Какие основные типы сигнальных молекул используются для передачи информации между клетками. Как различные сигнальные молекулы взаимодействуют с клетками-мишенями. Какие механизмы передачи сигнала внутри клетки запускаются при связывании сигнальных молекул с рецепторами. Как регулируется передача сигналов между клетками.
Основные типы сигнальных молекул в межклеточной коммуникации
Для передачи информации между клетками многоклеточные организмы используют множество различных типов сигнальных молекул. Основные классы сигнальных молекул включают:
- Стероидные гормоны
- Пептидные гормоны
- Факторы роста
- Нейромедиаторы
- Эйкозаноиды
- Газообразные молекулы (оксид азота, монооксид углерода)
Каждый тип сигнальных молекул имеет свои особенности структуры и механизма действия на клетки-мишени. Рассмотрим их подробнее.
Стероидные гормоны и их рецепторы
Стероидные гормоны представляют собой липофильные молекулы, синтезируемые из холестерина. К ним относятся:
- Половые гормоны (тестостерон, эстрогены, прогестерон)
- Глюкокортикоиды (кортизол)
- Минералокортикоиды (альдостерон)
Благодаря своей гидрофобности, стероидные гормоны способны проникать через плазматическую мембрану клеток-мишеней путем простой диффузии. Внутри клетки они связываются со специфическими ядерными рецепторами, которые относятся к суперсемейству стероидных рецепторов.
Как действуют стероидные гормоны на уровне клетки?
- Гормон проникает в клетку
- Связывается с рецептором в цитоплазме или ядре
- Комплекс гормон-рецептор активируется
- Активированный комплекс связывается с ДНК
- Запускается транскрипция генов-мишеней
Таким образом, стероидные гормоны напрямую регулируют экспрессию генов в клетках-мишенях, что приводит к изменению синтеза белков и физиологического ответа клетки.
Пептидные гормоны и факторы роста
Пептидные гормоны и факторы роста представляют собой водорастворимые белковые молекулы различного размера — от нескольких аминокислот до крупных полипептидов. К этой группе относятся:
- Инсулин
- Гормон роста
- Пролактин
- Эпидермальный фактор роста (EGF)
- Тромбоцитарный фактор роста (PDGF)
- Нейротрофины
В отличие от стероидных гормонов, пептидные гормоны и факторы роста не способны проникать через плазматическую мембрану. Как они передают сигнал в клетку?
- Связываются с рецепторами на поверхности клетки
- Активируют рецептор
- Запускают внутриклеточные сигнальные каскады
- Это приводит к изменениям метаболизма, экспрессии генов и других клеточных процессов
Многие рецепторы факторов роста обладают собственной ферментативной активностью (например, тирозинкиназной). Их активация запускает сложные сети внутриклеточной передачи сигнала.
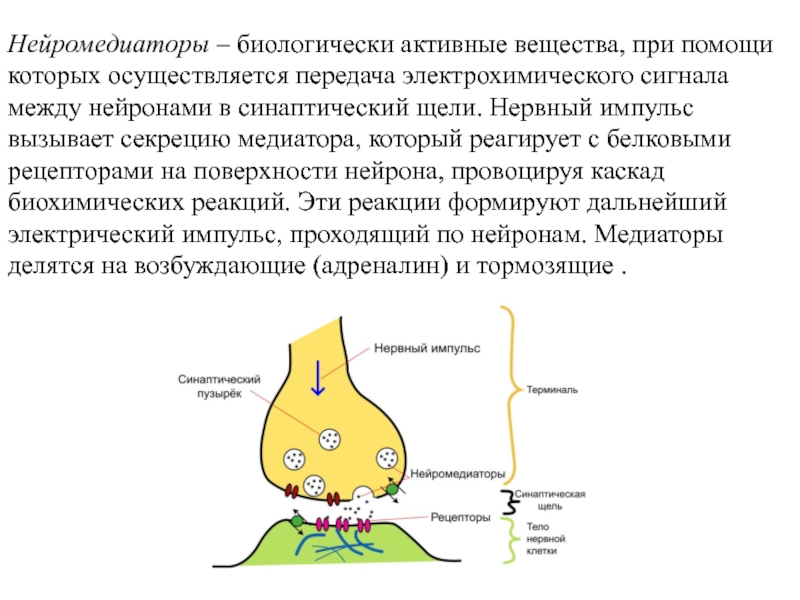
Нейромедиаторы и их рецепторы
Нейромедиаторы — это химические вещества, передающие сигналы между нейронами через синапсы. Основные классы нейромедиаторов включают:
- Аминокислоты (глутамат, ГАМК, глицин)
- Моноамины (дофамин, серотонин, норадреналин)
- Пептиды (эндорфины, энкефалины)
- Ацетилхолин
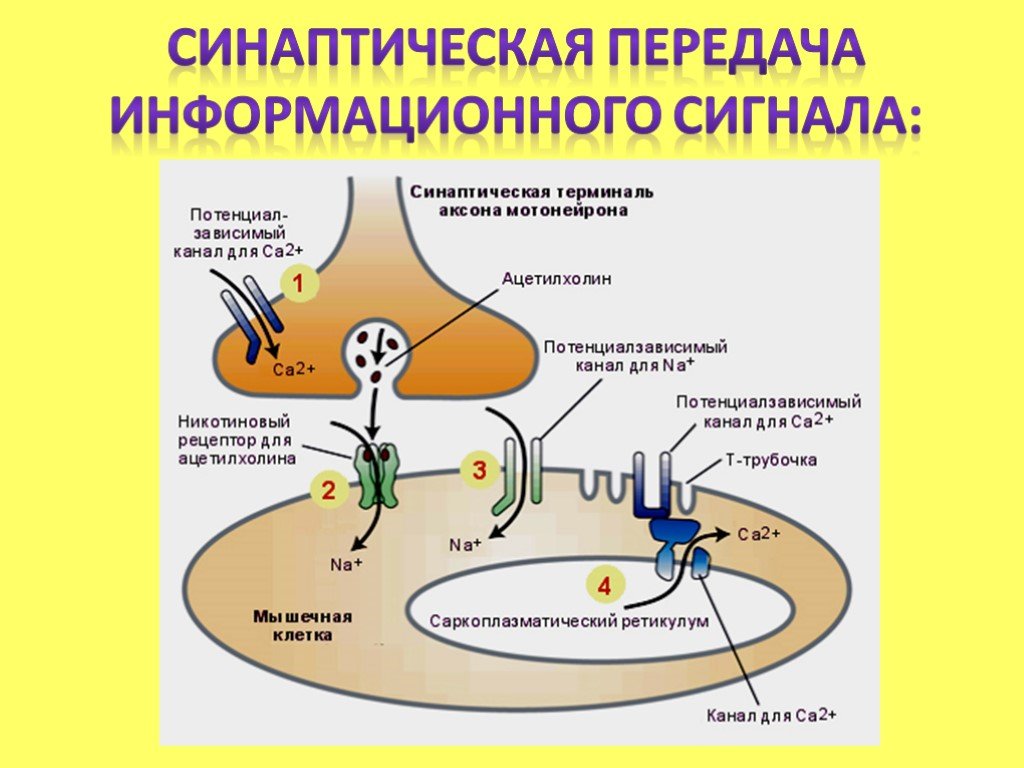
Как происходит передача сигнала с помощью нейромедиаторов?

- Нейромедиатор высвобождается в синаптическую щель
- Связывается с рецепторами на постсинаптической мембране
- Активирует ионные каналы или метаботропные рецепторы
- Изменяет мембранный потенциал или запускает внутриклеточные сигнальные каскады
- Вызывает возбуждение или торможение нейрона
Рецепторы нейромедиаторов делятся на два основных типа:
- Ионотропные — это лиганд-зависимые ионные каналы
- Метаботропные — связаны с G-белками и вторичными мессенджерами
Газообразные сигнальные молекулы
Оксид азота (NO) и монооксид углерода (CO) — это газообразные сигнальные молекулы, обладающие уникальными свойствами:
- Легко диффундируют через мембраны
- Имеют очень короткое время жизни
- Действуют локально
- Не требуют рецепторов на поверхности клетки
Как работает передача сигнала с помощью NO?
- NO синтезируется ферментом NO-синтазой
- Диффундирует в соседние клетки
- Активирует фермент гуанилатциклазу
- Повышается уровень циклического GMP
- Запускаются физиологические эффекты (например, расслабление гладких мышц сосудов)
NO играет важную роль в регуляции тонуса сосудов, нейротрансмиссии и иммунных реакциях.
