Что такое потенциал человека. Какие бывают виды потенциалов личности. Как оценить и развить свой внутренний потенциал. На что влияет уровень развития различных потенциалов человека.
Понятие и сущность потенциала человека
Потенциал человека представляет собой совокупность его скрытых возможностей, способностей и ресурсов, которые могут быть реализованы при определенных условиях. Это внутренний резерв личности, который позволяет ей развиваться, достигать поставленных целей и самореализовываться в различных сферах жизни.
Ключевые характеристики потенциала человека:
- Является врожденным, но может развиваться на протяжении жизни
- Включает как реализованные, так и нереализованные возможности
- Проявляется в конкретных видах деятельности
- Зависит от внутренних качеств личности и внешних условий
- Имеет индивидуальный характер у каждого человека
Раскрытие и реализация потенциала позволяет человеку наиболее полно проявить свои таланты и способности, достичь успеха и самореализации. Поэтому понимание структуры собственного потенциала и работа над его развитием крайне важны для каждой личности.

Основные виды потенциалов человека
Выделяют следующие основные виды потенциалов человека:
1. Интеллектуальный потенциал
Включает умственные способности, интеллект, креативность, способность к обучению и освоению новых знаний. Влияет на успешность в образовании, профессиональной и творческой деятельности.
2. Творческий потенциал
Способность человека к созданию нового, оригинального в различных сферах деятельности. Проявляется в генерации идей, нестандартном мышлении, стремлении к самовыражению.
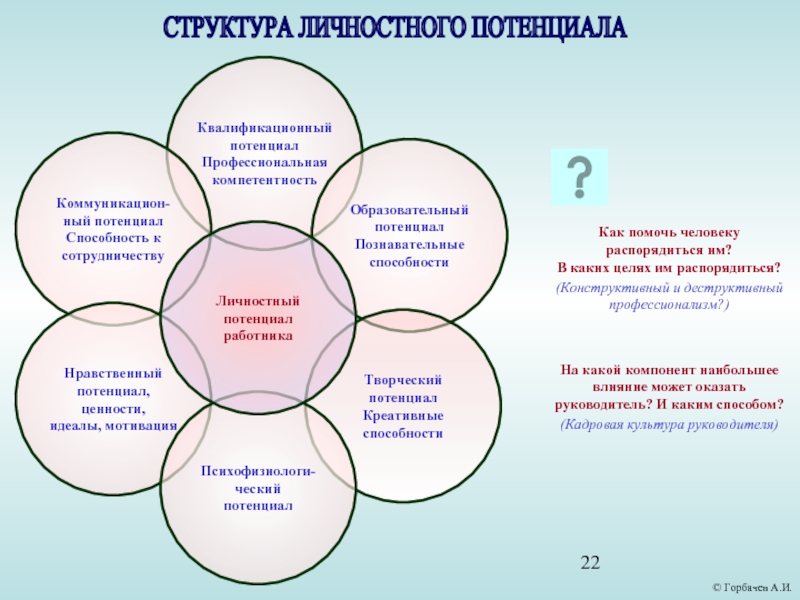
3. Личностный потенциал
Комплекс психологических свойств и качеств личности — воля, мотивация, ценности, убеждения. Определяет способность к саморазвитию и самореализации.
4. Физический потенциал
Возможности организма человека, его физическое здоровье, выносливость, работоспособность. Влияет на активность и продуктивность во всех сферах жизни.
5. Коммуникативный потенциал
Способности в сфере общения и взаимодействия с другими людьми. Включает навыки коммуникации, эмпатию, умение работать в команде.

Оценка внутреннего потенциала человека
Для оценки собственного потенциала можно использовать следующие методы:
- Психологическое тестирование для выявления способностей и личностных качеств
- Анализ достижений и успехов в различных сферах жизни
- Оценка уровня знаний, умений и навыков
- Изучение интересов, увлечений и предпочтений
- Обратная связь от окружающих людей
- Самоанализ и рефлексия
Важно оценивать не только актуальный уровень развития потенциала, но и перспективы его раскрытия. Это позволит определить направления для дальнейшего личностного роста и развития.
Способы развития потенциала личности
Основные способы развития и реализации потенциала человека включают:
- Постоянное обучение и получение новых знаний
- Практическое применение способностей в деятельности
- Постановка амбициозных целей
- Выход из зоны комфорта, преодоление трудностей
- Творческая самореализация
- Развитие необходимых личностных качеств
- Расширение круга общения
- Поддержание физического и психического здоровья
При этом важно развивать потенциал комплексно, уделяя внимание всем его аспектам. Только гармоничное развитие различных видов потенциала позволит достичь максимальной самореализации.

Влияние потенциала на жизнь и успех человека
Уровень развития потенциала оказывает значительное влияние на различные аспекты жизни человека:
- Успешность в профессиональной деятельности
- Возможности карьерного роста
- Материальное благосостояние
- Качество отношений с окружающими
- Удовлетворенность жизнью
- Способность справляться со стрессом и трудностями
- Творческая самореализация
- Достижение поставленных целей
Люди с высоким уровнем развития потенциала, как правило, более успешны, уверены в себе и удовлетворены жизнью. Они легче адаптируются к изменениям и находят возможности для роста в любых ситуациях.
Роль окружения в раскрытии потенциала человека
Окружение играет важную роль в раскрытии и реализации потенциала личности. Оно может как способствовать развитию способностей, так и препятствовать ему.
Факторы окружения, влияющие на раскрытие потенциала:
- Семья и воспитание
- Образовательная среда
- Профессиональное окружение
- Круг общения
- Социально-экономические условия
- Культурный контекст
Благоприятная среда, поддержка близких людей, наличие возможностей для развития способствуют более полному раскрытию потенциала человека. При этом даже в неблагоприятных условиях сильная личность способна преодолеть ограничения и реализовать свой потенциал.
Ограничения в развитии потенциала и пути их преодоления
Существует ряд факторов, которые могут ограничивать развитие потенциала человека:
- Неуверенность в себе, заниженная самооценка
- Страх неудачи, нежелание выходить из зоны комфорта
- Отсутствие четких целей и мотивации
- Негативное влияние окружения
- Недостаток знаний и навыков
- Проблемы со здоровьем
Для преодоления этих ограничений важно:
- Работать над повышением самооценки и уверенности в себе
- Ставить конкретные цели и планировать их достижение
- Развивать необходимые навыки и получать новые знания
- Окружать себя людьми, которые поддерживают ваше развитие
- Заботиться о физическом и психическом здоровье
- Учиться на ошибках и не бояться трудностей
Осознание собственных ограничений и целенаправленная работа над их преодолением позволяют существенно расширить возможности для реализации потенциала.
Взаимосвязь различных видов потенциала человека
Все виды потенциала человека тесно взаимосвязаны и влияют друг на друга. Развитие одного вида потенциала часто способствует росту других. Например:

- Развитие интеллектуального потенциала повышает творческие способности
- Укрепление физического здоровья положительно влияет на умственную активность
- Развитие коммуникативных навыков расширяет возможности для самореализации
- Личностный рост способствует раскрытию всех видов потенциала
Поэтому для максимальной реализации заложенных в человеке возможностей важно развивать потенциал комплексно, уделяя внимание всем его аспектам. Такой подход позволяет достичь синергетического эффекта и наиболее полно раскрыть свои таланты и способности.
Потенциал человека и его скрытые способности
«Если вы можете это вообразить, вы можете это и сделать» .
Уолт Дисней
Определение внутреннего потенциала человека – это не самая простая задача. Первое, что необходимо сделать, так это понять, что потенциал – это способность человека реализовать собственные скрытые возможности. У каждого человека внутренний потенциал разный, как и каждый человек индивидуален. Потенциал представляет собой здоровую основу для самореализации в мире и обществе, и является внутренним резервом. В мире, где главенствует левое полушарие, успеха добиваются те, кто умеет использовать как левое(логическое) , так и правое (творческое) полушарие мозга.
Потенциал личности складывается в целостную картину из множества других потенциалов. Именно их уровень развития определяет, насколько человек будет здоров психологически и успешен, счастлив. В целом, можно выделить следующие виды потенциалов человека:
- Потенциал разума. Его можно охарактеризовать, как способность человека совершенствовать собственное мышление. Когда приходится решать различного рода жизненные задачи, ум становится подвижным, задействуется реалистичное, творческое, системное, дивергентное мышление. Однако человек с неразвитым потенциалом ума при возникновении проблем часто впадает в депрессию и у него могут возникать ощущения усталости от жизни.
- Потенциал воли. Он отражает то, насколько человек способен к самореализации, ставить перед собой цели и какие адекватные пути их достижения он выберет. Как утверждают ученые, в случаях, когда потенциал воли развит слабо, у человека возникают проблемы с психикой (чаще всего неврозы). Однако когда удается взрастить в себе волю, то людям становится проще жить, им удается определить свое место в жизни, создать личную систему ценностей и выстраивать стратегию, которая помогает добиваться поставленных целей.
