Как устроена сотовая сеть и обеспечивается мобильная связь. Что такое базовые станции и соты. Как происходит подключение и передача сигнала в сети GSM. Какие стандарты связи существуют и чем отличается 5G.
Основные принципы работы сотовой связи
Сотовая связь — это вид мобильной радиосвязи, основанный на принципе деления территории на небольшие зоны покрытия, называемые сотами. Каждая сота обслуживается базовой станцией, которая поддерживает связь с мобильными устройствами в пределах своей зоны действия.
Ключевые элементы сотовой сети включают:
- Базовые станции с антеннами
- Контроллеры базовых станций
- Центры коммутации
- Мобильные устройства абонентов
Как происходит соединение при звонке с мобильного телефона?
- Телефон связывается с ближайшей базовой станцией по радиоканалу
- Сигнал передается от базовой станции в центр коммутации
- Центр коммутации определяет местоположение вызываемого абонента
- Устанавливается соединение через соответствующие базовые станции
Устройство и принцип работы базовых станций
Базовая станция — это основной элемент сотовой сети, обеспечивающий радиосвязь с мобильными устройствами. Она состоит из следующих компонентов:

- Приемо-передающие антенны (обычно 3-6 штук)
- Радиомодули
- Контроллер базовой станции
- Источники питания
- Система охлаждения
Антенны базовых станций обычно размещают на специальных мачтах или крышах высоких зданий. Радиус действия одной станции в городе составляет 300-1000 метров, вне города — до 35 км.
Как работает базовая станция?
- Антенны постоянно сканируют эфир, принимая сигналы от мобильных устройств
- При обнаружении сигнала станция выделяет свободный канал связи
- Происходит обмен служебными данными между станцией и телефоном
- Устанавливается голосовое или data-соединение
- Станция поддерживает связь, пока устройство находится в зоне ее действия
Принцип сотовой структуры сети
Сотовая структура позволяет эффективно использовать выделенный частотный диапазон. Основные принципы:
- Вся территория покрытия делится на небольшие соты
- Каждая сота обслуживается отдельной базовой станцией
- Соседние соты используют разные частоты во избежание помех
- Частоты повторно используются в удаленных сотах
- При перемещении абонента происходит переключение между сотами
Такая структура позволяет:

- Увеличить емкость сети
- Снизить мощность передатчиков
- Обеспечить непрерывность связи при перемещении
Процесс подключения мобильного телефона к сети
При включении мобильного устройства происходит следующий процесс подключения к сотовой сети:
- Телефон сканирует эфир в поисках сигнала базовой станции
- Устройство отправляет свой уникальный идентификатор IMSI
- Базовая станция передает IMSI в центр коммутации
- Проверяется наличие абонента в базе HLR и его права на обслуживание
- При успешной авторизации устройство регистрируется в VLR
- Телефону назначается временный идентификатор TMSI
- Устройство получает доступ к сетевым сервисам
Весь процесс занимает доли секунды и происходит автоматически при каждом включении телефона или переходе в зону действия нового оператора.
Эволюция стандартов сотовой связи
За время существования сотовой связи сменилось несколько поколений технологий:
- 1G — аналоговая связь (AMPS, NMT)
- 2G — цифровая связь (GSM, CDMA)
- 3G — мобильный широкополосный доступ (UMTS, CDMA2000)
- 4G — высокоскоростной мобильный интернет (LTE)
- 5G — сверхвысокие скорости и минимальные задержки
Каждое новое поколение обеспечивало более высокие скорости передачи данных и новые возможности для пользователей. Если 1G позволяла только голосовые вызовы, то 5G открывает путь к интернету вещей и беспилотному транспорту.
Особенности технологии 5G
5G — это новейший стандарт сотовой связи, обладающий рядом революционных преимуществ:
- Скорость передачи данных до 20 Гбит/с
- Сверхнизкие задержки менее 1 мс
- Возможность подключения до 1 млн устройств на 1 км²
- Повышенная энергоэффективность
- Улучшенное покрытие и доступность сигнала
Области применения 5G:
- Беспилотный транспорт
- Промышленный интернет вещей
- Дополненная и виртуальная реальность
- Телемедицина
- Умные города
Внедрение 5G потребует существенной модернизации сетевой инфраструктуры, но откроет возможности для инновационных сервисов и приложений.
Проблемы безопасности в сотовых сетях
- Перехват и прослушивание разговоров
- Клонирование SIM-карт
- Подмена базовых станций
- DDoS-атаки на сетевую инфраструктуру
- Утечка персональных данных абонентов
Для защиты от этих угроз применяются различные методы:
- Шифрование передаваемых данных
- Аутентификация абонентов и устройств
- Анализ аномалий в сети
- Изоляция критически важных элементов инфраструктуры
- Строгий контроль доступа к абонентским данным
Тем не менее, появление новых технологий вроде 5G создает и новые векторы атак, требуя постоянного совершенствования систем безопасности.
Как работает мобильная связь: соты, стандарты и возможности 5G — Технологии
Сотовая связь считается одним из самых полезных изобретений человечества — наряду с колесом, электричеством, интернетом и компьютером. И лишь за несколько десятилетий эта технология пережила целый ряд революций. С чего начиналось беспроводное общение, как работают соты и какие возможности откроет новый мобильный стандарт
5G?- Первое использование подвижной телефонной радиосвязи относится к 1921 году — тогда в США полиция Детройта использовала одностороннюю диспетчерскую связь в диапазоне 2 МГц для передачи информации от центрального передатчика к приемникам в автомобилях полицейских.
Как появилась сотовая связь
Впервые идея сотовой связи была выдвинута в 1947 году — над ней работали инженеры из Bell Labs Дуглас Ринг и Рэй Янг. Однако реальные перспективы ее воплощения стали вырисовываться только к началу 1970-х годов, когда сотрудники компании разработали рабочую архитектуру аппаратной платформы сотовой связи.
Так, американские инженеры предложили размещать передающие станции не в центре, а по углам «ячеек», а чуть позже была придумана технология, позволяющая абонентам передвигаться между этими «сотами», не прерывая связи. После этого осталось разработать действующее оборудование для такой технологии.
Задачу успешно решила компания Motorola — ее инженер Мартин Купер 3 апреля 1973 года продемонстрировал первый работающий прототип мобильного телефона. Он позвонил начальнику исследовательского отдела компании-конкурента прямо с улицы и рассказал ему о собственных успехах.
Руководство Motorola немедленно вложило в перспективный проект 100 миллионов долларов, однако на коммерческий рынок технология вышла только через десять лет. Такая задержка связана с тем, что сначала требовалось создать глобальную инфраструктуру базовых станций сотовой связи.
На территории США этой работой занялась компания AT&T — телекоммуникационный гигант добился от федерального правительства лицензирования нужных частот и построил первую сотовую сеть, которая охватила крупнейшие американские города. В качестве первого мобильника выступила знаменитая модель Motorola DynaTAC 8000. В продажу первый сотовый телефон поступил 6 марта 1983 года. Он весил почти 800 граммов, мог работать на одном заряде 30 минут в режиме разговора и заряжался около 10 часов. При этом аппарат стоил 3995 долларов — баснословную сумму по тем временам. Несмотря на это, мобильник мгновенно стал популярен.
Почему связь называется сотовой
Принцип мобильной связи прост — территория, на которой обеспечивается соединение абонентов, разбивается на отдельные ячейки или «соты», каждую из которых обслуживает базовая станция. При этом в каждой «соте» абонент получает идентичные услуги, поэтому сам он никак не чувствует пересечения этих виртуальных границ.
Обычно базовая станция в виде пары железных шкафов с оборудованием и антенн размещается на специально построенной вышке, однако в городе их нередко размещают на крышах высотных зданий. В среднем каждая станция ловит сигнал от мобильных телефонов на удалении до 35 километров.
Для улучшения качества обслуживания операторы также устанавливают фемтосоты — маломощные и миниатюрные станции сотовой связи, предназначенные для обслуживания небольшой территории. Они позволяют резко улучшить покрытие в тех местах, где это необходимо.Сотовую связь в России объединят с космосом
Находящийся в сети мобильник прослушивает эфир и находит сигнал базовой станции. В современную SIM-карту, кроме процессора и оперативки, вшит уникальный ключ, позволяющий авторизоваться в сотовой сети. Связь телефона со станцией может осуществляться по разным протоколам — например, цифровым DAMPS, CDMA, GSM, UMTS.
Сотовые сети разных операторов соединены друг с другом, а также со стационарной телефонной сетью. Если телефон выходит из поля действия базовой станции, аппарат налаживает связь с другими — установленное абонентом соединение незаметно передается другим «сотам», что обеспечивает непрерывную связь при перемещениях.
В России для вещания сертифицированы три диапазона — 800 МГц, 1800 МГц и 2600 МГц. Диапазон 1800 МГц считается самым популярным в мире, так как сочетает высокую емкость, большой радиус действия и высокую проникающую способность. Именно в нем сейчас работают большинство мобильных сетей.
Какие стандарты мобильной связи бывают
Первые мобильники работали с технологий 1G — это самое первое поколение сотовой связи, которое опиралось на аналоговые телекоммуникационные стандарты, главным из которых стал NMT — Nordic Mobile Telephone. Он предназначался исключительно для передачи голосового трафика.
К 1991 году относят рождение 2G — главным стандартом нового поколения стал GSM (Global System for Mobile Communications). Данный стандарт поддерживается до сих пор. Связь в этом стандарте стала цифровой, появилась возможность шифрования голосового трафика и отправки СМС.
Скорость передачи данных внутри GSM не превышала 9,6 кбит/с, что делало невозможной передачу видео или высококачественного звука. Проблему был призван решить стандарт GPRS, известный как 2.5G. Он впервые позволил пользоваться сетью Интернет владельцам мобильных телефонов.
Такой стандарт уже обеспечил скорость передачи данных до 114 Кбит/c. Однако вскоре он также перестал удовлетворять постоянно растущие запросы пользователей. Для решения этой проблемы в 2000 году был разработан стандарт 3G, который обеспечивал доступ к услугам Сети на скорости передачи данных в 2 Мбита.
Еще одним отличием 3G стало присвоение каждому абоненту IP-адреса, что позволило превратить мобильники в маленькие компьютеры, подключенные к интернету. Первая коммерческая сеть 3G была запущена 1 октября 2001 года в Японии. В дальнейшем пропускная способность стандарта неоднократно увеличивалась.
Наиболее современный стандарт — связь четвертого поколения 4G, которая предназначена только для высокоскоростных сервисов передачи данных. Пропускная способность сети 4G способна достигать 300 Мбит/сек, что дает пользователю практически неограниченные возможности работы в интернете.
Сотовая связь будущего
Стандарт 4G заточен на непрерывную передачу гигабайтов информации, в нем даже отсутствует канал для передачи голоса. За счет чрезвычайно эффективных схем мультиплексирования загрузка фильма высокого разрешения в такой сети займет у пользователя 10-15 минут. Однако даже его возможности уже считаются ограниченными.
В 2020 году ожидается официальный запуск нового поколения связи стандарта 5G, который позволит передачу больших объемов данных на сверхвысоких скоростях до 10 Гбит/сек. Кроме этого, стандарт позволит подключить к высокоскоростному интернету до 100 миллиардов устройств.
Именно 5G позволит появиться настоящему интернету вещей — миллиарды устройств будут обмениваться информацией в реальном времени. По оценке экспертов, сетевой трафик скоро вырастет на 400%. Например, автомобили начнут постоянно находиться в глобальной Сети и получать данные о дорожной обстановке.
ЕЩЕ ПО ТЕМЕ:
Первый смартфон 5G появится раньше необходимых сетейНизкая степень задержки обеспечит связь между транспортными средствами и инфраструктурой в режиме реального времени. Ожидается, что надежное и постоянно действующее соединение впервые откроет возможность для запуска на дорогах полностью автономных транспортных средств.
Российские операторы уже экспериментируют с новыми спецификациями — например, работы в этом направлении ведет «Ростелеком». Компания подписала соглашение о строительстве сетей 5G в инновационном центре «Сколково». Реализация проекта входит в государственную программу «Цифровая экономика», недавно утвержденную правительством.
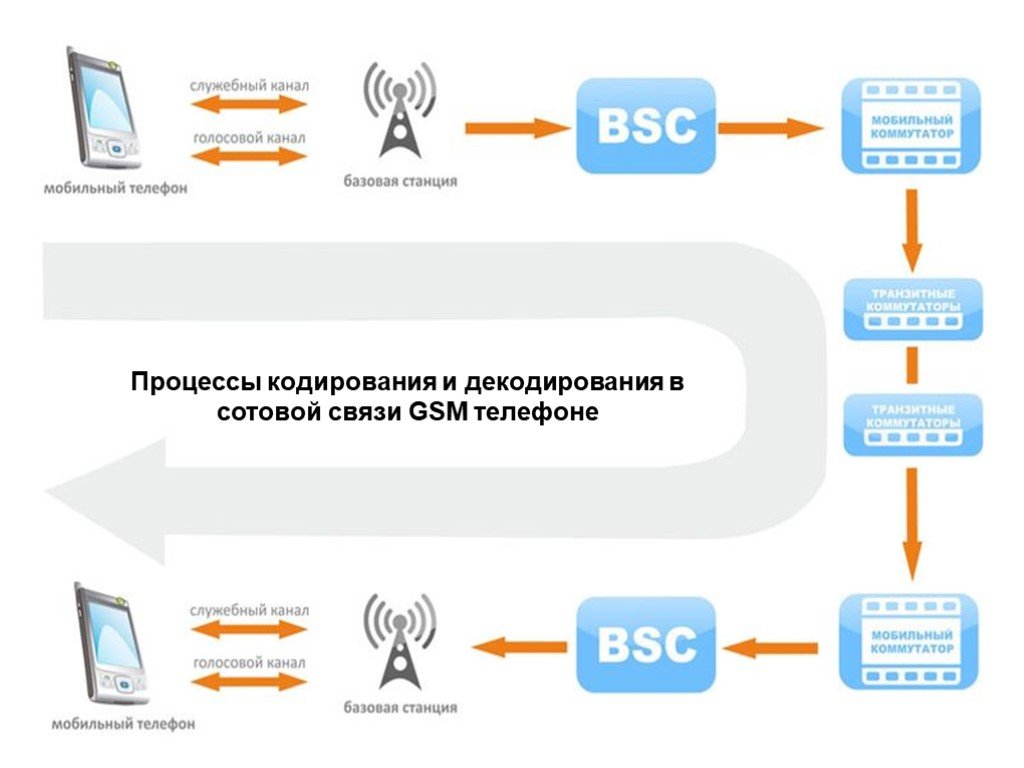
Как работает сотовая связь
Многие задумываются о том, как работают сотовые сети, что происходит, когда мы нажимаем на кнопку вызова? Звоня друг другу, зачастую вызываемый абонент находится не только в пределах одного города, но даже и на другом континенте. Как же работает сотовая связь?ПРИНЦИП ДЕЙСТВИЯ СОТОВОЙ СВЯЗИ
На большей части территории нашей страны размещается оборудование для сотовой связи, называется оно базовые станции. Их хорошо заметно на открытых площадях – в полях, между населенными пунктами. В городской черте их часто размещают на крышах зданий. Базовая станция способна уловить сигнал от смартфона на расстоянии до тридцати пяти километров, контакт между вышками осуществляется посредством специального служебного или голосового сигнала.Активное развитие мобильной связи породило проблему, заключающуюся в ограничении частоты, а именно, рабочие каналы, расположенные близко, начали перекликаться, создавая помехи. Много лет наза была предложена идея, по которой определенный участок обслуживания оператором сотовой связи необходимо разбить на ячейки. Каждая ячейка обслуживается специальным передатчиком, предполагающим фиксированный частотный диапазон и радиус действия. Такая система исключает помехи при использовании той же частоты, но уже в другой соте. Чтобы разделить определенную площадь на равные участки наиболее оптимальной является фигура с шестью углами, напоминающая пчелиную соту, так как установленная в центе соты антенна с круговой диаграммой будет обеспечивать свободный устойчивый доступ ко всем точкам ячейки. У всех сот есть собственная полоса частот и обслуживающая базовая станция. Ячейки смежного расположения не используют одинаковые частоты, тем самым исключая перекрестные помехи и интерференции, и наоборот, соты, располагающиеся далеко друг от друга могут использовать идентичные частоты.
КАК РАБОТАЮТ БАЗОВЫЕ СТАНЦИИ
Когда смартфон пребывает в режиме ожидания, его приёмный механизм сканирует каналы системы. Если пользователь собираясь совершить звонок набирает номер аппарат автоматически находит станцию, которая располагается к нему ближе и посылает запрос о выделении голосового канала. Те базовые станции, которые принимают ответный сигнал, перенаправляют его данные в центр коммутации, где происходит переключение разговора на ближайшую станцию к вызываемому абоненту с более высоким уровнем сигнала. В центре коммутации, также, определяют, какой оператор мобильной связи используется вызываемым абонентом.В том случае, если звонок осуществляется между абонентами внутри одной сети, то в центре коммутации сразу происходит идентификация месторасположения вызываемого абонента, причем, неважно где находится человек: дома, в транспорте или в командировке в другой стране. Физическое месторасположение абонента ни коим образом не помешает соединению и осуществлению звонка. Если в центр коммутации поступает информация о том, что вызываемый абонент использует оператора другой связи, тогда запрос будет отправлен в центр коммутации другой сети. В общем-то, выходит, что система довольно проста, и как работает сотовая связь понятно. Интересным остается вопрос, как же выглядит устройство базовой станции: и здесь все просто – это всего лишь несколько металлических тумб, располагающихся на крышах зданий и для бесперебойной их работы достаточно качественной вентиляции.
ПРОБЛЕМЫ УСТОЙЧИВОСТИ СВЯЗИ
Понятно, что в момент набора номера аппарат занимает незанятый канал с максимально возможным уровнем сигнала. Но, если в процессе разговора абонент начинает удаляться от базовой станции или условия расширения радиоволн ухудшатся – все это неблагоприятным образом скажется на связи и ее качестве. Логично, что ее улучшение происходит после переключения абонентов на другие, более устойчивые, каналы связи.Каждая базовая станция имеет антенну, состоящую из нескольких элементов, так называемых секторов, отвечающих за «свою» площадь. Вертикальная составляющая антенны ответственна за связь с мобильными аппаратами, круглая – с контроллером. С учетом того, что одна станция чаще всего состоит из 6-ти секторов, и каждый из них способен принять минимум 70 звонков, после нехитрых вычислений выходит, что обслужить более 400 абонентов одновременно для нее не проблема. Такой производительности, зачастую, вполне достаточно. Но, случаются и внештатные ситуации, когда все абоненты всех операторов мобильной связи начинают звонить, например, на большие праздники (Новый Год), и базовые станции просто не справляются — начинаются перебои и помехи. Тем не менее для средней загрузки шести секторов более чем достаточно.
Следует отметить, что в зависимости от площади населенного пункта и плотности населения операторы мобильной связи устанавливают базовые станции с разным диапазоном частот:
• 900 МГц. Установка такой станции более целесообразна в небольших городках, поселках городского типа и т.д. В данном режиме базовая станция охватывает площадь радиусом порядка 35 км, или даже 70 км если на данный момент она обслуживает малое количество мобильных устройств.
• 1 800 МГц. Оптимальный вариант для больших городов, когда необходимо проникнуть сквозь толщину бетонной стены, однако, даже при таком диапазоне частот в городской черте базовых станций понадобится намного больше, чем в малонаселенных пунктах.
• 2 100 МГц. Это связь нового, более современного поколения 3G.
Одна базовая станция способна поддерживать сразу все возможные частотные диапазоны. Основная задача базовых станций заключается в том, чтобы покрыть максимальную площадь земли и обеспечить большое количество абонентов качественной связью. То есть улавливать сигналы на таких же расстояниях, но не на земле, а в воздушном пространстве базовые станции не могут.
Официальный магазин смартфонов Highscreen
Каталог мобильных телефонов Хайскрин
Принцип работы сетей GSM. Часть 1
Часть 1: структура мобильных сетей
Все мы пользуемся мобильными телефонами, но при этом редко кто задумывается — как же они работают? В данной статье мы постараемся разобраться, как, собственно, реализуется связь относительно вашего мобильного оператора.
