-microscope-p-12731.html
Все хорошо, увеличивает, можно менять кратность, но проблема что на телефон зацепить возможности нет, поэтому приобрел еще один, с креплением на айфон, думал переделаю крепление под свой, и буду цеплять свой черный микроскопик, как оказалось я ошибался:)
ограничюсь общим фото:
И наконецто перейдем к сути:
Для начала попробуем примастырить микроскопы к телефону, дабы облегчить наглядное измерение кратности.
В комплекте с первым минископоп (назовем его условно «Белый» для простоты восприятия)
идет вот такой фиксатор переходник:
Телефон у меня devdb.ru/motorola_atrix_4g
Камера расположена с той же стороны что и у айфона, но чуть смещена вниз, и сам телефон толще и шире.
Для подгонки нам понадобиться
нож
и моя любимая горелочка с динодиректа.
Принцип простой, лишнее срезаем, оставшиеся нагреваем гладящими движениями, вытягиваем руками, опять нагреваем, быстро одеваем на телефон, пальцами пригибаем где надо, главный нюанс,
Подгоняем к окошку камеры, по возможности центруем:
Получаем вот такую кракозяблю в сравнении с новой:
(у меня два минископа таких, есть еще один переходник для сравнения.
В процессе слегка заплавил и деформировал разьем куда вставляется минископ (Держиться просто трением) Но не проблема, нагреваем, вставляем, все норм:)
В результате должно получить вот так:
Пол дела сделано, теперь можно без проблем получить вот такие фото:
Переходим к сложному этапу, придумать как зафиксировать мощный «черный» минископ, изначальная задумка не получилось, в фиксатор от белого не влазит, «разьемы» разные по диаметру:
Да еще и форма у черного какаято овальная. Чещем в затылки, думаем, думаем, придумали:) ищем старый жесткий чехол, сразу же прицеливаемся:
В принципе можно просто приклеить к чехлу минископ, но тогда не получиться использовать комфортно отдельно от телефона его. Так что делаем другой вариант: приклеиваем разьем к чехлу.
Нашел вот такую полителеновую хреновинку,
но она слишком мягкая, решил пойти нестандартным способом и «закалить» ее, для этого нагрел в кипятке, потом бросил в воду с льдом, и на удивлении получилось, полителен стал жестким:)
Обрезаем лишнее
Безопасность превыше всего, одеваем перчатки, подлаживаем на стол целофан, сверху бумагу т. к. цианокрилатный клей не застывает на целофане,
к. цианокрилатный клей не застывает на целофане,
Сам процесс наклейки:
Телефон кутаем в целофан, одеваем сверху чехол, капаем клей, центруем разьем, дышим на него, чтобы быстрее застыл.
и клееим, по кругу армируем изолентой и цианокрилатом, на чехол для жесткости клеим кусочек пластика от обычной бутылки.
Вставляем минископ:
Держиться собака:)))
Теперь можно попробывать оценить кратность, фотаем линейку:
Делаем снимок на минимальном увелечении черного на той же высоте:
на максимальном:
Белый минископ:
Собстно расчеты (возможно я не прав, кто лучше разбирается поправьте):
Без увеличения мы видим 120мм линейки, в белом минископе видно 5мм, 120/5=24 получаем кратность 24? для черного 120/2,5 = 48 для черного с выдвинутым фокусом: 120/1,5 = 80. В прочем, есть еще зум у телефона, с зумом можно получить вот такую картинку с белого:
Практическое применение можно посмотреть в моем прошлом обзоре:
mysku. club/blog/tinydeal/16507.html
club/blog/tinydeal/16507.html
Спасибо за внимание:)
Понравился опус:)? хотите сделать автору приятно? нажмите «обзор понравился» и кнопку «планирую купить» вам мелочь, а мне приятно:)
Обзор монокуляра STEINER Miniscope 8 x 22
Здравствуйте, друзья! Сегодня мы познакомимся с весьма интересным биноклем. По сути даже не биноклем, а бинокликом, а если правильно его называть, то монокуляром, т.к. «глаз» у него один и картинка, соответственно не бинокулярная (не объемная). Но это не уменьшает его достоинств, т.к. это тот случай, когда важен маленький размер, ведь монокуляр STEINER Miniscope 8 x 22 предназначен для того, чтобы всегда иметь его под рукой.
STEINER miniscope 8 x 22
STEINER — немецкая компания, которая производит бинокли и оптические прицелы как для военного, так и для гражданского применения.
Внешний вид и конструктив
В комплекте к монокуляру идёт удобный чехольчик на липучке, что позволяет защищать монокуляр от лишних царапин. Особенно это важно т.к. у такого «малыша» нет защитных крышек на окуляр и на переднюю линзу.
Особенно это важно т.к. у такого «малыша» нет защитных крышек на окуляр и на переднюю линзу.
Монокуляр чудесным образом складывается и становится похож на мини-бинокль, что делает его более компактным и удобным для удержания.
Технические характеристики
| Увеличение | 8x |
| Объектив | 22 мм |
| Тип призмы | Porro |
| Выходной зрачок | 2,85 мм |
| Система фокусировки | Sports-Auto-Focus |
| Оптика | High-Contrast (высококонтрастная оптика) |
| Освещенность | 7,6 |
| Сумеречный фактор | 13,3 |
| Поле зрения на 1000 м | 115 м |
| Размеры | 51x49x46 мм |
| Минимальная дистанция фокусировки | 4 м |
| Рабочие температуры | — 10°C to + 55°C |
| Вес | 80 гр |
Из технических характеристик становится ясно, что увеличение этого монокуляра весьма приличное. Морской бинокль из предыдущей статьи имел увеличение 7х, а тут целых 8.
Морской бинокль из предыдущей статьи имел увеличение 7х, а тут целых 8.
Поскольку фокусировка здесь спортивного типа, мы имеем рабочий диапазон от 20 м до бесконечности, что весьма удобно для наблюдения за спортивными соревнованиями, например. Потому система фокусировки и называется Sports-Auto-Focus.
Второй момент касается выходного зрачка.
Здесь он не очень большой, потому монокуляр не предназначен для работы в сумерках, когда человеческий зрачок может достигать 7 мм и изображение будет темнее, чем мы видим глазами.
Впрочем, это больше актуально для молодых, у которых зрачок глаза способен менять размер в широком диапазоне, а для людей старше сорока этот параметр все равно около 4 мм и потому ограничение можно считать не таким уж существенным.
Сфера применения монокуляра STEINER miniscope 8 x 22
Сфер применения монокуляра STEINER Miniscope 8 x 22 огромное множество, потому что в нашей жизни очень много сюжетов, когда хочется рассмотреть что-то удалённое.
Идете на соревнования, на концерт или в театр (если говорить про театр, то увеличение театральных биноклей, как правило, не превышает 4х, а тут целых 8, можно хоть с «галёрки» смотреть)! Едете на экскурсию? Монокуляр удобнейший и полезнейший инструмент.
пример экскурсии
А кто лезет на дом и с какой целью?
А может вы любите наблюдать за птицами или животными в естественной среде обитания?
Или, например, вы, также как и мы, любите ездить на железнодорожную станцию смотреть с детьми поезда.
Не подумайте, что они «кислые» 🙂 Они меня и вытаскивают всё время смотреть на поезда 🙂 Это у нас «выражение лица для фото».
При наличии монокуляра STEINER Miniscope 8 x 22 я беру его с собой на встречи в незнакомых местах, чтобы легче найти того, с кем встречаешься. Даже в метро удобно углядеть своего товарища через всю станцию.
Минископ станет полезным и в оживленных местах больших городов. Недавно он очень выручил меня на остановке в дороге при наблюдениях в зеркало заднего вида.
Недавно он очень выручил меня на остановке в дороге при наблюдениях в зеркало заднего вида.
Наблюдая в зеркало заднего вида, я мог прекрасно различать убирают машины в конце улицы или нет.
Я попробовал смотреть в зеркало заднего вида через бинокль STEINER Miniscope 8 x 22, но сфокусироваться на бесконечность не смог. Секундная заминка, пока до меня дошло, что я фокусироваться должен не на бесконечность, а вовсе даже на макро дистанцию. Но бинокль на то и бинокль, чтобы иметь минимальная дистанцию фокусировки больше, чем 0,5 м, которые до зеркала. Но поскольку я не первый день держу в руках оптику, то в голову пришло решение. Нужно отдалить одну линзу от светоприёмника (глаза). Благо передний линзоблок просто вкручен, я стал выкручивать его и смотреть на изображение. В какой-то момент (почти выкрутил линзоблок) бинокль стал макрообъективом и я смог спокойно наблюдать за дальним концом улицы через зеркало заднего обзора. Вот такой маленькой и полезной штучкой оказался монокуляр STEINER Miniscope 8 x 22!
Также не так давно приезжал мой друг Андрей Орлов из Дагестана со своим коллегой и поступило неожиданное предложение залезть на колоннаду Исаакиевского собора. Наверху я сначала пожалел, что не взял бинокль, а потом вспомнил, что монокуляр-то всегда со мной 🙂 И сам посмотрел, и другим дал посмотреть!
Наверху я сначала пожалел, что не взял бинокль, а потом вспомнил, что монокуляр-то всегда со мной 🙂 И сам посмотрел, и другим дал посмотреть!
Резюме
Монокуляр STEINER Miniscope 8 x 22 хотя и маленький, но удаленький. Оптика хорошая, контрастная (хорошая немецкая оптика с мультипросветлением). Рекомендован всем без исключения!
Он не имеет такого широкого обзора, как большие бинокли типа STEINER Сommander 7×50 , нужно держать правильное расстояние до глаз, не так удобен в сумерках и картинка не бинокулярная, но это с лихвой окупается его малым размером.
Увеличение ведь даже больше!
Как выяснилось, у меня много в жизни случаев, когда требуется монокуляр. Большой бинокль с собой брать неудобно на каждый выход из дома (у меня даже в фоторюкзаке не всегда есть свободное место под большой бинокль), а STEINER Miniscope 8 x 22 помещается в любой карман. Да и цена в разы ниже, что тоже немаловажно.
Автор: Дмитрий Евтифеев
|
⇐ ПредыдущаяСтр 6 из 6 Проверка подлинности таможенных документов предполагает проведение экспресс-исследований на предмет выявления фактов изменений первоначального содержания или вида документов, представленных в таможенные органы. Прежде всего, при раздельном анализе и изучении представленных на перемещаемый через таможенную границу объект документов необходимо обратить внимание на соответствие документа установленной форме — наличие или отсутствие требуемых реквизитов: штампов, печатей, подписей, обозначений, дат и т.д., на соответствие содержания документа существующим нормативным актам и правилам, причём необходимо иметь образцы правильно оформленных документов, соответствующих видов бланков, оттисков подлинных печатей, штампов и подписей должностных лиц. Как правило, эта проверка осуществляется визуально путём сравнения с имеющимися образцами в условиях естественного или искусственного освещения. Следующим этапом является этап исследования внешних признаков документа, который осуществляется уже с применением технических средств таможенного контроля криминалистического направления — этими техническими средствами являются: наблюдательные лупы, микроскопы, ультрафиолетовые осветители, настольные приборы проверки документов. Наблюдательные лупы. Одним из самых распространённых видов оптических приборов, применяемых для увеличения и более детального рассмотрения слабо различимых глазом фрагментов документов или атрибутов, являются наблюдательные лупы. Лупа — это собирательная положительная линза или система линз с небольшим увеличением и фокусным расстоянием порядка 40-70 мм, заключённая в специальную оправу. На вооружении таможен имеется несколько моделей таких приборов. Две лупы западногерманского производства, приобретённые для таможен у фирмы «Helling». «Большая» лупа имеет увеличение наблюдаемого изображения в 3,5 раза, фокусное расстояние — 69 мм, линейное поле зрения — 97 мм, размеры: 230х34х67 мм, массу — 240 г. Лупа снабжена устройством подсветки исследуемой зоны наблюдения — миниатюрной лампочкой. «Малая» лупа — по конструкции аналогична «большой» и обеспечивает увеличение в 6,5 раз, имеет фокусное расстояние — 38 мм, линейное поле зрения — 50 мм. Из отечественных оптических средств в практике таможенных органов может применяться наблюдательная лупа типа ЛПП-1, которая, также как и ранее названные модели, имеет возможность местной подсветки, увеличение — 3,5 раза, фокусное расстояние — 71 мм, линейное поле зрения — 65 мм. Микроскопы. При исследовании документов и атрибутов таможенного обеспечения к микроскопам прибегают в тех случаях, когда увеличение, создаваемое лупой, — недостаточно. Микроскоп — это комбинация двух оптических систем — объектива и окуляра. В практике работы таможенных органов применяются в основном две модели миниатюрных микроскопов. Это — «Минископ», модель 1171, имеет 30-ти кратное увеличение, линейное поле зрения — 5 мм, размеры — длина 125 мм, диаметр 15 мм, встроенной подсветки — не имеет. В случае применения «Минископа» для контроля целостности пломб исследование вынуждено проводиться на весу, т.к. минимальная высота объекта, который может быть рассмотрен при установке пломбы и прибора на одну плоскость, составляет всего 5 мм при выдвинутом окуляре до 9 мм, что требует достаточно хорошей практической подготовки оперативных работников при работе с этого вида техникой. По отзывам сотрудников таможенных служб, «Минископ» не находит широкого применения из-за указанного недостатка, а также из-за отсутствия возможности подсветки исследуемого объекта; неудобно для восприятия и перевёрнутое изображение. Более совершенной и удобной в эксплуатации моделью миниатюрного микроскопа является модель «FF-393», японского производства. Микроскоп «FF-393» имеет увеличение 30 крат, габариты 140х48х22 мм, вес всего 30 г. Питание встроенной лампочки подсветки обеспечивает непрерывную работу в течение 3-х часов. Особенность конструкции данной модели заключается в том, что исследование участка документа можно осуществлять как при естественном освещении, так и без него, используя возможность включения лампочки местной подсветки. Для фокусировки наблюдаемого изображения используется фокусирующее кольцо. Ультрафиолетовые осветители. Исследование документов таможенного обеспечения в невидимых, не воспринимаемых человеческим глазом лучах, к которым относятся и ультрафиолетовые лучи, является достаточно эффективным средством проверки их подлинности и целостности, т.к. оптические свойства веществ в этих лучах отличаются от их свойств в видимом свете. УФ-лучи в спектре электромагнитных волн занимают интервал длин волн от 10 миллимикрон до 400 нм. Настольные приборы проверки документов. В стационарных условиях, когда оперативный работник таможни имеет возможность работать с представленными служебными документами непосредственно на своём рабочем месте, неудобно использовать все виды технических средств для проверки документов — лупы, микроскопы, электро- и ультрафиолетовые фонари — по отдельности. Технические средства инспекции объектов таможенного контроля
Системы для досмотра грузовых автомобилей: Rapiscan 2xxx — системы, предназначенные для быстрого и эффективного инспектирования грузовых контейнеров и автомобилей. Они способны инспектировать авиационные грузовые контейнеры, большегрузные автомобили с максимальными габаритами 4,5 м по высоте, 3,5 м шириной, 25 м длиной и массой до 60 т. Системы Rapiscan 2 ххх соответствуют стандартам и требованиям IAEA и WHO. После досмотра грузов отсутствует остаточная радиация, вредные токсикологические, пищевые или микробиологические последствия при инспектировании продуктов питания. Rapiscan 2100. Система низкой пропускной способности, в которой грузовой автомобиль паркуется внутри камеры досмотра и рентгеновская система перемещается вдоль автомобиля, генерируя изображение. Максимальная пропускная способность — около 10 грузовых автомобилей в час. Главное преимущество системы — небольшая площадь земли, требуемая для установки — 25х30 м. Rapiscan 2200. Система стандартной пропускной способности со стационарной рентгеновской установкой и перемещением автомобиля конвейером низкой стоимости. Максимальная пропускная способность — около 10 грузовых автомобилей в час. Rapiscan 2300. Система высокой пропускной способности со стационарной рентгеновской установкой и перемещением автомобиля конвейером высокого качества. Максимальная пропускная способность — около 35 грузовых автомобилей в час, что делает ее более эффективной в ситуациях, требующих высокого уровня досмотра. Rapiscan 2400. Система для досмотра авиационных контейнеров. Пропускная способность может достигать 60 и более инспекций в час Мобильная система досмотра Rapiscan 3000. Пропускная способность для грузовика, контейнера, вагона или машины 1-3 минуты, она зависит от размера досматриваемого транспортного средства и площади установки. Пропускная способность до 5 миль в час достигается при должных условиях зоны исключения и участка контроля для досмотра транспортного средства. Технические средства наблюдения за оперативной обстановкой в зонах таможенного контроля Первая — это телевизионный контроль внутренних таможенных зон. Применение здесь ТСТК для телевизионного наблюдения сведено к необходимости ведения контроля за оперативной обстановкой во внутренних таможенных помещениях — пассажирские залы, комплектовки, грузовые пакгаузы и т.п., имеющего целью получения информации о поведении конкретных лиц, находящихся в этих помещениях, их действиях, контактах, попытках к сокрытию или передаче другим лицам каких-либо предметов, поведение отдельных пассажиров, прибывающих из «взрывоопасных» или «наркотикоопасных» регионов. Вторая составляющая задачи — визуальный и телевизионный контроль таможенных территорий — предполагает осуществление наблюдения с помощью ТСТК за опасными с точки зрения контрабандной деятельности или криминогенной обстановки зонами, участками, объектами. Речь идет, например, о наблюдении за действиями отдельных членов экипажа судна, на подходе к порту или на рейде с целью выявления фактов сброса предметов контрабанды в воду, или возможного сноса с борта судна, уже прошедшего таможенный досмотр, не декларированных, контрабандных товаров, выявление потенциальных контактов их с лицами, занимающимися контрабандной деятельностью. Техника должна обеспечивать возможность независимого контроля одновременно всех оперативно необходимых участков таможенной зоны, обеспечивать возможность просмотра зон путем поворота ТВ — камер в горизонтальной, и вертикальной плоскостях, возможности трансфоцирования изображения, видеозапись изображения с любого видеоконтрольного устройства — телемонитора на видеомагнитофон. Технические средства контроля носителей аудиоинформации и видеоинформации Аудиоинформация, которая запрещена к перемещению через границу, может сохраняться на звуковых бобинах, катушках, компакт-кассетах стандартного бытового образца или на специальных микрокассетах от миниатюрных диктофонов. Для прослушивания информации, записанной на этих звуковых магнитных носителях, используются обычные бытовые магнитофоны, компакт-кассетные плееры и магнитофоны, а также минидиктофоны под различные «фирменные» микрокассеты. Микрокассетный диктофон типа RN-502EZ-K фирмы «Панасоник» предназначен для записи и воспроизведения микрокассет. Диктофон RN-502EZ-K оснащен системой активации при звуке т.е. звук записывается автоматически, что позволяет избежать лишних затрат пленки. Также имеет функцию двух скоростей записи и воспроизведения. Микрокассетными диктофонами фирмы «Панасоник» оснащались и оснащаются таможенные службы. Контроль видеоинформации, записанной на видеокассетах, требует оснащения участков таможенного контроля видеомагнитофонами или плеерами всех существующих в настоящее время видеосистем и форматов видеозаписи, которые, естественно, должны приобретаться за рубежом, т. В качестве телевизионных видеоконтрольных устройств используются мультисистемные мониторы в основном японского производства, также приобретаемые за рубежом. В отдельных случаях для просмотра видеокассет могут использоваться и отечественные телевизоры, имеющие специальные декодеры. В Rolsen R2V 400 предусмотрена как поддержка основных цифровых форматов — проигрыванием всех основных форматов, — DVD, Video CD, Super Video CD, Audio CD, MP3, так и поддержка воспроизведения видеофоматов PALMesecam. Мощный аудио-ЦАП обеспечивает высококлассное Hi-Fi-стереозвучание, а 6 видеоголовок с лазерной юстировкой обеспечат и высокое качество просмотра обычного видео. Проигрыватель Dream X-108 — это практически универсальный DVD проигрыватель компакт-дисков. Не подлежащая перемещению через госграницу визуальная информация может вместе с иной информацией храниться на экспонированных фото и кинопленках, слайдах, микрофильмах, микрофишах. Просмотр микрофильмов и микрофишей может проводиться на специальных компактных просмотровых устройствах, которые выпускаются серийно пока только за рубежом. Контроль носителей ЭВМ т.е. предметов звукозаписи, видеозаписи и носителей информации, относящиеся к ЭВМ, осуществляется после соответствующего таможенного контроля с применением технических средств таможенного контроля. Заполненные носители информации в силу своей специфики пропускаются при наличии сведений о типе ЭВМ и операционной системе, с использованием которых произведена запись. При его отсутствии и при невозможности расшифровки носителя информации ЭВМ инспектор таможни вправе с разрешения начальника таможни вернуть заполненные носители информации владельцу и отказать в перемещении через государственную границу. Информация на магнитных носителях в основном храниться в виде файлов. Файл — это совокупность записей, единиц информации, отождествляющих собой единое целое. В файлах могут храниться текстовые, графические документы, выполняемые программы и т.д. Имена файлов состоят непосредственно из имени и его расширения, которое определяет его принадлежность к тому или иному типу или программе. Расширение файлов вовсе необязательны и поэтому они могут и отсутствовать, но для правильной и корректной работы их употребляют, т.к. они отождествляют определенную принадлежность данного файла к определенной программе в которой, этот файл был создан.
⇐ Предыдущая123456 Читайте также: |
 Из сложившейся практики работы правоохранительных органов, имеющих значительный опыт в области исследований различных видов документов, проверка подлинности таможенных документов строится по следующей схеме.
Из сложившейся практики работы правоохранительных органов, имеющих значительный опыт в области исследований различных видов документов, проверка подлинности таможенных документов строится по следующей схеме.

 Благодаря способности вызывать люминесценцию, использование УФ-лучей позволяет различать материалы, имеющие одинаковый внешний вид при наблюдении в видимом свете. Из-за различного химического состава по-разному люминесцируют некоторые сходные, близкие по цвету красители, а также места вытравливании записей. Ультрафиолетовый осветитель представляет собой наполненный инертным газом кварцевый сосуд, на внутреннюю поверхность которого нанесено небольшое количество распылённой ртути и в который вмонтированы электроды. При подаче напряжения на электроды образуются пары ртути, через которые проходят электроны, вызывая световой поток, богатый УФ-лучами. Поскольку обычное стекло в значительной степени поглощает УФ-лучи, то фильтр, поглощающий видимый свет, делают, как правило, из кварца или специального вида стекла. В качестве ультрафиолетовых осветителей таможенные органы в настоящее время используют три модификации фонарей. Одна из этих моделей — УФ-фонарь «Паломар», которой оснащались таможни в период 1981-82 гг.
Благодаря способности вызывать люминесценцию, использование УФ-лучей позволяет различать материалы, имеющие одинаковый внешний вид при наблюдении в видимом свете. Из-за различного химического состава по-разному люминесцируют некоторые сходные, близкие по цвету красители, а также места вытравливании записей. Ультрафиолетовый осветитель представляет собой наполненный инертным газом кварцевый сосуд, на внутреннюю поверхность которого нанесено небольшое количество распылённой ртути и в который вмонтированы электроды. При подаче напряжения на электроды образуются пары ртути, через которые проходят электроны, вызывая световой поток, богатый УФ-лучами. Поскольку обычное стекло в значительной степени поглощает УФ-лучи, то фильтр, поглощающий видимый свет, делают, как правило, из кварца или специального вида стекла. В качестве ультрафиолетовых осветителей таможенные органы в настоящее время используют три модификации фонарей. Одна из этих моделей — УФ-фонарь «Паломар», которой оснащались таможни в период 1981-82 гг. Это достаточно габаритный фонарь размерами: 380х60х90 мм и весом 710 г. Расстояние, с которого обеспечивается нормальная освещённость объекта, составляет 16 см. На защитных баллонах ламп и крышках батарейного отсека установлены резиновые уплотнители, что позволяет использовать фонарь во влажных и взрывоопасных средах. На корпусе фонаря установлен достаточно сильный магнит, с помощью которого фонарь можно крепить к металлическим конструкциям, чтобы освободить руки оперативного работника. Из-за достаточно больших габаритов и веса, фонарь практически очень редко использовался в оперативных условиях. Второй моделью УФ-фонаря, применяемой в оперативной работе таможенных органов, является модель «81600», фирма-поставщик «Helling». Этот УФ-фонарь, — карманного типа, работает в длинноволновом диапазоне ультрафиолетового излучения. Малогабаритный, лёгкий, с достаточной мощностью УФ-излучения УФ-фонарь модели «81600» отлично зарекомендовал себя в оперативной практике проверки подлинности таможенных документов и целостности атрибутов таможенного обеспечения.
Это достаточно габаритный фонарь размерами: 380х60х90 мм и весом 710 г. Расстояние, с которого обеспечивается нормальная освещённость объекта, составляет 16 см. На защитных баллонах ламп и крышках батарейного отсека установлены резиновые уплотнители, что позволяет использовать фонарь во влажных и взрывоопасных средах. На корпусе фонаря установлен достаточно сильный магнит, с помощью которого фонарь можно крепить к металлическим конструкциям, чтобы освободить руки оперативного работника. Из-за достаточно больших габаритов и веса, фонарь практически очень редко использовался в оперативных условиях. Второй моделью УФ-фонаря, применяемой в оперативной работе таможенных органов, является модель «81600», фирма-поставщик «Helling». Этот УФ-фонарь, — карманного типа, работает в длинноволновом диапазоне ультрафиолетового излучения. Малогабаритный, лёгкий, с достаточной мощностью УФ-излучения УФ-фонарь модели «81600» отлично зарекомендовал себя в оперативной практике проверки подлинности таможенных документов и целостности атрибутов таможенного обеспечения. Вместе с тем, опыт оперативной работы показывает, что и модель «81600» не во всех случаях удовлетворяет условиям, в которых работают сотрудники таможен. Зачастую при проверке таможенных документов и атрибутов таможенного обеспечения в сочетании с ультрафиолетовым излучением требуется проведение осмотра в видимых лучах. Это заставляет использовать отдельно ещё и карманные электрофонари, что вызывает определенные затруднения и неудобства при проведении контроля. Эти недостатки полностью исключены в новой модели комбинированного УФ-фонаря типа «BHW», которым будут снабжаться таможенные органы. Фонарь выполнен в виде комбинации 3-х осветителей: собственно ультрафиолетового, имеющего полностью аналогичные технические параметры УФ-фонарей модели «81600»; фонаря видимого освещения, выполненного в виде поворачивающейся головки с лампочкой и отражателем и мигающего фонаря — «маячка».
Вместе с тем, опыт оперативной работы показывает, что и модель «81600» не во всех случаях удовлетворяет условиям, в которых работают сотрудники таможен. Зачастую при проверке таможенных документов и атрибутов таможенного обеспечения в сочетании с ультрафиолетовым излучением требуется проведение осмотра в видимых лучах. Это заставляет использовать отдельно ещё и карманные электрофонари, что вызывает определенные затруднения и неудобства при проведении контроля. Эти недостатки полностью исключены в новой модели комбинированного УФ-фонаря типа «BHW», которым будут снабжаться таможенные органы. Фонарь выполнен в виде комбинации 3-х осветителей: собственно ультрафиолетового, имеющего полностью аналогичные технические параметры УФ-фонарей модели «81600»; фонаря видимого освещения, выполненного в виде поворачивающейся головки с лампочкой и отражателем и мигающего фонаря — «маячка». Исключить это неудобство позволяет специально разработанный отечественной промышленностью по заданию таможенного комитета комбинированный настольный прибор для проверки подлинности «таможенных» документов. Приборы «Версия — М» уже начинают поступать в таможенные органы, что позволит в значительной степени повысить эффективность работы по проверке подлинности таможенных документов. Они станут неотъемлемой частью рабочего места оперативного работника таможни, а благодаря своим техническим возможностям с успехом будут эксплуатироваться работниками подразделений дознания.
Исключить это неудобство позволяет специально разработанный отечественной промышленностью по заданию таможенного комитета комбинированный настольный прибор для проверки подлинности «таможенных» документов. Приборы «Версия — М» уже начинают поступать в таможенные органы, что позволит в значительной степени повысить эффективность работы по проверке подлинности таможенных документов. Они станут неотъемлемой частью рабочего места оперативного работника таможни, а благодаря своим техническим возможностям с успехом будут эксплуатироваться работниками подразделений дознания. Такие системы используют унифицированные составляющие подсистемы для комбинирования их в различных конфигурациях для строительства оптимальных для любого применения систем. Для получения изображения досматриваемого объекта используются высокоэнергетические рентгеновские лучи. Изображение сохраняется в компьютере системы вместе с данными товарно-транспортной накладной и другими данными, подлежащими детальному изучению или сравнению.
Такие системы используют унифицированные составляющие подсистемы для комбинирования их в различных конфигурациях для строительства оптимальных для любого применения систем. Для получения изображения досматриваемого объекта используются высокоэнергетические рентгеновские лучи. Изображение сохраняется в компьютере системы вместе с данными товарно-транспортной накладной и другими данными, подлежащими детальному изучению или сравнению.
 Система имеет проем, размеры которого 3,5 м Х 4,5 м для сканирования больших транспортных средств, размеры которых превышают стандартные, обычно встречающихся при работе в портах. На приемной площадке сканируется декларация карго-груза, информация поступает в компьютер контроля системы вместе с другими необходимыми данными, а во время процесса сканирования оператор может сравнить содержимое с заявленным.
Система имеет проем, размеры которого 3,5 м Х 4,5 м для сканирования больших транспортных средств, размеры которых превышают стандартные, обычно встречающихся при работе в портах. На приемной площадке сканируется декларация карго-груза, информация поступает в компьютер контроля системы вместе с другими необходимыми данными, а во время процесса сканирования оператор может сравнить содержимое с заявленным. Кроме этого, что в последнее время стало весьма актуальным, необходимо с помощью техники получать информацию о поведении отдельных пассажиров, грузовых агентов, служащих транспортных зон непосредственно в процессе таможенного контроля, из взаимоотношений с оперативным составом таможен, осуществляющим этот контроль, с целью предотвращения противоправных действий и исключения влияния этих людей на безупречное выполнение таможенниками своих обязанностей.
Кроме этого, что в последнее время стало весьма актуальным, необходимо с помощью техники получать информацию о поведении отдельных пассажиров, грузовых агентов, служащих транспортных зон непосредственно в процессе таможенного контроля, из взаимоотношений с оперативным составом таможен, осуществляющим этот контроль, с целью предотвращения противоправных действий и исключения влияния этих людей на безупречное выполнение таможенниками своих обязанностей. Кроме этого, ТСТК, обеспечивающие телевизионное наблюдение, предназначаются для контроля периметров таможенных территорий, отдельных грузовых досмотровых площадок, зон, где хранятся грузы, контейнеры, транспортные средства, требующие обеспечения их безопасности.
Кроме этого, ТСТК, обеспечивающие телевизионное наблюдение, предназначаются для контроля периметров таможенных территорий, отдельных грузовых досмотровых площадок, зон, где хранятся грузы, контейнеры, транспортные средства, требующие обеспечения их безопасности. Но звуковая информация может быть записана и на обычных грампластинках и на сверхсовременных аудионосителях — лазерных дисках. Для их прослушивания в практике таможен применяются обычные проигрыватели грампластинок отечественного производства и закупленные за рубежом плееры лазерных аудиодисков.
Но звуковая информация может быть записана и на обычных грампластинках и на сверхсовременных аудионосителях — лазерных дисках. Для их прослушивания в практике таможен применяются обычные проигрыватели грампластинок отечественного производства и закупленные за рубежом плееры лазерных аудиодисков. к. отечественных аналогов нет и в обозримом будущем не предвидится.
к. отечественных аналогов нет и в обозримом будущем не предвидится. Для ее просмотра применяются устройство просмотра фотопленок «ДЭФИ», диапроекторы «Пеленг», диапроектор с просветным складным экраном «Экран-универсал», просмотрово-монтажный столик узкоформатных кинофильмов.
Для ее просмотра применяются устройство просмотра фотопленок «ДЭФИ», диапроекторы «Пеленг», диапроектор с просветным складным экраном «Экран-универсал», просмотрово-монтажный столик узкоформатных кинофильмов. Носителями информации в основном являются персональные, портативные компьютеры их устройства такие, как модули памяти SIMM, DIMM, RIMM, SOJ с электронной поддержкой питания, жесткие диски, а также магнитные и электронные накопители т.е. флоппи-диски, CD, DVD-диски, магнитные и ленточные накопители, USB накопители и т.д.
Носителями информации в основном являются персональные, портативные компьютеры их устройства такие, как модули памяти SIMM, DIMM, RIMM, SOJ с электронной поддержкой питания, жесткие диски, а также магнитные и электронные накопители т.е. флоппи-диски, CD, DVD-диски, магнитные и ленточные накопители, USB накопители и т.д.Приборы ночного видения
|
||||||||||||
|
Главная Приборы ночного видения Вернуться в раздел технические средства |
|||||||||||
|
www. |
||||||||||||
 12.2017
12.2017 0″
0″
Copyright 2010 www.sis-tss.ru
|
Заглавная страница
КАТЕГОРИИ: Археология ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрации Техника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ? Влияние общества на человека Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. |
⇐ ПредыдущаяСтр 15 из 20Следующая ⇒
Раньше квалифицированный настройщик электронного оборудования был вынужден иметь целый арсенал измерительных приборов — тестер или мультиметр, электронный осциллограф, частотомер, измеритель емкости и индуктивности, анализатор спектров и т. д. Новую концепцию инструментальных средств и приборов для отладки электронных устройств предложила фирма Fluke (США). Помимо выпуска стандартных приборов (например, мультиметров разного класса) фирма создала новый тип приборов -.скопметры. Из самого названия этих приборов ясно, что они предназначены для измерения скопом всего, что только можно измерить: напряжения, тока, частоты, длительности импульсов и их времени нарастания, наблюдения и запоминания формы электрических и радиотехнических сигналов и т. д. Для промышленности выпускаются скопметры Fluke 105B/99B/96B/92B. Достаточно широкую номенклатуру портативных осциллографов выпускает объединение АКТАКОМ. Под его маркой выпускаются осциллографы ряда зарубежных фирм, имеющие уровень параметров соответствующий мировому уровню работки и производства осциллографов. АСК-4201 — портативный цифровой осциллограф в виде мультиметра с ручкой (рис. 8.3). Рисунок 8.3 – Мультископ АСК-4201 Основные характеристики: 2 канала с гальванической развязкой, полоса пропускания 20МГц, ЖКИ 58×38 мм с разрешением 128×64 точки, чувствительность 0,1В/дел …5 В/дел, коэффициенты развертки 0,5 мкс/дел…0,2 с/дел, максимальное входное напряжение 40 В, время установления 17,5 нс, входное сопротивление МОм, входная емкость 60 пФ. Режимы разверток: автоколебательный, ждущий, однократный, Х-У, предварительная запись осциллограмм. Прибор позволяет выполнять измерения в полевых и цеховых условиях. Малые габариты и вес позволяют носить его в кейсе или дипломате. Открытая модульная конструкция позволяет наращивать возможности прибора. Цифровой портативный осциллограф-щуп (минископ) АСК-40П один из самых миниатюрных осциллографов ((рис.8.4). Рисунок 8.4 – Минископ АСК-4011 Он имеет следующие характеристики: 2 канала, лоса пропускания 5 МГц, ЖКИ 35×12 (2м), коэффициенты развертки 0,05 мкс/дел…2 мс/дел, максимальное входное напряжение 80 В. Режимы разверток: автоколебательный, однократный. Источники синхронизации: внутренний, внешний. Дополнительные функции; вольтметр, интерфейс RS-232, программное обеспечение. Питание через сетевой адаптер. Габариты 165x33x20, масса 86 г.
Тема 9. Цифровые приборы измерения частоты Общие сведения
Во многих областях науки и техники измерение частоты электромагнитных колебаний является одним из самых распространенных видов измерений. Частотой колебаний f называют число полных колебаний в единицу времени f = n/t, где t — время существования n колебаний. Для периодических сигналов частота f = 1/Т, где Т — период колебаний, который определяется как наименьший интервал времени, через который повторяются мгновенные значения периодического сигнала. Таким образом, частота — это физическая величина, численно равная числу идентичных событий в единицу времени. Частота электромагнитных колебаний однозначно связана не только с периодом колебания Т, но и с длиной однородной плоской волны в свободном пространстве λ: f = c/ λ, где с ≈ 3 ∙ 108 м/с — скорость света в свободном пространстве; λ — длина волны, м. Теоретически измерения частоты, времени и длины волны равноценны, но практически в большинстве случаев измеряют частоты, реже — интервалы времени. Длина волны при необходимости легко вычисляется, а на сверхвысоких частотах и измеряется. Диапазон частот электрорадиотехнических сигналов простирается от долей герца до тысяч гигагерц. Например, спектр частот, применяемых для радиосвязи, радиовещания и телевидения упорядочен. В соответствии с ГОСТ 24375-80 такое разделение частот по полосам и их метрические наименования приведены в таблице 9.1. Таблица 9.1 – Стандартные полосы частот
Измерение частоты в основном осуществляется двумя основными методами: — преобразованием частоты в постоянное напряжение, измеряемое стрелочным или цифровым вольтметром; — дискретного счета – подсчетом цифровым счетчиком числа периодов сигнала за эталонный промежуток времени. Первый способ широко применялся в аналоговых приборах. При высоких требованиях к точности всегда используется второй метод. Метод дискретного счета лежит в основе построения электронно-счетных частотомеров (ЭСЧ), используемых для измерения частотно-временных параметров электрических сигналов.
Цифровой частотомер
Упрощенная структурная схема ЭСЧ в режиме измерения частоты представлена на (рис.9.1). Схема содержит следующие элементы: входное устройство (ВУ), формирующее устройство (ФУ), временной селектор (ВС), опорный генератор (ОГ), делитель частоты (ДЧ), устройство формирования и управления (УФУ), электронный счетчик (ЭС), дешифратор (Дш) и цифровой индикатор (ЦИ). Рисунок 9.1 – Структурная схема ЭСЧ в режиме измерения частоты Входное устройство обеспечивает усиление или, наоборот, ослабление входного сигнала и его фильтрацию. Формирующее устройство преобразует исследуемый сигнал в последовательность импульсов, частота следования которых равна частоте исследуемого сигнала. Временной селектор представляет собой логическую схему, которая обеспечивает пропускание на электронный счетчик сформированные импульсы измеряемой частоты только при наличии на управляющем входе стробирующего импульса, длительность которого определяет время измерения. Опорный генератор является источником сигнала калиброванного временного интервала. В этих целях, как правило, используется высокостабильный термостатированный кварцевый генератор частотой 1 или 5 МГц. С помощью формирующего устройства опорного генератора вырабатывается последовательность импульсов, поступающих на делитель частоты. Делитель частоты формирует последовательность импульсов, частота следования которых в 10n (n = 1, 2, 3…) раз ниже частоты сигнала опорного генератора. Устройство формирования и управления на основе сигнала, поступающего с делителя частоты, обеспечивает получение прямоугольного строб-импульса, длительность которого определяет время счета и соответственно время измерения. Электронный счетчик обеспечивает подсчет и запоминание числа импульсов, прошедших через селектор за время строб-импульса. Информация с электронного счетчика через дешифратор поступает на цифровой индикатор, на табло которого появляется информация в единицах частоты. Перед началом нового цикла измерений необходимо подготовить счетчик, сбросив показания прошлого цикла. Это делается через цепь сброса от устройства управления. Таким образом, как следует из описания структурной схемы прибора, при измерении частоты на первый вход ВС поступает последовательность импульсов с периодом Тх, определяемым частотой исследуемого сигнала fх, причем Тх = 1/fx. На второй вход ВС поступает строб-импульс длительностью ∆Т=10nТкв, (9.1) где Ткв — период следования импульсов с опорного кварцевого генератора. На электронный счетчик проходит группа импульсов, число которых (9.2) Если не учитывать погрешность дискретизации и сравнивать значения для ∆Т из равенств (9. откуда (9.3) Таким образом, измеряемая частота равна числу импульсов N, образованных из измеряемого сигнала, а fкв ∙ 10—n — коэффициент, определяющий единицу измерения частоты и число значащих цифр при отсчете (рис.9.2). Рисунок 9.2 – Временная диаграмма работы ЭСЧ в режиме измерения частоты Принято выделять две основные составляющие погрешности измерения частоты δf. Первая составляющая — это погрешность формирования образцового интервала времени ∆Т, в течение которого временной селектор пропускает импульсы, т.е. проводится измерение. Эта погрешность определяется погрешностью меры, т.е. нестабильностью частоты кварцевого генератора δкв. Вторая составляющая – погрешность дискретизации δД, возникает за счет потери части периода измеряемых импульсов ∆t1, ∆t2 и приблизительно составляет ∆N = ±1 импульс. Относительная погрешность дискретизации (9.