- Потенциал чувств. Он характеризует то, насколько эмоциональная сфера человека богата и насколько адекватно он умеет выражать чувства, а также понимать эмоции других людей. От реализации этого потенциала зависит, насколько человек ощущает себя счастливым. В случае если потенциал чувств не был реализован, возникает ряд психосоматических расстройств. Профилактикой от различных психических болезней и психологических расстройств является умение позитивно смотреть на жизнь.
- Потенциал тела. Каждый человек должен понимать, что его тело – это часть его личности. Поэтому умение совершенствовать его и прислушиваться к нему является крайне важной частью в развитии потенциала в целом.
- Общественный потенциал. От уровня его развития зависит то, насколько человеку легко дается налаживание контактов с окружающими его людьми, развитие коммуникативной культуры и адаптация в социальных условиях.
- Креативный (творческий) потенциал. Он определяет, насколько человек способен к активности, самовыражению, преобразованию мира. Созидательная деятельность и реализация этого потенциала – основа психологической зрелости.
- Духовный потенциал. Как правило, он определяет стиль жизнедеятельности человека и то, как он развивает в себе духовную природу.
Все, перечисленное выше, является основой психологической устойчивости человека. Если какой-либо из потенциалов развит слишком слабо, возможен дисбаланс, который приводит к внутренним конфликтам.
Оценка внутренних резервов
Предполагается , что навыки родителей передаются ребенку в момент зачатия. Таким образом, можно утверждать, что многое в человеке зависит от его генетической предрасположенности. Поэтому, определяя потенциал кого-либо, следует изучить качества и способности его родителей. Именно таким образом и формируется природный потенциал – та основа, которая лежит в психологической составляющей здоровья человека.
Второй аспект оценки внутренних резервов человека заключается в определении его способностей к концентрации. Стоит отметить, что эта способность играет большую роль, так как именно она позволяет человеку решать сложные жизненные задачи. Умение сконцентрироваться, позволяет развивать и такие качества, как устойчивость в стрессовых ситуациях и способность выживать в сложных условиях.
При оценке потенциала личности, необходимо учитывать и то, насколько человек коммуникабелен, а также как легко ему удается располагать людей к себе и влиять на них. При этом следует понимать, что оценивать нужно только то общение, которое имеет позитивный характер (то есть нормальное, непринужденное и бесконфликтное). Обращают внимание и на то, умеет ли находить человек компромиссы и «сглаживать углы». Этот момент важен при оценке потенциала тем, что он дает возможность понять, насколько гармонична жизнь человека и определить, насколько легко ему будет добиться поставленных целей. Помимо перечисленного выше, при оценке потенциала необходимо учитывать и удачливость человека, его отношение к неудачам, привычки, увлечении и хобби.
Развитие потенциала
Следует учитывать, что те внутренние резервы, которые были заложены природой, можно не только укреплять, но и развивать. Иногда этот процесс происходит сам собой, под влиянием сложных жизненных обстоятельств. Раскрыть потенциал – это задача, которая состоит из двух составляющих – понять, что было упущено, и выстроить план дальнейших действий. К примеру, девушка в детстве имела способность к рисованию, но жизненные обстоятельства заставили ее забыть об этом увлечении. Она поступила в ВУЗ, получила образование по финансовой специальности и устроилась бухгалтером. Со временем ей стало тяжело, и она осознала, что та работа, которой она занимается, не приносит ей удовольствия. Она собрала последние силы, и начала по вечерам рисовать картины. Удовольствие от этого процесса начали перекрывать все негативное, что происходит вокруг. Это произошло под влиянием того, что девушка дала волю потенциалу, находящемуся «под замком» отсутствия времени и других дел. Развивайте в себе способности смотреть на мир по новому, возможно, не каждый угол обзора будет полезен, но этот навык в итоге окрасит вашу жизнь новыми красками. Мы уже много знаем про работу сознания, и точно можно сказать: сначала появляется идея в творческом полушарии, а потом логическое левое воплащает ее в жизнь.
Когда нужно заняться раскрытием внутренних резервов
Существует несколько признаков того, что человеку крайне необходимо заняться раскрытием собственного потенциала. К ним относятся следующие:
- нехватка энергии на то, чтобы что-то предпринять для изменения своей жизни в лучшую сторону;
- неполадки дома, на работе или в отношениях с близкими;
- возникновение мыслей, что жизнь проходит мимо.
Это все признаки того, что тот внутренний резерв, который был в человеке, исчерпал себя. В такие моменты следует найти источники и начать развивать свой потенциал, для того чтобы исправить ситуацию. Следует начать читать различную духовную литературу (это могут быть книги по психологии или эзотерике, учения философов.) Нужно находить время на медитацию. Необходимо начать вести личный дневник , следует вспомнить свои прошлые успехи.
Внешние источники
Источники извне – это обстоятельства, люди и ситуации, которые заставят понять, чего не хватает для раскрытия собственного потенциала. Это может быть общение с психологом, тренером или духовным наставников, посещение различных тренингов. Стоит отметить, что хорошо помогает общение с детьми и животными. Помимо этого, личностный потенциал хорошо развивается через помощь нуждающимся.
Если человеку удалось предпринять хотя бы первые шаги для развития личного потенциала, ему становится намного проще и легче жить дальше. Всегда стоит помнить, что никогда не поздно приступить к поискам внутренних резервов. Этот момент очень важен для того, чтобы постоянно улучшать качество своей жизни.
УЗ «4-я городская поликлиника» валеолог Наталья Дударчик
содержание и классификация – тема научной статьи по экономике и бизнесу читайте бесплатно текст научно-исследовательской работы в электронной библиотеке КиберЛенинка
В условиях рыночной экономики знание производственных, финансовых и иных возможностей предприятия необходимо по многим причинам, например, для разработки его стратегии, оценки стоимости, качества менеджмента, устойчивости развития в кризисной ситуации. Для характеристики возможностей предприятия в разных сферах деятельности используется понятие «потенциал», содержание которого раскрывается в энциклопедических изданиях. Потенциал — это « …средства, запасы, источники, имеющиеся в наличии и могущие быть мобилизованы, приведены в действие, использованы для достижения определенных целей, осуществления плана, решения какой-либо задачи; возможности отдельного лица, общества, государства в определенной области» [1, т.20, с.428].
Необходимо подчеркнуть, что речь здесь, очевидно, идет ресурсах, которые уже имеются у предприятия и которые определяют его возможности. Следовательно, в понятие потенциала не включены те средства, которые могут быть привлечены в будущем.
Отметим, далее, что в экономической и финансовой литературе рассматриваются различные виды потенциала: экологический, экономический, финансовый, рыночный, производственный, трудовой, кадровый, интеллектуальный, научно-технический, управленческий, организационный, инновационный, инвестиционный, информационный и др. [3,7,8]. Однако при этом, как правило, не указываются классификационные признаки их группировки, не определяются взаимосвязи и соподчиненность этих понятий. Иными словами, в настоящее время отсутствует системный подход к классификации видов потенциала, что затрудняет определение их места и роли.
Систематизация различных подходов к группировке видов потенциала требует использования, прежде всего, такого признака классификации, как уровень иерархии. В зависимости от него следует выделять потенциал страны, региона, населения, корпорации или предприятия в ее составе, которые взаимосвязаны между собой как часть и целое.
Далее, в зависимости от сферы, к которой относится потенциал, можно рассматривать такие его виды, как экологический, оборонный, трудовой, интеллектуальный, научный и другие. Очевидно, что эти и подобные им виды потенциала относятся в большей степени к потенциалу страны или иной территории, так как характеризуют, прежде всего, ресурсы и возможности внешней среды
хозяйствующего субъекта, а не самого предприятия.
Используя системный подход, из всей совокупности видов потенциала предприятия следует, по нашему мнению, выделить первоначально общие его виды, свойственные любому хозяйствующему субъекту: ресурсный, управленческий, инновационный, информационный потенциал. Это — первый уровень в структуре экономического потенциала предприятия. Очевидно, что, не располагая
соответствующей ресурсной базой или всей необходимой информацией о рынках, конкурентах и т.п., любое предприятие не может эффективно функционировать, равно как и при отсутствии — в условиях конкурентного рынка — адекватной системы управления или инновационной деятельности.
Первостепенное значение здесь имеет ресурсный потенциал, т.к. без финансовых или материальных ресурсов предприятие вообще не может начать свою деятельность. Именно по этой причине учредители предприятия сначала формируют его уставный капитал.
Далее, на втором уровне, следует выделить специфические виды потенциала -производственный, рыночный и финансовый, — соответствующие особым видам деятельности предприятия: производственной, коммерческой и финансовой, а также основным стадиям воспроизводственного цикла. Эти виды потенциала базируются на специфических ресурсах,
которыми располагает предприятие — производственных и финансовых ресурсах, ресурсах рынка.
В силу особенностей деятельности предприятий различных отраслей эти виды потенциала имеют свою специфику, отражающую особенности отраслевой принадлежности используемых ресурсов. Это касается, прежде всего, производственного и рыночного потенциалов, хотя и структура финансового потенциала в разных отраслях может значительно отличаться, например, большим удельным весом оборотных средств (легкая, пищевая промышленность) или денежных средств и товарных запасов (например, торговля).