Когда вы осуществляете звонок своему собеседнику, или кто-то звонит вам, ваш телефон соединяется по радиоканалу с одной из антенн соседней базовой станции (БС, BS, Base Station).Каждая базовая станция сотовой связи (в простонародье — вышки сотовой связи) включает в себя от одной до двенадцати приемо-передающих антенн, имеющих направления в разные стороны с целью обеспечения качественной связью абонентов в радиусе своего действия. Такие антенны специалисты на своем жаргоне называют «секторами», представляющими собой серые прямоугольные конструкции, которые вы можете практически каждый день видеть на крышах зданий или специальных мачтах.
Сигнал от такой антенны поступает по кабелю прямо в
управляющий блок базовой станции. Базовая станция является совокупностью
секторов и управляющего блока. При этом определенную часть населенного пункта
или территории обслуживают сразу несколько базовых станций, подключенных к
специальному блоку – контроллеру локальной зоны (сокращенно LAC, Local Area
Controller или просто «контроллер»). Как правило, один контроллер объединяет до
15 базовых станций определенного района.
Со своей стороны, контроллеры (их также может быть несколько) соединены с самым главным блоком — Центром управления мобильными услугами (MSC, Mobile services Switching Center), который для упрощения восприятия принято называть просто «коммутатором». Коммутатор, в свою очередь, осуществляет вход и выход на любые линии связи – как сотовой, так и проводной.
Если отобразить написанное в виде схемы, то получится следующее:GSM-сети небольшого масштаба (как правило, региональные) могут использовать всего один коммутатор. Крупные же, такие как наши операторы «большой тройки» МТС, Билайн или МегаФон, обслущивающие одновременно миллионы абонентов, используют сразу несколько объединенный между собой устройств MSC.
Давайте разберемся, зачем нужна столь сложная система и почему нельзя подключить антенны базовых станций к коммутатору напрямую? Для этого нужно рассказать про еще один термин, называемый на техническом языке handover (хэндовер). Он характеризует собой передачу обслуживания в мобильных сетях по эстафетному принципу. Иными словами, когда вы перемещаетесь по улице пешком или в транспортном средстве и говорите при этом по телефону, то, чтобы ваш разговор при этом не прерывался, следует своевременно переключать ваш аппарат из одного сектора БС в другой, из зоны действия одной базовой станции или контроллера локальной зоны в другую и т.д. Следовательно, если бы сектора базовых станций подключались к коммутатору напрямую, ему бы пришлось самому осуществлять данную процедуру хендовера всех своих абонентов, а у коммутатора и без того хватает задач. Поэтому для уменьшения вероятности отказов оборудования, связанных с его перегрузками, схема построения сотовых сетей GSM реализуется по многоуровнему принципу.
В итоге, если вы со своим телефоном перемещаетесь из зоны обслуживания одного сектора БС в зону действия другого, то данное перемещение осуществляет блок управления данной базовой станции, не касаясь при это более «высокостоящих» устройств – LAC и MSC. Если же хэндовер происходит между разными БС, то за него берется уже LAC и т. д.
Коммутатор – ни что иное, как основной «мозг» сетей GSM, поэтому его работу следует рассмотреть более детально. Коммутатор сотовой сети берет на себя примерно те же задачи, что и АТС в сетях проводных операторов. Именно он понимает, куда вы осуществляете звонок или кто звонит вам, регулирует работу дополнительных услуг и, собственно, решает – можете ли вы в настоящее время осуществить свой звонок или нет.
Теперь давайте разберемся, что же происходит, когда вы включаете свой телефон или смартфон?
Итак, вы нажали «волшебную кнопку» и ваш телефон включился. На SIM-карте вашего сотового оператора находится специальный номер, который носит название IMSI – International Subscriber Identification Number (Международный опознавательный номер абонента). Он является уникальным номером для кажой SIM-карты не только у вашего оператора МТС, Билайн, МегаФон и т.п., а уникальным номером для всех мобильных сетей в мире! Именно по нему операторы отличают абонентов между собой.
В момент включения телефона ваш аппарат посылает данный код IMSI на базовую станцию, которая передает его далее на LAC, он же, в свою очередь, отсылает его на коммутатор. При этом в нашу игру вступают два дополнительных устройства, свзанных непосредственно с коммутатором – HLR (Home Location Register) и VLR (Visitor Location Register). В переводе на русский это, соответственно, Регистр домашних абонентов и Регистр гостевых абонентов. HLR хранит в себе IMSI всех абонентов своей сети. В VLR же содержится информация о тех абонентах, которые пользуются сетью данного оператора в настоящее время.
Номер IMSI передается в HLR с помощью системы шифрования (за этот процесс отвечает еще одно устройство AuC — Центр аутентификации). HLR при этом проверяет, существует ли в его базе абонент с данным номером, и если факт его наличия подтверждается, система смотрит, может ли он в настоящее время пользоваться услугами связи или, скажем, имеет финансовую блокировку. Если все нормально, то данный абонент отправляется в VLR и после этого получает возможность звонить и пользоваться другими услугами связи.
Для наглядности отобразим данную процедуру с помощью схемы:
Таким образом, мы коротко описали принцип работы сотовых сетей GSM. На самом деле, это описание достаточно поверхностно, т.к. если углубиться в технические детали подробнее, то материал бы получился во много раз объемнее и гораздо менее понятным для большинства читателей.
Во второй части мы продолжим знакомство с работой сетей GSM и рассмотрим, как и за что оператор списывает средства с нашего с вами счета.
Перейти к части 2 >>
Перейти к части 3 >>
Сотовая связь — это… Что такое Сотовая связь?
Сотовая связь, сеть подвижной связи — один из видов мобильной радиосвязи, в основе которого лежит сотовая сеть. Ключевая особенность заключается в том, что общая зона покрытия делится на ячейки (соты), определяющиеся зонами покрытия отдельных базовых станций (БС). Соты частично перекрываются и вместе образуют сеть. На идеальной (ровной и без застройки) поверхности зона покрытия одной БС представляет собой круг, поэтому составленная из них сеть имеет вид сот с шестиугольными ячейками (сотами).
Сеть составляют разнесённые в пространстве приёмопередатчики, работающие в одном и том же частотном диапазоне, и коммутирующее оборудование, позволяющее определять текущее местоположение подвижных абонентов и обеспечивать непрерывность связи при перемещении абонента из зоны действия одного приёмопередатчика в зону действия другого.
История
Первое использование подвижной телефонной радиосвязи в США относится к 1921 г.: полиция Детройта использовала одностороннюю диспетчерскую связь в диапазоне 2 МГц для передачи информации от центрального передатчика к приёмникам, установленным на автомашинах. В 1933 г. полиция Нью-Йорка начала использовать систему двусторонней подвижной телефонной радиосвязи также в диапазоне 2 МГц. В 1934 г. Федеральная комиссия связи США выделила для телефонной радиосвязи 4 канала в диапазоне 30—40 МГц, и в 1940 г. телефонной радиосвязью пользовались уже около 10 тысяч полицейских автомашин. Во всех этих системах использовалась амплитудная модуляция. Частотная модуляция начала применяться с 1940 г. и к 1946 г. полностью вытеснила амплитудную. Первый общественный подвижный радиотелефон появился в 1946 г. (Сент-Луис, США; фирма Bell Telephone Laboratories), в нём использовался диапазон 150 МГц. В 1955 г. начала работать 11-канальная система в диапазоне 150 МГц, а в 1956 г. — 12-канальная система в диапазоне 450 МГц. Обе эти системы были симплексными, и в них использовалась ручная коммутация. Автоматические дуплексные системы начали работать соответственно в 1964 г. (150 МГц) и в 1969 г. (450 МГц).
В СССР в 1957 г. московский инженер Л. И. Куприянович создал опытный образец носимого автоматического дуплексного мобильного радиотелефона ЛК-1 и базовую станцию к нему. Мобильный радиотелефон весил около трех килограммов и имел радиус действия 20—30 км. В 1958 году Куприянович создаёт усовершенствованные модели аппарата весом 0,5 кг и размером с папиросную коробку. В 1960-х гг. Христо Бочваров в Болгарии демонстрирует свой опытный образец карманного мобильного радиотелефона. На выставке «Интероргтехника-66» Болгария представляет комплект для организации местной мобильной связи из карманных мобильных телефонов РАТ-0,5 и АТРТ-0,5 и базовой станции РАТЦ-10, обеспечивающей подключение 10 абонентов.
В конце 50-х гг в СССР начинается разработка системы автомобильного радиотелефона «Алтай», введённая в опытную эксплуатацию в 1963 г. Система «Алтай» первоначально работала на частоте 150 МГц. В 1970 г. система «Алтай» работала в 30 городах СССР и для неё был выделен диапазон 330 МГц.
Аналогичным образом, с естественными отличиями и в меньших масштабах, развивалась ситуация и в других странах. Так, в Норвегии общественная телефонная радиосвязь использовалась в качестве морской мобильной связи с 1931 г.; в 1955 г. в стране было 27 береговых радиостанций. Наземная мобильная связь начала развиваться после второй мировой войны в виде частных сетей с ручной коммутацией. Таким образом, к 1970 г. подвижная телефонная радиосвязь, с одной стороны, уже получила достаточно широкое распространение, но с другой — явно не успевала за быстро растущими потребностями, при ограниченном числе каналов в жёстко определённых полосах частот. Выход был найден в виде системы сотовой связи, что позволило резко увеличить ёмкость за счёт повторного использования частот в системе с ячеистой структурой.
Сотовые системы
Отдельные элементы системы сотовой связи существовали и раньше. В частности, некоторое подобие сотовой системы использовалось в 1949 г. в Детройте (США) диспетчерской службой такси — с повторным использованием частот в разных ячейках при ручном переключении каналов пользователями в оговорённых заранее местах. Однако архитектура той системы, которая сегодня известна как система сотовой связи, была изложена только в техническом докладе компании Bell System, представленном в Федеральную комиссию связи США в декабре 1971 года. С этого времени начинается развитие собственно сотовой связи.
В 1974 г. Федеральная комиссия связи США приняла решение о выделении для сотовой связи полосы частот в 40 МГц в диапазоне 800 МГц; в 1986 г. к ней было добавлено ещё 10 МГц в том же диапазоне. В 1978 г. в Чикаго начались испытания первой опытной системы сотовой связи на 2 тыс. абонентов. Поэтому 1978 год можно считать годом начала практического применения сотовой связи. Первая автоматическая коммерческая система сотовой связи была введена в эксплуатацию также в Чикаго в октябре 1983 г. компанией American Telephone and Telegraph (AT&T). В Канаде сотовая связь используется с 1978 г., в Японии — с 1979 г., в североевропейских странах (Дания, Норвегия, Швеция, Финляндия) — с 1981 г., в Испании и Англии — с 1982 г. По состоянию на июль 1997 г. сотовая связь работала более чем в 140 странах всех континентов, обслуживая более 150 млн абонентов.
Первой коммерчески успешной сотовой сетью была финская сеть Autoradiopuhelin (ARP). Это название переводится на русский как «Автомобильный радиотелефон». Запущенная в 1971 г., она достигла 100%-го покрытия территории Финляндии в 1978 году, а в 1986 году в ней было более 30 тыс. абонентов. Работала сеть на частоте 150 МГц, размер соты — около 30 км.
Принцип действия сотовой связи
Основные составляющие сотовой сети — это сотовые телефоны и базовые станции, которые обычно располагают на крышах зданий и вышках. Будучи включённым, сотовый телефон прослушивает эфир, находя сигнал базовой станции. После этого телефон посылает станции свой уникальный идентификационный код. Телефон и станция поддерживают постоянный радиоконтакт, периодически обмениваясь пакетами. Связь телефона со станцией может идти по аналоговому протоколу (AMPS, NAMPS, NMT-450) или по цифровому (DAMPS, CDMA, GSM, UMTS). Если телефон выходит из поля действия базовой станции (или качество радиосигнала сервисной соты ухудшается), он налаживает связь с другой (англ. handover).
Сотовые сети могут состоять из базовых станций разного стандарта, что позволяет оптимизировать работу сети и улучшить её покрытие.
Сотовые сети разных операторов соединены друг с другом, а также со стационарной телефонной сетью. Это позволяет абонентам одного оператора делать звонки абонентам другого оператора, с мобильных телефонов на стационарные и со стационарных на мобильные.
Операторы могут заключать между собой договоры роуминга. Благодаря таким договорам абонент, находясь вне зоны покрытия своей сети, может совершать и принимать звонки через сеть другого оператора. Как правило, это осуществляется по повышенным тарифам. Возможность роуминга появилась лишь в стандартах 2G и является одним из главных отличий от сетей 1G.[1]
Операторы могут совместно использовать инфраструктуру сети, сокращая затраты на развертывание сети и текущие издержки.
Сотовая связь в России
В России сотовая связь начала внедряться с 1990 г., коммерческое использование началось с 9 сентября 1991 г., когда в Санкт-Петербурге компанией «Дельта Телеком» была запущена первая в России сотовая сеть (работала в стандарте NMT-450) и был совершён первый символический звонок по сотовой связи мэром Санкт-Петербурга Анатолием Собчаком.
Руководитель Клуба региональной журналистики Ирина Ясина вспоминает[2]:
| Первый мобильный телефон был вот такой величины и назывался Nokia. Он стоил 5 тыс. долл. Его, как чемодан, надо было за собой носить. Когда мы выходили из машины, мы снимали дворники всегда, потому что их воруют. А крутой брал телефон из салона, шел в кафе, клал перед собой телефон, который занимал полстола. Это было смешно |
.
К июлю 1997 г. общее число абонентов в России составило около 300 тысяч. На 2007 год основные протоколы сотовой связи, используемые в России — GSM-900 и GSM-1800. Помимо этого, работают и CDMA-сети, в стандарте CDMA-2000, он же IMT-MC-450. Также GSM-операторами ведётся плавный переход на стандарт UMTS. В частности, первый фрагмент сети этого стандарта в России был введён в эксплуатацию 2 октября 2007 года в Санкт-Петербурге компанией «МегаФон».
Компания IDC на основе исследования российского рынка сотовой связи сделала вывод, что в 2005 году общая продолжительность разговоров по сотовому телефону жителей РФ достигла 155 миллиардов минут, а текстовых сообщений было отправлено 15 миллиардов штук.
Согласно данным британской исследовательской компании Informa Telecoms & Media за 2006 год, средняя стоимость минуты сотовой связи для потребителя в России составила $0,05 — это самый низкий показатель из стран «большой восьмёрки».[3]
В декабре 2007 года число пользователей сотовой связи в России выросло до 172,87 млн абонентов, в Москве — до 29,9, в Санкт-Петербурге — до 9,7 млн. Уровень проникновения в России — до 119,1 %, Москве — 176 %, Санкт-Петербурге — 153 %. В декабре 2011 года уровень проникновения в России — до 156 %, Москве — 212,1 %, Санкт-Петербурге — 215,6 %[4]. Доля рынка крупнейших сотовых операторов на декабрь 2007 года составила: МТС 30,9 %, «ВымпелКом» 29,2 %, «МегаФон» 19,9 %, другие операторы 20 %.[5]
Согласно исследованию компании J’son & Partners, количество зарегистрированных в России сим-карт по состоянию на конец ноября 2008 года достигло 183,8 млн[6]. Эта цифра обусловлена отсутствием абонентской платы на популярных тарифных планах у российских операторов сотовой связи и низкой ценой подключения к сети. Абоненты в ряде случаев имеют сим-карты разных операторов, при этом могут ими не пользоваться продолжительное время, либо использовать одну сим-карту в служебном мобильном телефоне, а другую — для личных разговоров[7].
В России в декабре 2008 г. насчитывалось 187,8 млн пользователей сотовой связи (по числу проданных сим-карт). Уровень проникновения сотовой связи (количество SIM-карт на 100 жителей) на эту дату составил, таким образом, 129,4 %. В регионах, без учёта Москвы, уровень проникновения превысил 119,7 %.[8]
Доля рынка крупнейших сотовых операторов на декабрь 2008 года составила: 34,4 % у МТС, 25,4 % у «Вымпелкома» и 23,0 % у «МегаФона».[8]
Уровень проникновения на конец 2009 года достиг 162,4 %.
По состоянию на апрель 2010 г. доля рынка в России по абонентам: МТС — 32,9 %, МегаФон — 24,6 %, Вымпелком — 24,0 %, Tele2 — 7,5 %, другие операторы — 11,0 %
Услуги сотовой связи
Операторы сотовой связи предоставляют следующие услуги:
Интересные факты
| В этом разделе не хватает ссылок на источники информации.
Информация должна быть проверяема, иначе она может быть поставлена под сомнение и удалена. Вы можете отредактировать эту статью, добавив ссылки на авторитетные источники. Эта отметка установлена 12 мая 2011. |
- Для работы первой автоматической системы сотовых телефонов требовался человек-оператор, вручную подключавший пользователей к внешней телефонной линии.
- Первые базовые станции для «КБ „Импульс“» (ныне «ВымпелКом») были созданы «с нуля» в Радиотехническом институте им. А. Л. Минца и по своим характеристикам не уступали зарубежным аналогам.
- Первая система сотовой связи появилась в СССР в 1950-е годы.
- Обычно телефонный номер, не используемый в течение 3-6 месяцев (в зависимости от правил оператора) с момента последнего исходящего звонка у абонента отбирают, он помещается в «отстойник» (необходимо, чтобы новому владельцу номера не звонили знакомые старого владельца — этот срок примерно или ровно три месяца), далее его выставляют на продажу. Некоторые операторы по истечении определённого срока отсутствия платных исходящих звонков (например, 90 дней), когда оператор имеет право изъять номер, сначала включают «услугу сохранения номера» (фактически — ежедневную абонентскую плату) и лишь тогда, когда баланс близок к нулю, номер изымается. В этом случае можно обратиться в офис своего сотового оператора с просьбой вернуть номер и, если номер ещё не продан другому человеку, то его вернут.[9] Однако оператор не обязан включать «сохранение номера» или возвращать утраченный номер.
См. также
Примечания
- ↑ О сотовой связи
- ↑ Семинары
- ↑ Тимофей Дзядко. Дешевле, чем в России, почти не бывает. Минута разговора по сотовому обходится в среднем в $0,05 // Ведомости, № 164 (1938), 3 сентября 2007
- ↑ Уровень проникновения сотовой связи в России составляет 156,8% | OSP News | Издательство «Открытые системы»
- ↑ NEWSru.com | Технологии | Число пользователей сотовой связи в России выросло до 172,87 млн абонентов
- ↑ Количество зарегистрированных в России сим-карт приблизилось к 184 миллионам
- ↑ В России на 100 человек приходится 140 сим-карт
- ↑ 1 2 Число пользователей сотовой связи в России в 2008 г выросло на 8,7 проц до 187,8 млн абонентов, проникновение увеличилось до 129,4 проц — AC&M Consulting /расширенная версия/
- ↑ Это подтверждают справочные службы сотовых операторов (Билайн, Мегафон).
Ссылки
Как зарождалась связь | Мир ПК
Фото: Wikipedia
Когда говорят об истории сотовой связи, прежде всего вспоминается 3 апреля 1973 г. Именно в этот день Мартин Купер, глава подразделения мобильной связи американской компании Motorola, совершил первый в мире звонок по мобильному телефону. И теперь он считается днем рождения мобильной связи в том виде, к какому мы привыкли. Но ее история началась гораздо раньше.
Начало пути
Наверное, первой и самой важной датой в истории мобильной связи следует считать 7 мая 1895 г., когда известный русский ученый Александр Степанович Попов продемонстрировал прибор, предназначенный для регистрации электромагнитных волн. Что интересно, изначально Попов не планировал создавать какие-либо средства радиосвязи, а разрабатывал «грозоотметчик», прибор для регистрации молний. Но, по сути, прибор Попова стал первым в мире радиоприемником, источником сигнала для которого служили грозовые разряды. Позднее, в сентябре 1895 г., вместо метрологического регистратора Попов подключил к своему «грозоотметчику» телеграфный аппарат Морзе, что еще больше приблизило его к средству для беспроводной передачи информации.
Следующим шагом к мобильной связи стали сеансы беспроводной телеграфной связи, проводимые Гульельмо Маркони. Причем если в 1896 г. информация передавалась на расстояние нескольких километров, то к концу 1901 г. сообщение Маркони было получено по ту сторону Атлантического океана. Свою роль сыграло и то, что Маркони обладал коммерческой жилкой, благодаря чему разработанная им технология стала коммерчески успешной, а компания «Маркони и К°» — известной на весь мир.