Тогда, относительная погрешность измерения будет равна (9.5) ⇐ Предыдущая10111213141516171819Следующая ⇒ Читайте также: Организация работы процедурного кабинета Статус республик в составе РФ Понятие финансов, их функции и особенности Сущность демографической политии |
||||||
|
Последнее изменение этой страницы: 2016-04-07; просмотров: 708; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. |

 Определение реакций опор и моментов защемления
Определение реакций опор и моментов защемления

 Прибор можно использовать как цифровой частотомер и мультиметр.
Прибор можно использовать как цифровой частотомер и мультиметр.
 Этот спектр разбит на девять полос, внутри которых частоты изменяются в 10 раз.
Этот спектр разбит на девять полос, внутри которых частоты изменяются в 10 раз.


 1) и (9.2), то имеем
1) и (9.2), то имеем 4)
4) Обратная связь — 176.9.44.166 (0.008 с.)
Обратная связь — 176.9.44.166 (0.008 с.) Осцилограф в Киеве (Осциллографы) — Авител — электронные компоненты на Bizorg.su
Цена: Цену уточняйте
за 1 ед.
Компания Авител — электронные компоненты (Киев) является зарегистрированным поставщиком на сайте BizOrg.su. Вы можете приобрести товар Осцилограф с доставкой по одному региону Украины, расчеты производятся в ₽. Если у вас возникли проблемы при заказе товара, пожалуйста, сообщите об этом нам через форму обратной связи.
Описание товара
Все лабораторное и измерительное оборудование
Измерительные приборы, большой выбор, со склада в Киеве и под заказ, скидки.
Минископ, осциллограф – мультиметр, портативный осциллограф HPS-10
Минископ, осциллограф – мультиметр, портативный осциллограф HPS-40
Минископ, осциллограф – мультиметр, портативный осциллограф FLUKE-123
Минископ, осциллограф – мультиметр, портативный осциллограф Осциллограф С1-137/1
Минископ, осциллограф – мультиметр, портативный осциллограф FLUKE-124
Минископ, осциллограф – мультиметр, портативный осциллограф ОМЦ-200
Минископ, осциллограф – мультиметр, портативный осциллограф U1562A
Минископ, осциллограф – мультиметр, портативный осциллограф U1590A
Минископ, осциллограф – мультиметр, портативный осциллограф U1602A
Минископ, осциллограф – мультиметр, портативный осциллограф FLUKE-225C
Минископ, осциллограф – мультиметр, портативный осциллограф FLUKE-215C
Минископ, осциллограф – мультиметр, портативный осциллограф АСК-2068
Минископ, осциллограф – мультиметр, портативный осциллограф 2000/E
Минископ, осциллограф – мультиметр, портативный осциллограф АКИП-4113/1 с LAN
Минископ, осциллограф – мультиметр, портативный осциллограф Fluke-215C/S
Минископ, осциллограф – мультиметр, портативный осциллограф АКИП-4113/6
Осциллограф аналогово-цифровой АСК-22020
Осциллограф аналогово-цифровой АСК-22060
Осциллограф аналогово-цифровой АСК-2021
Осциллограф аналогово-цифровой АСК-2031
Осциллограф аналогово-цифровой GOS-6051
Осциллограф аналогово-цифровой GOS-620
Осциллограф аналогово-цифровой GOS-620FG
Осциллограф аналогово-цифровой GOS-6050
Осциллограф аналогово-цифровой GOS-6030
Осциллограф аналогово-цифровой GRS-6052A
Осциллограф аналогово-цифровой GOS-6031
Осциллограф аналогово-цифровой GOS-6112
Осциллограф аналогово-цифровой GOS-653G
Осциллограф аналогово-цифровой GOS-622G
Осциллограф аналогово-цифровой ОСУ-10В
Осциллограф аналогово-цифровой ОСУ-10А
Осциллограф аналогово-цифровой ОСУ-20
Осциллограф аналогово-цифровой GOS-6200
Осциллограф аналогово-цифровой GOS-652G
Осциллограф аналогово-цифровой С1-120
Осциллограф аналогово-цифровой MSO 6032A
Осциллограф аналогово-цифровой Осциллограф С1-137/2
Осциллограф аналогово-цифровой MSO4034
Осциллограф аналогово-цифровой АКИП-4104
Осциллограф аналогово-цифровой MSO4034B
Осциллограф аналоговый АСК-1102
Осциллограф аналоговый АСК-24020
Осциллограф аналоговый АСК-1021
Осциллограф аналоговый АСК-21060
Осциллограф аналоговый АСК-1051
Осциллограф аналоговый АСК-1052
Осциллограф аналоговый GOS-6103C
Осциллограф аналоговый GOS-6103
Осциллограф аналоговый АСК-1053
Осциллограф аналоговый АСК-7022
Осциллограф аналоговый АСК-7203
Осциллограф аналоговый АСК-7304
Осциллограф аналоговый АСК-7404
Осциллограф аналоговый АСК-7474
Осциллограф аналоговый АСК-8064
Осциллограф аналоговый АСК-8104
Осциллограф аналоговый С1-220
Осциллограф аналоговый АСК-2150-РО
Осциллограф аналоговый GOS-7630FC
Осциллограф аналоговый MOS-620
Осциллограф цифровой GRS-6032A
Осциллограф цифровой TDS-3034B+TDS3GV+WSTRO
Осциллограф цифровой WS 452
Осциллограф цифровой TDS-5034B
Осциллограф цифровой GDS-806S
Осциллограф цифровой FLUKE-196B
Осциллограф цифровой FLUKE-196C
Осциллограф цифровой FLUKE-199B/S
Осциллограф цифровой FLUKE-199C
Осциллограф цифровой TDS3014B
Осциллограф цифровой TDS-3052C
Осциллограф цифровой WJ 314A
Осциллограф цифровой DSO 6012A
Осциллограф цифровой DSO 6014A
Осциллограф цифровой DSO 6032A
Осциллограф цифровой DSO 6052A
Осциллограф цифровой DSO 6054A
Осциллограф цифровой DSO 6102A
Осциллограф цифровой MSO 6014A
Осциллограф цифровой MSO 6052A
Осциллограф цифровой GDS-2104
Осциллограф цифровой GDS-2202
Осциллограф цифровой GDS-2064
Осциллограф цифровой GDS-2204
Осциллограф цифровой GDS-2062
Осциллограф цифровой TDS-1002B
Осциллограф цифровой WJ 332A
Осциллограф цифровой WJ 322A
Осциллограф цифровой GDS-2102
Осциллограф цифровой WJ 334A
Осциллограф цифровой WJ 352
Осциллограф цифровой WJ 354A
Осциллограф цифровой WS 64XS
Осциллограф цифровой WR 104Xi-A
Осциллограф цифровой WS 104XS
Осциллограф цифровой TDS-1012B
Осциллограф цифровой TDS-2012C
Осциллограф цифровой АСК-2150 с калибровкой
Осциллограф цифровой АСК-4166-РО5
Осциллограф цифровой TDS-1001B
Осциллограф цифровой PCSU1000
Осциллограф цифровой 1690A
Осциллограф цифровой 34901A
Осциллограф цифровой 34903A
Осциллограф цифровой 34970A
Осциллограф цифровой DSO 5014A
Осциллограф цифровой DSO 5032A
Осциллограф цифровой N2861A
Осциллограф цифровой TDS-3012B
Осциллограф цифровой WJ 312
Осциллограф цифровой DPO4032
Осциллограф цифровой АСК-2034
Осциллограф цифровой АСК-2028
Осциллограф цифровой АСК-2067
Осциллограф цифровой GDS-71022
Осциллограф цифровой WS 44XS
Осциллограф цифровой DPO4104
Осциллограф цифровой GDS-71102
Осциллограф цифровой GDS-71062
Осциллограф цифровой GDS-71042
Осциллограф цифровой WP 7100A
Осциллограф цифровой WP 7200A
Осциллограф цифровой WP 7300A
Осциллограф цифровой WM 8600A XXL
Осциллограф цифровой WM 8620A
Осциллограф цифровой DS1062CA
Осциллограф цифровой DS1102CA
Осциллограф цифровой DS1202CA
Осциллограф цифровой DS1302CA
Осциллограф цифровой DS5202M
Осциллограф цифровой DPO3014
Осциллограф цифровой TDS-3012C
Осциллограф цифровой WS 62XS
Осциллограф цифровой DPO3032
Осциллограф цифровой DPO3034
Осциллограф цифровой DPO3052
Осциллограф цифровой DPO3054
Осциллограф цифровой TDS-3054C
Осциллограф цифровой TDS-3034C
Осциллограф цифровой WA 222
Осциллограф цифровой TDS-3014C
Осциллограф цифровой TDS-3032C
Осциллограф цифровой UT81B
Осциллограф цифровой DPO3012
Осциллограф цифровой АСК-2042
Осциллограф цифровой АСК-2043
Осциллограф цифровой АСК-2062
Осциллограф цифровой АСК-2063
Осциллограф цифровой АСК-2065
Осциллограф цифровой АСК-2103
Осциллограф цифровой АСК-2104
Осциллограф цифровой АСК-2105
Осциллограф цифровой АСК-2203
Осциллограф цифровой АСК-2205
Осциллограф цифровой АСК-6022
Осциллограф цифровой WA 102
Осциллограф цифровой WA 112
Осциллограф цифровой WS 104XS-A
Осциллограф цифровой GDS-1102
Осциллограф цифровой WA 202
Осциллограф цифровой WA 212
Осциллограф цифровой GDS-71062A
Осциллограф цифровой GDS-71102A
Осциллограф цифровой GDS-71152A
Осциллограф цифровой DSO 1002A
Осциллограф цифровой DSO 1004A
Осциллограф цифровой DSO 1012A
Осциллограф цифровой DSO 1014A
Осциллограф цифровой DSO 1022A
Осциллограф цифровой WA 232
Осциллограф цифровой WS 24Xs-A
Осциллограф цифровой WS 24MXs-A
Осциллограф цифровой WS 44Xs-A
Осциллограф цифровой WS 44MXs-A
Осциллограф цифровой WS 42Xs-A
Осциллограф цифровой WS 64Xs-A
Осциллограф цифровой WS 62Xs-A
Осциллограф цифровой WS 64MXs-A
Осциллограф цифровой UTD2025C
Осциллограф цифровой DS1052D
Осциллограф цифровой DS1052E
Осциллограф цифровой DS1102D
Осциллограф цифровой DS1102E
Осциллограф цифровой DS1104B
Осциллограф цифровой DS1204B
Осциллограф цифровой АСК-2167
Осциллограф цифровой АСК-2064
Осциллограф цифровой DSO 8064A
Осциллограф цифровой АСК-5065
Осциллограф цифровой АСК-5105
Осциллограф цифровой GDS-73104
Осциллограф цифровой GDS-73202
Осциллограф цифровой GDS-73204
Осциллограф цифровой GDS-73302
Осциллограф цифровой GDS-73304
Осциллограф цифровой GDS-73502
Осциллограф цифровой GDS-73504
Осциллограф цифровой АОС-3182
Осциллограф цифровой UTD2052CL-R
Осциллограф цифровой АКИП-4115/5
Осциллограф цифровой TDS-2024C
Осциллограф цифровой TDS-2014C
Осциллограф цифровой TDS-2004C
Осциллограф цифровой TDS-2002C
Осциллограф цифровой TDS-2001C
Осциллограф цифровой АОС-5062
Осциллограф цифровой АОС-5064
Осциллограф цифровой АОС-5102
Осциллограф цифровой АОС-5104
Осциллограф цифровой АОС-5202
Осциллограф цифровой АОС-5204
Осциллограф цифровой АОС-5302
Осциллограф цифровой АОС-5304
Осциллограф цифровой АОС-5115
Осциллограф цифровой АОС-5110
Осциллограф цифровой АОС-5103
Осциллограф цифровой АКИП-4115/6
Осциллограф цифровой PDS-6042
Осциллограф цифровой PDS-6062T
Осциллограф цифровой UTD2102CEL-R
Осциллограф цифровой ADS-2061M
Осциллограф цифровой ADS-2061MV
Осциллограф цифровой ADS-2111M
Осциллограф цифровой ADS-2111MV
Осциллограф цифровой ADS-2121M
Осциллограф цифровой ADS-2121MV
Осциллограф цифровой АКИП-4116/1
Осциллограф цифровой GDS-73152
Осциллограф цифровой АОС-5074
Осциллограф цифровой GDS-73154
Осциллограф цифровой GDS-73252
Осциллограф цифровой GDS-73254
Осциллограф цифровой GDS-73352
Осциллограф цифровой GDS-73354
Осциллограф цифровой AFG-72125
Товары, похожие на Осцилограф
Вы можете заказать товар Осцилограф в компании Авител — электронные компоненты через нашу систему. В текущий момент товар находится в статусе «в наличии».
В текущий момент товар находится в статусе «в наличии».
Компания Авител — электронные компоненты является зарегистрированным поставщиком на сайте BizOrg.su.
Служебная информация:
На нашей площадке для удобства, каждой компании присвоен уникальный ID. Авител — электронные компоненты имеет ID 177409. Осцилограф имеет ID на сайте — 4906790. Если у вас возникли сложности при работе с компанией Авител — электронные компоненты – сообщите идентификаторы компании и товара/услуги в нашу службу технической поддержки.
Товар был добавлен на сайт 04/09/2013, дата последнего изменения — 09/04/2020. За все время товар был просмотрен 429 раз.
Обращаем ваше внимание на то, что торговая площадка BizOrg.su носит исключительно информационный характер и ни при каких условиях не является публичной офертой.
Заявленная компанией Авител — электронные компоненты цена товара «Осцилограф» может не быть окончательной ценой продажи. Для получения подробной информации о наличии и стоимости указанных товаров и услуг, пожалуйста, свяжитесь с представителями компании Авител — электронные компоненты по указанным телефону или адресу электронной почты.
Часы работы:
Телефоны:
+380 (44) 360-79-83
+380 (50) 061-63-98
+380 (97) 545-03-13
+380 (63) 744-32-07
+380 (89) 250-00-03
Купить осцилограф в Киеве:
Здолбуновская 9, Киев, Украина
Условия доставки из другого региона:
Доставка в страны:
– Россия
– Украина
NINscope, универсальный минископ для исследования цепей в нескольких областях
Abstract
Миниатюрные флуоресцентные микроскопы (минископы) сыграли важную роль в мониторинге нейронных сигналов во время неконтролируемого поведения, а их версии с открытым исходным кодом сделали их доступными. Часто размер и вес минископов с открытым исходным кодом приносятся в жертву дополнительным функциональным возможностям. Здесь мы представляем NINscope: легкий минископ с небольшой площадью основания, который объединяет высокочувствительный датчик изображения, инерциальный измерительный блок и светодиодный драйвер для внешнего оптогенетического зонда. Мы используем его, чтобы выполнить первые одновременные записи клеточного разрешения из мозжечка и коры головного мозга у мышей без ограничений, продемонстрировать его возможности оптогенетической стимуляции для изучения мозжечково-церебральной или кортико-стриарной связи, а также воспроизвести результаты кодирования действия в дорсальном стриатуме. В сочетании с кросс-платформенным программным обеспечением для сбора и управления наш минископ является универсальным дополнением к расширяющемуся набору инструментов минископов с открытым исходным кодом, которые расширят доступ к исследованиям цепей в нескольких регионах во время неограниченного поведения.
Мы используем его, чтобы выполнить первые одновременные записи клеточного разрешения из мозжечка и коры головного мозга у мышей без ограничений, продемонстрировать его возможности оптогенетической стимуляции для изучения мозжечково-церебральной или кортико-стриарной связи, а также воспроизвести результаты кодирования действия в дорсальном стриатуме. В сочетании с кросс-платформенным программным обеспечением для сбора и управления наш минископ является универсальным дополнением к расширяющемуся набору инструментов минископов с открытым исходным кодом, которые расширят доступ к исследованиям цепей в нескольких регионах во время неограниченного поведения.
Введение
Визуализация с клеточным разрешением с использованием миниатюрных флуоресцентных микроскопов (минископов) позволяет контролировать топологию активности в мозговых цепях во время неограниченного поведения. В то время как достижения в электрофизиологии теперь позволяют регистрировать одновременно многие тысячи нейронов у бодрствующих животных (Juavinett et al. , 2018; Jun et al., 2017), подходы к визуализации могут анализировать активность отдельных нейронов и сохранять информацию об их активности. пространственно распределены в крупную сеть (Terada et al., 2018; Stirman et al., 2016; Kim et al., 2016). Часто для кластерной активности существует анатомический субстрат, как в случае с мозжечком, где близлежащие клетки Пуркинье получают входные данные от лазающих волокон, происходящих из соседних нейронов нижнего ядра ствола мозга (Ruigrok, 2011). Таким образом, методы визуализации могут показать, как отдельные клетки, встроенные в более крупную сеть, проявляют скоординированную активность на разных этапах поведения или обучения (Wagner et al., 2017; Heffley et al., 2018; Galiñanes et al., 2018; Giovannucci et al. , 2017; Костадинов и др., 2019.). Более того, из-за их способности записывать свободно движущихся животных минископы сыграли важную роль в выявлении паттернов нейронной активности, возникающих во время естественного поведения и связанных с ним состояний мозга, включая социальные взаимодействия (Murugan et al.
, 2018; Jun et al., 2017), подходы к визуализации могут анализировать активность отдельных нейронов и сохранять информацию об их активности. пространственно распределены в крупную сеть (Terada et al., 2018; Stirman et al., 2016; Kim et al., 2016). Часто для кластерной активности существует анатомический субстрат, как в случае с мозжечком, где близлежащие клетки Пуркинье получают входные данные от лазающих волокон, происходящих из соседних нейронов нижнего ядра ствола мозга (Ruigrok, 2011). Таким образом, методы визуализации могут показать, как отдельные клетки, встроенные в более крупную сеть, проявляют скоординированную активность на разных этапах поведения или обучения (Wagner et al., 2017; Heffley et al., 2018; Galiñanes et al., 2018; Giovannucci et al. , 2017; Костадинов и др., 2019.). Более того, из-за их способности записывать свободно движущихся животных минископы сыграли важную роль в выявлении паттернов нейронной активности, возникающих во время естественного поведения и связанных с ним состояний мозга, включая социальные взаимодействия (Murugan et al. , 2017; Remedios et al., 2017; Liang). et al., 2018; Kingsbury et al., 2019) или спать (Chen et al., 2018; Cox et al., 2016) с полностью интактным вестибулярным входом.
, 2017; Remedios et al., 2017; Liang). et al., 2018; Kingsbury et al., 2019) или спать (Chen et al., 2018; Cox et al., 2016) с полностью интактным вестибулярным входом.
Минископы с открытым исходным кодом являются доступными инструментами для исследования клеточной активности у грызунов (Ghosh et al., 2011; Cai et al., 2016) и птиц (Liberti et al., 2016; Liberti et al., 2017) во время безудержного поведения и до недавнего времени ограничивались записями из одного региона, но см. Gonzalez et al. (2019). Чтобы понять, как мозговые цепи вызывают поведение, необходимо исследовать мультирегиональные взаимодействия во время спонтанного или обученного поведения. Если для этой цели используются минископы, они должны быть достаточно легкими и компактными, чтобы можно было делать записи с более чем одного места без ущерба для качества изображения, позволять прямое отслеживание поведения и иметь возможность оптогенетически управлять цепями. Чтобы удовлетворить все эти потребности в одном устройстве, мы разработали универсальный и компактный минископ (NINscope, названный в честь института происхождения) с чувствительным датчиком CMOS, встроенным инерциальным измерительным блоком (IMU) и точным светодиодным драйвером для оптогенетического запуска других области мозга с помощью изготовленного на заказ светодиодного зонда.
Используя возможности NINscope, мы демонстрируем его способность отслеживать функциональные взаимодействия между мозжечком и корой головного мозга у мышей без ограничений, носящих двойные минископы, одну из возможных конфигураций двойных эндоскопов с нашим минископом. Сложная спайковая активность в дендритах клеток Пуркинье коррелирует с нейронной активностью, измеренной в коре в периоды ускорения движения, в соответствии с ожиданиями, основанными на предыдущих анатомических и функциональных исследованиях мозжечково-таламо-кортикальной связи (Badura et al., 2018; Bostan et al. ., 2013; Аккал и др., 2007; Гувер и Стрик, 19 лет.99; Гао и др., 2018 г.; Вагнер и др., 2019). Используя встроенные в NINscope возможности оптогенетической стимуляции в сочетании с показаниями акселерометра, мы показываем, что стимуляция мозжечка вызывает четко различимые поведенческие реакции и активацию корковых нейронов.
Далее мы демонстрируем применимость NINscope для получения изображения нейронов дорсального полосатого тела мыши, глубокой области мозга, доступной только после имплантации релейной линзы GRIN. Встроенный акселерометр позволил нам идентифицировать клетки в дорсальном стриатуме, активность которых модулируется исключительно, когда мыши совершают повороты, противоположные месту записи, повторяя латерализацию в дорсальном стриатуме и роль этих нейронов в представлении пространства действия (Klaus et al., 2017). ; Barbera et al., 2016; Cui et al., 2013). Наконец, мы используем NINscope, чтобы показать, что оптогенетическая активация двух разных проекционных путей приводит к дифференциальной модуляции активности в нейронах дорсального полосатого тела.
Встроенный акселерометр позволил нам идентифицировать клетки в дорсальном стриатуме, активность которых модулируется исключительно, когда мыши совершают повороты, противоположные месту записи, повторяя латерализацию в дорсальном стриатуме и роль этих нейронов в представлении пространства действия (Klaus et al., 2017). ; Barbera et al., 2016; Cui et al., 2013). Наконец, мы используем NINscope, чтобы показать, что оптогенетическая активация двух разных проекционных путей приводит к дифференциальной модуляции активности в нейронах дорсального полосатого тела.
Наши проверочные эксперименты показывают, что NINscope позволяет проводить новые типы записи на мышах без ограничений. Мы демонстрируем возможность одновременной записи из двух областей одной и той же мыши, использования встроенного светодиодного драйвера в сочетании со светодиодным зондом для оптогенетической стимуляции и анализа поведенческих состояний с помощью включения акселерометра. Интеграция этих компонентов в одном устройстве облегчает сбор и анализ данных. NINscope — это проект с открытым исходным кодом, позволяющий другим использовать его дизайн и функциональные возможности, тем самым способствуя расширению диапазона инструментов с открытым исходным кодом для изучения нейронных цепей во время неконтролируемого поведения.
NINscope — это проект с открытым исходным кодом, позволяющий другим использовать его дизайн и функциональные возможности, тем самым способствуя расширению диапазона инструментов с открытым исходным кодом для изучения нейронных цепей во время неконтролируемого поведения.
Результаты
Дизайн и функциональность NINscope
NINscope (рис. 1A–C) отличается от других минископов тем, что сохраняет малый вес и небольшой форм-фактор, что позволяет вести запись на двух площадках, а также предлагает ряд новых интегрированных функций и межплатформенное взаимодействие (для сравнения с другими минископами см. Таблицу 1). ). Использовали более тонкий оптический эмиссионный фильтр и дихроичное зеркало (оба 500 мкм), а эмиссионный фильтр (1 мм) приклеивали к плосковыпуклой линзе с помощью оптического связующего клея (NOA81, Norland Products). Были разработаны изготовленные на заказ датчики межсоединительных разъемов высокой плотности (HDI) и интерфейсные печатные платы (10 на 10 мм, рис. 1B), которые были оснащены электронными компонентами с помощью устройства для захвата и установки (NeoDen4, NeoDen Tech, Ханчжоу, Китай) и уложены друг на друга для поддержания небольшой след. Это позволило интегрировать на плате IMU и несколько драйверов светодиодов, в том числе один для оптогенетического срабатывания и два для возбуждения светодиодов возбуждения. Последняя функция позволяет в будущем расширить режим с одинарного до двойного возбуждения.
1B), которые были оснащены электронными компонентами с помощью устройства для захвата и установки (NeoDen4, NeoDen Tech, Ханчжоу, Китай) и уложены друг на друга для поддержания небольшой след. Это позволило интегрировать на плате IMU и несколько драйверов светодиодов, в том числе один для оптогенетического срабатывания и два для возбуждения светодиодов возбуждения. Последняя функция позволяет в будущем расширить режим с одинарного до двойного возбуждения.
NINscope, компактный, легкий и универсальный минископ.
( A ) Схема NINscope с размерами в мм. Две печатные платы HDI размером 10 на 10 мм (PCB), одна для взаимодействия с блоком сбора данных (Interface PCB), а другая содержит датчик изображения CMOS (Sensor PCB), уложены друг на друга и смонтированы в корпусе, напечатанном на 3D-принтере. Возбуждающий свет от светодиода коллимируется полусферической линзой, проходит через возбуждающий фильтр и отражается дихроичным зеркалом на образец. Испускаемая флуоресценция собирается через линзу объектива GRIN и проходит через дихроичную и плосковыпуклую линзы, которые фокусируют изображение на датчике CMOS. На плосковыпуклую линзу с помощью оптического клея наклеен эмиссионный фильтр. ( B ) KiCad визуализирует специально созданный интерфейс и печатные платы датчиков с видами сверху и снизу. Плата интерфейса содержит инерциальный измерительный блок (IMU) для измерения ускорения и ориентации головы, три драйвера светодиодов, в том числе один для оптогенетического (стробирующего) управления и два для светодиодов возбуждения (один используется), а также красный следящий светодиод, сериализатор, и расширитель ввода-вывода. Печатная плата датчика содержит CMOS-сенсор PYTHON480, тактовый генератор и блок управления питанием, который обеспечивает датчик изображения необходимыми напряжениями, а также их синхронизацией и последовательностью. ( C ) Фотография NINscope с коаксиальным и натяжным кабелями, светодиодами возбуждения и оптогенетическими светодиодными кабелями.
Испускаемая флуоресценция собирается через линзу объектива GRIN и проходит через дихроичную и плосковыпуклую линзы, которые фокусируют изображение на датчике CMOS. На плосковыпуклую линзу с помощью оптического клея наклеен эмиссионный фильтр. ( B ) KiCad визуализирует специально созданный интерфейс и печатные платы датчиков с видами сверху и снизу. Плата интерфейса содержит инерциальный измерительный блок (IMU) для измерения ускорения и ориентации головы, три драйвера светодиодов, в том числе один для оптогенетического (стробирующего) управления и два для светодиодов возбуждения (один используется), а также красный следящий светодиод, сериализатор, и расширитель ввода-вывода. Печатная плата датчика содержит CMOS-сенсор PYTHON480, тактовый генератор и блок управления питанием, который обеспечивает датчик изображения необходимыми напряжениями, а также их синхронизацией и последовательностью. ( C ) Фотография NINscope с коаксиальным и натяжным кабелями, светодиодами возбуждения и оптогенетическими светодиодными кабелями. ( D ) Специально изготовленный имплантируемый светодиодный зонд для оптогенетической стимуляции, который подключается к NINscope с помощью оптогенетического светодиодного кабеля. ( E ) Карта UCLA Miniscope DAQ v3.2 использовалась с небольшими модификациями, включая 256 КБ x 8-бит I2C EEPROM (STMicroelectronics) и проводное соединение, соединяющее вход/выход общего назначения 2 (GPIO2) с контрольной точкой 4 ( ТР4). Сигналы последовательного периферийного интерфейса (SPI): ведущий выход, подчиненный вход (MOSI), последовательный сигнал (SCK) и выбор подчиненного устройства (SSN) подключаются к GPIO0, GPIO1 и GPIO3 через перемычки.
( D ) Специально изготовленный имплантируемый светодиодный зонд для оптогенетической стимуляции, который подключается к NINscope с помощью оптогенетического светодиодного кабеля. ( E ) Карта UCLA Miniscope DAQ v3.2 использовалась с небольшими модификациями, включая 256 КБ x 8-бит I2C EEPROM (STMicroelectronics) и проводное соединение, соединяющее вход/выход общего назначения 2 (GPIO2) с контрольной точкой 4 ( ТР4). Сигналы последовательного периферийного интерфейса (SPI): ведущий выход, подчиненный вход (MOSI), последовательный сигнал (SCK) и выбор подчиненного устройства (SSN) подключаются к GPIO0, GPIO1 и GPIO3 через перемычки.
Особенности NINscope и других минископов.
Обзор функций выпускаемых в настоящее время минископов с открытым исходным кодом и имеющихся в продаже (широкоугольное освещение). Опто-дистанционный: способность оптогенетически стимулировать вне поля зрения. Opto in-field: способность оптогенетически стимулировать поле визуализации. IMU: инерциальная единица измерения. eFocus: электрическая фокусировка с использованием электросмачиваемой линзы (EWL). 1: (Cai et al., 2016), 2: (Shuman et al., 2020), 3: (Leman et al., 2018), 4: (Jacob et al., 2018), 5: (Zhang et al. ., 2019), 6: (Барбера и др., 2019), 7: (Гонсалес и др., 2019). н.д. = данные недоступны. *Система nVoke. ** Оптогенетически синхронизированный флуоресцентный микроскоп.
Opto in-field: способность оптогенетически стимулировать поле визуализации. IMU: инерциальная единица измерения. eFocus: электрическая фокусировка с использованием электросмачиваемой линзы (EWL). 1: (Cai et al., 2016), 2: (Shuman et al., 2020), 3: (Leman et al., 2018), 4: (Jacob et al., 2018), 5: (Zhang et al. ., 2019), 6: (Барбера и др., 2019), 7: (Гонсалес и др., 2019). н.д. = данные недоступны. *Система nVoke. ** Оптогенетически синхронизированный флуоресцентный микроскоп.
| Минископ | Вес (г) | 3D-печать | Кроссплатформенный? | Два прицела? | Опто-пульт | Опто-дистанционный | Микрофон | IMU | eFocus |
|---|---|---|---|---|---|---|---|---|---|
| NINscope | 1.6 | yes | yes | yes | yes | no | no | yes | no |
| UCLA Miniscope v3 1 | 3. 2 2 | no | no | no | no | no | no | no | no |
| UCLA Miniscope wireless 2 | 4. 5 | no | no | no | no | № | NO | NO | NO |
| FINCHSCOPE 3 | 1,8 | NO | NO | .A.A.A.A. | no | no | yes | no | no |
| CHEndoscope 4 | 4.5 | yes | no | no | no | no | no | no | no |
| miniScope Lin 5 | 2. 4 4 | yes | yes | no | no | no | no | no | no |
| miniScope Lin Wireless 6 | scope: 3.9 battery:2.2– 5 | да | нет | нет | нет | нет | нет | нет | нет |
| н.д. | № | н.д. | yes | no | no | no | no | no | |
| Inscopix (commercial) | 1.8 and up | no | yes | n.a. | нет | да* | нет | нет | да |
| Линзы Doric (коммерческие) | 2,2, 27,0 кроме канюля | нет | нет | нет | нет | да** | нет | нет | да |
Чтобы обеспечить оптогенетическую стимуляцию вдали от места визуализации, мы разработали простой светодиодный зонд, состоящий из разъема Mill-Max, изолированных эмалевых проводов и SMD-светодиода с размером корпуса 402, который был загерметизирован эпоксидной смолой (рис. 1D).
1D).
Датчик PYTHON480 (ON Semiconductor) был выбран как компактный, но чувствительный датчик CMOS (размер пикселя: 4,8 мкм, динамический диапазон > 59 дБ) со скромными требованиями к мощности. В NINscope используется линза GRIN диаметром 1,8 мм (числовая апертура 0,55, #64–519)., Edmund Optics) в качестве объектива и имеет увеличение ~4,6x (рис. 1 — дополнение к рисунку 1). Это дало нам приблизительные размеры поля 786 на 502 мкм с использованием области интереса (ROI) 752 × 480 пикселей. В программном обеспечении мы реализовали возможность перевода этой области интереса на площадь сенсора CMOS 800 × 600 пикселей (836 на 627 мкм).
Из-за широкой доступности первого поколения UCLA Miniscope мы сохранили модуль сбора данных (DAQ V3.2) проекта UCLA Miniscope с небольшими изменениями, которые включали EEPROM для хранения увеличенной модифицированной версии последней версии Cypress EZ- Прошивка USB FX3 и проводное соединение от входа/выхода общего назначения 2 (GPIO2) к контрольной точке 4 (TP4), обеспечивающее точность синхронизации 1 мс для оптогенетического драйвера светодиодов (рис. 1E). Прошивка модуля сбора данных была изменена, чтобы обеспечить последовательное управление яркостью оптогенетического и возбуждающего светодиодов, а также усилением, экспозицией и уровнем черного CMOS-сенсора.
1E). Прошивка модуля сбора данных была изменена, чтобы обеспечить последовательное управление яркостью оптогенетического и возбуждающего светодиодов, а также усилением, экспозицией и уровнем черного CMOS-сенсора.
Корпус микроскопа был напечатан на 3D-принтере (принтер EnvisionTec Micro Plus Advantage, смола RCP30 M и принтер Formlabs Form 2, черная смола RS-F2-GPBK-04), что позволяет быстро создавать прототипы различных конструкций минископов. Точность печати оказалась достаточной для нашей окончательной конструкции, что позволило нам снизить вес минископа до 1,6 грамма (корпус + оптика + печатные платы), при этом позволяя использовать два минископа одновременно на одной мыши. В верхней половине NINscope есть корпус для сенсорных и интерфейсных печатных плат, чтобы защитить их от повреждений во время бесконтрольного поведения животных. NINscope крепится на малогабаритной опорной плите (внешние размеры 6,5 на 7,5 мм, рисунок 1 — дополнение к рисунку 2) с помощью установочного винта. Нижняя половина корпуса NINscope имеет выступ, который входит в выемку на опорной плите для повышения устойчивости.
Нижняя половина корпуса NINscope имеет выступ, который входит в выемку на опорной плите для повышения устойчивости.
На языке Processing было разработано удобное для пользователя программное обеспечение (http://processing.org), обеспечивающее настоящую межплатформенную совместимость и контроль над экспериментальными записями (рис. 1 — дополнение к рис. 3). Была включена возможность записи как в режиме с одной, так и с двумя головками (два минископа) в сочетании с дополнительной веб-камерой USB для видеосъемки поведения. Кольцевые буферы были реализованы в программном обеспечении, чтобы избежать временных задержек кадров и пропусков кадров во время сбора данных. Временные метки полученных кадров регистрировались для постфактум синхронизации. Мы протестировали сбор данных на ряде конфигураций компьютеров с различными типами операционных систем (Windows, MacOS, Linux), процессорами (i5, i7) и жесткими дисками (SATA, SSD). Данные акселерометра IMU могут отображаться в реальном времени во время записи минископа. Кроме того, паттерны оптогенетической стимуляции и ток светодиодного зонда можно регулировать через интерфейс, обеспечивающий интегрированный контроль всех аспектов эксперимента (рис. 1 — дополнение к рис. 3). Более подробное описание аппаратного и программного обеспечения представлено в разделе «Материалы и методы». Файлы дизайна и инструкции по сборке оборудования (для минископов и светодиодных датчиков), программированию прошивки и установке программного обеспечения можно найти на нашем сайте GitHub: https://github.com/ninscope.
Кроме того, паттерны оптогенетической стимуляции и ток светодиодного зонда можно регулировать через интерфейс, обеспечивающий интегрированный контроль всех аспектов эксперимента (рис. 1 — дополнение к рис. 3). Более подробное описание аппаратного и программного обеспечения представлено в разделе «Материалы и методы». Файлы дизайна и инструкции по сборке оборудования (для минископов и светодиодных датчиков), программированию прошивки и установке программного обеспечения можно найти на нашем сайте GitHub: https://github.com/ninscope.
Чтобы проверить наш минископ, мы сделали изображения в различных конфигурациях записи и стимуляции, включая режимы одиночного и двойного эндоскопа в разных областях мозга. За поведением животных следили с помощью веб-камеры USB (рис. 2А), светодиодного индикатора минископа (рис. 2В) и бортового акселерометра. В показанном примере клетки Пуркинье в дольке V мозжечка (AP: -6,4, ML: 0 мм) были трансдуцированы с помощью GCaMP6f, их дендриты были визуализированы с помощью NINscope и сегментированы с использованием пакета CNMF-E (Zhou et al. , 2018). ) (рис. 2C,D, рис. 2—видео 1). Для всех анализов переходные процессы деконволюции кальция (выходные данные нейрона C, полученные после запуска CNMF-E; см. Vogelstein et al., 2009).) использовались для извлечения времени события (рис. 2C). Лазающие волокна, происходящие от нижней оливы в вентральной части продолговатого мозга, вызывают сложные спайки в клетках Пуркинье; они вызывают приток кальция по всему дендритному дереву клеток Пуркинье вплоть до поверхности мозга, а частота их возбуждения стабильно поддерживается на уровне около 1 Гц (Ju et al., 2019). Парасагиттальное расположение дендритов клеток Пуркинье, каждый из которых имеет ширину до ~ 150 мкм и толщину ~ 4–6 мкм (Nedelescu and Abdelhack, 2013), делает их пригодными для визуализации в минископе (Ghosh et al., 2011). . Таким образом, мы смогли извлечь парасагиттально выровненные пространственные следы с помощью CNMF-E (см. Материалы и методы для конкретных настроек параметров).
, 2018). ) (рис. 2C,D, рис. 2—видео 1). Для всех анализов переходные процессы деконволюции кальция (выходные данные нейрона C, полученные после запуска CNMF-E; см. Vogelstein et al., 2009).) использовались для извлечения времени события (рис. 2C). Лазающие волокна, происходящие от нижней оливы в вентральной части продолговатого мозга, вызывают сложные спайки в клетках Пуркинье; они вызывают приток кальция по всему дендритному дереву клеток Пуркинье вплоть до поверхности мозга, а частота их возбуждения стабильно поддерживается на уровне около 1 Гц (Ju et al., 2019). Парасагиттальное расположение дендритов клеток Пуркинье, каждый из которых имеет ширину до ~ 150 мкм и толщину ~ 4–6 мкм (Nedelescu and Abdelhack, 2013), делает их пригодными для визуализации в минископе (Ghosh et al., 2011). . Таким образом, мы смогли извлечь парасагиттально выровненные пространственные следы с помощью CNMF-E (см. Материалы и методы для конкретных настроек параметров).
Визуализация мозжечка с помощью NINscope.

( A ) Мышь с одним NINscope, установленным над V долькой мозжечка, где клетки Пуркинье были селективно трансдуцированы с помощью GCaMP6f. ( B ) Положение животного можно определить с помощью светодиода слежения в сочетании с одновременными записями с веб-камеры. Цвета представляют время с начала записи, а дорожка представляет исследование клетки в течение ~ 2 минут. ( C ) Пространственные следы дендритов клеток Пуркинье (A), необработанные (C_raw) и деконволютированные сигналы ( C ) были извлечены с использованием CNMF-E (Zhou et al., 2018) после коррекции движения с помощью NoRMCorre (Pnevmatikakis and Giovannucci, 2017). ). Время события (Events) было извлечено из сигналов после деконволюции. Показанный сигнал изображен в рамочной области на D. ( D ) Пространственные следы 15 дендритных деревьев клеток Пуркинье (A) и соответствующие им переходные процессы кальция (C_raw), растр событий по всем (168) извлеченным сигналам, x, y , z каналов акселерометра, а также средний необработанный сигнал (C_raw mean) и сумму всех событий (Event sum). ( E ) Переходные процессы дендритов клеток Пуркинье были выровнены по началу ускорения в заштрихованной красным области на D. В этом примере рефлекторное движение (подергивание) было вызвано громким хлопком. ( F ) Спонтанное поведение, отслеживаемое с помощью веб-камеры, может быть связано с отчетливыми сигнатурами в данных акселерометра, например, во время подъема. В этой записи подмножество клеток Пуркинье продемонстрировало значительный ответ во время выращивания животных с временными подъемами, связанными с фазами подъема и падения мыши (среднее значение 4 подъемов, среднее ± SEM, N = 1 мышь). Пунктирные линии указывают моменты времени, в которые были захвачены изображения с веб-камеры.