Отметим, далее, что производственный и другие специфические виды потенциала, в свою очередь, могут быть разделены на частные виды. Это — третий уровень в системе экономического потенциала предприятия.
Так, в рамках производственного потенциала могут быть выделены такие его частные виды, как технико-технологический потенциал, материальный, кадровый, инвестиционный и т.п.
Что касается инвестиционного потенциала, то он может быть включен в производственный потенциал только в той части, в какой в него были направлены реальные инвестиции, предусматривавшие, например, реконструкцию или техническое перевооружение производства. После завершения последних возрастает, в первую очередь, величина производственного потенциала, а затем — на этой основе — могут увеличиться рыночный и финансовый потенциалы. Если же часть инвестиций предприятие направляет в финансовые активы (приобретение акций и
облигаций, выпущенных другими эмитентами), то это влечет увеличение размера его финансового потенциала за счет финансовых вложений.
Выделение трех уровней в структуре экономического потенциала предприятия (общих, специфических и частных видов потенциала) не исключает, а, напротив, предполагает возможность дальнейшей их дифференциации и интеграции друг с другом. Иными словами, экономический потенциал как совокупность различных видов потенциала является открытой системой.
Например, в рамках технико-технологического потенциала могут быть выделены технический и технологический потенциалы, характеризующие возможности соответственно техники и технологии. Инновационный потенциал предприятия, базирующийся на новейших результатах исследования и опытно-конструкторских разработках, может включать его научный потенциал. В свою очередь, научный потенциал, направленный на увеличение потенциала техники и технологии, формирует научно-технический потенциал предприятия, о котором упоминают многие авторы, например, Р.В. Окороков [6, с.23].
То же можно сказать и относительно других видов потенциала. Так, в литературе часто используют понятие организационного потенциала [5], который, «соединяя воедино» различные ресурсы, приводит к появлению нового качества этих ресурсов — приросту результата их использования, т.е. синергическому эффекту. По нашему мнению, понятие организационного потенциала входит в более широкое понятие «Управленческий потенциал». Сочетание возможностей организационного потенциала с другими специфическими видами потенциала формирует понятие организационно-производственного потенциала, который, в свою очередь, увеличивает возможности экономического потенциала как интегральной характеристики величины всех видов потенциала хозяйствующего субъекта.
Подводя итог изложенному выше, можно представить его содержание в виде трехуровневой схемы, систематизирующей иерархическую структуру экономического потенциала предприятия (рис. (П Пп, П|||, П|У). Рыночный потенциал является, на его взгляд, суммой этих ресурсов, обеспечивающих рыночную деятельность [9, с.42].
Из рис. 1 также видно, что финансовый потенциал предприятия напрямую связан с производственным и рыночным потенциалами и формируется его финансовыми ресурсами, являющимися составной частью ресурсного потенциала. Финансовый потенциал опосредованно связан с производственными, рыночными и другими ресурсами предприятия, а также с общими видами его потенциала, определяя тем самым возможности инновационной деятельности, управления, информатизации.
Однако, несмотря на первостепенную роль, которую играет финансовый потенциал в экономике предприятия, ему в литературе уделяется гораздо меньшее внимание по сравнению с другими видами потенциала — производственным, рыночным, инновационным, информационным. Содержание финансового потенциала трактуется разными авторами различным образом, до настоящего времени отсутствует общепринятая формулировка этого понятия.
Так, Е.П. Енина и С.В. Овсянников считают, что финансовый потенциал отражает возможность привлекать финансовые ресурсы определенного состава, стоимости и качества, а также эффективность их использования. Ресурсные же возможности в области финансов отражают совокупность денежных средств, которые используются в механизме финансирования и являются результатом формирования, распределения и использования денежных доходов [2, с.54-57].
Соглашаясь в принципе с таким подходом, следует отметить, однако, некоторую расплывчатость и неопределенность этой формулировки.
В то же время, по мнению В.В. Ковалева, финансовый потенциал предприятия — это характеристика его финансового положения и финансовых возможностей. Количественно он характеризуется показателями ликвидности, платежеспособности, финансовой устойчивости, внутрифирменной эффективности, прибыльности, рентабельности и инвестиционной привлекательности. В совокупности с имущественным потенциалом он отражает экономический потенциал предприятия [4, с.3б8].
Признавая, что финансовый потенциал предприятия — это, действительно, характеристика его финансовых возможностей, заметим, что он в то же время не может характеризовать финансовое положение предприятия, т.к. последнее отражает лишь использование имеющихся возможностей. К
тому же относительные показатели (ликвидности, финансовой устойчивости и т.п.) не могут отражать абсолютную величину финансового потенциала, т.е. давать ему количественную оценку.
Экономический потенциал предприятия
Г
I уровень (общие виды потенциала)
Инновационный
потенциал
V.
Управленческий
потенциал
Информационный
потенциал
Инновационные ресурсы Ресурсы управления Информационные ресурсы
Ресурсный потенциал
Производственные ресурсы Ресурсы рынка Финансовые ресурсы
III уровень (частные виды потенциала)
материальный
потенциал
кадровый
потенциал
потенциал
реальных
инвестиций
потенциал трудовых ресурсов, связанных с
маркетингом
потенциал материальных ресурсов рынка
потенциал
рыночных
информационных
ресурсов
II уровень Производственный Рыночный Финансовый
(специфические ^ потенциал потенциал потенциал
виды потенциала)
Г технико- методический потенциал
—► технологический (инструменталь ^ собственных
потенциал ный) потенциал финансовых
маркетинга ресурсов
1
потенциал
финансовых
инвестиций
потенциал заемных и привлеченных финансовых ресурсов
Частные виды рыночного потенциала приведены согласно группировке Е.В. Попова [9]
Рисунок 1. Трехуровневая схема потенциала предприятия
Т.Н. Толстых и Е.М. Уланова не совсем удачно, по нашему мнению, формулируют понятие финансового потенциала предприятия как обеспеченность его финансовыми ресурсами, необходимыми для нормального функционирования, а также целесообразность их размещения и эффективность использования [11].
Следует подчеркнуть при этом, что финансовый потенциал предприятия — это абсолютная величина его возможностей, тогда как обеспеченность, по смысловому значению этого слова, является относительной характеристикой степени использования этих возможностей. Кроме того, как справедливо отмечено в одной из статей [3], здесь имеет место смешение потенциала как совокупных возможностей предприятия с его фактическим состоянием и финансовой устойчивостью.
Таким образом, анализ имеющихся точек зрения позволяет сделать вывод, что сложившаяся противоречивая ситуация требует уточнения понятия «Финансовый потенциал» применительно к предприятию. В связи с этим необходимо сделать несколько предварительных замечаний.
Прежде всего, рассматривая содержание и понятие «потенциал», следует разграничивать, с одной стороны, наличный (фактический) потенциал, определяемый ресурсами, имеющимися в наличии у предприятия на данный момент времени (как собственными, так и привлеченными; с другой стороны — перспективный потенциал, который определяется не только имеющимися у предприятия ресурсами, но и теми средствами, которые оно может нарастить самостоятельно, а также привлечь со стороны в будущем.
Далее, говоря о потенциале предприятия с позиции его возможностей, необходимо иметь в виду, что в этом случае речь должна идти не о любых возможностях, а о максимальных, т.е. таких, которыми может располагать хозяйствующий субъект в данный момент при наиболее полном и наилучшем использовании имеющихся у него ресурсов. Именно такой подход в наибольшей мере отвечает смысловому содержанию термина «потенциал» и по этой причине используется сейчас при определении, например, производственного потенциала предприятия, т.е. его производственной мощности [12, с.459].
Важно подчеркнуть при этом, что термины «наиболее полное» и «наилучшее» имеют, по нашему мнению, различное смысловое содержание. Наиболее полное использование ресурсов означает, что в деятельности предприятия нашли применение все имеющиеся ресурсы, и каждый ресурс используется с наименьшим остатком или без остатка. Наилучшее использование ресурсов предполагает достижение наибольшего результата при использовании ресурсов наиболее рациональным из имеющихся способов, без совершенствования последних, т.е. за счет лучшей организации труда и производства.
С учетом вышеизложенного можно сформулировать понятие «финансовый потенциал» применительно к предприятию. Финансовый потенциал предприятия — это максимально возможная стоимостная величина финансовых ресурсов, находящихся в его распоряжении, при условии наиболее полного и наилучшего использования всего ресурсного потенциала предприятия.
В отличие от известных формулировок предлагаемое понятие финансового потенциала предприятия четко указывает критерий этого потенциала — максимально возможная и количественно измеряемая в стоимостном выражении величина имеющихся в данный момент на предприятии финансовых ресурсов, которую он может достичь при наилучшем и наиболее полном использовании всех ресурсов. Важно подчеркнуть, что последнее предполагает эффективно использование и других видов экономического потенциала предприятия.