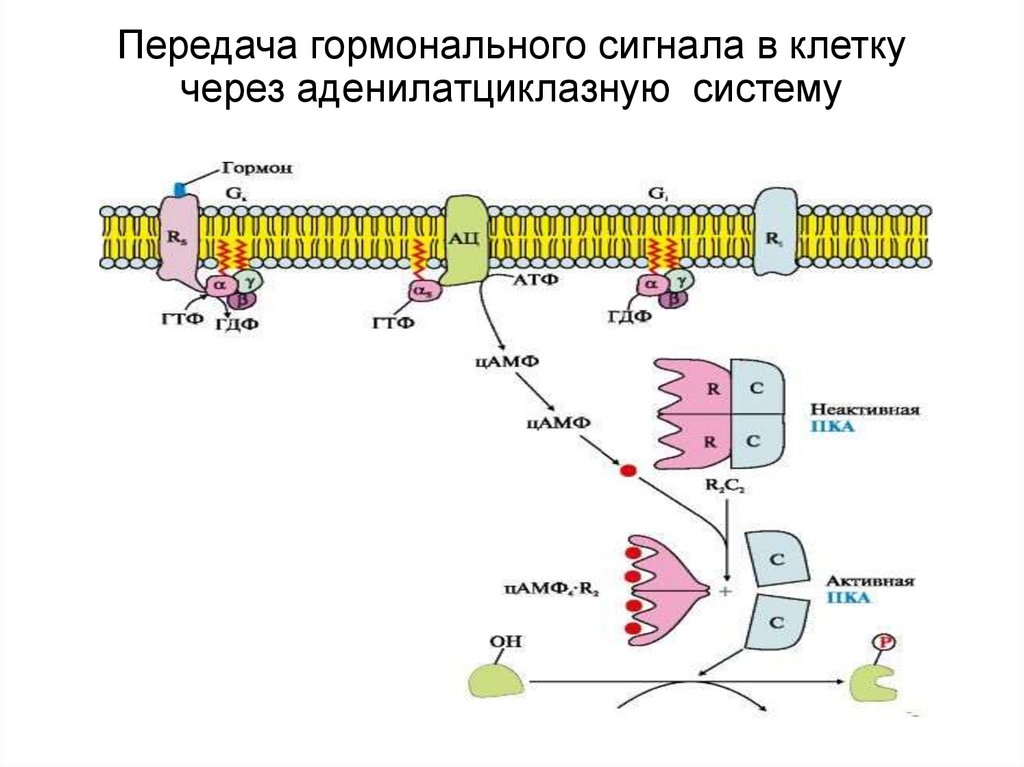
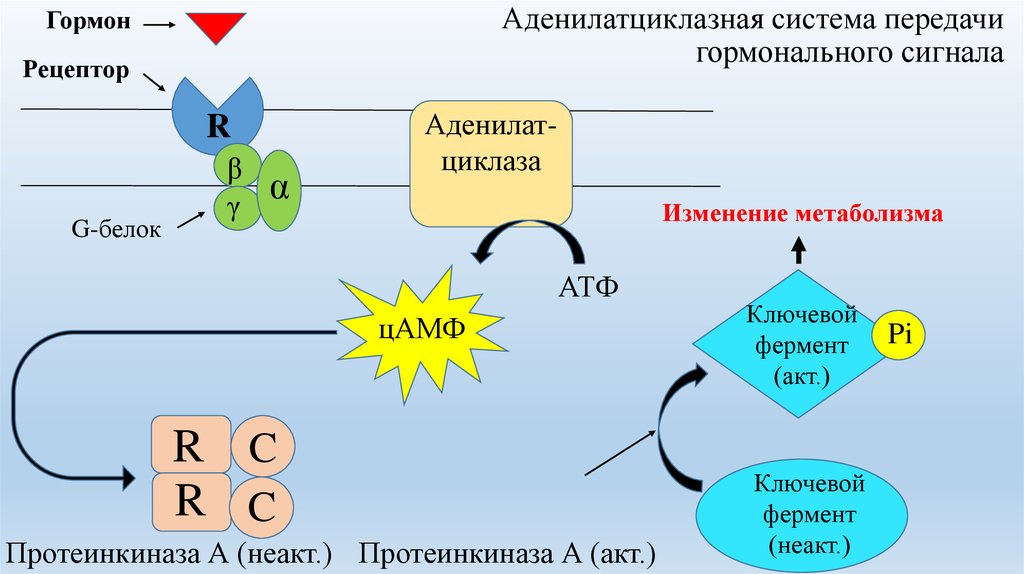
Механизмы передачи сигнала внутрь клетки
При связывании сигнальных молекул с рецепторами на поверхности клетки запускаются различные механизмы передачи сигнала внутрь:
- Активация G-белков и вторичных мессенджеров (цАМФ, IP3, DAG)
- Активация рецепторных тирозинкиназ
- Активация ионных каналов
- Активация JAK-STAT сигнального пути
Эти механизмы приводят к изменениям активности ферментов, транскрипционных факторов и других внутриклеточных белков, что в итоге определяет клеточный ответ на внешний сигнал.
Регуляция межклеточной сигнализации
Передача сигналов между клетками жестко регулируется на разных уровнях:
- Синтез и секреция сигнальных молекул
- Экспрессия рецепторов на клетках-мишенях
- Десенситизация рецепторов
- Деградация сигнальных молекул
- Обратная связь через внутриклеточные сигнальные пути
Нарушения в регуляции межклеточной сигнализации могут приводить к различным патологическим состояниям, включая рак, аутоиммунные и эндокринные заболевания.
Заключение
Межклеточная сигнализация — это сложный и многоуровневый процесс, обеспечивающий согласованную работу клеток в многоклеточном организме. Понимание механизмов передачи сигналов между клетками имеет огромное значение для биологии, медицины и разработки новых лекарственных препаратов.
Передача сигнала по кабелю — Актив СБ
- Каталог
- Передача сигнала по кабелю
(707)
Приемопередатчики видеосигнала
Разветвители и усилители видеосигнала
Удлинители интерфейса
Усилители и разветвители интерфейсов
Модуляторы видеосигнала на антенный вход
Удлинители Ethernet
Коммутаторы
Устройства грозозащиты
Тестеры для камер
Изоляторы кабеля, фильтры
Коммутаторы матричные
Медиаконвертеры
Преобразователи (конверторы) интерфейсов
Преобразователи интерфейса беспроводные
Приемопередатчики аудиосигнала
Трансиверы (SFP-модули)
Доп.
 оборудование для передачи данных
оборудование для передачи данных
 оборудование для передачи данных
оборудование для передачи данныхСистема приема и передачи сигналов состоит из:
- Передатчик сигнала — устройство, осуществляющее передачу какого-либо сигнала;
- Приемник сигнала — устройство, осуществляющее прием какого-либо сигнала;
- Приемопередатчик — устройство, способное осуществлять как прием, так и передачу сигнала;
- Разветвитель — устройство, имеющее на один сигнальный вход несколько выходов;
- Усилитель — устройство, увеличивающее уровень сигнала для передачи его на большее расстояние;
- Пассивное устройство передачи сигнала — устройство, не требующее для работы внешнего питания;
- Активное устройство передачи сигнала — устройство, требующее подачи внешнего элеткропитания.
Передача сигнала по кабелю полезная информация:
Пять вопросов по передаче видеосигнала
Нетривиальными вопросами при построении системы передачи видеосигнала могут стать выбор типа кабеля и сетевого стандарта, развитие существующей кабельной инфраструктуры на объекте, организация беспроводных сегментов сети и совместимость сетевого оборудования
Особенности передачи аналогового видеосигнала и организации питания видеокамер
Многие, скрупулёзно выбирая по характеристикам видеокамеры и видеорегистраторы, не думают о том, какие линии передачи будут использоваться, чтобы донести все эти телевизионные линии до средств видеорегистрации и отображения в высоком качестве и без помех. В данной статье рассмотрим вопросы, которые возникают при организации питания и передачи аналогового видеосигнала, на которые часто не обращают внимания, особенно если система видеонаблюдения, как часто бывает в последнее время, создается без этапа проектирования
В данной статье рассмотрим вопросы, которые возникают при организации питания и передачи аналогового видеосигнала, на которые часто не обращают внимания, особенно если система видеонаблюдения, как часто бывает в последнее время, создается без этапа проектирования
Сеть передачи данных: транспортная инфраструктура или «страшилка» для безопасников?
Глядя на отчеты компаний-производителей IP-видеонаблюдения за последний год, кажется, что всеобщая IP-революция, стремительно «бежавшая» по стране, сейчас идет медленно и неуверенно.
Передача сигналов на большие расстояния в условиях электромагнитных помех
В последние десятилетия проблема взаимного воздействия на радиоэлектронные устройства непреднамеренных помех (специалисты говорят о проблеме электромагнитной совместимости, ЭМС) стала настолько острой, что иногда для обеспечения ЭМС приходится искусственно снижать технические характеристики аппаратуры.
Передача сигнала по кабелю витой пары
Специалисты в области видеонаблюдения знают, что получить качественный видео- и аудио сигнал – это еще полдела. Сложности возникают, если необходимо передать данные системы видеонаблюедния на десятки и сотни метров без искажений и помех.
Сложности возникают, если необходимо передать данные системы видеонаблюедния на десятки и сотни метров без искажений и помех.
Помехи, шумы, наводки и как с ними бороться.
Построение любой системы вещательного уровня обязательно требует обеспечения достаточно низкого уровня шумов как в тракте звуковых, так и видеосигналов. Как всякие электрические сигналы, помехи, наводки и шумы имеют вполне конкретные источники и пути проникновения в тракт сигнала. Попробуем разобраться с основными видами помех и типичными методами борьбы с ними.
Бесконтактная индуктивная передача сигналов | Каталог
Каталог товаров
- Аксессуары
- Блоки питания для ПЛК и датчиков
- Датчики и преобразователи давления промышленные
- Датчики линейных перемещений и расстояний
- Датчики оптические для специальных задач
- Датчики положения
- Датчики, реле потока воздуха и промышленных газов
- Датчики, реле потока жидкости
- Датчики температуры промышленные
- Датчики ускорения — акселерометры
- Датчики угла наклона — инклинометры
- Датчики угловых перемещений, энкодеры, потенциометры
- Датчики щелевые
- Индикаторы, преобразователи и регуляторы
- Мониторинг и диагностика
- Расходомеры жидкости
- Расходомеры сжатого воздуха и промышленных газов
- Сигнализаторы уровня жидкости и сыпучих веществ
- Сканеры штрих-кодов промышленные
- RFID компоненты
- Уровнемеры
- Сигнальное оборудование
- Безопасность на производстве
- Промышленные интерфейсы
- Системы освещения на производстве
-
Бесконтактные системы передачи данных
- Оптическая передача данных
- Индуктивная передача сигналов
Тип элемента
Выберитевторичный — удаленныйпервичный — база
Расстояние передачи
Выберитедо 3 ммдо 5 ммдо 10 мм
Сигнал передачи
ВыберитеPNP
Интерфейс
ВыберитеIO-Link
Сбросить фильтр
Выбор производителя
- Все
- Balluff
- Pepperl+Fuchs
- Turck
Каталог индуктивных систем связи, предназначенных для бесконтактной передачи сигналов от датчиков положения в условиях промышленного производства на расстояния до нескольких миллиметров. Индуктивные устройства связи Balluff, Pepperl+Fuchs позволяют принимать и передавать сигнал от нескольких датчиков.
Индуктивные устройства связи Balluff, Pepperl+Fuchs позволяют принимать и передавать сигнал от нескольких датчиков.