( E ) Переходные процессы дендритов клеток Пуркинье были выровнены по началу ускорения в заштрихованной красным области на D. В этом примере рефлекторное движение (подергивание) было вызвано громким хлопком. ( F ) Спонтанное поведение, отслеживаемое с помощью веб-камеры, может быть связано с отчетливыми сигнатурами в данных акселерометра, например, во время подъема. В этой записи подмножество клеток Пуркинье продемонстрировало значительный ответ во время выращивания животных с временными подъемами, связанными с фазами подъема и падения мыши (среднее значение 4 подъемов, среднее ± SEM, N = 1 мышь). Пунктирные линии указывают моменты времени, в которые были захвачены изображения с веб-камеры.
Для всех записей для получения сигнала высокого качества требовалась минимальная мощность света (~120–240 мкВт до, 30–140 мкВт после входа в объектив GRIN, в пределах линейного диапазона драйвера светодиода возбуждения, рис. 2 — дополнение к рис. 1). записи отношения шума к шуму с использованием объектива GRIN, установленного на поверхности мозга (рис. 2D, рис. 2 — видео 1). Во время наших сеансов визуализации не наблюдалось обесцвечивания сигнала или фотоповреждений (обычно 5 000–20 000 кадров, то есть 2–11 минут на сеанс, повторяемый с короткими перерывами не менее 10 раз). Каналы акселерометра x, y и z использовались для обнаружения начала движения (рис. 2E) и определения специфического поведения, такого как вставание на дыбы (рис. 2F).
1). записи отношения шума к шуму с использованием объектива GRIN, установленного на поверхности мозга (рис. 2D, рис. 2 — видео 1). Во время наших сеансов визуализации не наблюдалось обесцвечивания сигнала или фотоповреждений (обычно 5 000–20 000 кадров, то есть 2–11 минут на сеанс, повторяемый с короткими перерывами не менее 10 раз). Каналы акселерометра x, y и z использовались для обнаружения начала движения (рис. 2E) и определения специфического поведения, такого как вставание на дыбы (рис. 2F).
Визуализация двух областей с помощью NINscope
Стимулом для создания более легких и компактных минископов является возможность одновременной записи из двух областей у мышей без ограничений, не нарушая спонтанного поведения. Например, можно было бы получить одновременные записи клеточного разрешения мозжечка и коры головного мозга. Существует достаточно анатомических данных о мозжечково-таламо-церебральных петлях (Hoover and Strick, 1999; Akkal et al., 2007; Bostan et al. , 2013), и все большее число исследований предполагает, что функциональные взаимодействия внутри таких петель важны для правильное выражение социального, когнитивного и моторного (планирующего) поведения (Badura et al., 2018; Stoodley et al., 2017; Gao et al., 2018). Возможность одновременной записи как мозжечка, так и коры головного мозга у мышей без ограничений открывает возможность изучения того, как эти взаимодействия проявляются во время естественного спонтанного поведения. Более того, двухполушарные записи гиппокампа (Gonzalez et al., 2019), неокортекс или мозжечок могут выявить стабильность, избыточность и латерализацию нейронных представлений во время обучения и контроля поведения.
, 2013), и все большее число исследований предполагает, что функциональные взаимодействия внутри таких петель важны для правильное выражение социального, когнитивного и моторного (планирующего) поведения (Badura et al., 2018; Stoodley et al., 2017; Gao et al., 2018). Возможность одновременной записи как мозжечка, так и коры головного мозга у мышей без ограничений открывает возможность изучения того, как эти взаимодействия проявляются во время естественного спонтанного поведения. Более того, двухполушарные записи гиппокампа (Gonzalez et al., 2019), неокортекс или мозжечок могут выявить стабильность, избыточность и латерализацию нейронных представлений во время обучения и контроля поведения.
Учитывая уменьшенные размеры и вес NINscope, мы смогли установить два минископа на одну мышь (рис. 3A). Мы выбрали запись из мозжечка и коры, чтобы определить, в какой степени ускорение движения коррелирует с нейронной активностью в этих двух областях, что можно было бы напрямую решить с помощью NINscope, учитывая его встроенный акселерометр.
Визуализация двух областей с помощью NINscope.
( A ) Мышь с двумя NINscopes, установленными над мозжечком и корой. ( B ) Поведение не нарушалось, что определялось количественно путем подсчета количества вставаний и их продолжительности у мышей с одинарным или двойным минископом (подъемы в минуту, p = 0,7864, нс; продолжительность вставания, p = 0,4244, нс; N = 4 мыши). ). ( C ) Череп мыши с красными кружками, указывающими конфигурацию записи (позиции краниотомии). Для других возможных конфигураций см. Рисунок 3 — дополнение к рисунку 1 и Рисунок 3 — дополнение к рисунку 2. ( D ) Визуализация CAD, показывающая ростро-каудальное размещение двух NINscopes для одновременного изображения мозжечка и коры (расстояние между опорными пластинами ~ 8 мм) под углом 15–20°). ( E ) GCaMP6f селективно трансдуцировался в клетки Пуркинье мозжечка (долька VI или простая долька) и в нейроны моторной коры. ( F ) Двойные записи из мозжечковой дольки VI и двигательной коры, показывающие ответы сегментированных нейронов (пространственные следы, A) в каждой области с сигналом Z-среднего значения, числом коактивных дендритов клеток Пуркинье и составным сигналом ускорения (a xyz , √(x 2 +y 2 +z 2 )). Голубые линии представляют эпохи, когда синхронные паттерны (СП) были обнаружены в мозжечке и коре. ( G ) Комбинированные дуговые графики для этого набора данных, визуализирующие внутримозжечковые, внутримозговые (внутри) и мозжечково-мозговые (поперек) SP. Радиусы узлов масштабируются по количеству ячеек, с которыми соединяется узел. В этом примере нейроны мозжечка с высоким уровнем SP также демонстрируют значительные SP в разных регионах. ( H ) SP использовались для срабатывания составного сигнала ускорения. Ускорение поведения может быть отнесено к четырем категориям, состоящим из отсутствия изменений (отсутствие Δ, 64%, отсортировано по пиковой реакции), ускорение поведения после ПП (31%), до ПП (4%) или около ПП (1%).
( F ) Двойные записи из мозжечковой дольки VI и двигательной коры, показывающие ответы сегментированных нейронов (пространственные следы, A) в каждой области с сигналом Z-среднего значения, числом коактивных дендритов клеток Пуркинье и составным сигналом ускорения (a xyz , √(x 2 +y 2 +z 2 )). Голубые линии представляют эпохи, когда синхронные паттерны (СП) были обнаружены в мозжечке и коре. ( G ) Комбинированные дуговые графики для этого набора данных, визуализирующие внутримозжечковые, внутримозговые (внутри) и мозжечково-мозговые (поперек) SP. Радиусы узлов масштабируются по количеству ячеек, с которыми соединяется узел. В этом примере нейроны мозжечка с высоким уровнем SP также демонстрируют значительные SP в разных регионах. ( H ) SP использовались для срабатывания составного сигнала ускорения. Ускорение поведения может быть отнесено к четырем категориям, состоящим из отсутствия изменений (отсутствие Δ, 64%, отсортировано по пиковой реакции), ускорение поведения после ПП (31%), до ПП (4%) или около ПП (1%). , не показаны). ( I ) В разных регионах SP связаны со значительными отклонениями от базовой линии в составном сигнале акселерометра. Отзывчивые клетки показаны в мозжечке (долька VI) и коре головного мозга, запускаемые SP. Мышь, отдыхающая перед SP, совершила движение (влево, вверх) вокруг начала SP. Движение животного, визуализируемое с помощью оптического потока, имеет цветовую маркировку. ( J ) Усредненные по популяции ответы, запускаемые вокруг обнаруженных SP, обнаруживают реакции в мозжечке и коре во время подъема акселерометра. ( K ) Пример, показывающий нейроны мозжечка и коры, которые участвовали в SP (красный), и клетки, которые не участвовали (голубой).
, не показаны). ( I ) В разных регионах SP связаны со значительными отклонениями от базовой линии в составном сигнале акселерометра. Отзывчивые клетки показаны в мозжечке (долька VI) и коре головного мозга, запускаемые SP. Мышь, отдыхающая перед SP, совершила движение (влево, вверх) вокруг начала SP. Движение животного, визуализируемое с помощью оптического потока, имеет цветовую маркировку. ( J ) Усредненные по популяции ответы, запускаемые вокруг обнаруженных SP, обнаруживают реакции в мозжечке и коре во время подъема акселерометра. ( K ) Пример, показывающий нейроны мозжечка и коры, которые участвовали в SP (красный), и клетки, которые не участвовали (голубой).
С помощью двух минископов мыши исследовали клетку, ухаживали за ней, ели, прыгали (рис. 3 — видео 1) и вставали на дыбы, как это наблюдалось у мышей с одним минископом. Мы количественно оценили количество подъемов, а также их продолжительность у мышей, носящих один или два минископа (рис.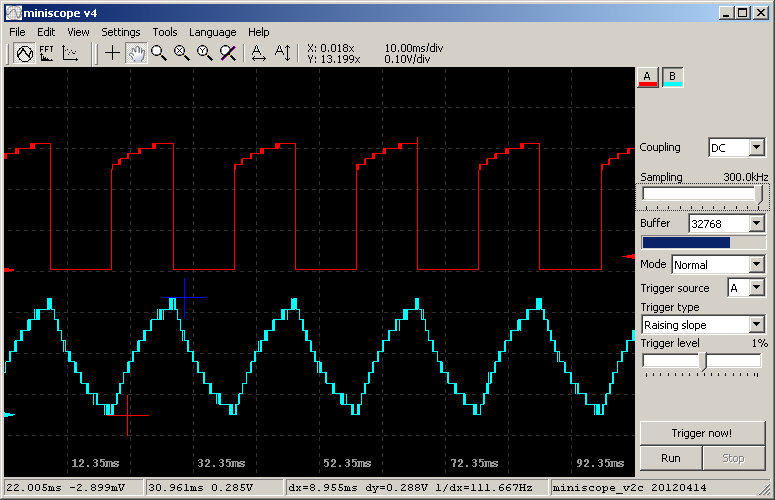 3B). У мышей, носивших два минископа, ни количество подергиваний, ни их продолжительность не изменились (количество подергиваний на один эндоскоп = 1,47 ± 1,07 подергивания в минуту, среднее значение ± SD; количество подергиваний на два эндоскопа: 1,36 ± 0,75 подергивания в минуту, среднее значение ± SD, p=0,7864, нс; продолжительность выращивания на одном эндоскопе = 2,21 ± 1,11 с, среднее ± SD; продолжительность выращивания на двух эндоскопах = 1,84 ± 0,77 с, среднее ± SD, p = 0,4244, нс; N = 4 мыши, критерий Колмогорова-Смирнова). Помимо установки одного эндоскопа над мозжечком и одного над корой, мы проверили способность мышей носить двойные эндоскопы в двух альтернативных конфигурациях (рис. 3 — дополнение к рисунку 1). Одна конфигурация с минископами, установленными над двумя полушариями зрительной коры, что также открывает возможность записи из подлежащей области CA1 гиппокампа (количество поднятий: 1,5 ± 0,9).5, среднее ± стандартное отклонение; продолжительность выращивания: 2,4 ± 1,15, среднее значение ± стандартное отклонение, N = 1 мышь).
3B). У мышей, носивших два минископа, ни количество подергиваний, ни их продолжительность не изменились (количество подергиваний на один эндоскоп = 1,47 ± 1,07 подергивания в минуту, среднее значение ± SD; количество подергиваний на два эндоскопа: 1,36 ± 0,75 подергивания в минуту, среднее значение ± SD, p=0,7864, нс; продолжительность выращивания на одном эндоскопе = 2,21 ± 1,11 с, среднее ± SD; продолжительность выращивания на двух эндоскопах = 1,84 ± 0,77 с, среднее ± SD, p = 0,4244, нс; N = 4 мыши, критерий Колмогорова-Смирнова). Помимо установки одного эндоскопа над мозжечком и одного над корой, мы проверили способность мышей носить двойные эндоскопы в двух альтернативных конфигурациях (рис. 3 — дополнение к рисунку 1). Одна конфигурация с минископами, установленными над двумя полушариями зрительной коры, что также открывает возможность записи из подлежащей области CA1 гиппокампа (количество поднятий: 1,5 ± 0,9).5, среднее ± стандартное отклонение; продолжительность выращивания: 2,4 ± 1,15, среднее значение ± стандартное отклонение, N = 1 мышь). Другая конфигурация с двумя минископами, установленными над полушариями мозжечка (Crus I) (количество подъемов: 1,8 ± 0,6, среднее ± SD; продолжительность подъема: 1,84 ± 0,82, среднее ± SD, N = 1 мышь). Для этих двух дополнительных конфигураций количество подъемов и их продолжительность были сопоставимы с конфигурациями с одним эндоскопом или корой мозжечка (рис. 3 — дополнение к рисунку 1). Гнездовой дисперсионный анализ для сравнения всех конфигураций минископа (одинарного и двойного) показал, что ни продолжительность выращивания (F (3,6) :1,859, p=0,23), ни частота выращивания (F (3,6) :0,221, p=0,88) существенно не различались. Апостериорный тест Тьюки для множественных сравнений подтвердил, что ни одно из взаимодействий между различными конфигурациями не было значительным (рис. 3 — исходные данные 1).
Другая конфигурация с двумя минископами, установленными над полушариями мозжечка (Crus I) (количество подъемов: 1,8 ± 0,6, среднее ± SD; продолжительность подъема: 1,84 ± 0,82, среднее ± SD, N = 1 мышь). Для этих двух дополнительных конфигураций количество подъемов и их продолжительность были сопоставимы с конфигурациями с одним эндоскопом или корой мозжечка (рис. 3 — дополнение к рисунку 1). Гнездовой дисперсионный анализ для сравнения всех конфигураций минископа (одинарного и двойного) показал, что ни продолжительность выращивания (F (3,6) :1,859, p=0,23), ни частота выращивания (F (3,6) :0,221, p=0,88) существенно не различались. Апостериорный тест Тьюки для множественных сравнений подтвердил, что ни одно из взаимодействий между различными конфигурациями не было значительным (рис. 3 — исходные данные 1).
Наша конструкция минископа позволяет проводить записи из двух областей у мышей с расстоянием между базальными пластинками ~8 мм и минископами, расположенными под углом 15–20° в случае одновременных записей из мозжечка и коры (или, альтернативно, из дорсального полосатого тела, рис. 3C,D) и ~ 6–7 мм под углом 45–50° при расположении рядом друг с другом для двухполушарных записей мозжечка или коры. Эти значения были получены с использованием точных трехмерных моделей черепа мыши C57BL/6 и двух NINscopes с базовыми пластинами с использованием программного обеспечения для моделирования САПР. Таким образом, основываясь на наших выводах, NINscope позволяет проводить записи в двух местах с несколькими конфигурациями у мышей без ограничений (рис. 3 — дополнение к рисунку 2).
3C,D) и ~ 6–7 мм под углом 45–50° при расположении рядом друг с другом для двухполушарных записей мозжечка или коры. Эти значения были получены с использованием точных трехмерных моделей черепа мыши C57BL/6 и двух NINscopes с базовыми пластинами с использованием программного обеспечения для моделирования САПР. Таким образом, основываясь на наших выводах, NINscope позволяет проводить записи в двух местах с несколькими конфигурациями у мышей без ограничений (рис. 3 — дополнение к рисунку 2).
В нашей конкретной демонстрации визуализации с двумя участками были сделаны инъекции вируса для селективной трансдукции GCaMP6f в мозжечковые клетки Пуркинье простой мозжечковой дольки (центр объектива GRIN AP: -5,8, ML: 2,2 мм) или дольки VI (центр объектива GRIN AP: −7,4, ML:0,0 мм), а также глобально в нейронах моторной коры (AP объективного центра GRIN: +1,4, ML: 1,5), охватывающих большие участки каудальной области передних конечностей и небольшую часть M2 на ростральной границе линзы объектива GRIN (рис.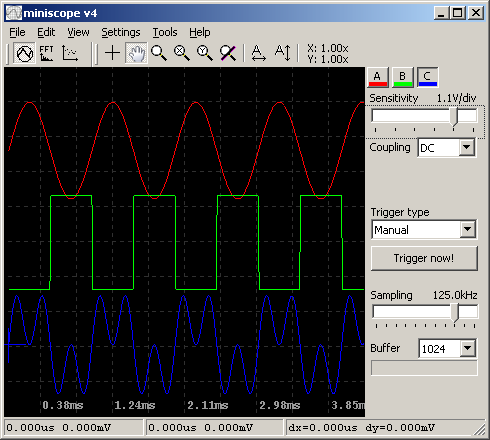 3E).
3E).
Артефакты движения мозга часто встречаются во время визуализации in vivo как у животных с фиксированной головой, так и у животных без фиксации, и для их коррекции были разработаны алгоритмические подходы (Pnevmatikakis and Giovannucci, 2017). Артефакты движения в наших данных были, по-видимому, незначительными даже у мышей, носящих двойные минископы (рис. 3 — видео 2), что мы дополнительно проверили, введя статические флуоресцентные шарики в мозжечок и кору головного мозга, чтобы отслеживать движение шариков во время ничем не ограниченного поведения (рис. 3 — дополнение к рисунку 3). , Рисунок 3—видео 3).
Сигналы могут быть извлечены из сотен клеток мозжечка и коры (мозжечок: 141 ± 53, диапазон 62–200, кора: 201 ± 90, диапазон 89–361, среднее ± SD, N = 4 мыши, рис. 3F) . Переходные процессы кальция из дендритов клеток Пуркинье, экспрессирующих GCaMP6f, демонстрировали более быструю кинетику распада, чем клетки коры, экспрессирующие тот же сенсорный белок кальция (t 1/2 мозжечка: 0,217 ± 0,12 с, 15784 транзиентов; t 1/2 коры: 0,488 ± 0,34). с, 9424 переходных процесса, среднее ± стандартное отклонение, G Хеджа = 1,19), вероятно, отражающие различия в буферизации эндогенного кальция (Celio, 1990; Baimbridge et al., 1992). Частота событий, измеренная на дендритах клеток Пуркинье, соответствовала основной частоте поступления лазящих волокон (0,62 ± 0,39 Гц, среднее значение ± стандартное отклонение, 1629 клеток, N = 4 мыши), в то время как нейроны коры запускали события с более низкой частотой (0,22 ± 0,28 Гц, среднее ± SD, 2212 клеток, N = 4 мыши).
с, 9424 переходных процесса, среднее ± стандартное отклонение, G Хеджа = 1,19), вероятно, отражающие различия в буферизации эндогенного кальция (Celio, 1990; Baimbridge et al., 1992). Частота событий, измеренная на дендритах клеток Пуркинье, соответствовала основной частоте поступления лазящих волокон (0,62 ± 0,39 Гц, среднее значение ± стандартное отклонение, 1629 клеток, N = 4 мыши), в то время как нейроны коры запускали события с более низкой частотой (0,22 ± 0,28 Гц, среднее ± SD, 2212 клеток, N = 4 мыши).
Во всех записях мы наблюдали отчетливые периоды повышенной активности в разных регионах (рис. 3 — видео 4) вокруг периодов движения животных (рис. 3F), судя по комплексному сигналу ускорения и проверке необработанных данных. Для количественной оценки корреляций в мозжечке (долька VI, N = 2, простая долька, N = 2; N = 4 мыши) и коре головного мозга время начала кальциевого переходного процесса, извлеченное из деконволютивных переходных процессов, свертывалось с ядром Епанечникова и суммировалось по всем клеткам. Это привело к сумме ядер, в которой глобальные синхронные паттерны (SP) были определены как случаи, когда сумма ядер превышала среднее значение +2σ и где эти паттерны встречались не менее пяти раз (голубые линии на рисунке 3F). Визуализация функциональной связности клеток показана на комбинированных дуговых графиках на рисунке 3G, где размер узла зависит от количества SP в мозжечке (долька VI в этом примере), в коре или в двух областях. Как клетки Пуркинье мозжечка, так и нейроны коры демонстрировали SP внутри и между областями, при этом клетки Пуркинье имели значительно больше внутри-SP, чем нейроны коры (доля общей дольки VI: 66,4 ± 18,9).%, простая долька: 73,3 ± 27,4%, кора: 35,7 ± 17%, среднее ± SD, CBL vs CTX p = 2,7801e-17, критерий Колмогорова-Смирнова). Значительно больше клеток Пуркинье участвовало в кросс-СП, чем нейроны коры (доля общей дольки VI: 42,3 ± 29,6%; фракция простой дольки: 47,4 ± 31,6%; кора: 26,7 ± 18,5%, среднее ± SD; CBL против CTX p = 1,8409 е-15, тест Колмогорова-Смирнова).
Это привело к сумме ядер, в которой глобальные синхронные паттерны (SP) были определены как случаи, когда сумма ядер превышала среднее значение +2σ и где эти паттерны встречались не менее пяти раз (голубые линии на рисунке 3F). Визуализация функциональной связности клеток показана на комбинированных дуговых графиках на рисунке 3G, где размер узла зависит от количества SP в мозжечке (долька VI в этом примере), в коре или в двух областях. Как клетки Пуркинье мозжечка, так и нейроны коры демонстрировали SP внутри и между областями, при этом клетки Пуркинье имели значительно больше внутри-SP, чем нейроны коры (доля общей дольки VI: 66,4 ± 18,9).%, простая долька: 73,3 ± 27,4%, кора: 35,7 ± 17%, среднее ± SD, CBL vs CTX p = 2,7801e-17, критерий Колмогорова-Смирнова). Значительно больше клеток Пуркинье участвовало в кросс-СП, чем нейроны коры (доля общей дольки VI: 42,3 ± 29,6%; фракция простой дольки: 47,4 ± 31,6%; кора: 26,7 ± 18,5%, среднее ± SD; CBL против CTX p = 1,8409 е-15, тест Колмогорова-Смирнова).
Затем мы оценили, связана ли коррелирующая активность в мозжечке и коре с поведенческим ускорением. Чтобы определить, предшествовал он или следовал за SP в разных регионах, составной сигнал ускорения запускался для разных SP. Мы можем отнести поведенческое ускорение к четырем категориям (рис. 3H), состоящим из отсутствия существенных изменений (64%, 206/321 среди SP), поведенческого ускорения после SP (31%, 99/321), до SP (4%, 13/321) или около SP (1%, 3/321). Таким образом, большинство (86%, 99/115) СП связаны со значительным ускорением после прохождения через СП. Мы также подтвердили, что СП связаны с событиями ускорения, сравнив долю случайного ускорения по сравнению с ускорением, вызванным СП, превышающим среднее значение +2σ от исходного уровня, чтобы показать, что доля событий ускорения, вызванных СП, не могла возникнуть случайно (случайное по сравнению с СП-ускорением). триггерный, р=0,0017, критерий Колмогорова-Смирнова).
Как и ожидалось, переходные процессы кальция в клетках Пуркинье мозжечка и корковых нейронах происходили вокруг поведенческого ускорения (рис. 3I, J). В показанном примере (запущенном поперек SP, обозначенном голубыми треугольниками на рисунках 3F и I) в составном сигнале акселерометра произошло большое отклонение вокруг начала SP. Это движение также было замечено при просмотре параллельных кадров изображения с веб-камеры и визуализировано здесь с помощью оптического потока с цветами, представляющими направление движения до (вверху) и вокруг начала SP (внизу).
3I, J). В показанном примере (запущенном поперек SP, обозначенном голубыми треугольниками на рисунках 3F и I) в составном сигнале акселерометра произошло большое отклонение вокруг начала SP. Это движение также было замечено при просмотре параллельных кадров изображения с веб-камеры и визуализировано здесь с помощью оптического потока с цветами, представляющими направление движения до (вверху) и вокруг начала SP (внизу).
Сопоставление записей (N = 4 животных, девять записей) с началом после SP выявило явный подъем сигнала акселерометра, совпадающий с увеличением содержания кальция как в клетках Пуркинье мозжечка, так и в корковых нейронах (рис. 3J, K). Хотя электрические записи обеспечивают более точную оценку латентности, чем визуализация кальция, мы исследовали относительную синхронизацию ответов кальция в мозжечке и коре по отношению к ускорению движения. Латентность пикового ответа переходных процессов кальция в мозжечке, коре и составного сигнала ускорения показала, что максимальное ускорение наступало после пикового ответа как в мозжечке, так и в коре (CBL: 444 ± 280 мс, CTX: 505 ± 260 мс, xyz : 700 ± 269 мс, критерий Краскела-Уоллиса p = 1,6553e-08). Апостериорный тест с использованием S Шеффе показал, что средние ранги пиковых латентностей переходных процессов кальция значительно различались по сравнению с задержанными пиковыми задержками сигнала акселерометра, в то время как это не применялось при сравнении средних рангов переходных процессов кальция в мозжечке и коре. . Таким образом, на основе наших предварительных данных, зарегистрированных с помощью NINscope, мы показываем, что координированная мозжечково-мозговая активность обычно предшествует пиковому ускорению движения.
Апостериорный тест с использованием S Шеффе показал, что средние ранги пиковых латентностей переходных процессов кальция значительно различались по сравнению с задержанными пиковыми задержками сигнала акселерометра, в то время как это не применялось при сравнении средних рангов переходных процессов кальция в мозжечке и коре. . Таким образом, на основе наших предварительных данных, зарегистрированных с помощью NINscope, мы показываем, что координированная мозжечково-мозговая активность обычно предшествует пиковому ускорению движения.
Мультисайтовая оптогенетическая стимуляция
До сих пор минископическая оптогенетика фокусировалась на стимуляции в пределах одного и того же поля зрения (Stamatakis et al., 2018). Стимуляция в пределах одного и того же поля зрения накладывает ограничения на конструкцию минископа, требуя дополнительной оптики, которая увеличивает вес и площадь основания минископа. Это также требует использования опсина, который может быть спектрально отделен от индикатора активности. Более того, часто желательно стимулировать участок, удаленный от минископа, например, для активации непрямых проекционных путей. Путем непосредственного управления светодиодом от минископа количество кабелей может быть уменьшено, а начало стимуляции может быть напрямую записано на диск вместе с кадрами изображения и данными акселерометра для облегчения последующего анализа. Кроме того, можно использовать несколько зондов для стимуляции более чем в одном месте у одного и того же животного. Используя встроенный оптогенетический светодиодный драйвер NINscope, мы демонстрируем его использование путем стимуляции клеток Пуркинье мозжечка в трансгенных Pcp2 -Cre Jdhu x Ai32(RCL-ChR2(h234R)/EYFP) мышей при выполнении визуализации кальция из нейронов в моторной коре, трансдуцированных с помощью GCaMP6f (рис. 4A–B, центр объектива GRIN AP: +1,4, ML: 1,5) . Мы выбрали четыре местоположения в мозжечке для размещения синих (470 нм) оптогенетических светодиодных зондов: II голени ипси- и контралатеральнее места визуализации в правом полушарии, контралатеральной симплексной дольки и дольки VI в медиальном черве мозжечка.
Более того, часто желательно стимулировать участок, удаленный от минископа, например, для активации непрямых проекционных путей. Путем непосредственного управления светодиодом от минископа количество кабелей может быть уменьшено, а начало стимуляции может быть напрямую записано на диск вместе с кадрами изображения и данными акселерометра для облегчения последующего анализа. Кроме того, можно использовать несколько зондов для стимуляции более чем в одном месте у одного и того же животного. Используя встроенный оптогенетический светодиодный драйвер NINscope, мы демонстрируем его использование путем стимуляции клеток Пуркинье мозжечка в трансгенных Pcp2 -Cre Jdhu x Ai32(RCL-ChR2(h234R)/EYFP) мышей при выполнении визуализации кальция из нейронов в моторной коре, трансдуцированных с помощью GCaMP6f (рис. 4A–B, центр объектива GRIN AP: +1,4, ML: 1,5) . Мы выбрали четыре местоположения в мозжечке для размещения синих (470 нм) оптогенетических светодиодных зондов: II голени ипси- и контралатеральнее места визуализации в правом полушарии, контралатеральной симплексной дольки и дольки VI в медиальном черве мозжечка. Штыри разъема светодиодного зонда (рис. 4C) позволяли переключать соединение драйвера светодиода минископа с одного места имплантации на другое (рис. 4D).
Штыри разъема светодиодного зонда (рис. 4C) позволяли переключать соединение драйвера светодиода минископа с одного места имплантации на другое (рис. 4D).
Мультисайтовая оптогенетическая стимуляция с помощью NINscope.
( A ) Мышей Pcp2 -Cre Jdhu скрещивали с мышами Ai32 для получения селективной экспрессии ChR2 (h234R) в клетках Пуркинье мозжечка. Нейроны моторной коры у этих мышей трансдуцировали с помощью GCaMP6f. ( B ) Экспериментальная конфигурация для объединения оптогенетической стимуляции мозжечка с визуализацией в коре головного мозга. ( C ) Мышь с опорной пластиной над корой и четырьмя светодиодными датчиками, установленными над мозжечком (1: II ипсиловая голень, 2: VI долька, 3: II контра, 4: Simplex долька). Контакты разъема использовались для подключения драйвера светодиодов NINscope к каждому из четырех датчиков. ( D ) Мышь с NINscope и подключением к одному из четырех мест стимуляции. ( E ) Оптогенетическая стимуляция клеток Пуркинье (50 мс, 22 мА, 2,3 мВт) вызывала четко различимое увеличение как среднего ответа (C_raw, среднее), так и количества коактивных клеток (пространственные следы, A) в моторной коре (все события, сумма событий). Повторная стимуляция вызывала в коре скачкообразную активность. ( F ) В этом примере большая часть клеток ответила (респондеры) на оптогенетическую стимуляцию контралатерального полушария мозжечка (Crus II). Слияние цветов показывает пространственную локализацию респондеров (красный) и нереспондеров (голубой). Реагирующие клетки отбирали по критерию, согласно которому постстимульный сигнал должен был превышать среднее значение +2σ исходного уровня до стимула. ( G ) Переходные процессы кальция (серый: отдельные переходные процессы, черный: среднее значение) и изменение суммированных событий (Δ событий), вызванных началом стимула в четырех разных местах на поверхности мозжечка, и соответствующие x (красный), y (зеленый) и z (синий) данные акселерометра канала.
( D ) Мышь с NINscope и подключением к одному из четырех мест стимуляции. ( E ) Оптогенетическая стимуляция клеток Пуркинье (50 мс, 22 мА, 2,3 мВт) вызывала четко различимое увеличение как среднего ответа (C_raw, среднее), так и количества коактивных клеток (пространственные следы, A) в моторной коре (все события, сумма событий). Повторная стимуляция вызывала в коре скачкообразную активность. ( F ) В этом примере большая часть клеток ответила (респондеры) на оптогенетическую стимуляцию контралатерального полушария мозжечка (Crus II). Слияние цветов показывает пространственную локализацию респондеров (красный) и нереспондеров (голубой). Реагирующие клетки отбирали по критерию, согласно которому постстимульный сигнал должен был превышать среднее значение +2σ исходного уровня до стимула. ( G ) Переходные процессы кальция (серый: отдельные переходные процессы, черный: среднее значение) и изменение суммированных событий (Δ событий), вызванных началом стимула в четырех разных местах на поверхности мозжечка, и соответствующие x (красный), y (зеленый) и z (синий) данные акселерометра канала. Стимуляция полушарий мозжечка выявляет латерализацию поведенческого ответа со стимуляцией слева или справа, вызывая движения головы влево и вправо соответственно. Вызванные поведенческие рефлексы, как правило, начинались до появления транзиторных кальциевых рефлексов в коре головного мозга.
Стимуляция полушарий мозжечка выявляет латерализацию поведенческого ответа со стимуляцией слева или справа, вызывая движения головы влево и вправо соответственно. Вызванные поведенческие рефлексы, как правило, начинались до появления транзиторных кальциевых рефлексов в коре головного мозга.
Стимуляция каждого из участков мозжечка (50 мс, ток 22 мА, мощность света 2,3 мВт) приводила к увеличению количества активированных нейронов в моторной коре правого полушария (рис. 4E–G, рис. 4 — видео 1). Повторяющаяся стимуляция мозжечка (0,3 Гц, 4-5 раз) может вызывать линейно-подобную активность, как видно при усреднении ответов по клеткам моторной коры (рис. 4Е). Данные акселерометра минископа зарегистрировали движение, которое произошло с задержкой после смещения стимула (ножка II ипси: 76 ± 23 мс; ножка II контра: 80 ± 30 мс; долька VI: 79).± 34 мс; simplex lobule: 73 ± 32 мс, среднее ± SD, N = 2 мыши) в соответствии с более ранними наблюдениями о том, что растормаживание клеток Пуркинье и связанное с ним обратное возбуждение нейронов в ядрах мозжечка могут вызывать замедленные двигательные рефлексы (Hoebeek et al. , 2010; Виттер и др., 2013). Значительное количество клеток соответствовало критерию увеличения содержания кальция, превышающего среднее значение +2σ исходного уровня до стимула (называемые ответчиками; см. также рисунок 4F). Во всех записях и местах стимуляции число респондеров составляло примерно половину всех сегментированных клеток моторной коры (49).0,9 ± 14,2%, среднее ± стандартное отклонение, 2227/4614 клеток, N = 2 мыши). Средние амплитуды кальциевых переходных процессов с Z-оценкой, измеренных в нейронах моторной коры, варьировались в зависимости от стимулируемой области (ножка II ипси: 2,1 ± 1,3 Z; ножка II контра: 2,9 ± 1,5 Z; долька VI: 2,9 ± 1,6 Z; простая долька: 2,0 ± 1,3 Z; критерий Крускала-Уоллиса p = 1,1239e-70 и апостериорные множественные сравнения с использованием S Шеффе выявили значительно отличающиеся средние ранги для стимуляции контралатеральной дольки VI и ножки II по сравнению с другими группами) (рис. 4G), но все показали сопоставимое время начала (ножка II ипси: 218 ± 120 мс; ножка II контра: 221 ± 118 мс; долька VI: 233 ± 123 мс; простая долька: 212 ± 130 мс, среднее значение ± стандартное отклонение; тест Крускала-Уоллиса, p =0,1, нс) относительно смещения стимула.
, 2010; Виттер и др., 2013). Значительное количество клеток соответствовало критерию увеличения содержания кальция, превышающего среднее значение +2σ исходного уровня до стимула (называемые ответчиками; см. также рисунок 4F). Во всех записях и местах стимуляции число респондеров составляло примерно половину всех сегментированных клеток моторной коры (49).0,9 ± 14,2%, среднее ± стандартное отклонение, 2227/4614 клеток, N = 2 мыши). Средние амплитуды кальциевых переходных процессов с Z-оценкой, измеренных в нейронах моторной коры, варьировались в зависимости от стимулируемой области (ножка II ипси: 2,1 ± 1,3 Z; ножка II контра: 2,9 ± 1,5 Z; долька VI: 2,9 ± 1,6 Z; простая долька: 2,0 ± 1,3 Z; критерий Крускала-Уоллиса p = 1,1239e-70 и апостериорные множественные сравнения с использованием S Шеффе выявили значительно отличающиеся средние ранги для стимуляции контралатеральной дольки VI и ножки II по сравнению с другими группами) (рис. 4G), но все показали сопоставимое время начала (ножка II ипси: 218 ± 120 мс; ножка II контра: 221 ± 118 мс; долька VI: 233 ± 123 мс; простая долька: 212 ± 130 мс, среднее значение ± стандартное отклонение; тест Крускала-Уоллиса, p =0,1, нс) относительно смещения стимула. Стимуляция клеток Пуркинье над обоими полушариями мозжечка вызывала противоположные движения головы, как определено по нашим данным акселерометра, с движениями влево при стимуляции над левой и движениями вправо при стимуляции над правой ножкой II (рис. 4G). Эти данные согласуются с выводами о том, что голени I и II у грызунов не только получают информацию, связанную с орофациальным поведением и поведением взмахов (Ju et al., 2019).; Romano et al., 2018), но также информацию о голове и шее (Quy et al., 2011; Huang et al., 2013), и что активация полушарий мозжечка у людей связана с движениями головы (Prudente et al., 2015). Латеральная стимуляция левой простой дольки вызывала более умеренные латеральные движения по сравнению со стимуляцией ножки II, тогда как при стимуляции медиальной доли червя VI, где движения вперед/назад были более выражены, они в основном отсутствовали. Поведенческие рефлексы, зарегистрированные с помощью акселерометра при стимуляции клеток Пуркинье, по-видимому, не коррелировали напрямую с активацией коры головного мозга, что указывает на наличие других мишеней ниже по течению, лежащих в основе этих рефлексов. Мы не наблюдали увеличения числа активированных нейронов в моторной коре при стимуляции с той же интенсивностью и продолжительностью у мышей дикого типа, лишенных ChR2, и такие стимуляции не были связаны с вызванными стимулом отклонениями акселерометра (рис. 4 — дополнение к рисунку 1). ).
Стимуляция клеток Пуркинье над обоими полушариями мозжечка вызывала противоположные движения головы, как определено по нашим данным акселерометра, с движениями влево при стимуляции над левой и движениями вправо при стимуляции над правой ножкой II (рис. 4G). Эти данные согласуются с выводами о том, что голени I и II у грызунов не только получают информацию, связанную с орофациальным поведением и поведением взмахов (Ju et al., 2019).; Romano et al., 2018), но также информацию о голове и шее (Quy et al., 2011; Huang et al., 2013), и что активация полушарий мозжечка у людей связана с движениями головы (Prudente et al., 2015). Латеральная стимуляция левой простой дольки вызывала более умеренные латеральные движения по сравнению со стимуляцией ножки II, тогда как при стимуляции медиальной доли червя VI, где движения вперед/назад были более выражены, они в основном отсутствовали. Поведенческие рефлексы, зарегистрированные с помощью акселерометра при стимуляции клеток Пуркинье, по-видимому, не коррелировали напрямую с активацией коры головного мозга, что указывает на наличие других мишеней ниже по течению, лежащих в основе этих рефлексов. Мы не наблюдали увеличения числа активированных нейронов в моторной коре при стимуляции с той же интенсивностью и продолжительностью у мышей дикого типа, лишенных ChR2, и такие стимуляции не были связаны с вызванными стимулом отклонениями акселерометра (рис. 4 — дополнение к рисунку 1). ).
Глубокая визуализация мозга и анализ поведения
Полосатое тело — это подкорковая структура, недоступная для визуализации без опускания релейной линзы GRIN на интересующее место (рис. 5А). Из-за повреждения тканей вдоль и ниже дорожки линзы требуется более длительное время восстановления, прежде чем можно будет начать визуализацию (Bocarsly et al., 2015). Значительное количество света теряется из-за комбинации двух линз GRIN вдоль оптического пути (реле GRIN и линза объектива GRIN), что делает эти эксперименты более сложными, чем получение изображений с дендритов клеток Пуркинье мозжечка или поверхностных слоев коры головного мозга. Чтобы подтвердить NINscope для изучения полосатого тела у непристегнутых животных и, в частности, доказать его эффективность для глубокой визуализации мозга, мы пересмотрели предыдущую работу, в которой была предложена роль дорсального полосатого тела (DS) в инициации контраверсивного движения и кодировании действия (Клаус и др. , 2017; Куи и др., 2013).
Глубокая визуализация мозга и анализ поведения с помощью NINscope.