Финансовые ресурсы предприятия как совокупность всех видов денежных средств, финансовых активов, находящихся в его распоряжении [10, с.38], включают в себя собственные и привлеченные (прежде всего, заемные) средства. Максимизация величины финансовых ресурсов предприятия означает также и одновременное достижение им максимального финансового результата, поскольку в состав этих ресурсов включается и полученная прибыль.
Несмотря на то, что финансовый потенциал является частью экономического потенциала, ему принадлежит особая роль в деятельности предприятия. С момента формирования финансовых ресурсов в процессе создания уставного капитала начинается функционирование любого предприятия. После приобретения на эти средства оборудования, материалов и производственных запасов, найма персонала развивается производственная и коммерческая деятельность, в результате которой произведённая продукция (услуги) реализуется на рынке. Из вырученных средств — после компенсации затрат — погашаются финансовые обязательства перед государством, кредиторами, формируются и пополняются различного рода фонды и резервы. Нераспределенная часть прибыли вместе с накопленной амортизацией служит финансовой основой для дальнейшего развития предприятия.
Таким образом, финансовые ресурсы и соответствующие денежные отношения опосредуют всю деятельность предприятия, им принадлежит первостепенная роль в формировании и наращивании производственного, рыночного, инновационного и других видов потенциала предприятия, что впоследствии ведет к дальнейшему увеличению самих финансовых ресурсов. Поэтому финансовый потенциал предприятия как максимально возможная величина его финансовых ресурсов определяет финансовые возможности развития предприятия, и в свою очередь, отражает результаты использования всех других видов ресурсов. По этой причине возможно, на наш взгляд, рассматривать финансовый потенциал предприятия как потенциал особого рода, имеющий двойственное содержание. С одной стороны, он в качестве ресурсной составляющей — входит в экономический потенциал предприятия (это его узкая трактовка). С другой стороны, он, отражая отношения по поводу формирования, распределения и использования денежных средств, опосредует движение всех видов ресурсов, формирующих экономический потенциал предприятия, в процессе производства и реализации товаров и услуг, определяет получение синергетического эффекта. Следовательно, в нем отражаются возможности и конечные результаты всей экономической деятельности предприятия. Это дает основания рассматривать финансовый потенциал как денежную форму экономического потенциала предприятия (это — широкая трактовка финансового потенциала).
Вызванные потенциалы | Официальный сайт Научного центра неврологии
Вызванные потенциалы (ВП) – метод выделения слабых и сверхслабых изменений электрической активности мозга в ответ на стимул различной модальности. Метод позволяет получить объективную информацию о состоянии периферических и центральных звеньев различных сенсорных систем таких как зрение, слух и т.д. Является неинвазивным и объективным методом тестирования функций ЦНС, для которого не имеется противопоказаний. Исследование ВП является неоценимым средством объективизации состояния сенсорных функций при различных неврологических заболеваниях, таких как острое нарушение мозгового кровообращения (ОНМК), опухоли головного и спинного мозга, последствия черепно-мозговой травмы, рассеянный склероз и другие демиелинизирующие заболевания, болезнь Паркинсона, наследстенные атаксии, хорея Гентингтона, болезнь Вильсона-Коновалова (гепатолентикулярная дегененрация), хронические цереброваскулярные заболевания, комы, персистирующие вегетативные состояния, тазовая боль, расстройства мочеиспускания, невралгия тройничного нерва.В Научном центре неврологии проводится исследование вызванных потенциалов различных модальностей:
- зрительные ВП на шахматный реверсивный паттерн (чёрно-белый и цветной паттерн) и на светодиодную вспышку (позволяют оценить состояние зрительного анализатора на разных уровнях: зрительного нерва, зрительного тракта, коркового звена зрительного анализатора и их динамику при лечении и прогрессировании заболевания)
- акустические стволовые ВП (позволяют оценить функциональное состояние периферического и внутристволового звеньев слухового анализатора)
- миогенные вестибулярные ВП (позволяют оценить функцию вестибулярного анализатора)
- соматосенсорные ВП при стимуляции n.medianus, n.tibialis, n.pudendus (позволяют выявить нарушения проведения на периферическом, стволовом и корковом уровнях)
- когнитивные ВП (Р300) (позволяют объективизировать состояние когнитивных функций)
- вегетативные ВП (позволяют выявить нарушения в вегетативной нервной системе).
Вызванные потенциалы — Первый МГМУ им. Сеченова
Вызванные потенциалы (ВП) — это метод исследования нервной системы, основанный на регистрации электрических потенциалов мозга в ответ на какое-либо воздействие.
Обращаться по телефону
+7 (985) 876-18-22 (с 9:00 до 16:00)
Включает несколько методик:
| ЗВП – зрительные ВП на вспышку света или шахматный паттерн | – используются для исследования зрительных функций |
| СВПСМ – слуховые ВП ствола мозга на звуковые стимулы | – используются в диагностике поражений слухового нерва и ствола головного мозга |
| СССВП – соматосенсорные ВП на стимуляцию слабым электрическим током периферических нервов рук и ног | – используются для тестирования проводящих функций нервной системы от периферического нерва до коры головного мозга |
| ТВП – тригеминальные ВП на стимуляцию тройничного нерва | – используются для исследования чувствительности в области лица |
| Термические ВП – ВП на термическую стимуляцию кожи с помощью прибора CHEPS | – используются для тестирования тонких нервных волокон, ответы регистрируют с любой части тела |
| КСП – кожный симпатический потенциал | – используются для тестирования функций волокон вегетативной нервной системы |
| Р300 – когнитивный потенциал | – тестируют функции внимания и памяти |
| ТКМС – транскраниальная магнитная стимуляция | – используется для исследований центральных двигательных проводников |
Исследование зрительных ВП проводит профессор Торопина Г.Г.
Техника регистрации: Для ВП проводится стимуляция различных сенсорных систем, регистрируют ответы мозга с помощью электродов, которые крепятся липкой лентой или пастой на кожу туловища и головы на разных уровнях прохождения вызванного импульса. Для ТКМС проводится магнитная стимуляция двигательных центров, регистрируют ответы мышц. Информация от регистрирующих электродов поступает в компьютер, где специальным образом обрабатывается и в виде графиков выводится на экран.
Процедура занимает от 30 мин до 2 часов в зависимости от вида ВП и количества исследований. Методика абсолютно безопасна.
Подготовка: чистая кожа головы и туловища, при исследовании ЗВП в случае ношения очков или линз необходимо иметь их при себе
Показания: используется в диагностике
- рассеянного склероза (тримодальные ВП: ЗВП+СВПСМ+ССВП)
- опухолей, травм, врожденных аномалий, инфекционно-аллергических и др. патологических процессов в спинном и головном мозге
- заболеваний периферических нервов – диабетических, токсических и др. полиневропатий, а также травм периферических нервов
- для диагностики болевых синдромов
- для уточнения уровня поражения нервной системы
- для дифференциальной диагностики органических и функциональных заболеваний нервной системы
Аппаратура: в отделении имеются 2 кабинета, оборудованных приборами производства фирм «Николет» (США) и «Нейрософт» (Россия)
ТКМС проводит врач Клишевская ЛА. (за компьютером), ассистирует медсестра Борзова М.А.
Описание и оценка технологий, практики, вариантов, потенциалов смягчения последствий и затрат на…
Рабочей группой III — Смягчение последствий изменения климата
Транспорт отличается от других энергопотребляющих секторов своей преобладающей зависимостью от одного ископаемого ресурса и невозможностью улавливания выбросов углерода на средствах транспорта никакими известными технологиями. Важно также рассматривать сокращение выбросов ПГ в связи с проблемами загрязнения воздуха, дорожных пробок и энергобезопасности (импорта нефти). Поэтому решения должны пытаться оптимизировать смягчение проблем транспорта в целом, а не только проблем выбросов ПГ [5.5.4].
Со времени Третьего доклада об оценках (ТДО) достигнуты значительные успехи в технологиях смягчения последствий, и по всему миру начаты масштабные программы научных исследований, разработок и демонстрации в области транспортных средств на водородных топливных батареях. Кроме того, продолжает оставаться множество возможностей совершенствования традиционных технологий. Биотоплива продолжают занимать важное место на определенных рынках и обладают гораздо большим потенциалом на будущее. Что касается выбросов других газов, кроме CO2, то разработаны автомобильные системы кондиционирования воздуха на основе хладагентов с низким ПГП [5.3].
Дорожное движение: эффективные технологии и альтернативные виды топлива
Со времени ТДО энергоэффективность дорожных транспортных средств повысилась благодаря успеху на рынке экологически более чистых дизельных двигателей с прямым впрыском и турбонаддувом (ТПВ), а также вследствие продолжающегося проникновения на рынок многих технологий с повышенной эффективностью; свою роль сыграли и гибридные двигатели, хотя степень их проникновения на рынок в настоящее время мала. Ожидаются дальнейшие технические усовершенствования в гибридных двигателях и дизельных двигателях с ТПВ. Сочетание этих усовершенствований с другими технологиями, включая замену материалов, снижение аэродинамического сопротивления, уменьшение сопротивления качению, уменьшение трения в двигателе и сокращение насосных потерь, может к 2030 году приблизительно удвоить топливную экономичность «новых» легких транспортных средств, благодаря чему выбросы углерода на 1 милю пробега сократились бы приблизительно наполовину (отметим, что это относится только к новым автомобилям, а не является средним значением по парку) (средняя степень согласия, средний объем доказательств) [5.3.1].
