Что представляет собой белок PLK. Какие функции выполняет PLK в клетке. На какие процессы влияет сигнальный путь PLK. Как регулируется активность PLK. Каково значение PLK для нормального деления клеток.
Что такое PLK и его роль в регуляции клеточного цикла
PLK (Polo-like kinase) — это семейство серин/треониновых протеинкиназ, играющих ключевую роль в регуляции клеточного цикла и митоза. Основной представитель этого семейства — PLK1 — активируется в начале M-фазы клеточного цикла и участвует в координации многих важных процессов деления клетки.
Основные функции PLK1 включают:
- Активацию циклин-зависимых киназ и продвижение клетки в митоз
- Регуляцию центросом и формирования митотического веретена
- Контроль выхода клетки из митоза и цитокинеза
- Участие в репарации повреждений ДНК
Активация и регуляция PLK в клетке
Активность PLK1 строго регулируется на протяжении клеточного цикла. Основные механизмы активации PLK1 включают:
- Фосфорилирование активационной петли киназы Aurora A
- Связывание с белками-партнерами через PBD-домен
- Убиквитин-зависимая деградация ингибиторов PLK1
Активированная PLK1 фосфорилирует множество белков-мишеней, запуская каскад сигнальных путей, контролирующих вход и прохождение клетки через митоз.
Роль PLK в формировании и функционировании митотического веретена
Одна из ключевых функций PLK1 — регуляция динамики центросом и митотического веретена деления. PLK1 участвует в следующих процессах:
- Созревание и разделение центросом в начале митоза
- Нуклеация и стабилизация микротрубочек веретена
- Ориентация и прикрепление хромосом к нитям веретена
- Контроль сборки кинетохоров
Нарушение этих функций PLK1 приводит к дефектам формирования веретена и неправильному расхождению хромосом при делении клетки.
Участие PLK в контроле выхода из митоза
PLK1 играет важную роль в регуляции выхода клетки из митоза и инициации цитокинеза. Ключевые функции PLK1 на этом этапе включают:
- Активацию комплекса APC/C, запускающего деградацию циклина B
- Фосфорилирование белков центральной части веретена
- Регуляцию сборки сократительного кольца при цитокинезе
- Контроль абсцизии — финального этапа разделения дочерних клеток
Таким образом, PLK1 координирует завершение митоза и цитокинез, обеспечивая корректное разделение генетического материала и органелл между дочерними клетками.

PLK и репарация повреждений ДНК
Помимо регуляции митоза, PLK1 участвует в клеточном ответе на повреждения ДНК:
- Активирует контрольные точки клеточного цикла при повреждениях ДНК
- Фосфорилирует белки, участвующие в репарации двунитевых разрывов ДНК
- Регулирует процесс возобновления клеточного цикла после устранения повреждений
Эти функции PLK1 важны для поддержания стабильности генома и предотвращения накопления мутаций при делении клеток.
Значение PLK для нормального деления клеток
PLK1 является ключевым регулятором множества процессов, обеспечивающих правильное прохождение митоза и цитокинеза. Нарушение функций PLK1 может приводить к серьезным последствиям:
- Остановке клеточного цикла и блоку деления клеток
- Нарушениям сегрегации хромосом и анеуплоидии
- Дефектам цитокинеза и образованию многоядерных клеток
- Накоплению повреждений ДНК и геномной нестабильности
Поэтому строгая регуляция активности PLK1 критически важна для нормального деления клеток и поддержания стабильности генома.
Роль PLK в канцерогенезе и перспективы терапии
Нарушение регуляции PLK1 часто наблюдается в раковых клетках и ассоциировано с плохим прогнозом многих видов рака. Повышенная экспрессия PLK1 способствует:
- Усиленной пролиферации опухолевых клеток
- Резистентности к апоптозу
- Геномной нестабильности и прогрессии опухоли
- Метастазированию
Это делает PLK1 перспективной мишенью для противоопухолевой терапии. Ряд ингибиторов PLK1 в настоящее время проходят клинические испытания как потенциальные противораковые препараты.
Заключение
PLK1 является ключевым регулятором клеточного цикла, митоза и цитокинеза. Сигнальный путь PLK координирует множество важнейших процессов деления клетки — от созревания центросом до разделения дочерних клеток. Строгий контроль активности PLK1 необходим для нормального деления клеток и поддержания стабильности генома. Нарушение регуляции PLK1 ассоциировано с развитием рака, что делает этот белок перспективной мишенью для разработки новых противоопухолевых препаратов.

Документы, инструкции и сертификаты на камнерезный станок DIAM PLK-250/1.8 PROLINE 600101
1800
Тип двигателя
электрический
Длина реза, мм
610
Max глубина пропила, мм
38/60/90
Число оборотов, об/мин
4000
25.4
Диаметр диска, мм
250
Беговые лыжи SALOMON Aero 7 Skin X-Stiff + PLK Access (19/20)
Беговые лыжи SALOMON Aero 7 Skin X-Stiff + PLK Access (19/20) классическая прогулочная лыжа от компании SALOMON. Беговые лыжи SALOMON Aero 7 Skin X-Stiff + PLK Access (19/20) модель обладает повышенной жесткостью в колодке для возможности эксплуатации лыж тяжелыми лыжниками. Благодаря использованию под колодкой приклеенного синтетического камуса SALOMON Aero 7 Skin (19/20) не требуют нанесения мазей и обладают эффективным держанием лыжи при отталкивании. Особенностью конструкции Skin является наличие синтетического камуса под колодкой лыжи, камус обеспечивает отличное удержание лыжи при отталкивании и не мешает паре скользить вперед. Данная особенность позволяет использовать лижу в широчайшем диапазоне температур и снега и освобождает лыжника от использования температурных мазей держания. 100 % мохер для камуса это идеальный баланс между сцеплением и скольжением. Камус приклеен к лыже.Эти любительские лыжи для классического стиля, обладают высокими характеристиками скольжения и держания. Благодаря сердечнику Densolite 2000, новая модель Aero 7 Skin является крайне легкой. Эти лыжи обеспечивают высокую передачу толчка от ноги к лыже и более динамичны в фазе скольжения. Такие качества позволяют начинающим лыжникам довольно быстро освоить технику классических ходов.
В сезоне 19-20 на лыжах предустановлены новые крепления SALOMON Prolink Shift, данные крепления имеют возможность смещения по длине лыжи относительно точки баланса, что позволяет лыжнику более точно настроить лыжи индивидуально для себя смещая баланс между скольжением или держанием.
Вы получите искреннее удовольствие от катания на этих лыжах.
SALOMON Aero 7 Skin X-Stiff модель рассчитанная на тяжелых лыжников.
ТЕХНОЛОГИИ:
Геометрия бокового выреза: Parallel Classic Race
Конструкция лыж: Propulsion, Digital Dynamic Flex Control
Сердечник: Densolite 2000
Скользящая поверхность: G3
Шлифовка скользящей поверхности: Race base, DGM — Digital Grinding Measurement
Защита задней кромки: Rental specific
Вес 1760 гр.
Kermi Line PLV-PLK линейный стальной радиатор
Линейные радиаторы Kermi Line PLV-PLK производятся с боковым или нижним подключением. Устанавливаются на пол или стену. Характерной особенностью германских моделей является эстетичность и экономичность. Рассчитаны на эксплуатацию в отопительных системах с давлением до 10 атмосфер. Максимально допустимое значение (испытательное) составляет 16 атмосфер.
Предлагаем ознакомиться с другими вариантами радиаторов Керми:
Радиаторы Kermi Line PLV-PLK с пролинееной декоративной накладкой поддерживаются в складской программе в высоте 200 мм и 500 мм с нижним подключением.