Товаров на странице: 24 48 96 Все
Сортировать: По наличию По умолчанию
BIC 1I1-P2A05-M12MM-BPX0-003-M45A | BIC0077 элемент связи индуктивный
Срок поставки:
4-7 недель
BIC 2I1-P2A05-M12MF-BPX0-003-M44A | BIC0078 элемент связи индуктивный
Срок поставки:
4-7 недель
BIC 1I3-P2A50-M30MI3-SM4ACA | BIC0009 элемент связи индуктивный
Срок поставки:
4-7 недель
BIC 2I3-P2A50-M30MI3-SM4ACA | BIC000A элемент связи индуктивный
Срок поставки:
4-7 недель
BIC 2I22-P2A02-M18MF2-EPX07-050 | BIC007U элемент связи индуктивный
Срок поставки:
4-7 недель
BIC 1I22-P2A02-M18MN2-EPX07-050 | BIC007T элемент связи индуктивный
Срок поставки:
4-7 недель
BIC 2I3-P2A50-M30MI3-BPX0C-002-M4CA | BIC005J элемент связи индуктивный
Срок поставки:
4-7 недель
BIC 1I3-P2A50-Q40KFU-EPX0-002-M4CA | BIC007J элемент связи индуктивный
Срок поставки:
4-7 недель
NICP-M30-IOL2P8X-h2141 | 4300101 элемент связи индуктивный
Срок поставки:
2-4 недели
NICS-M30-IOL2P8-0.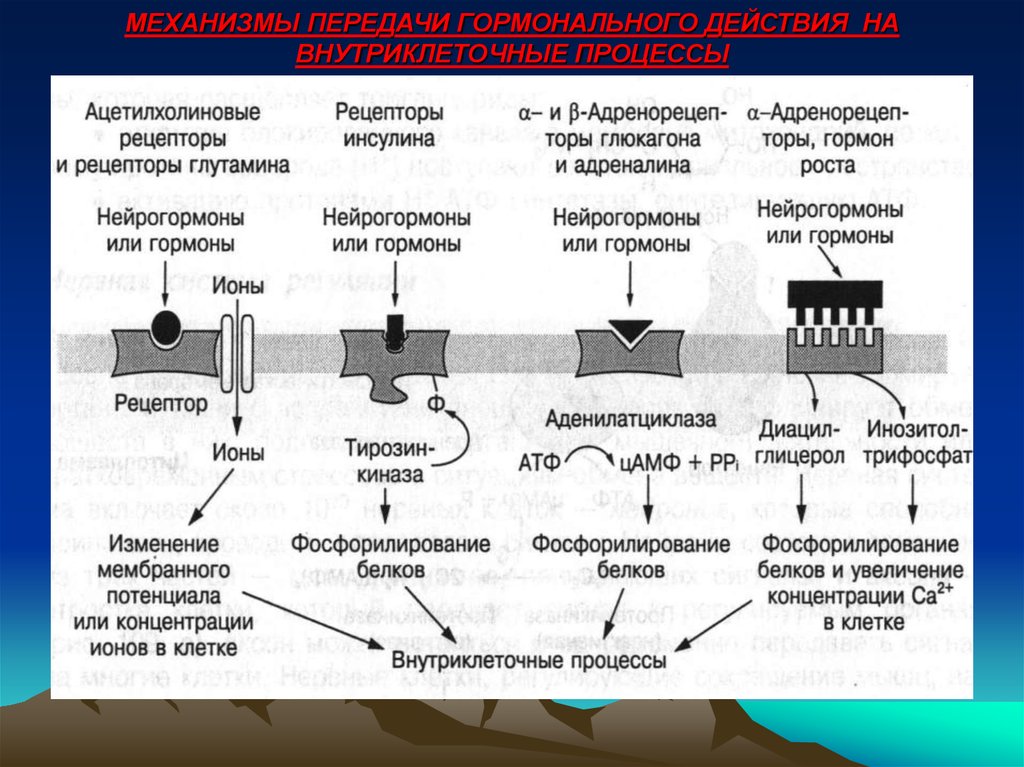 3-RKC4.4T | 4300301 элемент связи индуктивный
3-RKC4.4T | 4300301 элемент связи индуктивный
Срок поставки:
2-4 недели
BIC 2B0-ITA50-M30MF1-SM4A5A | BIC007E элемент связи индуктивный
Срок поставки:
4-6 недель
BIC 1B0-ITA50-M30MF1-SM4A5A | BIC007L элемент связи индуктивный
Срок поставки:
4-7 недель
дополнительная информация
|
Индуктивная система передачи сигнала решает частую проблему на автоматизированном производстве, которая заключается в соединении датчиков с подвижными элементами механизмов машин. Стандартные решения в виде кабелей обладают ограниченным сроком службы, так как постоянное механическое воздействие, сгибание проводов и периодическое движение выводит из строя проводники и контакты. При помощи беспроводной связи для подключения датчиков можно избежать проблем, связанных с кабелями. Системы работают по принципу индуктивной связи и состоят из передатчика и приемника с модулем. |
 Датчики положения, подключенные к передатчику, получают от него питание. Для передачи электроэнергии на стороне главного компонента создаётся индуктивное переменное поле. Когда вторичная цепь входит в это поле, напряжение индуцируется через отрицательную обратную связь, и таким образом происходит передача электроэнергии. Величины энергии вполне достаточно для функционирования индуктивных, оптических или ультразвуковых датчиков прямо на подвижных элементах механизмов. Сигналы датчиков передаются на сторону приемника посредством отдельных импульсов передачи и, таким образом, становятся доступными системе управления. Для заказа доступны индуктивные системы передачи сигналов Balluff BIC, Pepperl+Fuchs WIS.
Датчики положения, подключенные к передатчику, получают от него питание. Для передачи электроэнергии на стороне главного компонента создаётся индуктивное переменное поле. Когда вторичная цепь входит в это поле, напряжение индуцируется через отрицательную обратную связь, и таким образом происходит передача электроэнергии. Величины энергии вполне достаточно для функционирования индуктивных, оптических или ультразвуковых датчиков прямо на подвижных элементах механизмов. Сигналы датчиков передаются на сторону приемника посредством отдельных импульсов передачи и, таким образом, становятся доступными системе управления. Для заказа доступны индуктивные системы передачи сигналов Balluff BIC, Pepperl+Fuchs WIS.Сигнальные молекулы и их рецепторы — Клетка
Множество различных видов молекул передают информацию между клетками многоклеточных организмов. Хотя все эти молекулы действуют как лиганды, которые связываются с рецепторами, экспрессируемыми их клетками-мишенями, существуют значительные различия в структуре и функциях различных типов молекул, которые служат передатчиками сигналов.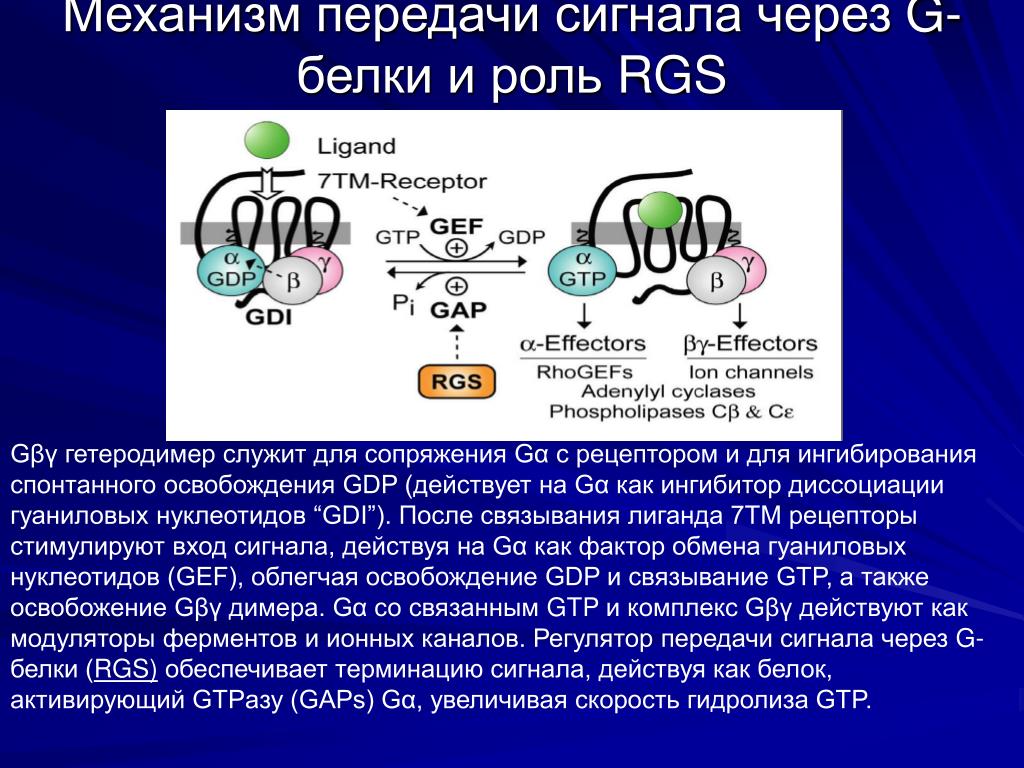 Структурно сигнальные молекулы, используемые растениями и животными, варьируются по сложности от простых газов до белков. Некоторые из этих молекул передают сигналы на большие расстояния, тогда как другие действуют локально, передавая информацию между соседними клетками. Кроме того, сигнальные молекулы различаются по способу действия на клетки-мишени. Некоторые сигнальные молекулы способны пересекать плазматическую мембрану и связываться с внутриклеточными рецепторами в цитоплазме или ядре, тогда как большинство связывается с рецепторами, экспрессируемыми на поверхности клетки-мишени. В следующих разделах обсуждаются основные типы сигнальных молекул и рецепторы, с которыми они взаимодействуют. Последующее обсуждение в этой главе сосредоточено на механизмах, с помощью которых рецепторы клеточной поверхности функционируют, чтобы регулировать поведение клеток.
Структурно сигнальные молекулы, используемые растениями и животными, варьируются по сложности от простых газов до белков. Некоторые из этих молекул передают сигналы на большие расстояния, тогда как другие действуют локально, передавая информацию между соседними клетками. Кроме того, сигнальные молекулы различаются по способу действия на клетки-мишени. Некоторые сигнальные молекулы способны пересекать плазматическую мембрану и связываться с внутриклеточными рецепторами в цитоплазме или ядре, тогда как большинство связывается с рецепторами, экспрессируемыми на поверхности клетки-мишени. В следующих разделах обсуждаются основные типы сигнальных молекул и рецепторы, с которыми они взаимодействуют. Последующее обсуждение в этой главе сосредоточено на механизмах, с помощью которых рецепторы клеточной поверхности функционируют, чтобы регулировать поведение клеток.
Способы передачи сигналов между клетками
Передача сигналов между клетками может быть результатом либо прямого взаимодействия клетки с ее соседом, либо действия секретируемых сигнальных молекул (). Передача сигналов посредством прямых межклеточных (или клеточно-матриксных) взаимодействий играет критическую роль в регуляции поведения клеток в тканях животных. Например, интегрины и кадгерины (которые обсуждались в предыдущей главе) функционируют не только как молекулы клеточной адгезии, но и как сигнальные молекулы, которые регулируют пролиферацию и выживание клеток в ответ на межклеточные и межклеточные контакты. Кроме того, клетки экспрессируют множество рецепторов клеточной поверхности, которые взаимодействуют с сигнальными молекулами на поверхности соседних клеток. Передача сигналов посредством таких прямых межклеточных взаимодействий играет критическую роль в регуляции многих взаимодействий между различными типами клеток, происходящих во время эмбрионального развития, а также в поддержании тканей взрослого организма.
Передача сигналов посредством прямых межклеточных (или клеточно-матриксных) взаимодействий играет критическую роль в регуляции поведения клеток в тканях животных. Например, интегрины и кадгерины (которые обсуждались в предыдущей главе) функционируют не только как молекулы клеточной адгезии, но и как сигнальные молекулы, которые регулируют пролиферацию и выживание клеток в ответ на межклеточные и межклеточные контакты. Кроме того, клетки экспрессируют множество рецепторов клеточной поверхности, которые взаимодействуют с сигнальными молекулами на поверхности соседних клеток. Передача сигналов посредством таких прямых межклеточных взаимодействий играет критическую роль в регуляции многих взаимодействий между различными типами клеток, происходящих во время эмбрионального развития, а также в поддержании тканей взрослого организма.