Биотоплива обладают потенциалом замены значительной части – но не всего – потребления нефти транспортом. В одном из последних докладов МЭА отмечено, что доля биотоплив может к 2030 году возрасти приблизительно до 10% при цене 25 долларов за тонну CO2-экв, что включает незначительный вклад биотоплив, полученных из целлюлозной биомассы. Этот потенциал сильно зависит от эффективности производства, разработки передовых технологий, например, переработки целлюлозы энзиматическими процессами или путем газификации и синтеза, затрат и конкуренции с другими направлениями землепользования. В настоящее время стоимость и производительность этанола с точки зрения предотвращения выбросов CO2 неблагоприятны, за исключением варианта производства из сахарного тростника в странах с низким уровнем заработной платы (рис. TS.16) (средняя степень согласия, средний объем доказательств) [5.3.1].
Рис. TS.16. Сравнение текущих и будущих затрат на производство биотоплива в сравнении с заводскими ценами (ФОБ) на бензин и дизельное топливо в диапазоне цен на сырую нефть [Figure 5.9].
Экономический и рыночный потенциал водородных транспортных средств остается неопределенным. Электрические транспортные средства с высоким кпд (более 90%), но с малой дальностью пробега и малым временем работы от батарей имеют ограниченную степень проникновения на рынок. Для обоих вариантов выбросы определяются производством водорода и электроэнергии. Если производить водород из угля или газа с УХУ (сейчас это самый дешевый способ) либо из биомассы, солнечной, атомной или ветровой энерии, то сопутствующие выбросы углерода можно было бы почти устранить. Потребовались бы дальнейшие технические усовершенствования в топливных элементах, хранении водорода, производстве водорода или электроэнергии с низким или нулевым уровнем выбросов углерода и батареях и (или) соответствующее снижение затрат (высокая степень согласия, средний объем доказательств) [5.3.1].
Совокупный потенциал смягчения, которым обладают энергоэффективные меры, применяемые к легким транспортным средствам, в 2030 году составил бы около 0,7–0,8 Гт СO2-экв при ценах ниже 100 долларов за тонну CO2. Данных для того, чтобы получить аналогичную оценку для тяжелых транспортных средств, недостаточно. Использование нынешних и новейших видов биотоплива, как упоминалось выше, дало бы в 2030 году дополнительный потенциал смягчения в сумме 600-1500 Мт CO2-экв при ценах ниже 25 долларов за тонну CO2 (низкая степень согласия, ограниченный объем доказательств) [5.4.2].
Критическая угроза для потенциала будущего сокращения выбросов CO2 от использования топливосберегающих технологий состоит в том, что с их помощью можно увеличить мощность и габариты автомобилей, но не улучшить общую топливную экономичность и не снизить выбросы углерода. Предпочтение рынком мощности и габаритов поглотило большую часть потенциала уменьшения последствий выбросов ПГ, достигнутого за последние два десятилетия. Если эта тенденция сохранится, это значительно уменьшит потенциал уменьшения последствий выбросов ПГ, которым обладают вышеописанные передовые технологии (высокая степень согласия, много доказательств) [5.2; 5.3].
Воздушный транспорт
Топливную экономичность гражданской авиации можно повысить самыми разными средствами, включая технологию, эксплуатацию и управление воздушным движением. Технологические усовершенствования могут обеспечить к 2015 году повышение топливной экономичности на 20% по сравнению с уровнем 1997 года, а к 2050 году возможно повышение на 40-50%. Поскольку объем перевозок в гражданской авиации продолжает расти приблизительно на 5% ежегодно, такие усовершенствования вряд ли смогут воспрепятствовать росту выбросов углерода от воздушных перевозок в глобальном масштабе. Внедрение биотоплива могло бы смягчить некоторые последствия выбросов углерода в авиации, однако на данный момент ни о стоимости такого топлива, ни об объеме выбросов в процессе их производства ничего определенного сказать нельзя (средняя степень согласия, средний объем доказательств) [5.3.3].
Воздушные перевозки можно оптимизировать на предмет энергопотребления (с минимальными выбросами CO2) путем сокращения до минимума времени руления, полетов на оптимальных крейсерских высотах, полетов по ортодромических маршрутам с минимальным расстоянием, сокращения до минимума удерживания и накопления вокруг аэропортов. Потенциал сокращения выбросов ПГ таких стратегий оценивается в 6-12%. В последнее время исследователи начали заниматься вопросом потенциала минимизации общего воздействия воздушных перевозок на климат, включая воздействие на озоновый слой, конденсационные следы и выбросы оксидов азота. Потенциал смягчения последствий в авиации на 2030 год составляет 280 Мт CO2/год при затратах менее 100 долларов на тонну CO2 (средняя степень согласия, средний объем доказательств) [5.4.2].
Морской транспорт
После ТДО в одной из оценок Международной морской организации (ММО) было обнаружено, что сочетание технических могло бы сократить выбросы углерода на 4-20% на старых кораблях и на 5-30% на новых кораблях путем применения современных знаний, в частности, в области проектирования и технического обслуживания корпуса и винта. Учитывая, однако, длительный срок службы двигателей, пройдут десятилетия, прежде чем эти меры будут реализованы в значительном масштабе на действующих кораблях. Краткосрочный потенциал оперативных мер, включая планирование маршрутов и снижение скорости, составляет от 1 до 40%. По данным исследования, максимальное сокращение выбросов от эксплуатации мирового флота составит к 2010 году около 18%, а к 2020 году, когда должны быть реализованы все меры, – около 28%. Эти данные не позволяют оценить абсолютный потенциал смягчения последствий, и, как ожидается, этот потенциал не будет достаточным для компенсации роста объема морских перевозок за этот же период (средняя степень согласия, средний объем доказательств) [5.3.4].
Железнодорожный транспорт
Главные возможности смягчения последствий выбросов ПГ, связанных с железнодорожным транспортом, состоят в улучшении аэродинамики, уменьшении веса составов, внедрении рекуперативного торможения, внедрении бортового аккумулирования энергии и, конечно, сокращении выбросов ПГ от выработки электроэнергии. Оценок совокупного потенциала смягчения и затрат на смягчение нет [5.3.2].
Смена видов транспорта и общественный транспорт
Создание систем общественного транспорта и сопутствующей инфраструктуры, а также содействие развитию немеханизированного транспорта может способствовать сокращению выбросов ПГ. При этом, однако, местные условия определяют, какое количество транспорта может быть переведено на менее энергоемкие виды. Кроме того, потенциал смягчения зависит от показателей заполненности и источников первичной энергии видов транспорта [5.3.1].
На энергетические потребности городского транспорта сильно влияет плотность и пространственная структура застройки, а также расположение, объем и характер транспортной инфраструктуры. Для расширения общественного транспорта все больше используются автобусы большой вместимости, трамвайные транзитные сети, метро, пригородные железные дороги. Для скоростных автобусных сетей характерны относительно низкие капитальные и эксплуатационные затраты, но не ясно, можно ли их внедрить в развивающихся странах с таким же успехом, как это сделано в Южной Америке. Если бы доля автобусов в пассажирском транспорте возросла на 5-10%, то выбросы CO2 снизились бы на 4-9% при затратах порядка 60–70 долларов на тонну CO2 [5.3.1].
Более чем 30% поездок автомобилей в Европе совершаются на расстояние менее 3 км, а 50% — менее 5 км. Хотя эти цифры для других континентов могут быть разными, существует потенциал смягчения последствий путем перехода с автомобилей на немеханизированный транспорт (пеший и велосипедный) или посредством предотвращения роста количества автомобильного транспорта за счет немеханизированного. Потенциал смягчения последствий сильно зависит от местных условий, однако есть существенные сопутствующие выгоды в плане качества воздуха, уменьшения пробок и повышения безопасности движения (высокая степень согласия, много доказательств) [5.3.1].
Совокупный потенциал смягчения последствий в транспортном секторе
Общий потенциал смягчения последствий выбросов CO2 и затраты на это смягчение можно оценить лишь частично, поскольку нет данных по тяжелым транспортным средствам, железнодорожному транспорту, морскому транспорту, а также по содействию смене видов транспорта и развитию общественного транспорта. Совокупный экономический потенциал повышения эффективности легких транспортных средств и самолетов, а также замены традиционного ископаемого топлива биотопливом составляет, по оценкам, около 1600-2550 Мт CO2 при цене углерода до 100 долларов за тонну CO2-экв. Это – недооценка потенциала смягчения в транспортном секторе (высокая степень согласия, средний объем доказательств) [5.4.2].