Особенности:
- строгий дизайн. Эстетический минимализм относится к числу достоинств: модель органично вписывается в интерьер. Она дополняет обстановку, не привлекая к себе избыточного внимания;
- быстрый обогрев помещения. Благодаря продуманному инженерному решению радиаторы обогревают помещение за короткий промежуток времени;
- динамичность температурного режима. Для нагрева приборов отопления требуется мало воды. Небольшая инертность позволяет быстро изменить температурный режим;
- простота монтажа. Для установки требуется минимум времени, поскольку радиаторы поставляются в собранном виде и укомплектованы необходимыми крепежами.
Производитель поставляет на рынок модели с разными размерами, что дает возможность выбрать наиболее подходящий вариант под конкретные требования. Высота радиатора варьируется от 200 до 900 мм, длина от 400 до 3000 мм. По глубине они деляться на типы с помощью которых можно определить не только глубину, но и конструкцию прибора — первое число отвечают за количество панелей по каналам, которых протекает теплоноситель, второе число говорит о количестве конвекционных решеток внутри батареи.
Типы стального радиатора Керми Лайн:
- 10 тип (63 мм) — 1 передняя панель с пролинееной декоративной накладкой, 0 пластин с конвекционным оребрением
- 11 тип (63 мм) — 1 передняя панель с пролинееной декоративной накладкой, 1 конвекционная пластина (без задней панели)
- 12/21 тип (66 мм) — 2 панели (спереди с пролинееной декоративной накладкой и сзади), 1 конвекционная пластина
- 22 тип (102 мм) — 2 панели (спереди с пролинееной декоративной накладкой и сзади), 2 конвекционные пластины
- 33 тип (157 мм) — 3 панели (спереди с пролинееной декоративной накладкой, посредине и сзади), 3 конвекционные пластины
Монтаж стального радиатора очень прост и легок в понимание, по умолчанию предусматривает настенное крепление. У моделей высотой от 300 мм в комплекте идет двухсторонний кронштейн (одна сторона — 30, вторая — 50 мм), это означает, что вы сами можете выбрать какой стороной крепить держатель к стене — на расстояние 30 или 50 мм. На задней панели приварены ушки, на которые и будут цепляться кронштейны.
Для радиатора длиной до 1600 мм (включительно) предусмотрено 2 крепления, от 1800 мм (включительно) — 3 крепления.
Подключение у представленной модели может быть друх типов: боковое (PLK) и нижнее (PLV). При боковом подключение допускается несколько вариантов подвода труб: левое, правое, диагональное или седловидное (снизу по бокам). Нижнее подключение по умолчанию сделано с правой стороны батареи с межосевым растоянием 50 мм, по желанию клиенту может быть прибор может быть с левым подключением. Радиаторы Kermi с нижним подключением имеют уже встроенную вентильную вставку с резьбовым соединением М30х1,5, на которую можно монтировать термостатическую головку.
РАЗМЕРЫ ПОДКЛЮЧЕНИЯ ДЛЯ РАДИАТОРОВ КЕРМИ ДЛЯ БОКОВОГО ПОДКЛЮЧЕНИЯ
РАЗМЕРЫ ПОДКЛЮЧЕНИЯ ДЛЯ РАДИАТОРОВ КЕРМИ ДЛЯ НИЖНЕГО ПОДКЛЮЧЕНИЯ
|
[PLK] J’pilote la caisse, ma chérie, t’en fais pas J’me demandais : comment on s’connait ? Elle veut qu’on quitte le pays, qu’on aille se balader J’ai mis la dernière re-pai, frais, toujours opé’ Elle veut qu’on quitte le pays, qu’on aille se balader J’pilote la caisse, ma chérie, t’en fais pas J’me demandais : comment on s’connait ? [Hamza] Elle veut qu’on quitte le pays, qu’on aille se balader [PLK] J’me demandais : comment on s’connait ? |
[PLK] Я за рулем, дорогая, ни о чем не беспокойся; Я спрашиваю себя: как мы сошлись? Она хочет, чтобы мы поехали за границу, отправились бродить. Я надеваю последнюю пару обуви, свежий, всегда в форме, Она хочет, чтобы мы уехали за границу, отправились бродить. Я за рулем, дорогая, ни о чем не беспокойся; Я спрашиваю себя: как мы сошлись? [Hamza] Она хочет, чтобы мы уехали за границу, отправились бродить. [PLK] Я спрашиваю себя: как мы сошлись? |
Как открыть файл PLK? Расширение файла .PLK
Что такое файл PLK?
PLK суффикс имени файла в основном используется для COREX Part-of-speech Tag файлов. Файлы с расширением PLK могут использоваться программами, распространяемыми для платформы Windows. PLK файл относится к категории Document Files так же, как #NUMEXTENSIONS # других расширений файлов, перечисленных в нашей базе данных. Для управления файлами PLK рекомендуется COREX.
Программы, которые поддерживают PLK расширение файла
Следующий список функций PLK -совместимых программ. Файлы с суффиксом PLK могут быть скопированы на любое мобильное устройство или системную платформу, но может быть невозможно открыть их должным образом в целевой системе.
Как открыть файл PLK?
Причин, по которым у вас возникают проблемы с открытием файлов PLK в данной системе, может быть несколько. К счастью, наиболее распространенные проблемы с файлами PLK могут быть решены без глубоких знаний в области ИТ, а главное, за считанные минуты. Приведенный ниже список проведет вас через процесс решения возникшей проблемы.
Шаг 1. Скачайте и установите COREXОсновная и наиболее частая причина, препятствующая открытию пользователями файлов PLK, заключается в том, что в системе пользователя не установлена программа, которая может обрабатывать файлы PLK. Решение этой проблемы очень простое. Загрузите COREX и установите его на свое устройство. В верхней части страницы находится список всех программ, сгруппированных по поддерживаемым операционным системам. Если вы хотите загрузить установщик COREX наиболее безопасным способом, мы рекомендуем вам посетить сайт и загрузить его из официальных репозиториев.
Шаг 2. Обновите COREX до последней версииЕсли у вас уже установлен COREX в ваших системах и файлы PLK по-прежнему не открываются должным образом, проверьте, установлена ли у вас последняя версия программного обеспечения. Разработчики программного обеспечения могут реализовать поддержку более современных форматов файлов в обновленных версиях своих продуктов. Если у вас установлена более старая версия COREX, она может не поддерживать формат PLK. Последняя версия COREX должна поддерживать все форматы файлов, которые совместимы со старыми версиями программного обеспечения.
Шаг 3. Свяжите файлы COREX Part-of-speech Tag с COREXЕсли проблема не была решена на предыдущем шаге, вам следует связать PLK файлы с последней версией COREX, установленной на вашем устройстве. Процесс связывания форматов файлов с приложением по умолчанию может отличаться в деталях в зависимости от платформы, но основная процедура очень похожа.
Процедура изменения программы по умолчанию в Windows
- Выберите пункт Открыть с помощью в меню «Файл», к которому можно щелкнуть правой кнопкой мыши файл PLK.
- Нажмите Выбрать другое приложение и затем выберите опцию Еще приложения
- Наконец, выберите Найти другое приложение на этом… , укажите папку, в которой установлен COREX, установите флажок Всегда использовать это приложение для открытия PLK файлы свой выбор, нажав кнопку ОК
Процедура изменения программы по умолчанию в Mac OS
- Щелкните правой кнопкой мыши на файле PLK и выберите Информация.
- Найдите опцию Открыть с помощью — щелкните заголовок, если он скрыт
- Выберите COREX и нажмите Изменить для всех …
- Если вы выполнили предыдущие шаги, должно появиться сообщение: Это изменение будет применено ко всем файлам с расширением PLK. Затем нажмите кнопку Вперед», чтобы завершить процесс.