«Грозоотметчик» Попова – прибор, с которого началась
беспроводная радиосвязь
Не менее важен был и переход от использования абстрактных «точек-тире» к передаче живого человеческого голоса. Для исследователей-радиотехников тех лет это была одна из наиболее актуальных задач, в процессе решения которой были проведены сотни исследований и получены десятки патентов. Но наибольшего успеха добился Реджинальд Фессенден, в 1900 г. впервые передавший голос по радиоканалу, а к 1903 г. – получивший вполне приемлемое его качество. Датой «мобилизации» беспроводной радиосвязи стал 1901 г., когда Маркони установил приемо-передающее устройство на паровой автомобиль марки «Тоникрофт».
Так выглядел первый автомобиль,
оснащенный системой подвижной радиосвязи
Следующим ключевым стал 1921 г., когда в американском Детройте была запущена первая в мире диспетчерская система телеграфной подвижной связи, предназначенная для нужд местной полиции. Обмен информацией был односторонним – получив сигнал (азбукой Морзе), полицейские связывались с участком по обычному телефону. Фактически построенная в Детройте система была прототипом уже позабытой многими пейджинговой связи. Двусторонняя подвижная радиосвязь для помощи полиции появилась в 1933 г. в Нью-Йорке. Причем она уже была не телеграфной, а голосовой, хотя и работающей в полудуплексном режиме, т.е. для переключения между приемом и передачей нужно было нажимать кнопку.
Америка и Европа
Частным клиентам мобильная радиосвязь впервые стала доступна 17 июня 1946 г., когда в американском Сент-Луисе (штат Миссури) совместными усилиями AT&T и Bell Telephone Laboratories была запущена сеть стандарта MTS, работавшая на частоте 150 МГц. Принцип действия MTS-сети отличался от современной мобильной связи – для покрытия определенной территории использовался один мощный передатчик, а для регистрации сигнала от абонентских устройств – сеть приемников. Вызов в MTS-сети осуществлялся в ручном режиме – сначала абонент выбирал свободный канал, а затем устанавливал связь с оператором, соединявшим его с нужным абонентом. Причем изначально МТS-сеть работала в полудуплексном режиме, что позволяло решить проблему эха. Полнодуплексный режим (т.е. как в обычном телефоне) и автоматический выбор каналов появились лишь в 1964 г. Кстати, уже к концу 40-х гг. прошлого века AT&T и Bell Telephone Laboratories, не были самым передовыми – в 1948 г. Радиотелефонной компанией Ричмонда (штат Индиана) была запущена полностью автоматическая система подвижной радиосвязи, в которой вызов абонента осуществлялся без помощи оператора.
Один из первых автомобильных радиотелефонов
Все первые системы подвижной радиосвязи тех лет имели серьезное ограничение в виде частотного ресурса с ограниченным числом каналов. Это мешало обеспечить полное покрытие значительной территории и не позволяло двум сетям работать в одном частотном диапазоне – минимальное расстояние между двумя радиосистемами должно было составлять не менее 100 км. Решение данной проблемы было найдено сотрудником Bell Laboratories Д. Рингом, предложившим всю зону покрытия разделить на ячейки (соты), образуемые базовыми станциями, работающими в различающихся частотных диапазонах. И именно сотовый принцип стал основополагающим для современных мобильных сетей. Практическая реализация идеи появилась в 1969 г. в поездах Metroliner, курсировавших между Нью-Йорком и Вашингтоном – весь маршрут поезда (255 миль) был разделен на девять зон, в каждой из которых было доступно по шесть каналов на частоте 450 МГц, а центр управления системой находился в Филадельфии.
Схематичное изображение сотовой сети
Одновременно с США системы подвижной радиосвязи развивались и в Европе, где основные работы вели компании «Эрикссон» и «Маркони». Первые испытания европейских систем радиосвязи состоялись в 1951 г., а японских – в 1967 г. Кстати, именно японцы установили, что в условиях городской застройки для подвижной радиосвязи больше всего подходят диапазоны в районе 400 и 900 МГц. Среди европейских стран первая коммерчески успешная сеть сотовой связи была развернута в Финляндии в 1971 г., а к 1978 г. ей была покрыта вся территория страны. Естественно, речь шла об автомобильной радиосвязи, что даже нашло отражение в ее названии – Autoradiopuhelin (ARP, «Автомобильный радиотелефон»). Аналогично позиционировалась и сеть Autotel. Однако несмотря на аналоговую передачу голоса, в стандарте Autotel вся служебная информация, в отличие от других систем подвижной радиосвязи тех лет, передавалась уже в цифровой форме.
Велись разработки в области подвижной радиосвязи и в нашей стране, но о них будет рассказано немного ниже, а пока вернемся в США, где развернулась яростная борьба между компаниями AT&T Bell Labs и Motorola, стремившимися стать лидерами на зарождающемся рынке мобильной связи. Причем AT&T Bell Labs делала ставку на автомобильную радиосвязь, а Motorola – на компактные устройства, которые можно было носить с собой. Конкуренция была достаточно жесткой, предпринимались даже попытки задействовать административный ресурс в лице FCC (Федеральной комиссии по коммуникациям). Победителем в борьбе вышла Motorola, а главным направлением дальнейшего развития мобильной связи стало создание компактных устройств, которые можно было просто носить с собой. Коммерческая сеть, основанная на предложенных Motorola принципах, была запущена в 1983 г., через десятилетие после того исторического звонка.
Первый мобильный телефон Motorola DynaTAC 8000X
(Dynamic adaptive total area coverage)
Если обсуждать стандарты сотовой связи тех лет, то следует напомнить, что в Америке начинал набирать популярность аналоговый стандарт AMPS (Advanced mobile phones service — усовершенствованная подвижная телефонная служба), впоследствии усовершенствованный до цифрового D-AMPS. В Европе появилась целая россыпь различных несовместимых между собой стандартов, а наибольшее распространение получили скандинавский NMT (Nordic mobile telephony) и развернутый в ряде европейских стран TACS (Total access communications system, аналог AMPS). В Японии наиболее популярными стали NTT (Nippon telephone and telegraph system) и модифицированный вариант TACS, получивший имя JTACS(NTACS). Все перечисленные стандарты, как и AMPS, были аналоговыми, а построенные сети относились к первому поколению мобильной связи.
Одновременно с ростом количества абонентов мобильных сетей перед европейцами встал вопрос создания единого стандарта мобильной связи, для чего в 1982 г. была создана группа Groupe Spécial Mobile, включавшая 26 европейских телефонных компаний. На разработку одноименного стандарта ушло девять лет – его первая спецификация была опубликована в 1991 г., а первая в мире коммерческая GSM-сеть была запущена в 1992 г. в Финляндии. Альтернативой GSM стал стандарт CDMA, распространенный в США и странах Азии. Первая коммерческая CDMA-сеть появилась в 1995 г. в Гонконге, а первая спутниковая система связи коммерческого назначения (основанная на технологии CDMA Omni TRACKC) была запущена в 1980 г. Кстати, теоретические основы CDMA заложил еще в 1935 г. русский ученый Д. В. Агеев.
Наша история
Сотовая связь в современном понимании пришла в нашу страну в 1991 г., когда компания «Дельта Телеком» развернула сеть стандарта NMT-450i, а первый звонок с ее использованием состоялся 9 сентября 1991 г. Первая российская GSM-сеть была запущена в 1994 г., одновременно с появлением оператора «Северо-Западный GSM».
Однако история развития мобильной связи в нашей стране имеет более глубокие корни. Все началось с того, что во время Великой Отечественной войны советский ученый Георгий Ильич Бабат предложил идею устройства под названием «монофон», представлявшего собой переносной телефонный аппарат, работающий полностью в автоматическом режиме. Рабочий диапазон частот устройства должен был находиться в районе 1—2 ГГц, но в отличие от современных средств сотовой связи, в «монофоне» для передачи голоса планировалось использовать не радиоканал, а разветвленную сеть волноводов.
Г.И. Бабат, изобретатель «монофона»
Следующий шаг к отечественной мобильной связи был сделан Г. Шапиро и И. Захарченко, предложившими в 1946 г. систему автомобильной радиотелефонной связи. Ее принцип был прост и гениален – городские телефонные станции предполагалось дополнить радиоприемной аппаратурой, а каждому автомобилю, оснащенному радиосвязью, – выделить индивидуальные позывные. Для совершения вызова достаточно было передать в эфир свои позывные, после чего автоматически включался установленный в автомобиле телефон, пользоваться которым можно было, как обыкновенным телефонным аппаратом. При поступлении на номер мобильного абонента входящего звонка установка связи с ним осуществлялась также посредством позывных. На первых порах даже радиус действия системы Шапиро — Захарченко составлял примерно 20 км, но впоследствии изобретатели смогли увеличить его до 150 км, причем сам прибор был весьма компактным. Изначально систему Шапиро — Захарченко предполагалось использовать для координации работы милиции, пожарных, медиков и других экстренных служб. Однако идея не прижилась в первую очередь из-за нежелания этих служб быть привязанными к городской телефонной сети.
Но действительно сенсационным можно считать то, что в 1957 г. Л. И. Куприянович создал прототип мобильного телефона, получившего имя ЛК-1. Что интересно, до разработки ЛК-1 сферой деятельности Куприяновича было создание портативных раций, так же как и у его заокеанского коллеги Мартина Купера. Сопряжение ЛК-1 с городской телефонной сетью осуществлялось через «Автоматическую телефонную радиостанцию» (АТР), с которой «мобильная» трубка была связана четырьмя частотными каналами: прием звука, передача звука, передача сигналов набора номера и отправка сигнала завершения вызова. Причем был продуман и вопрос массового использования ЛК-1 – в этом случае управляющие сигналы различались по тональности, а для передачи голоса использовались разные частотные каналы. Радиус действия аппарата составлял несколько десятков километров.
Заметка в журнале «Наука и жизнь», №10, 1958 г..
Обратите внимание – в СССР изначально ставка делалась именно на создание систем подвижной радиосвязи, использование которых максимально похоже на использование обычных городских телефонов, причем эти системы должны были максимально просто интегрироваться с действующей городской телефонной сетью. Также понималась и важность компактных размеров – если первые варианты ЛК-1 весили около 3 кг (напомню, вес автомобильных радиотелефонов составлял 10—20 кг.), то уже в 1958 г. Куприяновичу удалось изготовить телефон весом всего 500 гр. А в 1959 г. он выдвинул предложение установить АТР на высотном задании, т.е. реализовать то же самое, что сделал Мартин Купер спустя 14 лет. Но изобретение Л.И. Куприяновича хода не получило, и к 1960—1961 гг. в своих статьях он рассказывает о портативных рациях и новостях электроники, но ни словом не упоминает о радиотелефоне.
И это не случайно — в конце 50-х гг. прошлого века по заказу высшего руководства страны в СССР началась разработка системы подвижной автоматической радиосвязи «Алтай». Причем одно из главных требований состояло в том, чтобы ее использование было максимально схоже с применением обычной телефонной сети, т.е. ручное переключение каналов и необходимость вызова диспетчера были исключены. И эта задача была решена – уже в 1963 г. система была запущена в опытную эксплуатацию на территории Москвы. Рабочий диапазон «Алтая» находился в районе 150 МГц, а позднее был задействован и диапазон 330 МГц. К середине 70-х под покрытием этой системы оказались уже 114 городов СССР, а на Олимпиаде 1980 г. она стала основным средством для связи освещавших ее журналистов. Причем качество связи на «Алтае» было не хуже, чем на лучших проводных телефонных линиях, а проблемы со связью возникали достаточно редко. В эпоху своего расцвета она стала доступна не только партийным и государственным деятелям, но и руководителям предприятий – к началу 80-х гг. ею пользовались около 25 тыс. абонентов. Для высшего руководства страны и нужд спецслужб также была создана «Роса», представлявшая собой вариант «Алтая», дополненный средствами шифрования.
Абонентское оборудование «Алтай» образца 1960-х гг
Были у СССР и планы по развертыванию сети мобильной связи, доступной для рядового человека. В начале 1980-х годов была начата работа над системой «ВоЛеМоТ», название которой состояло из первых букв городов, где велась ее разработка: Воронеж, Ленинград, Молодечно, Тернополь. Причем в систему изначально закладывались возможность использования множества базовых станций с целью покрыть всю территорию страны и поддержка автоматического перехода между базовыми станциями без прерывания разговора. Таким образом, «ВоЛеМоТ» могла стать полноценной сотовой сетью, и если бы не бюрократические проволочки и недостаточное финансирование работ, то ее запустили бы уже к середине 1980-х гг. В качестве рабочего диапазона в ней планировалось использовать частоту 330 МГц, что давало возможность покрыть одной базовой станцией большие расстояния. Кстати, запуск системы в эксплуатацию в некоторых городах все же состоялся, но произошло это лишь в середине 1990-х гг., когда технологическое лидерство было упущено, а на рынке доминировали NMT- и GSM-сети.
Резюме
История не имеет сослагательного наклонения. Мы упустили возможность стать лидерам в деле строительства мобильных сетей, а ведь шансы для этого у нашей страны были. В 1959 г. болгарский ученый Христо Бачваров создал мобильный телефон, концептуально схожий с аппаратом Л.И. Куприяновича, и получил соответствующий патент. Более того, на выставке «Интероргтехника-66» засветились РАТ-0,5 и АТРТ-0,5, компактные мобильные телефоны промышленного производства, а также базовая станция РАТЦ-10, способная одновременно связать шесть мобильных абонентов с городской телефонной сетью. Но в серию все эти наработки так и не пошли, а днем рождения мобильной связи все признали 3 апреля 1973 г., когда Мартин Купер совершил свой исторический звонок.
1)Принцип работы сотовой связи
Лекция: Принципы работы сотовой связи. Стандарты, способы реализации и перспективы развития. (4 часа)
Содержание:
2) История стандартов сотовой связи
3) Стандарты сотовой связи. Перспективы
4) Топология сотовой сети на примере сети GSM
1)Принцип работы сотовой связи
Основой любой сотовой сети является сота (ячейка), в центре которой находится базовая станция. При вызове, поступающем с базовой станции, сотовый телефон соединяется с ней и ему выделяется специальный радиоканал для разговора. Размеры соты определяются максимальной дальностью связи телефона с базовой станцией, типом сети, мощностью базовой станции и рядом других факторов. Максимальная дальность — радиус соты — может быть в диапазоне от нескольких метров до многих десятков километров.
Телефон и мобильный пользователь, еще не выйдя из зоны действия одной базовой станции, попадают в зону действия следующей, и так далее, пока совсем не выйдет из зоны обслуживания сети. Сотовая связь совсем не обязательно должна быть мобильной: на сегодня большое распространение получает так называемая стационарная сотовая связь. Такое решение часто является удобным и экономически выгодным — отпадает необходимость в дорогостоящей прокладке телефонного кабеля, а одной мощной базовой станции вполне достаточно для телефонизации целого микрорайона. На предприятиях и больших фирмах возможна установка собственных микросотовых систем связи стандарта DECT.
1.1)Аналоговая сотовая связь
На начальном этапе развития сотовой связи важнее всего было обеспечить максимально возможную зону охвата при минимально возможном количестве базовых станций. С этой задачей успешно справлялись аналоговые системы стандарта AMPS (Advanced Mobile Phone Service), в которых каждому абоненту на время разговора выделялась определенная полоса частот. При небольшом числе абонентов это было удобно и не вызывало проблем-свободных частот хватало на всех, а базовые аналоговые станции диапазона 450 Мгц можно было размещать на значительном (порядка десятков километров) расстоянии друг от друга. Первые сотовые телефоны весили чуть ли не пять килограммов, имели форму небольшого чемодана и стоили достаточно дорого. Однако по мере роста числа абонентов начали возникать проблемы перегрузки сети, так как ограниченного диапазона частот перестало хватать на всех абонентов, которые желали поговорить одновременно в зоне действия одной и той же базовой станции.
Основой любой сотовой системы связи является обеспечение «многостанционного (множественного) доступа». Это означает, что система поддерживает одновременную работу в сети нескольких абонентов. В аналоговой же сотовой связи значительное количество абонентов делят между собой ограниченное число радиоканалов. Канал в этой ситуации может рассматриваться как часть ограниченного частотного диапазона, выделенного для данной сети связи.
1.2)Цифровая сотовая связь.
Интенсивный процесс развития новых технологий мобильной телефонной связи можно считать начавшимся с того момента, когда рост числа абонентов сотовых сетей связи натолкнулся на нехватку частот. Несмотря на значительные усовершенствования, стандарт аналоговой передачи AMPS начал отживать свое, и начался переход на цифровые стандарты, позволяющие при прочих равных условиях кардинально увеличить количество одновременно разговаривающих абонентов.
В разных системах сотовой связи используются разные технологии множественного доступа. Традиционные аналоговые системы сотовой связи, основанные на стандартах AMPS и TACS (Total Access Communications System), используют технологию частотного разделения каналов. AMPS — системы делят частотный диапазон на участки шириной 30 КГц на канал, узкополосные системы AMPS (NAMPS) используют участки спектра 10 КГц на каждый канал, а системы TACS — участки спектра 25 КГц.
Новую струю в развитие сотовой связи внесли цифровые системы. Самой распространенной является технологией многостанционного доступа с временным разделением каналов (TDMA — Time Division Multiple Access). В группу стандартов цифровой связи TDMA входят Североамериканская цифровая сотовая связь (известная также под именем используемого ею стандарта IS-54/IS-136), Глобальная система мобильной связи (GSM — Global System for Mobile Communications) и Персональная цифровая сотовая связь (PDC — Personal Digital Cellular). Стандарт IS-54, в настоящее время IS-136, разработан в 1990 году для стран Северной Америки. Эта технология, которую еще называют D-AMPS (Цифровая AMPS), разрабатывалась для увеличения емкости существующих аналоговых сетей AMPS, для которых уже не хватало частот в условиях быстрого роста числа абонентов. Одновременно в США начала быстрыми темпами развиваться еще одна система сотовой цифровой связи, основанная на технологии кодового разделения каналов — стандарт CDMA.
Принцип сотовой связи
Как функционирует сотовая связь? Почему называется «сотовая»? Все эти вопросы изучаются в университете! Но скучно будет просто цитировать учебник… Я расскажу вам обо всём в более простом и понятном варианте. Итак, ПРИНЦИП СОТОВОЙ СВЯЗИ.
С развитием систем радиотелефонной связи возникла проблема — ограниченность частотного ресурса. То есть телефоны с близкими рабочими каналами начали создавать помехи друг другу.
По данным на май 2011 года число абонентов сотовой связи в России составляло 222 миллиона человек. Эта цифра подразумевает число рабочих СИМ-карт. В полтора раза больше населения нашей страны!!!
В середине 1940-х годов исследовательский центр Bell Laboratories предложил идею разбиения всей обслуживаемой территории на небольшие участки, которые стали называться сотами (от английского CELL – ячейка, сота). Каждая сота должна обслуживаться передатчиком с ограниченным радиусом действия и фиксированной частотой. Это позволило бы без взаимных помех использовать ту же самую частоту повторно в другой соте.
Чтобы была понятна суть этого абзаца, приведу рисунок.
Для оптимального разделения территории на соты могут быть использованы только три геометрических фигуры: треугольник, квадрат и шестиугольник. Наиболее подходящей фигурой является шестиугольник, так как если антенну с круговой диаграммой направленности установить в его центре, то будет обеспечен доступ почти ко всей соте.
Каждая ячейка, которой выделяется своя полоса частот, обслуживается базовой станцией, состоящей из передатчика, приёмника и модуля управления. Смежные ячейки используют разные частоты, чтобы избежать интерференции или перекрёстных помех. В то же время ячейки, находящиеся на большом расстоянии друг от друга, могут использовать одинаковые полосы частот. Таким образом, основной принцип сотовой связи — это повторное использование частот в несмежных сотах.
Когда телефон находится в режиме ожидания, его приёмное устройство постоянно сканирует все каналы системы. Для вызова абонента всеми базовыми станциями передаётся сигнал вызова. Сотовый телефон вызываемого абонента при получении этого сигнала отвечает по свободному каналу управления. Базовые станции, принявшие ответный сигнал, передают информацию о его параметрах в центр коммутации, который, в свою очередь, переключает разговор на ту базовую станцию, где зафиксирован максимальный уровень сигнала сотового радиотелефона вызываемого абонента.