( A ) Схема конфигурации NINscope, в которой объектив GRIN сочетается с релейной линзой GRIN (600 мкм) для получения изображения дорсального полосатого тела (DS) правого полушария. ( B ) Корональный срез, показывающий дорожку ретрансляционной линзы GRIN и нейроны в правом DS, экспрессирующие GCaMP6f (желтый). ( C ) Левые и правые повороты были количественно определены путем объединения видеонаблюдений или светодиодного отслеживания с анализом данных G-сенсора. Сверху показаны примеры мыши, совершающей левый и правый поворот на открытой арене, и путь, полученный от светодиода отслеживания NINscope, где прогрессия времени в течение одной секунды обозначена цветом. Ниже показаны пространственные компоненты (A) и переходные процессы (C_raw) 15 нейронов, извлеченные с помощью CNMF-E, время начала всех событий, извлеченных из C, а также x-канал акселерометра. Вертикальные столбики показывают, что животное сначала повернулось вправо (желтый цвет), а затем влево (синий цвет), при этом модуляция активности в правой ДС совпадала с контраверсивными движениями. ( D ) Данные акселерометра, показывающие среднее левое и правое ускорение канала x вокруг начала движения (среднее ± SEM). Пунктирные серые линии представляют противоположное ускорение. ( E ) Средние переходные реакции кальция за одну секунду до и после начала движения (N = 5 мышей) обнаруживают четкую модуляцию активности во время левых поворотов (среднее ± SEM) при визуализации правого DS. ( F ) Количественная оценка переходных реакций кальция до и после начала движения для поворотов влево и вправо соответственно. Правый DS показал значительное транзиторное увеличение кальция только для левых поворотов (p<0,05).
Используя вирусный вектор с промотором синапсина человека, мы трансдуцировали все полосатые нейроны с GCaMP6s или GCaMP6f в каудальной, дорсальной части правого полосатого тела (рис. 5B). Непосредственно после трансдукции вируса над интересующей областью имплантировали релейную линзу GRIN диаметром 600 мкм. NINscope был установлен на опорной плите, к которой был приклеен объектив GRIN. Вся сборка была опущена чуть выше релейной линзы GRIN, чтобы сфокусировать клетки. В DS мы извлекли сигналы от 84 клеток (62 ± 16,70, среднее значение ± стандартное отклонение, диапазон 38–84; рис. 5C), которые, основываясь на переходных процессах кальция, имели частоту ~ 1 Гц (1,106 ± 0,9).3 Гц, среднее ± SD, n = 308 клеток, N = 5 мышей) (рис. 5D). Несмотря на использование относительно низкой мощности света (~300 мкВт после объектива и перед релейной линзой GRIN), мы получили хорошее соотношение сигнал/шум. Используя как светодиод слежения NINscope, так и данные акселерометра, мы обнаружили эпохи, когда мыши совершали спонтанные повороты туловища и головы (рис. 5E, рис. 5 — видео 1). Такие повороты были связаны с отклонениями вверх или вниз в x-канале нашего акселерометра, отражающими повороты влево или вправо соответственно (рис. 5C, D). Во время левых поворотов (контралатерально к месту визуализации в DS) большинство нейронов в правом DS (85%, n = 308 клеток, N = 5 мышей) имели пиковые ответы после начала действия, а 20% продемонстрировали значительно повышенные ответы для продолжительность выполнения действия (парный базовый уровень t-теста по сравнению с активностью выполнения действия, p <0,05) (рис. 5G, H). Самые большие из них начались после начала движения, что указывает на преобладающую связь с выполнением действия, а не с подготовкой (латентность начала, 310 ± 45 мс, среднее значение ± стандартное отклонение). Ни одна из клеток, которые мы записали, не реагировала во время движений вперед-назад или движений ипсилатерально к месту записи (рис. 5G, H), что подтверждает латерализацию сигналов движения в полосатом теле. Когда сигналы усреднялись за одну секунду до и после начала движения (рис. 5I, J), повторные измерения ANOVA выявили значительный основной эффект (F (1,18, 4,7) =16,48, р=0,01) выполнения действия. Апостериорная поправка Бонферрони для множественных сравнений продемонстрировала, что эффект возникал исключительно в эпохи контралатерального (t = 9,45, df = 4, p = 0,001), но не ипсилатерального начала движения (t = 3,07, df = 4, нс).
Комбинация глубокой визуализации мозга с оптогенетической стимуляцией
Чтобы показать полезность NINscope для комбинирования оптогенетической стимуляции с глубокой визуализацией мозга, мы изучили влияние корковых входов на активность нейронов при ДС у животных без ограничений. Опсин ChrimsonR был преобразован либо в орбитофронтальную кору (OFC), либо во вторичную моторную кору (M2), а светодиод (с пиковым излучением 645 нм) помещали над корой (рис. 6А). OFC и M2 могут по-разному регулировать активность нейронов в определенных областях DS in vitro (Corbit et al., 2019).), при этом влияние OFC на DS сильнее, чем у M2. Мы стремились подтвердить эти выводы in vivo. Сначала мы оценили прямые терминальные поля OFC и M2 до DS и нанесли их на репрезентативное изображение атласа головного мозга (рис. 6B). Вход M2 в DS был более рассеянным, чем проекции OFC, что согласуется с предыдущими выводами (Hintiryan et al., 2016; Corbit et al., 2019; Hunnicutt et al., 2016). Во время сеансов визуализации DS, трансдуцированных с помощью GCaMP6f (используемая мощность света: 170 мкВт после релейной линзы GRIN, рисунок 2 — дополнение к рисунку 1), животные могли свободно исследовать арену открытого поля, в то время как OFC или M2 были оптогенетически стимулированы (10 с). , 20 Гц, длительность импульса 5 мс, светодиод мощностью 3,4 мВт). Во время такой стимуляции OFC и M2 ответы нейронов DS можно было разделить на три различных типа, в том числе те, которые были снижены, повышены или не изменились по сравнению с исходной активностью (парный t-критерий базовой линии по сравнению с активностью, вызванной стимулом, p <0,05, N = 4 мыши) (рис. 6С). Частота срабатывания кальциевых переходных процессов во время стимуляции различалась между этими кластерами (уменьшилась: 0,23 ± 0,25 Гц, среднее ± SD, n = 33 клетки, N = 4 мыши, не изменилась: 0,77 ± 0,68 Гц, среднее ± SD, n = 199 клеток, N = 4 мыши, увеличение: 1,63 ± 1,23 Гц, среднее ± SD, n = 22 клетки, N = 4 мыши).
Сочетание глубокой визуализации мозга с оптогенетической стимуляцией с использованием NINscope.
( A ) Схема, показывающая размещение NINscope с объективом GRIN и релейной линзой GRIN для записи с дорсального полосатого тела (DS), а также расположение оптогенетического светодиодного датчика, управляемого встроенным светодиодным драйвером. Вирусные векторы вводили либо в орбитофронтальную кору (OFC), либо во вторичную моторную кору (M2) для преобразования нейронов с помощью ChrimsonR, либо в DS для преобразования нейронов с помощью GCaMP6f для визуализации кальция. ( B ) Терминальные поля OFC и M2, нанесенные на шаблон Allen Brain Atlas, показывают их перекрытие в DS под линзой реле GRIN. ( C ) Ответы для каждого нейрона усреднялись по 10 испытаниям для 254 клеток у четырех мышей. Различные типы ответов обнаруживаются при СД, когда либо OFC, либо M2 оптогенетически стимулируются. Нейроны демонстрировали снижение активности (синий кластер), отсутствие видимых изменений (оранжевый кластер) или повышенный ответ (красный кластер). ( D ) Переходные процессы кальция с Z-оценкой во время стимуляции OFC (слева, N = 2 мыши) и M2 (справа, N = 2 мыши) (импульс 10 с, 20 Гц). Для каждого нейрона усреднялось 10 проб. ( E ) Ответы, усредненные по всем нейронам DS для всех 10 испытаний, выявили сходные типы модуляции при стимуляции OFC и M2 (среднее ± SEM). Круглые вставки обозначают долю клеток, которые показали подавление, отсутствие изменений или повышенный ответ.
Стимуляцию повторяли в течение 10 испытаний, а ответы усредняли по испытаниям (рис. 6D). Наблюдалась модуляция активности субпопуляций нейронов ДС при стимуляции ОФК: 20% нейронов (26/133) демонстрировали значительное снижение активности, 69% не показали изменений (93/133), а 11% (15/133) увеличились. Стимуляция М2 оказывала сопоставимое, но более слабое влияние на активность субпопуляций нейронов ДС: 7% нейронов (7/107) показали снижение, 86% (93/107) не показали изменений и 7% (7/107) клеток. показали повышение активности. Хотя доля реагирующих нейронов DS (т. е. тех, которые показывают уменьшение или увеличение) во время стимуляции коры значительно различалась между OFC (31%) и M2 (14%) (хи-квадрат (1) = 13,85, p <0,001), среднее значение ответ для каждой категории нейронов DS во время стимуляции был сопоставим (рис. 6E). Пониженные нейроны демонстрировали постепенное снижение активности во время стимуляции, тогда как нейроны, реагирующие на стимуляцию повышенной активностью, демонстрировали прогрессивное увеличение, пока стимуляция была предоставлена, что позволяет предположить, что входная модуляция активности в нейронах DS масштабировалась с продолжительностью стимуляции.
Обсуждение
NINscope
Мы продемонстрировали применимость NINscope для выполнения записей в двух точках у мышей и комбинирования поверхностной и глубокой визуализации мозга с оптогенетической стимуляцией при анализе движений с помощью встроенного акселерометра.
Наш минископ расширяет функциональные возможности, сохраняя при этом небольшие размеры, что делает его ценным вкладом в расширяющийся набор инструментов для минископов с открытым исходным кодом. Программное обеспечение для сбора данных и управления NINscope не зависит от платформы и может использоваться во всех основных операционных системах с использованием различных аппаратных конфигураций, включая ноутбуки, доступные большинству пользователей.
Во время прототипирования NINscope необходимо было сделать ряд вариантов дизайна. Мы напечатали корпус микроскопа на 3D-принтере, чтобы конструкция была легкой, но воздержались от использования линз с электросмачиванием, которые сделали бы фокусировку практичной, но минископ слишком тяжелый и громоздкий для записи в двух местах. Включение светодиодного драйвера высокого разрешения для оптогенетической стимуляции вместе с использованием специального светодиодного зонда предоставило возможность напрямую управлять проекциями в месте их происхождения, а не их окончаниями в поле зрения изображения, что является особенностью. который раньше не интегрировался в минископ. Кроме того, мы представляем первый минископ со встроенным акселерометром, который позволяет анализировать поведение во время визуализации. Наконец, мы включили второй драйвер светодиода возбуждения на интерфейсную плату, который позволит использовать двухцветное возбуждение флуоресценции в будущих конструкциях.
Учитывая, что наши строительные планы имеют открытый исходный код, основные функциональные возможности нашей области могут быть расширены в зависимости от конкретных исследовательских вопросов, которые необходимо решить, что в конечном итоге диктует ограничения по размеру, весу и функциональности.
Визуализация двух областей и многоцентровая оптогенетика
Насколько нам известно, мы представляем первые одновременные записи клеточного разрешения мозжечка и коры головного мозга у мышей без ограничений. Мы обнаружили, что синхронные паттерны активности в этих регионах являются общими для нескольких поведенческих состояний и коррелируют с ускорением движения. Одновременная визуализация мозжечка и коры головного мозга может помочь выяснить, как активность в этих двух областях коррелирует на этапах планирования, обучения и выполнения движения (Guo et al. , 2015; Kawai et al., 2015; Sauerbrei et al., 2020). ; Гао и др., 2018). Используя NINscope, мы также выполнили грубую оценку функциональной связи между мозжечком и корой головного мозга путем оптогенетической стимуляции клеток Пуркинье в нескольких местах мозжечка у одного и того же животного. Такие стимулы вызывали рефлекторные движения и последующую активацию нейронов коры головного мозга с задержками, подобными тем, о которых мы сообщали ранее (Witter et al., 2013). Таким образом, NINscope может облегчить исследования, требующие оптогенетической стимуляции нескольких отдельных участков с считыванием активности в одном или двух удаленных регионах. Двойная визуализация с использованием NINscope не ограничивается записями из мозжечка и коры или подкорковых структур, таких как ростральное полосатое тело. Мы показываем, что возможны и другие конфигурации, включая двухполушарные записи коры, гиппокампа и мозжечка. Такие типы записей могут показать, в какой степени нейронная активность латерализована во время поведения и обучения, а также могут отслеживать стабильность мультирегиональных нейронных сигналов с течением времени (Gonzalez et al. , 2019).). Благодаря интегрированному контролю над оптогенетикой и считыванию сигналов ускорения NINscope позволяет в будущем контролировать активность с обратной связью, запускаемую либо поведенческим состоянием, либо уровнями активности, измеряемыми в одной или двух областях.
Глубокая визуализация мозга
КМОП-сенсор PYTHON480, встроенный в NINscope, позволяет регистрировать сигналы от дорсального полосатого тела (DS) с хорошим соотношением сигнал/шум при мощности в несколько сотен мкВт перед релейной линзой GRIN, что подтверждает применимость нашего минископа для получения изображений глубоких структур мозга . Анализируя данные, собранные с помощью видеокамеры и встроенного акселерометра, в сочетании с изображениями, полученными с помощью DS, мы смогли подтвердить предыдущие выводы, согласно которым полосатое тело было идентифицировано как структура, представляющая пространство действия (Klaus et al., 2017; Cui et al., 2013). ; Барбера и др., 2016; Текуапетла и др. , 2014).
Используя NINscope для регистрации DS в сочетании с оптогенетической стимуляцией OFC или M2 в месте их происхождения, мы показываем, что эти два входных пути по-разному влияют на активность DS, как показано ранее на срезах мозга (Corbit et al., 2019). ). Независимо от знака модуляции, мы обнаружили ответы, масштаб которых зависел от продолжительности стимула, что еще раз подчеркивает эффективность NINscope для решения вопросов, требующих комбинированного подхода к стимуляции и визуализации.
В совокупности NINscope представляет собой универсальный минископ с открытым исходным кодом, который может соответствовать нише пользователей, которым требуются записи в двух областях у животных без привязи, совместимость с ОС, оптогенетическое манипулирование областями на расстоянии от места изображения, поведенческий анализ с использованием акселерометра или сочетание всего вышеперечисленного.
Материалы и методы
| Тип реагента (виды) или ресурс | Обозначение | Источник или Ссылка | Идентификаторы | Дополнительная Информация | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Генетический реагент ( M. Musculus ) | B6.CG-TG (PCP2-CRE) 35555JJDHUID: | . JAX:010536, RRID:IMSR_JAX:010536 | https://www.jax.org/strain/010536 | |||||||
| Genetic reagent ( M. musculus ) | Ai32 | PMID: 22446880 | IMSR Cat # JAX:012569, RRID:IMSR_JAX:012569 | https://www.jax.org/strain/012569 | ||||||
| Рекомбинантный реагент ДНК | AAV1.CAG.FLEX.GCAMP6F.WPRE.SV40 | 707.LED | .WPERID .WPERID .WPERID .WPRID. .addgene.org/100835/||||||||
| Рекомбинантный реагент ДНК | AAV1.CMV.PI.CRE.RBG | ADDGENE | RRID: ADDGENI_105530 | HTTTPS: ADDGENI_105530 | 7070707070707. | Реактив рекомбинантной ДНК | AAV. Syn.GCaMP6f.WPRE.SV40 | Addgene | RRID:Addgene_100837 | https://www.addgene.org/100837/ |
| Recombinant DNA reagent | AAV.Syn.GCaMP6f.WPRE.SV40 | Addgene | RRID:Addgene_100843 | https://www.addgene.org/100843/ | ||||||
| Other | NINscope | This paper | RRID:SCR_017628 | https://github.com/ninscope | ||||||
| Software, algorithm | FIJI | PMID: 22743772 | RRID: SCR_002285 | https://imagej.net/Fiji/Downloads | ||||||
| Software, algorithm | MATLAB | MathWorks | RRID: SCR_001622 | https://uk. mathworks .com/products/matlab.html | ||||||
| Программное обеспечение, алгоритм | Bonsai | PMID: 25 1 | RRID: SCR_017218 | https:/bonsai-rx.117777777777777777777777777777.rxai-rx.milsai-rx.milsai-rx.milsai-rx.milsai-rx.mil.0070 R | R Project | RRID: SCR_001905 | http://www.r-project.org/ | |||
| , Algorithm | Python | Python | Python | Python | Python | . .python.org/ | ||||
| Программное обеспечение, алгоритм | SPSS | IBM | RRID:SCR_002865 | , алгоритм | Призма | Графический планшет 9 | Пользовательские скрипты Python данные времени |
Конструкция печатной платы
Запросить подробный протокол В NINscope используются две печатные платы (PCB): датчик изображения CMOS и интерфейсная плата размером 10 на 10 мм (толщина 0,6 мм, стандарт HDI), разработанные с использованием кросс-платформенного пакета автоматизации электронного проектирования с открытым исходным кодом KiCad (http: //kicad-pcb. org). Плата интерфейса включает в себя DS9Серийный преобразователь 0UR913A (Cypress Semiconductor) для подключения через FPD-link III к плате сбора данных. Плата интерфейса размещается поверх платы датчика, и две платы соединяются проводами, припаянными к зубчатым отверстиям на краях платы. Печатная плата датчика содержит регуляторы напряжения, регулятор мощности и генератор, необходимые для инициализации и работы датчика изображения. Регуляторы напряжения представляют собой регуляторы LDO со сверхнизким уровнем шума (NCP163, On Semiconductor) и предназначены для использования в камерах. Датчик изображения нуждается в трех источниках питания, которые предоставляются последовательно, что мы достигаем с помощью секвенсора питания LM3880 (Texas Instruments). Для работы датчика изображения используется маломощный КМОП-тактовый генератор с частотой 66,6667 МГц (Kyocera Electronics). В дополнение к сериализатору интерфейсная печатная плата имеет датчик IMU (LSM6DSLTR, STMicroelectronics), 2 драйвера светодиодов (один светодиод для оптогенетических стимулов: LM36011; двойной светодиод для одного светодиода возбуждения флуоресценции: LM3643, Texas Instruments) и I2C I/O. расширитель (FXL6408UMX, ON Semiconductor). Расширитель ввода/вывода управляет сигналами, которые не являются критичными по времени для датчика изображения и светодиодного индикатора слежения/индикатора в верхней части интерфейсной печатной платы. Драйвер светодиода и акселерометр более подробно описаны ниже.
КМОП-датчик изображения
Запросить подробный протокол Печатная плата датчика включает датчик изображения PYTHON480 (ON Semiconductor) CMOS SVGA. Датчик изображения PYTHON480 CMOS является гибким и имеет множество возможных конфигураций. Задача состояла в том, чтобы настроить его под наши требования. Как только мы выяснили, как запустить датчик в режиме CMOS вместо режима LVDS, мы смогли подавать сигналы данных на общий программируемый интерфейс (GPIF) USB-контроллера Cypress для передачи данных изображения. Для оптимизации энергопотребления и ограничения перегрева мы решили отключить фазовую автоподстройку частоты (PLL), чтобы снизить потребление тока на 30%. Без PLL выход параллельного тактового сигнала CMOS в 4 раза ниже, что ограничивает количество пиксельных данных, которые могут быть считаны для данной частоты кадров. Мы установили скорость сбора данных в текущем NINscope на 30 кадров в секунду и размер кадра на 752 × 480 пикселей, чтобы обеспечить возможность считывания как данных пикселей, так и трех каналов акселерометра из IMU. Улучшенное рассеивание тепла в будущих итерациях NINscope сделает доступными данные с более высокой частотой кадров до 120 Гц и позволит дополнительно считывать данные гироскопа с нашего IMU.
Оборудование для сбора данных
Запросить подробный протокол Проект UCLA Miniscope имеет крупнейшее сообщество минископов с открытым исходным кодом с пользовательской базой из сотен пользователей. Мы использовали существующее оборудование для сбора данных (версия DAQ 3.2) из проекта UCLA Miniscope (http://www.miniscope.org) и представили небольшие модификации, состоящие из 256-килобайтной x 8-битной EEPROM I2C (STMicroelectronics) для хранения данных. наша более крупная модифицированная прошивка и соединения на плате DAQ между GPIO2 и TP4 и сигналами SPI на GPO0-GPO3. Датчик PYTHON480 CMOS подключается через FPD-Link III и коаксиальный кабель к USB-контроллеру Cypress на плате сбора данных. Датчик изображения имеет интерфейс SPI вместо интерфейса I2C для настройки датчика. Выходы общего назначения (GPO) используются, поскольку канал FPD в UCLA DAQ поддерживает шину I2C, а не шину SPI. Для этого требуется маршрутизация сигналов от MOSI, SCK и SS к GPO0, GPO1 и GPO3, позволяющая полностью управлять датчиком изображения из прошивки USB-контроллера. Прошивка NINscope основана на примере прошивки, взятой из примечания к применению Cypress AN75779.(дата изменения: 30.10.2017). КМОП-датчик изображения, зарегистрированный как USB-устройство обработки изображений, расширяется для работы в качестве составного USB-устройства, что позволяет добавить виртуальный последовательный порт. Виртуальный последовательный порт используется для связи и настройки КМОП-сенсора и драйверов светодиодов (например, для установки коэффициента усиления сенсора изображения, яркости, уровней черного, настроек светодиодов, параметров оптогенетического стимула), а также для регистрации данных акселерометра.
Маломощный (0,65 мА в высокопроизводительном режиме) трехмерный акселерометр и гироскоп инерциальный модуль iNemo (LSM6DSLTR, STMicroelectronics), набор драйверов светодиодов (один светодиод для оптогенетических стимулов: LM36011; двойной светодиод для одного светодиода возбуждения флуоресценции: LM3643 , Texas Instruments) и расширитель ввода-вывода (FXL6408, ON Semiconductor) управляются по шине I2C. Оставшийся GPO используется для управления генерацией импульсов и точной синхронизацией оптогенетических стимулов путем введения проводного соединения между GPO2 и TP4 в UCLA DAQ (рис. 1E).
Акселерометр IMU имеет диапазон +/- 2 g, где каждый бит соответствует 0,061 мг. Установлена частота дискретизации 104 Гц, которая заполняет буфер FIFO данными x, y, z, которые считываются в конце каждого кадра изображения и передаются по виртуальному последовательному соединению с компьютером.
Светодиодные драйверы и светодиоды
Запросить подробный протокол Для управления генерацией оптогенетических импульсов в светодиодном зонде с длиной волны 470 нм (LED 150040BS73240, размер корпуса 402, Wurth Electronics) соответственно ) или светодиодный зонд 630 нм (APHHS1005SURCK, Kingbright) и светодиод возбуждения флуоресценции 470 нм (светодиод возбуждения LXML-PB02 470 нм, Lumileds). Оптогенетический светодиод можно регулировать с шагом 11,725 мА, а светодиод возбуждения — с шагом 1,4 мА. Максимальный ток каждого светодиода зависит от общего потребления тока камерой и типа/длины используемого кабеля. Светодиод с длиной волны 621 нм (SML-P11UTT86R, Rohm) был встроен в верхнюю часть платы интерфейса, чтобы обеспечить отслеживание положения животного с помощью камеры и уведомление о подключении минископа.
Корпус микроскопа
Запросить подробный протокол Прототипы корпуса микроскопа были разработаны с использованием Inventor (AutoDesk), а первоначальные прототипы были напечатаны на принтере Micro Plus Advantage (EnvisionTec) с использованием смолы RCP30 M, позволяющей печатать мелкие детали. Самая тонкая функциональная стенка окончательного прототипа имела толщину 500 мкм. Принтер Form 2 (Formlabs) впоследствии использовался для печати корпуса микроскопа черной смолой (РС-Ф2-ГПБК-04). Микроскоп состоит из трех частей: верхней части для размещения печатных плат, плосковыпуклой линзы и эмиссионного фильтра, нижней части для размещения оптики, включая кристалл светодиода, полушаровидную линзу, фильтр возбуждения и дихроичное зеркало, и сдвижной крышки для крепления. светодиод и защитить оптические фильтры. Нижняя часть микроскопа имеет небольшой выступ, который фиксируется в выемке специальной металлической опорной плиты (рис. 1 — дополнение к рисунку 2) и фиксируется установочным винтом для сведения к минимуму перемещения микроскопа. После печати и очистки корпуса изопропиловым спиртом и пескоструйной обработки корпус внутри и снаружи, напечатанный на принтере EnvisionTec, был обработан аэрографом (Infinity CR Plus, Harder and Steenbeck, Германия) тонким слоем черной краски (h22 Flat Black, Гунзе, Япония).
Оптика
Запросить подробный протокол Оптические конструкции были смоделированы с помощью Zemax Optic Studio (рис. 1 — дополнение к рисунку 1). Мы использовали специальные фильтры возбуждения (ET470/40 × 3,5 × 3,5 × 0,5 мм, Chroma), дихроичные (T495lpxr 3,5 × 5 × 0,5 мм, Chroma) и эмиссионные фильтры (ET525/50 m, 4 × 4 × 1 мм, Chroma). ), полусферическую линзу N-BK7 (диаметр 3 мм, 47–269, Edmund Optics) и плосковыпуклую линзу (диаметр 4,0 мм, фокусное расстояние 10,0 мм, 45–429, Edmund Optics) для фокусировки на CMOS-изображение. датчик. Эмиссионный фильтр был прикреплен к этой линзе с помощью оптического клея (NOA81, Norland Products). Объектив ГРИН (ЧА 0,55, 64–519, Edmund Optics) имплантировали для поверхностной визуализации или вклеивали в базовую пластину для глубокой визуализации мозга, а минископ устанавливали на базовую пластину, которая была приклеена к черепу.
Кабели
Запросить подробный протокол Для подключения осциллографа NIN к блоку сбора данных тонкий (0,101 мм) коаксиальный провод (38awg A9438W-10-nd, Alpha Wire) длиной 50 см был присоединен к минископу одним концом и подключен через набор разъемов (ED -ND и ED8250-ND, Digi-Key) к более толстому (1,17 мм FEP мм) коаксиальному проводу (VMTX Mini,Pro Power) длиной 2 м на другом конце. Провод длиной 2 м был припаян к 150-мм коаксиальному кабелю RG174 с прямой вилкой SMA (2096227, LPRS), который можно было напрямую подключить к плате сбора данных. Драйвер светодиода возбуждения был подключен к светодиоду с помощью ультратонкого провода длиной 22 мм (UT3607, Habia). Такой же провод длиной 25 мм был подключен к оптогенетическому стимулирующему светодиодному драйверу на интерфейсной плате. На одном конце был прикреплен разъем (851-43-050-10-001000, Mill-Max или ED-ND, Digi-Key) для подключения к оптогенетическому зонду.
Программное обеспечение для сбора данных и управления
Запросить подробный протокол Программное обеспечение для сбора данных и управления было разработано на кроссплатформенном языке Processing (https://processing.org/). Программное обеспечение поддерживает сбор данных с двух минископов и одной веб-камеры USB. NINscope управляется через последовательный коммуникационный порт с использованием последовательной библиотеки Processing, которая поддерживает пользовательские микроконтроллерные устройства. Модуль захвата в видеобиблиотеке для Processing версии 2.0 был изменен путем преобразования YUV в необработанный формат, чтобы использовать преимущества полной 8-битной шкалы. Чтобы различать камеры и NINscopes одного типа, к строковому списку всех подключенных устройств добавляется префикс. G4P, библиотека обработки (http://www.lagers.org.uk/g4p/), была использована для создания пользовательского интерфейса программного обеспечения, обеспечивающего доступ к наиболее распространенным элементам управления для изменения настроек микроскопа и обновления таких параметров, как интенсивность света, усиление , уровень черного, продолжительность оптогенетического стимула или регистрация G-сенсора. В скетче обработки функция события захвата передает каждое захваченное изображение в кольцевой буфер для быстрой обработки изображения и во избежание длительного времени доступа к жесткому диску (HD), которое может привести к пропуску кадров. Поток используется для сохранения всех изображений из кольцевого буфера в HD. Размер кольцевого буфера можно изменить в скетче, но он установлен на 60. Последние 4 пикселя кадра зарезервированы для счетчика кадров для отслеживания пропущенных кадров. Каждый кадр сохраняется как изображение в формате TIFF в градациях серого с наименьшим возможным заголовком для уменьшения размера файла.
Мыши
Мы использовали как самцов, так и самок мышей C57BL/6 (диапазон веса: 22–28 г, возраст:> 6 недель). В экспериментах с оптогенетической стимуляцией клеток Пуркинье мы использовали двух трансгенных мышей-самцов от скрещивания линии Pcp2 -Cre Jdhu (JAX # 010536) и линии мышей Ai32 (JAX # 012569) на фоне C57BL/6. Мышей размещали группами до пяти мышей до имплантации линз GRIN, после чего их размещали поодиночке. Мышей содержали на цикле свет:темнота 12:12 с включенным светом с 7:00 до 19:00.:00. Они получили свободный доступ к воде и пище (Teklad, Envigo). Был предоставлен подстилочный материал для строительства гнезда. Все проведенные эксперименты были лицензированы компетентным органом Нидерландов и одобрены местным органом по защите животных в соответствии с Европейскими рекомендациями по уходу и использованию лабораторных животных Директивой 2010/63/ЕС.
Имплантаты линз GRIN и инъекции вируса
Запросить подробный протокол Перед операцией мышей анестезировали 3% изофлураном перед переносом в стереотаксический аппарат, после чего анестезию поддерживали 1,5% изофлураном (скорость потока: 0,3 мл/мин O 2 ). Для визуализации головного мозга и коры мозжечка на поверхность головного мозга помещали объектив GRIN (диаметр 1,8 мм, шаг 0,25, 64–519, Edmund Optics). После удаления волос и дезинфекции кожи раствором йода (5%) и спиртом (70%) на коже делали небольшой разрез. Затем на обнаженный череп наносили лидокаин (100 мг/мл, Astra Zeneca, Великобритания) и удаляли надкостницу. Были определены центральные координаты для размещения линзы GRIN, и небольшая чернильная точка была помещена в правильном месте относительно брегмы (мозжечковая мозжечковая мозжечковая мозжечковая долька, AP: -5,8 мм ML: 2,2 мм; долька VI, AP: -7,4 мм, ML: 0,0 мм; кора, AP: 1,4 мм, ML: 1,5 мм). Координаты масштабировались относительно среднего расстояния между брегмой и лямбдой (4,21 мм), как указано в атласе мозга мышей Paxinos. Перед сверлением кости мышам вводили внутрибрюшинно. Инъекции 15% D-маннитола в физиологическом растворе (0,55 мл/25 г) для облегчения диффузии вирусных частиц после инъекции вируса. Затем была просверлена круговая трепанация черепа диаметром 2 мм по центру вокруг отмеченного места. В промежутках между сверлением череп увлажняли стерильным физиологическим раствором. Затем удаляли лоскут черепа и твердую мозговую оболочку и вирус (мозжечок: AAV1.CAG.FLEX.GCaMP6f/AAV1.CMV.PI.Cre.rBG смешивали 1:1 и разводили в физиологическом растворе 1:3; кору головного мозга: AAV1.Syn.GCaMP6f. WPRE.SV40, разбавленный в физиологическом растворе 1:3, UPenn Vector Core) вводили в четыре места. В каждое место вводили 25 нл вируса один раз на глубине 350, два раза на 300 и один раз на глубину 250 мкм со скоростью 25 нл/мин с помощью автонанолитрового инжектора Nanoject II (Drummond Scientific Company, США). Место краниотомии покрывали гелевой пеной (Pfizer, США), смоченной в стерильном физиологическом растворе (0,9% NaCl, B. Braun Medical Inc, США). Линзу GRIN опускали с помощью вакуумного держателя, помещенного в стереотаксический аппарат, до соприкосновения поверхности линзы с мозгом, а затем опускали еще на 50 мкм. Края краниотомии герметизировали Kwik-Sil (WPI, США). Затем вокруг линзы был нанесен стоматологический цемент (Super-Bond C и B, Sun Medical, Япония), чтобы зафиксировать ее на месте. Для покрытия и защиты линзы использовали Kwik-Cast (WPI, США). В конце операции животным вводили подкожно. введение 5 мг/кг метакама.
Для доступа к области глубокого мозга в дорсальном полосатом теле были имплантированы релейные линзы GRIN диаметром 0,6 мм (длина: 7,3 мм, числовая апертура: 0,45, Inscopix, CA) (AP: 0,2, ML: 1,5, DV: -3,1 ). Чтобы облегчить имплантацию и уменьшить повреждение ткани выше дорсального полосатого тела, с помощью иглы 25G создали дорожку. Используя моторизованную стереотаксическую руку, иглу медленно опускали в течение 10 минут, оставляли на месте на 10 минут, а затем удаляли с той же скоростью с постоянной скоростью 0,31 мм/мин. Вирус доставляли путем опускания шприца Гамильтона в течение 4 минут для инъекции 500 нл AAVdj.hSyn.GCaMP6s (N = 2 животных, рис. 5, N = 4 мышей, рис. 6) или AAV1.hSyn.GCaMP6f (N = 3 мышей, рисунок 5) в дорсальное полосатое тело со скоростью 100 нл/мин, после чего шприц оставляли на месте для увеличения диффузии вируса (5 мин) и затем втягивали (5 мин). Затем релейную линзу GRIN опускали (0,10 мм/мин) в дорсальный стриатум (DV: -3,1). Зазор между релейной линзой GRIN и черепом был покрыт цианоакрилатным клеем, а линза была закреплена на черепе с помощью стоматологического цемента. Линза реле GRIN была покрыта и защищена с помощью Kwik-Cast (World Precision Instruments, США). Объектив GRIN 1,8 мм (64–519, Edmund Optics, Великобритания) прикрепляли к основанию минископа с помощью цианоакрилатного клея, а затем минископ устанавливали на основание. Голова мышей была ограничена с помощью специально изготовленного ремня для бега, чтобы обеспечить передвижение, прицел с установленной опорной пластиной и объективом GRIN опускался над релейной линзой GRIN до тех пор, пока клетки в поле зрения не становились видимыми, опорная пластина крепилась к черепу с помощью стоматологический цемент (покрытый черным лаком для ногтей) и минископ удалены.
Светодиодные датчики
Запросить подробный протокол В экспериментах по визуализации дорсального полосатого тела AAV5. hSyn.ChrimsonR.tdTomato вводили либо в орбитофронтальную кору (AP: 2,8, ML: 1, DV: -2,2), либо во вторичную моторную кору (AP: 2,3, ML: 0,7 DV: −1,2). Светодиодный зонд, покрытый биосовместимой эпоксидной смолой, предназначенный для подключения к NINscope с красным светодиодом (пиковое излучение 645 нм, KingBright Inc, США), помещали на поверхность мозга, а светодиод закрепляли на черепе с помощью стоматологического цемента.
Для стимуляции мозжечка над интересующей областью мозжечка была просверлена небольшая краниотомия диаметром 1 мм (долька Simplex, AP: -5,8 мм, ML: 2,2 мм; долька VI, AP: -7,4 мм, ML: 0,0 мм; Crus II , AP: -7,3 мм, ML: + и -2,7 мм), следуя аналогичным стерильным процедурам, описанным для имплантата объектива GRIN. Светодиод, покрытый слоем биосовместимой эпоксидной смолы, был вставлен с правильной ориентацией в краниотомию, которая затем была герметизирована слоем Kwik-Sil (WPI, США), а затем дополнительным слоем Kwik-Cast. Оставшуюся проволоку датчика и соединитель датчика впоследствии прикрепили к черепу стоматологическим цементом (Super-Bond C и B, Sun Medical, Япония).
Визуализация
Запросить подробный протоколЗаписи мозжечка и коры головного мозга были выполнены в домашней клетке для мышей, в то время как записи, полученные из дорсального стриатума, были получены в специальной открытой арене (30 на 30 на 45 см). Минископы присоединяли либо после кратковременной анестезии (индукция 3% изофлюрана, время восстановления после анестезии 30 минут), чтобы можно было подключить разъем оптогенетического светодиодного драйвера к светодиодному зонду, установить один или два эндоскопа и отрегулировать фокус, либо — когда с использованием модифицированной опорной плиты или специальной перекладины для головы — в состоянии с фиксированной головой, когда животные могут бегать по беговой дорожке.
Гистология
Запросить подробный протокол Мышей транскардиально перфузировали 4% PFA в 1x PBS (10 мМ PO43-, 137 мМ NaCl, 2,7 мМ KCl). После фиксации мозг удаляли и оставляли на ночь в 10% сахарозе в PBS, после чего их заключали в желатин и оставляли на ночь в 30% сахарозе в PBS. Срезы головного мозга (50 мкм) вырезали в криостате (LEICA CM3050 S), окрашивали DAPI и затем монтировали (Dako Fluorescence Mounting Medium, S3023). Конфокальные изображения были собраны на конфокальном микроскопе SP8 (Leica, Германия), а изображения всего мозга были получены путем объединения нескольких (1024 × 1024 пикселей) снимков в окончательное изображение.
Инъекции бусин
Запросить подробный протокол Для оценки артефактов движения с помощью двух NINscope мы сделали круговую краниотомию диаметром 2 мм, затем ввели 51 нл флуоресцентных шариков диаметром 6 мкм (InSpeck Green (505/515) Набор для калибровки интенсивности изображения для микроскопа, 6 мкм, компонент G, ThermoFisher Scientific, США) разводят до концентрации 50% (соответствует примерно 102 шарикам) в мозжечок и кору на глубину 150–350 мкм с помощью Nanoject II (см. выше) перед ретракцией и перед размещением объективов GRIN как описано выше. Животных помещали на основу и визуализировали так же, как мышей, которым вводили вирусные инъекции. Сдвиги в плоскости x-y определялись с помощью функции motion_metrics в NoRMCorre.
Визуальный анализ кальция
Запросить подробный протокол Временные метки для всех осциллографов были записаны на диск, а в случае записи двух осциллографов кадры видео были выровнены перед коррекцией движения с помощью NoRMCorre (Pnevmatikakis and Giovannucci, 2017) и извлечением сигнала с помощью CNMF-E (Zhou et al., 2018). ). Мы использовали следующие параметры в CNMF-E для сегментации дендритов клеток Пуркинье и нейронов коры головного мозга соответственно (дендриты клеток Пуркинье: gSig = 7, gSiz = 40; нейроны коры головного мозга: gSig = 7, gSiz = 10, для обоих merge_thr = [ 1д-1, 0,85, 0,5]). В экспериментах по стимуляции мозжечка нейроны в коре классифицировались как реагирующие, если постстимульный сигнал превышал среднее значение +2σ до стимула. Время начала этих кальциевых переходных процессов определяли путем подгонки сигмовидной функции к переходным процессам, а время начала устанавливали таким образом, чтобы подогнанная функция превышала среднее значение + σ базового уровня до стимула. Для экспериментов с дорсальным полосатым телом нейроны классифицировали как респондеры, если сигнал во время периода стимуляции значительно отличался от исходного сигнала до стимула. Для каждой клетки активность до стимула и во время стимуляции усреднялась по испытанию и статистически тестировалась с использованием парных t-тестов. Фиджи (Schindelin et al., 2012) использовался для проверки необработанных данных и создания видеороликов. Анализы проводились в Matlab (Mathworks, Nantucket), Python 3.7 (Rossum, 1995) и (R Development Core Team, 2019).
Обнаружение внутрирегионального и межрегионального синхронного шаблона (SP)
Запросить подробный протокол Переходные явления кальция были выведены с использованием алгоритма конечной скорости обновления для быстрого и точного обнаружения всплесков (Oñativia et al., 2013), установив τ = 1 для получения времени начала переходных процессов кальция с разрешением субзаписи скорости из переходных процессов кальция. на ячейку. Эти времена начала затем были свернуты с ядром Епанечникова (крутизна = 0,1) и суммированы по всем ячейкам, что привело к сумме ядер. Все временные интервалы, для которых сумма ядер превышала среднее значение на два стандартных отклонения, считались значимыми глобальными синхронными событиями. Если время начала двух ячеек попадало в одно и то же синхронное событие, считалось, что эти ячейки сработали синхронно один раз. Для всех пар ячеек мы подсчитали, сколько раз эти ячейки срабатывали в одном и том же синхронном событии. Впоследствии мы отобрали все синхронные пары (СП), которые срабатывали вместе не менее пяти раз.
Эти пары были сохранены в графе, где каждый узел представляет собой ячейку, а каждое ребро — количество общих синхронных событий запуска. Эти графики были преобразованы в дуговую диаграмму с помощью пакета arcdiagram (Gaston Sanchez, https://github.com/gastonstat/arcdiagram) в R. Клетки были сгруппированы по областям мозга (мозжечок, кора) и отсортированы внутри группы по степени графа. , то есть количество ячеек, с которыми они связаны. Степень корреляции представлена радиусом узла. Написанные на заказ скрипты для извлечения SP были написаны на Python и могут быть загружены по адресу: https://github.com/Romanovg185/sps-continuous-time-data.