Диагностика когнитивных вызванных потенциалов (ВП) головного мозга в Москве
Диагностика когнитивных вызванных потенциалов — инструментальная неинвазивная диагностическая процедура, позволяющая оценить функции высшей нервной системы. Метод используют в детской практике в рамках комплексной диагностики заболеваний неврологического и психического профиля, а также для оценки ранних когнитивных нарушений, снижения памяти и внимания. В клинике «СМ-Доктор» данное обследование проводится детям с 6-ти лет.Особенности диагностической процедуры
Анализ когнитивных (умственных) функций подразумевает оценку способности человека к восприятию, запоминанию и воспроизведению информации, принятию решений и прогнозированию ситуаций. Все это высшие функции нервной системы, которые влияют на личностные характеристики и социальные процессы. Объективные данные о когнитивных функциях мозга позволяют выявить и оценить последствия органических изменений в нервной ткани.Метод подразумевает регистрацию и последующий анализ электрических импульсов мозга, возникающих в ответ на определенные раздражители и постановку ментальных задач. Методика проведения диагностики когнитивных вызванных потенциалов Р 300 у детей похожа на процедуру электроэнцефалографии. Во время исследования ребенку на голову надевают специальную шапочку и устанавливают электроды. Раздражающие сигналы поступают через наушник. Перед маленьким пациентом ставят задачи (например, посчитать количество разных сигналов, найти различия в картинке). Показатели прибора анализируют в совокупности с ответами ребенка. Результаты исследования позволяют поставить более точный диагноз.
Преимущества проведения теста в «СМ-Доктор»
- Опытные специалисты. Тест проводит сертифицированный врач функциональной диагностики, имеющий большой опыт работы с детьми.
- Современное оборудование. Исследование осуществляют с применением приборов Нейро Спектр КВП, Нейро Софт, 2019 года выпуска.
- Комфортные условия проведения. Исследование не сопровождается болью или другими видами дискомфорта, не требуются какие-либо инвазивные вмешательства.
Показания для проведения диагностики вызванных когнитивных потенциалов Р 300 у детей
Исследование назначают детям с уже диагностированными заболеваниями, чтобы объективно оценить когнитивные способности. Диагностику когнитивных вызванных потенциалов Р 300 назначают в следующих ситуациях:- перенесенные заболевания или травмы головного мозга;
- хроническая ишемия мозга, подтвержденная результатами нейровизуализационных диагностических процедур;
- острые нарушения мозгового кровообращения в анамнезе;
- ухудшение памяти;
- синдром дефицита внимания с гиперактивностью;
- головные боли;
- рассеянность, неспособность сконцентрировать внимание;
- снижение успеваемости в учебном заведении и пр.
Процедуру могут назначить при подозрении на неврологические и психические расстройства.
Как проходит исследование?
Диагностику вызванных когнитивных потенциалов Р 300 ребенку может назначить врач (невролог, психотерапевт, нейрохирург) в рамках консультативного осмотра. В «СМ-Доктор» исследование проводят по направлению как собственных специалистов, так и врачей из других медицинских учреждений.Диагностику когнитивных вызванных потенциалов Р300 выполняют в амбулаторном порядке без специальных подготовительных мероприятий.
Результаты диагностики вызванных когнитивных потенциалов Р 300 у детей
В процессе теста анализируют высшие функции нервной системы, включая внимание, распознавание, запоминание и воспроизведения информации, способность к обучению и принятию решений, а также ориентировочные реакции. Сведения, полученные в ходе исследования, используют для выявления причин снижения успеваемости, оценки последствий травм и ишемических процессов. Метод позволяет установить нарушения работы нервной системы и выявить заболевания еще на доклинической стадии.Противопоказания
Исследование не проводят при наличии открытых ран и инфекционно-воспалительных процессов в области головы, поскольку это затрудняет использование оборудования.Снижение успеваемости и рассеянность ребенка могут быть ранними признаками серьезного заболевания. Диагностика когнитивных вызванных потенциалов Р 300 у детей позволяет объективно оценить, есть ли начальные симптомы снижения памяти и внимания.
Звоните по телефону или заполняйте форму обратной связи на сайте, чтобы записать ребенка на диагностику когнитивных вызванных потенциалов Р300. Исследование проводится в удобное время!
Записаться на приём или задать вопросы можно круглосуточно по телефону +7 (495) 292-59-86
Вызванные потенциалы мозга
Вызванные потенциалы мозга(ВПМ) — это объективный неинвазивный метод тестирования центральной нервной системы. Использование вызванных потенциалов (ВП) – акустических, зрительных, соматосенсорных — является средством для раннего обнаружения и прогноза неврологических нарушений при различных заболеваниях, таких как рассеянный склероз, черепно-мозговые травмы, опухоли.
Основные области применения:
-
оценка расстройств зрительного пути
-
объективное тестирование функций слуха
-
оценка состояния сенсомоторной области
-
нарушение функций коры мозга, в том числе деменции различного генеза
-
метаболические и токсические повреждения центральной нервной системы
-
локализация нарушений ствола мозга
-
оценка развития мозгового ствола и коры
-
состояние когнитивных функций мозга
-
нарушения периферических нервов
-
нарушения проводящих путей спинного мозга
-
оценка комы и смерть мозга
Диагностические возможности отдельных методик:
Зрительные ВП:
-
оценка состояния при патологии зрительного нерва (невриты, демиелинизирующие заболевания)
-
объективная оценка зрительных нарушений и дифференциальная диагностика функциональных и органических состояний
-
объективная оценка состояния зрительных функций у маленьких детей
-
диагностика нарушений полей зрения (объективная периметрия)
Акустические ВП:
-
объективная аудиометрия
-
локализации патологии ствола мозга при опухоли, инфарктах и др.
-
диагностика демиелинизирующих заболеваний, прежде всего рассеянного склероза
-
ранняя диагностика невриномы слухового нерва
-
оценка развития мозгового ствола
-
оценка слуха у новорожденного
-
мониторинг состояния стволовых структур при операциях на задней черепной ямке
-
прогноз у больных, находящихся в коме
-
подтверждение диагноза смерти мозга при изоэлектрической ЭЭГ
Соматосенсорные ВП:
-
мониторинг функций спинного мозга и сенсорной коры
-
повреждение на уровне шейного и поясничного утолщений спинного мозга
-
поражение корешков и сплетений
-
демиелинизирующие заболевания центральной нервной системы
-
оценка нарушений сенсорных функций у истеричных больных
-
травматические повреждения спинного мозга
Мембранные потенциалы — WikiLectures
Мембранные потенциалы описываются различными конфигурациями ионной концентрации снаружи и внутри мембраны клетки. Эти потенциалы таковы:
- Мембранный потенциал покоя: мембранный потенциал в состоянии покоя, стационарных условиях.
- Потенциал действия: не оцениваемый потенциал, очень похожий на двоичный код (вкл / выкл).
- Постсинаптические потенциалы: градуированные потенциалы, которые можно суммировать / вычитать путем модуляции пресинаптических нейронов.
[редактировать часть] Человеческий организм состоит из множества клеток, каждая из которых имеет разные компоненты и, следовательно, имеет разные мембранные потенциалы покоя . Некоторые из этих клеток возбудимы (например, клетки; нейроны; мышечные волокна), генерируя потенциал действия при воздействии внешнего раздражителя, вызывая деполяризацию мембраны. Мембранный потенциал покоя (RMP) возникает из-за изменений проницаемости мембраны для калия, натрия, кальция и хлорида, которая возникает в результате движения этих ионов через нее.Как только мембрана поляризована и , она приобретает напряжение, которое представляет собой разность потенциалов между внутриклеточным и внеклеточным пространствами.
Что такое RMP?
Мембранный потенциал покоя составляет:
- — неравномерное распределение ионов по обеим сторонам клеточной мембраны;
- разность напряжений покоящихся ячеек;
- мембранный потенциал, который поддерживался бы, если бы через нее не было никаких стимулов или проводящих импульсов;
- определяется концентрацией ионов по обе стороны мембраны;
- — отрицательное значение . означает, что внутри элемента имеется избыток отрицательного заряда по сравнению с внешним.
- во многом зависит от уровня внутриклеточного калия, так как проницаемость мембран для калия примерно в 100 раз выше, чем для натрия.
Производство и обслуживание RMP
RMP производится и обслуживается:
- Эффект Доннана
- описывается как большие непроницаемые отрицательно заряженные внутриклеточные молекулы, притягивающие положительно заряженные ионы (например: Na + и K + ) и отталкивающие отрицательные (например.г .: Класс — )
- Селективность мембраны
- — разница проницаемости между разными ионами
- Активный транспорт (Na + / K + ATPase насос)
- — это опосредованный процесс перемещения частиц через биологическую мембрану против градиента концентрации.
- Первичный активный транспорт — если он тратит энергию. Так работает насос Na + / K + АТФазы.
- Вторичный активный транспорт — если он включает электрохимический градиент.Это не связано с поддержанием RMP.