Если вы выполнили инструкции из предыдущих шагов, но проблема все еще не решена, вам следует проверить файл PLK, о котором идет речь. Вероятно, файл поврежден и, следовательно, недоступен.
1. Убедитесь, что PLK не заражен компьютерным вирусомЕсли случится так, что PLK инфицирован вирусом, это может быть причиной, которая мешает вам получить к нему доступ. Рекомендуется как можно скорее сканировать систему на наличие вирусов и вредоносных программ или использовать онлайн-антивирусный сканер. Если файл PLK действительно заражен, следуйте инструкциям ниже.
2. Проверьте, не поврежден ли файлЕсли файл PLK был отправлен вам кем-то другим, попросите этого человека отправить вам файл. Возможно, что файл не был должным образом скопирован в хранилище данных и является неполным и поэтому не может быть открыт. Это может произойти, если процесс загрузки файла с расширением PLK был прерван и данные файла повреждены. Загрузите файл снова из того же источника.
3. Проверьте, есть ли у вашей учетной записи административные праваИногда для доступа к файлам пользователю необходимы права администратора. Выйдите из своей текущей учетной записи и войдите в учетную запись с достаточными правами доступа. Затем откройте файл COREX Part-of-speech Tag.
4. Убедитесь, что ваше устройство соответствует требованиям для возможности открытия COREXОперационные системы могут иметь достаточно свободных ресурсов для запуска приложения, поддерживающего файлы PLK. Закройте все работающие программы и попробуйте открыть файл PLK.
5. Проверьте, есть ли у вас последние обновления операционной системы и драйверовСовременная система и драйверы не только делают ваш компьютер более безопасным, но также могут решить проблемы с файлом COREX Part-of-speech Tag. Возможно, что одно из доступных обновлений системы или драйверов может решить проблемы с файлами PLK, влияющими на более старые версии данного программного обеспечения.
PLK Сообщество Отзыв
При застройке роскошных квартир и кондоминиумов планы могут меняться вплоть до начала строительства проекта. Сообщества PLK нуждались в банке, который мог бы не отставать.
«LCNB очень оперативно реагирует на запросы», — сказал финансовый директор Джон Блэкхэм. «Мы можем изменить внешний вид здания, или мы можем решить построить 130 единиц вместо 120. Они смогли двигаться так же быстро, как и мы».
Компания PLK, базирующаяся в Цинциннати, может похвастаться 25 офисами в Огайо и Кентукки, и многие другие находятся в стадии разработки.Проекты такого масштаба зависят от множества факторов, и их реализация требует открытого общения и надежности. LCNB сначала выделился для Блэкхэма как потенциальный партнер своей готовностью задавать вопросы и понимать проект.
«Когда вы получаете обязательства от банкиров, а они не задают никаких вопросов, это красный флаг», — говорит Блэкхэм. «Вы не хотите сотрудничать с банком, который не понимает риска. LCNB делает все возможное, чтобы проявить должную осмотрительность.”
То, как LCNB выполняет свои условия, позволяет PLK уверенно продвигать свои проекты. Банкиры LCNB не предлагают условия без одобрения всей команды. Таким образом, PLK знает, что LCNB будет соблюдать свои условия на протяжении всего закрытия.
«В случае с другими банками банкир дает нам перечень условий, но у них нет возможности сотрудничать с руководителями банков», — сказал Блэкхэм. «Затем мы приближаемся к завершению, и кто-то, исключенный из первоначального процесса, в последнюю минуту испытывает затруднения.С LCNB этого не происходит ».
Широкий спектр знанийLCNB помогает PLK достигать множества целей. LCNB помогает во всех аспектах бизнеса — от квартир и кондоминиумов до зданий розничной торговли и складских помещений.
«У меня прекрасные отношения с моим банкиром, и я уверен, что в его распоряжении вся команда LCNB». — говорит Блэкхэм. «Они быстро разбирают сделку, оценивают риски и принимают решение. Их своевременность помогает мне уменьшить стресс и осложнения.”
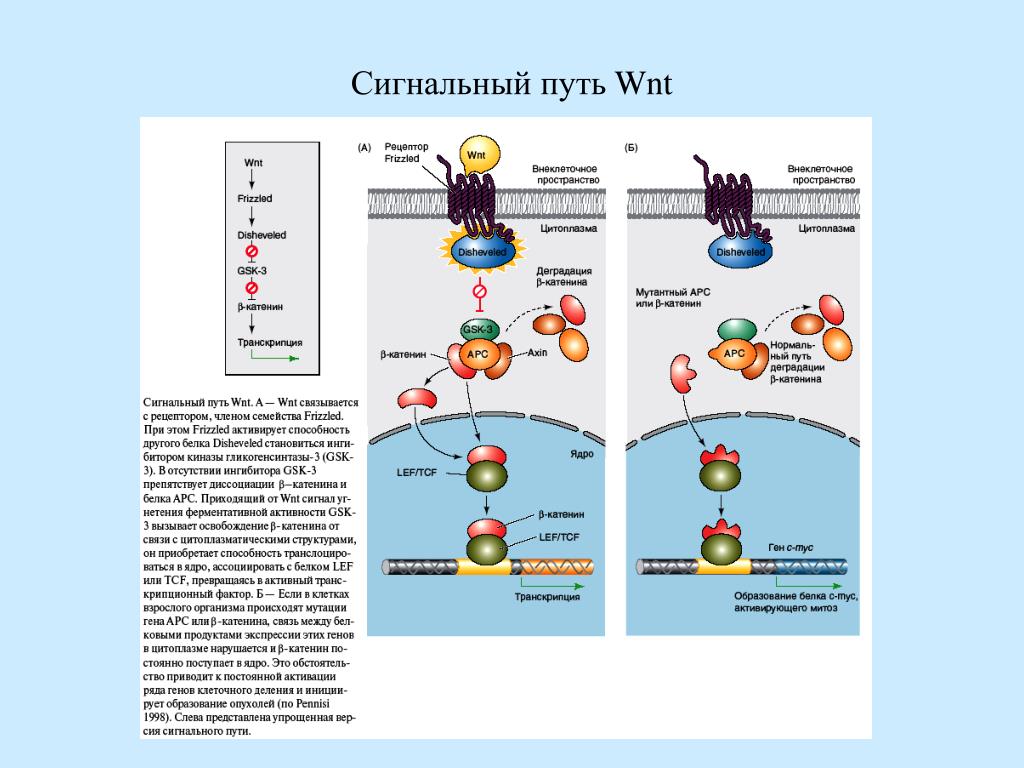
Plk является функциональным гомологом Saccharomyces cerevisiae Cdc5, и повышенная активность Plk вызывает множественные перегородочные структуры
Plk представляет собой серин / треониновую протеинкиназу млекопитающих, пик активности которой приходится на начало M-фазы. Он тесно связан с другими киназами млекопитающих, Snk, Fnk и Prk, а также с Xenopus laevis Plx1, Drosophila melanogaster polo, Schizosaccharomyces pombe Plo1 и Saccharomyces cerevisiae Cdc5.М-фаза клеточного цикла — это хорошо скоординированный процесс, который обеспечивает равнораспределение генетического и клеточного материалов во время деления клеток. Чтобы сделать возможным понимание функции Plk во время прогрессирования M фазы, различные мутанты Plk были созданы и экспрессированы в клетках Sf9 и почкующихся дрожжах. Анализы киназы in vitro с иммунопреципитатами Plk, полученными из клеток Sf9, показывают, что Glu206 и Thr210 играют одинаково важные роли для активности Plk и что замена Thr210 отрицательно заряженным остатком повышает специфическую активность Plk.Эктопическая экспрессия Plk дикого типа (Plk WT) дополняет дефект клеточного деления, связанный с мутацией cdc5-1 в S. cerevisiae. Степень комплементации тесно коррелирует с активностью Plk, измеренной in vitro, поскольку она усиливается мутационно активированным Plk, T210D, но не наблюдается с неактивными формами K82M, D194N и D194R. На фоне CDC5 дикого типа экспрессия Plk WT или T210D, но не неактивных форм, вызвала резкое накопление клеток в G1. В соответствии с повышенной активностью Plk, этот феномен усиливается за счет удаленных на С-конце форм WT deltaC и T210D deltaC.Экспрессия T210D также индуцировала класс клеток с необычно удлиненными зачатками, в которых развились множественные перегородочные структуры. Однако этого не наблюдалось с удаленной на С-конце формой T210D deltaC. Оказывается, что С-конец Plk не требуется для наблюдаемого влияния клеточного цикла, но может быть важным для роста поляризованных клеток и формирования перегородочной структуры.