Во время набора номера сотовый телефон занимает один из свободных каналов, уровень сигнала базовой станции в котором в данный момент максимален. По мере удаления абонента от базовой станции или в связи с ухудшением условий распространения радиоволн уровень сигнала уменьшается, что ведёт к ухудшению качества связи. Улучшение качества связи достигается путём автоматического переключения абонента на другой канал связи.
Узнайте, что такое чувствительность телефона и как она влияет на качество связи.
Как вам статья?
ПомоглаНе помоглаЗадайте вопрос специалисту в комментариях
Нужна консультация
Общие принципы клеточной коммуникации — молекулярная биология клетки
Механизмы, позволяющие одной клетке влиять на поведение другой, почти наверняка существовали в мире одноклеточных организмов задолго до появления многоклеточных организмов на Земле. Доказательства получены из исследований современных одноклеточных эукариот, таких как дрожжи. Хотя эти клетки обычно ведут независимый образ жизни, они могут общаться и влиять на поведение друг друга при подготовке к половому спариванию.В почкующихся дрожжах Saccharomyces cerevisiae, , например, когда гаплоидная особь готова к спариванию, она секретирует пептид , фактор спаривания , который сигнализирует клеткам противоположного типа спаривания о прекращении пролиферации и подготовке к спариванию (). Последующее слияние двух гаплоидных клеток противоположных типов спаривания дает диплоидную клетку, которая затем может подвергаться мейозу и споруляции, генерируя гаплоидные клетки с новым набором генов.
Рисунок 15-2
Клетки почкующихся дрожжей реагируют на фактор спаривания.(A) Клетки обычно имеют сферическую форму. (B) В ответ на фактор спаривания, секретируемый соседними клетками дрожжей, они выступают в направлении источника фактора при подготовке к спариванию. (Любезно предоставлено (подробнее …)
Исследования дрожжевых мутантов, которые не могут спариваться, идентифицировали многие белки, которые необходимы в процессе передачи сигналов. Эти белки образуют сигнальную сеть, которая включает в себя белки рецепторов клеточной поверхности, GTP-связывающие белки, и протеинкиназы, каждая из которых имеет близких родственников среди белков, передающих сигналы в клетках животных.Однако из-за дупликации и дивергенции генов сигнальные системы у животных стали намного более сложными, чем у дрожжей.
Внеклеточные сигнальные молекулы связываются со специфическими рецепторами
Дрожжевые клетки связываются друг с другом для спаривания, секретируя несколько видов небольших пептидов. Напротив, клетки высших животных общаются с помощью сотен видов сигнальных молекул. К ним относятся белки, небольшие пептиды, аминокислоты, нуклеотиды, стероиды, ретиноиды, производные жирных кислот и даже растворенные газы, такие как оксид азота и оксид углерода.Большинство этих сигнальных молекул секретируются из сигнальной клетки во внеклеточное пространство посредством экзоцитоза (обсуждается в главе 13). Другие высвобождаются путем диффузии через плазматическую мембрану, а некоторые попадают во внеклеточное пространство, оставаясь при этом прочно связанными с поверхностью сигнальной клетки.
Независимо от природы сигнала, клетка-мишень отвечает посредством специфического белка, называемого рецептором, который специфически связывает сигнальную молекулу и затем инициирует ответ в клетке-мишени.Внеклеточные сигнальные молекулы часто действуют при очень низких концентрациях (обычно ≤ 10 -8 M), и рецепторы, которые их распознают, обычно связывают их с высоким сродством (константа сродства K α ≥ 10 8 литров / моль ; видеть ). В большинстве случаев эти рецепторы представляют собой трансмембранные белки на поверхности клетки-мишени. Когда они связываются с внеклеточной сигнальной молекулой (лиганд), , они активируются и генерируют каскад внутриклеточных сигналов, которые изменяют поведение клетки.В других случаях рецепторы находятся внутри клетки-мишени, и сигнальная молекула должна войти в клетку, чтобы активировать их: для этого требуется, чтобы сигнальные молекулы были достаточно маленькими и гидрофобными, чтобы диффундировать через плазматическую мембрану ().
Рисунок 15-3
Связывание внеклеточных сигнальных молекул либо с рецепторами на поверхности клетки, либо с внутриклеточными рецепторами. Большинство сигнальных молекул гидрофильны и поэтому не могут напрямую пересекать плазматическую мембрану; вместо этого они связываются с рецепторами клеточной поверхности (подробнее…)
Внеклеточные сигнальные молекулы могут действовать как на коротких, так и на больших расстояниях
Многие сигнальные молекулы остаются связанными с поверхностью сигнальной клетки и влияют только на клетки, которые с ней контактируют (). Такая контактно-зависимая передача сигналов особенно важна во время развития и при иммунных ответах. Однако в большинстве случаев сигнальные молекулы секретируются. Секретируемые молекулы могут уноситься далеко, чтобы воздействовать на удаленные мишени, или они могут действовать как локальные медиаторы, воздействуя только на клетки в непосредственном окружении сигнальной клетки.Этот последний процесс называется паракринной передачей сигналов (). Для того чтобы паракринные сигналы доставлялись только к их собственным клеткам-мишеням, секретируемым молекулам нельзя позволять диффундировать слишком далеко; по этой причине они часто быстро захватываются соседними клетками-мишенями, разрушаются внеклеточными ферментами или иммобилизуются внеклеточным матриксом.
Рисунок 15-4
Формы межклеточной сигнализации. (A) Контакт-зависимая передача сигналов требует, чтобы клетки находились в прямом мембранно-мембранном контакте.(B) Паракринная передача сигналов зависит от сигналов, которые высвобождаются во внеклеточное пространство и действуют локально на соседние клетки. (подробнее …)
Для большого, сложного многоклеточного организма передачи сигналов ближнего действия недостаточно для координации поведения его клеток. У этих организмов развивались наборы специализированных клеток, играющих особую роль в связи между широко отдельными частями тела. Самыми сложными из них являются нервные клетки или нейроны, которые обычно расширяют длинные отростки (аксоны), которые позволяют им контактировать с клетками-мишенями на большом расстоянии.При активации сигналами из окружающей среды или других нервных клеток нейрон быстро посылает электрические импульсы (потенциалы действия) вдоль своего аксона; когда такой импульс достигает конца аксона, он заставляет расположенные там нервные окончания выделять химический сигнал, называемый нейротрансмиттером. Эти сигналы секретируются в специализированных клеточных соединениях, называемых химическими синапсами , , которые предназначены для обеспечения доставки нейромедиатора специфически к постсинаптической клетке-мишени ().Детали этого синаптического сигнального процесса обсуждаются в главе 11.
Второй тип специализированной сигнальной клетки, которая контролирует поведение организма в целом, — это эндокринная клетка. Эти клетки выделяют в кровоток свои сигнальные молекулы, называемые гормонами, которые передают сигнал клеткам-мишеням, широко распространенным по всему телу ().
Сравниваются механизмы, позволяющие эндокринным и нервным клеткам координировать клеточное поведение животных.Поскольку эндокринная передача сигналов зависит от диффузии и кровотока, она является относительно медленной. Синаптическая передача сигналов, напротив, может быть намного быстрее и точнее. Нервные клетки могут передавать информацию на большие расстояния с помощью электрических импульсов, которые распространяются со скоростью до 100 метров в секунду; После выхода из нервного окончания нейротрансмиттер должен диффундировать к клетке-мишени на расстояние менее 100 нм, а этот процесс занимает менее миллисекунды. Еще одно различие между эндокринной и синаптической передачей сигналов заключается в том, что, хотя гормоны сильно разбавлены в кровотоке и интерстициальной жидкости и, следовательно, должны действовать при очень низких концентрациях (обычно <10 -8 M), нейротрансмиттеры разбавлены гораздо меньше и могут добиться высоких местных концентраций.Концентрация ацетилхолина в синаптической щели активного нервно-мышечного соединения, например, составляет около 5 × 10 -4 М. Соответственно, рецепторы нейротрансмиттеров имеют относительно низкое сродство к своему лиганду, что означает, что нейромедиатор может диссоциировать. быстро от рецептора, чтобы прекратить ответ. Более того, после выхода из нервного окончания нейротрансмиттер быстро удаляется из синаптической щели либо специфическими гидролитическими ферментами, которые его разрушают, либо специфическими мембранными транспортными белками, которые перекачивают его обратно либо в нервное окончание, либо в соседние глиальные клетки.Таким образом, синаптическая передача сигналов намного точнее, чем эндокринная передача сигналов как во времени, так и в пространстве.
Рисунок 15-5
Контраст между эндокринной и синаптической передачей сигналов. У сложных животных эндокринные клетки и нервные клетки работают вместе, чтобы координировать разнообразную деятельность миллиардов клеток. В то время как разные эндокринные клетки должны использовать разные гормоны для связи (подробнее …)
Скорость ответа на внеклеточный сигнал зависит не только от механизма доставки сигнала, но и от природы ответа в клетке-мишени.Если для ответа требуются только изменения в белках, уже присутствующих в клетке, это может произойти за секунды или даже миллисекунды. Однако, когда ответ включает изменения в экспрессии генов и синтез новых белков, обычно требуются часы, независимо от способа доставки сигнала.
Автокринная сигнализация может координировать решения групп идентичных ячеек
Все формы сигнализации, обсужденные до сих пор, позволяют одной клетке влиять на другую. Часто сигнальная клетка и мишень относятся к разным типам клеток.Однако клетки могут также посылать сигналы другим клеткам того же типа, а также себе. При такой аутокринной передаче сигналов клетка выделяет сигнальные молекулы, которые могут обратно связываться с ее собственными рецепторами. Во время развития, например, как только клетка была направлена по определенному пути дифференцировки, она может начать секретировать аутокринные сигналы себе, которые подкрепляют это решение, связанное с развитием.
Аутокринная передача сигналов наиболее эффективна, когда осуществляется одновременно соседними клетками одного и того же типа, и она, вероятно, будет использоваться для поощрения групп идентичных клеток к принятию одинаковых решений в отношении развития.Таким образом, аутокринная передача сигналов считается одним из возможных механизмов, лежащих в основе «эффекта сообщества», который наблюдается на раннем этапе развития, во время которого группа идентичных клеток может реагировать на сигнал, индуцирующий дифференцировку, но одна изолированная клетка того же типа не может ( ).
Рисунок 15-6
Автокринная сигнализация. Группа идентичных клеток производит более высокую концентрацию секретируемого сигнала, чем одна клетка. Когда этот сигнал связывается обратно с рецептором того же типа клеток, он побуждает клетки координированно реагировать как группа.(подробнее …)
К сожалению, раковые клетки часто используют аутокринную передачу сигналов для преодоления нормального контроля над пролиферацией и выживанием клеток, которые мы обсудим позже. Секретируя сигналы, которые воздействуют на собственные рецепторы клетки, раковые клетки могут стимулировать свое собственное выживание и пролиферацию и, таким образом, выживать и размножаться в местах, где нормальные клетки того же типа не могут. Как возникает это опасное нарушение нормального поведения ячеек, обсуждается в главе 23.
Щелевые соединения позволяют передавать сигнальную информацию соседним ячейкам
Другой способ координировать действия соседних ячеек — это щелевые соединения.Это специализированные межклеточные соединения, которые могут образовываться между близко расположенными плазматическими мембранами и напрямую соединять цитоплазмы соединенных клеток через узкие заполненные водой каналы (см.). Каналы позволяют обмениваться небольшими внутриклеточными сигнальными молекулами (внутриклеточные медиаторы), , такими как Ca 2+, и циклический АМФ (обсуждается позже), но не макромолекулами, такими как белки или нуклеиновые кислоты. Таким образом, клетки, соединенные щелевыми контактами, могут общаться друг с другом напрямую, без необходимости преодолевать барьер, создаваемый промежуточными плазматическими мембранами ().
Рисунок 15-7
Сигнализация через щелевые соединения. Клетки, соединенные щелевыми контактами, имеют общие небольшие молекулы, в том числе небольшие внутриклеточные сигнальные молекулы, и поэтому могут реагировать на внеклеточные сигналы скоординированным образом.
Как обсуждалось в главе 19, структура соединений щелевых соединений в ткани может быть обнаружена либо электрически, с помощью внутриклеточных электродов, либо визуально после микроинъекции небольших водорастворимых красителей. Исследования такого рода показывают, что клетки в развивающемся эмбрионе создают и разрывают соединения щелевых соединений в определенных и интересных паттернах, что убедительно указывает на то, что эти соединения играют важную роль в процессах передачи сигналов, которые происходят между этими клетками.Например, мыши и люди, у которых отсутствует один конкретный белок щелевых соединений (коннексин 43), имеют серьезные дефекты в развитии сердца. Подобно аутокринной передаче сигналов, описанной выше, коммуникация по щелевому соединению помогает соседним клеткам схожего типа координировать свое поведение. Однако до сих пор неизвестно, какие именно небольшие молекулы важны как переносчики сигналов через щелевые соединения, а конкретные функции связи щелевых соединений в развитии животных остаются неопределенными.
Каждая клетка запрограммирована так, чтобы реагировать на определенные комбинации внеклеточных сигнальных молекул.
Типичная клетка многоклеточного организма подвергается воздействию сотен различных сигналов в окружающей среде. Эти сигналы могут быть растворимыми, связаны с внеклеточным матриксом или с поверхностью соседней клетки, и они могут действовать во многих миллионах комбинаций. Клетка должна избирательно реагировать на этот поток сигналов в соответствии со своим собственным специфическим характером, который она приобрела в результате прогрессивной клеточной специализации в ходе развития.Клетка может быть запрограммирована так, чтобы реагировать на одну комбинацию сигналов путем дифференцирования, на другую комбинацию путем умножения и на еще одну посредством выполнения некоторой специализированной функции, такой как сокращение или секреция.
Большинство клеток сложного животного также запрограммированы на то, чтобы зависеть от определенной комбинации сигналов, просто чтобы выжить. Когда клетка лишена этих сигналов (например, в чашке для культивирования), клетка активирует программу самоубийства и убивает себя — процесс, называемый запрограммированной смертью клеток , или апоптоз ().Поскольку разные типы клеток требуют разных комбинаций сигналов выживания, каждый тип клеток ограничен различными средами в организме. Способность подвергаться апоптозу — фундаментальное свойство клеток животных, и оно обсуждается в главе 17.
Рисунок 15-8
Зависимость животной клетки от множественных внеклеточных сигналов. Каждый тип клеток отображает набор рецепторов, которые позволяют ему реагировать на соответствующий набор сигнальных молекул, производимых другими клетками.Эти сигнальные молекулы работают в комбинациях для регулирования (подробнее …)
В принципе, сотни сигнальных молекул, производимых животными, можно использовать для создания почти неограниченного числа сигнальных комбинаций. Использование этих комбинаций для управления поведением клеток позволяет животному управлять своими клетками весьма специфическими способами, используя ограниченное разнообразие сигнальных молекул.
Различные клетки могут по-разному реагировать на одну и ту же внеклеточную сигнальную молекулу
Конкретный способ, которым клетка реагирует на окружающую среду, варьируется.Он варьируется в зависимости от набора рецепторных белков, которыми обладает клетка, который определяет конкретное подмножество сигналов, на которые она может реагировать, и варьируется в зависимости от внутриклеточного механизма, с помощью которого клетка интегрирует и интерпретирует сигналы, которые она получает (см.). Таким образом, одна сигнальная молекула часто по-разному воздействует на разные клетки-мишени. Например, нейромедиатор ацетилхолин стимулирует сокращение клеток скелетных мышц, но снижает скорость и силу сокращения клеток сердечной мышцы.Это связано с тем, что белки рецепторов ацетилхолина на клетках скелетных мышц отличаются от белков на клетках сердечной мышцы. Но различия рецепторов не всегда являются объяснением различных эффектов. Во многих случаях одна и та же сигнальная молекула связывается с идентичными рецепторными белками, но вызывает очень разные ответы в разных типах клеток-мишеней, отражая различия во внутреннем механизме, с которым связаны рецепторы ().
Рисунок 15-9
Различные ответы, индуцированные нейромедиатором ацетилхолином.Различные типы клеток по-разному реагируют на ацетилхолин. (A и B) Для этих двух типов клеток ацетилхолин связывается с аналогичными рецепторными белками, но внутриклеточные (подробнее …)
Концентрация молекулы может быть скорректирована быстро, только если время жизни молекулы короткое
It Естественно думать о сигнальных системах с точки зрения изменений, производимых при доставке сигнала. Но не менее важно учитывать, что происходит, когда сигнал отменяется.Во время развития временные сигналы часто вызывают длительные эффекты: они могут вызвать изменение в развитии клетки, которое сохраняется бесконечно, благодаря механизмам клеточной памяти, таким как те, которые обсуждались в главах 7 и 21. Однако в большинстве случаев во взрослых тканях реакция исчезает, когда сигнал прекращается. Эффект временный, потому что сигнал проявляет свои эффекты, изменяя набор молекул, которые нестабильны, претерпевая непрерывный оборот. Таким образом, как только сигнал отключен, замена старых молекул новыми стирает все следы его действия.Отсюда следует, что скорость, с которой клетка реагирует на удаление сигнала, зависит от скорости разрушения или оборота молекул, на которые воздействует сигнал.
Также верно, хотя и менее очевидно, что эта скорость оборота также определяет быстроту реакции при включении сигнала. Рассмотрим, например, две внутриклеточные сигнальные молекулы X и Y, каждая из которых обычно поддерживается в концентрации 1000 молекул на клетку. Молекула Y синтезируется и разлагается со скоростью 100 молекул в секунду, при этом каждая молекула имеет среднее время жизни 10 секунд.Скорость обновления молекулы X в 10 раз ниже, чем у молекулы Y: она синтезируется и разлагается со скоростью 10 молекул в секунду, так что каждая молекула имеет среднее время жизни в клетке 100 секунд. Если сигнал, воздействующий на клетку, увеличивает скорость синтеза как X, так и Y в десять раз без какого-либо изменения времени жизни молекул, в конце 1 секунды концентрация Y увеличится почти на 900 молекул на клетку (10 × 100 — 100), тогда как концентрация X увеличится всего на 90 молекул на клетку.Фактически, после того, как скорость синтеза молекулы резко увеличилась или уменьшилась, время, необходимое для того, чтобы молекула сместилась наполовину от ее старой до новой равновесной концентрации, равно ее нормальному периоду полураспада, то есть времени, которое потребовалось бы, чтобы его концентрация упала вдвое, если бы весь синтез был остановлен ().
Рисунок 15-10
Важность быстрой текучести. Графики показывают прогнозируемые относительные скорости изменения внутриклеточных концентраций молекул с различным временем оборота, когда скорость их синтеза либо (A) снижается, либо (B) внезапно увеличивается на (больше…)
Те же принципы применимы к белкам и маленьким молекулам, а также к молекулам во внеклеточном пространстве и внутри клеток. Многие внутриклеточные белки имеют короткий период полураспада, некоторые живут менее 10 минут. В большинстве случаев это белки, выполняющие ключевые регуляторные роли, концентрация которых в клетке быстро регулируется путем изменения скорости их синтеза. Аналогично, любые ковалентные модификации белков, которые происходят как часть процесса быстрой передачи сигналов — чаще всего, добавление фосфатной группы к боковой цепи аминокислоты — должны постоянно удаляться с высокой скоростью, чтобы сделать возможной быструю передачу сигналов.
Мы обсудим некоторые из этих молекулярных событий подробно позже для сигнальных путей, которые действуют через рецепторы на поверхности клетки. Но эти принципы применимы довольно широко, как показывает следующий пример.