Ионное воздействие на мембранный потенциал покоя
RMP создается за счет распределения ионов и их диффузии через мембрану. Ионы калия важны для RMP из-за его активного транспорта , который еще больше увеличивает его концентрацию внутри клетки. Однако калий-селективные ионные каналы всегда открыты, производя накопление отрицательного заряда внутри элемента.Его движение наружу обусловлено случайным движением молекул и продолжается до тех пор, пока внутри клетки не накопится достаточно избыточного отрицательного заряда, чтобы сформировать мембранный потенциал.
Na
+ / K + Поражение АТФазной помпы RMPNa + / K + АТФазный насос создает градиент концентрации, перемещая 3Na + из клетки и 2K + в клетку. Na + откачивается, а K + закачивается против их градиентов концентрации.Поскольку этот насос перемещает ионы против градиентов их концентрации, ему требуется энергия .
Влияние ионных каналов на мембранный потенциал покоя
Клеточная мембрана содержит белковых каналов , которые позволяют ионам пассивно диффундировать без прямых затрат метаболической энергии. Эти каналы позволяют Na + и K + перемещаться через клеточную мембрану от более высокой концентрации к более низкой. Поскольку эти каналы обладают селективностью по отношению к определенным ионам, существует калий- и натрий-селективных ионных каналов .Все клеточные мембраны более проницаемы для K + , чем для Na + , потому что у них больше каналов K + , чем для Na + .
Уравнение Нернста
Это математическое уравнение, применяемое в физиологии для расчета равновесных потенциалов для определенных ионов.
[математика] Ei = (\ frac {R · T} {F · z}) \ cdot ln \ frac {[X] 1} {[X] 2} [/ math]
- R = Газовая постоянная
- T = Абсолютная температура (K)
- E = разность потенциалов на мембране
- F = Константа Фарадея (96 500 кулонов / моль)
- z = Валентность иона
Уравнение Гольдмана-Ходжкина-Каца
— математическое уравнение, применяемое в физиологии для определения потенциала через клеточную мембрану с учетом всех проницаемых ионов.
[математика] Em = 58 log (\ frac {PNa \ cdot [Na] out + PK \ cdot [K] out} {PNa \ cdot [Na] in + PK \ cdot [K] in}) [/ math ]
- E = разность потенциалов на мембране
- P = проницаемость мембраны для натрия или калия
- [] = Концентрация натрия или калия внутри или снаружи
Измерение потенциалов покоя
В некоторых ячейках постоянно меняется об / мин. . Для этого никогда не существует потенциала покоя , что является лишь теоретической концепцией.Другие клетки с мембранными транспортными функциями, которые меняют потенциал со временем, обладают потенциалом покоя. Это можно измерить, вставив электрод в ячейку. Трансмембранные потенциалы также можно измерить оптически с помощью красителей, которые изменяют свои оптические свойства в соответствии с мембранным потенциалом.
Мембранный потенциал покоя варьируется в зависимости от типа клеток
- Например:
- Клетки скелетных мышц: −95 мВ
- Гладкомышечные клетки: −50 мВ
- Астроциты: −80 / −90 мВ
- Нейроны: −70 мВ
- Эритроциты: −12 мВ
[редактировать часть] Потенциал действия — это событие, при котором мембранный потенциал клетки быстро повышается и понижается.Траектория следует постоянной схеме.
Компоненты
Потенциал покоя
Потенциал покоя в значительной степени определяется разницей концентраций ионов K + и имеет значение от -70 до -90 мВ, внутренняя часть ячейки имеет отрицательный заряд.
Активный потенциал
Если ввести один электрод внутрь аксона, а другой — на цитоплазматическую поверхность аксона, происходит гиперполяризация (в случае отрицательных внутренних электродов) или деполяризация (в случае отрицательных внешних).
Если мы увеличим мембранный потенциал до порогового потенциала (в мембране с мембранным потенциалом покоя , от -70 мВ до примерно -55 мВ), нервное волокно ответит появлением потенциала действия (внезапное открытие, управляемое напряжением. каналы ионов натрия, таким образом позволяя ионам натрия проходить через мембрану, заставляя внутреннюю часть клеток становиться положительной — имеется трансполяризация ().
Если приращение мембранного потенциала не достигает «порогового потенциала», натриевый потенциал-зависимый канал не открывается.В этом случае потенциал действия не создается.
На следующем этапе мембрана снова становится проницаемой для ионов калия, и потенциал возвращается к исходному значению, несмотря на небольшую гиперполяризацию.
Различия с постсинаптическим потенциалом
| Параметр | AP | PSP |
|---|---|---|
| Характеристика | Все или ничего | баллов |
| амплитуда | около 100 мВ | 1–10 мВ |
| продолжительность | ок. 10–40 мс | 1–5 мс |
| Где? | по аксону | постсинаптическая мембрана (тело нейрональной клетки, дендриты) |
| разворот | без декремента | с декрементом |
Постсинаптические потенциалы
Ионные основы постсинаптического возбуждения
В возбуждающем синапсе нейромедиатор открывает каналы в постсинаптической мембране, которые проницаемы для катионов, в основном для Na + и K + .Чистый ионный ток (IEPSP), который вызывает EPSP, представляет собой сумму отдельных токов, переносимых всеми ионами, пронизывающими канал.
Возможности разворота ВПСП. Обратный потенциал (потенциал Нернста) — это мембранный потенциал Em, при котором чистый ионный ток через канал равен нулю.
I EPSP = I Na + I K
E rev = g Na E Na + g K E K / g Na + g K
- г Na , г K — проводимости, пропорциональные проницаемости канала для Na + и K +
- E Na , E K — потенциалы Нернста для этих ионов .
- Перестановка
[математика] \ dfrac {g_ {Na}} {g_K} = \ dfrac {E_K — E_ {rev}} {E_ {rev} — E_K} [/ math]
Ионные основы постсинаптического ингибирования
Когда мембранный потенциал покоя составляет -65 мВ, IEPSP направлен наружу; Направленный наружу ток Cl — представляет собой приток отрицательно заряженного Cl — , который гиперполяризует клетку. Внешний синаптический ток будет увеличиваться, когда мембранный потенциал становится менее отрицательным (деполяризованным), и уменьшаться, когда потенциал становится более отрицательным (гиперполяризованным).
Другая возможность состоит в том, что канал тормозного синапса проницаем для K + с потенциалом Нернста -90 мВ. Повышенная проницаемость для K + может приблизить мембранный потенциал к потенциалу Нернста для K + и, таким образом, гиперполяризовать клетку.
Общие свойства постсинаптических потенциалов
- Постсинаптические потенциалы (например, потенциалы концевой пластинки и рецептора) представляют собой постепенное изменение потенциала. Амплитуда постсинаптического потенциала увеличивается с увеличением количества высвобождаемого нейромедиатора.Другими словами, он увеличивается с количеством рецепторов, связанных нейромедиатором, учитывая, что эти рецепторы приведут к деполяризации клетки , что приведет к притоку Ca 2+ , что в конечном итоге вызовет экзоцитоз нейротрансмиттера в следующих случаях: синаптическая щель.
- Поскольку постсинаптические потенциалы градуированы, индивидуальные потенциальные изменения могут суммироваться. Суммирование постсинаптических потенциалов может быть либо временным, либо пространственным:
- Временное суммирование возникает, когда один и тот же вход стимулируется повторно, так что второй синаптический потенциал наступает до того, как постсинаптическая клетка восстановится от первого синаптического потенциала.Существует запас в 15 мс с момента появления потенциала действия, в течение которого второй постсинаптический потенциал может быть наложен и добавлен к первому, прежде чем он исчезнет до -65 мВ (в состоянии покоя). Это приводит к постепенному увеличению изменения постсинаптического потенциала.
- Во время пространственного суммирования второй синаптический потенциал обеспечивается вторым входом одновременно, что создает синаптический контакт в непосредственной близости от первого входа. Если два входа имеют одинаковый знак (оба возбуждающие или оба тормозящие), постсинаптический ответ, вызванный стимуляцией, будет больше, чем если бы любой вход стимулировался отдельно.
- Одиночные возбуждающие PSP не превышают 0,5–1 мВ.
- И в возбуждающих, и в тормозных PSP изменение ионной проводимости (пик) длится 1-2 мс, а затем требуется до 15 мс для восстановления потенциала покоя, потому что это время, необходимое для избыточного положительного / отрицательного заряды утекают / поступают из / в ячейку.
- Порог возбуждения в среднем составляет около -45 мВ. Чтобы достичь этого порога, нам потребуется суммарная ВПСП +20 мВ. Пространственное и временное суммирование отдельных PSP (40-80 EPSP) из дендритов и сомы может дать такое значение.
- Однако, даже если достигается это значение (+20 мВ) и потенциал сомы> -45 мВ (порог), потенциал действия не может быть выявлен. Этот порог применяется только для бугорка аксона , где наблюдается максимальная плотность Na + -VGC. Чтобы потенциал действия мог быть вызван сомой, суммарная величина ВПСП должна находиться в пределах от +30 мВ до 40 мВ.