Анализ поведенческого ускорения
Запросить подробный протоколЧтобы различить, превышали ли сигналы акселерометра средний порог +2σ до или после SP, мы сначала определили, где возник наибольший средний сигнал. Затем мы отобрали сигналы до или после SP, которые превысили пороговое значение. Для остальных сигналов мы искали 300 мс вокруг SP (-150, +150 мс) для повышения выше порога. Все остальные сигналы были классифицированы как не демонстрирующие изменения в поведенческом ускорении, связанного с SP.
Анализ выращивания
Запросить подробный протокол Мы изучили видеозаписи поведения животных, чтобы определить приблизительное время выращивания (и получить количество посиделок на период записи), а затем использовали соответствующие данные акселерометра для определения продолжительности выращивания. Для всех подъемов, отмеченных в наших видеоданных, можно было различить четко различимый рост и падение данных акселерометра y и z (см. Рисунок 2F). Канал ускорения y использовался для оценки времени начала и окончания периода выращивания. Для оценки поведения использовали разных мышей для всех конфигураций с одним и двумя минископами.
Наличие файлов проекта и программного обеспечения
Запросить подробный протоколPCB и механические конструкции, микропрограммы и программное обеспечение для сбора данных можно найти на GitHub по адресу: https://github.com/ninscope.
Доступность данных
Оборудование, микропрограммы и программное обеспечение размещены на GitHub под лицензией MIT.
Ссылки
-
- Аккал Д
- Дум РП
- Стрик PL
(2007) Дополнительная моторная зона и предварительная дополнительная моторная зона: мишени для базальных ганглиев и выходной сигнал мозжечка
Journal of Neuroscience 27 :10659–10673.

https://doi.org/10.1523/JNEUROSCI.3134-07.2007
- пабмед
- Google ученый
-
- Бадура А
- Verpeut JL
- Мецгер JW
- Перейра ТД
- Пизано TJ
- Деверетт Б
- Бакшинская ДЭ
- Ван СС
(2018) Нормальное когнитивное и социальное развитие требует задней мозжечковой активности
eLife 7 :e36401.
https://doi.org/10.7554/eLife.36401
- пабмед
- Google ученый
-
- Беймбридж КГ
- Целио MR
- Роджерс JH
(1992) Белки, связывающие кальций, в нервной системе
Trends in Neurosciences 15 :303–308.
https://doi.org/10.1016/0166-2236(92)-I
- пабмед
- Google ученый
-
- Барбера Г
- Лян Б
- Чжан Л
- Герфен CR
- Кулурчелло E
- Чен Р
- Ли Ю
- Лин ДТ
(2016) Пространственно компактные нейронные кластеры в дорсальном полосатом теле кодируют информацию, относящуюся к передвижению
Нейрон 92 :202–213.
https://doi.org/10.1016/j.
neuron.2016.08.037
- пабмед
- Google ученый
-
- Барбера Г
- Лян Б
- Чжан Л
- Ли Ю
- Лин ДТ
(2019) Беспроводной минископ для глубокой визуализации мозга у свободно передвигающихся мышей
Journal of Neuroscience Methods 323 :56–60.
https://doi.org/10.1016/j.jneumeth.2019.05.008
- пабмед
- Google ученый
-
- Бокарсли ME
- Цзян туалет
- Ван С
- Дудман JT
- Джи Н
- Апонте Y
(2015) Минимально инвазивная система микроэндоскопии для функциональной визуализации in vivo глубоких ядер мозга мыши
Biomedical Optics Express 6 :4546–4556.
https://doi.org/10.1364/BOE.6.004546
- пабмед
- Google ученый
-
- Бостан AC
- Дум РП
- Стрик PL
(2013) Сети мозжечка с корой головного мозга и базальными ганглиями
Trends in Cognitive Sciences 17 :241–254.
https://doi.org/10.1016/j.tics.2013.03.003
- пабмед
- Google ученый
-
- Кай диджей
- Аарони Д
- Шуман Т
- Шобе J
- Биан Дж
- Песня W
- Вэй Б
- Вешкини М
- Ла-Ву М
- Лу Дж
- Флорес ЮВ
- Ким я
- Сано Y
- Чжоу М
- Баумгартель К
- Лави А
- Камата М
- Тушинский М
- Мэйфорд М
- Гольшани П
- Сильва AJ
(2016) Общий нейронный ансамбль связывает отдельные контекстуальные воспоминания, закодированные близко во времени
Природа 534 :115–118.
https://doi.org/10.1038/nature17955
- Google ученый
-
- Целио MR
(1990) Кальбиндин D-28k и парвальбумин в нервной системе крыс
Неврология 35 :375–475.
https://doi.org/10.1016/0306-4522(90)
-H - пабмед
- Google ученый
- Чен КС
- Сюй М
- Чжан Z
- Чанг туалет
- Гай Т
- Шаффер ДВ
- Дэн Y
(2018) Гипоталамический переключатель для быстрого и медленного сна
Нейрон 97 :1168–1176.
https://doi.org/10.1016/j.neuron.2018.02.005
- пабмед
- Google ученый
- Корбит ВЛ
- Мэннинг EE
- Гиттис AH
- Ахмари SE
(2019) Усиление входных сигналов от вторичной моторной коры к полосатому телу в мышиной модели компульсивного поведения
Журнал неврологии 39 :2965–2975.
https://doi.org/10.1523/JNEUROSCI.1728-18.2018
- пабмед
- Google ученый
- Кокс Дж
- Пинто L
- Дэн Y
(2016) Кальций-визуализация нейронной активности, связанной со сном и бодрствованием, в дорсальном мосту
Nature Communications 7 :10763.
https://doi.org/10.1038/ncomms10763
- пабмед
- Google ученый
- Цуй Г
- июнь SB
- Джин Х
- Фам MD
- Фогель СС
- Любовник ДМ
- Коста ринггитов
(2013) Одновременная активация стриарных прямых и непрямых путей во время инициации действия
Природа 494 : 238–242.
https://doi.org/10.1038/nature11846
- пабмед
- Google ученый
- Галинанес GL
- Бонарди С
- Хубер Д
(2018) Направленное стремление к воде как поведенческая структура, зависящая от кортекса, у мышей
Мобильные отчеты 22 :2767–2783.
https://doi.org/10.1016/j.celrep.2018.02.042
- пабмед
- Google ученый
- Гао Z
- Дэвис С
- Томас AM
- Экономо МН
- Абрего AM
- Свобода К
- Де Зеув CI
- Ли Н
(2018) Корково-мозжечковая петля для планирования движений
Природа 563 :113–116.
https://doi.org/10.1038/s41586-018-0633-x
- пабмед
- Google ученый
- Гош КК
- Бернс LD
- Кокер ЭД
- Ниммерьян А
- Зив И
- Гамаль АЕ
- Шницер MJ
(2011) Миниатюрная интеграция флуоресцентного микроскопа
Nature Methods 8 :871–878.
https://doi.org/10.1038/nmeth.1694
- пабмед
- Google ученый
- Джованнуччи А
- Бадура А
- Деверетт Б
- Наджафи F
- Перейра ТД
- Гао Z
- Озден I
- Клот г. н.э.
- Пневматикакис E
- Панинский Л
- Де Зеув CI
- Медина JF
- Ван СС
(2017) Зернистые клетки мозжечка приобретают широко распространенный прогностический сигнал обратной связи во время двигательного обучения
Nature Neuroscience 20 :727–734.
https://doi.org/10.1038/nn.4531
- пабмед
- Google ученый
- Гонсалес WG
- Чжан Х
- Арутюнян А
- Лоис С
(2019) Сохранение представлений нейронов во времени и при повреждении гиппокампа
Наука 365 :821–825.
https://doi.org/10.1126/science.aav9199
- Google ученый
- Го JZ
- Грейвс AR
- Го WW
- Чжэн Дж
- Ли А
- Родригес-Гонсалес J
- Ли Н
- Маклин JJ
- Филлипс JW
- Меньш БД
- Брэнсон К
- Хантман AW
(2015) Cortex управляет выполнением квалифицированного движения
eLife 4 :e10774.
. https://doi.org/10.7554/eLife.10774
- пабмед
- Google ученый
- Хеффли W
- Песня EY
- Сюй Z
- Тейлор Б.Н.
- Хьюз Массачусетс
- МакКинни А
- Джошуа М
- Корпус С
(2018) Координированная активность лазающих волокон мозжечка сигнализирует о выученных сенсомоторных предсказаниях
Nature Neuroscience 21 :1431–1441.
https://doi.
org/10.1038/s41593-018-0228-8
- пабмед
- Google ученый
- Интирян Г
- Фостер NN
- Боуман I
- Бэй М
- Песня МОЙ
- Гоу Л
- Ямасита С
- Биенковски MS
- Зинг Б
- Чжу М
- Ян XW
- Ши JC
- Тога AW
- Донг Х-В
(2016) Корково-стриарная проекция мыши
Nature Neuroscience 19 :1100–1114.
https://doi.org/10.1038/nn.4332
- Google ученый
- Хобик FE
- Виттер Л
- Руигрок TJH
- Де Зеув CI
(2010) Дифференциальный оливо-мозжечковый корковый контроль рикошетной активности в ядрах мозжечка
PNAS 107 :8410–8415.
https://doi.org/10.1073/pnas.0
8107 - Google ученый
- Гувер JE
- Стрик PL
(1999) Организация выходов мозжечка и базальных ганглиев в первичную моторную кору по данным ретроградного транснейронального транспорта вируса простого герпеса 1-9 типа0005
Журнал неврологии 19 :1446–1463.
https://doi.org/10.1523/JNEUROSCI.19-04-01446.1999
- пабмед
- Google ученый
- Хуан CC
- Сугино К
- Сима Y
- Го С
- Бай С
- Меньш БД
- Нельсон СБ
- Хантман AW
(2013) Конвергенция мостового и проприоцептивного потоков на мультимодальных гранулярных клетках мозжечка
eLife 2 :e00400.
https://doi.org/10.7554/eLife.00400
- пабмед
- Google ученый
- Ханникатт БЖ
- Jongbloets BC
- Песня птиц WT
- Герц KJ
- Чжун Х
- Мао Т
(2016) Подробная карта возбуждающих входов полосатого тела показывает новую функциональную организацию
eLife 5 :e19103.
https://doi.org/10.7554/eLife.19103
- пабмед
- Google ученый
- Джейкоб г. н.э.
- Рамсаран АИ
- Мокле AJ
- Транс ЛМ
- Ян С
- Франкленд PW
- Джосселин С.А.
(2018) Компактный эндоскоп с креплением на голове для визуализации кальция in vivo у свободно ведущих себя мышей
Current Protocols in Neuroscience 84 :e51.
https://doi.org/10.1002/cpns.51
- пабмед
- Google ученый
- Ю С
- Босман LWJ
- Хугланд ТМ
- Велаутапиллай А
- Муругасан П
- Варнаар P
- Ван Гендерен RM
- Негрелло М
- Де Зеув CI
(2019) Нейроны нижней оливы реагируют на широкий спектр сенсорной информации, но находятся под гомеостатическим контролем
The Journal of Physiology 597 :2483–2514.
https://doi.org/10.1113/JP277413
- пабмед
- Google ученый
Препринт
- Юавинетт AL
- Бехит Г
- Черчленд AK
(2018) Хронически имплантированные нейропиксельные зонды обеспечивают высокую производительность записи у свободно движущихся мышей
биоRxiv.
https://doi.org/10.1101/406074
- Google ученый
- июнь JJ
- Штейнмец Н.А.
- Зигле JH
- Денман DJ
- Бауза М
- Барбаритс Б
- Ли АК
- Анастассиу CA
- Андрей А
- Айдын Ç
- Барбик М
- Бланш TJ
- Бонин В
- Коуто J
- Датта Б
- Граций СЛ
-
Гутниский Д. А.
- Хойссер М
- Карш Б
- Ледохович П
- Лопес см
- Мителут C
- Муса С
- Окунь М
- Пачитариу М
- Путцейс J
- Рич ПД
- Россант С
- Солнце WL
- Свобода К
- Карандини М
- Харрис КД
- Кох С
- О’Киф J
- Харрис ТД
(2017) Полностью интегрированные кремниевые зонды для регистрации нейронной активности с высокой плотностью
Nature 551 :232–236.
https://doi.org/10.1038/nature24636
- пабмед
- Google ученый
- Каваи Р
- Маркман Т
- Поддар R
- Ко Р
- Фантана AL
- Дхавале АК
- Кампф АР
- Ольвечский BP
(2015) Моторная кора необходима для обучения, но не для выполнения моторного навыка
Нейрон 86 :800–812.
https://doi.org/10.1016/j.neuron.2015.03.024
- пабмед
- Google ученый
- Ким TH
- Чжан И
- Лекок J
- Юнг JC
- Ли Дж
- Цзэн Х
- Найл CM
- Шницер MJ
(2016) Долгосрочный оптический доступ к приблизительно одному миллиону нейронов в коре головного мозга живой мыши
Cell Reports 17 : 3385–3394.
https://doi.org/10.1016/j.celrep.2016.12.004
- Google ученый
- Кингсбери Л
- Хуан С
- Ван Дж
- ГУ К
- Гольшани П
- У Е
- Хонг W
(2019) Коррелированная нейронная активность и кодирование поведения в мозгу социально взаимодействующих животных
Клетка 178 :429–446.
https://doi.org/10.1016/j.cell.2019.05.022
- пабмед
- Google ученый
- Клаус А
- Мартинс GJ
- Пайшао VB
- Чжоу П
- Панинский Л
- Коста ринггитов
(2017) Пространственно-временная организация полосатого тела кодирует пространство действия
Нейрон 95 :1171–1180.
https://doi.org/10.1016/j.neuron.2017.08.015
- пабмед
- Google ученый
- Костадинов Д
- Бо М
- Позо МБ
- Хойссер М
(2019) Прогностические и реактивные сигналы вознаграждения, передаваемые входами восходящих волокон к клеткам Пуркинье мозжечка
Nature Neuroscience 22 :950–962.
https://doi.org/10.1038/s41593-019-0381-8
- пабмед
- Google ученый
Конференц-связь
- Леман ДП
- Чен ИА
- Йена WW
- Круз-Мартин А
- Перкинс N
- Либерти WA
- Гарднер TJ
- Отчи ТМ
- Дэвисон IG
(2018)
Расширенный набор инструментов с открытым исходным кодом для широкопольной визуализации кальция у свободно ведущих себя животных
Общество нейробиологов, 2018 г.
- Google ученый
- Лян Б
- Чжан Л
- Барбера Г
- Клык W
- Чжан Дж
- Чен Х
- Чен Р
- Ли Ю
- Лин ДТ
(2018) Отдельные и динамичные нейронные ансамбли ВКЛ и ВЫКЛ в префронтальной коре кодируют социальное исследование
Нейрон 100 :700–714.
https://doi.org/10.1016/j.neuron.2018.08.043
- пабмед
- Google ученый
- Либерти WA
- Марковиц JE
- Перкинс ЛН
- Либерти DC
- Леман ДП
- Гитчаунтс G
- Велью Т
- Коттон ДН
- Лоис С
- Гарднер TJ
(2016) Нестабильные нейроны лежат в основе стабильного усвоенного поведения
Nature Neuroscience 19 :1665–1671.
https://doi.org/10.1038/nn.4405
- пабмед
- Google ученый
- Либерти WA
- Перкинс ЛН
- Леман ДП
- Гарднер TJ
(2017) Миниатюрная микроскопическая система с открытым исходным кодом и возможностью беспроводной связи
Journal of Neural Engineering 14 :045001.
https://doi.org/10.1088/1741-2552/aa6806
- пабмед
- Google ученый
- Муруган М
- Чан ХДж
- Парк М
- Миллер ЭМ
- Кокс Дж
- Талиаферро JP
- Паркер НФ
- Бхаве В
- Гур H
- Лян И
- Нектоу AR
- Подушка JW
- Виттен IB
(2017) Комбинированное социальное и пространственное кодирование в нисходящей проекции из префронтальной коры
Сотовый 171 :1663–1677.
https://doi.org/10.1016/j.cell.2017.11.002
- пабмед
- Google ученый
- Неделеску H
- Абдельхак М
(2013) Сравнительная морфология дендритных стволов в популяциях клеток Пуркинье в борозде мыши и верхушке
Нейронная пластичность 2013 :1–12.
https://doi.org/10.1155/2013/948587
- Google ученый
- Онативиа J
- Шульц SR
- Драготти PL
(2013) Алгоритм с конечной скоростью обновления для быстрого и точного обнаружения спайков с помощью двухфотонной визуализации кальция
Журнал нейронной инженерии 10 :046017.
https://doi.org/10.1088/1741-2560/10/4/046017
- пабмед
- Google ученый
- Пневматикакис EA
- Джованнуччи А
(2017) NoRMCorre: онлайн-алгоритм кусочно-жесткой коррекции данных визуализации кальция
Journal of Neuroscience Methods 291 :83–94.
https://doi.org/10.1016/j.jneumeth.2017.07.031
- пабмед
- Google ученый
- Пруденте CN
- Стилла R
- Бютефиш СМ
- Сингх С
- Гесс EJ
- Ху Х
- Сатиан К
- Джинна HA
(2015) Нервные субстраты движений головы у человека: исследование функциональной магнитно-резонансной томографии
Journal of Neuroscience 35 :9163–9172.
https://doi.org/10.1523/JNEUROSCI.0851-15.2015
- пабмед
- Google ученый
- Quy PN
- Фуджита H
- Сакамото Y
- На Дж
- Сугихара I
(2011) Паттерны проекций одиночных аксонов мшистых волокон, происходящих из ядер дорсального столба, нанесенных на карту в компартментах альдолазы С в коре мозжечка крысы
The Journal of Comparative Neurology 519 :874–899.
https://doi.org/10.1002/cne.22555
- пабмед
- Google ученый
Программное обеспечение
- Основная группа разработки R
(2019) R: Язык и среда для статистических вычислений
Основная группа разработчиков R, Вена, Австрия.
https://www.R-project.org
- Ремедиос R
- Кеннеди А
- Зеликовский М
- Греве BF
- Шницер MJ
- Андерсон DJ
(2017) Социальное поведение формирует представления гипоталамического нейронного ансамбля о половой принадлежности
Природа 550 :388–392.
https://doi.org/10.1038/nature23885
- пабмед
- Google ученый
- Романо В
- Де Пропри L
- Босман LW
- Варнаар P
- Тен Бринке MM
- Линдеман С
- Ю С
- Велаутапиллай А
- Спанке JK
- Миддендорп Герра E
- Хугланд ТМ
- Негрелло М
- Д’Анджело E
- Де Зеув CI
(2018) Потенцирование клеток Пуркинье мозжечка облегчает адаптацию усикового рефлекса за счет повышения активности простых спайков
eLife 7 :e38852.
https://doi.org/10.7554/eLife.38852
- пабмед
- Google ученый
Книга
- Россум G
(1995)
Учебное пособие по Python
Амстердам: Centrum voor Wiskunde en Informatica (CWI).
- Google ученый
- Руигрок TJH
(2011) Входы и выходы модулей мозжечка
Мозжечок 10 :464–474.
https://doi.org/10.1007/s12311-010-0164-y
- Google ученый
- Зауэрбрей BA
- Го JZ
- Коэн JD
- Мискиати М
- Го W
- Кабра М
- Верма N
- Меньш Б
- Брэнсон К
- Хантман AW
(2020) Генерация корковых паттернов во время ловких движений управляется вводом
Природа 577 :386–391.
https://doi.org/10.1038/s41586-019-1869-9
- пабмед
- Google ученый
- Шинделин J
- Арганда-Каррерас I
- Фризе Е
- Кайниг V
- Лонгэйр М
- Пицш Т
- Прейбиш S
- Рюден C
- Заальфельд С
- Шмид Б
- Тиневез JY
- Белый ди-джей
- Хартенштейн V
- Элисейри К
- Томанчак П
- Кардона А
(2012) Фиджи: платформа с открытым исходным кодом для анализа биологических изображений
Природные методы 9 :676–682.
https://doi.org/10.1038/nmeth.2019
- пабмед
- Google ученый
- Шуман Т
- Аарони Д
- Кай диджей
- Ли КР
- Чавлис S
- Пейдж-Harley L
- Ветере LM
- Фэн Ю
- Ян CY
- Моллинедо-Гахате I
- Чен Л
- Пеннингтон ZT
- Таксидис J
- Флорес SE
- Ченг К
- Джавахериан М
- Каба CC
- Рао Н
- Ла-Ву М
- Панди я
- Штрахман М
-
Бахурин К. И.
- Масманидис SC
- Хах БС
- Поирази P
- Сильва AJ
- Гольшани П
(2020) Нарушение пространственного кодирования и синхронизации интернейронов у мышей с эпилепсией
Nature Neuroscience 7 :0559.
https://doi.org/10.1038/s41593-019-0559-0
- Google ученый
- Стаматакис AM
- Шахтер MJ
- Гулати С
- Зителли КТ
- Малановский S
- Таджик А
- Фриц С
- Трулсон М
- Отте SL
(2018) Одновременная оптогенетика и визуализация кальция с клеточным разрешением во время активного поведения с использованием миниатюрного микроскопа
Frontiers in Neuroscience 12 :496.
https://doi.org/10.3389/fnins.2018.00496
- пабмед
- Google ученый
- Стирман JN
- Смит IT
- Куденов М.В.
- Смит СЛ
(2016) Широкоугольная, многозонная, двухфотонная визуализация активности нейронов в мозге млекопитающих
Природа Биотехнология 34 :857–862.
https://doi.org/10.1038/nbt.3594
- пабмед
- Google ученый
- Студли CJ
- Д’Мелло AM
- Эллегуд J
- Джаккамсетти V
- Лю П
- Небель МБ
- Гибсон JM
- Келли Э
- Мэн Ф
- Кано CA
- Паскуаль JM
- Мостофский СХ
- Лерх JP
- Цай PT
(2017) Изменение связности мозжечка при аутизме и опосредованное мозжечком восстановление поведения, связанного с аутизмом, у мышей
Nature Neuroscience 20 :1744–1751.
https://doi.org/10.1038/s41593-017-0004-1
- пабмед
- Google ученый
- Текуапетла F
- Матиас С
- Дуг GP
- Майнен ZF
- Коста ринггитов
(2014) Сбалансированная активность в проекционных путях базальных ганглиев имеет решающее значение для контраверсивных движений
Nature Communications 5 :4315.
https://doi.org/10.1038/ncomms5315
- пабмед
- Google ученый
- Терада СИ
- Кобаяши К
- Окура М
- Накаи Дж
- Мацудзаки М
(2018) Сверхширокопольная двухфотонная визуализация с помощью микрооптического устройства, перемещающегося в постобъективном пространстве
Nature Communications 9 :3550.
https://doi.org/10.1038/s41467-018-06058-8
- пабмед
- Google ученый
- Фогельштейн JT
- Уотсон БО
- Пакер АМ
- Юсте Р
- Джедынак Б
- Панинский Л
(2009) Определение всплесков на основе изображений кальция с использованием последовательных методов Монте-Карло
Биофизический журнал 97 : 636–655.
https://doi.org/10.1016/j.bpj.2008.08.005
- Google ученый
- Вагнер MJ
- Ким TH
- Савалл J
- Шницер MJ
- Ло Л
(2017) Зернистые клетки мозжечка кодируют ожидание вознаграждения
Природа 544 :96–100.
https://doi.org/10.1038/nature21726
- пабмед
- Google ученый
- Вагнер МЮ
- Ким TH
- Кадмон Дж
- Нгуен ND
- Гангули С
- Шницер MJ
- Ло Л
(2019) Общая корково-мозжечковая динамика при выполнении и изучении двигательной задачи
Ячейка 177 :669–682.
https://doi.org/10.1016/j.cell.2019.02.019
- пабмед
- Google ученый
- Виттер Л
- Песнь CB
- Хугланд ТМ
- де Груйл JR
- Де Зеув CI
(2013) Сила и время двигательных реакций, опосредованных рикошетным возбуждением в ядрах мозжечка после активации клеток Пуркинье
Frontiers in Neural Circuits 7 :133.
https://doi.org/10.3389/fncir.2013.00133
- пабмед
- Google ученый
- Чжан Л
- Лян Б
- Барбера Г
- Хоуз С
- Чжан И
- Пень К
- Баум I
- Ян Ю
- Ли Ю
- Лин ДТ
(2019) Система Miniscope GRIN Lens для кальциевой визуализации активности нейронов в глубоких структурах головного мозга у животных с поведением
Current Protocols in Neuroscience 86 :e56.
https://doi.org/10.1002/cpns.56
- Google ученый
- Чжоу П
- Резендез СЛ
- Родригес-Ромагера J
- Хименес JC
- Нойфельд SQ
- Джованнуччи А
- Фридрих J
- Пневматикакис EA
- Штубер ГД
- Курица R
- Хейрбек М.А.
- Сабатини BL
- Касс RE
- Панинский Л
(2018) Эффективное и точное извлечение сигналов кальция in vivo из микроэндоскопических видеоданных
eLife 7 :e28728.
https://doi.org/10.7554/eLife.28728
- пабмед
- Google ученый
Письмо-решение
Сводка о принятии:
Рецензенты и редакторы высоко оценили универсальность и практичность общедоступной системы NINscope. По сравнению с другими системами, NINscope имеет преимущества в весе и размере, что позволяет использовать акселерометры, оптогенетическую стимуляцию и возможность получения изображений с двух участков. Вместе они открывают более широкие возможности, чем существуют с другими системами. Поскольку она находится в открытом доступе, мы ожидаем, что эта система будет особенно полезным дополнением для лабораторий, у которых нет возможности разрабатывать эти дополнительные функции самостоятельно путем модификации существующих систем.
Письмо с решением после экспертной оценки:
Благодарим вас за представление вашей статьи «NINscope: универсальный минископ для исследований в нескольких регионах» на рассмотрение в eLife .
Ваша статья была проверена тремя рецензентами, а за оценкой наблюдали редактор-рецензент и Кейт Вассум в качестве старшего редактора. Следующее лицо, участвовавшее в рассмотрении вашей заявки, согласилось раскрыть свою личность: Дениз Джей Кай (Рецензент №1). Рецензенты обсудили обзоры друг с другом, и редактор-рецензент подготовил это решение, чтобы помочь вам подготовить исправленное представление.
Резюме:
Ваш документ вызвал довольно много дискуссий на стадии консультаций. С одной стороны, рецензенты считали, что отдельные технические достижения здесь были относительно небольшими. С другой стороны, было достигнуто согласие в том, что, в принципе, небольшие преимущества в весе и размере, хотя и увеличивающиеся по отдельности, в сочетании с акселерометрами, оптогенетической стимуляцией и возможностью получения изображений с двух участков, вместе могут открыть более широкие возможности, чем существующие с другими системами. . Основное преимущество NINscope было замечено в универсальности и практическом удобстве использования, а не в технических характеристиках как таковых.
Выделение того, КАК комбинировать визуализацию с меньшими размерами с другими приложениями (опто, двойная визуализация), является хорошим вкладом. Если бы эта система стала общедоступной, она могла бы стать полезным дополнением для лабораторий, у которых нет возможности разрабатывать эти дополнительные функции самостоятельно путем модификации существующих систем. В статье «Инструменты и ресурсы» не требуется сообщать о новых важных биологических открытиях или механизмах, но должно быть ясно, что они позволят осуществить такие достижения. Поскольку цель здесь не в том, чтобы ответить на биологические вопросы, необходимо более четко указать, что эксперименты являются демонстрационными экспериментами (т. е. объем и связанные с ними технологические работы), И данные должны быть высокого качества.
Были подняты ключевые вопросы, которые необходимо решить, чтобы убедительно продемонстрировать эти моменты, кратко изложенные здесь:
1) Самое главное, есть опасения по поводу качества данных изображения, представленных в дополнительных видеороликах, включая явный артефакт движения по оси Z, деконволюцию сигналов GCaMP и сегментацию дендритов клеток Пуркинье (по-разному затрагиваемые всеми 3 рецензента, но особенно рецензента 3).
Видео также сильно обработано, красные вспышки указывают на активность, что скрывает сегментацию и извлечение сигнала. Чтобы гарантировать, что меньший размер NINscope не принесет в жертву стабильность и качество изображения, мы просим вас: – Обеспечьте средства управления YFP (или другим статическим флуорофором) для артефактов движения. Это особенно (но не только) актуально для случаев повышенной активности в регионах, которые коррелируют с ускорением головы.
— Предоставьте больше необработанных данных и информации о сегментации. В видео, пожалуйста, предоставьте те же данные изображения кальция в необработанном виде, после коррекции движения, а затем снова после сегментации. Это можно сделать, показав одно и то же видео для этих трех шагов.
2) Высказывались опасения, что слишком много делается с точки зрения биологических результатов, полученных в результате показанных разрозненных экспериментальных экспериментов. Эти исследования считались недостаточно мощными, в них не было экспериментальных деталей и контрольных экспериментов, чтобы сделать убедительные биологические выводы.
Более того, подробный ответ на все эти биологические вопросы выходит за рамки инструментов и справочного документа. Пожалуйста, сосредоточьтесь на аспектах инструментов и методов (и объясните это) и смягчите выводы о результатах скоординированной нейронной активности в разных областях мозга. 3) Чтобы подтвердить полезность установки 2 эндоскопов на поведение животных, пожалуйста, предоставьте
– более показательные изображения конфигурации с двумя креплениями (как предложил рецензент 1)
– количественная оценка поведения с установленными 0,1,2 прицелами ( предложено рецензентом 2).
4) Чтобы можно было напрямую оценить характеристики NINscope по сравнению с другими существующими вариантами (минископ UCLA, Inscopix), приложите сравнительную таблицу, предложенную рецензентом 2.
5) Пожалуйста, предоставьте подтверждение истинного разделения стимуляции и места регистрации для оптогенетики (Рецензент 3, пункт 2).
Поэтому мы приглашаем вас представить исправленную рукопись, которая полностью решает эти проблемы, а также полный набор комментариев рецензентов, приведенных ниже.
Рецензент №1:
Groot et al. сообщают о конструкции и применении нового минископа — NINscope, основным преимуществом которого по сравнению с существующими технологиями минископов является его малый размер и малый вес, что позволяет имплантировать два из них для одновременной визуализации двух областей мозга у мышей. Кроме того, NINscope способен интегрироваться с оптогенетикой для одновременной визуализации и оптогенетического манипулирования цепями мозга. Авторы демонстрируют, что NINscope способен получать видео высокого разрешения, лежащую в основе нейронную активность которого можно деконволюционировать с помощью обычных методов обработки данных (например, CNMF). Затем они обнаруживают синхронные паттерны активности (СП) в коре и мозжечке во время начала движения, используя NINscope для записи в двух областях и встроенный акселерометр для измерения поведенческого движения. Чтобы продемонстрировать, что NINscope поддается интеграции с оптогенетикой, авторы затем визуализируют активность кальция в коре при стимуляции клеток Пуркинье в 4 субрегионах мозжечка и показывают, что стимуляция каждой субобласти увеличивает активность кальция в коре.
Более того, оценка поведенческого ускорения свидетельствует о том, что стимуляция одного полушария Crus II приводит к движению головы в противоположном направлении. Чтобы продемонстрировать способность NINscope получать изображения из глубоких структур мозга, авторы имплантировали линзу GRIN в дорсальную часть полосатого тела с помощью системы релейных линз и регистрировали кальциевую активность полосатых нейронов, когда мыши исследовали открытое поле. С помощью этой системы авторы наблюдают за отдельными нейронами, настроенными на повороты в сторону, противоположную регистрируемому полушарию. Наконец, авторы используют глубокую визуализацию мозга и оптогенетику, чтобы продемонстрировать, что оптогенетическая стимуляция OFC и M2 по-разному влияет на активность полосатых нейронов. В целом, NINscope служит ценным и гибким инструментом, позволяющим исследовать динамику популяций нейронов в разных областях мозга. Из-за своего небольшого размера он допускает гибкое использование, например, в сочетании с дополнительным NINscope или оптогенетикой. Этот инструмент, безусловно, принесет пользу сообществу нейронауков в целом, однако некоторые дополнительные детали методологии и разъяснения некоторых областей (рассмотренные ниже) значительно улучшат эту рукопись. И хотя то, что авторы показывают несколько примеров того, как NINscope можно использовать в экспериментах, похвально, неясно, какие биологические открытия заключаются в этих разных экспериментах, которые, кажется, решают разрозненные исследовательские вопросы. Авторы могут захотеть более четко указать, какие гипотезы основаны на предыдущих исследованиях, какие результаты согласуются с предыдущими исследованиями, а какие являются новыми или ставят под сомнение предыдущие взгляды. Я понимаю, что это может слишком расширить рамки этой статьи, и я хотел бы предложить авторам рассмотреть возможность сосредоточения своих экспериментальных вопросов. Рецензент №2:
de Groot et al. представить новый миниатюрный флуоресцентный микроскоп с открытым исходным кодом.
В прямом сравнении с наиболее широко используемыми миниатюрными флуоресцентными микроскопами новый прицел (получивший название NINscope) имеет несколько дополнительных функций/свойств, которые сделают его привлекательным для некоторых групп. В частности, NINscope занимает меньше места, легче и содержит встроенные акселерометры и драйверы светодиодов для управления внешними светодиодами. Эти функции хорошо интегрированы в дизайн, и их полезность продемонстрирована в рукописи. Чтобы продемонстрировать функциональность этого нового микроскопа и связанного с ним аппаратного и программного обеспечения, авторы подтвердили множество предыдущих выводов из литературы по движению. А именно, авторы используют NINscope для успешного отслеживания активности дендритов клеток Пуркинье и нейронов дорсального полосатого тела во время движения, для изучения функциональной связи между мозжечком и корой и для подтверждения выводов о том, что орбитофронтальная и моторная кора обеспечивают нисходящий контроль дорсального полосатого тела. На мой взгляд, основным результатом этой статьи является сам микроскоп, и для демонстрации его функций и полезности требуется совсем немногое. Однако некоторые модификации усилят распространение соответствующей информации о NINscope. Нейробиологические эксперименты, возможно, недостаточно мощные, но не настолько, чтобы подорвать сам микроскоп. Например, известные ограничения обнаружения снижения сигнала от GCaMP могут обеспечить некоторую погрешность на рисунке 6, но эта возможность не умаляет сделанных здесь технических достижений. Одним дополнением, которое усилит эту рукопись, является более подробное сравнение между NINscope и другими миниатюрными микроскопами с открытым исходным кодом (возможно, коммерческими, когда это применимо). Это может быть текст или рисунок, но исчерпывающая таблица может быть наиболее кратким способом передачи информации, которая будет полезна будущим пользователям. Я понимаю, что у последнего автора есть обзор на эту тему, но в этом обзоре не содержится ни самого NINscope, ни подробностей о беспроводном эндоскопе от Shuman et al.
(биоРксив, 2018). Прямое сравнение веса, размера основания, схем управления, механизма фокусировки, особенностей программного обеспечения, вспомогательного оборудования и возможностей стимуляции — это лишь некоторые из переменных/ограничений, которые могут склонить пользователя от одного прицела к другому. Другим основным сравнением, которое укрепило бы заявления о самом аппаратном обеспечении прицела (должно быть легко собрано или из существующих данных), является количественное сравнение двигательного поведения с нулевым, одним и двумя прицелами NIN, установленными на голове. Авторы предполагают, что наблюдается нормальное поведение, и относятся к исследованию клетки, уходу, еде и выращиванию, но на самом деле это не продемонстрировано. Эпоха выращивания определяется видеоданными и данными IMU в 2G, предположительно количество тылов может быть собрано из установок с одним и двумя телескопами и сравнено.
Было бы полезно включить дополнительную информацию о светодиодах для оптической стимуляции.
Это очень простое применение стандартных светодиодов, и другим может быть полезно использовать их с NINscope или без него. Дополнительные фотографии и/или схемы помогут. Мощность света для визуализации не указана для рисунка 6. Была ли она такой же, как на рисунке 5? В связи с возможностью активации синего хвоста ChrimsonR, пожалуйста, включите эту информацию.
Reviewer #3:
В этой рукописи de Groot et al. авторы разрабатывают и описывают миниатюрный эпифлюоресцентный микроскоп, который будет использоваться для визуализации кальция у свободно ведущих себя грызунов. Микроскоп похож на ранее опубликованные минископы (минископ UCLA, Inscopix nVista и Finchscope) с некоторыми незначительными отличиями. А именно, он немного легче, чем прицел Inscopix, и содержит акселерометр/светодиод для отслеживания положения. Он также имеет дополнительный светодиод, который можно использовать для фотостимуляции. В то время как другие системы не предлагают этого, кажется, что это было бы легко реализовать с другими системами, если это необходимо.
Конкретные комментарии:
1) Одним из основных моментов, отмеченных на протяжении всей рукописи, является то, что эта система допускает многосайтовую запись. Хотя это продемонстрировано в мозжечке и лобной коре, маловероятно, что это может быть достигнуто во многих других областях. Фронтальная кора и мозжечок находятся довольно далеко друг от друга (таким образом, пока это возможно в этом примере), но это будет сильно ограничено другими местами дуэли.
2) Интересна оптогенетическая стимуляция в сочетании с визуализацией, и ранее о ней сообщалось с использованием системы Inscopix nVoke (где стимуляция и запись происходят в одном и том же поле зрения). Преимущество этой статьи в том, что стимуляция может быть отделена от места записи. Хотя это интересно, похоже, не проводились контрольные эксперименты, в которых мыши не экспрессировали ChR2. Таким образом, вполне возможно, что нейронная активность, вызванная стимуляцией, может быть связана с каким-либо артефактом, вызванным нагревом или светом.
3) У меня есть некоторые опасения по поводу качества данных, представленных в дополнительных видеороликах. Многие из них показывают неисправимый артефакт z-движения, а записи дендритов мозжечка, по-видимому, показывают много флуоресценции не в фокусе, что затрудняет извлечение сигналов от отдельных дендритов. Авторы использовали недавно опубликованный алгоритм CMNF-E для извлечения сигналов, который извлекает что-то из данных, но неясно, действительно ли извлеченные сигналы представляют собой отдельные дендриты.
https://doi.org/10.7554/eLife.49987.sa1
Ответ автора
Рецензент №1:
[…] В целом, NINscope служит ценным и гибким инструментом, позволяющим исследовать динамику популяций нейронов в разных областях мозга. Из-за своего небольшого размера он допускает гибкое использование, например, в сочетании с дополнительным NINscope или оптогенетикой. Этот инструмент, безусловно, принесет пользу сообществу нейронауков в целом, однако некоторые дополнительные детали методологии и разъяснения некоторых областей (рассмотренные ниже) значительно улучшат эту рукопись.
И хотя то, что авторы показывают несколько примеров того, как NINscope можно использовать в экспериментах, похвально, неясно, какие биологические открытия заключаются в этих разных экспериментах, которые, кажется, решают разрозненные исследовательские вопросы. Авторы могут захотеть более четко указать, какие гипотезы основаны на предыдущих исследованиях, какие результаты согласуются с предыдущими исследованиями, а какие являются новыми или ставят под сомнение предыдущие взгляды. Я понимаю, что это может слишком расширить рамки этой статьи, и я хотел бы предложить авторам рассмотреть возможность сосредоточения своих экспериментальных вопросов. Мы благодарим рецензента за то, что он уделил время внимательному прочтению нашей рукописи, ценные предложения по ее улучшению и общую положительную оценку. Мы согласны с выводом о том, что продемонстрированные эксперименты не обязательно связаны с всеобъемлющей гипотезой, но нашей целью в этой рукописи «Инструменты и ресурсы» было, прежде всего, показать универсальность и применимость NINscope, что привело к разнородному набору экспериментов.
Признавая вышеизложенное, мы смягчили выводы, сделанные в отношении биологических данных. Рецензент №2:
[…] Однако некоторые модификации усилят распространение соответствующей информации о NINscope. Нейробиологические эксперименты, возможно, недостаточно мощные, но не настолько, чтобы подорвать сам микроскоп. Например, известные ограничения обнаружения снижения сигнала от GCaMP могут обеспечить некоторую погрешность на рисунке 6, но эта возможность не умаляет сделанных здесь технических достижений.