Сигналы газа оксида азота путем прямого связывания с ферментом внутри клетки-мишени
Хотя большинство внеклеточных сигналов представляют собой гидрофильные молекулы, которые связываются с рецепторами на поверхности клетки-мишени, некоторые сигнальные молекулы достаточно гидрофобны и / или достаточно малы для прохождения легко через плазматическую мембрану клетки-мишени.Попав внутрь, они напрямую регулируют активность определенного внутриклеточного белка. Важным и замечательным примером является газ оксид азота (NO) , который действует как сигнальная молекула как у животных, так и у растений. У млекопитающих одна из его функций — регулировать сокращение гладких мышц. Например, ацетилхолин высвобождается вегетативными нервами в стенках кровеносного сосуда и заставляет гладкомышечные клетки в стенке сосуда расслабляться. Ацетилхолин действует опосредованно, побуждая близлежащие эндотелиальные клетки производить и высвобождать NO, который затем дает сигнал расслаблению нижележащих гладкомышечных клеток.Этот эффект NO на кровеносные сосуды объясняет механизм действия нитроглицерина, который уже около 100 лет используется для лечения пациентов со стенокардией (боль, возникающая из-за недостаточного притока крови к сердечной мышце). Нитроглицерин превращается в NO, который расслабляет кровеносные сосуды. Это снижает нагрузку на сердце и, как следствие, снижает потребность сердечной мышцы в кислороде.
Многие типы нервных клеток используют NO для передачи сигналов своим соседям.NO, выделяемый вегетативными нервами полового члена, например, вызывает локальное расширение кровеносных сосудов, которое отвечает за эрекцию полового члена. NO также вырабатывается в качестве местного медиатора активированными макрофагами и нейтрофилами, помогая им уничтожать вторгшиеся микроорганизмы. У растений NO участвует в защитных реакциях на травму или инфекцию.
Газ NO образуется в результате дезаминирования аминокислоты аргинина, катализируемого ферментом NO-синтазой. Поскольку растворенный NO легко проходит через мембраны, он быстро диффундирует из клетки, где он вырабатывается, в соседние клетки.Он действует только локально, потому что у него короткий период полураспада — около 5–10 секунд — во внеклеточном пространстве, прежде чем он преобразуется в нитраты и нитриты кислородом и водой. Во многих клетках-мишенях, включая эндотелиальные клетки, NO связывается с железом в активном центре фермента гуанилилциклазы, стимулируя этот фермент продуцировать небольшой внутриклеточный медиатор cyclic GMP, , который мы обсудим позже (). Эффект NO может проявиться в течение нескольких секунд, потому что нормальная скорость оборота циклического GMP высока: быстрое разложение до GMP фосфодиэстеразой постоянно уравновешивает производство циклического GMP из GTP гуанилилциклазой.Препарат Виагра ингибирует эту циклическую фосфодиэстеразу GMP в половом члене, тем самым увеличивая период времени, в течение которого уровни циклического GMP остаются повышенными после того, как продукция NO индуцируется местными нервными окончаниями. Циклический GMP, в свою очередь, поддерживает расслабление кровеносных сосудов и эрекцию полового члена.
Рисунок 15-11
Роль оксида азота (NO) в расслаблении гладких мышц стенки кровеносного сосуда. Ацетилхолин, высвобождаемый нервными окончаниями в стенке кровеносного сосуда, активирует NO-синтазу в эндотелиальных клетках, выстилающих кровеносный сосуд, заставляя эндотелиальные клетки производить (больше…)
Окись углерода (CO) — еще один газ, который используется в качестве межклеточного сигнала. Он может действовать так же, как NO, стимулируя гуанилилциклазу. Эти газы — не единственные сигнальные молекулы, которые могут проходить непосредственно через плазматическую мембрану клетки-мишени. Группа небольших гидрофобных негазообразных гормонов и местных медиаторов также проникает в клетки-мишени таким образом. Но вместо связывания с ферментами они связываются с белками внутриклеточных рецепторов, которые напрямую регулируют транскрипцию генов, как мы обсудим далее.
Ядерные рецепторы являются лиганд-активируемыми белками-регуляторами генов
Ряд небольших гидрофобных сигнальных молекул диффундируют непосредственно через плазматическую мембрану клеток-мишеней и связываются с внутриклеточными рецепторными белками. Эти сигнальные молекулы включают стероидных гормонов, гормонов щитовидной железы, ретиноиды, и витамина D. Хотя они сильно отличаются друг от друга как по химической структуре (), так и по функциям, все они действуют по схожему механизму. Когда эти сигнальные молекулы связываются со своими рецепторными белками, они активируют рецепторы, которые связываются с ДНК, чтобы регулировать транскрипцию определенных генов.Все рецепторы структурно родственны и являются частью надсемейства ядерных рецепторов. Это очень большое суперсемейство также включает некоторые рецепторные белки, которые активируются внутриклеточными метаболитами, а не секретируемыми сигнальными молекулами. Многие члены семьи были идентифицированы только путем секвенирования ДНК, и их лиганд еще не известен; эти белки поэтому упоминаются как орфанных ядерных рецептора. На важность таких ядерных рецепторов у некоторых животных указывает тот факт, что 1-2% генов у нематоды C.elegans , хотя у людей их меньше 50 (см.).
Рисунок 15-12
Некоторые сигнальные молекулы, которые связываются с ядерными рецепторами. Обратите внимание, что все они маленькие и гидрофобные. Показана активная гидроксилированная форма витамина D 3 . Эстрадиол и тестостерон — стероидные половые гормоны.
Стероидные гормоны, в том числе кортизол, стероидные половые гормоны, витамин D (у позвоночных) и гормон линьки (у насекомых), производятся из холестерина. Кортизол вырабатывается в коре надпочечников и влияет на метаболизм многих типов клеток. Стероидные половые гормоны вырабатываются в яичках и яичниках, и они отвечают за вторичные половые признаки, отличающие мужчин от женщин. Витамин D синтезируется в коже в ответ на солнечный свет; после преобразования в активную форму в печени или почках он регулирует метаболизм Ca 2+ , способствуя поглощению Ca 2+ в кишечнике и уменьшая его выведение почками.Гормоны щитовидной железы, которые состоят из аминокислоты тирозина, действуют, увеличивая скорость метаболизма в самых разных типах клеток, в то время как ретиноиды, такие как ретиноевая кислота, производятся из витамина А и играют важную роль в качестве местных медиаторов у позвоночных. разработка. Хотя все эти сигнальные молекулы относительно нерастворимы в воде, они становятся растворимыми для транспорта в кровотоке и других внеклеточных жидкостях за счет связывания со специфическими белками-носителями, от которых они диссоциируют перед попаданием в клетку-мишень (см.).
Помимо фундаментального различия в способах передачи сигналов своим клеткам-мишеням, большинство нерастворимых в воде сигнальных молекул отличаются от водорастворимых по продолжительности пребывания в кровотоке или тканевых жидкостях. Большинство водорастворимых гормонов удаляются и / или расщепляются в течение нескольких минут после попадания в кровь, а местные медиаторы и нейротрансмиттеры удаляются из внеклеточного пространства еще быстрее — за секунды или миллисекунды. Стероидные гормоны, напротив, сохраняются в крови часами, а гормоны щитовидной железы — днями.Следовательно, водорастворимые сигнальные молекулы обычно опосредуют ответы короткой продолжительности, тогда как нерастворимые в воде имеют тенденцию опосредовать ответы, которые более продолжительны.
Внутриклеточные рецепторы стероидных и тироидных гормонов, ретиноидов и витамина D связываются со специфическими последовательностями ДНК, соседними с генами, регулируемыми лигандом. Некоторые рецепторы, такие как рецепторы кортизола, расположены в основном в цитозоле и проникают в ядро после связывания лиганда; другие, такие как рецепторы щитовидной железы и ретиноидов, связаны с ДНК в ядре даже в отсутствие лиганда.В любом случае неактивные рецепторы связаны с ингибирующими белковыми комплексами, а связывание лиганда изменяет конформацию рецепторного белка, вызывая диссоциацию ингибиторного комплекса. Связывание лиганда также заставляет рецептор связываться с белками-коактиваторами, которые индуцируют транскрипцию генов (). Транскрипционный ответ обычно происходит последовательно: прямая активация небольшого числа специфических генов происходит в течение примерно 30 минут и составляет первичный ответ ; белковые продукты этих генов, в свою очередь, активируют другие гены, вызывая отложенный вторичный ответ ; и так далее.Таким образом, простой гормональный триггер может вызвать очень сложное изменение характера экспрессии генов ().
Рисунок 15-13
Надсемейство ядерных рецепторов. Все рецепторы ядерных гормонов связываются с ДНК либо как гомодимеры, либо как гетеродимеры, но для простоты мы показываем их здесь как мономеры. (A) Все рецепторы имеют родственную структуру. Короткий ДНК-связывающий домен в каждом рецепторе (подробнее …)
Рисунок 15-14
Ответы, вызванные активацией рецептора ядерного гормона.(A) ранний первичный ответ и (B) отложенный вторичный ответ. На рисунке показаны реакции на стероидный гормон, но те же принципы применимы ко всем лигандам, активирующим это семейство (подробнее …)
Ответы на стероидные гормоны и гормоны щитовидной железы, витамин D и ретиноиды, такие как ответы на внеклеточные сигналы в в целом, определяются как природой клетки-мишени, так и природой сигнальной молекулы. Многие типы клеток имеют идентичный внутриклеточный рецептор, но набор генов, регулируемых рецептором, различен для каждого типа клеток.Это связано с тем, что более одного типа регуляторного белка гена обычно должны связываться с эукариотическим геном, чтобы активировать его транскрипцию. Таким образом, внутриклеточный рецептор может активировать ген только в том случае, если существует правильная комбинация других белков, регулирующих ген, и многие из них специфичны для клеточного типа. Таким образом, каждый из этих гормонов вызывает у животного набор характерных реакций по двум причинам. Во-первых, рецепторы к нему есть только у определенных типов клеток. Во-вторых, каждый из этих типов клеток содержит различную комбинацию регулирующих белков генов, специфичных для других типов клеток, которые взаимодействуют с активированным рецептором, чтобы влиять на транскрипцию определенных наборов генов.
Молекулярные детали того, как ядерные рецепторы и другие регуляторные белки генов контролируют транскрипцию конкретных генов, обсуждаются в главе 7.
Три самых больших класса рецепторных белков клеточной поверхности связаны с ионными каналами, G-белками и Ферментно-связанные рецепторы
Как упоминалось ранее, все водорастворимые сигнальные молекулы (включая нейротрансмиттеры и все сигнальные белки) связываются со специфическими рецепторными белками на поверхности клеток-мишеней, на которые они влияют.Эти рецепторные белки клеточной поверхности действуют как преобразователи сигналов . Они преобразуют событие связывания внеклеточного лиганда во внутриклеточные сигналы, которые изменяют поведение клетки-мишени.
Большинство белков рецепторов клеточной поверхности принадлежат к одному из трех классов, определяемых механизмом трансдукции, который они используют. Рецепторы, связанные с ионным каналом , также известные как управляемых трансмиттером ионных каналов или ионотропных рецепторов, участвуют в быстрой синаптической передаче сигналов между электрически возбудимыми клетками ().Этот тип передачи сигналов опосредуется небольшим количеством нейротрансмиттеров, которые временно открывают или закрывают ионный канал, образованный белком, с которым они связываются, на короткое время изменяя ионную проницаемость плазматической мембраны и тем самым возбудимость постсинаптической клетки. Рецепторы, связанные с ионным каналом, принадлежат к большому семейству гомологичных многопроходных трансмембранных белков. Поскольку они подробно обсуждаются в главе 11, мы не будем их здесь рассматривать.
Рисунок 15-15
Три класса рецепторов клеточной поверхности.(A) рецепторы, связанные с ионным каналом, (B) рецепторы, связанные с G-белком, и (C) рецепторы, связанные с ферментом. Хотя многие ферментно-связанные рецепторы обладают внутренней ферментативной активностью, как показано слева, многие другие полагаются на (подробнее …)
Рецепторы, связанные с G-белком действуют опосредованно, регулируя активность отдельного целевого белка, связанного с плазматической мембраной, который может быть либо ферментом, либо ионным каналом. Взаимодействие между рецептором и этим белком-мишенью опосредуется третьим белком, называемым тримерным GTP-связывающим белком (G-белок) ().Активация целевого белка может изменить концентрацию одного или нескольких внутриклеточных медиаторов (если целевой белок является ферментом) или может изменить ионную проницаемость плазматической мембраны (если целевой белок является ионным каналом). Затронутые внутриклеточные медиаторы, в свою очередь, действуют, изменяя поведение других сигнальных белков в клетке. Все рецепторы, связанные с G-белком, принадлежат к большому семейству гомологичных трансмембранных белков с семью проходами.
Связанные с ферментом рецепторы, при активации либо действуют непосредственно как ферменты, либо напрямую связаны с ферментами, которые они активируют ().Они образованы однопроходными трансмембранными белками, у которых есть сайт связывания лиганда вне клетки и сайт связывания каталитического или фермента внутри. Связанные с ферментом рецепторы неоднородны по структуре по сравнению с двумя другими классами. Однако подавляющее большинство из них являются протеинкиназами или связаны с протеинкиназами, и связывание с ними лиганда вызывает фосфорилирование определенных наборов белков в клетке-мишени.
Есть некоторые рецепторы клеточной поверхности, которые не входят ни в один из вышеперечисленных классов.Некоторые из них зависят от внутриклеточных протеолитических событий, передающих сигнал клетке, и мы обсудим их только после того, как подробно объясним, как действуют рецепторы, связанные с G-белком, и рецепторы, связанные с ферментом. Мы начнем с некоторых общих принципов передачи сигналов через рецепторы на поверхности клетки.
Наиболее активированные рецепторы на поверхности клетки передают сигналы через небольшие молекулы и сеть внутриклеточных сигнальных белков
Сигналы, принимаемые на поверхности клетки рецепторами, связанными либо с G-белком, либо с ферментативными рецепторами, передаются внутрь клетки посредством комбинация малых и больших внутриклеточных сигнальных молекул. Результирующая цепочка внутриклеточных сигнальных событий в конечном итоге изменяет белков-мишеней, , и эти измененные белки-мишени ответственны за изменение поведения клетки (см.).
Маленькие внутриклеточные сигнальные молекулы называются малыми внутриклеточными посредниками, или вторыми мессенджерами («первые мессенджеры» являются внеклеточными сигналами). Они генерируются в большом количестве в ответ на активацию рецептора и быстро диффундируют от своего источника, передавая сигнал другим частям клетки.Некоторые, например циклический AMP и Ca 2+ , растворимы в воде и диффундируют в цитозоле, в то время как другие, такие как диацилглицерин, , растворимы в липидах и диффундируют в плоскости плазматической мембраны. В любом случае они передают сигнал, связываясь с выбранными сигнальными белками или белками-мишенями и изменяя их поведение.
Большие внутриклеточные сигнальные молекулы являются внутриклеточными сигнальными белками. Многие из них передают сигнал в клетку, либо активируя следующий сигнальный белок в цепи, либо генерируя небольшие внутриклеточные медиаторы.Эти белки можно классифицировать в соответствии с их конкретной функцией, хотя многие из них относятся к более чем одной категории ():
Рисунок 15-16
Различные виды внутриклеточных сигнальных белков на пути передачи сигналов от рецептора на поверхности клетки к ядру. В этом примере ряд сигнальных белков и небольших внутриклеточных медиаторов передают внеклеточный сигнал в клетку, (подробнее …)
- 1.
-
Релейные белки просто передают сообщение следующему сигнальному компоненту в цепи.
- 2.
-
Белки-мессенджеры переносят сигнал от одной части клетки к другой, например, от цитозоля к ядру.
- 3.
-
Адаптерные белки связывают один сигнальный белок с другим, не передавая сигнал сами.
- 4.
-
Белки-усилители, , которые обычно являются либо ферментами, либо ионными каналами, значительно усиливают получаемый ими сигнал, либо производя большие количества небольших внутриклеточных медиаторов, либо активируя большое количество нижестоящих внутриклеточных сигнальных белков.Когда в релейной цепи есть несколько этапов усиления, цепь часто называют сигнальным каскадом .
- 5.
-
Белки-преобразователи преобразуют сигнал в другую форму. Примером может служить фермент, производящий циклический АМФ: он как преобразует сигнал, так и усиливает его, тем самым действуя как преобразователь, так и усилитель.
- 6.
-
Белки бифуркации распространяют сигнал от одного сигнального пути к другому.
- 7.
-
Интегрирующие белки получают сигналы от двух или более сигнальных путей и интегрируют их перед передачей сигнала вперед.
- 8.
-
Скрытые регуляторные белки генов активируются на поверхности клетки активированными рецепторами, а затем мигрируют в ядро, чтобы стимулировать транскрипцию гена.
Как показано в blue in, другие типы внутриклеточных белков также играют важную роль во внутриклеточной передаче сигналов. Модуляторные белки модифицируют активность внутриклеточных сигнальных белков и, таким образом, регулируют силу передачи сигналов по пути. Заякоренные белки поддерживают определенные сигнальные белки в определенном месте в клетке, привязывая их к мембране или цитоскелету. Каркасные белки представляют собой адаптерные и / или закрепляющие белки, которые связывают несколько сигнальных белков вместе в функциональный комплекс и часто удерживают их в определенном месте.
Некоторые внутриклеточные сигнальные белки действуют как молекулярные переключатели
Многие внутриклеточные сигнальные белки ведут себя как молекулярные переключатели: при получении сигнала они переключаются из неактивного в активное состояние, пока другой процесс не отключит их.Как мы обсуждали ранее, выключение так же важно, как и включение. Если сигнальный путь должен восстановиться после передачи сигнала, чтобы он мог быть готов передать другой, каждая активированная молекула в этом пути должна быть возвращена в исходное инактивированное состояние.
Молекулярные переключатели делятся на два основных класса, которые действуют по-разному, хотя в обоих случаях именно усиление или потеря фосфатных групп определяет, является ли белок активным или неактивным.Самый большой класс состоит из белков, которые активируются или инактивируются фосфорилированием (обсуждается в главе 3). Для этих белков переключатель включается в одном направлении протеинкиназой, которая добавляет одну или несколько фосфатных групп к сигнальному белку, и в другом направлении протеинфосфатазой, которая удаляет фосфатные группы из протеина (). Подсчитано, что одна треть белков в эукариотической клетке фосфорилируется в любой момент времени.
Рисунок 15-17
Два типа внутриклеточных сигнальных белков, которые действуют как молекулярные переключатели.В обоих случаях сигнальный белок активируется добавлением фосфатной группы и инактивируется удалением фосфата. (A) Фосфат ковалентно добавляется к (подробнее …)
. Многие сигнальные белки, контролируемые фосфорилированием, сами по себе являются протеинкиназами, и они часто организованы в каскады фосфорилирования . Одна протеинкиназа, активируемая фосфорилированием, фосфорилирует следующую протеинкиназу в последовательности и т. Д., Передавая сигнал дальше и, в процессе, усиливая его, а иногда и распространяя по другим сигнальным путям.Два основных типа протеинкиназ действуют как внутриклеточные сигнальные белки. Подавляющее большинство — серин / треониновых киназ, фосфорилирующих белки по серинам и (реже) треонинам. Другие — тирозинкиназы, фосфорилирующие белки по тирозинам. Случайная киназа может делать и то, и другое. Секвенирование генома показывает, что около 2% наших генов кодируют протеинкиназы, и считается, что в типичной клетке млекопитающих присутствуют сотни различных типов протеинкиназ.
Другим основным классом молекулярных переключателей, участвующих в передаче сигналов, являются GTP-связывающие белки (обсуждаемые в главе 3). Они переключаются между активным состоянием, когда привязан GTP, и неактивным состоянием, когда привязан GDP. После активации они обладают внутренней активностью GTPase и отключаются путем гидролиза связанного GTP до GDP (). Существует два основных типа GTP-связывающих белков — большие тримерных GTP-связывающих белков (также называемых G-белками), , которые передают сигналы от рецепторов, связанных с G-белками (см.), И маленькие мономерные GTPases ( также называется мономерных GTP-связывающих белков). Последние также помогают передавать внутриклеточные сигналы, но, кроме того, они участвуют в регулировании везикулярного движения и многих других процессов в эукариотических клетках.
Как обсуждалось ранее, сложное поведение клеток, такое как выживаемость и пролиферация клеток, обычно стимулируется конкретными комбинациями внеклеточных сигналов, а не одним сигналом, действующим в одиночку (см.). Следовательно, клетка должна интегрировать информацию, поступающую от отдельных сигналов, чтобы дать соответствующий ответ — жить или умереть, делиться или нет и так далее.Эта интеграция обычно зависит от белков-интеграторов (см.), Которые эквивалентны микропроцессорам в компьютере: им требуется несколько входных сигналов для получения выходного сигнала, вызывающего желаемый биологический эффект. Два примера, которые показывают, как могут работать такие белки-интеграторы, проиллюстрированы на.