Сигнальный трактPLK — Creative Diagnostics
Рисунок 1.Сигнальный путь PLK.
Обзор ПЛК
Поло-подобная киназа (PLK) — это высококонсервативная серин / треониновая протеинкиназа с высокогомологичным серин / треонинкиназным доменом на N-конце, который регулирует активность PLK и субклеточную динамику на C-конце и целевом полобоксовом домене ( PBD). Существует много членов семейства PLK, и в организме человека существует четыре подтипа, а именно PLK1, PLK2, PLK3 и PLK4, которые играют важную роль в регуляции различных фаз клеточного цикла.Среди четырех членов семьи текущее исследование PLK1 является наиболее тщательным. Белки PLK регулируют многие ключевые этапы клеточного цикла, включая образование биполярных веретен, сегрегацию хромосом, позднюю регуляцию комплексов и цитокинез. PLK представляет собой очень эффективную противораковую стратегию в качестве мишени для ингибирования образования веретена во время митоза, тем самым предотвращая деление раковых клеток.
Семейство ПЛК
Самая тщательная проработка в семействе PLK — это PLK1.В 1994 г. было сообщено о первом гене PLK, который расположен в 16p12, с длиной мРНК около 2,3 т.п.н., кодирующей белок с молекулярной массой около 66000. Его аминокислотная последовательность, а также CDC5 дрозофилы и дрожжей CDC5 и другие гены имеют большая гомология, и есть высококонсервативная каталитическая область на N-конце. С-концевой PBD состоит из двух консервативных областей, называемых поло-боксами. PLK1 играет важную роль в нормальном функционировании клеточного цикла, созревании центросом и цитоплазматическом разделении, но он сверхэкспрессируется в большинстве опухолевых клеток человека, и эта сверхэкспрессия часто сопровождается плохим прогнозом.Следовательно, изучение уровня экспрессии PLK1 в клетках имеет большое значение для ранней диагностики опухолей и прогнозирования прогноза пациентов с опухолями. Помимо ПЛК1, наиболее изученным является ПЛК4. В 1994 году в результате скрининга библиотеки кДНК учеными был обнаружен ген, кодируемый продукт которого очень похож на белки семейства POLO (Snk и PLK1) и содержит высокогомологичную N-концевую киназу. Поэтому snk / Plk, которая является протеинкиназой того же типа, позже была названа PLK4.PLK4 широко встречается у эукариот, таких как нематоды, дрозофилы и приматы, но не у прокариот. Ген мыши PLK4 расположен на хромосоме 13, а ген PLK4 человека расположен на хромосоме 4q28, которая часто подвергается хромосомным перестройкам или делециям в опухолевых клетках человека. Ген PLK 4 у мышей кодирует два подтипа белка, PLK-a и PLK-b. Длина PLK-b составляет 464 аминокислоты, а N-сегменты идентичны по 416 аминоконцам, а C-область отличается. Белок PLK4 имеет общую длину 970 аминокислот с высокой гомологией с мышиным PLK4-a.В отличие от других белков семейства PLK, PLK4 имеет только один поло бокс, который может образовывать межмолекулярные двойные димеры. Эти димеры подобны двум полобоксам PLK1, образующим внутримолекулярную гетеродимерную структуру.
Сигнальный путь PLK
- Каскад сигнального пути PLK
- Регуляция пути
- Связь с болезнью
PLK1 в основном способствует функциональному созреванию центросомы и формированию веретена на поздних стадиях G2 до ранней стадии M, тем самым способствуя вступлению клеток в фазу M; и он одновременно участвует в активации регулятора клеточного цикла CDC25, который, в свою очередь, способствует образованию цилиндрической дентинкиназы 2 (активация CDK2).Исследования соматических клеток показали, что центросома регулирует сборку веретена в соответствующее время и в нужном месте. При трансформации G2-M центросома постепенно созревает, проявляясь в виде кольцевого комплекса γ-tubulin (γ-TuRCs) и др. Регуляции микротрубочек. Факторы накапливаются, и две центросомы отделяются друг от друга, образуя полюса веретена. Этот ряд процессов регулируется тремя белками: TPX2, белком ткани микротрубочек, PLK1 и AuroraA. Эксперименты по интерференции антисмысловой РНК показывают, что эти три ключевых фермента могут регулировать созревание центросом по следующему механизму: PLK1 находится на самом верхнем конце этого каскада, который, с одной стороны, прикрепляет Aurora A к центросоме и способствует созреванию центросомы и правильной поляризации; Другая сторона регулирует TPX2, который накапливается на полюсах веретена и привлекает тубулин для формирования веретена.В то же время TPX2 также заставляет Aurora A локализоваться на шпинделе. В клетках Hela, если Aurora A полностью блокируется, образуется мультиполяризованная центросома, но тубулин все еще может накапливаться вокруг центросомы, и клетки могут вступать в фазу митоза; однако, если экспрессия PLK1 подавлена, клетки будут застаиваться в периоде G2. Это указывает на то, что помимо своей роли в созревании центросом, PLK1 может регулировать прогрессию клеточного цикла с помощью других механизмов. В настоящее время известно, что конкурирующее взаимодействие между CDK1 и его антагонистическим ферментом WEE1 является еще одним ключевым фактором, определяющим трансформацию клеток из фазы G2 в M-фазу.CDK1 инициирует конденсацию хромосом и процесс ядерной мембраны посредством фосфорилирования, в то время как CDC25 активирует CDK1, ключевой фермент, который вызывает его миграцию в ядро; в то время как WEE1 ингибирует его активность за счет фосфорилирования аминокислоты в сайте CDK1, он задерживает или даже задерживает митоз клеток. Напротив, PLK1 катализирует локализацию CDC25 в ядре, активирует CDK1, а активированный CDK1 фосфорилирует больше CDC25 за счет положительной обратной связи, ускоряя процесс петли митотического расширения; в то же время PLK1 может также определять ячейку, управляя относительным содержанием WEE1 и CDC25.Исследование также показало, что когда аномальная хромосомная репликация происходит в клетках, таких как опухолевые клетки, CDC25 и PLK1 сверхэкспрессируются, а активность WEE1 подавляется. Следовательно, раковые клетки могут размножаться за счет регуляции контрольной точки G2-M и злокачественной пролиферации. На средней и поздней стадиях митоза, с деградацией связывающего белка-адгезина между центромерами, делением центромеры и разделением сестринских хроматид, на этот процесс влияет раннее митотическое ингибирование 1 (Emi1) и фактор позднего промотирования (APC).На ранней стадии митоза APC существует как неактивный предшественник, а зрелый промотор-промоторный фактор (MPF), остающийся во время трансформации G2-M, зависит от cyclinB, способствующего активации фосфорилирования путем переноса фосфатной группы АТФ на APC. Таким образом, он может гидролизовать адгезин. В этом процессе PLK1 фосфорилирует Emi1 и MPF, что приводит к ингибированию активности Emi1, активации APC, активации APC-опосредованного убиквитинирования цитокина linB и деградации соответствующей протеазой; поскольку активность MPF зависит от циклина B, активность MPF исчезает и ядрышко восстанавливается.Таким образом, клетки обычно выходят из митоза, в противном случае митоз будет остановлен на более поздней стадии, в конечном итоге образуя многоядерные клетки. Исследование показало, что PLK1 локализуется в середине веретена на поздних стадиях митоза, что позволяет предположить, что он может играть важную роль в цитокинезе. Один из экспериментов показал, что в доминантном мутанте гомологичного гена polo PLK1 дрожжей S. pombe ген polo не мог нормально экспрессироваться, что не только ингибировало образование веретена, но также приводило к уменьшению усадочного кольца и бороздки деления. быть в М.Позднее формирование предотвращает образование новой мембраны. Другой эксперимент показал, что в клетках животных сверхэкспрессия PLK1 сопровождается образованием многоядерных клеток. Два экспериментальных результата показывают, что точная экспрессия PLK1 может играть важную роль в нормальном цитокинезе. В последние годы гипотеза о том, что PLK1 может косвенно активировать G-белок и опосредовать образование плазматической мембраны, может быть продвинута путем содействия созреванию тела веретена.