Одним дополнением, которое усилит эту рукопись, является более подробное сравнение между NINscope и другими миниатюрными микроскопами с открытым исходным кодом (возможно, коммерческими, когда это применимо). Это может быть текст или рисунок, но исчерпывающая таблица может быть наиболее кратким способом передачи информации, которая будет полезна будущим пользователям. Я понимаю, что у последнего автора есть обзор на эту тему, но в этом обзоре не содержится ни самого NINscope, ни подробностей о беспроводном эндоскопе от Shuman et al.
(биоРксив, 2018). Прямое сравнение веса, размера основания, схем управления, механизма фокусировки, особенностей программного обеспечения, вспомогательного оборудования и возможностей стимуляции — это лишь некоторые из переменных/ограничений, которые могут склонить пользователя от одного прицела к другому. Спасибо за это предложение. Мы добавили таблицу, в которой мы сравниваем NINscope с другими доступными в настоящее время минископами (с открытым исходным кодом и коммерческими).
Другим основным сравнением, которое укрепило бы заявления о самом аппаратном обеспечении эндоскопа (должно быть легко собрано или из существующих данных), является количественное сравнение двигательного поведения с нулевым, одним и двумя прицелами NIN, установленными на голове. Авторы предполагают, что наблюдается нормальное поведение, и относятся к исследованию клетки, уходу, еде и выращиванию, но на самом деле это не продемонстрировано. Эпоха выращивания определяется видеоданными и данными IMU в 2G, предположительно количество тылов может быть собрано из установок с одним и двумя телескопами и сравнено.
Было бы полезно включить дополнительную информацию о светодиодах для оптической стимуляции. Это очень простое применение стандартных светодиодов, и другим может быть полезно использовать их с NINscope или без него. Дополнительные фотографии и/или схемы помогут.
Это отличное предложение. По предложению рецензента мы количественно оценили количество вставаний на дыбы у мышей с одинарным и двойным минископом (конфигурация CBL-CTX) для 4 мышей. Мы обнаружили, что на частоту и продолжительность выращивания не влияет добавление дополнительного минископа.
Мы обновили рисунок 1: теперь мы включили отдельную панель (1D), чтобы показать светодиодный датчик, который мы создали для использования в сочетании с NINscope, и добавили короткий абзац к тексту, описывающему его части. Сборка зондов подробно описана на нашей аппаратной вики GitHub: https://github.com/ninscope/Hardware/wiki/8.-LED-probe.
Мощность света для визуализации не указана для рисунка 6.
Была ли она такой же, как на рисунке 5? В связи с возможностью активации синего хвоста ChrimsonR, пожалуйста, включите эту информацию. По запросу мы добавили информацию о мощности света. Рецензент поднимает обоснованный вопрос о возможности активации ChrimsonR синим светом. Мы полагаем, что активация синего (возбуждающего) хвоста ChrimsonR, вероятно, смягчается следующими факторами:
1) Мощность света, используемая для визуализации полосатого тела, измеренная под релейной линзой GRIN — теперь представлена на рисунке 2 — дополнение к рисунку 1 соответствует примерно до 170 мкВт. Эта световая мощность более чем на порядок ниже, чем световая мощность красного светодиода, используемого для стимуляции OFC и M2 в коре головного мозга.
2) Мы провели эти эксперименты через четыре недели после инъекции, когда терминальная экспрессия ChrimsonR была еще относительно слабой.
3) Даже если синий свет возбуждает терминали, экспрессирующие ChrimsonR, мы все равно наблюдаем дифференциальный постсинаптический ответ при стимуляции M2 и OFC.
Рецензент №3:
В этой рукописи de Groot et al. авторы разрабатывают и характеризуют миниатюрный эпифлуоресцентный микроскоп, который будет использоваться у свободно ведущих себя грызунов для визуализации кальция. Микроскоп похож на ранее опубликованные минископы (минископ UCLA, Inscopix nVista и Finchscope) с некоторыми незначительными отличиями. А именно, он немного легче, чем прицел Inscopix, и содержит акселерометр/светодиод для отслеживания положения. Он также имеет дополнительный светодиод, который можно использовать для фотостимуляции. В то время как другие системы не предлагают этого, кажется, что это было бы легко реализовать с другими системами, если это необходимо.
Пункты, сделанные выше, подчеркивают, что, возможно, нам следовало более подробно рассказать об усилиях, которые прилагаются к проектированию и созданию интегральных схем, позволяющих программно управлять всеми аспектами экспериментальных записей. Мы спроектировали и изготовили две компактные интегральные схемы для NINscope, уменьшив вес минископа, интегрировав акселерометр, добавив возможности визуализации с двух участков и разрешив оптогенетическую стимуляцию.
Добавление функциональности с помощью отдельных компонентов приведет к увеличению ненужного веса или кабелей, что может повлиять на поведение животных, и, прежде всего, потребует выполнения трудоемких шагов для обеспечения постфактум синхронизации данных. Даже если бы эти функции могли быть добавлены, это потребовало бы определенных технических навыков, к которым не у всех есть доступ.
Мы считаем, что, делясь этим ресурсом как инструментом с открытым исходным кодом, мы можем гарантировать, что большая группа пользователей сможет извлечь выгоду из его специфических функций и позволит им решать новые исследовательские вопросы.
Особые комментарии:
1) Одним из основных моментов, отмеченных в рукописи, является то, что эта система допускает многосайтовую запись. Хотя это продемонстрировано в мозжечке и лобной коре, маловероятно, что это может быть достигнуто во многих других областях. Фронтальная кора и мозжечок находятся довольно далеко друг от друга (таким образом, пока это возможно в этом примере), но это будет сильно ограничено другими местами дуэли.
Спасибо, что указали на это. Чтобы более четко продемонстрировать пригодность NINscope для записи в нескольких местах, мы теперь демонстрируем дополнительные конфигурации с двумя эндоскопами на рисунке 3 — дополнения к рисунку 1 и 2, включая изображения двух полушарий из соматосенсорной и зрительной коры, гиппокампа или двух полушарий мозжечка. .
2) Интересна оптогенетическая стимуляция в сочетании с визуализацией, и ранее о ней сообщалось с использованием системы Inscopix nVoke (где стимуляция и запись происходят в одном и том же поле зрения). Преимущество этой статьи в том, что стимуляция может быть отделена от места записи. Хотя это интересно, похоже, не проводились контрольные эксперименты, в которых мыши не экспрессировали ChR2. Таким образом, вполне возможно, что нейронная активность, вызванная стимуляцией, может быть связана с каким-либо артефактом, вызванным нагревом или светом.
Теперь мы включаем рисунок 4 — приложение к рисунку 1, которое демонстрирует, что стимуляция в нескольких местах мозжечка не вызывает ни ответа в нейронах коры, ни поведенческого ответа.
3) У меня есть некоторые опасения по поводу качества данных, представленных в дополнительных видеороликах. Многие из них показывают неисправимый артефакт z-движения, а записи дендритов мозжечка, по-видимому, показывают много флуоресценции не в фокусе, что затрудняет извлечение сигналов от отдельных дендритов. Авторы использовали недавно опубликованный алгоритм CMNF-E для извлечения сигналов, который извлекает что-то из данных, но неясно, действительно ли извлеченные сигналы представляют собой отдельные дендриты.
Мы согласны с рецензентом в том, что визуализация в представленных видеороликах была нечеткой. Красные вспышки представляли необработанную флуоресценцию с вычитанием среднего, наложенную на необработанные данные, а видео были сохранены в сжатом формате, который не четко представлял данные. Поэтому теперь мы предоставляем дополнительные видео более высокого качества, которые включают пошаговый процесс от необработанных данных до коррекции движения и сегментации на протяжении всей рукописи.
Эти видеоролики в сочетании с экспериментами с двумя минископами, в которых мы вводили флуоресцентные шарики в мозжечок и кору головного мозга, показывают, что артефакты движения на самом деле довольно скромны. Чтобы показать, что CNMF-E может успешно сегментировать дендриты клеток Пуркинье, мы обновили рисунок 2 и другие рисунки, чтобы показать извлеченные пространственные следы. Наши пространственные следы дендритов клеток Пуркинье соответствуют критериям парасагиттально выровненных удлиненных структур, а извлеченные сигналы соответствуют тому, что мы ожидаем от сложных переходных процессов, вызванных спайками кальция, с частотой возбуждения около 1 Гц. Более того, учитывая анатомию мозжечка и тот факт, что мы получаем изображение с поверхности коры мозжечка в сочетании с селективной трансдукцией GCaMP6f клетками Пуркинье, мы уверены, что сообщаем о сигналах от отдельных дендритов клеток Пуркинье.
https://doi.org/10.7554/eLife.
49987.sa2
Статья и информация об авторе
Сведения об авторе
-
Андрес де Гроот
Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, НидерландыВклад
Концептуализация, Ресурсы, Курирование данных, Программное обеспечение, Формальный анализ, Валидация, Методология, Написание — первоначальный проект, Написание — просмотр и редактированиеКонкурирующие интересы
Конкурирующие интересы не заявлены -
Bastijn JG van den Boom
- Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, Нидерланды
- Кафедра психиатрии, Амстердамский университет UMC, Амстердамский университет, Амстердам, Нидерланды
Пожертвование
Концептуализация, Курирование данных, Формальный анализ, Проверка, Исследование, Визуализация, Написание — первоначальный проект, Написание — просмотр и редактированиеКонкурирующие интересы
Конкурирующие интересы не заявлены «Этот ORCID iD идентифицирует автора этой статьи:» 0000-0002-0853-3763 -
Романо М.
ван Гендерен - Факультет прикладных наук Делфтского технического университета, Делфт, Нидерланды
- Департамент неврологии, Erasmus MC, Роттердам, Нидерланды
Пожертвование
Концептуализация, Ресурсы, Программное обеспечение, Формальный анализ, Методология, Написание — обзор и редактированиеКонкурирующие интересы
Конкурирующие интересы не заявлены -
Йорис Коппенс
Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, НидерландыВклад
Концептуализация, Формальный анализ, Валидация, Методология, Написание — обзор и редактированиеКонкурирующие интересы
Конкурирующие интересы не заявлены -
Джон ван Вельдхуйзен
Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, НидерландыВклад
Концептуализация, ресурсы, методологияКонкурирующие интересы
Конкурирующие интересы не заявлены -
Юп Бос
Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, НидерландыВклад
Концептуализация, ресурсы, методологияКонкурирующие интересы
Конкурирующие интересы не заявлены -
Хьюго Худемакер
Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, НидерландыВклад
Ресурсы, проверка, исследование, методология, написание — обзор и редактированиеКонкурирующие интересы
Конкурирующие интересы не заявлены -
Марио Негрелло
- Факультет прикладных наук Делфтского технического университета, Делфт, Нидерланды
- Департамент неврологии, Erasmus MC, Роттердам, Нидерланды
Пожертвование
Супервизия, методология, написание — обзор и редактированиеКонкурирующие интересы
Конкурирующие интересы не заявлены «Этот ORCID iD идентифицирует автора этой статьи:» 0000-0002-8527-4259 -
Ingo Willuhn
- Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, Нидерланды
- Кафедра психиатрии, Амстердамский университет UMC, Амстердамский университет, Амстердам, Нидерланды
Пожертвование
Надзор, получение финансирования, написание — обзор и редактированиеКонкурирующие интересы
Конкурирующие интересы не заявлены «Этот ORCID iD идентифицирует автора этой статьи:» 0000-0001-6540-6894 -
Chris I De Zeeuw
- Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, Нидерланды
- Департамент неврологии, Erasmus MC, Роттердам, Нидерланды
Пожертвование
Концептуализация, Привлечение финансирования, Написание — обзор и редактированиеДля корреспонденции
c. [email protected]
Конкурирующие интересы
Конкурирующие интересы не заявлены «Этот ORCID iD идентифицирует автора этой статьи:» 0000-0001-5628-8187 -
Тихо М Хугланд
- Нидерландский институт неврологии, Королевская нидерландская академия искусств и наук, Амстердам, Нидерланды
- Департамент неврологии, Erasmus MC, Роттердам, Нидерланды
Пожертвование
Концептуализация, Ресурсы, Курирование данных, Формальный анализ, Надзор, Получение финансирования, Валидация, Исследование, Визуализация, Методология, Написание — первоначальный проект, Администрирование проекта, Написание — обзор и редактированиеДля корреспонденции
[email protected]Конкурирующие интересы
Конкурирующие интересы не заявлены «Этот ORCID iD идентифицирует автора этой статьи:» 0000-0002-7444-9279
Финансирование
Koninklijke Nederlandse Akademie van Wetenschappen (240-840100)
- Крис И Де Зеув
- Тихо М Хугланд
h3020 Европейский исследовательский совет (ERC-2014-STG 638013)
- Инго Виллун
Nederlandse Organisatie voor Wetenschappelijk Onderzoek (2015/06367/ALW 864.
14.010)
- Инго Виллун
Nederlandse Organisatie voor Wetenschappelijk Onderzoek (ALWOP.2015.076)
- Крис И Де Зеув
- Тихо М Хугланд
h3020 Европейский исследовательский совет (ERC-adv ERC-POC)
- Крис И Де Зеув
Topsector Life Sciences & Health (LSH) (LSHM18001)
- Тихо М Хугланд
Спонсоры не участвовали в разработке исследования, сборе и интерпретации данных или принятии решения о представлении работы для публикации.
Этика
Эксперименты на животных: Все проведенные эксперименты были лицензированы Компетентным органом Нидерландов и одобрены местным органом по защите животных в соответствии с Европейскими рекомендациями по уходу и использованию лабораторных животных Директивой 2010/63/ЕС.
Старший редактор
- Кейт М Вассум, Калифорнийский университет, Лос-Анджелес, США
Редактор-рецензент
- Меган Р. Кэри, Фонд Шампалимо, Португалия
Рецензент
- Дениз Дж. Кай, Медицинская школа Маунт-Синай, США
История публикаций
- Получено: 10 июля 2019 г.
- Принято: 13 января 2020 г.
- Принятая рукопись опубликована: 14 января 2020 г. (версия 1)
- Версия записи опубликована: 29 января 2020 г. (версия 2)
Авторское право
© 2020, de Groot et al.
Эта статья распространяется в соответствии с лицензией Creative Commons Attribution License, которая разрешает неограниченное использование и распространение при условии указания оригинального автора и источника.
Метрики
Число цитирований статей, полученное путем опроса самых высоких значений из следующих источников: Scopus, Crossref, PubMed Central.
Ссылки для скачивания
Список ссылок, состоящий из двух частей, для загрузки статьи или частей статьи в различных форматах.
Открытые цитаты (ссылки для открытия цитат из этой статьи в различных онлайн-сервисах управления ссылками)
- Менделей
- ЧитатьКуб»>
Процитировать эту статью (ссылки для загрузки цитат из этой статьи в форматах, совместимых с различными инструментами управления ссылками)
- Андрес де Гроот
- Bastijn JG ван ден Бум
- Романо М ван Гендерен
- Йорис Коппенс
- Джон ван Вельдхуйзен
- Юп Бос
- Хьюго Худемакер
- Марио Негрелло
- Инго Вилун
- Крис I Де Зеув
- Тихо М Хугланд
(2020)
NINscope, универсальный минископ для исследования цепей в нескольких регионах
eLife 9 :e49987.
https://doi.org/10.7554/eLife.49987
- Скачать БибТекс
- Скачать .RIS
Категории и теги
- Инструменты и ресурсы
- Неврология
- минископ
- мозжечок
- кора головного мозга
- полосатое тело
- мышь
- визуализация
Исследовательский организм
- Мышь
исследований цепей с помощью миниатюрных микроскопов с открытым исходным кодом: прошлое, настоящее и будущее
Введение
За последние 5 лет мы стали свидетелями шквала разработки с открытым исходным кодом миниатюрных флуоресцентных микроскопов (минископов) для нейробиологических приложений, дальнейшего совершенствования уже существующих технологий и расширения доступа к гораздо более широкой базе научных пользователей (Cai et al.
., 2016; Либерти и др., 2016, 2017; Джейкоб и др., 2018b; Джуно и др., 2018; Лян и др., 2018; Скотт и др., 2018; Скочек и др., 2018; Чжан и др. др., 2018). Открытый исходный код текущих проектов miniscope позволяет быстро обмениваться проектами, модификациями и идеями в нейробиологическом сообществе. Это привело к резкому ускорению инноваций, в результате чего появились минископы, способные контролировать активность клеточных популяций при отображении нейронной активности свободно движущихся позвоночных, различая отдельные клеточные популяции с использованием двухцветных версий (Jacob et al., 2018b; Leman et al. ., 2018), картирование больших полей зрения (Leman et al., 2018), визуализация объемов с частотой кадров видео (Skocek et al., 2018) и одновременная запись вокализации животных (Liberti et al., 2017) или ускорение головы (De Groot et al., 2018). В этой обзорной статье мы обобщаем и сравниваем различные текущие разработки с открытым исходным кодом и их потенциал для нейробиологических исследований. Основной движущей силой разработки миниатюрных микроскопов является, прежде всего, их способность регистрировать активность многих нейронов с определенной топологией у животного, которое ничем не ограничено и может демонстрировать свое естественное врожденное поведение. Факторы, влияющие на поведение, такие как повышенный уровень стресса у животных с фиксированной головой и отсутствие вестибулярного сигнала, вероятно, влияют на зарегистрированные паттерны нейронной активности и, следовательно, на выводы, которые можно сделать о функционировании мозга в более естественных состояниях (Терли и Аяз, 2017). Кроме того, минископы дают представление о нейронной активности, лежащей в основе поведения, в большом количестве поведенческих тестов, разработанных за последние десятилетия (Моррис, 19).84; Графф и др., 1998; Надлер и др., 2004; Гомес-Марин и др., 2014). В результате для понимания того, как активность, зарегистрированная в целевых областях, может быть коррелирована с поведением. Кроме того, возможность записи из более чем одного региона либо путем выборки из более крупных областей изображения, либо с использованием нескольких устройств визуализации с меньшими размерами и весом (De Groot et al. , 2018) должна показать, как межрегиональная передача сигналов проявляется в течение более длительного времени. естественное поведение. Миниатюрные микроскопы с однофотонным возбуждением
Было испробовано и протестировано множество подходов к визуализации нейронной активности свободно ведущих себя животных. Многие из первоначальных разработанных систем были основаны на оптических волокнах (Helmchen et al., 2001; Helmchen, 2002; Göbel et al., 2004; Flusberg et al., 2005, 2008) — с вводом возбуждающего света и сбором флуоресцентного света. из корпуса микроскопа — до разработки миниатюрного микроскопа с креплением на голову, в котором возбуждение и детектирование флуоресценции были объединены на корпусе микроскопа (Ghosh et al., 2011). Компоненты такого миниатюрного микроскопа преодолели стоимостные ограничения настольных лазеров и дорогих детекторов и вместо этого использовали относительно доступные технологии, такие как датчики изображения CMOS, светодиоды в качестве источника света, стандартные оптические компоненты и градиентный показатель преломления.
(или GRIN) линзы. Типичная конструкция такого миниатюрного микроскопа показана на рис. 1. Оптический путь очень похож на оптический путь обычного широкопольного флуоресцентного микроскопа, с заметным отличием в использовании одной или нескольких линз GRIN. Линзы GRIN обеспечивают оптический интерфейс с мозгом благодаря короткому рабочему расстоянию, плоскому дну и различным длинам и диаметрам. Для поверхностной визуализации головного мозга однообъективная линза GRIN помещается непосредственно на поверхность мозга. Для доступа к более глубоким областям объективная линза GRIN комбинируется с релейной линзой GRIN меньшего диаметра, имплантируемой над интересующими нейронами, как правило, за счет более ограниченного поля зрения. По сравнению с двухфотонным возбуждением однофотонное возбуждение особенно восприимчиво к флуоресценции вне фокуса и плохому оптическому секционированию. Несмотря на этот очевидный недостаток, оказалось возможным извлекать сигналы от отдельных нейронов с помощью аналитических методов, включая комбинацию анализа основных и независимых компонентов (PCA/ICA) и недавней неотрицательной матричной факторизации с ограничениями (CNMF) с добавленным членом для смоделировать изменяющийся во времени фоновый сигнал флуоресценции (Mukamel et al. , 2009; Лу и др., 2018; Чжоу и др., 2018). Таким образом, миниатюрные флуоресцентные микроскопы с головным креплением используют дешевые компоненты, но при этом позволяют получать изображения с клеточным разрешением у бодрствующих животных. Доступность таких минископов у коммерческих поставщиков уже привела к множеству исследований, которые позволили лучше понять активность нейронных цепей, лежащую в основе последовательности действий, беспокойства, вокализации (у птиц), социальной памяти и сна (Markowitz et al., 2015; Okuyama). и др., 2016; Клаус и др., 2017; Чен и др., 2018; Хименес и др., 2018). Выпуск планов создания минископов с открытым исходным кодом привел к еще более широкому применению этой технологии в последние несколько лет и, вероятно, обеспечит консолидированную платформу для будущих итераций инструментов, чтобы понять, как топологически определенные паттерны активности в мозгу влияют на поведение. (Ахарони и др., 2019 г.). Более широкое применение минископов для визуализации нейронной активности бодрствующих животных было бы невозможно без достижений в разработке репортеров флуоресцентной активности. Например, производные генетически кодируемого белка-индикатора кальция (GECI) GCaMP (Chen et al., 2013) позволили осуществлять клеточно-специфическое нацеливание на сенсоры активности в сочетании с улучшенным обнаружением нейронной активности и субклеточным нацеливанием на такие сенсоры (Dana et al. ., 2016). Кроме того, предпринимаются шаги по разработке более чувствительных датчиков напряжения, которые могут быть нацелены на клеточную специфичность 9.0413 in vivo (Bando et al., 2019; Quicke et al., 2019). Также стали доступны флуоресцентные датчики для переходных процессов глутамата и дофамина, позволяющие визуализировать высвобождение нейротрансмиттеров у бодрствующих животных (Marvin et al., 2018; Patriarchi et al., 2018). Таким образом, различные аспекты функционирования схемного уровня теперь могут быть исследованы с помощью минископов во время ничем не ограниченного поведения. Существуют предостережения, связанные с использованием миниатюрных флуоресцентных микроскопов. Использование линз GRIN для доступа к более глубоко расположенным структурам требует введения линз в мозг, что связано с повреждением тканей, несмотря на некоторые признаки того, что осторожные хирургические процедуры могут ограничивать влияние на поведение — по крайней мере, у мышей (Lee et al. , 2016). ). В любом случае следует позаботиться о том, чтобы воспалительные реакции уменьшились после краниотомии для доступа к мозгу и установки линз GRIN (Bocarsly et al., 2015). Более того, из соображений стоимости, а также из соображений простоты использования в большинстве миниатюрных флуоресцентных микроскопов используется однофотонное возбуждение, что ограничивает оптические срезы и затрудняет извлечение сигналов из одного источника. Последнее требует постфактум алгоритмическое извлечение сигналов, включающее модели флуоресценции вне фокуса, которые сами по себе основаны на предположениях. Несмотря на эти очевидные недостатки, минископы и их версии с открытым исходным кодом открыли возможности для изучения больших ансамблей нейронов во время естественного поведения. Поскольку стоимость больше не является ограничением для проведения таких экспериментов, самое время представить обзор доступности и применимости проектов с открытым исходным кодом, которые сделали эту технологию доступной. Потребность в поведенческом картировании, прямом контроле активности на уровне цепи, а также в более детальном считывании субпопуляций нейронов в цепи побудила несколько лабораторий разработать новые версии минископов для удовлетворения этих потребностей. Поэтому мы не только обсудим существующие в настоящее время проекты с открытым исходным кодом, но и подробно расскажем о будущих итерациях и технологиях, направленных на улучшение и расширение общей функциональности минископов. Рисунок 1 . Миниатюрная конструкция микроскопа с однофотонным возбуждением. (A) Типичная конструкция миниатюрного микроскопа с однофотонным возбуждением, используемого в сочетании с линзами с градиентным показателем преломления (GRIN). Он состоит из возбуждающего светодиодного источника света, коллиматора света с полусферической линзой, возбуждающего фильтра (Exc. filter) и дихроичного зеркала для отражения возбуждающего света вниз к образцу и передачи излучаемой флуоресценции на детектор изображения.
Излучаемый свет фокусируется на датчике изображения CMOS с помощью ахроматической линзы после прохождения через эмиссионный фильтр (эмиссионный фильтр). Использование линз GRIN позволяет получать изображения как поверхностных (слева), так и глубоко расположенных (справа) структур головного мозга. Регулировка фокальной плоскости в образце достигается путем перемещения датчика изображения к ахроматической линзе или от нее. Для поверхностной визуализации линза объектива GRIN помещается непосредственно на поверхность мозга, для глубокой визуализации мозга линза объектива GRIN устанавливается внутри эндоскопа и комбинируется с более тонкой релейной линзой GRIN, которая имплантируется в мозг для получения изображения клеток, трансдуцированных с помощью репортер флуоресцентной активности (зеленые точки). (B) Минископ первого поколения Калифорнийского университета в Лос-Анджелесе с открытым исходным кодом, который устанавливается через опорную плиту на мышь на время сеанса записи. Мыши носят эти 3-граммовые минископы без какого-либо явного влияния на общее поведение, хотя версии с кабелем могут влиять на социальное взаимодействие с другими мышами. Масштабная линейка ~10 мм. Проекты минископов с открытым исходным кодом
Минископ Калифорнийского университета в Лос-Анджелесе
Несколько лабораторий занимались разработкой уникальных версий миниатюрных микроскопов (рис. 2), которые впоследствии были выпущены в общественное достояние. охват, более 400 лабораторий по всему миру создают и применяют эти устройства визуализации в своих исследованиях за последние 3 года. Это широкое распространение было достигнуто с помощью онлайн-документации и руководств по закупке деталей, сборке и экспериментальному применению, а также с помощью многочисленных личных семинаров (подробности см. на сайте: http://miniscope.org). Первое поколение минископов Калифорнийского университета в Лос-Анджелесе, которые впервые были использованы для демонстрации того, что воспоминания, сформированные близко во времени, демонстрируют большее перекрытие нейронных ансамблей CA1 (Cai et al.
, 2016), могут быть построены за небольшую часть стоимости их коммерческих аналогов. весит около 3 г, имеет поле зрения 700 на 450 мкм (752 на 480 пикселей, размер пикселя 6 мкм), позволяет получать изображения одноканальной флуоресценции с максимальной частотой 60 Гц и одновременный мониторинг поведения с помощью дополнительного USB-камера. Оптическая схема включает ахроматическую линзу (Edmund Optics 49-923), который фокусирует почти коллимированный свет от объектива GRIN на датчик CMOS и набор настраиваемых оптических фильтров (Chroma ET470/40x, T495lpxr, ET525/50 m). Корпус изготовлен на станке с ЧПУ из черного делрина, а специальная электроника используется для управления и считывания данных со светодиодного источника возбуждения (Luxeon LXML-PB01-0030) и датчика изображения CMOS (On Semiconductor MT9V032C12STM, динамический диапазон > 55 дБ). Этот датчик изображения CMOS был выбран из-за его относительно небольшого размера на момент проектирования, низкой стоимости, большого размера пикселя и доступности при небольшом количестве заказа. Кроме того, ранее было показано, что аналогичный датчик изображения CMOS способен разрешать флуоресцентную динамику GECI (Ghosh et al., 2011). Для поверхностной визуализации линза GRIN имплантируется поверх поверхности мозга. Металлическая опорная пластина, прикрепленная к черепу с помощью стоматологического цемента, позволяет установить минископ. Для визуализации более глубоких областей мозга может быть имплантирована вторая, более тонкая релейная линза GRIN. В этом случае линза объектива GRIN устанавливается внутри минископа и располагается над релейной линзой, чтобы сфокусировать клетки (рис. 1). Фокусировка происходит с помощью линейного ползунка, высота которого регулируется вручную, сохраняя ориентацию нейронов между сеансами визуализации. Уникальным для проекта UCLA Miniscope является кабель, соединяющий головной прицел и внешнее оборудование для сбора данных. Питание, связь и передача данных осуществляются по одному гибкому коаксиальному кабелю в сочетании с вспомогательным оборудованием (TI DS9). 0UB913A/DS90UB914A и фильтры Power-Over-Coax). Благодаря уменьшению общего диаметра кабеля до 0,3 мм (Molex 100065-0023) и совместимости с пассивными коммутаторами с низким крутящим моментом эта конструкция сводит к минимуму влияние кабелей на поведение животных. Рисунок 2 . Минископы с открытым исходным кодом выпущены в общественное достояние. (A) FinchScope (https://github.com/gardner-lab/FinchScope), изображение предоставлено: WA Liberti III. (B) miniScope (https://github.com/giovannibarbera/miniscope_v1.0). (C) UCLA Miniscope (http://www.miniscope.org). (D) CHEndoscope (https://github.com/jf-lab/chendoscope), изображение предоставлено: A. Jacob, лаборатория Josselyn.
Также была разработана беспроводная версия минископа UCLA (4,5 г), которая использовалась для регистрации клеток места CA1 во время навигации по лабиринту у мышей с эпилепсией (Shuman et al., 2018). Эта система включает в себя литий-полимерный аккумулятор (~ 1 г) для питания, карту MicroSD для локального хранения данных (~ 0,5 г) и энергоэффективный датчик изображения (E2V JADE EV76C454).
Чтобы сохранить низкое энергопотребление, данные записываются с разрешением 320 на 320 пикселей с использованием двухкратной подвыборки пикселей. Несмотря на то, что конструкция без проводов тяжелее, чем ее проводной аналог, использование конструкции без проводов привело к социальному поведению, сравнимому с поведением мышей без Miniscope, тогда как более легкая проводная версия на самом деле продемонстрировала снижение социальных исследований (Дополнительные данные в Шуман и др., 2018). ). FinchScope
Проект FinchScope, начатый в Бостонском университете (Liberti et al., 2017), привел к созданию микроскопа с открытым исходным кодом примерно в то же время, что и Miniscope Калифорнийского университета в Лос-Анджелесе. В этом проекте используются 3D-печатные детали с высоким разрешением (низкофлуоресцентные смолы FGPBLK01 и FGPBLK02 на настольном принтере Form 2) и дешевые готовые аппаратные компоненты, такие как интегрированная система камер с датчиком CMOS с микрофоном (MC900, третий глаз).
электроника, динамический диапазон: 48 дБ) и плата Arduino Mega для аппаратного управления. Он имеет поле зрения 800 на 600 мкм (640 на 480 пикселей) с использованием оптической схемы, аналогичной конструкции минископа UCLA первого поколения с линзой GRIN в качестве объектива и ахроматической линзой для фокусировки изображения на Датчик изображения CMOS. Датчик камеры позволяет получать данные с частотой 30 Гц, а использование компонентов, напечатанных на 3D-принтере, позволяет быстро создавать прототипы и создавать более легкие конструкции с меньшими затратами по сравнению с машинными деталями. Этот FinchScope включает в себя турель с резьбой для фокусировки, весит примерно 1,8 г при использовании в проводной конфигурации и может быть объединен со специально разработанным активным коммутатором, который позволяет проводить продольные наблюдения за пением птиц у зебровых вьюрков. Программное обеспечение обеспечивает обратную связь с малой задержкой для запуска записи во время вокализации птиц и сыграло решающую роль в открытии нейронной динамики, лежащей в основе стабильных двигательных паттернов (Liberti et al. , 2016). Благодаря дополнительному беспроводному передатчику весом 2 г и литий-полимерному аккумулятору FinchScope может осуществлять запись и передачу данных в полностью беспроводном режиме. Дополнительный вес благоприятствует его применению на более крупных животных, способных нести дополнительный вес, и разработчики заявили об успехе, используя беспроводную версию на мышах. miniScope
Проект miniScope, разработанный в Национальном институте по борьбе со злоупотреблением наркотиками (NIDA), представляет собой минископ массой 2,4 г, который использовался для изучения прямого и непрямого пути прохождения полосатого тела с целью изучения того, как активность коррелирует с текущим двигательным поведением (Barbera et al. ., 2016), а также то, как паттерны активности в медиальной префронтальной коре связаны с социальной значимостью и новизной поведения (Liang et al., 2018; Zhang et al., 2018). Корпус miniScope был напечатан на 3D-принтере с использованием прочного, легкого и светонепроницаемого материала SLArmor с никелированным покрытием (Protolabs).
Как и у FinchScope, у miniScope есть резьбовая головка для фокусировки. В нем используется ахроматическая дуплетная линза для фокусировки на CMOS-датчик изображения и асферическая линза в сочетании с релейной линзой GRIN для достижения глубоких целей. Поле зрения большое 1,1 на 1,1 мм и проецируется на поверхность 400 на 400 пикселей стандартного датчика CMOS (MT9V022IA7ATM, для полупроводников, динамический диапазон > 55 дБ) с частотой кадров 10 Гц. Сбор данных происходит через плату Opal Kelly с программируемой вентильной матрицей (FPGA), которая обеспечивает быстрое управление считыванием видеоданных и управление светодиодным источником света возбуждения. CHEndoscope
В Университете Торонто был разработан минископ под названием CHendoscope. Этот минископ имеет корпус, который можно распечатать на 3D-принтере (Jacob et al., 2018b), и используется с 5-мегапиксельным интегрированным модулем камеры весом 1,5 г (MU9).PC-MBRD, Ximea, динамический диапазон: 38 дБ), доведя общий вес прицела до 4,5 г.
Для достижения частоты дискретизации 20 Гц используется объединение пикселей для достижения эффективной области изображения 648 на 486 пикселей, что соответствует полю зрения примерно 500 мкм в поперечнике. Фокусировка осуществляется с помощью турели с резьбой, как в FinchScope и miniScope. Источник питания минископа необходимо регулировать вручную, в то время как видеоданные собираются через программный интерфейс Python, установленный на стандартном ПК. Простота конструкции, в которой используется коммерчески доступное оборудование для обработки изображений, такое как FinchScope, делает его привлекательным вариантом для исследователей, которые предпочитают использовать готовые компоненты, а не изготовленные на заказ печатные платы (PCB). Описанные выше минископы отличаются простотой сборки, форм-фактором и функциональностью, удовлетворяя особые потребности исследователей. Из-за того, что эти проекты имеют открытый исходный код и их относительно простые компоненты, это не является серьезной проблемой, поскольку модификации могут быть легко реализованы, что сокращает цикл разработки.
Ниже мы сосредоточимся на некоторых способах дальнейшего улучшения функциональности минископа. Улучшение функциональности минископов с открытым исходным кодом
Линзы электросмачивания
Линзы электросмачивания (EWL, также известные как жидкие линзы) состоят из поверхности раздела двух несмешивающихся жидкостей (обычно воды и масла), которые под напряжением деформируются и действуют как линза, позволяющая осуществлять электронную фокусировку без необходимости ручной регулировки (Рисунок 3). Это особенно важно для лонгитюдных исследований, когда экспериментатору необходимо восстановить ранее использовавшуюся фокальную плоскость, а один и тот же минископ используется либо для нескольких животных, либо для разных областей одного и того же животного. Более того, снижение потребности в механическом обращении с чувствительными компонентами минископа обеспечивает более длительный срок хранения. Дополнительным преимуществом является то, что оптическая мощность EWL может быть быстро отрегулирована в пределах типичной частоты кадров, используемой в минископах (30 Гц), что можно использовать, если нужно выполнить запись с чередованием из разных фокальных плоскостей.
Коммерческие поставщики начали внедрять электронную фокусировку в свои минископы, а миниатюризация и доступность этих линз (например, Varioptic Arctic 16F0) позволили использовать их в минископах с открытым исходным кодом. Например, установленный на голове микроскоп с двухфотонным возбуждением и оптоволоконной связью включает EWL, позволяющую проводить многоплоскостные записи у мышей без ограничений (Ozbay et al., 2018), и минископы на основе однофотонного возбуждения последуют этому примеру. Рисунок 3 . Электросмачивание линз (EWL). (A) Изображение объекта, сфокусированного с помощью EWL. (B) Имеющиеся в продаже миниатюрные EWL весят всего 0,2 г с внешним диаметром ~6 мм. (C) Пример использования EWL (Varioptic Arctic 25H0) для фокусировки на испытательной цели ВВС США 1951 года со сдвигом фокусного расстояния ±200 мкм.
Двухцветная флуоресценция
С появлением генетически закодированных репортеров активности, которые флуоресцируют в спектрально разделяемых длинах волн (Chen et al.
, 2013; Dana et al., 2016), становится возможным нацеливать отдельные клеточные популяции для визуализации, обеспечивая понимание того, как динамика двух популяций нейронов коррелирует с конкретным поведением (Jennings et al., 2019). Одним из способов реализации двухцветного изображения в минископе является добавление дополнительного дихроичного зеркала на пути излучения для разделения флуоресцентного света от двух флуорофоров на два детектора, которые могут располагаться независимо друг от друга. Это решение, реализованное коммерческим поставщиком миниатюрных микроскопов, позволяет обойти проблему хроматических аберраций, присущую объективам GRIN. Такие аберрации вносят фокусный сдвиг (до ~ 100 мкм для линз GRIN большего диаметра и в зависимости от оптической конфигурации), что приводит к выборке флуоресценции с разными длинами волн из разных фокальных плоскостей (рис. 4). В качестве альтернативы для коррекции аберраций можно использовать оптические элементы, либо располагая плосковыпуклую линзу непосредственно за укороченной линзой GRIN (Leiner and Prescott, 19). 83), полностью заменив объектив GRIN ахроматическим набором линз или используя дифракционные оптические элементы (требующие специальной оптики). Одновременное двухцветное изображение также может быть получено с использованием одного датчика изображения с модифицированным оптическим путем минископа. Два чередующихся источника света в сочетании с двумя полосовыми фильтрами и дихроичным фильтром используются для создания и объединения спектрально разделенного возбуждения. Двухдиапазонный дихроичный и эмиссионный фильтр используются на пути излучения для передачи испускаемой флуоресценции. Обнаружение различных флуоресцентных длин волн достигается за счет чередующихся считываний с одного датчика, синхронизированного с переменными источниками света. Датчики цветного изображения, которые используют фильтр Байера RGB, также могут использоваться для визуализации двух флуорофоров, но с компромиссом разрешения изображения для спектральной информации и часто требуют дополнительной автономной обработки для выделения различных длин волн из спектрально перекрывающейся записи RGB. Первые попытки разработки двухцветных минископов с открытым исходным кодом уже начались и привели к созданию двухцветных минископов с открытым исходным кодом с некоторой коррекцией хроматических аберраций (Jacob et al., 2018a; Leman et al., 2018). Хотя коррекция хроматических аберраций может быть реализована путем замены линзы GRIN объектива ахроматической оптикой, для доступа к более глубоким структурам может по-прежнему потребоваться релейная линза GRIN, которая вносит свои собственные аберрации. Рисунок 4 . Фокусное смещение, вызванное хроматическими аберрациями. Схема, показывающая смещение фокуса в стержневой линзе GRIN при отображении двух цветов.
Оптогенетика
Используя управляемые светом каналы, которые имеют минимально перекрывающиеся спектры с репортером флуоресцентной активности, можно создать минископ с одновременными возможностями нейровизуализации и манипулирования в поле зрения (Stamatakis et al., 2018 ). Этого можно добиться, используя двойной тракт возбуждения, аналогичный описанному в предыдущем разделе.