Рисунок 15-18
Интеграция сигналов. (A) Внеклеточные сигналы A и B активируют разные серии фосфорилирования белка, каждое из которых приводит к фосфорилированию белка Y, но в разных участках белка.Белок Y активируется только тогда, когда оба (подробнее …)
Внутриклеточные сигнальные комплексы увеличивают скорость, эффективность и специфичность ответа
Даже один тип внеклеточного сигнала, действующий через единственный тип G-белка, связанный с или связанный с ферментом рецептор обычно активирует множественные параллельные пути передачи сигналов и тем самым может влиять на множественные аспекты поведения клетки, такие как форма, движение, метаболизм и экспрессия генов. В самом деле, эти два основных класса рецепторов клеточной поверхности часто активируют одни и те же сигнальные пути, и обычно нет очевидной причины, почему конкретный внеклеточный сигнал использует один класс рецепторов, а не другой.
Сложность этих систем «сигнал-ответ» с множеством взаимодействующих релейных цепочек сигнальных белков устрашает. Неясно, как отдельной клетке удается отображать специфические ответы на такое множество различных внеклеточных сигналов, многие из которых связываются с одним и тем же классом рецепторов и активируют многие из одних и тех же сигнальных путей. Одна стратегия, которую клетка использует для достижения специфичности, включает каркасные белки (см.), Которые организуют группы взаимодействующих сигнальных белков в сигнальных комплексов ().Поскольку каркас управляет взаимодействиями между последовательными компонентами в таком комплексе, сигнал может передаваться с точностью, скоростью и эффективностью; более того, предотвращается нежелательное перекрестное взаимодействие между сигнальными путями. Однако, чтобы усилить сигнал и распространить его на другие части клетки, по крайней мере, некоторые из компонентов в большинстве сигнальных путей, вероятно, будут свободно диффундировать.
Рисунок 15-19
Два типа внутриклеточных сигнальных комплексов. (A) Рецептор и некоторые из внутриклеточных сигнальных белков, которые он последовательно активирует, предварительно собираются в сигнальный комплекс с помощью большого каркасного белка.(B) Собирается большой сигнальный комплекс (подробнее …)
В других случаях сигнальные комплексы образуются временно, например, когда сигнальные белки собираются вокруг рецептора после того, как его активировала внеклеточная сигнальная молекула. В некоторых из этих случаев цитоплазматический хвост активированного рецептора фосфорилируется во время процесса активации, и фосфорилированные аминокислоты затем служат в качестве стыковочных сайтов для сборки других сигнальных белков (). В других случаях активация рецептора приводит к продукции модифицированных молекул фосфолипидов в прилегающей плазматической мембране, и эти липиды затем привлекают специфические внутриклеточные сигнальные белки к этой области мембраны.Все такие сигнальные комплексы образуются лишь временно и быстро распадаются после того, как внеклеточный лиганд отделяется от рецептора.
Взаимодействия между внутриклеточными сигнальными белками опосредуются модульными связывающими доменами
Сборка как стабильных, так и временных сигнальных комплексов зависит от множества высококонсервативных малых связывающих доменов , которые обнаруживаются во многих внутриклеточных сигнальных белках. Каждый из этих компактных белковых модулей связывается с определенным структурным мотивом в белке (или липиде), с которым взаимодействует сигнальный белок.Из-за этих модульных доменов сигнальные белки связываются друг с другом во множестве комбинаций, как кирпичики Lego, при этом белки часто образуют трехмерную сеть взаимодействий, которая определяет маршрут, по которому следует сигнальный путь. Объединяя существующие домены вместе в новые комбинации, использование таких модульных связывающих доменов, по-видимому, облегчило быструю эволюцию новых сигнальных путей.
Домены гомологии 2 Src (Sh3) и фосфотирозин-связывающих (PTB) доменов , например, связываются с фосфорилированными тирозинами в конкретной пептидной последовательности на активированных рецепторах или внутриклеточных сигнальных белках. Домены гомологии 3 Src (Sh4) связываются с короткой, богатой пролином аминокислотной последовательностью. Гомология плэкстрина (PH) доменов (впервые описанных в белке плэкстрина в тромбоцитах крови) связываются с заряженными головными группами специфических фосфорилированных фосфолипидов инозитола, которые продуцируются в плазматической мембране в ответ на внеклеточный сигнал; тем самым они позволяют белку, частью которого они являются, закрепляться на мембране и взаимодействовать с другими рекрутированными сигнальными белками. Некоторые сигнальные белки функционируют только как адаптеры для связывания двух других белков вместе в сигнальном пути, и они состоят исключительно из двух или более связывающих доменов ().
Рисунок 15-20
Гипотетический путь передачи сигналов с использованием модульных связывающих доменов. Сигнальный белок 1 содержит три разных связывающих домена, а также домен каталитической протеинкиназы. Он перемещается к плазматической мембране, когда внеклеточные сигналы приводят к созданию различных (подробнее …)
Каркасных белков часто содержат несколько PDZ-доменов (первоначально обнаруженных в области синапса, называемой постсинаптической плотностью), каждый из которых связывается со специфическим мотивом рецептора или сигнального белка.Ярким примером является каркасный белок InaD в фоторецепторных клетках Drosophila . Он содержит пять доменов PDZ, один из которых связывает активируемый светом ионный канал, а остальные связываются с разными сигнальными белками, участвующими в реакции клетки на свет. Если какой-либо из этих PDZ-доменов отсутствует, соответствующий сигнальный белок не может собираться в комплекс, и зрение мухи является дефектным.
Считается, что некоторые рецепторы клеточной поверхности и внутриклеточные сигнальные белки временно группируются в определенные микродомены в липидном бислое плазматической мембраны, которые обогащены холестерином и гликолипидами.Некоторые из белков направляются к этим липидным рафтам ковалентно прикрепленными липидными молекулами. Подобно каркасным белкам, эти липидные каркасы могут способствовать скорости и эффективности сигнального процесса, выступая в качестве сайтов, где сигнальные молекулы могут собираться и взаимодействовать (см.).
Клетки могут резко реагировать на постепенно увеличивающуюся концентрацию внеклеточного сигнала
Некоторые клеточные ответы на внеклеточные сигнальные молекулы плавно изменяются в простой пропорции к концентрации молекулы.Первичные реакции на стероидные гормоны (см.) Часто следуют этой схеме, предположительно потому, что белок рецептора ядерного гормона связывает единственную молекулу гормона, и каждая конкретная последовательность распознавания ДНК в гене, отвечающем на стероидные гормоны, действует независимо. По мере увеличения концентрации гормона концентрация активированных комплексов рецептор-гормон пропорционально увеличивается, как и количество комплексов, связанных со специфическими последовательностями распознавания в ответных генах; поэтому ответ клетки постепенный и линейный.
Однако многие ответы на внеклеточные сигнальные молекулы начинаются более резко, когда концентрация молекулы увеличивается. Некоторые из них могут даже происходить практически по принципу «все или ничего», будучи необнаруживаемой ниже пороговой концентрации молекулы, а затем достигая максимума, как только эта концентрация будет превышена. Что может быть молекулярной основой таких резких или даже переключающихся ответов на ступенчатые сигналы?
Одним из механизмов усиления ответа является требование, чтобы более одной внутриклеточной эффекторной молекулы или комплекса связывались с некоторой макромолекулой-мишенью, чтобы вызвать ответ.Например, в некоторых ответах, вызванных стероидными гормонами, кажется, что более одного активированного комплекса рецептор-гормон должны одновременно связываться со специфическими регуляторными последовательностями в ДНК, чтобы активировать конкретный ген. В результате, когда концентрация гормона повышается, активация гена начинается более резко, чем если бы для активации было достаточно только одного связанного комплекса (). Подобный кооперативный механизм часто действует в сигнальных каскадах, активируемых рецепторами клеточной поверхности. Как мы обсудим позже, четыре молекулы небольшого внутриклеточного медиатора циклического АМФ, например, должны связываться с каждой молекулой циклической-АМФ-зависимой протеинкиназы, чтобы активировать киназу.Такие ответы становятся более острыми по мере увеличения количества взаимодействующих молекул, и если их количество достаточно велико, могут быть достигнуты ответы, приближающиеся к типу «все или ничего» (и).
Рисунок 15-21
Первичная реакция клеток яйцевода цыплят на стероидный половой гормон эстрадиол. При активации рецепторы эстрадиола включают транскрипцию нескольких генов. Показаны кривые доза-ответ для двух из этих генов, один кодирует яичный белок кональбумин (подробнее …)
Рисунок 15-22
Кривые активации как функция концентрации сигнальных молекул.Кривые показывают, как резкость ответа увеличивается с увеличением количества эффекторных молекул, которые должны связываться одновременно, чтобы активировать целевую макромолекулу. Кривые (подробнее …)
Рисунок 15-23
Один тип сигнального механизма, который, как ожидается, покажет крутой пороговый отклик. Здесь одновременное связывание восьми молекул сигнального лиганда с набором из восьми белковых субъединиц требуется для образования активного белкового комплекса. Способность (подробнее …)
Ответы также обостряются, когда внутриклеточная сигнальная молекула активирует один фермент и в то же время ингибирует другой фермент, который катализирует противоположную реакцию. Хорошо изученным примером этого распространенного типа регуляции является стимуляция распада гликогена в клетках скелетных мышц, вызванная гормоном адреналином (адреналином). Связывание адреналина с рецептором клеточной поверхности, связанным с G-белком, приводит к увеличению внутриклеточной концентрации циклического АМФ, который одновременно активирует фермент, способствующий распаду гликогена, и ингибирует фермент, который способствует синтезу гликогена.
Все эти механизмы могут вызывать очень резкие ответы, но, тем не менее, всегда плавно меняющиеся в зависимости от концентрации внеклеточной сигнальной молекулы. Другой механизм, однако, может производить истинные ответы типа «все или ничего», так что повышение сигнала выше критического порогового уровня вызывает внезапное переключение в отвечающей ячейке. Пороговые отклики типа «все или ничего» этого типа обычно зависят от положительной обратной связи ; по этому механизму нервные и мышечные клетки генерируют потенциалов действия по принципу «все или ничего» в ответ на нейротрансмиттеры (обсуждается в главе 11).Активация связанных с ионным каналом ацетилхолиновых рецепторов в нервно-мышечном соединении, например, приводит к чистому притоку Na + , который локально деполяризует плазматическую мембрану мышц. Это заставляет управляемые по напряжению каналы Na + открываться в той же области мембраны, производя дополнительный приток Na + , который дополнительно деполяризует мембрану и тем самым открывает больше каналов Na + . Если начальная деполяризация превышает определенное пороговое значение, эта положительная обратная связь имеет взрывной эффект «убегания», создавая потенциал действия, который распространяется на всю мышечную мембрану.
Ускоряющий механизм положительной обратной связи может также работать через сигнальные белки, которые являются ферментами, а не ионными каналами. Предположим, например, что конкретный внутриклеточный сигнальный лиганд активирует фермент, расположенный ниже по ходу сигнального пути, и что две или более молекулы продукта ферментативной реакции связываются обратно с одним и тем же ферментом для его дальнейшей активации (). Следствием этого является очень низкая скорость синтеза продукта в отсутствие лиганда. Скорость медленно увеличивается с концентрацией лиганда до тех пор, пока на некотором пороговом уровне лиганда не будет синтезировано достаточное количество продукта для активации фермента самоускоряющимся, неуправляемым образом.Затем концентрация продукта внезапно повышается до гораздо более высокого уровня. Посредством этих и ряда других механизмов, не обсуждаемых здесь, клетка часто переводит постепенное изменение концентрации сигнального лиганда в переключательное изменение, создавая ответ «все или ничего».
Рисунок 15-24
Ускоряющий механизм положительной обратной связи. В этом примере начальное связывание сигнального лиганда активирует фермент с образованием продукта, который обратно связывается с ферментом, дополнительно увеличивая активность фермента.
Клетка может запоминать влияние некоторых сигналов
Влияние внеклеточного сигнала на целевую клетку в некоторых случаях может сохраняться и после того, как сигнал исчез. Только что описанная ферментативная ускоряющая система положительной обратной связи представляет собой один из типов механизмов, демонстрирующих такую устойчивость. Если такая система была включена путем повышения концентрации внутриклеточного активирующего лиганда выше порогового значения, она, как правило, останется включенной даже после исчезновения внеклеточного сигнала; вместо того, чтобы точно отражать текущий уровень сигнала, система отклика отображает память.Мы встретимся с конкретным примером этого позже, когда будем обсуждать протеинкиназу, которая активируется Ca 2+ для фосфорилирования себя и других белков; аутофосфорилирование сохраняет активность киназы еще долгое время после того, как уровни Ca 2+ вернутся к норме, обеспечивая след в памяти исходного сигнала.
Временные внеклеточные сигналы часто вызывают гораздо более долгосрочные изменения в клетках во время развития многоклеточного организма. Некоторые из этих изменений могут сохраняться на протяжении всей жизни организма.Обычно они зависят от самоактивирующихся механизмов памяти, которые действуют ниже по сигнальному пути, на уровне транскрипции генов. Например, сигналы, запускающие детерминацию мышечных клеток, включают серию регулирующих белков специфичных для мышц генов, которые стимулируют транскрипцию собственных генов, а также генов, продуцирующих многие другие белки мышечных клеток. Таким образом, решение стать мышечной клеткой становится постоянным (см.).
Клетки могут регулировать свою чувствительность к сигналу
В ответ на многие типы стимулов клетки и организмы могут обнаруживать одинаковый процент изменения сигнала в очень широком диапазоне интенсивности стимула.Это требует, чтобы клетки-мишени претерпели обратимый процесс адаптации или десенсибилизации , в результате чего длительное воздействие стимула снижает реакцию клеток на этот уровень воздействия. В химической передаче сигналов адаптация позволяет клеткам реагировать на изменений концентрации сигнального лиганда (а не на абсолютную концентрацию лиганда) в очень широком диапазоне концентраций лиганда. Общий принцип — отрицательная обратная связь, которая действует с задержкой.Сильный ответ модифицирует механизм, отвечающий за этот ответ, таким образом, что механизм возвращается в исходное положение. Однако из-за задержки внезапное изменение стимула может дать о себе знать в течение короткого периода времени, прежде чем сработает отрицательная обратная связь.
Десенсибилизация к сигнальной молекуле может происходить по-разному. Например, связывание лиганда с рецепторами клеточной поверхности может вызывать их эндоцитоз и временную секвестрацию в эндосомах. Такой индуцированный лигандом рецепторный эндоцитоз может приводить к разрушению рецепторов в лизосомах, процесс, называемый понижающей регуляцией рецептора . В других случаях десенсибилизация является результатом быстрой инактивации рецепторов — например, в результате фосфорилирования рецептора, которое следует за его активацией с задержкой. Десенсибилизация также может быть вызвана изменением белка, участвующего в передаче сигнала, или выработкой ингибитора, блокирующего процесс трансдукции ().
Рисунок 15-25
Пять способов, которыми клетки-мишени могут стать десенсибилизированными к сигнальной молекуле. Механизмы инактивации, показанные здесь как для рецептора, так и для внутриклеточного сигнального белка, часто включают фосфорилирование белка, который инактивируется, хотя (more…)
Обсудив некоторые общие принципы клеточной передачи сигналов, мы теперь обратимся к рецепторам, связанным с G-белком. Это, безусловно, самый большой класс рецепторов клеточной поверхности, и они опосредуют ответы на подавляющее большинство внеклеточных сигналов. Это суперсемейство рецепторных белков не только обеспечивает межклеточную коммуникацию; это также центральное место для зрения, обоняния и вкусового восприятия.
Резюме
Каждая клетка в многоклеточном животном была запрограммирована во время развития, чтобы реагировать на определенный набор внеклеточных сигналов, производимых другими клетками.Эти сигналы действуют в различных комбинациях, чтобы регулировать поведение клетки. Большинство сигналов опосредуют форму передачи сигналов, при которой локальные медиаторы секретируются, но затем быстро захватываются, разрушаются или иммобилизуются, так что они действуют только на соседние клетки. Другие сигналы остаются связанными с внешней поверхностью сигнальной клетки и опосредуют контактно-зависимую передачу сигналов. Централизованный контроль осуществляется как с помощью эндокринной передачи сигналов, при которой гормоны, секретируемые эндокринными клетками, переносятся в кровь к клеткам-мишеням по всему телу, так и с помощью синаптической передачи сигналов, при которой нейротрансмиттеры, секретируемые аксонами нервных клеток, действуют локально на постсинаптические клетки, которые аксоны контакт.
Для передачи сигналов в клетке требуются не только внеклеточные сигнальные молекулы, но и дополнительный набор рецепторных белков в каждой клетке, которые позволяют ей связываться и отвечать на сигнальные молекулы характерным образом. Некоторые небольшие гидрофобные сигнальные молекулы, включая стероидные и тироидные гормоны, диффундируют через плазматическую мембрану клетки-мишени и активируют внутриклеточные рецепторные белки, которые непосредственно регулируют транскрипцию определенных генов. Растворенные газы оксид азота и оксид углерода действуют как местные медиаторы, диффундируя через плазматическую мембрану клетки-мишени и активируя внутриклеточный фермент — обычно гуанилилциклазу, которая продуцирует циклический GMP в клетке-мишени.Но большинство внеклеточных сигнальных молекул гидрофильны и могут активировать рецепторные белки только на поверхности клетки-мишени; эти рецепторы действуют как преобразователи сигналов, преобразовывая событие внеклеточного связывания во внутриклеточные сигналы, которые изменяют поведение клетки-мишени.
Существует три основных семейства рецепторов клеточной поверхности, каждое из которых по-своему передает внеклеточные сигналы. Рецепторы, связанные с ионным каналом, представляют собой управляемые передатчиком ионные каналы, которые на короткое время открываются или закрываются в ответ на связывание нейромедиатора.Связанные с G-белком рецепторы косвенно активируют или инактивируют связанные с плазматической мембраной ферменты или ионные каналы через тримерные GTP-связывающие белки (G-белки). Связанные с ферментом рецепторы либо действуют непосредственно как ферменты, либо связаны с ферментами; эти ферменты обычно представляют собой протеинкиназы, которые фосфорилируют определенные белки в клетке-мишени.
После активации рецепторы, связанные с ферментом и G-белками, передают сигнал внутрь клетки, активируя цепи внутриклеточных сигнальных белков; некоторые преобразовывают, усиливают или распространяют сигнал по мере его передачи, в то время как другие интегрируют сигналы от различных путей передачи сигналов.Многие из этих сигнальных белков функционируют как переключатели, которые временно активируются фосфорилированием или связыванием GTP. Функциональные сигнальные комплексы часто образуются посредством модульных связывающих доменов в сигнальных белках; эти домены позволяют сложным белковым сборкам функционировать в сигнальных сетях.
Клетки-мишени могут использовать различные внутриклеточные механизмы для резкого ответа на постепенно увеличивающуюся концентрацию внеклеточного сигнала или для преобразования кратковременного сигнала в длительный ответ.Кроме того, посредством адаптации они часто могут обратимо регулировать свою чувствительность к сигналу, чтобы позволить клеткам реагировать на изменения концентрации конкретной сигнальной молекулы в большом диапазоне концентраций.