Самым последним исследованием регуляции PLK1 является разработка родственных ингибиторов: как и большинство противоопухолевых препаратов, нацеленных на протеинкиназы, текущая разработка ингибиторов PLK1 осуществляется в основном за счет конкурентного связывания сайтов АТФ для прямого ингибирования каталитической активности PLK1.Недавно ученые открыли ряд производных хиназолина, которые могут конкурентно связываться с центральной областью АТФ с помощью высокопроизводительных методов скрининга, тем самым подавляя киназную активность PLK1. Мы хотели бы резюмировать текущий прогресс в исследованиях ингибиторов PLK: a. GSK461364 — это АТФ-конкурентный ингибитор PLK1, разработанный Glaxo Smith Kline. Он может быстро образовывать обратимый комплекс с PLK1. Ингибирующее действие на PLK1 в 400 раз больше, чем у PLK2 и PLK3. Эксперименты in vitro показывают, что GSK461364 подходит для пролиферации 120 линий опухолевых клеток и обладает ингибирующей активностью, а в более чем 83% протестированных клеточных линий IC50 составляет менее 50 нмоль / л.Было проведено клиническое исследование фазы I (солидная опухоль), которое показало, что у 20% субъектов развился ответ, и продолжаются последующие клинические исследования фазы II. б. LFM-A13 был впервые признан специфическим блокатором Btk (тирозинкиназы Bruton S), и недавно было обнаружено, что он также может быть потенциальным низкомолекулярным ингибитором PLK1. c. LFM-A13, низкомолекулярный химический ингибитор PLK1, действует на PTP-область Plx1 (гомологичный ген PLK1) на сайте связывания АТФ шелкового / треонинкиназного домена Plx1 через киназный домен.Связывание сайта АТФ приводит к изменению конформации Plx1, что ингибирует активность киназы PLK1, тем самым вызывая стагнацию митоза опухолевых клеток с образованием монополярного или мультиполярного аномального веретена. И 10 тирозинкиназ, таких как ABL, BRK, BMX, c-KIT, FYN, IGF1R, PDGFR, JAK2, MET и YES, не вызывали ингибирования. d. GW843682X — это новый конкурентный ингибитор PLK с АТФ, который избирательно подавляет активность PLK1 и PLK3 и оказывает широкий ингибирующий эффект на большинство опухолей.В клеточной линии аденокарциномы легкого NCI-h560 GW843682X может вызывать временную остановку G2-M, потерю митотического веретена, образование многоядерных клеток и, в конечном итоге, приводить к апоптозу раковых клеток, в то время как нормальные диплоидные фибробласты умирают редко. BI 2536 представляет собой дигидроптериридоновое соединение, которое связывается с доменом PLK1 и ингибирует активность PLK1, вызывая нарушения митоза опухолевых клеток, такие как остановка клеток в фазе G2 / M, тем самым вызывая апоптоз. IC50 для ингибирования активности PLK1 со стороны BI2536 составляет 0.83 нмоль / л, а ингибирование PLK1 в 10000 раз больше, чем других 63 протеинкиназ. BI 2536 может подавлять пролиферацию 32 линий раковых клеток человека (EC50 = 2 ~ 25 нмоль / л). Клетки, обработанные BI 2536, задерживались в фазе G2-M, образуя монополярное веретено, подобное изменению после интерференции siRNA с PLK1. После инъекции мышей с опухолями (включая клетки A549 и NCI-h560) два раза в неделю с помощью BI 2536 40 ~ 50 мг / кг, количество блокированных поло-блокировкой раковых клеток постепенно увеличивалось с последующим множественным апоптозом. Описание BI 2536 может опосредовать остановку митоза и апоптоз в раковых клетках.RO3280 также является новым АТФ-конкурентным ингибитором PLK1 с сильным ингибированием и хорошей селективностью в отношении PLK1, который не только обладает противоопухолевой активностью in vitro , но также оказывает сильное противораковое действие на моделях опухоли ксенотрансплантата у мышей.
Опухоль
Как упоминалось выше, сигнальный путь PLK играет важную роль в развитии и распространении рака, и текущие исследования также продемонстрировали, что ингибиторы PLK могут лечить рак.
Список литературы
- Талати Ч., Гриффитс Э.А., Ветцлер М. и др. Ингибиторы поло-подобных киназ при гематологических злокачественных новообразованиях. Критические обзоры в онкологии. 2016, 98: 200-210.
- Бусси К. Дж., Бапат А., Линнехан С. и др. Ориентация на поло-подобную киназу 1, регулятор p53, при лечении карциномы коры надпочечников. Клиническая и трансляционная медицина. 2016, 5 (1): 1.
- Кан Г. И, Ли Э. Р., Ким Дж. Х. и др. Подавление экспрессии PLK-1 при апоптозе клеток MCF-7, вызванном кемпферолом. Европейский журнал фармакологии. 2009, 611 (1): 17-21.
- Джанг И Дж, Джи Дж Х, Чой Й Си и др. Регулирование поло-подобной киназы 1 повреждением ДНК в митозе. Ингибирование митотической PLK-1 протеинфосфатазой 2A. Журнал биологической химии. 2007, 282 (4): 2473.