Рисунок 13.1
Режимы передачи сигналов между ячейками. Передача сигналов между клетками может происходить либо посредством прямых межклеточных контактов, либо посредством действия секретируемых сигнальных молекул.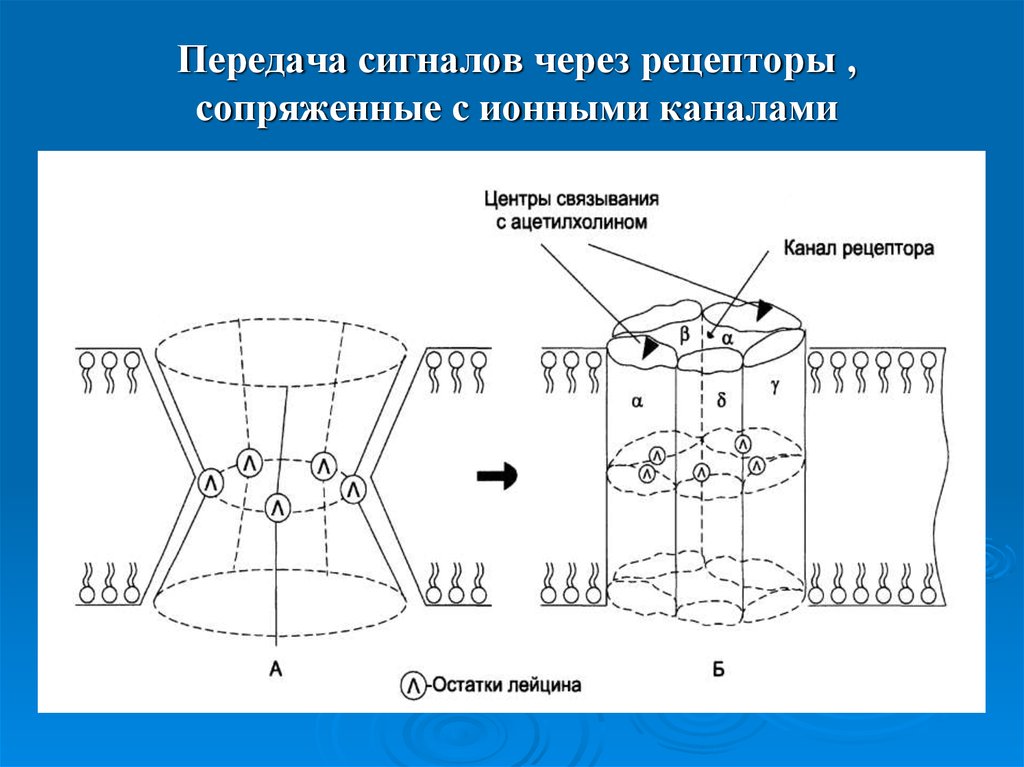 (A) При эндокринной передаче гормоны переносятся через систему кровообращения для воздействия на отдаленные (далее…)
(A) При эндокринной передаче гормоны переносятся через систему кровообращения для воздействия на отдаленные (далее…)
Многочисленные разновидности передачи сигналов секретируемыми молекулами часто делятся на три основные категории в зависимости от расстояния, на которое передаются сигналы . При эндокринной передаче сигналов сигнальные молекулы (гормоны) секретируются специализированными эндокринными клетками и переносятся кровотоком, воздействуя на клетки-мишени в отдаленных участках тела. Классический пример — стероидный гормон эстроген, вырабатываемый яичниками и стимулирующий развитие и поддержание женской репродуктивной системы и вторичных половых признаков. У животных более 50 различных гормонов вырабатываются эндокринными железами, включая гипофиз, щитовидную железу, паращитовидную железу, поджелудочную железу, надпочечники и половые железы.
В отличие от гормонов, некоторые сигнальные молекулы действуют локально, воздействуя на поведение близлежащих клеток. При паракринной передаче сигналов молекула, высвобождаемая одной клеткой, действует на соседние клетки-мишени. Примером может служить действие нейротрансмиттеров при переносе сигналов между нервными клетками в синапсе. Наконец, некоторые клетки реагируют на сигнальные молекулы, которые они сами производят. Одним из важных примеров такой аутокринной передачи сигналов является ответ клеток иммунной системы позвоночных на чужеродные антигены. Определенные типы Т-лимфоцитов реагируют на антигенную стимуляцию, синтезируя фактор роста, который стимулирует их собственную пролиферацию, тем самым увеличивая количество реагирующих Т-лимфоцитов и усиливая иммунный ответ. Следует также отметить, что аномальная аутокринная передача сигналов часто способствует неконтролируемому росту раковых клеток (см. главу 15). В этой ситуации раковая клетка вырабатывает фактор роста, на который она также реагирует, тем самым непрерывно стимулируя собственную нерегулируемую пролиферацию.
Примером может служить действие нейротрансмиттеров при переносе сигналов между нервными клетками в синапсе. Наконец, некоторые клетки реагируют на сигнальные молекулы, которые они сами производят. Одним из важных примеров такой аутокринной передачи сигналов является ответ клеток иммунной системы позвоночных на чужеродные антигены. Определенные типы Т-лимфоцитов реагируют на антигенную стимуляцию, синтезируя фактор роста, который стимулирует их собственную пролиферацию, тем самым увеличивая количество реагирующих Т-лимфоцитов и усиливая иммунный ответ. Следует также отметить, что аномальная аутокринная передача сигналов часто способствует неконтролируемому росту раковых клеток (см. главу 15). В этой ситуации раковая клетка вырабатывает фактор роста, на который она также реагирует, тем самым непрерывно стимулируя собственную нерегулируемую пролиферацию.
Стероидные гормоны и суперсемейство стероидных рецепторов
Как уже отмечалось, все сигнальные молекулы действуют путем связывания с рецепторами, экспрессируемыми их клетками-мишенями.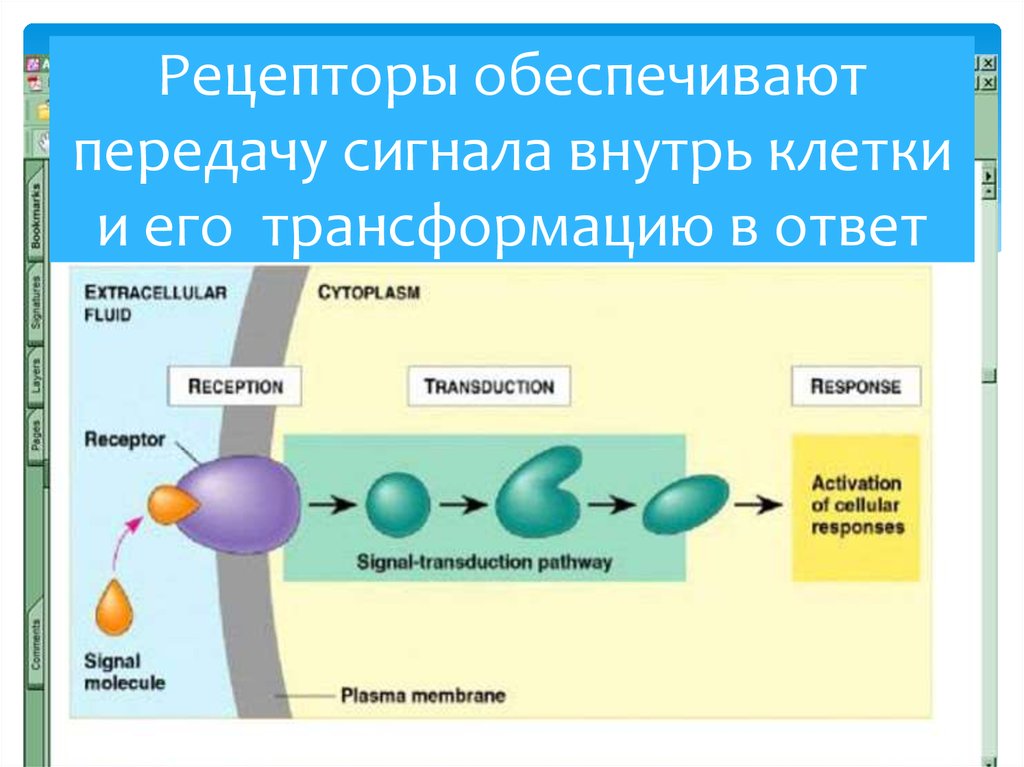 Во многих случаях эти рецепторы экспрессируются на поверхности клетки-мишени, но некоторые рецепторы представляют собой внутриклеточные белки, расположенные в цитозоле или ядре. Эти внутриклеточные рецепторы реагируют на небольшие гидрофобные сигнальные молекулы, способные диффундировать через плазматическую мембрану. Стероидные гормоны являются классическими примерами этой группы сигнальных молекул, в которую также входят гормоны щитовидной железы, витамин D 3 и ретиноевая кислота ().
Во многих случаях эти рецепторы экспрессируются на поверхности клетки-мишени, но некоторые рецепторы представляют собой внутриклеточные белки, расположенные в цитозоле или ядре. Эти внутриклеточные рецепторы реагируют на небольшие гидрофобные сигнальные молекулы, способные диффундировать через плазматическую мембрану. Стероидные гормоны являются классическими примерами этой группы сигнальных молекул, в которую также входят гормоны щитовидной железы, витамин D 3 и ретиноевая кислота ().
Рисунок 13.2
Структура стероидных гормонов, гормона щитовидной железы, витамина D 3 и ретиноевой кислоты. К стероидам относятся половые гормоны (тестостерон, эстроген и прогестерон), глюкокортикоиды и минералокортикоиды.
Все стероидные гормоны (включая тестостерон, эстроген, прогестерон, кортикостероиды и экдизон) синтезируются из холестерина. Тестостерон, эстроген и прогестерон — это половые стероиды, которые вырабатываются гонадами. Кортикостероиды вырабатываются надпочечниками. К ним относятся глюкокортикоиды , которые действуют на различные клетки, стимулируя выработку глюкозы, и минералокортикоиды , которые действуют на почки, регулируя солевой и водный баланс. Экдизон — это гормон насекомых, который играет ключевую роль в развитии, запуская метаморфоз личинок во взрослых особей.
К ним относятся глюкокортикоиды , которые действуют на различные клетки, стимулируя выработку глюкозы, и минералокортикоиды , которые действуют на почки, регулируя солевой и водный баланс. Экдизон — это гормон насекомых, который играет ключевую роль в развитии, запуская метаморфоз личинок во взрослых особей.
Хотя гормон щитовидной железы, витамин D 3 и ретиноевая кислота структурно и функционально отличаются от стероидов, они имеют общий механизм действия в клетках-мишенях. Гормон щитовидной железы синтезируется из тирозина в щитовидной железе; он играет важную роль в развитии и регуляции метаболизма. Витамин D 3 регулирует метаболизм Ca 2+ и рост костей. Ретиноевая кислота и родственные соединения ( ретиноиды ), синтезируемые из витамина А, играют важную роль в развитии позвоночных.