Сотовая связь — обзор
2.3.4 Стратегическое развертывание сети
Очевидно, что стратегическое развертывание сети определенно сможет снизить потребление энергии в сотовой связи. В большинстве случаев исследователи в основном сосредотачиваются на характеристиках сети, таких как покрытие, спектральная эффективность и емкость (Hanly and Mathar, 2002).Другие исследователи сосредотачиваются на оптимальном размере соты (Chen et al., 2010), возникающих гетерогенных сетях (смесь макросот, микросот, пикосот и фемтосот), различных релейных и кооперативных связях и т. Д. Richter et al. достигли ряда значительных результатов (Richter and Fettweis, 2009, 2010; Fehske et al., 2009; Richter et al., 2009) для энергоэффективных приложений в гетерогенных сетях. Они изучили оптимальную компоновку микроячеек поверх обычных макроэлементов.В частности, многие исследователи сосредотачиваются на простых моделях энергопотребления различных BS, рассматривая меньшее количество сценариев в качестве тематических исследований. В их исследованиях мощность передачи была разделена на зависимые части (усилитель, фидер, охлаждающие устройства, связанные с передачей) и независимые части (мощность схемы для обработки сигналов, резервная батарея, потребление охлаждения объекта и т. Д.). Энергопотребление на БС моделировалось как сумма этих двух типов мощности. Обратите внимание, что потребление энергии зависимой частью линейно зависит от средней излучаемой мощности.Кроме того, также учитывалось влияние межсайтового расстояния и среднего количества микросайтов на макроячейку на энергопотребление площади. В своих более поздних работах они исследовали потенциальное снижение энергии за счет изменения количества микросайтов и размера макроячеек для получения заданных целевых значений спектральной эффективности в условиях полной нагрузки. Их результаты показали, что развертывание микросайтов было намного предпочтительнее из-за значительного сокращения энергопотребления области в сети при сохранении требуемой целевой пропускной способности области.Кроме того, они провели сравнение энергопотребления области со спектральной эффективностью области однородных макросайтов, однородных микросайтов и гетерогенных сетей. Их результаты указывают на улучшение EE, в то время как развертывание дополнительных микросайтов необходимо при условии более высокой целевой пропускной способности с более высокой плотностью пользователей. Помимо этого, они пришли к выводу, что развертывание пикосот и фемтосот является хорошей стратегией для предоставления экономически эффективных услуг.
Кроме того, может быть выгодно комбинировать технологии беспроводной и оптической связи, чтобы снизить общее энергопотребление (Zhang et al., 2010). Эту технологию можно рассматривать как распределенную антенную систему , определенную в системе Long-term Evolution Advanced (LTE-A). Эта идея выделяет два основных блока для BS: блок основной полосы частот (BBU) и блок удаленной радиосвязи (RRU). RRU — это географически разделенные точки в координированной многоточечной системе , которая подключается к BBU через оптическое волокно. Цель этой технологии — децентрализовать развертывание антенн для расширения зоны покрытия соты.Таким образом, это увеличивает пропускную способность системы из-за того, что расстояние между пользователем и антеннами было сокращено, тем самым уменьшая потребление энергии между пользователем и антеннами. Поскольку расстояние между пользователем и антенной варьируется в широких пределах, это может вызвать колебания мощности передачи. Следовательно, для достижения лучшего EE необходим эффективный алгоритм развертывания RRU.
С другой стороны, зеленая сотовая связь — это еще одна новая архитектура, предложенная Эзри и Шило (2009), нацеленная на минимальное излучение от мобильных станций без каких-либо дополнительных источников излучения.Эта новая архитектура оснащена «дублированными зелеными антеннами» на каждом приемопередатчике базовой станции. Мобильные пользователи, расположенные рядом с зелеными антеннами, могут передавать с меньшей мощностью передачи и, таким образом, снижать энергопотребление с меньшим количеством проблем с помехами. Помимо этих преимуществ, зеленые антенны не создают дополнительного излучения, поскольку ретранслятор трафика касается только восходящего канала.
Одной из актуальных тем, касающихся EE, является архитектура ретрансляции и совместной связи.Архитектура с большим количеством узлов ретрансляции может сэкономить энергию, поскольку она снижает потери на пути из-за более короткого диапазона передачи. Следовательно, эта архитектура создает меньше помех из-за низкой мощности передачи (Bae and Stark, 2009; Miao et al., 2009). В отличие от обычных ретрансляционных систем, каждый кооперативный узел в кооперативных коммуникациях способен генерировать источник информации, а также ретранслировать информацию. Эта архитектура использует разнесение каналов для потенциальной экономии энергии.Cui et al. (2004) показали, что в некоторых диапазонах расстояний совместная передача и прием с множеством входов и множеством выходов (MIMO) также может способствовать экономии энергии. Более того, они также показали, что размер созвездия для разных расстояний передачи может улучшить EE совместной связи, которая лучше, чем прямая связь. Однако недостатком как ретрансляционной, так и совместной связи является то, что выбор оптимальных партнеров может быть чрезвычайно сложной задачей.Еще одна важная проблема — это распределение ресурсов для достижения минимального потребления энергии.
Технология мобильных телефонов »Электроника
Мобильный телефон или сотовая технология широко используются и основаны на концепции повторного использования частоты приложением в серии ячеек покрытия.
Основы сотовой / мобильной связи Включает:
Что такое сотовая связь
Понятие сотовой системы
Методы множественного доступа
Дуплексные методы
Что внутри мобильного телефона
Сдавать
Обратный рейс
Мобильные телефоны или сотовые телекоммуникационные технологии широко используются с начала 1980-х годов.
С момента своего первого внедрения, его использование очень быстро увеличилось до такой степени, что большая часть населения мира имеет доступ к технологии.
От развитой страны к развивающейся стране мобильные телефоны или сотовая связь используются во всех странах по всему миру.
Отрасль сотовой связи была основным двигателем роста радио и электронной промышленности.
Развитие сотовой связи
Хотя сотовая связь сейчас принята в повседневной жизни, потребовалось много лет, чтобы они начали развиваться.
Хотя основные концепции технологии сотовой связи были предложены в 1940-х годах, только в середине 1980-х годов технологии и системы радиосвязи были развернуты, чтобы обеспечить широкую доступность.
Использование систем сотовой связи быстро росло, и, например, было подсчитано, что в Соединенном Королевстве к 2011 году было совершено больше звонков с мобильных телефонов, чем с проводных устройств.
Другой пример роста систем сотовой связи произошел в 2004 году, когда GSMA объявила на Всемирном мобильном конгрессе в феврале 2004 года, что количество абонентов мобильной связи стандарта GSM превышает 1 миллиард — прошло 12 лет с момента запуска первой сети.Для сравнения, чтобы достичь того же показателя по проводным телефонным соединениям, потребовалось более 100 лет.
Тогда к 2015 году было активным более 7 миллиардов мобильных подписок (по всем технологиям). Это большой подвиг, если учесть, что население Земли составляло чуть более 7 миллиардов человек. Это означало, что у многих людей было более одной подписки, хотя проникновение на рынок, очевидно, было очень значительным.
Поколения сотовой связи
О поколениях мобильных телефонов много говорят.3G переходит на 4G, а затем на 5G.
Каждое поколение мобильных телефонов преследовало свои цели и могло обеспечивать разные уровни функциональности.
Также могло существовать несколько различных конкурирующих стандартов в пределах разных поколений. Для сотовой связи 3G существовало два основных стандарта, а для 4G — только один, поскольку существовал глобальный консенсус по использованию системы, и это способствовало глобальному роумингу.
| Поколение | Примерный год выпуска | Фокус |
|---|---|---|
| 1 г | 1979 | Мобильный голос |
| 2G | 1991 | Мобильный голос |
| 3G | 2001 | Мобильная широкополосная связь |
| 4G | 2009 | Мобильная широкополосная связь |
| 5G | 2020 (ожидается) | Повсеместное подключение |
Ключевые концепции сотовой связи
Как видно из названия, технология сотовой связи основана на концепции использования большого количества базовых станций, каждая из которых покрывает небольшую территорию или соту.Поскольку каждая базовая станция обменивается данными с разумным количеством пользователей, это означает, что вся система может поддерживать огромное количество подключений, а уровни использования частот являются хорошими.
Система сотовой связи имеет ряд различных областей, каждая из которых выполняет свою функцию. Основные области, подробно описанные ниже, являются основными, на которые обычно ссылаются при обсуждении систем сотовой связи. Каждую из этих областей часто можно разделить на разные сущности.
- Мобильный телефон или пользовательское оборудование, UE: Пользовательское оборудование или мобильный телефон — это элемент системы мобильной связи, который видит пользователь. Он подключается к сети и позволяет пользователю получать доступ к услугам передачи голоса и данных. Пользовательское оборудование также может быть ключом, используемым для доступа к данным на ноутбуке, или модемом на другом устройстве — например, сотовая связь начинает использоваться для Интернета вещей, приложений IoT и, как следствие, может быть подключен к интеллектуальному счетчику для автоматической отправки показаний счетчика или может использоваться для любого из множества других приложений.
- Сеть радиодоступа, RAN: Сеть радиодоступа находится на периферии системы сотовой связи. Он обеспечивает связь с пользовательским оборудованием из сотовой сети. Он состоит из ряда элементов и в целом включает в себя базовую станцию и контроллер базовой станции. С развитием технологий сотовой связи используемые термины и то, что они содержат, меняются, но их основная функция остается, по сути, той же.
- Базовая сеть: Базовая сеть является центром системы сотовой связи.Он управляет всей системой, а также хранит пользовательские данные, управляет контролем доступа, связывает с внешним миром и предоставляет множество других функций.
Темы беспроводного и проводного подключения:
Основы мобильной связи
2G GSM
3G UMTS
4G LTE
5G
Вай фай
IEEE 802.15.4
Беспроводные телефоны DECT
NFC — связь ближнего поля
Основы сетевых технологий
Что такое облако
Ethernet
Серийные данные
USB
SigFox
LoRa
VoIP
SDN
NFV
SD-WAN
Вернуться к беспроводному и проводному подключению
«Глава 11 — Принципы передачи сигналов клетками» в «Основах клеточной биологии» на OpenALG
Как клетки общаются?
Передача сигналов клеток (также называемая сигнальной трансдукцией) описывает способность клеток реагировать на стимулы из окружающей среды.Некоторые примеры включают заживление ран после травмы, активацию иммунной системы в ответ на патогены и изменения экспрессии генов на разных стадиях развития. Связь между ячейками называется межклеточной передачей сигналов , а связь внутри ячейки называется внутриклеточной передачей сигналов . Легкий способ запомнить различие — это понять латинское происхождение префиксов: inter — означает «между» (например, пересекающиеся линии — это те, которые пересекают друг друга) и intra — означает «внутри» (например, внутривенный).
Сигнальные молекулы и рецепторы
Передача сигналов в клетке инициируется сигнальными молекулами (также называемыми лигандами ). Сигнальными молекулами могут служить разные типы молекул. Это могут быть белки (например, инсулин, гормон роста человека), липиды (например, простагландины, тестостерон, кортизол), метаболиты аминокислот (например, гистамин, дофамин), газы (например, оксид азота) и многие другие. Сигнальные молекулы взаимодействуют с белками в клетках-мишенях или на них, которые являются клетками, на которые воздействуют химические сигналы; белки, которые отвечают на лиганды, также называются рецепторами .Сигнальные молекулы и рецепторы существуют в нескольких разновидностях; однако сигнальная молекула будет связываться со специфическим рецептором, который обычно связывает только один тип сигнальной молекулы. Эта специфичность возникает из-за того, что сигнальные молекулы имеют уникальный заряд, форму, размер и гидрофобность, которые позволяют им связываться только с конкретными, комплементарными областями на рецепторах. Таким образом, можно думать о взаимодействиях сигнальных молекул и рецепторов как о «ключах» и «замках».
Типы передачи сигналов клеток
Передача сигналов клеток может быть разделена на четыре основные категории в зависимости от того, насколько далеко сигнальная молекула находится от рецептора и где находится рецептор (Рисунок 11-1):
-
Передача сигналов на большие расстояния ( эндокринный)
-
Передача сигналов ближнего действия ( паракрин)
-
Самоактивация ( аутокрин)
-
Прямая активация ( контактно-зависимая t, также называется юкстакрин 906
Рисунок 11-1: Четыре типа передачи сигналов соты. Передача сигналов клетки делится на четыре основные категории в зависимости от местоположения рецептора и расстояния, пройденного сигнальной молекулой. К этим категориям относятся эндокринная (передача сигналов дальнего действия), паракринная (локальная передача сигналов), аутокринная (самостимуляция клеток) и контактно-зависимая / юкстакринная передача сигналов (передача сигналов, требующая прямого контакта между клетками).
Эндокринные сигналы
Сигналы от удаленных клеток называются эндокринными сигналами , и они исходят от эндокринных клеток.В организме многие эндокринные клетки расположены в эндокринных железах, таких как щитовидная железа, гипоталамус и гипофиз. Эти типы сигналов обычно вызывают более медленную реакцию, но имеют более продолжительный эффект. Лиганды, высвобождаемые при эндокринной передаче сигналов, называются гормонами, сигнальными молекулами, которые вырабатываются в одной части тела, но воздействуют на другие области тела на некотором расстоянии (рис. 11-1).
Гормоны перемещаются на огромные расстояния между эндокринными клетками и их клетками-мишенями через кровоток, что является относительно медленным способом перемещения по телу.Из-за своей формы транспорта гормоны разбавляются и присутствуют в низких концентрациях, когда они действуют как лиганды для рецепторов на своих клетках-мишенях.
Паракринная передача сигналов
Сигналы, которые действуют локально между клетками, которые расположены близко друг к другу, называются паракринными сигналами (рисунок 11-1). Паракринные сигналы распространяются через внеклеточный матрикс. Эти типы сигналов обычно вызывают быстрые реакции, которые длятся недолго.Чтобы реакция оставалась локализованной, молекулы паракринного лиганда обычно быстро разрушаются ферментами или удаляются соседними клетками. Удаление сигналов восстановит градиент концентрации сигнала, позволяя им диффундировать через внутриклеточное пространство, если они быстро высвобождаются снова.
Воспалительная реакция — один из примеров паракринной передачи сигналов. В ответ на травму (например, укус насекомого) клетки выделяют сигнальные молекулы, называемые цитокинами. Цитокины привлекают и активируют иммунные клетки рядом с местом повреждения, что приводит к покраснению и отеку.Это первый шаг к заживлению ран.
Другой пример паракринной передачи сигналов — передача сигналов через синапсы между нервными клетками (рис. 11-2). Нервная клетка состоит из тела клетки, нескольких коротких разветвленных отростков, называемых дендритами, которые получают стимулы, и длинного отростка, называемого аксоном, который передает сигналы другим нервным клеткам или мышечным клеткам. Соединение между нервными клетками, где происходит передача сигнала, называется синапсом. Синаптический сигнал — это химический сигнал, который проходит между нервными клетками.Быстро движущиеся электрические импульсы распространяют сигналы внутри нервных клеток. Когда эти импульсы достигают конца аксона, сигнал продолжается к дендриту следующей клетки путем высвобождения химических лигандов, называемых нейротрансмиттерами , пресинаптической клеткой (клеткой, излучающей сигнал). Нейромедиаторы переносятся на очень небольшие расстояния между нервными клетками, которые называются химическими синапсами. Небольшое расстояние между нервными клетками позволяет сигналу быстро распространяться; это позволяет мгновенно отреагировать, например убрать руку с горячей поверхности плиты!
Когда нейромедиатор связывает рецептор на поверхности постсинаптической клетки, электрохимический потенциал клетки-мишени изменяется, и запускается следующий электрический импульс.Нейротрансмиттеры, которые высвобождаются в химический синапс, быстро разрушаются или реабсорбируются пресинаптической клеткой, так что нервная клетка-реципиент может быстро восстановиться и быть подготовленной к быстрому ответу на следующий синаптический сигнал.
Рисунок 11-2: Передача нейронных сигналов. В нейрональной передаче сигналов сигнальные молекулы высвобождаются пресинаптической клеткой и связываются с рецепторами на соседней постсинаптической клетке. Это пример паракринной передачи сигналов.
Автокринная передача сигналов
Аутокринные сигналы производятся сигнальными клетками, которые также могут связываться с высвобождаемым лигандом (рис. 11-1). Это означает, что сигнальная ячейка и целевая ячейка — это одно и то же (префикс автоматически означает себя, напоминание о том, что сигнальная ячейка отправляет сигнал самой себе). Этот тип передачи сигналов часто возникает на раннем этапе развития организма, чтобы гарантировать, что клетки развиваются в правильные ткани и принимают правильные функции.Аутокринная передача сигналов также регулирует болевые ощущения и воспалительные реакции. Кроме того, если клетка инфицирована вирусом, клетка может подать себе сигнал о запрограммированной гибели клетки, убивая вирус в процессе. В некоторых случаях соседние клетки того же типа также подвержены влиянию высвобождаемого лиганда, поэтому также индуцируют паракринную передачу сигналов. В эмбриологическом развитии этот процесс стимуляции группы соседних клеток может помочь направить дифференцировку идентичных клеток в один и тот же тип клеток, обеспечивая тем самым правильный результат для развития.
Контактно-зависимая передача сигналов
В контактно-зависимой передаче сигналов (также называемой передачей сигналов juxtacrine ) клетки должны физически взаимодействовать, чтобы инициировать передачу сигналов клеток (рис. 11-1). Это может происходить через щелевые соединения и / или через взаимодействия трансмембранных белков. В этом случае сигнальная молекула / лиганд обычно связана с мембраной одной клетки, поэтому клетке необходимо вступить в прямой контакт с рецептором на клетке-мишени для доставки сигнала.Одним из примеров контактно-зависимой передачи сигналов является путь Delta-Notch, используемый в эмбриональном развитии. Дельта, трансмембранный белок сигнальной клетки, связывается с Notch, рецептором на принимающей клетке. Notch изменяет свою конформацию, позволяя его цитоплазматическому домену быть отрезанным ферментом, называемым секретазой. Затем цитоплазматический домен перемещается в ядро, где он действует как активирующий фактор транскрипции для усиления экспрессии генов, необходимых на следующей стадии развития.
Компоненты сигнального пути клетки
Как упоминалось ранее, передача сигнала клетки инициируется, когда сигнальная молекула / лиганд связывается с соответствующим ей рецептором. После активации рецептора в клетке запускается серия событий (рис. 11-3), которые в конечном итоге приводят к результирующему клеточному ответу (например, изменению экспрессии генов или изменению метаболизма). Эти события описаны в следующих разделах.
Внутриклеточные сигнальные каскады и эффекторы
Рецепторы, даже при высокой плотности, составляют лишь мельчайшую долю площади поверхности клетки и, следовательно, еще меньшую долю объема клетки.Следовательно, активация рецептора должна быть усилена, чтобы он инициировал клеточную активность (например, движение, рост, прогрессирование клеточного цикла). Таким образом, одно из первых действий рецептора при активации или связывании лиганда — это инициировать внутриклеточный сигнальный каскад (рис. 11-3). Эта точно названная последовательность событий начинается с того, что рецептор активирует фермент. Фермент может быть цитоплазматическим доменом самого рецептора или может быть независимым белком, но тесно связанным с рецептором.Фермент делает то же, что и фермент: он быстро превращает молекулы субстрата в молекулы продукта. В этом случае иногда продукт является активатором другого фермента, а иногда субстратом является неактивный фермент, а продукт — активированный фермент. В любом случае, из-за высоких показателей активности однократная активация рецептора увеличивала десятки или сотни активаций ферментов. Каждый из них активирует еще сотню и так далее, так что действие рецептора может быстро распространяться по клетке.
В некоторых случаях молекулы внутриклеточного сигнального каскада будут производить небольшие внутриклеточные молекулы, которые служат для усиления клеточного ответа на первичные сигнальные молекулы. Эти молекулы называются вторичными посредниками (рис. 11-3).
Рисунок 11-3: Общий путь передачи сигналов ячейки. Передача сигналов в клетке начинается, когда сигнальная молекула связывается со своим рецептором. Рецепторы находятся как внутри клетки, так и на ее поверхности.После стимуляции сигнальной молекулой рецептор инициирует внутриклеточный сигнальный каскад, который представляет собой серию ферментативных реакций. Многие из этих ферментов представляют собой киназы, фосфатазы или GTPases. В некоторых случаях внутриклеточные сигнальные каскады продуцируют вторичные мессенджеры, небольшие внутриклеточные молекулы, которые служат для усиления клеточного ответа на первичную сигнальную молекулу. Результатом каскада является активация эффекторных белков. Эффекторные белки облегчают клеточный ответ на сигнальную молекулу.Эффекторами, участвующими в «быстрых» ответах (от секунды до нескольких минут), обычно являются метаболические ферменты и / или белки ремоделирования цитоскелета. Факторы транскрипции служат эффекторами для более медленных ответов (от нескольких минут до часов), которые зависят от изменений в экспрессии генов.
Двумя наиболее распространенными ферментами внутриклеточных сигнальных каскадов являются киназы и ГТФазы. Киназы (рис. 11-4) способствуют добавлению фосфатной группы (PO 4 –3 ) к молекуле, такой как белок, в процессе, называемом фосфорилированием .Киназы добавляют фосфатные группы к остаткам серина, треонина и тирозина белков, где они заменяют гидроксильную группу аминокислоты. Различные киназы названы в честь субстрата, который они фосфорилируют. Фосфорилирование остатков серина и треонина часто активирует ферменты. Фосфорилирование остатков тирозина может либо влиять на активность фермента, либо создавать сайт связывания, который взаимодействует с нижележащими компонентами сигнального каскада. Фосфорилирование может активировать или инактивировать ферменты.Обращение фосфорилирования, называемое дефосфорилированием , завершается ферментом, называемым фосфатазой . Дефосфорилирование обратит эффект, который фосфорилирование оказало на его субстрат.