В зависимости от выбранного репортера опсина и флуоресцентной активности эти оптогенетические минископы могут уменьшить, но не полностью устранить перекрестные помехи между каналами. Например, Stamatakis et al. показали в своем коммерческом внедрении миниатюрного оптогенетического микроскопа, что источник изображения света ослабляет оптогенетическую реакцию, которая может быть вызвана в препарате для теста на срезе. Это продемонстрировало, что при настройке источника возбуждающего света для визуализации следует соблюдать осторожность, чтобы обеспечить хороший компромисс между сигналом и шумом и минимальным пересечением сигнала. Альтернативный подход заключается в стимуляции вне поля зрения изображения, что возможно путем имплантации оптических волокон или беспроводных имплантируемых µLED (Shin et al., 2017). Однако для этого требуется отдельный драйвер светодиодов, синхронизированный с минископом. Альтернативная реализация может использовать дополнительный светодиодный драйвер на самом минископе в сочетании с имплантатом для оптогенетической стимуляции (De Groot et al. , 2018). Wide-Field Imaging
До недавнего времени поля зрения существующих минископов с открытым исходным кодом с разрешением в одну ячейку расширялись до максимального заявленного поля зрения ~ 1 на 1 мм с использованием ахроматического объектива (Zhang и др., 2018). В области относительно небольших микроскопов, которые могут носить мыши, датчики изображения большего размера можно комбинировать с набором плосковыпуклых и двойных вогнутых линз для достижения большего (3 на 4 мм 2 ) поля зрения. вид (Leman et al., 2018). Для визуализации позвоночных животных крупнее мышей в Принстонском университете разработан cScope (33 г) 1 дает возможность значительно расширить поле зрения (до 7,8 мм × 4 мм) и исследовать паттерны внутрикортикальной активности во время свободного перемещения с разрешением менее одной клетки (Scott et al., 2018). Используя электронику обработки изображений и сбора данных UCLA Miniscope, большое поле зрения cScope было достигнуто за счет оптимизированной оптики макроскопа и сопоставления изображений, полученных при различных угловых положениях датчика изображения (рис.
5). В целом, заменив линзу объектива GRIN и выбрав линзы с коррекцией аберраций, можно получить более широкое поле зрения: тенденция, которую мы увидим в будущих минископах с открытым исходным кодом. Рисунок 5 . Макроскоп cScope разработан в Принстонском университете. Схема, показывающая оптику, используемую для увеличения поля зрения наголовного микроскопа. Схема предоставлена С.Ю. Тиберге, Принстонский университет.
Нейроваскулярное сопряжение
Гемодинамические сигналы искажают сигналы флуоресценции, отображаемые с помощью эпифлуоресцентного микроскопа. В дополнение к светодиодному источнику света 480 нм для флуоресцентной визуализации конструкция микроскопа cScope включает светодиодную подсветку (530 нм) для измерения собственного оптического сигнала (OIS). Последний обеспечивает меру оксигенации крови, которую можно использовать для коррекции флуоресцентного сигнала, возникающего в результате нервной активности. Таким образом, cScope можно использовать не только для мониторинга активности нейронов в больших областях мозга и вычитания гемодинамического сигнала, но и для получения информации о нервно-сосудистых связях.
Чтобы изучить это на мышах, 9Недавно был разработан минископ -g с системой облегчения веса, ограничивающей вес головы до 3 г (Senarathna et al., 2019). В этом микроскопе используется светодиод 452 нм для возбуждения флуоресценции, светодиод 570 нм для визуализации OIS и лазерный диод 680 нм для измерения поглощения дезоксигемоглобина. Набор оранжевых светодиодов используется для синхронизации сигналов прицела с периферийным оборудованием. Объемная визуализация
Принцип, лежащий в основе микроскопии светового поля, заключается в сборе угловой информации вместе с информацией о положении световых лучей, исходящих из объема (например, их интенсивности, направлении, длине волны или поляризации), и использовании вычислительных методов для восстановления объемной информации постфактум из информации, содержащейся в 2D-изображении (Cohen et al., 2014; Prevedel et al., 2016). В одной распространенной реализации микроскопии светового поля используется матрица микролинз (MLA) для фокусировки света, идущего под разными углами, на разные пиксели датчика изображения.
MLA был реализован в реконфигурированном минископе Калифорнийского университета в Лос-Анджелесе (MiniFLM) для достижения объема 700 на 600 на 360 мкм 3 , который можно было получить на частоте 16 Гц (Skocek et al., 2018). Процедура итеративного извлечения источника (Seed-Iterative Demixing, SID) была разработана специально для использования с визуализацией светового поля в рассеивающей ткани (Nöbauer et al., 2017) и может использоваться в сочетании с микроскопом MiniLFM для надежного извлечения сигналов от нейронов. разделены на 15 мкм и более. При использовании MiniFLM возникают некоторые сложности, учитывая, что сложнее исправить артефакты движения, а некоторая пространственная информация заменяется угловой информацией, но общее увеличение объема данных, которые могут быть собраны при отображении объемов с частотой кадров видео (например, в сочетании с клеточно-специфическое нацеливание на репортеры активности) может быть значительным по сравнению с широкоугольной визуализацией. Беспроводная и беспроводная запись
Были разработаны беспроводные (Liberti et al., 2017) и беспроводные (Shuman et al., 2018) минископы с открытым исходным кодом. Оба питаются от встроенных литий-полимерных аккумуляторов, которые значительно утяжеляют конструкцию микроскопа. Энергопотребление зависит от полосы пропускания данных и является одним из ограничивающих факторов при разработке миниатюрного беспроводного микроскопа из-за требований к размеру батареи и весу. Энергоэффективные датчики изображения CMOS можно использовать вместе с пиксельным биннингом или подвыборкой пикселей, чтобы минимизировать энергопотребление при сохранении поля зрения, сравнимого с проводными минископами. Большинство датчиков изображения, используемых в миниатюрных микроскопах, имеют более высокую плотность пикселей, чем требуется, по сравнению с расчетной функцией рассеяния точек для их оптического пути и уменьшенное пространственное разрешение из-за рассеяния в ткани. Таким образом, для увеличения времени записи при сохранении приемлемых требований к весу выбирается более низкое или пониженное разрешение датчика.
Кроме того, возможно беспроводное питание миниатюрной конструкции с низким энергопотреблением благодаря беспроводной передаче энергии в ближнем поле, подобно тем, которые уже были реализованы для беспроводной оптогенетической стимуляции (Montgomery et al., 2015; Park et al., 2015; Shin). и др., 2017). Электрофизиология
В рамках проекта UCLA Miniscope разрабатываются различные реализации минископов, которые поддерживают одновременную внеклеточную запись и визуализацию. Это может быть достигнуто с помощью автономной системы записи ephys, синхронизированной с минископом, или путем полного включения оборудования для записи ephys в систему Miniscope с использованием чипов цифрового интерфейса электрофизиологии (например, Intan RHD2132/64). Для крыс в UCLA 9 разработана сборка для записи микродисков до 18 каналов (OvalDrive 18-ES).0093 2 , что позволяет одновременно проводить электрофизиологические записи или вольтамперометрию и визуализацию у крыс. Кроме того, канюли для инъекций жидкости и оптические волокна также могут быть вставлены, что позволяет манипулировать активностью во время визуализации с помощью минископа у свободно движущихся крыс.
Считывание данных о поведении
Записи с минископа можно комбинировать с камерами, записывающими поведение животных, но хранение, обработка и аннотирование таких наборов данных о поведении часто являются нетривиальными и требуют много времени. Фреймворк для обработки потоков данных под названием Bonsai (Lopes et al., 2015) помогает автоматизировать части такого анализа, и был разработан модуль для поддержки UCLA Miniscope 9.0093 3 . Альтернативный способ регистрировать поведенческое состояние животного — интегрировать инерциальный измерительный блок (IMU) в минископ для измерения трехмерного ускорения и ориентации головы (De Groot et al., 2018). Поведение, связанное с уходом за собой, приемом пищи и воспитанием детей, связано с отчетливыми моделями ускорения, которые можно использовать для классификации поведения (Venkatraman et al., 2010; Pasquet et al., 2016; Wilson et al., 2018). Для более сложных систем отслеживания поведения использовались многокамерные системы, например, для отслеживания трехмерной осанки (Mimica et al.
, 2018), но для этого требуются значительные ресурсы, которые могут быть недоступны для всех лабораторий. Другая система отслеживания (3D Tracker) 4 использует четыре камеры измерения глубины RealSense TM , относительно недорог и эффективен для отслеживания позы и социальных взаимодействий у крыс, но требует значительной постобработки (Matsumoto et al., 2013; Nakamura et al., 2016). ). Достижения в области машинного обучения приведут к автоматизированному анализу поведенческих мотивов, зафиксированных в данных камеры, например, к извлечению временной структуры позы животного (Wiltschko et al., 2015; Pereira et al., 2019) или к безмаркерному отслеживанию животных ( Матис и др., 2018). Open Ephys, которая разрабатывает инструменты с открытым исходным кодом для нейронауки 5 , выступила с решением для отслеживания поведения с использованием систем отслеживания, изначально разработанных для потребительских игр: отслеживание Valve SteamVR TM . Он использует ортогональные лазерные излучатели на базовой станции, которые просматривают пространство (два развертки 2pi/360°), а также фотодиоды и IMU на отслеживаемом объекте (например, минископ) для получения информации об ориентации отслеживаемых объектов, скорости и угловой скорости. в настоящее время. Преимущество этой системы в том, что она не требует такого же объема постобработки, как в вышеупомянутых системах, доступна по цене и может быть интегрирована с другими периферийными устройствами Open Ephys. Двухфотонное возбуждение
Двухфотонное возбуждение обладает хорошо известными преимуществами по сравнению с однофотонным возбуждением, включая меньший объем возбуждения, повышенное проникновение в ткани и снижение фототоксичности благодаря использованию более длинных волн возбуждения. Небольшой фемтолитровый объем возбуждения значительно улучшает оптическое сечение образца и, таким образом, гарантирует, что собранный свет исходит только от клеточных и субклеточных структур, расположенных вдоль фокальной плоскости. Первая задокументированная попытка построить миниатюрный микроскоп с двухфотонным возбуждением на основе волокон (Helmchen et al., 2001) продемонстрировала свою общую осуществимость, даже несмотря на то, что во время поведения не было получено функционального изображения нейронов, отчасти из-за сильных артефактов движения.
Микроскоп также был сравнительно тяжелым и большим (25 г, высота 70 мм), что ограничивало возможности его применения, связанные с естественным поведением мелких животных. Первоначально пьезоэлектрический элемент использовался для возбуждения вибраций в усиленном волокне для создания схемы сканирования Лиссажу (Helmchen, 2002; Flusberg et al., 2005, 2008). Дальнейшие усовершенствования, основанные на аналогичной конструкции, привели к значительно более легким портативным микроскопам с двухфотонным возбуждением (Göbel et al., 2004; Flusberg et al., 2008; Sawinski et al., 2009).), используемые для визуализации свободно движущихся мышей и крыс (см. также Ozbay et al., 2018). Также были разработаны альтернативные схемы сканирования, основанные на зеркалах микроэлектромеханических систем (МЭМС) (Piyawattanametha et al., 2006; Zong et al., 2017). Среди них портативный сканирующий микроскоп с двухфотонным возбуждением весом около 2 г использовался для изучения энторинальной коры у свободно ведущих себя мышей (Zong et al. , 2017; Obenhaus et al., 2018). Дорогие настольные лазеры используются для двухфотонного возбуждения и соединяются в одно оптическое волокно, а флуоресценция собирается через оптическое волокно для обнаружения. Стоимость высокочастотного импульсного лазера и других периферийных устройств (например, фотоумножителей, изготовленных на заказ оптических волокон для повышения эффективности сбора данных и гибкости) может ограничивать повсеместное использование двухфотонных микроскопов по сравнению с миниатюрными микроскопами с однофотонным возбуждением. Кроме того, недавнее всестороннее исследование двухфотонного и однофотонного возбуждения визуализации кальция с использованием, соответственно, настольного двухфотонного и миниатюрного однофотонного микроскопа показало, что настройка ориентации одних и тех же идентифицированных нейронов была сопоставима независимо от типа используемого флуоресцентного возбуждения. (Глас и др., 2018). Таким образом, несмотря на ключевые преимущества микроскопии с двухфотонным возбуждением, миниатюрные флуоресцентные микроскопы с однофотонным возбуждением остаются полезными и доступными инструментами для визуализации клеток с разрешением во время неограниченного поведения. В еще большей степени это относится к высокоподвижным животным, таким как птицы и летучие мыши, у которых волоконная связь возбуждающего и испускаемого света нецелесообразна. Анализ данных
Мозг животных движется во время свободного передвижения, а это означает, что перед извлечением сигнала требуется коррекция движения. Хотя были реализованы различные типы алгоритмов коррекции движения (Dombeck et al., 2007; Greenberg and Kerr, 2009; Dubbs et al., 2016; Mitani and Komiyama, 2018), кусочно-жесткая реализация 6 адаптирована для флуоресцентных эндоскопических данных. оказался особенно эффективным при исправлении нежестких деформаций (Pnevmatikakis and Giovannucci, 2017). Более поздний подход, использующий иерархическую структуру регистрации видео, чтобы отличать стабильные кадры от кадров с большими межкадровыми сдвигами, может обеспечить еще более надежную регистрацию изображений (Lu et al., 2018). После того, как кадры зарегистрированы, необходимо извлечь и разделить сигналы из обнаруженных ячеек.
Первоначально подход PCA/ICA использовался для данных, полученных с помощью миниатюрных флуоресцентных микроскопов (Mukamel et al., 2009).), но этот подход в основном был заменен более точными методами, основанными на CNMF, с добавленной моделью для оценки и учета изменяющихся во времени фоновых флуктуаций флуоресценции под названием CNMF-E (Zhou et al., 2018). Было высказано предположение, что, несмотря на точность обнаружения клеток, алгоритм CNMF-E 7 может иметь более высокий уровень ложных срабатываний, чем подход MIN1PIPE 8 , разработанный Lu et al. (2018), в котором используется альтернативный метод для устранения фоновых колебаний флуоресценции и усиления нейронных сигналов. Выходной сигнал обычно представляет собой масштабированную версию частичного изменения флуоресценции, которую можно нормализовать путем деления на стандартное отклонение или базовую оценку шума. Деконволюция необработанного сигнала часто выполняется для устранения шума в сигнале и позволяет оценить время события, которое отражает лежащую в основе нейронную активность. Будущие разработки
Сообщество разработчиков минископов с открытым исходным кодом находится на пороге выпуска широкого спектра минископов второго поколения, которые включают ETL для фокусировки, обнаружения двухцветной флуоресценции, большего поля зрения, меньшей занимаемой площади и оптогенетических и возможности считывания поведения. Какие разработки ждут впереди?
Эксперименты с обратной связью
Доступность плат ПЛИС (Ciliberti and Kloosterman, 2017; Siegle et al., 2017; Buccino et al., 2018) позволяет проводить быстрые эксперименты с обратной связью (Ahrens et al., 2012; Clancy et al., 2014; Packer et al., 2014; Prsa et al., 2017), предоставляя информацию, например, о сенсомоторном и протезном обучении в нейронных цепях, которые важны для разработки интерфейсов мозг-машина. Обнаружение в реальном времени определенного типа поведения или модели активности можно использовать для прямого управления входными путями или целевыми клеточными сборками. Затем можно было бы исследовать клеточные субстраты, лежащие в основе выражения поведения или взаимодействия между мозгом, телом и окружающей средой (Buckley and Toyoizumi, 2018).
Замкнутые системы также можно использовать для обнаружения и подавления развития эпилептиформной активности (Sorokin et al., 2017) и определения клеточных механизмов, лежащих в основе эффективной глубокой стимуляции мозга (Parastarfeizabadi and Kouzani, 2017; Ghasemi et al., 2018). . Платформы с открытым исходным кодом, такие как Open Ephys, уже разработали аппаратную интеграцию FPGA для экспериментов с замкнутым контуром с малой задержкой (Siegle et al., 2017), и такие возможности замкнутого цикла могут быть перенесены на минископы с открытым исходным кодом. Учитывая расширение функциональности минископа и сопутствующее увеличение количества каналов данных, которые необходимо обрабатывать, использование FPGA упростит обработку данных и управление ими. Благодаря минископам, позволяющим получать объемные изображения, и необходимости демультиплексирования сигналов, такая технология также может способствовать эффективному извлечению данных. Custom Optics
Использование набора ахроматических линз вместо объектива GRIN помогает уменьшить, но не полностью устранить хроматические аберрации в большинстве конфигураций двухцветных минископов.
Одним из решений является использование дифракционного элемента или аксикона, который исправляет хроматическую аберрацию на оптическом пути. Нанолитография позволяет печатать на 3D-принтере специальные оптические элементы, в том числе те, которые корректируют хроматические аберрации (Schmid et al., 2018), коллимируют свет (Thiele et al., 2016) и действуют как линзы с высоким разрешением (Gissibl et al., 2016). ). Миниатюризация этих оптических компонентов в сочетании с малогабаритным КМОП-датчиком может привести к значительно более легким и компактным минископам, позволяющим вести одновременную запись из нескольких областей с использованием более одного минископа, устанавливаемого на голову. Более того, резонансные плазмонные метаповерхности в сочетании с материалами с фазовым переходом можно использовать для создания миниатюрных нанооптических элементов, которые действуют как бифокальные зум-линзы и могут управлять светом (Yin et al., 2017). Трехмерная печать оптики по-прежнему зависит от дорогих двухфотонных лазеров, которые наращивают компоненты слой за слоем в процессе трудоемкого процесса, что ограничивает их широкое распространение в минископах с открытым исходным кодом. Тем не менее, ряд академических учреждений начали предоставлять платную инфраструктуру нанолитографии, которая могла бы сделать эту технологию достаточно распространенной, чтобы начать интеграцию 3D-печатных оптических элементов в минископы. Более того, после того, как окончательная оптическая конструкция, напечатанная на 3D-принтере, будет протестирована, изготовленную по индивидуальному заказу оптику, вдохновленную этими конструкциями, можно будет заказать через специализированных подрядчиков по микрооптике. Безлинзовая визуализация
Важные успехи в дальнейшей миниатюризации устройств визуализации для флуоресцентной микроскопии были достигнуты путем замены фокусирующих элементов в минископах рассеивателем или маской на пути излучения флуоресценции (Adams et al., 2017; Antipa et al. , 2017). Диффузионный элемент с оптимизированной амплитудной маской в сочетании с вычислительным разделением сигналов позволяет реконструировать сигналы из объема на основе информации, содержащейся в 2D-изображениях (аналогично тому, что использовалось для визуализации светового поля с использованием MLA).
Безлинзовые минископы могут значительно уменьшить вес и занимаемую площадь головных систем визуализации, а также увеличить их поле зрения. Одно из предостережений в настоящее время заключается в том, что источники света должны быть интегрированы в систему, чтобы обеспечить возбуждение флуоресценции. В принципе, это можно сделать, расположив микросветодиоды вокруг датчика изображения или имплантировав оптические волноводы, которые могут создавать управляемые световые лучи и листы. Еще одно предостережение заключается в том, что, как и в случае с микроскопом MiniLFM, коррекцию движения может быть сложно реализовать. Тем не менее, идея безлинзового минископа является многообещающей и может привести к созданию гораздо более легких портативных флуоресцентных микроскопов с большим полем зрения. Сбор данных с высокой частотой кадров
Большинство минископов в настоящее время используются в сочетании с GECI, которые имеют — в дополнение к Ca 2+ , обеспечивающий косвенное измерение текущей активности — относительно медленную кинетику.
Таким образом, здесь обычно достаточно типичной частоты кадров 30 Гц. С быстрым улучшением генетически кодируемых индикаторов напряжения и их потенциала для клеточно-специфического нацеливания in vivo на (Flytzanis et al., 2014; Marshall et al., 2016; Piatkevich et al., 2018; Bando et al., 2019).; Quicke et al., 2019) становится актуальным достижение более высокой частоты кадров с помощью высокочувствительных датчиков изображения. Имеющиеся в настоящее время датчики изображения CMOS, достаточно малые для интеграции в минископы, часто могут достигать частоты кадров в несколько сотен Гц, что достаточно для наблюдения за колебаниями напряжения на уровне населения (Marshall et al., 2016) за счет комбинации подвыборки пикселей, объединения пикселей и уменьшения количество активных строк пикселей. Для достижения разрешения с одной ячейкой и с одним потенциалом действия, вероятно, потребуется разработать как более яркие индикаторы напряжения, так и меньшие по размеру научные КМОП-датчики с задней подсветкой. Учитывая дальнейшую миниатюризацию электроники, нагрев является проблемой, которую необходимо решить. Тепло можно рассеивать, обеспечивая достаточную конвекцию воздуха или через металлические радиаторы, оба из которых могут значительно увеличить вес конструкции минископа. Работа с более высокими частотами дискретизации потребляет больше энергии и, как правило, требует дополнительного рассеивания тепла, устанавливая ограничение на использование этих высоких частот кадров, особенно в беспроводных или беспроводных конфигурациях. Заключительные замечания
Разработка минископов с открытым исходным кодом дала большой толчок нейробиологам, которым нужны доступные, недорогие и понятные инструменты для визуализации мозга позвоночных во время естественного поведения. Проекты с открытым исходным кодом обычно имеют относительно простые планы проектирования с модульными компонентами, которые легко доступны или могут быть изготовлены на заказ с использованием 3D-принтеров. Низкая стоимость этих компонентов и сравнительная простота сборки таких прицелов привели к их широкому распространению.
Хотя многие из этих систем достаточно просты, чтобы их можно было довольно легко распространять и внедрять, остается проблемой масштабная интеграция более сложных, менее популярных технологий в минископы с открытым исходным кодом. К счастью, доступ к недорогим, высококачественным и малым партиям производственных линий для нестандартных печатных плат, оптики, станков с ЧПУ и литьевых деталей значительно вырос в последние годы и, вероятно, продолжится. Компоненты, которые были громоздкими и дорогими всего несколько лет назад, теперь миниатюризированы и доступны по цене, такие как EWL для быстрой электрической фокусировки, высокоскоростные CMOS-датчики с задней подсветкой и маломощные IMU, которые можно интегрировать в минископы для считывания ускорения головы. и ориентация. Следуя этой тенденции, даже такие технологии, как нанолитография, которые в настоящее время кажутся несколько экзотическими для проекта разработки с открытым исходным кодом, через несколько лет могут стать более обычным явлением. Возможность создавать легкие минископы небольшого размера будет способствовать благополучию животных и улучшит считывание активности нейронов в естественных условиях. Сообщество открытого исходного кода имеет силу в количестве, и постоянный открытый обмен новыми идеями приведет к улучшению минископов (например, чувствительности, поля зрения и экспериментального контроля с обратной связью), более широкому распространению и лучшему пониманию мозга. как он взаимодействует с окружающей средой. Вклад авторов
DA и TH подготовили рисунки и написали рукопись.
Финансирование
Работа выполнена при поддержке Health~Holland (LSHM18001-H001; TH) и NSF Neurotech Hub (1700408; DA).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
9 https://github. com/JinghaoLu/MIN1PIPE
Ссылки
Adams, J.K., Boominathan, V., Avants, B.W., Vercosa, D.G., Ye, F., Baraniuk, R.G., et al. (2017). Однокадровая трехмерная флуоресцентная микроскопия с ультраминиатюрным безлинзовым прибором FlatScope. Науч. Доп. 3:e1701548. doi: 10.1126/sciadv.1701548
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ахарони Д., Хах Б. С., Сильва А. Дж. и Гольшани П. (2019). Весь свет, который мы можем видеть: новая эра в миниатюрной микроскопии. Нац. Методы 16, 11–13. doi: 10.1038/s41592-018-0266-x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ahrens, M.B., Li, J.M., Orger, M.B., Robson, D.N., Schier, A.F., Engert, F., et al. (2012). Динамика нейронов всего мозга во время двигательной адаптации у рыбок данио. Природа 485, 471–477. doi: 10.1038/nature11057
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Антипа Н.
, Куо Г., Хеккель Р., Милденхолл Б., Бостан Э., Нг Р. и др. (2017). DiffuserCam: безлинзовое 3D-изображение с одной экспозицией. arXiv [препринт] 1710.02134 [cs.CV]. Доступно в Интернете по адресу: http://arxiv.org/abs/1710.02134 Google Scholar
Бандо Ю., Сакамото М., Ким С., Айзенштат И. и Юсте Р. (2019) . Сравнительная оценка генетически закодированных показателей напряжения. Cell Rep. 26, 802.e4–813.e4. doi: 10.1016/j.celrep.2018.12.088
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Barbera, G., Liang, B., Zhang, L., Gerfen, C.R., Culurciello, E., Chen, R., et al. (2016). Пространственно компактные нейронные кластеры в дорсальном полосатом теле кодируют информацию, относящуюся к передвижению. Нейрон 92, 202–213. doi: 10.1016/j.neuron.2016.08.037
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бокарсли, М. Э., Цзян, В.-К., Ван, К., Дадман, Дж. Т., Джи, Н., и Апонте, Ю.
(2015). Минимально инвазивная система микроэндоскопии для функциональной визуализации in vivo глубоких ядер мозга мыши. Биомед. Опц. Экспресс 6, 4546–4556. doi: 10.1364/boe.6.004546 PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Буччино, А. П., Лепперод, М. Э., Драгли, С.-А., Хефлигер, П., Фюн, М., и Хафтинг, Т. (2018). Модули с открытым исходным кодом для отслеживания поведения животных и стимуляции с обратной связью на основе Open Ephys и Bonsai. J. Нейронная инженерия. 15:055002. doi: 10.1088/1741-2552/aacf45
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бакли, К.Л., и Тойоидзуми, Т. (2018). Теория того, как активное поведение стабилизирует нейронную активность: модуляция нейронного усиления за счет обратной связи с окружающей средой с обратной связью. Вычисл. PLoS. биол. 14:e1005926. doi: 10.1371/journal.pcbi.1005926
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Cai, D.
J., Aharoni, D., Shuman, T., Shobe, J., Biane, J., Song, W., et al. (2016). Общий нейронный ансамбль связывает различные контекстуальные воспоминания, закодированные близко во времени. Природа 534, 115–118. doi: 10.1038/nature17955 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Chen, T.-W., Wardill, T.J., Sun, Y., Pulver, S.R., Renninger, S.L., Baohan, A., et al. (2013). Сверхчувствительные флуоресцентные белки для визуализации активности нейронов. Природа 499, 295–300. doi: 10.1038/nature12354
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Chen, K.-S., Xu, M., Zhang, Z., Chang, W.-C., Gaj, T., Schaffer, D.V., et al. (2018). Гипоталамический переключатель для быстрого и медленного сна. Нейрон 97, 1168.e4–1176.e4. doi: 10.1016/j.neuron.2018.02.005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чилиберти, Д., и Клоостерман, Ф. (2017). Falcon: очень гибкое программное обеспечение с открытым исходным кодом для нейробиологии с обратной связью.
J. Нейронная инженерия. 14:045004. doi: 10.1088/1741-2552/aa7526 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Клэнси, К.Б., Коралек, А.С., Коста, Р.М., Фельдман, Д.Е., и Кармена, Дж.М. (2014). Волевая модуляция оптически регистрируемых сигналов кальция при нейропротезном обучении. Нац. Неврологи. 17, 807–809. doi: 10.1038/nn.3712
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Коэн Н., Ян С., Андалман А., Брокстон М., Гросеник Л., Дейссерот К. и др. (2014). Повышение производительности микроскопа светового поля с помощью кодирования волнового фронта. Опц. Экспресс 22, 24817–24839. doi: 10.1364/OE.22.024817
CrossRef Полный текст | Google Scholar
Дана Х., Мохар Б., Сун Ю., Нараян С., Гордус А., Хассеман Дж. П. и др. (2016). Чувствительные индикаторы кальция с красным белком для визуализации нервной активности. Элайф 5:e12727. doi: 10.7554/eLife.12727
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Де Гроот, А.
, Бос, Дж., Ван Вельдхуйзен, Дж., Коппенс, Дж., Ходемакер, Х., Де Зеув, К.И., и др. (2018). «Минископы уменьшенного размера для визуализации нескольких областей мозга во время естественного поведения», в Номер реферата: F18–1717. Fens Forum 2018 Берлин, Германия. Домбек, Д. А., Хаббаз, А. Н., Коллман, Ф., Адельман, Т. Л., и Танк, Д. В. (2007). Визуализация крупномасштабной нейронной активности с клеточным разрешением у бодрствующих подвижных мышей. Нейрон 56, 43–57. doi: 10.1016/j.neuron.2007.08.003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Даббс А., Гевара Дж. и Юсте Р. (2016). moco: быстрая коррекция движения для визуализации кальция. Перед. Нейроинформ. 10:6. doi: 10.3389/fninf.2016.00006
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Флюсберг Б. А., Юнг Дж. К., Кокер Э. Д., Андерсон Э. П. и Шнитцер М. Дж. (2005). Визуализация головного мозга in vivo с использованием портативного 3,9-граммового двухфотонного флуоресцентного микроэндоскопа.
Опц. лат. 30, 2272–2274. doi: 10.1364/ol.30.002272 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Flusberg, B.A., Nimmerjahn, A., Cocker, E.D., Mukamel, E.A., Barretto, R.P.J., Ko, T.H., et al. (2008). Высокоскоростная миниатюрная флуоресцентная микроскопия на свободно движущихся мышах. Нац. Методы 5, 935–938. doi: 10.1038/nmeth.1256
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Flytzanis, N.C., Bedbrook, C.N., Chiu, H., Engqvist, M.K.M., Xiao, C., Chan, K.Y., et al. (2014). Варианты архаэрродопсина с усиленной вольтаж-чувствительной флуоресценцией в нейронах млекопитающих и Caenorhabditis elegans . Нац. коммун. 5:4894. doi: 10.1038/ncomms5894
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Гасеми П., Сахраи Т. и Мохаммади А. (2018). Замкнутая и открытая глубокая стимуляция мозга: методы, проблемы, текущие и будущие аспекты. Дж. Биомед.
физ. англ. 8, 209–216. doi: 10.31661/jbpe.v8i2.898 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гош К.К., Бернс Л.Д., Кокер Э.Д., Ниммерьян А., Зив Ю., Гамаль А.Е. и др. (2011). Миниатюрная интеграция флуоресцентного микроскопа. Нац. Методы 8, 871–878. doi: 10.1038/nmeth.1694
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Гиссибль Т., Тиле С., Херкоммер А. и Гиссен Х. (2016). Двухфотонная прямая лазерная запись сверхкомпактных многолинзовых объективов. Нац. Фотоника 10, 554–560. doi: 10.1038/nphoton.2016.121
CrossRef Полный текст | Google Scholar
Glas, A., Huebener, M., Bonhoeffer, T., and Goltstein, P.M. (2018). Сравнение миниатюрной микроскопии с двухфотонной визуализацией кальция с использованием настройки ориентации одиночных клеток в зрительной коре мыши. bioRxiv [Препринт] 4
. doi: 10.1101/4 CrossRef Полный текст | Google Scholar
Гёбель В.
, Керр Дж. Н. Д., Ниммерьян А. и Хельмхен Ф. (2004). Миниатюрный двухфотонный микроскоп на основе гибкого пучка когерентных волокон и линзового объектива с градиентным показателем преломления. Опц. лат. 29, 2521–2523. doi: 10.1364/ol.29.002521 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гомес-Марин А., Патон Дж. Дж., Кампфф А. Р., Коста Р. М. и Майнен З. Ф. (2014). Большие поведенческие данные: психология, этология и основы нейронауки. Нац. Неврологи. 17, 1455–1462. doi: 10.1038/nn.3812
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Graeff, F.G., Netto, C.F., and Zangrossi, H. Jr. (1998). Приподнятый Т-образный лабиринт как экспериментальная модель тревоги. Неврологи. Биоповедение. Ред. 23, 237–246. doi: 10.1016/s0149-7634(98)00024-4
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гринберг, Д. С., и Керр, Дж. Н. Д. (2009). Автоматическая коррекция артефактов быстрого движения для двухфотонной визуализации бодрствующих животных.
J. Neurosci. Методы 176, 1–15. doi: 10.1016/j.jneumeth.2008.08.020 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хельмхен Ф. (2002). Миниатюризация флуоресцентных микроскопов с использованием оптоволокна. Экспл. Физиол. 87, 737–745. doi: 10.1113/eph8702478
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Helmchen, F., Fee, M.S., Tank, D.W., and Denk, W. (2001). Миниатюрный налобный двухфотонный микроскоп позволяет получать изображения головного мозга с высоким разрешением у свободно движущихся животных. Нейрон 31, 903–912. doi: 10.1016/S0896-6273(01)00421-4
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Джейкоб А. Д., Рамсаран А. И., Мокле А. Дж., Тран Л. М., Ян К., Франкленд П. В. и др. (2018а). Двухцветная система для визуализации популяций инграмм с кальцием in vivo. Программа № 511.05. Планировщик встреч по нейробиологии на 2018 год. Сан-Диего, Калифорния: Общество неврологии.
Джейкоб, А. Д., Рамсаран, А. И., Мокле, А. Дж., Тран, Л. М., Ян, К., Франкленд, П. В., и др. (2018б). Компактный эндоскоп с головным креплением на in vivo визуализация кальция у свободно ведущих себя мышей. Курс. протокол Неврологи. 84:e51. doi: 10.1002/cpns.51
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дженнингс Дж. Х., Ким С. К., Маршел Дж. Х., Раффи М., Йе Л., Куирин С. и др. (2019). Взаимодействие нейронных ансамблей в орбитофронтальной коре для социального и пищевого поведения. Природа 565, 645–649. doi: 10.1038/s41586-018-0866-8
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Хименес, Дж. К., Су, К., Голдберг, А. Р., Луна, В. М., Биан, Дж. С., Ордек, Г., и др. (2018). Тревожные клетки в гиппокампально-гипоталамическом контуре. Нейрон 97, 670.e6–683.e6. doi: 10.1016/j.neuron.2018.01.016
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Джуно, Дж.
, Дюре, Г., Робинсон, Дж., и Кемер, К. (2018). «Улучшенный модуль датчика изображения для головных микроскопов», в материалах Proceedings of the 40th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC) (IEEE: Гонолулу, Гавайи), 826–829. Google Scholar
Клаус А., Мартинс Г. Дж., Пайшао В. Б., Чжоу П., Панински Л. и Коста Р. М. (2017). Пространственно-временная организация полосатого тела кодирует пространство действия. Нейрон 95, 1171.e7–1180.e7. doi: 10.1016/j.neuron.2017.08.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ли С. А., Холли К. С., Возиянов В., Виллаба С. Л., Тонг Р., Григсби Х. Э. и др. (2016). Микролинза с градиентным индексом, имплантированная в префронтальную кору мышей, не влияет на выполнение поведенческих тестов с течением времени. PLoS ONE 11: e0146533. doi: 10.1371/journal.pone.0146533
CrossRef Полный текст | Google Scholar
Лейнер, Д.
К., и Прескотт, Р. (1983). Коррекция хроматических аберраций в эндоскопах GRIN. Заяв. Опц. 22, 383–386. doi: 10.1364/ao.22.000383 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Леман Д. П., Чен И. А., Йен В. В., Марковиц Дж. Э., Перкинс Л. Н., Либерти В. А. и др. (2018). Расширенный набор инструментов с открытым исходным кодом для визуализации кальция в широком поле у свободно ведущих себя животных. Программа № 338.20. Планировщик встреч по нейробиологии на 2018 год. Сан-Диего, Калифорния: Общество неврологии.
Лян, Б., Чжан, Л., Барбера, Г., Фанг, В., Чжан, Дж., Чен, X., и др. (2018). Отдельные и динамичные нейронные ансамбли ВКЛ и ВЫКЛ в префронтальной коре кодируют социальное исследование. Нейрон 100, 700.e9–714.e9. doi: 10.1016/j.neuron.2018.08.043
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Liberti, W. A. III., Markowitz, J. E., Perkins, L. N., Liberti, D. C., Leman, D.
P., Guitchounds, G., et al. (2016). Нестабильные нейроны лежат в основе стабильного усвоенного поведения. Нац. Неврологи. 19, 1665–1671. doi: 10.1038/nn.4405 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Либерти, В. А., Перкинс, Л. Н., Леман, Д. П., и Гарднер, Т. Дж. (2017). Миниатюрная микроскопическая система с открытым исходным кодом и беспроводной связью. J. Нейронная инженерия. 14:045001. doi: 10.1088/1741-2552/aa6806
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лопес Г., Бонакки Н., Фразао Дж., Нето Дж. П., Аталлах Б. В., Соареш С. и др. (2015). Бонсай: основанная на событиях структура для обработки и управления потоками данных. Фронт. Нейроинформ. 9:7. doi: 10.3389/fninf.2015.00007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Lu, J., Li, C., Singh-Alvarado, J., Zhou, Z.C., Fröhlich, F., Mooney, R., et al. (2018). MIN1PIPE: конвейер извлечения сигнала для визуализации кальция на основе однофотонного минископа.
Cell Rep. 23, 3673–3684. doi: 10.1016/j.celrep.2018.05.062 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Марковиц Дж. Э., Либерти В. А. III., Гитчаунтс Г., Вельо Т., Лоис К. и Гарднер Т. Дж. (2015). Мезоскопические паттерны нейронной активности поддерживают корковые последовательности певчих птиц. PLoS Биол. 13:e1002158. doi: 10.1371/journal.pbio.1002158
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Маршалл Дж. Д., Ли Дж. З., Чжан Ю., Гонг Ю., Сен-Пьер Ф., Лин М. З. и др. (2016). Оптическая запись динамики мембранного напряжения у свободно движущихся мышей в зависимости от типа клеток. Сотовый 167, 1650.e15–1662.e15. doi: 10.1016/j.cell.2016.11.021
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Марвин Дж. С., Шолль Б., Уилсон Д. Э., Подгорски К., Каземипур А., Мюллер Дж. А. и др. (2018). Стабильность, аффинность и хроматические варианты глутаматного сенсора iGluSnFR.
Нац. Методы 15, 936–939. doi: 10.1038/s41592-018-0171-3 CrossRef Полный текст | Google Scholar
Матис А., Мамиданна П., Кьюри К. М., Эйб Т., Мурти В. Н., Матис М. В. и др. (2018). DeepLabCut: безмаркерная оценка позы определяемых пользователем частей тела с глубоким обучением. Нац. Неврологи. 21, 1281–1289. doi: 10.1038/s41593-018-0209-y
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мацумото Дж., Уракава С., Такамура Ю., Малчер-Лопес Р., Хори Э., Томаз С. и др. (2013). Компьютеризированный анализ социальных и сексуальных взаимодействий у крыс на основе 3D-видео. PLoS One 8:e78460. doi: 10.1371/journal.pone.0078460
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Mimica, B., Dunn, B.A., Tombaz, T., Bojja, V.P.T.N.C.S., and Whitlock, J.R. (2018). Эффективное корковое кодирование трехмерной позы у свободно ведущих себя крыс. Наука 362, 584–589. doi: 10.1126/science.
aau2013 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Митани А. и Комияма Т. (2018). Обработка данных двухфотонной визуализации кальция в режиме реального времени, включая коррекцию артефактов бокового движения. Фронт. Нейроинформ. 12:98. doi: 10.3389/fninf.2018.00098
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Монтгомери К.Л., Йех А.Дж., Хо Дж.С., Цао В., Мохан Айер С., Гросеник Л. и др. (2015). Полностью внутренняя оптогенетика с беспроводным питанием для мозга, спинного мозга и периферических цепей у мышей. Нац. Методы 12, 969–974. doi: 10.1038/nmeth.3536
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Моррис Р. (1984). Разработка процедуры водного лабиринта для изучения пространственного обучения у крыс. J. Neurosci. Методы 11, 47–60. doi: 10.1016/0165-0270(84)-4
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мукамель, Э.
А., Ниммерьян, А., и Шнитцер, М. Дж. (2009). Автоматизированный анализ клеточных сигналов из крупномасштабных данных визуализации кальция. Нейрон 63, 747–760. doi: 10.1016/j.neuron.2009.08.009 PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Надлер, Дж. Дж., Мой, С. С., Долд, Г., Транг, Д., Симмонс, Н., Перес, А., и др. (2004). Автоматизированный аппарат для количественной оценки поведения социального подхода у мышей. Гены Поведение мозга. 3, 303–314. doi: 10.1111/j.1601-183x.2004.00071.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Накамура Т., Мацумото Дж., Нисимару Х., Бретас Р. В., Такамура Ю., Хори Э. и др. (2016). Безмаркерная трехмерная компьютеризированная система захвата движения, включающая модель скелета обезьяны. PLoS One 11:e0166154. doi: 10.1371/journal.pone.0166154
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Нёбауэр Т., Скочек О.