Из-за своего гидрофобного характера стероидные гормоны, гормоны щитовидной железы, витамин D 3 и ретиноевая кислота способны проникать в клетки путем диффузии через плазматическую мембрану ().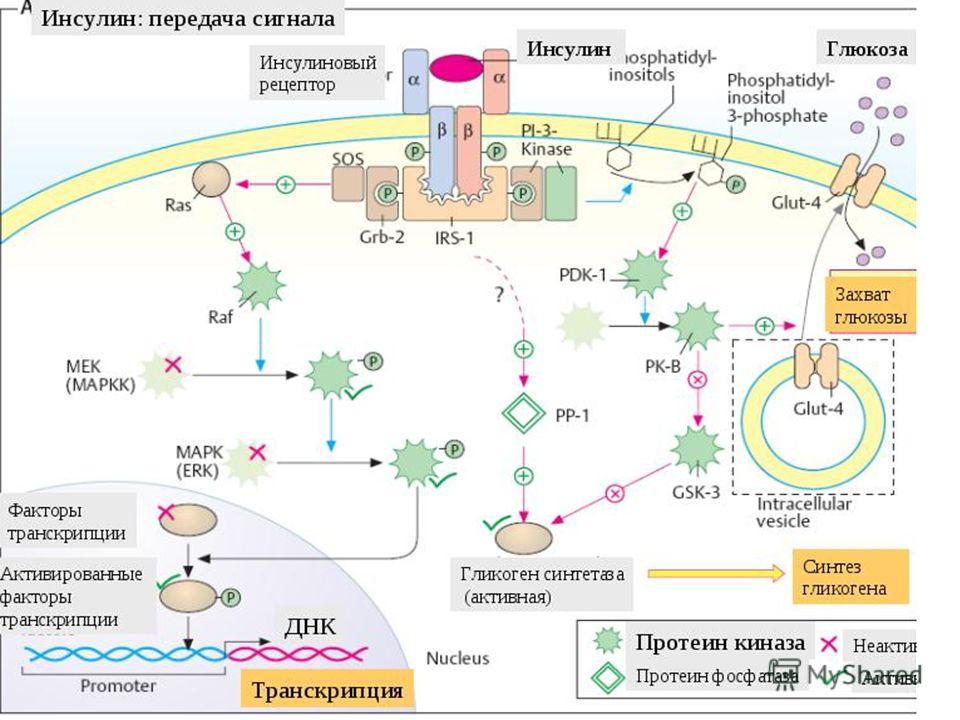 Оказавшись внутри клетки, они связываются с внутриклеточными рецепторами, которые экспрессируются гормонально чувствительными клетками-мишенями. Эти рецепторы, которые являются членами семейства белков, известного как суперсемейство стероидных рецепторов, являются факторами транскрипции, которые содержат родственные домены для связывания лиганда, связывания ДНК и активации транскрипции. Связывание лигандов регулирует их функцию активаторов или репрессоров генов-мишеней, поэтому стероидные гормоны и родственные им молекулы напрямую регулируют экспрессию генов.
Оказавшись внутри клетки, они связываются с внутриклеточными рецепторами, которые экспрессируются гормонально чувствительными клетками-мишенями. Эти рецепторы, которые являются членами семейства белков, известного как суперсемейство стероидных рецепторов, являются факторами транскрипции, которые содержат родственные домены для связывания лиганда, связывания ДНК и активации транскрипции. Связывание лигандов регулирует их функцию активаторов или репрессоров генов-мишеней, поэтому стероидные гормоны и родственные им молекулы напрямую регулируют экспрессию генов.
Рисунок 13.3
Действие стероидных гормонов. Стероидные гормоны диффундируют через плазматическую мембрану и связываются с ядерными рецепторами, которые непосредственно стимулируют транскрипцию генов-мишеней. Рецепторы стероидных гормонов связываются с ДНК в виде димеров.
Связывание лиганда оказывает различное воздействие на разные рецепторы. Некоторые члены надсемейства стероидных рецепторов, такие как эстрогеновые и глюкокортикоидные рецепторы, неспособны связываться с ДНК в отсутствие гормона. Связывание гормона вызывает конформационные изменения рецептора, позволяя ему связываться с регуляторными последовательностями ДНК и активировать транскрипцию генов-мишеней. В других случаях рецептор связывается с ДНК либо в присутствии, либо в отсутствие гормона, но связывание гормона изменяет активность рецептора как молекулы, регулирующей транскрипцию. Например, рецептор тиреоидного гормона действует как репрессор в отсутствие гормона, но связывание гормона превращает его в активатор, который стимулирует транскрипцию генов, индуцируемых тиреоидным гормоном.
Связывание гормона вызывает конформационные изменения рецептора, позволяя ему связываться с регуляторными последовательностями ДНК и активировать транскрипцию генов-мишеней. В других случаях рецептор связывается с ДНК либо в присутствии, либо в отсутствие гормона, но связывание гормона изменяет активность рецептора как молекулы, регулирующей транскрипцию. Например, рецептор тиреоидного гормона действует как репрессор в отсутствие гормона, но связывание гормона превращает его в активатор, который стимулирует транскрипцию генов, индуцируемых тиреоидным гормоном.
Рисунок 13.4
Генная регуляция рецептором гормона щитовидной железы. Рецептор гормона щитовидной железы связывается с ДНК как в присутствии, так и в отсутствие гормона. Однако связывание гормона меняет функцию рецептора с репрессора на активатор транскрипции гена-мишени. (подробнее…)
Окись азота и окись углерода
Простой газообразный оксид азота (NO) является основной паракринной сигнальной молекулой в нервной, иммунной и сердечно-сосудистой системах. Подобно стероидным гормонам, NO способен диффундировать непосредственно через плазматическую мембрану своих клеток-мишеней. Однако молекулярная основа действия NO отличается от действия стероидов; вместо связывания с рецептором, регулирующим транскрипцию, NO изменяет активность внутриклеточных ферментов-мишеней.
Подобно стероидным гормонам, NO способен диффундировать непосредственно через плазматическую мембрану своих клеток-мишеней. Однако молекулярная основа действия NO отличается от действия стероидов; вместо связывания с рецептором, регулирующим транскрипцию, NO изменяет активность внутриклеточных ферментов-мишеней.
Оксид азота синтезируется из аминокислоты аргинина с помощью фермента синтазы оксида азота (). После синтеза NO диффундирует из клетки и может действовать локально, воздействуя на близлежащие клетки. Его действие ограничивается такими локальными эффектами, потому что NO чрезвычайно нестабилен, а его период полураспада составляет всего несколько секунд. Одним хорошо охарактеризованным примером действия NO является сигнал о расширении кровеносных сосудов. Первым шагом в этом процессе является высвобождение нейротрансмиттеров, таких как ацетилхолин, из окончаний нервных клеток в стенке кровеносного сосуда. Эти нейротрансмиттеры действуют на эндотелиальные клетки, стимулируя синтез NO. Затем NO диффундирует в соседние гладкомышечные клетки, где вступает в реакцию с железом, связанным с активным центром фермента гуанилатциклазы. Это увеличивает ферментативную активность, что приводит к синтезу вторичного мессенджера циклического GMP (обсуждается далее в этой главе), который вызывает расслабление мышечных клеток и расширение кровеносных сосудов. Например, NO отвечает за передачу сигналов о расширении кровеносных сосудов, что приводит к эрекции полового члена. Интересно также отметить, что медицинское использование нитроглицерина для лечения сердечных заболеваний основано на его превращении в NO, который расширяет коронарные сосуды и увеличивает приток крови к сердцу.
Затем NO диффундирует в соседние гладкомышечные клетки, где вступает в реакцию с железом, связанным с активным центром фермента гуанилатциклазы. Это увеличивает ферментативную активность, что приводит к синтезу вторичного мессенджера циклического GMP (обсуждается далее в этой главе), который вызывает расслабление мышечных клеток и расширение кровеносных сосудов. Например, NO отвечает за передачу сигналов о расширении кровеносных сосудов, что приводит к эрекции полового члена. Интересно также отметить, что медицинское использование нитроглицерина для лечения сердечных заболеваний основано на его превращении в NO, который расширяет коронарные сосуды и увеличивает приток крови к сердцу.
Рисунок 13.5
Синтез оксида азота. Фермент синтаза оксида азота (NOS) катализирует образование оксида азота из аргинина.
Другой простой газ, окись углерода (СО), также действует как сигнальная молекула в нервной системе. CO тесно связан с NO и, по-видимому, действует аналогично как нейротрансмиттер и медиатор расширения кровеносных сосудов.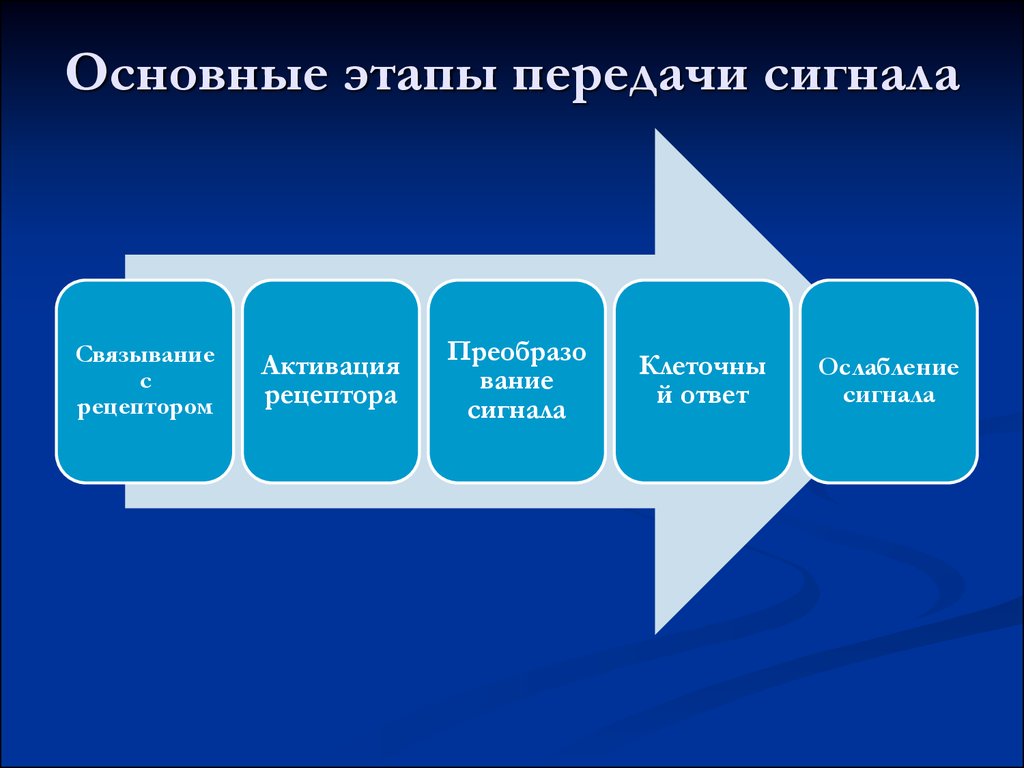 Синтез СО в клетках мозга, как и NO, стимулируется нейротрансмиттерами. Кроме того, CO может стимулировать гуанилатциклазу, которая также может представлять собой основную физиологическую мишень передачи сигналов CO.