Рисунок 11-4: Молекулярный переключатель киназы / фосфатазы. Киназы добавляют фосфатную группу (PO 4 –3 ) ко многим субстратам, включая белки, участвующие в передаче сигналов клетки. Фосфорилирование может изменять функцию белка, изменяя его ферментативную активность, расположение в клетке и / или форму.Эти функциональные изменения могут быть отменены ферментами, называемыми фосфатазами, которые удаляют фосфатные группы.
GTPases гидролизуют связанный GTP до GDP, где молекула GDP остается связанной с самой GTPase. Хотя GTPase может спонтанно гидролизовать GTP, белок, активирующий GTPase, GAP значительно ускоряет скорость гидролиза. Когда GDP связан, белок неактивен. Чтобы система вернулась к GTP и активировала белок, GDP не подвергается повторному фосфорилированию: скорее, GDP должен быть удален и заменен на новый GTP.Обмену в значительной степени способствует действие вспомогательного белка, фактора обмена гуаниновых нуклеотидов (GEF) (рис. 11-5).
Рисунок 11-5: Молекулярный переключатель GTPase. Активные GTPases связаны с GTP. ГТФазы быстро гидролизуют ГТФ до ВВП в присутствии соответствующих им белков GAP, делая ГТФазу неактивной. GTPases становятся активными, когда белок GEF обменивает связанный GDP на GTP.
Конечной точкой внутриклеточных каскадов является активация эффекторов .Эффекторы — это белки, которые вызывают клеточные ответы на сигнальные молекулы. Например, если клеточный ответ на конкретную сигнальную молекулу представляет собой изменение формы, эффекторные белки будут взаимодействовать с ферментами, которые реконструируют цитоскелет. Точно так же, если клеточный ответ на сигнальную молекулу представляет собой изменение экспрессии генов, факторы транскрипции, модулирующие экспрессию этих генов, будут эффекторами (рис. 11-3).
Эффекторы контролируют как быстрые (от секунды до нескольких минут), так и медленные (от нескольких минут до часов) клеточные ответы на сигнальные молекулы.Клетки могут быстро изменять свой метаболизм и форму. Следовательно, эффекторы, участвующие в быстрых клеточных ответах, обычно представляют собой метаболические ферменты или ферменты, регулирующие цитоскелет. Поскольку процессы транскрипции и трансляции требуют времени, клеточные ответы, которые включают изменения в экспрессии генов (то есть, когда эффекторы являются факторами транскрипции), считаются медленными ответами (рис. 11-3).
Рецепторы — это белковые молекулы в клетке-мишени или на ее поверхности, которые связывают сигнальные молекулы.Есть два типа рецепторов: внутренние рецепторы и рецепторы на поверхности клетки.
Внутренние рецепторы
Внутренние рецепторы , также известные как внутриклеточные или цитоплазматические рецепторы, обнаруживаются в цитоплазме клетки и отвечают на молекулы гидрофобного лиганда, которые могут перемещаться через плазматическую мембрану. Попав внутрь клетки, многие из этих молекул связываются с белками, которые действуют как регуляторы синтеза (транскрипции) мРНК, опосредуя экспрессию генов. Экспрессия генов — это клеточный процесс преобразования информации клеточной ДНК в последовательность аминокислот, которая в конечном итоге формирует белок.Когда лиганд связывается с внутренним рецептором, запускается конформационное изменение, которое открывает сайт связывания ДНК на белке. Комплекс лиганд-рецептор перемещается в ядро, затем связывается со специфическими регуляторными областями хромосомной ДНК и способствует инициации транскрипции (рис. 11-6). Внутренние рецепторы могут напрямую влиять на экспрессию генов, не передавая сигнал другим рецепторам или мессенджерам.
Рисунок 11-6: Внутриклеточные рецепторы .Внутриклеточные рецепторы связывают сигнальные молекулы, которые могут диффундировать через плазматическую мембрану. Многие внутриклеточные рецепторы представляют собой факторы транскрипции, которые связываются с промоторами, чтобы изменить экспрессию генов при связывании с соответствующим лигандом.
Сигнализация стероидного гормона
Небольшие гидрофобные лиганды могут напрямую диффундировать через плазматическую мембрану и взаимодействовать с внутренними рецепторами. Важными членами этого класса лигандов являются стероидные гормоны. Стероиды — это липиды, которые имеют углеводородный скелет с четырьмя конденсированными кольцами; разные стероиды имеют разные функциональные группы, прикрепленные к углеродному скелету (рис. 11-7).Стероидные гормоны включают женский половой гормон эстрадиол, который является разновидностью эстрогена; мужской половой гормон тестостерон; и холестерин, который является важным структурным компонентом биологических мембран и предшественником стероидных гормонов. Другие гидрофобные гормоны включают гормоны щитовидной железы и витамин D. Чтобы быть растворимыми в крови, гидрофобные лиганды должны связываться с белками-носителями, пока они транспортируются через кровоток.
Рисунок 11-7: Стероидные гормоны. Эстрадиол и тестостерон являются примерами стероидных гормонов. Они являются метаболитами холестерина и поэтому могут диффундировать через плазматическую мембрану для связывания с внутриклеточными рецепторами.
Сигнализация оксида азота
Оксид азота (NO) — это газ, который может диффундировать непосредственно через плазматическую мембрану. Благодаря этому свойству NO, как и стероидные гормоны, может связываться с внутриклеточным рецептором. NO играет важную роль в регулировании кровотока, расслабляя гладкие мышцы вокруг кровеносных сосудов (что приводит к увеличению кровотока).В ответ на нейрональную передачу сигналов NO высвобождается из эндотелиальных клеток и связывается со своим внутриклеточным рецептором, гуанилилциклазой (GC) , в соседних клетках гладких мышц. GC — рецептор с ферментативной активностью. Связываясь со своим лигандом NO, GC метаболизирует гуанозинтрифосфат (GTP) в циклический монофосфат гуанозина (cGMP) (рис. 11-8). цГМФ является вторичным мессенджером, потому что это небольшая молекула, которая усиливает ответ. Ферменты, называемые фосфодиэстеразами (PDE), превращают цГМФ в гуанозинмонофосфат (GMP).
Рисунок 11-8: Гуанилилциклаза и цГМФ . Гуанилилциклаза (GC) — это внутриклеточный рецептор оксида азота (NO) и фермент. Когда NO связан с GC, GC может превращать гуанозинтрифосфат (GTP) в циклический гуанозинмонофосфат (cGMP). cGMP является необходимым вторичным мессенджером в сигнальном пути NO. цГМФ может расщепляться на гуанозинмонофосфат (GMP) ферментами, называемыми фосфодиэстеразами (PDE).
Когда уровни вторичного мессенджера цГМФ повышаются, молекулы цГМФ связываются и активируют внутриклеточную сигнальную молекулу , протеинкиназу А (PKA) .Активированная PKA фосфорилирует многочисленные эффекторные белки, которые в совокупности расслабляют гладкие мышцы и увеличивают кровоток (рис. 11-9).
NO имеет очень короткий период полураспада и, следовательно, действует только на коротких расстояниях. Нитроглицерин, средство для лечения сердечных заболеваний, запускает высвобождение NO, что вызывает расширение (расширение) кровеносных сосудов, тем самым восстанавливая приток крови к сердцу. В последнее время NO стал более известен, потому что лекарства, отпускаемые по рецепту, нацелены на путь, на который он влияет на эректильную дисфункцию, например, виагра (при эрекции задействованы расширенные кровеносные сосуды) (рис. 11-9).
Рисунок 11-9: Сигнализация ячейки оксида азота . В ответ на нейрональную передачу сигналов эндотелиальные клетки, которые обвивают кровеносные сосуды, производят небольшой газ оксид азота (NO). NO диффундирует из эндотелиальных клеток в соседние гладкомышечные клетки. Внутри гладкомышечных клеток NO связывается со своим внутриклеточным рецептором, гуанилилциклазой (GC). В активном состоянии GC метаболизирует GTP во вторичный мессенджер cGMP. Повышение уровней цГМФ привело к активации эффекторных белков, таких как киназа легкой цепи миозина, что привело к расслаблению мышц.Когда сглаживающие мышцы вокруг кровеносных сосудов расслабляются, кровоток увеличивается. Лекарство от эректильной дисфункции Виагра нацелено на фосфодиэфиры (PDE), ферменты, которые расщепляют цГМФ до GMP. Ингибируя распад вторичного мессенджера цГМФ, Виагра поддерживает высокие клеточные уровни цГМФ, таким образом поддерживая расслабление гладких мышц, усиление кровотока и длительную эрекцию.
Рецепторы клеточной поверхности
Рецепторы клеточной поверхности , также известные как трансмембранные рецепторы или внеклеточные рецепторы, представляют собой заякоренные (интегральные) белки клеточной поверхности, которые связываются с молекулами внешнего лиганда.Этот тип рецептора охватывает плазматическую мембрану и выполняет сигнальную трансдукцию, при которой внеклеточный сигнал преобразуется в межклеточный сигнал. Лиганды, которые взаимодействуют с рецепторами клеточной поверхности, не должны проникать в клетку, на которую они воздействуют. Рецепторы клеточной поверхности также называют клеточно-специфическими белками или маркерами, поскольку они специфичны для отдельных типов клеток.
Поскольку белки рецепторов клеточной поверхности имеют фундаментальное значение для нормального функционирования клеток, неудивительно, что нарушение работы любого из этих белков может иметь серьезные последствия.Было показано, что ошибки в белковых структурах некоторых рецепторных молекул играют роль в гипертонии (повышенном кровяном давлении), астме, сердечных заболеваниях и раке.
Каждый рецептор клеточной поверхности состоит из трех основных компонентов: внешнего лиганд-связывающего домена, гидрофобного участка, охватывающего мембрану, и внутриклеточного домена внутри клетки. Лиганд-связывающий домен также называют внеклеточным доменом. Размер и степень каждого из этих доменов широко варьируются в зависимости от типа рецептора.
Управляемые лигандами ионных канала связывают сигнальные молекулы и открывают канал через мембрану, который позволяет специфическим ионам проходить через них. Чтобы сформировать канал, этот тип рецептора клеточной поверхности имеет обширную область, охватывающую мембрану (рис. 11-10). Чтобы взаимодействовать с хвостами фосфолипидных жирных кислот, которые образуют центр плазматической мембраны, многие аминокислоты в области, охватывающей мембрану, являются гидрофобными. И наоборот, аминокислоты, выстилающие внутреннюю часть канала, являются гидрофильными, что позволяет проходить полярным молекулам, таким как вода или ионы.Когда лиганд связывается с внеклеточной областью канала, происходит конформационное изменение в структуре белка, которое позволяет проходить ионам, таким как натрий, кальций, магний и водород. рецепторы, сопряженные с G-белком (GPCR) и рецепторные тирозинкиназы (RTK), два дополнительных типа рецепторов на поверхности клетки, более подробно рассматриваются в следующей главе.
Рис. 11-10. Ионные каналы . Поверхность клетки Ионные каналы, управляемые лигандами, открываются, когда они связываются со специфическими сигнальными молекулами.После открытия ионы втекают в ячейку или выходят из нее, в зависимости от конкретного канала.
Связанные видео:
Клеточная коммуникация — Принципы биологии
Цели обучения
Результаты курса для этого раздела:
Объясните, как основные элементы клеточной структуры определяют функции всех живых существ.
г. Объясните, как структура клеточных мембран приводит к ее различным функциям, включая избирательную проницаемость и транспорт, а также передачу клеточных сигналов.
Представьте, какой была бы жизнь, если бы вы и окружающие не могли общаться. Вы не сможете выразить свои пожелания другим или задать вопросы, чтобы узнать больше о вашем окружении. Социальная организация зависит от общения между людьми, составляющими это общество; без общения общество развалится.
Как и в случае с людьми, для отдельных клеток жизненно важно иметь возможность взаимодействовать с окружающей средой. Это верно независимо от того, растет ли клетка сама по себе в пруду или является одной из многих клеток, образующих более крупный организм.Чтобы правильно реагировать на внешние раздражители, клетки разработали сложные механизмы коммуникации, которые могут принимать сообщение, передавать информацию через плазматическую мембрану, а затем производить изменения внутри клетки в ответ на сообщение.
В многоклеточных организмах клетки постоянно отправляют и получают химические сообщения, чтобы координировать действия отдаленных органов, тканей и клеток. Возможность быстро и эффективно отправлять сообщения позволяет ячейкам координировать и настраивать свои функции.
Хотя необходимость клеточной коммуникации у более крупных организмов кажется очевидной, даже одноклеточные организмы общаются друг с другом. Клетки дрожжей сигнализируют друг другу, чтобы способствовать спариванию. Некоторые формы бактерий координируют свои действия, чтобы сформировать большие комплексы, называемые биопленками, или организовать производство токсинов для удаления конкурирующих организмов. Способность клеток общаться с помощью химических сигналов возникла в отдельных клетках и была необходима для эволюции многоклеточных организмов.Эффективное и безошибочное функционирование систем связи жизненно важно для всей жизни, какой мы ее знаем.
OpenStax, Биология. OpenStax CNX. 13 октября 2017 г. https://cnx.org/contents/[email protected]:1e9l33C7@2/Introduction
как работает мобильная сеть?
Мобильная связь следует общему принципу телефонии: соединение двух удаленных пользователей через сетевое оборудование оператора, ответственного за управление услугой. Но в отличие от стационарных телефонов, в мобильной сети не медные провода или оптоволокно, а радиопередача обеспечивает окончательное соединение.Мобильный телефон пользователя связывается по воздуху с антенной базовой станции, которая, в свою очередь, подключается к центральной станции оператора — компьютеру. Это направляет связь к соответствующему абоненту в фиксированной сети или через другие базовые станции.
Для связи мобильный пользователь должен находиться в пределах досягаемости базовых станций. Он имеет ограниченный диапазон и охватывает только небольшую область вокруг себя, называемую «ячейкой» (отсюда и альтернативное название «сотовые сети», часто используемое для мобильных сетей).Чтобы покрыть максимальную территорию и гарантировать, что пользователи всегда могут звонить, операторы развертывают тысячи ячеек, каждая из которых оборудована антеннами, обеспечивая перекрытие их ячеек и, таким образом, никогда не теряйте текущее местоположение пользователей.
городские соты, сельские соты
Размер соты зависит от многих факторов, таких как тип используемых базовых станций, рельеф (равнины, горы, долины и т. Д.), Место установки (сельские или городские районы) и численность населения плотность. Размер ячейки также ограничен радиусом действия мобильного телефона, который должен иметь возможность делать обратную ссылку.
И, что важно, базовые станции имеют ограниченную пропускную способность и могут обрабатывать только определенное количество одновременных вызовов. По этой причине в городских районах, где плотность населения высока и имеется значительное количество коммуникаций, ячейки, как правило, бывают многочисленными и небольшими — сотни или даже десятки метров друг от друга. В сельской местности, где плотность населения намного ниже, размер ячейки намного больше, иногда до нескольких километров, но редко превышает десять километров.
Важно подчеркнуть, что уменьшение мощности сигнала, излучаемого базовыми станциями, в свою очередь, уменьшает покрытие сот. Таким образом, улучшение способности сети передавать голосовые вызовы или трафик данных обязательно требует увеличения количества базовых станций.
Основы проектирования сотовых систем
Несмотря на возрастающую сложность стандартов и устройств беспроводной связи, сотовые технологии поддерживают набор общих принципов, которые составляют основу разработки сотовых систем.
Основы радиосвязи для сотовых сетей : Сотовые сети позволяют таким устройствам, как смартфоны и устройства Интернета вещей (IoT), обмениваться данными по беспроводной сети. Сотовые технологии продвинулись от аналоговых технологий первого поколения (1G) до передовых высокопроизводительных систем четвертого (4G) и пятого поколений (5G) всего за четыре десятилетия.
На протяжении развития каждого поколения беспроводной связи эти сотовые сети разделяли ряд общих основных атрибутов.Многие из обменов связью на основе протоколов между устройством и базовой станцией следуют аналогичной философии идентификации потенциальной соты, регистрации и аутентификации в базовой сети и поддержки мобильности посредством передачи сигналов. Эти принципы, скорее всего, будут включены в системы 6G, каким бы ни был этот стандарт в будущем. Конечно, реализация этих основополагающих принципов будет варьироваться от одного стандарта к другому, а иногда даже в пределах редакции данного стандарта.
Основы
Хотя точная сетевая архитектура отличается от поколения к поколению, типичная сотовая сеть состоит из сети радиодоступа (RAN), базовой сети (CN) и сервисной сети. RAN содержит базовые станции (BS), которые обмениваются данными с беспроводными устройствами с помощью радиочастотных (RF) сигналов, и это интерфейс между базовой станцией и устройствами. RAN выделяет радиоресурсы устройствам, чтобы сделать беспроводную связь реальностью.CN выполняет такие функции, как аутентификация пользователя, авторизация услуг, активация безопасности, выделение IP-адресов и установка подходящих каналов для облегчения передачи пользовательского трафика, такого как голос и видео. Сервисная сеть включает в себя серверы конкретного оператора и подсистему IP-мультимедиа (IMS), чтобы предлагать абонентам беспроводной связи различные услуги, включая голосовые вызовы, текстовые сообщения (SMS) и видеозвонки.
T передатчики
Первый общий принцип сотовых сетей — это использование множества передатчиков меньшей мощности (100 Вт или меньше) меньшего размера с более узкими зонами покрытия вместо одного мощного передатчика с более широкой зоной покрытия.Эти передатчики размещаются на базовых станциях, более известных как вышки сотовой связи. Базовые станции также содержат приемники и дополнительные блоки управления.
Ячейки
Зоны покрытия разделены на соты, каждая из которых обслуживается собственной антенной (передатчиком). Полоса частот назначается передатчику / приемнику в зависимости от поставщика услуг связи. Ячейки расположены так, что антенны в зоне покрытия имеют шестиугольную диаграмму направленности. Это связано с тем, что для представления шестиугольника требуется меньше ячеек по сравнению с треугольником или квадратом, а это означает, что сетевые операторы могут покрывать более широкую область с меньшим количеством базовых станций.Еще одно преимущество гексагональной сотовой системы состоит в том, что с помощью этой формы возможно повторное использование частоты.
Повторное использование частоты
Второй общий принцип построения сотовых сетей — повторное использование частот. Повторное использование частот — это процесс использования одних и тех же радиочастот на базовых станциях и других радиопередатчиках в пределах географической области. Эти сайты разделены достаточным расстоянием, чтобы создавать минимальные помехи друг другу. Используя географически небольшие ячейки с низким энергопотреблением, частоты могут повторно использоваться несмежными ячейками.
Причиной повторного использования частот является ограниченное количество выделенных несущих частот, установленных регулирующими органами.
Разделение ячеек
Разделение ячеек — это процесс разделения перегруженной ячейки на меньшие ячейки таким образом, чтобы каждая меньшая ячейка имела свою собственную базовую станцию. Эти небольшие соты оснащены антеннами с уменьшенной высотой и мощностью передатчика. Две меньшие соты увеличивают пропускную способность сотовой сети, поскольку увеличивается количество повторных использований каналов.
В популярной конфигурации сотовой сети одна базовая станция контролирует три географических региона, называемых секторами (или сотами), где каждый сектор покрывает область 120 °. Три сектора вместе обеспечивают покрытие 3 × 120 ° = 360 ° вокруг базовой станции.
Передача сотовой связи
Когда мобильное устройство перемещается в заданной области, оно пересекает границы ячеек. Передача обслуживания — это процесс, при котором выделенное радиосоединение между устройством и сетью радиодоступа переключается с одной ячейки на другую.Сотовая передача данных гарантирует, что устройство имеет выделенное радиосвязь с наилучшей возможной линией связи. Кроме того, передача обслуживания может использоваться для балансировки нагрузки между обслуживающими базовыми станциями и между несущими частотами, доступными в соте или секторе.