Вернуться к ресурсам
Твиттер Facebook
| Кат. | Информация | Ссылки на использование продуктов | Подтверждения продукции |
|---|---|---|---|
| S1109 |
BI 2536BI-2536 представляет собой мощный ингибитор Plk1 с IC50 0,83 нМ в бесклеточном анализе. BI-2536 ингибирует бромодомен 4 (BRD4) с Kd 37 нМ и сильно подавляет экспрессию c-Myc . BI-2536 индуцирует апоптоз и ослабляет аутофагию . Этап 2. |
||
| S2235 |
Воласертиб (BI 6727)Воласертиб (BI 6727) является сильнодействующим ингибитором Plk1 с IC50 , равным 0.87 нМ в бесклеточном анализе. Он показывает в 6 и 65 раз большую селективность против Plk2 и Plk3. Воласертиб вызывает остановку клеточного цикла и апоптоз в различных раковых клетках. Этап 3. |
Вестерн-блоттинг. Клетки HeLa или MCF7 обрабатывали нокодазолом (noc, 50 нг / мл), паклитакселом (pacli, 50 нМ), ингибитором Plk1 BI 2536 (50 нМ) или BI 6727 (50 нМ) в течение 16 часов и получали клеточные экстракты для вестерн-блоттинг с указанными антителами.минус: клеточные экстракты из контрольных клеток без какой-либо обработки. β-актин служил контролем нагрузки. |
|
| S2758 |
Вортманнин (KY 12420)Вортманнин (KY 12420, SL-2052, BRN 0067676, NSC 627609) является первым описанным ингибитором PI3K с IC50 3 нМ в бесклеточном анализе с низкой селективностью в пределах семейства PI3K. Вортманнин блокирует образование аутофагосомы и эффективно ингибирует DNA-PK / ATM с IC50 16 нМ и 150 нМ в бесклеточных анализах.Вортманнин также подавляет активность PLK1 . |
клеток L3.6pl при 6000 клеток на лунку инкубировали в MEM с 5% FBS в трех экземплярах в 96-луночном культуральном планшете, а затем обрабатывали отдельно 5 мкмоль / л BMS-777607, 10 мкмоль / л вортманнина или BMS- 777607 в сочетании с индивидуальными ингибиторами. Полиплоидию исследовали под микроскопом BK71 Olympus и сфотографировали через 72 часа после обработки. |
|
| S1362 |
Ригосертиб (ON-01910)Ригосертиб (ON-01910) представляет собой неконкурентный с АТФ ингибитор PLK1 с IC50 9 нМ в бесклеточном анализе.Он показывает в 30 раз большую селективность против Plk2 и не проявляет активности к Plk3. Ригосертиб ингибирует путь PI3K / Akt и активирует сигналы окислительного стресса. Ригосертиб индуцирует апоптоз в различных раковых клетках. Этап 3. |
Наряду с гибелью клеток иммуноблоттинг показывает ON 01910.Na вызывает гиперфосфорилирование RanGAP1, повышение экспрессии RanGAP1.SUMO1, но снижение экспрессии свободного немодифицированного RanGAP1. Жизнеспособные клетки SU-DHL-5 не были доступны для иммуноблоттинга при 0.5 мкМ ПО 01910.Na. |
|
| S2193 |
ГСК461364GSK461364 (GSK461364A) ингибирует очищенный Plk1 с помощью K i 2,2 нМ в бесклеточном анализе. Он более чем в 1000 раз селективен против Plk2 / 3. Этап 1. |
Клетки со сниженной экспрессией GRK5 более чувствительны к ингибированию PLK1. A показана гибель клеток в контроле или клетках, стабильно трансфицированных GRK5-shRNA после 72 часов обработки 0-120 нМ BI 2536, как определено путем исключения трипанового синего (n = 6 ).B показана гибель клеток в контроле или клетках, стабильно трансфицированных GRK5-shRNA, после 72-часовой обработки 0–120 нМ GSK461364, как определено по исключению трипанового синего (n = 4). |
|
| S7837 Новый |
Центринон (LCR-263)Центринон (LCR-263) представляет собой селективный и обратимый ингибитор поло-подобной киназы 4 (PLK4) с Ki 0,16 нМ. |
||
| S1485 |
HMN-214HMN-214 является пролекарством HMN-176, которое изменяет пространственную ориентацию клеток Plk1. |
Внутриклеточный транспорт плазмидной ДНК в присутствии и в отсутствие ингибиторов PLK1. Клетки PC3-PSMA трансфицировали полиплексами плазмидной ДНК, меченной PEI: флуоресцеин (зеленый), в массовом соотношении 1: 4. Клетки, обработанные A) контрольным носителем (ДМСО) и B) 25 нМ BI 2536, показаны через 48 часов после совместной обработки полиплексами. Клетки, обработанные C) контрольным носителем (DMSO) и D) HMN-214, показаны через 24 часа после совместной обработки полиплексами. Масштабная линейка = 20 мкм.Синий сигнал указывает на ядра, окрашенные DAPI, а белые стрелки указывают на секвестрацию плазмидной ДНК в околоядерном отделении рециркуляции (PNRC). Красные стрелки указывают на измененную локализацию плазмидной ДНК после обработки ингибиторами PLK1. Изображения представляют три независимых эксперимента (N = 3). (Для интерпретации ссылок на цвет в легенде этого рисунка читателю отсылается к веб-версии этой статьи.) |
|
| S2898 |
MLN0905MLN0905 — мощный ингибитор PLK1 с IC50 2 нМ. |
||
| S7248 |
Ro3280RO3280 (Ro5203280) — мощный, высокоселективный ингибитор Polo-подобной киназы 1 (PLK1) с IC50 3 нМ. |
RO3280 способствует образованию многоядерных клеток в клеточных линиях рака мочевого пузыря. (A) Репрезентативные изображения клеток, окрашенных DAPI (синий) и альфа-тубулином (красный), были исследованы под флуоресцентным микроскопом (увеличение 400 ×).Увеличенные клетки, содержащие несколько равномерно окрашенных ядер (многоядерные клетки), характерны для митотической катастрофы (белая стрелка). (B) Количественная оценка процента многоядерных клеток на основе анализа флуоресцентной микроскопии (данные представлены в виде среднего значения ± S.E.M., * P |
|
| S5807 |
HMN-176HMN-176 является активным метаболитом HMN-214, который обладает сильной противоопухолевой активностью в моделях ксенотрансплантатов мышей. HMN-176 эффективно ингибирует PLK-1 путем вмешательства в его нормальное внутриклеточное пространственное распределение в центросомах и вдоль структуры цитоскелета. |
||
| S7255 |
Онвансертиб (NMS-P937)Онвансертиб (NMS-P937, PCM-075, NMS1286937) представляет собой доступный перорально селективный ингибитор Polo-подобной киназы 1 (PLK1) с IC50 2 нМ, 5000-кратная селективность по сравнению с PLK2 / PLK3. Онвансертиб (NMS-P937) мощно вызывает остановку митотического клеточного цикла с последующим апоптозом в линиях раковых клеток и подавляет рост опухоли. Этап 1. |
||
| S7552 |
CFI-400945CFI-400945 является орально активным, сильным и селективным ингибитором поло-подобной киназы 4 (PLK4) со значением Ki 0.26 нМ. |
||
| S7720 |
SBE 13 HClSBE 13 HCl является мощным и селективным ингибитором PLK1 с IC50 200 пМ,> 4000-кратной селективностью по сравнению с киназой Aurora A, Plk2 и Plk3. |
||
| Кат. № | Информация | Ссылки на использование продуктов | Подтверждения продукции |
| S1109 |
BI 2536BI-2536 представляет собой мощный ингибитор Plk1 с IC50 из 0.83 нМ в бесклеточном анализе. BI-2536 ингибирует бромодомен 4 (BRD4) с Kd 37 нМ и сильно подавляет экспрессию c-Myc . BI-2536 индуцирует апоптоз и ослабляет аутофагию . Этап 2. |
||
| S2235 |
Воласертиб (BI 6727)Volasertib (BI 6727) представляет собой высокоэффективный ингибитор Plk1 с IC50 0,87 нМ в бесклеточном анализе.Он показывает в 6 и 65 раз большую селективность против Plk2 и Plk3. Воласертиб вызывает остановку клеточного цикла и апоптоз в различных раковых клетках. Этап 3. |
Вестерн-блоттинг. Клетки HeLa или MCF7 обрабатывали нокодазолом (noc, 50 нг / мл), паклитакселом (pacli, 50 нМ), ингибитором Plk1 BI 2536 (50 нМ) или BI 6727 (50 нМ) в течение 16 часов и получали клеточные экстракты для вестерн-блоттинг с указанными антителами. минус: клеточные экстракты из контрольных клеток без какой-либо обработки.β-актин служил контролем нагрузки. |