Синтез СО в клетках мозга, как и NO, стимулируется нейротрансмиттерами. Кроме того, CO может стимулировать гуанилатциклазу, которая также может представлять собой основную физиологическую мишень передачи сигналов CO.
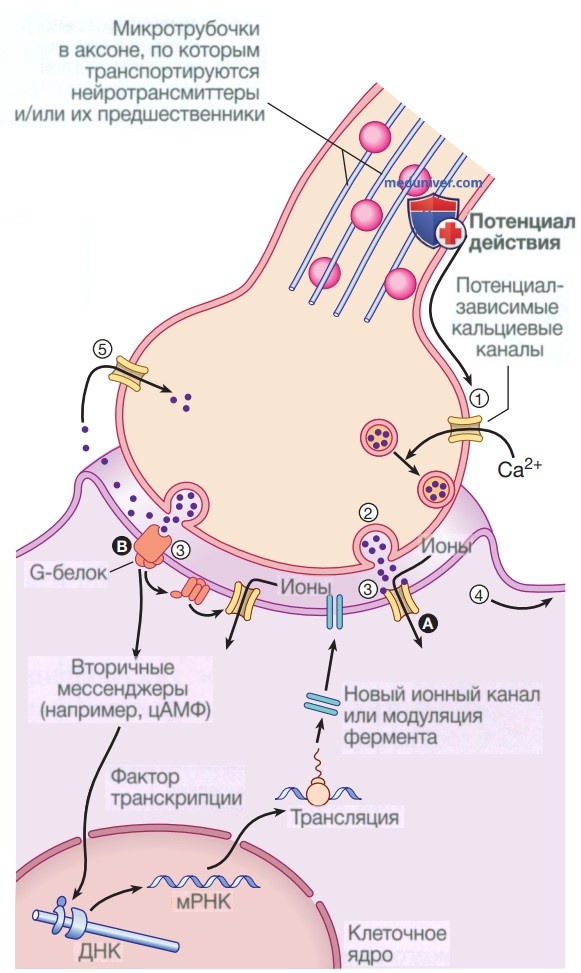
Нейротрансмиттеры
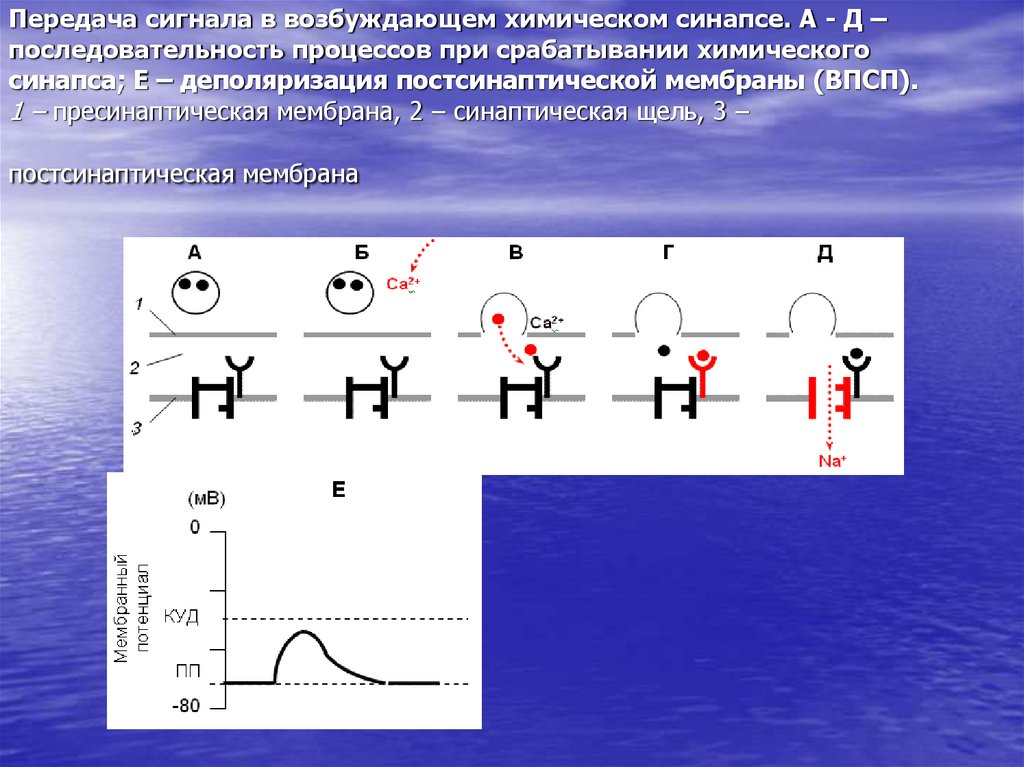
Нейротрансмиттеры передают сигналы между нейронами или от нейронов к другим типам клеток-мишеней (таким как мышечные клетки). Они представляют собой разнообразную группу небольших гидрофильных молекул, включая ацетилхолин, дофамин, эпинефрин (адреналин), серотонин, гистамин, глутамат, глицин и γ-аминомасляную кислоту (ГАМК) (). Высвобождение нейротрансмиттеров сигнализируется появлением потенциала действия на конце нейрона (см. Ресурсы). Затем нейротрансмиттеры диффундируют через синаптическую щель и связываются с рецепторами на поверхности клетки-мишени. Обратите внимание, что некоторые нейротрансмиттеры также могут действовать как гормоны. Например, адреналин действует как нейротрансмиттер и как гормон, вырабатываемый надпочечниками, сигнализирующий о распаде гликогена в мышечных клетках.
Рисунок 13.6
Структура репрезентативных нейротрансмиттеров. Нейротрансмиттеры представляют собой гидрофильные молекулы, которые связываются с рецепторами клеточной поверхности.
Поскольку нейротрансмиттеры представляют собой гидрофильные молекулы, они не могут проникать через плазматическую мембрану клеток-мишеней. Следовательно, в отличие от стероидных гормонов и NO или CO, нейротрансмиттеры действуют путем связывания с рецепторами клеточной поверхности. Многие рецепторы нейротрансмиттеров представляют собой лиганд-управляемые ионные каналы, такие как ацетилхолиновый рецептор, обсуждавшийся в предыдущей главе (см. Ресурсы). Связывание нейротрансмиттера с этими рецепторами вызывает конформационные изменения, открывающие ионные каналы, что непосредственно приводит к изменениям ионного потока в клетке-мишени. Другие рецепторы нейротрансмиттеров связаны с G-белками — основной группой сигнальных молекул (обсуждаемых далее в этой главе), которые связывают рецепторы клеточной поверхности с различными внутриклеточными реакциями. В случае рецепторов нейротрансмиттеров связанные с ними G-белки часто косвенно регулируют активность ионных каналов.
В случае рецепторов нейротрансмиттеров связанные с ними G-белки часто косвенно регулируют активность ионных каналов.
Пептидные гормоны и факторы роста
Самым большим разнообразием сигнальных молекул у животных являются пептиды, размер которых варьируется от нескольких до более сотни аминокислот. В эту группу сигнальных молекул входят пептидные гормоны, нейропептиды и разнообразные полипептидные факторы роста (14). Хорошо известные примеры пептидных гормонов включают инсулин, глюкагон и гормоны, продуцируемые гипофизом (гормон роста, фолликулостимулирующий гормон, пролактин и другие).
Таблица 13.1
Типичные пептидные гормоны, нейропептиды и факторы роста.
Нейропептиды секретируются некоторыми нейронами вместо низкомолекулярных нейротрансмиттеров, обсуждавшихся в предыдущем разделе. Некоторые из этих пептидов, такие как энкефалины и эндорфины , функционируют не только как нейротрансмиттеры в синапсах, но и как нейрогормоны , действующие на отдаленные клетки. Энкефалины и эндорфины широко изучались из-за их активности в качестве естественных анальгетиков, уменьшающих болевые реакции в центральной нервной системе. Обнаруженные в ходе изучения наркозависимости, они представляют собой встречающиеся в природе соединения, которые связываются с теми же рецепторами на поверхности клеток головного мозга, что и морфин.
Энкефалины и эндорфины широко изучались из-за их активности в качестве естественных анальгетиков, уменьшающих болевые реакции в центральной нервной системе. Обнаруженные в ходе изучения наркозависимости, они представляют собой встречающиеся в природе соединения, которые связываются с теми же рецепторами на поверхности клеток головного мозга, что и морфин.
Полипептидные факторы роста включают широкий спектр сигнальных молекул, которые контролируют рост и дифференцировку клеток животных. Первый из этих факторов ( фактор роста нервов, или NGF ) был обнаружен Ритой Леви-Монтальчини в 1950-х годах. NGF является членом семейства полипептидов (называемых нейротрофинами ), которые регулируют развитие и выживание нейронов. В ходе экспериментов с NGF Стэнли Коэн по счастливой случайности обнаружил неродственный фактор (называемый эпидермальным фактором роста, или 9).0029 EGF ), который стимулирует пролиферацию клеток. EGF, полипептид из 53 аминокислот, послужил прототипом большого количества факторов роста, которые играют решающую роль в контроле пролиферации клеток животных как во время эмбрионального развития, так и во взрослых организмах.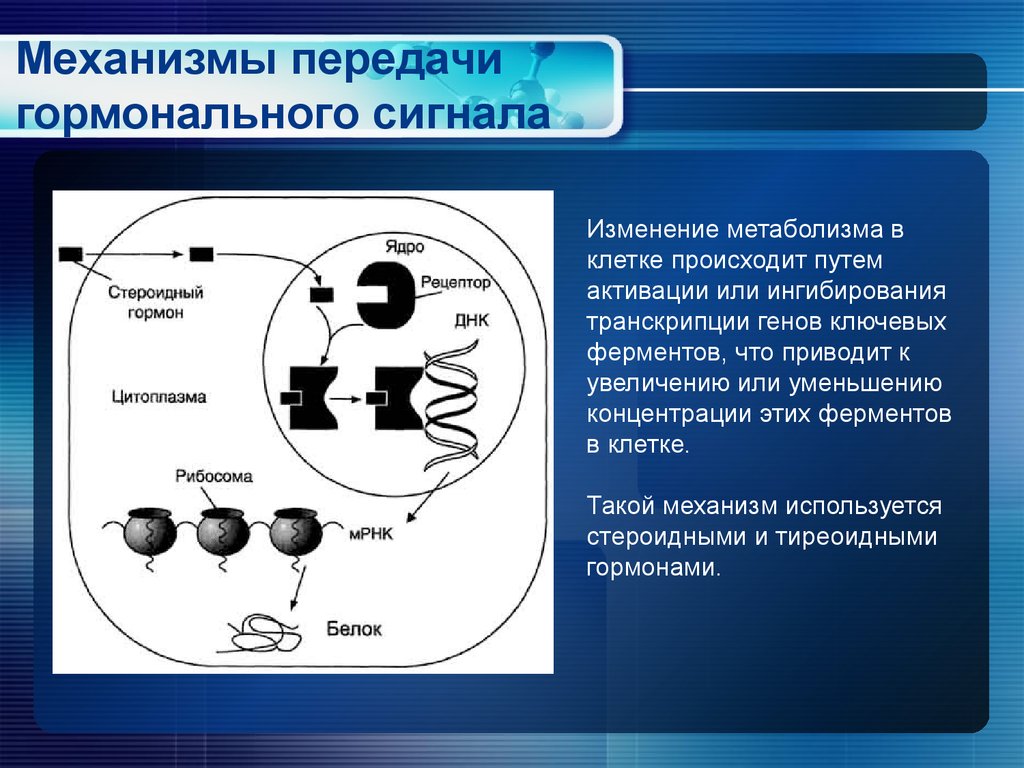
Рисунок 13.7
Структура эпидермального фактора роста (EGF). EGF представляет собой одну полипептидную цепь из 53 аминокислот. Указаны дисульфидные связи между остатками цистеина. (По Г. Карпентеру и С. Коэну, 1979. Энн. Преподобный Биохим. 48: 193.)