, Перния-Андраде А. Дж., Вейльгуни Л., Трауб Ф. М., Молодцов М. И. и др. (2017). Оцените видеообъемную визуализацию Ca2+ в коре головного мозга с помощью микроскопии с итеративным демиксированием (SID). Нац. Методы 14, 811–818. doi: 10.1038/nmeth.4341 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Обенхаус Х. А., Роуз Т., Цзун В., Цао А., Донато Ф., Хойдал А. и др. (2018). Миниатюрная двухфотонная микроскопия позволяет изучать топографию функциональной сети в медиальной энторинальной коре. Программа № 689.06. Планировщик встреч по нейробиологии на 2018 г. (Сан-Диего, Калифорния: Общество нейробиологов), доступно в Интернете по адресу: https://www.abstractsonline.com/pp8/#!/4649/presentation/9606
Okuyama, T., Kitamura, T. , Рой Д.С., Итохара С. и Тонегава С. (2016). Вентральные нейроны СА1 хранят социальную память. Наука 353, 1536–1541. doi: 10.1126/science.aaf7003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Озбай Б.
Н., Футия Г. Л., Ма М., Брайт В. М., Гопинат Дж. Т., Хьюз Э. Г. и др. (2018). Трехмерная двухфотонная визуализация мозга свободно движущихся мышей с использованием миниатюрного оптоволоконного микроскопа с активным аксиальным сканированием. Науч. Респ. 8:8108. doi: 10.1038/s41598-018-26326-3 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пакер, А. М., Рассел, Л. Э., Далглиш, Х. В. П., и Хойссер, М. (2014). Одновременная полностью оптическая манипуляция и запись активности нейронной цепи с клеточным разрешением in vivo . Нац. Методы 12, 140–146. doi: 10.1038/nmeth.3217
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Парастарфейзабади, М., и Кузани, А.З. (2017). Достижения в области устройств глубокой стимуляции мозга с обратной связью. Дж. Нейроинж. Реабилит. 14:79. doi: 10.1186/s12984-017-0295-1
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Park, S.
I., Brenner, D.S., Shin, G., Morgan, C.D., Copits, B.A., Chung, H.U., et al. (2015). Мягкие, растяжимые, полностью имплантируемые миниатюрные оптоэлектронные системы для беспроводной оптогенетики. Нац. Биотехнолог. 33, 1280–1286. doi: 10.1038/nbt.3415 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Паске, М. О., Тихи, М., Гуржон, А., Помпили, М. Н., Годсил, Б. П., Лена, К., и др. (2016). Беспроводное инерционное измерение кинематики головы свободно движущихся крыс. Науч. Респ. 6:35689. doi: 10.1038/srep35689
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Патриархи Т., Чо Дж. Р., Мертен К., Хоу М. У., Марли А., Сюн У.-Х. и др. (2018). Сверхбыстрая нейронная визуализация динамики дофамина с помощью разработанных генетически кодируемых сенсоров. Наука 360: eaat4422. doi: 10.1126/science.aat4422
Полный текст CrossRef | Google Scholar
Перейра Т. Д., Алдарондо Д. Э., Уиллмор Л.
, Кислин М., Ван С. С.-Х., Мурти М. и др. (2019). Быстрая оценка позы животного с использованием глубоких нейронных сетей. Нац. Методы 16, 117–125. doi: 10.1038/s41592-018-0234-5 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пяткевич К.Д., Юнг Э.Е., Штрауб К., Линху К., Парк Д., Сук Х.-Дж. и др. (2018). Роботизированный подход к многомерной направленной эволюции применительно к флуоресцентным репортерам напряжения. Нац. хим. биол. 14, 352–360. doi: 10.1038/s41589-018-0004-9
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Piyawattanametha, W., Barretto, R.P.J., Ko, T.H., Flusberg, B.A., Cocker, E.D., Ra, H., et al. (2006). Быстросканирующая двухфотонная флуоресцентная визуализация на основе микроэлектромеханических систем двумерного сканирующего зеркала. Опц. лат. 31, 2018–2020. doi: 10.1364/ol.31.002018
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пневматикакис, Э.
А., и Джованнуччи, А. (2017). NoRMCore: онлайн-алгоритм кусочно-жесткой коррекции данных визуализации кальция. J. Neurosci. Методы 291, 83–94. doi: 10.1016/j.jneumeth.2017.07.031 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Prevedel, R., Verhoef, A.J., Pernía-Andrade, A.J., Weisenburger, S., Huang, B.S., Nöbauer, T., et al. (2016). Быстрая объемная визуализация кальция в нескольких слоях коры с использованием скульптурного света. Нац. Методы 13, 1021–1028. doi: 10.1038/nmeth.4040
Полный текст CrossRef | Google Scholar
Прса, М., Галинанес, Г.Л., и Хубер, Д. (2017). Быстрая интеграция искусственной сенсорной обратной связи во время оперантного кондиционирования нейронов моторной коры. Нейрон 93, 929.e6–939.e6. doi: 10.1016/j.neuron.2017.01.023
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Quicke, P., Song, C., McKimm, E.J., Milosevic, M.M., Howe, C.L., Neil, M., et al.
(2019). Однофотонная визуализация напряжения на уровне одного нейрона с редко нацеленными генетически закодированными индикаторами напряжения. Перед. Клетка. Неврологи. 13:39. doi: 10.3389/fncel.2019.00039 PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Савински Дж., Уоллес Д. Дж., Гринберг Д. С., Гроссманн С., Денк В. и Керр Дж. Н. Д. (2009). Зрительно вызванная активность клеток коры головного мозга у свободно движущихся животных. Проц. Натл. акад. науч. США 106, 19557–19562. doi: 10.1073/pnas.00106
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шмид М., Тиле С., Херкоммер А. и Гиссен Х. (2018). Трехмерные прямые лазерные аксиконы и многокомпонентные микролинзы. Опц. лат. 43, 5837–5840. doi: 10.1364/ol.43.005837
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Скотт Б. Б., Тиберге С. Ю., Го К., Терво Д. Г. Р., Броди С. Д., Карпова А. Ю. и др.
(2018). Визуализация динамики коры у трансгенных крыс GCaMP с помощью широкопольного макроскопа, закрепленного на голове. Нейрон 100, 1045.e5–1058.e5. doi: 10.1016/j.neuron.2018.09.050 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сенаратна Дж., Ю Х., Денг К., Цзоу А. Л., Исса Дж. Б., Хаджиабади Д. Х. и др. (2019). Миниатюрный мультиконтрастный микроскоп для функциональной визуализации свободно ведущих себя животных. Нац. коммун. 10:99. doi: 10.1038/s41467-018-07926-z
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шин Г., Гомес А. М., Аль-Хасани Р., Чон Ю. Р., Ким Дж., Се З. и др. (2017). Гибкая беспроводная оптоэлектроника ближнего поля в качестве подкожных имплантатов для широкого применения в оптогенетике. Нейрон 93, 509.e3–521.e3. doi: 10.1016/j.neuron.2016.12.031
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Шуман Т., Ахарони Д., Кай Д. Дж.
, Ли С. Р., Чавлис С., Таксидис Дж. и др. (2018). Нарушение пространственного кодирования и нейронной синхронизации при эпилепсии. bioRxiv [Препринт]. 358580. doi: 10.1101/358580 CrossRef Full Text | Google Scholar
Зигле Дж. Х., Лопес А. К., Патель Ю. А., Абрамов К., Охайон С. и Фойгтс Дж. (2017). Open Ephys: платформа с открытым исходным кодом для многоканальной электрофизиологии. J. Нейронная инженерия. 14:045003. doi: 10.1088/1741-2552/aa5eea
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Скочек О., Нёбауэр Т., Вейлгуни Л., Мартинес Трауб Ф., Ся С. Н., Молодцов М. И. и др. (2018). Высокоскоростная объемная визуализация активности нейронов свободно движущихся грызунов. Нац. Методы 15, 429–432. doi: 10.1038/s41592-018-0008-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сорокин Дж. М., Дэвидсон Т. Дж., Фрешетт Э., Абрамян А. М., Дейссерот К., Хьюгенард Дж. Р. и др.
(2017). Двунаправленный контроль сетей генерализованной эпилепсии посредством быстрого переключения режима срабатывания в реальном времени. Нейрон 93, 194–210. doi: 10.1016/j.neuron.2016.11.026 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Стаматакис А. М., Шахтер М. Дж., Гулати С., Зителли К. Т., Малановский С., Таджик А. и др. (2018). Одновременная оптогенетика и визуализация кальция с клеточным разрешением во время активного поведения с использованием миниатюрного микроскопа. Перед. Неврологи. 12:496. doi: 10.3389/fnins.2018.00496
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Тиле, С., Гиссибл, Т., Гиссен, Х., и Херкоммер, А. М. (2016). Сверхкомпактная встроенная светодиодная коллимационная оптика с трехмерной фемтосекундной прямой лазерной записью. Опц. лат. 41, 3029–3032. doi: 10.1364/ol.41.003029
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Терли, К.
, и Аяз, А. (2017). Системы виртуальной реальности для грызунов. Курс. Зоол. 63, 109–119. doi: 10.1093/cz/zow070 PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Венкатраман, С., Джин, X., Коста, Р. М., и Кармена, Дж. М. (2010). Исследование нейронных коррелятов поведения свободно ведущих себя грызунов с помощью инерциальных датчиков. J. Нейрофизиол. 104, 569–575. doi: 10.1152/jn.00121.2010
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Уилсон, Дж. Дж., Александр, Н., Трентин, К., и Триподи, М. (2018). Трехмерное представление двигательного пространства в верхнем двухолмии мышей. Курс. биол. 28, 1744.e12–1755.e12. doi: 10.1016/j.cub.2018.04.021
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Вильчко А. Б., Джонсон М. Дж., Юрилли Г., Петерсон Р. Э., Катон Дж. М., Пашковски С. Л. и др. (2015). Отображение субсекундной структуры в поведении мыши. Нейрон 88, 1121–1135.
doi: 10.1016/j.neuron.2015.11.031 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Инь X., Штайнле Т., Хуанг Л., Таубнер Т., Вуттиг М., Зентграф Т. и др. (2017). Переключение луча и бифокальное зум-линзирование с использованием активных плазмонных метаповерхностей. Легкие науки. заявл. 6:e17016. doi: 10.1038/lsa.2017.16
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чжан Л., Лян Б., Барбера Г., Хоуз С., Чжан Ю., Стамп К. и др. (2018). Система линз Miniscope GRIN для визуализации кальциевой активности нейронов в глубоких структурах мозга у животных с поведением. Курс. протокол Неврологи. 86:e56. doi: 10.1002/cpns.56
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Zhou, P., Resendez, S.L., Rodriguez-Romaguera, J., Jimenez, J.C., Neufeld, S.Q., Giovannucci, A., et al. (2018). Эффективное и точное извлечение in vivo сигналы кальция из микроэндоскопических видеоданных.
Элиф 7:e28728. doi: 10.7554/elife.28728 PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Zong, W., Wu, R., Li, M., Hu, Y., Li, Y., Li, J., et al. (2017). Быстрая миниатюрная двухфотонная микроскопия с высоким разрешением для визуализации мозга свободно ведущих себя мышей. Нац. Методы 14, 713–719. doi: 10.1038/nmeth.4305
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Miniscope: Миниатюрный флуоресцентный микроскоп
Новый инструмент микроскопии дает неврологам невиданный ранее взгляд на клеточные сети, лежащие в основе поведения.
Модель миниатюрного люминесцентного микроскопа (минископ).
Источник: miniscope.orgЗакройте глаза и представьте любимое воспоминание из недавней поездки. Вполне вероятно, что другие воспоминания из той поездки начнут всплывать, воссоздавая в вашей голове небольшое домашнее видео. Но почему размышления об одном воспоминании вызывают воспоминание о другом событии? Связаны ли воспоминания, связанные во времени, также и физически? Или одна и та же группа нейронов кодирует оба воспоминания?
Невероятные достижения в области неврологии приблизили исследователей к ответам на эти вопросы, которым уже несколько десятков лет.
Однако методы манипулирования активностью нейронов, по-видимому, процветали быстрее, чем методы визуализации и мониторинга активности свободно ведущих себя животных. Хотя в настоящее время существуют способы мониторинга активности нейронов, когда мышь выполняет определенную задачу, по-прежнему сложно записывать ответы одной и той же клетки во времени. Теперь преобразующий инструмент визуализации под названием минископ, рожденный в результате сотрудничества нескольких групп, направлен на решение этих проблем. Переделка минископа
Минископ, миниатюрный флуоресцентный микроскоп, первоначально впервые разработанный в лаборатории Марка Шнитцера в Стэнфордском университете [1], маленький и портативный, что делает его идеальным инструментом для изучения активности мозга у мышей, занимающихся поведение. Ранее использовавшиеся микроскопы с фиксированной головой были большими и тяжелыми, обездвиживая крошечных мышей и не позволяя им выполнять многие из своих обычных действий.
«Минископ позволяет нам увидеть, как мозг меняется и адаптируется к миру», — сказал Альчино Сильва из Калифорнийского университета в Лос-Анджелесе (UCLA), соучредитель веб-сайта с открытым исходным кодом miniscope.org. В тесном сотрудничестве с группой Пеймана Гольшани, также работающей в Калифорнийском университете в Лос-Анджелесе, группа Сильвы переработала оригинальный минископ, создав гораздо более дешевую версию с открытым исходным кодом, тем самым расширив доступность этого нового инструмента для всего научного сообщества.
Минископы можно использовать практически в любом месте мозга для измерения флуоресценции, испускаемой генетически закодированным индикатором кальция; каждый раз, когда ячейка становится активной, она загорается, передавая эту информацию через минископ. «Это позволяет нам наблюдать за активностью одних и тех же нейронов в течение многих дней или недель, что является очень важной возможностью — увидеть, как активность одних и тех же клеток меняется в зависимости от поведения или обучения», — говорит Гольшани.
сказал. Минископы в движении
В прошлом году Сильва и его коллеги использовали минископ для изучения записи воспоминаний у активных мышей (2). Они поместили мышей в три разные по контексту камеры, поразив их током в одной из камер, чтобы вызвать пугающую реакцию замирания. Как и ожидалось, при повторном воздействии на камеры мыши замерзали в камере, где они получили удар током. Интересно, что мыши также демонстрировали аналогичные уровни замирания в камере, где их не подвергали шоку, но где они выжидали непродолжительное время в течение 5 часов до удара током. Этот эффект не наблюдался во время повторного воздействия камеры, которую мыши исследовали за 7 дней до получения удара током, что подтверждает гипотезу группы о том, что воспоминания, сформированные в непосредственной близости друг от друга, контекстуально связаны. Но что происходит на клеточном уровне?
Чтобы ответить на этот вопрос, исследователи ввели флуоресцентный индикатор кальция GCaMP6f в область CA1 гиппокампа, а затем имплантировали линзу микроскопа для доступа через минископ.
Затем они позволили мышам исследовать три разные камеры и варьировали время между посещением новой камеры. Используя минископ, исследователи измерили флуоресценцию и, следовательно, нейронную активность отдельных нейронов, когда мыши исследовали эти различные контексты. Они обнаружили значительно большее перекрытие в группе нейронов, реактивированных в результате воздействия камер, когда первоначальные события происходили с разницей в 5 часов, чем когда события происходили с разницей в 7 дней. Впервые исследователи смогли показать нейронную динамику связывания воспоминаний, закодированных близко во времени. «Видение — это часть веры», — сказал Сильва. «В этом сила технологий обработки изображений».
Модификации минископа
Сильва, Гольшани и их сотрудники неустанно работали над усовершенствованием минископа, чтобы сделать его еще дешевле и проще в использовании в лабораториях по всему миру. Есть несколько захватывающих дополнений к семейству минископов, которые они надеются представить в ближайшее время.
Текущая система минископа измеряет флуоресценцию, связанную с передачей сигналов кальция, классической системой вторичных мессенджеров. «В будущем мы надеемся, что сможем измерять флуоресцентные индикаторы для других транзакционных систем вторичных мессенджеров, чтобы мы могли следить не только за активностью конкретных клеток, но и за тем, активировался ли в этой клетке определенный путь передачи сигнала во время конкретное поведение», — сказал Гольшани.
Кроме того, в сотрудничестве с лабораторией Silva лаборатория Golshani оптимизирует беспроводной минископ с батарейным питанием, который позволит мышам двигаться в более широком диапазоне. В настоящее время с помощью этой системы они могут записывать только 20 минут за раз, поскольку исследователи ограничены весом батареи.
Наконец, команда приближается к проверке оптогенетического минископа, а также многоканального флуоресцентного минископа. Эти новые модификации позволят исследователям задавать вопросы о мозге и поведении, на которые до сих пор не было ответа.
Например, исследователи могли бы активировать или ингибировать определенные популяции клеток с помощью света, а затем регистрировать не только поведенческие результаты, но и активность отдельных клеток и локальных схем в ответ на внешние манипуляции. Источник
- (1) Ghosh et al. Миниатюрная интеграция флуоресцентного микроскопа. Природные методы. 2011 г., 11 сентября; 8, 871–878. doi:10.1038/nmeth.1694
- (2) Cai et al. Общий нейронный ансамбль связывает различные контекстуальные воспоминания, закодированные близко во времени. Природа. 2016 23 мая; 534, 115–118. дои: 10.1038 / природа17955
2021 Virtual Miniscope Workshop
General Miniscope Workshop
Записи первого дня
1/6)» src=»https://www.youtube.com/embed/3SX6VdKRiqg» allowfullscreen=»»/>
4/6)» src=»https://www.youtube.com/embed/STdfVPeKWvQ» allowfullscreen=»»/>
General Miniscope Workshop
Day 2 Recordings
1/9)» src=»https://www.youtube.com/embed/BON6VaCxf3E» allowfullscreen=»»/>
4/9)» src=»https://www.youtube.com/embed/gi6u-CaZqao» allowfullscreen=»»/>
7/9)» src=»https://www.youtube.com/embed/7icsleqBljw» allowfullscreen=»»/>
Hands-On Analysis Учебник
Записи 3-го дня
1/5)» src=»https://www.youtube.com/embed/617_n_YtR0w» allowfullscreen=»»/>
4/5)» src=»https://www.youtube.com/embed/oflOr4ioeh8″ allowfullscreen=»»/>
Продолжайте учиться
-
Пожалуйста, выберите «Индивидуальная поддержка» в этой форме, если вы хотите получить индивидуальную помощь для анализа данных визуализации кальция. Мы свяжемся с вами, чтобы организовать звонок для обсуждения ваших потребностей;
-
Выберите «Бесплатная пробная версия» (та же форма), если вы хотите в интерактивном режиме запустить ту же записную книжку, которая использовалась в демо-версии Практического семинара по анализу, без локальной настройки.
Вы получите логин в среду, дублирующую ту, что использовалась в мастерской; -
Выберите «Рабочее пространство для обучения» (та же форма), если вы заинтересованы в проведении семинара, который включает в себя практический компонент с использованием сред анализа данных с нулевой настройкой, подобных той, которую мы создали для практического руководства Virtual Miniscope.
О семинаре Virtual Miniscope Workshop 2021
Добро пожаловать на семинар Virtual Miniscope Workshop 2021! Мы рады провести виртуальный семинар с лекциями и практическими занятиями, которые научат участников, как использовать минископы с открытым исходным кодом для визуализации кальция in vivo. Этот семинар предназначен для ознакомления со всеми аспектами использования минископов, включая основные принципы проектирования и визуализации минископов, как собрать и прикрепить минископ, как имплантировать линзу GRIN для визуализации глубоких структур и как анализировать данные визуализации.
На семинаре также будут рассмотрены самые последние разработки в технологии Miniscope и отмечены некоторые из лучших достижений в этой захватывающей и развивающейся области. Мы предлагаем два отдельных курса для этого семинара:
-
Комплексный двухдневный семинар по Miniscope (29-30 ноября) охватит все аспекты визуализации Miniscope, от разработки оборудования до внедрения, и будет включать лекцию, посвященную последним достижениям. в анализе данных. Этот семинар подходит для всех уровней опыта, и участники должны покинуть семинар с достаточными знаниями, чтобы начать создавать и выполнять визуализацию в своей собственной лаборатории. Этот мастер-класс открыт для всех, и количество мест не будет ограничено, но необходима регистрация.
-
В практическом учебном пособии по анализу (30 ноября — 1 декабря) будет подробно рассказано, как анализировать данные с помощью Minian, пакета анализа на основе Python, разработанного Cai, Aharoni и Shuman Labs.
Участники пройдут анализ демонстрационных данных через облачную платформу, а затем будут работать над анализом своих собственных данных или дополнительных предоставленных данных. Чтобы обеспечить адекватное взаимодействие с инструкторами, этот трек ограничен 25 участниками, поэтому вы должны подать заявку на этот трек.
Участники могут зарегистрироваться/подать заявку на оба направления или выбрать одно направление. Оба трека абсолютно бесплатны для посещения и пройдут с 29 ноября по 1 декабря 2021 года. Это мероприятие организовано Даниэлем Ахарони, Дениз Кай и Тристаном Шуманом и проводится в рабочей области MetaCell для анализа визуализации кальция.
Структура виртуального семинара
Общий семинар по минископу
Дни 1-2-
Изучите основные принципы визуализации с помощью минископов, как построить минископ, установить линзы GRIN и получить данные во время поведения
-
Узнайте о конвейере анализа Minian и других инструментах анализа изображений кальция
-
Узнайте о последних разработках Miniscope, текущих проектах и будущих направлениях Miniscope
-
Узнайте, как исследователи во всем мире используют Miniscope и разрабатывают новые инновации в миниатюрной микроскопии
-
Общайтесь с другими учеными и разработчиками во время различных интерактивных секционных сессий
-
Открыто для всех, но регистрация требуется
Руководство по практическому анализу
Дни 2-3-
Учебный урок.
как взаимодействовать с Minian в облаке для выполнения анализа на серверах анализа -
Каждый участник запустит конвейер Minian на демонстрационных данных, а затем сможет провести анализ, используя свои собственные данные или дополнительные предоставленные наборы данных
-
Сеть с сообществом исследователей, работающих над анализом данных визуализации кальция в условиях небольшой группы
-
Количество мест ограничено 25 участниками, поэтому подавайте заявку заранее!
докладчики
Дениз CAI
Доцент, Школа медицины ICAHN на горе SINAI
CAI LAB
DANIEL AHARONI
.
0005 Aharoni Lab
Tristan Shuman
Доцент, Медицинский факультет ICAHN, 9,0977, 9000, 908, 908,
Tycho Hoogland,
,,
Tycho Hoogland
. КемереДоцент, Университет Райса
Лаборатория Кемере
Андреа Джованнуччи
Доцент, Университет Северной Каролины в Чапел-Хилл
Нейронная инженерная лаборатория
Лаура Уоллер
Доцент профессор UC Berkeley
LABINGING Imaging
YANIV ZIV
Аспирант.
Постдокторский научный сотрудник
Cai Lab
Закари Пеннингтон
Постдокторский научный сотрудник
zachpenn.
github.io William Mau
Postdoctoral Fellow
Cai Lab
Phil (Zhe) Dong
Graduate Student
Cai Lab
Joe (Yosif) Zaki
Graduate Student
Cai Lab
Susie (Yu) Feng
Аспирант
Лаборатория Шумана
Федерико Санджулиано
Лаборант
Лаборатория Ахарони
Стивен Ларсон 05Соучредитель и генеральный директор
MetaCell
Паоло Баззигалуппи
Старший консультант
MetaCell
Пожалуйста, напишите по адресу Workshop@metacell.
us Микроскопия своими руками с открытым исходным кодом | Edmund Optics
Доступная, гибкая технология, поддерживаемая сообществом пользователей
Сеть пользователей, сотрудничающих для разработки и улучшения технологии
Экономичные системы микроскопии, доступные широкой аудитории
Системы микроскопов, которые легко собрать и модифицировать
Способствует распространению жизненно важных передовых диагностических методов
Технология с открытым исходным кодом основана на сообществе пользователей, которые сотрудничают в разработке, тестировании, совместном использовании и улучшении технологии.
Предоставляется техническая информация, и пользователи технологии обмениваются идеями и разработками, связанными с продуктом. Наиболее распространенным продуктом с открытым исходным кодом является программное обеспечение, поскольку оно хорошо подходит для редактирования и манипулирования сетью людей. Тем не менее, другие технологии, такие как микроскопия, также могут быть открытыми, что делает очень дорогие продукты доступными для гораздо более широкой аудитории. Микроскопия «сделай сам» с открытым исходным кодом обеспечивает доступный, экономичный и гибкий способ использования микроскопических систем для исследований и прототипирования. UCLA Miniscope: флуоресцентный микроскоп с открытым исходным кодом
Для исследователей в области неврологии изучение нейронной активности на свободно ведущих себя животных имеет решающее значение для исследований, но традиционные микроскопы, используемые для изучения мозга животных, как правило, громоздки и дороги, и при использовании должны быть стационарными.
. Это делает их крайне неудобными для наблюдения за нейронами мыши, исследующей окружающую среду или взаимодействующей с другими животными. Лаборатории Khakh, Silva и Golshani в Калифорнийском университете в Лос-Анджелесе (UCLA) создали миниатюрный флуоресцентный микроскоп с открытым исходным кодом, устанавливаемый на голове, для изучения нейронной активности на свободно ведущих себя мышах. Любая нейробиологическая лаборатория может построить это оптическое устройство за долю стоимости коммерческого флуоресцентного микроскопа.
Рис. 2: Минископ в поперечном сеченииМинископ крепится на голову мыши для визуализации активности нейронов и выявления различий в нейронной активности между здоровым мозгом и мозгом с неврологическими заболеваниями. В соответствии со своей целью сделать исходный код открытым, исследовательская группа Калифорнийского университета в Лос-Анджелесе создала дизайн этого микроскопа и разместила список деталей, инструкции по сборке и использованию Miniscope, а также ссылки на видеоуроки на специальном вики-сайте.
Эта вики представляет собой централизованное место для обмена файлами дизайна, исходным кодом и другой соответствующей информацией, чтобы сообщество пользователей и ученых со всего мира могло обмениваться идеями и разработками, связанными с этим важным методом визуализации. Центральное расположение информации помогает распространять технологию среди более широкого сообщества нейробиологов, создавая базу пользователей, которые будут продолжать продвигать эту технологию и вносить свой вклад в проект.
Рис. 3: Минископ в разобранном видеЭтот микроскоп для самостоятельной сборки включает стандартные компоненты от Edmund Optics ® , включая ахроматические линзы, полусферические линзы и линзы GRIN. Miniscope представляет собой строительный блок стабильной платформы, на которую лаборатории могут добавлять компоненты для поддержки своего конкретного использования микроскопа. Сегодня Miniscope используют более 300 лабораторий, многие из которых адаптировали устройство к своим уникальным исследовательским потребностям.
Чтобы узнать больше или приобрести полный набор компонентов для Miniscope, посетите эту веб-страницу. Часто задаваемые вопросы
Продает ли Edmund Optics ® комплектные минископы?
EO продает несколько компонентов, которые входят в состав минископа, в том числе наши линзы с диаметром 5 мм и фокусным расстоянием 15 мм, MgF 2 Ахроматическая дуплетная линза с покрытием (#45-207), полушаровидная линза N-BK7 диаметром 3,0 мм (#47- 269) и нашу линзу GRIN без покрытия диаметром 1,8 мм, 670 нм DWL (№ 64-519).
Где я могу приобрести полный набор компонентов для Miniscope?
Чтобы узнать больше о Miniscope и приобрести полный набор компонентов, посетите эту веб-страницу.
Сколько весит Минископ?
Минископ в собранном виде весит 3 грамма. Эта малая масса позволяет устанавливать его на голове свободно ведущих себя мышей.
Где я могу найти все файлы дизайна, исходный код и другую важную информацию о Miniscope?
Всю информацию из открытых источников, связанную с Miniscope, можно найти на вики-странице по адресу www.miniscope.org.
Проект Miniscope все еще находится в разработке?
Да, по состоянию на январь 2018 года проект Miniscope все еще продолжается, и его сообщество пользователей продолжает делиться новыми идеями и разработками.
-
Примечания по применению -
Связанные страницы
Замечания по применению
Техническая информация и примеры применения, включая теоретические пояснения, уравнения, графические иллюстрации и многое другое.
Флуоресцентная микроскопия: встроенное освещение с визуализирующими фильтрами
ЧитатьФлуорофоры и оптические фильтры для флуоресцентной микроскопии
ЧитатьОптическая микроскопия.
Применение: Fluorescence
ReadСвязанные страницы
.
Расширенная диагностика: мозг
Узнать большеРождение минископа Дэвидом Алленом
Дополнительные фотографии и информация находятся на странице онлайн-выставки осциллографов серии 200. Этот рабочий прототип «минископа» ручной работы сыграл важную роль в начальной разработке серии 200. Дэйв Аллен восстановил его до рабочего состояния после почти 50 лет игнорирования.
Трудно вспомнить, прошло 50 лет. В 1968 году я пришел работать в группу Advanced Circuit Design Хиро Мориясу в качестве летнего студента. Я уже собрал осциллограф и осциллограф собственной разработки, прежде чем пришел в Tektronix. Я помню, как был впечатлен Sony/Tek 321, и меня очень заинтересовала идея осциллографа в зонде с этим 1-дюймовым ЭЛТ. Я уверен, что идея новой линейки миниатюрных осциллографов обсуждалась на собраниях сотрудников, в которых участвовали Говард, Билл, Хиро и другие.
Когда я вернулся в Tek после окончания учебы, мне, должно быть, поручили работать над новым проектом «минископ».0005 Будучи новым инженером в крупной компании, я чувствовал себя как в «раю» со складским помещением размером с мою гостиную со всеми компонентами, которые только мог пожелать инженер. У меня был лучший осциллограф, который можно было купить за деньги, и много других фантастических инструментов. Я начал собирать вещи, чтобы построить 1-дюймовый прицел в зонде. Я был готов разработать что-то великое.
Я очень мало знал обо всей поддержке, которую мог предоставить Tek, например, о проектировании ЭЛТ, промышленном дизайне, машиностроении, производстве прототипов, компоновке ECB, керамике, многих других группах поддержки и желающих поставщиках компонентов. Я уверен, что Хиро был тем, кто связался со всеми этими группами, чтобы все заработало (очевидно, под влиянием «Группы перспективного проектирования схем» и комментария типа «Ховард хочет этого»). Все концептуальные прототипы были сделаны Группа промышленного дизайна, и каждую неделю у нас была еще одна маленькая модель для рассмотрения.
Некоторые были сделаны из картона и пенопласта, а другие были сформированы, окрашены, с прозрачным верхом и имели настоящие поворотные ручки. Я точно не помню, как окончательный формат появилась серия 200, но у нее была панель управления хорошего размера, место для хранения зонда и наклонная подставка для удобства просмотра — более практичная версия футуристических предыдущих моделей 9.0005 В то время Тек только что запустил новую фабрику по производству аналоговых интегральных схем «LSI» и искал новые конструкции. Я начал проектировать микросхемы для небольшого объема. Лаборатория IC должна предоставить два или три настоящих IC-транзистора в восьмиконтактном корпусе. Я использовал много транзисторных корпусов для создания и тестирования предложенной конструкции, используя только транзисторы IC, несколько резисторов и конденсаторы NO. После демонстрации того, что схема прототипа работает, принципиальная схема была передана группе макетирования маски ИС, и через две или три недели все слои диффузионных масок были разрезаны на листы рубилита размером 4 фута на 6 футов для фотоуменьшения для диффузии.
маски нового ИС. Еще через неделю или около того мне дали несколько настоящих ИС, и они работали точно так же, как сделанный вручную прототип. Я разработал IC схемы полной развертки, IC вертикального усилителя и IC выхода, каждый в отдельных IC Единственным 16-выводным корпусом ИС, доступным в то время, был двухрядный корпус длиной около 1 дюйма, слишком большой для настоящего «минископа». сидел в отверстии в ECB и был впаян вручную. Tek не нравились впаянные ИС, поэтому в области упаковки ИС был разработан 16-контактный квадратный корпус, который можно было вставить, и мы согласились с этим. В то время компоненты для поверхностного монтажа только появлялись, но у Tek не было средств для их размещения, а Tek не думала, что они будут настолько надежными, поэтому мы использовали стандартные миниатюрные компоненты (для компонентов, которые раньше не использовались). ИС) и воспользовались преимуществами сетей керамических резисторов Tek для прецизионных резисторов.
Поскольку поворотные переключатели были слишком велики для такого маленького минископа, мы разработали небольшой поворотный переключатель с контактами, которые можно было использовать на золотых трассах ЕЦБ.
Думаю, в то время разрабатывался кулачковый переключатель. Мы сделали логику переключения выгравированной прямо на ECB. У нас были пластины коммутатора с обеих сторон ECB, и какое-то время у нас были сквозные проходы «на полпути», поэтому мы могли подключить второй уровень к коммутатору на первом уровне, а третий уровень соединить с четвертым уровнем, не испортив другой. стороны. ECB Fab не нравились «половинчатые проходы», поэтому мы изменили расположение переключателей, чтобы сделать их счастливыми. Макет ECB был прямо в коридоре, и со схемой, чертежом контура, опытом хорошего специалиста по компоновке и некоторыми предложениями через плечо, примерно через неделю четырехслойный ECB был полностью заклеен и готов к фотографированию. снижение. В то время в подвале 50-го здания располагалась фабрика ECB, и если во второй половине дня была сдана серия съемок, на следующее утро у вас был готовый ECB. Насколько это может быть хорошо?
Две вещи, которые я помню о магазине моделей в подвале 50-го дома, помимо всех фантастических моделей, которые они сделали для нас.
Когда мы, наконец, получили полноценный ЭЛТ для тестирования, ему понадобился магнитный экран. Обычно формируется экран из мю-металла, который затем сваривается точечной сваркой, а затем отжигается. Что ж, модельный цех решил посмотреть, смогут ли они вытянуть цельный экран ЭЛТ из цельного листа мю-металла. Для этого они сделали штампы, и я помню, как наблюдал, как они прижимали внутренний штамп к листу мю-металла и примерно за три прохода вытягивали полный ЭЛТ-экран. Это действительно сработало. Я не знаю, почему они не использовали этот процесс, вероятно, потому, что у Tek не было надлежащего оборудования для его производства, но экраны ЭЛТ серии 200, наконец, были изготовлены так же, как и другие, отформованы и сварены вместе точечной сваркой. Когда мы, наконец, получили все детали для первого (этап pre-A) 211, нам понадобился пластиковый корпус, чтобы поместить их в него. задолго до того, как можно было отлить первые детали. Модельный цех изготовил две боковые панели, выточив их из толстых пластиковых блоков.
Верхняя и нижняя части корпуса были сформированы в вакууме на алюминиевых пресс-формах, а затем обрезаны, чтобы лучше представить фактические части. Это выглядело очень грубо, но впервые у нас был настоящий «минископ» 211, который мы могли держать в руках, провести некоторые начальные тесты и похвастаться перед всеми теми, кто внес свой вклад в его рождение. Одной из первых целей проектирования было сделать осциллографы серии 200 двойной изоляцией, чтобы они были «плавающими». Мы рекламировали его как «плавучий», и в ответ последовало несколько ироничных комментариев о том, что кто-то бросил его в воду, но «ОНО СРАЗУ УТОЛО!». Благодаря двойной изоляции «минископа» он имеет пластиковый корпус. Каждая часть электроники внутри, подключенная к линиям электропередачи переменного тока, имеет по крайней мере два слоя изоляции между внутренними цепями и всем, к чему пользователь может прикоснуться. Цепь зарядки, аккумуляторы и первичная обмотка внутреннего источника питания подключены к сети переменного тока.
Поскольку заземление входного щупа можно было подключить к линиям электропередачи переменного тока, все схемы внутри устройства должны были иметь одинаковую изоляцию снаружи устройства, к которой пользователь мог бы прикоснуться. У нас должны были быть дополнительные изолированные крышки ползункового переключателя, пластиковые ручки без установочных винтов, а любое отверстие для доступа должно быть достаточно маленьким. Изоляция продукта для модели 211 была несложной, так как у нее был только один заземляющий провод для зонда. Но для двойной трассы 212 и 214 у нас должны были быть желтые провода заземления для двух датчиков со специальными инструкциями о том, что эти два провода внутренне соединены друг с другом и их следует использовать с осторожностью при плавании прибора. Я помню, как пошел в Plastics, чтобы посмотреть, как они вводят первые части корпуса 211. Белая боковая панель работала нормально, и мы получили несколько новых деталей. У них были проблемы с инъекцией верхней и нижней части корпуса, так как матрица прилипала, а не отделялась, как должна была.
Им было трудно извлечь готовую деталь из формы. Насколько я помню, проблема заключалась в том, что форма была не симметрична слева направо. В готовой детали отсутствовала вся сторона, куда подошла бы боковая панель. Крепление штампа в раме для впрыска должно было воспринимать боковое давление. Я не знаю, как они это исправили, но мы, наконец, вытащили верхнюю и нижнюю части из формы. Одна вещь, которую Tek всегда делала с новым продуктом, заключалась в тщательном тестировании его на соответствие EIS (Спецификация инженерного прибора), которая изначально была составлена и утверждена инженерами, производством, руководством и т. д. Я только что помню, как поместил этот маленький 211 в эту ужасную климатическую камеру и прокрутил его от 0 ° C до 80 ° C при относительной влажности 98% — и он все еще работал!
По мере того, как производились эти «минископы» серии 200, я следил за их количеством. Когда 1000 -й 211 как раз собирались построить, я спросил, могу ли я получить 1000 -й -й блок для передачи Говарду Воллуму.
Поскольку Electro-chem мог сделать почти все, я взял несколько корпусов 211 и попросил их покрыть их золотом. Что ж, процесс нанесения золота на пластик требовал немного большего нагрева, чем пластик мог выдержать. Несколько корпусов были сильно деформированы, поэтому я выбрал лучшие, применил немного тепла и давления, и один набор выглядел очень хорошо. Следующее препятствие вырисовывалось. Как мы все знаем, золото НЕ является изолятором. Теперь внутренняя часть корпуса была соединена с красивым золотым покрытием снаружи. Золотой 211 может быть смертельным, если вы подключите его, держа в руках. С помощью стандартного инструмента Dremel я удалил золотую дорожку на внутренней стороне корпуса шириной около ¼ дюйма по всему краю, чтобы внешняя часть была только косметической и не соединилась с внутренней частью. Это выглядело чудесно!. Мне всегда было интересно, что Ховард с ним сделал. Модель 211 была первой, модель 212 представляла собой обновление одного из электронных блоков управления и новую боковую панель.
Затем последовала модель 214, включающая в себя несколько специальных переключателей и две новые печатные платы. Также требовался новый ЭЛТ-накопитель для хранения данных, предоставленный инженерами ЭЛТ. 214 был немного больше усилий. Следующей была модель 221, и поскольку она была в десять раз быстрее, потребовалось гораздо больше инженерных разработок. Ни одна из микросхем 211 не была достаточно быстрой, поэтому вся 221 была сделана из незаметных деталей. Дискретные части были втиснуты, и это требовало немного больше энергии. К тому времени, когда мы приступили к работе над 213, я в основном руководил группой и занимался только дизайном зарядного устройства. Мы решили использовать две батареи размера D, поскольку в двух батареях D было примерно в два раза больше энергии, чем в батареях размера AA того же объема. Управлять целым прицелом от 2В было непросто. Поскольку у 213 было много разных функций, за каждой частью был закреплен отличный дизайнер. У модели 211 было два ECB, и чтобы втиснуть все функции 213 в один корпус, нам понадобилось шесть ECB.
Все детали находились внутри четырех основных ECB (ни одно свободное место не оставалось неиспользованным), и только ECB были видны, когда вы вынимали 213 из корпуса. Чтобы работать с 213 в производстве, нам пришлось разработать 3 небольших ECB и длинный кабель, который соединял бы четыре основных ECB вместе, располагая их плоско, чтобы были видны детали, над которыми нужно работать. Я уверен, что производство действительно ненавидело эту проблему. После того, как 213 был закончен, мы приступили к разработке 20-мегагерцового двухканального «минископа» с новой ЭЛТ с экраном восемь на десять делений с использованием недавно разработанной ЭЛТ с расширением сканирования сетки. Мы даже получили несколько новых ЭЛТ на пробу, но размер пятна в первых прототипах все еще был слишком большим. Пока мы разрабатывали спецификации для нового «минископа» с более высокой пропускной способностью, Tektronix заключила контракт с Burroughs на создание цифровой испытательной системы (в конечном итоге получившей название 851).
Похожие записи
 .
.