|
| S2758 |
Вортманнин (KY 12420)Вортманнин (KY 12420, SL-2052, BRN 0067676, NSC 627609) является первым описанным ингибитором PI3K с IC50 3 нМ в бесклеточном анализе с низкой селективностью в пределах семейства PI3K. Вортманнин блокирует образование аутофагосомы и эффективно ингибирует DNA-PK / ATM с IC50 16 нМ и 150 нМ в бесклеточных анализах.Вортманнин также подавляет активность PLK1 . |
клеток L3.6pl при 6000 клеток на лунку инкубировали в MEM с 5% FBS в трех экземплярах в 96-луночном культуральном планшете, а затем обрабатывали отдельно 5 мкмоль / л BMS-777607, 10 мкмоль / л вортманнина или BMS- 777607 в сочетании с индивидуальными ингибиторами. Полиплоидию исследовали под микроскопом BK71 Olympus и сфотографировали через 72 часа после обработки. |
|
| S1362 |
Ригосертиб (ON-01910)Ригосертиб (ON-01910) представляет собой неконкурентный с АТФ ингибитор PLK1 с IC50 9 нМ в бесклеточном анализе.Он показывает в 30 раз большую селективность против Plk2 и не проявляет активности к Plk3. Ригосертиб ингибирует путь PI3K / Akt и активирует сигналы окислительного стресса. Ригосертиб индуцирует апоптоз в различных раковых клетках. Этап 3. |
Наряду с гибелью клеток иммуноблоттинг показывает ON 01910.Na вызывает гиперфосфорилирование RanGAP1, повышение экспрессии RanGAP1.SUMO1, но снижение экспрессии свободного немодифицированного RanGAP1. Жизнеспособные клетки SU-DHL-5 не были доступны для иммуноблоттинга при 0.5 мкМ ПО 01910.Na. |
|
| S2193 |
ГСК461364GSK461364 (GSK461364A) ингибирует очищенный Plk1 с помощью K i 2,2 нМ в бесклеточном анализе. Он более чем в 1000 раз селективен против Plk2 / 3. Этап 1. |
Клетки со сниженной экспрессией GRK5 более чувствительны к ингибированию PLK1. A показана гибель клеток в контроле или клетках, стабильно трансфицированных GRK5-shRNA после 72 часов обработки 0-120 нМ BI 2536, как определено путем исключения трипанового синего (n = 6 ).B показана гибель клеток в контроле или клетках, стабильно трансфицированных GRK5-shRNA, после 72-часовой обработки 0–120 нМ GSK461364, как определено по исключению трипанового синего (n = 4). |
|
| S7837 Новый |
Центринон (LCR-263)Центринон (LCR-263) представляет собой селективный и обратимый ингибитор поло-подобной киназы 4 (PLK4) с Ki 0,16 нМ. |
||
| S1485 |
HMN-214HMN-214 является пролекарством HMN-176, которое изменяет пространственную ориентацию клеток Plk1. |
Внутриклеточный транспорт плазмидной ДНК в присутствии и в отсутствие ингибиторов PLK1. Клетки PC3-PSMA трансфицировали полиплексами плазмидной ДНК, меченной PEI: флуоресцеин (зеленый), в массовом соотношении 1: 4. Клетки, обработанные A) контрольным носителем (ДМСО) и B) 25 нМ BI 2536, показаны через 48 часов после совместной обработки полиплексами. Клетки, обработанные C) контрольным носителем (DMSO) и D) HMN-214, показаны через 24 часа после совместной обработки полиплексами. Масштабная линейка = 20 мкм.Синий сигнал указывает на ядра, окрашенные DAPI, а белые стрелки указывают на секвестрацию плазмидной ДНК в околоядерном отделении рециркуляции (PNRC). Красные стрелки указывают на измененную локализацию плазмидной ДНК после обработки ингибиторами PLK1. Изображения представляют три независимых эксперимента (N = 3). (Для интерпретации ссылок на цвет в легенде этого рисунка читателю отсылается к веб-версии этой статьи.) |
|
| S2898 |
MLN0905MLN0905 — мощный ингибитор PLK1 с IC50 2 нМ. |
||
| S7248 |
Ro3280RO3280 (Ro5203280) — мощный, высокоселективный ингибитор Polo-подобной киназы 1 (PLK1) с IC50 3 нМ. |
RO3280 способствует образованию многоядерных клеток в клеточных линиях рака мочевого пузыря. (A) Репрезентативные изображения клеток, окрашенных DAPI (синий) и альфа-тубулином (красный), были исследованы под флуоресцентным микроскопом (увеличение 400 ×).Увеличенные клетки, содержащие несколько равномерно окрашенных ядер (многоядерные клетки), характерны для митотической катастрофы (белая стрелка). (B) Количественная оценка процента многоядерных клеток на основе анализа флуоресцентной микроскопии (данные представлены в виде среднего значения ± S.E.M., * P |
|
| S5807 |
HMN-176HMN-176 является активным метаболитом HMN-214, который обладает сильной противоопухолевой активностью в моделях ксенотрансплантатов мышей. HMN-176 эффективно ингибирует PLK-1 путем вмешательства в его нормальное внутриклеточное пространственное распределение в центросомах и вдоль структуры цитоскелета. |
||
| S7255 |
Онвансертиб (NMS-P937)Онвансертиб (NMS-P937, PCM-075, NMS1286937) представляет собой доступный перорально селективный ингибитор Polo-подобной киназы 1 (PLK1) с IC50 2 нМ, 5000-кратная селективность по сравнению с PLK2 / PLK3. Онвансертиб (NMS-P937) мощно вызывает остановку митотического клеточного цикла с последующим апоптозом в линиях раковых клеток и подавляет рост опухоли. Этап 1. |
||
| S7552 |
CFI-400945CFI-400945 является орально активным, сильным и селективным ингибитором поло-подобной киназы 4 (PLK4) со значением Ki 0.26 нМ. |
||
| S7720 |
SBE 13 HClSBE 13 HCl является мощным и селективным ингибитором PLK1 с IC50 200 пМ,> 4000-кратной селективностью по сравнению с киназой Aurora A, Plk2 и Plk3. |
| Функциональный ключ | Позиция (я) | Описание Действия | Графическое изображение | Длина |
|---|---|---|---|---|
|
Этот подраздел «Патология и биотехнология» описывается влияние экспериментальной мутации одной или нескольких аминокислот на биологические свойства белка. < p> Подробнее … Мутагенез i | 67 | C → V в аналоговочувствительном мутанте; увеличенный каталитический карман для размещения аналогов пурина; когда он связан с G-130. Ручное утверждение, основанное на эксперименте i
| 1 | |
| Мутагенез i | 82 | K → M: потеря киназной активности.Не влияет на прогрессирование S-фазы. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 82 | K → R: потеря киназной активности. Не влияет на связывание RIOK2. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 130 | L → G у аналогово-чувствительного мутанта; увеличенный каталитический карман для размещения аналогов пурина; когда связан с V-67. Ручное утверждение, основанное на эксперименте i
| 1 | |
| Мутагенез i | 137 | S → A: Без изменений активности. Повышает активность и восстанавливает восстановление после контрольной точки повреждения ДНК; когда связан с D-210. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 137 | S → D: Повышает активность. Результат в блоке в G1 / S. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 176 | D → N: отменяет киназную активность. Ручное утверждение, основанное на эксперименте в i
| 1 | |
| Мутагенез i | 194 | D → A: Не мешает связыванию FRY. Ручное утверждение на основе эксперимента в i | 1 | |