Хорошим примером действия фактора роста является активность тромбоцитарного фактора роста ( PDGF ) при заживлении ран. PDGF хранится в тромбоцитах крови и высвобождается при свертывании крови в месте раны. Затем он стимулирует пролиферацию фибробластов вблизи тромба, тем самым способствуя повторному росту поврежденной ткани. Члены другой большой группы полипептидных факторов роста (называемых цитокинами) регулируют развитие и дифференцировку клеток крови и контролируют активность лимфоцитов во время иммунного ответа. Другие полипептидные факторы роста (заякоренные в мембране факторы роста) остаются связанными с плазматической мембраной, а не секретируются во внеклеточные жидкости, поэтому функционируют специфически как сигнальные молекулы во время прямых межклеточных взаимодействий.
Пептидные гормоны, нейропептиды и факторы роста не способны проникать через плазматическую мембрану своих клеток-мишеней, поэтому они действуют путем связывания с рецепторами клеточной поверхности, как описано далее в этой главе. Как и следовало ожидать, исходя из критической роли полипептидных факторов роста в контроле клеточной пролиферации, аномалии в передаче сигналов факторов роста являются основой для множества заболеваний, включая многие виды рака. Например, аномальная экспрессия близкого родственника рецептора EGF является важным фактором в развитии многих видов рака груди и яичников у человека.
Эйкозаноиды
Несколько типов липидов служат сигнальными молекулами, которые, в отличие от стероидных гормонов, действуют путем связывания с рецепторами клеточной поверхности. Наиболее важные из этих молекул относятся к классу липидов, называемому эйкозаноидами, который включает простагландинов , простациклинов , тромбоксанов и лейкотриенов (). Эйкозаноиды быстро расщепляются и поэтому действуют локально в аутокринных или паракринных сигнальных путях. Они стимулируют различные реакции в клетках-мишенях, включая агрегацию тромбоцитов, воспаление и сокращение гладкой мускулатуры.
Эйкозаноиды быстро расщепляются и поэтому действуют локально в аутокринных или паракринных сигнальных путях. Они стимулируют различные реакции в клетках-мишенях, включая агрегацию тромбоцитов, воспаление и сокращение гладкой мускулатуры.
Рисунок 13.8
Синтез и структура эйкозаноидов. К эйкозаноидам относятся простагландины, простациклин, тромбоксаны и лейкотриены. Они синтезируются из арахидоновой кислоты, которая образуется при гидролизе фосфолипидов, катализируемом фосфолипазой (подробнее…)
Все эйкозаноиды синтезируются из арахидоновой кислоты, которая образуется из фосфолипидов. Первым этапом пути, ведущего к синтезу либо простагландинов, либо тромбоксанов, является превращение арахидоновой кислоты в простагландин H 9.0019 2 . Интересно, что фермент, катализирующий эту реакцию (циклооксигеназа), является мишенью для аспирина и других нестероидных противовоспалительных препаратов. Ингибируя синтез простагландинов, аспирин уменьшает воспаление и боль.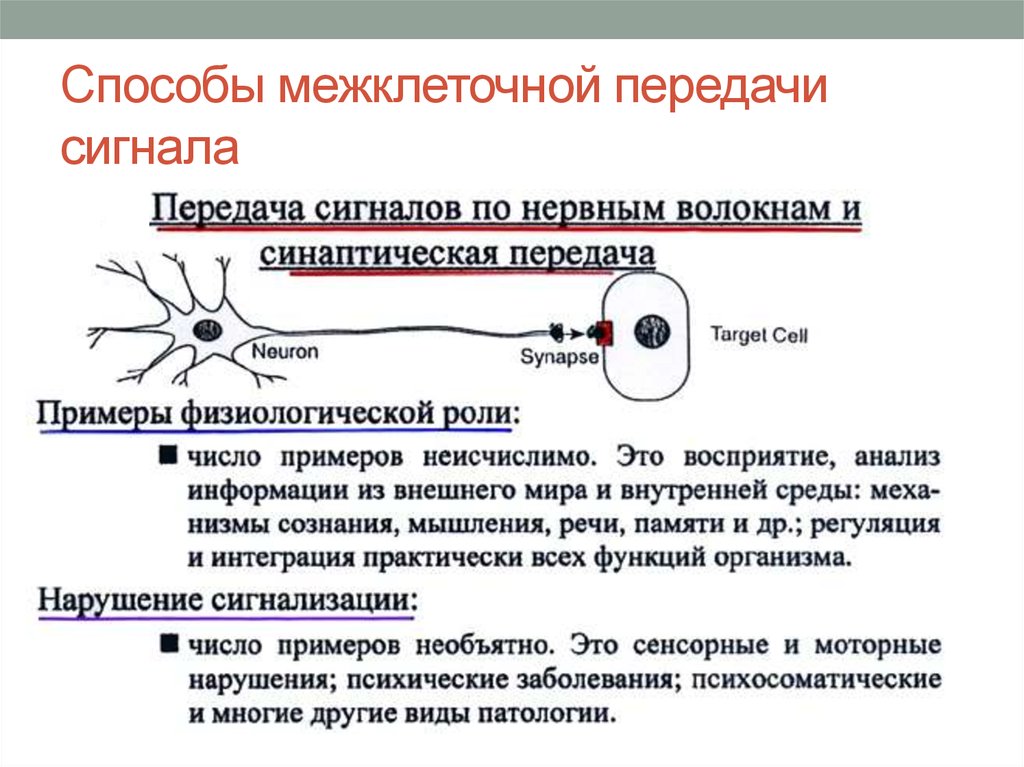 Ингибируя синтез тромбоксана, аспирин также снижает агрегацию тромбоцитов и свертываемость крови. Из-за этой активности для профилактики инсульта часто назначают небольшие суточные дозы аспирина. Кроме того, было обнаружено, что аспирин и нестероидные противовоспалительные препараты снижают частоту рака толстой кишки как у животных, так и у людей, по-видимому, за счет ингибирования синтеза простагландинов, которые стимулируют пролиферацию клеток и способствуют развитию рака.
Ингибируя синтез тромбоксана, аспирин также снижает агрегацию тромбоцитов и свертываемость крови. Из-за этой активности для профилактики инсульта часто назначают небольшие суточные дозы аспирина. Кроме того, было обнаружено, что аспирин и нестероидные противовоспалительные препараты снижают частоту рака толстой кишки как у животных, так и у людей, по-видимому, за счет ингибирования синтеза простагландинов, которые стимулируют пролиферацию клеток и способствуют развитию рака.
Гормоны растений
Рост и развитие растений регулируются группой небольших молекул, называемых растительными гормонами. Уровни этих молекул в растении обычно изменяются факторами окружающей среды, такими как свет или инфекция, поэтому они координируют реакцию тканей в разных частях растения на сигналы окружающей среды.
Гормоны растений обычно делятся на пять основных классов: ауксины , гиббереллины , цитокинины , абсцизовая кислота и этилен (), хотя недавно было обнаружено несколько дополнительных растительных гормонов. Первым идентифицированным растительным гормоном был ауксин, и первые эксперименты, приведшие к его открытию, были проведены Чарльзом Дарвином в 1880-х годах. Одним из эффектов ауксинов является индуцирование удлинения растительных клеток за счет ослабления клеточной стенки (см. Ресурсы). Кроме того, ауксины регулируют многие другие аспекты развития растений, включая деление и дифференцировку клеток. Другие растительные гормоны также оказывают множественное воздействие на ткани-мишени, включая удлинение стебля (гиббереллины), созревание плодов (этилен), деление клеток (цитокинины) и начало покоя (абсцизовая кислота).
Первым идентифицированным растительным гормоном был ауксин, и первые эксперименты, приведшие к его открытию, были проведены Чарльзом Дарвином в 1880-х годах. Одним из эффектов ауксинов является индуцирование удлинения растительных клеток за счет ослабления клеточной стенки (см. Ресурсы). Кроме того, ауксины регулируют многие другие аспекты развития растений, включая деление и дифференцировку клеток. Другие растительные гормоны также оказывают множественное воздействие на ткани-мишени, включая удлинение стебля (гиббереллины), созревание плодов (этилен), деление клеток (цитокинины) и начало покоя (абсцизовая кислота).
Рисунок 13.9
Структура фитогормонов.
Наше понимание молекулярных механизмов действия растительных гормонов менее продвинуто, чем сопоставимые исследования клеток животных, а рецепторы для растительных гормонов только начинают выявляться и охарактеризовываться. Одной из областей, заслуживающих внимания, стало понимание механизма, с помощью которого растительные клетки реагируют на этилен. Используя небольшой сорняк Arabidopsis в качестве модели, было идентифицировано несколько генов, необходимых для чувствительности к этилену. К ним относятся гены, кодирующие рецептор этилена, который похож на семейство рецепторов, обычно встречающихся у бактерий и дрожжей. Дополнительные гены, идентифицированные в сигнальном пути этилена, включают белок, родственный протеинкиназе Raf, который играет ключевую роль в сигнальных путях клеток животных (обсуждается далее в этой главе), и факторы транскрипции, которые регулируют экспрессию этилен- отзывчивые гены.