| Мутагенез i | 210 | T → A: Отменяет активность. Отменяет восстановление контрольной точки. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 210 | T → D: Повышает активность и восстанавливает восстановление после контрольной точки повреждения ДНК. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 210 | T → V: пониженная каталитическая активность, но не влияет на сродство к АТФ. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 337 | R → A: препятствует убиквитинированию и последующей протеасомной деградации в анафазе; при связке с А-340. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 340 | L → A: препятствует убиквитинированию и последующей протеасомной деградации в анафазе; когда связан с А-337. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 414 | W → F: устраняет взаимодействие с CDC25C и снижает центросомную локализацию. Ручное утверждение, основанное на эксперименте в i | 1 | |
| Мутагенез i | 414 | W → F: Не влияет ни на центросомную локализацию, ни на прогрессию S-фазы; при ассоциации с A-427. Утрата центросомной локализации и прогрессирования S-фазы; когда он связан с A-415 и A-427. Ручное утверждение, основанное на эксперименте в i | 1 | |
| Мутагенез i | 415 | V → A: потеря центросомной локализации и прогрессирования S-фазы; в сочетании с A-414 и A-427. Ручное утверждение, основанное на эксперименте в i | 1 | |
| Мутагенез i | 427 | L → A: не влияет ни на центросомную локализацию, ни на прогрессию S-фазы; когда связан с A-414. Утрата центросомной локализации и прогрессирования S-фазы; когда он связан с A-414 и A-415. Ручное утверждение, основанное на эксперименте в i | 1 | |
| Мутагенез i | 492 | K → R: Серьезные митотические дефекты, приводящие к задержке прометафазы.Повышенная локализация на кинетохорах, приводящая к повышению уровня фосфорилированного BUBR1. Ручное утверждение, основанное на эксперименте i
| 1 | |
| Мутагенез i | 538 | H → A в клещевом мутанте; потеря центросомного расположения и снижение взаимодействия с фосфорилированными CDC25C и BUB1; при связке с М-540. Ручное утверждение на основе эксперимента в i
| 1 | |
| Мутагенез i | 540 | K → M в клещевом мутанте; потеря центросомного расположения и снижение взаимодействия с фосфорилированными CDC25C и BUB1; когда связан с А-538. Ручное утверждение на основе эксперимента в i
| 1 |
Мотив связывания фосфо-PLK (ST * P) (D73F6) Кролик mAb
Ограниченное использование
Если иное прямо не согласовано в письменной форме, подписанной законным представителем CST, следующие условия: применяются к Продуктам, предоставляемым CST, ее аффилированными лицами или ее дистрибьюторами. Любые условия и положения Клиента, которые находятся в дополняют или отличаются от содержащихся в настоящем документе, если иное не принято в письменной форме юридически уполномоченным представитель CST, отклоняются и не имеют силы.
Продукты имеют маркировку «Только для исследовательского использования» или аналогичное заявление о маркировке и не были одобрены, одобрены или лицензированы. FDA или другой регулирующей иностранной или отечественной организацией для любых целей. Заказчик не должен использовать какой-либо Продукт для диагностики. или в терапевтических целях, или иным образом любым способом, который противоречит заявлению на этикетке. Продукты, продаваемые или лицензируемые CST предоставляются Заказчику как конечному пользователю и исключительно для использования в исследованиях и разработках.Любое использование Продукта для диагностики, в профилактических или терапевтических целях, или любая покупка Продукта для перепродажи (отдельно или в качестве компонента) или в других коммерческих целях, требуется отдельная лицензия от CST. Клиент обязуется (а) не продавать, лицензировать, предоставлять взаймы, дарить или иным образом передавать или делать доступными любой Продукт для любой третьей стороны, отдельно или в сочетании с другими материалами, или использовать Продукты для производства любых коммерческие продукты, (б) не копировать, изменять, реконструировать, декомпилировать, дизассемблировать или иным образом пытаться обнаружить лежащие в основе структуру или технологию Продуктов, или использовать Продукты с целью разработки любых продуктов или услуг, которые конкурировать с продуктами или услугами CST, (c) не изменять и не удалять из Продуктов какие-либо товарные знаки, торговые наименования, логотипы, патенты или уведомления об авторских правах или маркировка, (d) использовать Продукты исключительно в соответствии с Условия продажи продуктов CST и любые применимые документации, и (e) соблюдать любую лицензию, условия обслуживания или аналогичное соглашение в отношении любых сторонних продуктов или услуги, используемые Клиентом в связи с Продуктами.
Cell Signaling Technology является товарным знаком Cell Signaling Technology, Inc.
Для использования антител по мотивам технологии передачи сигналов (CST) в определенных методах (например, патенты США № 7,198,896 и 7,300,753) может потребоваться лицензия от CST. Для получения информации об условиях лицензирования для учебных заведений попросите ваш офис передачи технологий связаться с юридическим отделом CST по адресу [адрес электронной почты защищен]. Для получения информации об условиях коммерческого лицензирования свяжитесь с отделом CST Pharma Services по адресу [адрес электронной почты защищен]LANCE Ultra ULight ™ -PLK (Ser137) пептид, 0.125 нмоль
проверить количество ложныйТолько для исследовательских целей; не для диагностических процедур. Все продукты должны использоваться в соответствии с применимыми законами и правилами, включая, помимо прочего, требования к потреблению и утилизации в соответствии с европейскими правилами REACH (EC 1907/2006).
Запросить дополнительную информацию купить сейчас Добавить в корзину Запрос цитатыПожалуйста, введите действительное количество
Пожалуйста, авторизуйтесь, чтобы добавить в избранное.
НУЛЕВАЯ ИЛИ ПУСТАЯ КОРЗИНА
ОбзорLANCE Ultra TR-FRET технология обеспечивает двухкомпонентный анализ киназы, который сочетает прямое мечение субстрата киназы с U Light ™, инновационным эмиссионным красителем для оптимальной эффективности связывания, сокращения этапов и времени подготовки анализа. Подтвержденные для более чем 310 киназ, мы обеспечиваем прозрачность данных, предоставляя:
- Условия тестирования: установка для быстрого перехода к оптимизированному анализу
- Данные S: B: уверенность в доказанной эффективности киназы на определенных субстратах; не угадывать
- Фосфомотивы: предоставляются последовательности субстрата
- Варианты анализа: тестируются несколько субстратов, чтобы вы могли выбрать лучшую пептидную последовательность и S: B для своей киназы
Для субстратов U Light :
- 0.5 нмолей = 1000 точек анализа
- 5 нмолей = 10000 точек анализа
В анализах киназы LANCE Ultra связывание Eu-меченного антифосфосубстратного антитела с фосфорилированным субстратом, меченным U Light , приносит донора и молекулы акцептора в непосредственной близости. После облучения киназной реакции на длине волны 320 или 340 нм энергия донора Eu передается акцепторному красителю U Light , который, в свою очередь, генерирует свет на длине волны 665 нм.Интенсивность светового излучения пропорциональна уровню фосфорилирования субстрата U Light .
Характеристики
| Совместимость с автоматикой | Есть |
|---|---|
| Метод обнаружения | Флуоресценция с временным разрешением (TRF), TR-FRET |
| Экспериментальный тип | In vitro |
| Флуорофор | ULight |
| Молекулярная модификация | Фосфорилирование |
| Торговая марка продукта | LANCE Ультра |
| Состояние при доставке | Сухой лед |
| Размер блока | 0.
Похожие записи
|