Используя небольшой сорняк Arabidopsis в качестве модели, было идентифицировано несколько генов, необходимых для чувствительности к этилену. К ним относятся гены, кодирующие рецептор этилена, который похож на семейство рецепторов, обычно встречающихся у бактерий и дрожжей. Дополнительные гены, идентифицированные в сигнальном пути этилена, включают белок, родственный протеинкиназе Raf, который играет ключевую роль в сигнальных путях клеток животных (обсуждается далее в этой главе), и факторы транскрипции, которые регулируют экспрессию этилен- отзывчивые гены.
Сотовая связь и сигнализация | Домашняя страница
Цели и область применения
Cell Communication and Signaling (CCS) — это журнал с открытым доступом, который охватывает все основные и трансляционные аспекты клеточных коммуникаций и сигнальных путей в норме и при патологии. Журнал приветствует представление оригинальных статей, обзорных статей, кратких отчетов и комментариев, относящихся как к эукариотической, так и к прокариотической внутриклеточной и межклеточной передаче сигналов, включая, помимо прочего:
- Передача сигналов белок-белковыми взаимодействиями и модульными доменами
- Посттрансляционная модификация белков и эпигенетическая модификация нуклеиновых кислот
- Механизмы клеточной трансформации, метастазирования, старения и старения
- Гомеостатические и патофизиологические механизмы прогрессирования заболевания
- Механизмы вирусного и бактериального патогенеза и факторы вирулентности
Кроме того, учитывая принадлежность журнала к Международному обществу клеточной смерти, CCS приветствует материалы по всем аспектам клетки
смерть, включающая:
- Апоптотические и неапоптотические механизмы гибели клеток
- Гибель клеток в модельных системах
- Аутофагия
- Клиренс умирающих клеток
- Иммунологические и патологические последствия гибели клеток в тканевом микроокружении Подробнее
- Недавний
- Самые популярные
- Коллекции
-
RBCK1 является эндогенным ингибитором тройного негативного рака молочной железы через ось гиппо/YAP.

Авторы: Чжунбо Ли, Пэн Су, Иньлу Дин, Хунлэй Гао, Хуйцзе Ян, Синь Ли, Сяо Ян, Янь Ся, Ченмяо Чжан, Минси Фу, Дэхай Ван, Е Чжан, Шу Чжо, Цзянь Чжу и Тин Чжуан
Тип контента: Исследования 24 октября 2022 г.
-
Всегда ли один плюс один равняется двум? Структурные различия между несфатином-1, -2 и несфатином-1/2
Авторы: Рафал Ленда, Михал Паджасек, Артур Кренжель, Анджей Ожигар и Доминика Быстроновска
Тип контента: Исследование 24 октября 2022 г.
-
Дефицит
Lrpap1 приводит к миопии из-за TGF-β-индуцированного апоптоза у рыбок даниоАвторы: Шаньшань Лю, Тинг Чен, Бинхао Чен, Ицзюнь Лю, Сяохэ Лу и Цзяли Ли
Тип контента: Исследования 19Октябрь 2022
-
Ориентация на эндоцитоз и клеточные коммуникации в опухолевой иммунной микросреде
Авторы: Бо Ву, Цянь Ван, Сян Ши и Мэйси Цзян
Тип контента: Обзор 18 октября 2022 г.
-
Эндрю Х. Уилли, пионер в области апоптоза.
Авторы: Захра Закери и Ричард А. Локшин
Тип контента: Некролог 17 октября 2022 г.
-
МикроРНК-188 подавляет переход G
1 /S путем нацеливания на множественные комплексы циклин/CDKАвторы: Jiangbin Wu, Qing Lv, Jie He, Haoxiang Zhang, Xueshuang Mei, Kai Cui, Nunu Huang, Weidong Xie, Naihan Xu и Yaou Zhang
Тип контента: Исследования 11 октября 2014 г.
-
Роль сигнального пути JAK-STAT и его регуляторов в судьбе Т-хелперных клеток
Авторы: Фархад Сейф, Маджид Хошмирсафа, Хоссейн Аазами, Монире Мохсензадеган, Голамреза Седиги и Мохаммадали Бахар
Тип контента: Обзор 21 июня 2017 г.
-
Различные популяции и источники мезенхимальных стволовых клеток человека (МСК): сравнение МСК, полученных из тканей взрослых и новорожденных
Авторы: Ральф Хасс, Корнелия Каспер, Стефани Бём и Роланд Джейкобс
Тип контента: Обзор 14 мая 2011 г.

-
Rac и Rho GTPases в контроле подвижности раковых клеток
Авторы: Маттео Парри и Паола Кьяруги
Тип контента: Обзор 7 сентября 2010 г.
-
Домены PDZ и их партнеры по связыванию: структура, специфичность и модификация
Авторы: Хо-Джин Ли и Цзе Дж. Чжэн
Тип контента: Обзор 28 мая 2010 г.
Артикул
Последние статьи RSS
Посмотреть все статьи
Самые популярные статьи RSS
Посмотреть все статьи
2021
Киназы, адапторные белки и перестройки актинового цитоскелета при ЭМП и раке
Под редакцией Лешека Котулы и Патриции Дубельецкой-Щербы
Под редакцией Сары Бондос, А.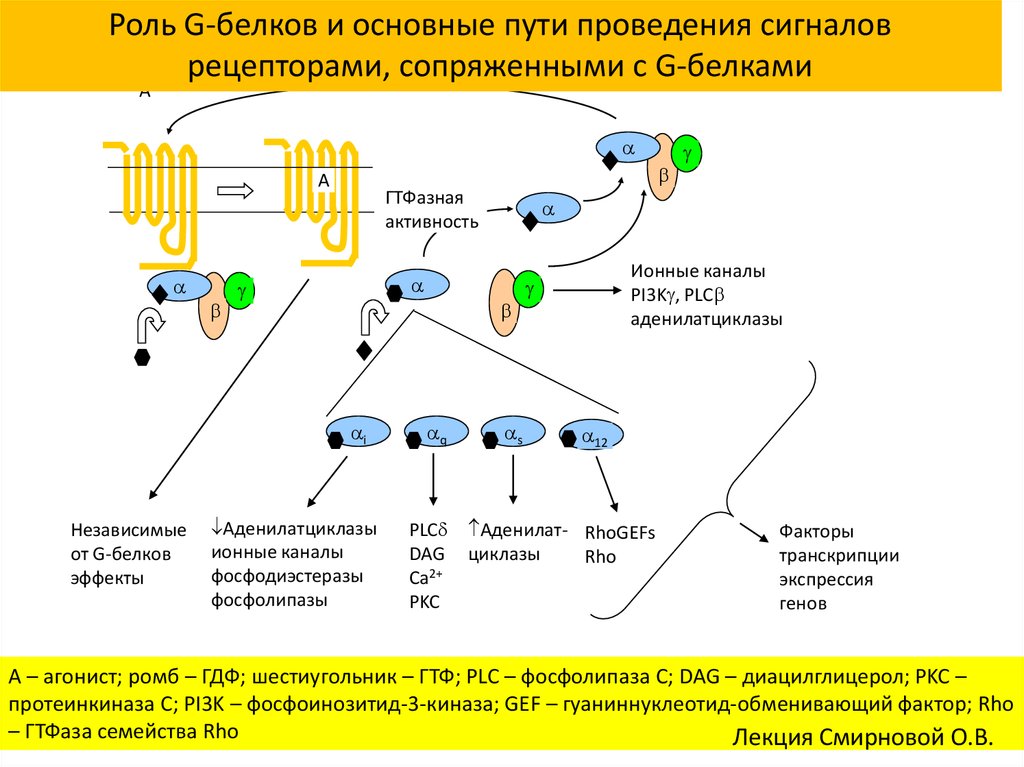 К. Дункера и Владимира Уверского
К. Дункера и Владимира Уверского
2020
Биология фосфатидилсерина: базовая физиология и последствия для иммунологии, инфекционных заболеваний и рака
, под редакцией Raymond Birge и David Calianese
2019
25th Anniversary of The Icds Dithed Ditedredge 9019 9019 3th Anniversary of The Icds Dithedge Diteledge 9013 9019 3 -й Annibrary of The Icds Dithed Diech 9013
.
2018
Плюрипотентные стволовые клетки: обзор исследований
2017
Связь опухолевых клеток с микроокружением
Под редакцией Silja Wessler
2016
Short linear motifs — the unexplored frontier of the eukaryotic proteome
Edited by Norman Davey
2014
Bacterial pathogen — host cell interactions
Edited by Silja Wessler
2013
Systems Biology and Medicine
Под редакцией Stephan Feller и Fred Schaper
2011
Взаимодействие Helicobacter pylori с клеткой-хозяином
Под редакцией Silja Wessler
2010
Cancer cell motility and microenvironment
Call for papers
Extracellular vesicles in infectious disease
Edited by Dr. Nicole-Meisner Kober and Dr. Silja Wessler
Nicole-Meisner Kober and Dr. Silja Wessler
Submission Deadline: January 31, 2023
Внутренние клеточные механизмы иммунного побега
Под редакцией доктора Ганапати Шрирама и доктора Раймонда Бирджа
Отправьте свою рукопись здесь.
Новости и обновления
Ежегодное собрание ICDS
Ежегодное собрание ICDS пройдёт 2–5 июля 2022 года в Стамбуле. Это будет личная встреча (требуется подтверждение вакцинации). Чтобы узнать больше, посетите веб-сайт конференции.
14 th Ежегодное ежегодное собрание ÖGMBT
Вскоре будет объявлено о собрании Австрийской ассоциации молекулярных наук о жизни и биотехнологии в 2022 году. Вернитесь для получения дополнительной информации или посетите веб-сайт OGMBT.
Новая тематическая серия открыта для заявок
Приглашенные редакторы Николь Мейснер-Кобер и Силья Весслер приветствуют материалы для своей новой серии статей о внеклеточных везикулах при инфекционных заболеваниях. Подробнее о сериале здесь.
Подробнее о сериале здесь.
Видеобайты здесь
В Cell Communication and Signaling, мы увлечены распространением исследований.
Мы рады сообщить, что с 12 августа 2019 года все авторы, чьи статьи будут приняты, получат видеобайт.
Созданный нашими партнерами Research Square, Video Byte представляет собой 1-2-минутное видеообзор статьи, которое будет включено в реферат статьи и опубликовано на веб-сайте журнала. Авторы также получат общедоступную версию своего видеобайта.
Профиль редактора
Раймонд Бирдж, главный редактор
Раймонд Бирдж, доктор философии, профессор Университета штата Нью-Джерси в Рутгерсе и член онкологического центра Медицинской школы Нью-Джерси. Доктор Бирдж поступил в Медицинскую школу UMDNJ-Нью-Джерси в 2000 году и в настоящее время занимает должность профессора и заместителя председателя по исследованиям на кафедре микробиологии, биохимии и молекулярной биологии. Доктор Бирдж также имеет совместные назначения в Онкологическом центре Медицинской школы Нью-Джерси (Ньюарк) и Институте рака Нью-Джерси (Нью-Брансуик).