Важность суммирования становится очевидной при рассмотрении модели спинномозгового мотонейрона: каждый мотонейрон несет около 20 000 индивидуальных синаптических бутонов, которые представляют около 6000 отдельных входов (каждый вход формирует более одного синапса).В реальной ситуации нейроны находятся под постоянным влиянием как тормозных, так и возбуждающих синаптических входов. В любой момент мембранный потенциал нейронов отражает сумму всех этих входов. Снижение уровня тонического возбуждающего воздействия приведет к гиперполяризации клетки, так же как и к увеличению тормозного воздействия. Точно так же увеличение тонического возбуждающего воздействия деполяризует клетку, что также может быть достигнуто за счет уменьшения мощности тонического подавляющего воздействия.
Облегчение нейронов
Облегчение нейронов означает приближение к порогу.Часто суммарный постсинаптический потенциал является возбуждающим, но не поднимается достаточно высоко, чтобы достичь порога активации постсинаптического нейрона. Когда это происходит, нейрон считается облегченным. То есть его мембранный потенциал ближе к порогу срабатывания, чем обычно, но еще не на уровне срабатывания. Следовательно, другой возбуждающий сигнал, поступающий в нейрон из другого источника, может очень легко возбудить нейрон. Распространенные сигналы в нервной системе часто действительно облегчают работу больших групп нейронов, так что они могут быстро и легко реагировать на сигналы, поступающие из других источников.
Возбужденное состояние
«Состояние возбуждения» нейрона определяется как суммарная степень возбуждающего воздействия на нейрон. Если существует более высокая степень возбуждения, чем ингибирование нейрона в любой данный момент, то говорят, что существует состояние возбуждения. И наоборот, если больше торможения, чем возбуждения, то говорят, что есть тормозящее состояние. Когда состояние возбуждения нейрона превышает пороговое значение для возбуждения, нейрон будет срабатывать повторно, пока состояние возбуждения остается на этом уровне.
Некоторые нейроны центральной нервной системы срабатывают непрерывно, потому что даже нормальное состояние возбуждения выше порогового уровня. Их частоту стрельбы обычно можно еще больше увеличить, еще больше усилив их возбуждающее состояние. Частоту можно уменьшить или даже остановить возбуждение, наложив на нейрон тормозящее состояние. Это похоже на модуляцию типа несущей на непрерывных скоростях.
Таким образом, разные нейроны по-разному реагируют, имеют разные пороги возбуждения и сильно различаются максимальные частоты разряда.
Пресинаптическое торможение
Пресинаптическое торможение вызывается высвобождением тормозящего вещества на внешней стороне пресинаптических нервных волокон до того, как их собственные окончания завершатся на постсинаптическом нейроне. В большинстве случаев ингибирующим передающим веществом является ГАМК (γ-аминомасляная кислота). Это имеет специфический эффект открытия анионных каналов, позволяя большому количеству хлорид-ионов диффундировать в концевые фибриллы. Отрицательные заряды этих ионов подавляют синаптическую передачу, потому что они нейтрализуют большую часть возбуждающего эффекта положительно заряженных ионов натрия, которые также входят в концевые фибриллы при достижении потенциала действия.Пресинаптическое торможение происходит во многих сенсорных путях нервной системы. Фактически, соседние сенсорные нервные волокна часто взаимно подавляют друг друга, что сводит к минимуму боковое распространение и смешивание сигналов в сенсорных трактах. Это основа бокового торможения.
Ссылки
Статьи по теме
Источники
- ПОКОРНЫЙ, Ярослав. Ионные каналы, мембранные потенциалы и их распространение [лекция по предмету «Физиология», специализация «Физиология», 1.LF UK Карлов университет в Праге. Прага. 2010.
Библиография
- HALL, John E — GUYTON, Arthur Clifton. Учебник медицинской физиологии Гайтона и Холла. 11. издание. Saunders / Elsevier, 2005. ISBN 0721602401.
- DESPOPOULOS, Agamnenon — SILBERNAGL, Stefan. Цветной атлас физиологии. 5. издание. Thieme, 2003. ISBN 3135450058.
Дополнительная литература
Связь между нейронами | Анатомия и физиология I
Цели обучения
- Объясните различия между типами градуированных потенциалов
- Классифицируйте основные нейротрансмиттеры по химическому типу и эффекту
Электрические изменения, происходящие в нейроне, как описано в предыдущем разделе, аналогичны включению выключателя света.Стимул запускает деполяризацию, но потенциал действия запускается сам по себе при достижении порога. Теперь возникает вопрос: «Что включает выключатель света?» Временные изменения напряжения клеточной мембраны могут быть результатом получения нейронами информации из окружающей среды или воздействия одного нейрона на другой. Эти особые типы потенциалов влияют на нейрон и определяют, возникнет ли потенциал действия. Многие из этих временных сигналов исходят из синапсов.
Градуированные потенциалы
Локальные изменения мембранного потенциала называются градуированными потенциалами и обычно связаны с дендритами нейрона. Величина изменения мембранного потенциала определяется размером стимула, который его вызывает. В примере тестирования температуры душа слегка теплая вода вызовет только небольшое изменение терморецептора, тогда как горячая вода вызовет большое изменение мембранного потенциала.
Градуированные потенциалы могут быть двух видов: деполяризующие или гиперполяризующие (рис. 1).Для мембраны с потенциалом покоя градиентный потенциал представляет изменение этого напряжения выше -70 мВ или ниже -70 мВ. Деполяризующие градиентные потенциалы часто возникают в результате попадания в клетку Na + или Ca 2+ . Оба этих иона имеют более высокие концентрации вне клетки, чем внутри; поскольку они имеют положительный заряд, они будут перемещаться в ячейку, делая ее менее отрицательной по сравнению с внешним миром. Гиперполяризующие градиентные потенциалы могут быть вызваны выходом K + из ячейки или проникновением Cl — в ячейку.Если положительный заряд выходит из ячейки, ячейка становится более отрицательной; если в ячейку попадает отрицательный заряд, происходит то же самое.
Рисунок 1. Градуированные потенциалы. Градуированные потенциалы — это временные изменения мембранного напряжения, характеристики которых зависят от размера стимула. Некоторые типы стимулов вызывают деполяризацию мембраны, тогда как другие вызывают гиперполяризацию. Это зависит от конкретных ионных каналов, которые активируются в клеточной мембране.
Типы градуированных потенциалов
Для униполярных клеток сенсорных нейронов — как со свободными нервными окончаниями, так и внутри инкапсуляций — в дендритах развиваются дифференцированные потенциалы, которые влияют на генерацию потенциала действия в аксоне той же самой клетки. Это называется потенциалом генератора . Для других сенсорных рецепторных клеток, таких как вкусовые клетки или фоторецепторы сетчатки, градиентные потенциалы в их мембранах приводят к высвобождению нейротрансмиттеров в синапсах с сенсорными нейронами.Это называется потенциалом рецептора .
Постсинаптический потенциал (PSP) — это градиентный потенциал дендритов нейрона, который получает синапсы от других клеток. Постсинаптические потенциалы могут быть деполяризующими или гиперполяризующими. Деполяризация постсинаптического потенциала называется возбуждающим постсинаптическим потенциалом (ВПСП) , потому что она заставляет мембранный потенциал приближаться к пороговому значению. Гиперполяризация постсинаптического потенциала — это ингибирующий постсинаптический потенциал (IPSP) , потому что он заставляет мембранный потенциал отклоняться от порогового значения.
Суммирование
Рисунок 2. Суммирование постсинаптического потенциала Результатом суммирования постсинаптических потенциалов является общее изменение мембранного потенциала. В точке А несколько различных возбуждающих постсинаптических потенциалов складываются в большую деполяризацию. В точке B сочетание возбуждающего и тормозного постсинаптических потенциалов приводит к другому конечному результату для мембранного потенциала.
Все типы градуированных потенциалов приводят к небольшим изменениям деполяризации или гиперполяризации в напряжении мембраны.Эти изменения могут привести к достижению нейроном порога, если изменения складываются вместе, или суммируют . Комбинированные эффекты различных типов градуированных потенциалов показаны на рисунке 2. Если общее изменение напряжения в мембране составляет положительное значение 15 мВ, что означает, что мембрана деполяризуется от -70 мВ до -55 мВ, то в результате будут получены градиентные потенциалы. в мембране, достигающей порога.
Для рецепторных потенциалов порог не является фактором, потому что изменение мембранного потенциала рецепторных клеток напрямую вызывает высвобождение нейромедиатора.Однако генераторные потенциалы могут инициировать потенциалы действия в аксоне сенсорного нейрона, а постсинаптические потенциалы могут инициировать потенциал действия в аксоне других нейронов. Градуированные потенциалы суммируются в определенном месте в начале аксона, чтобы инициировать потенциал действия, а именно в начальном сегменте. Для сенсорных нейронов, у которых нет тела клетки между дендритами и аксоном, начальный сегмент непосредственно примыкает к дендритным окончаниям. Для всех других нейронов бугорок аксона по существу является начальным сегментом аксона, и именно здесь происходит суммирование.Эти места имеют высокую плотность потенциалзависимых каналов Na + , которые инициируют фазу деполяризации потенциала действия.
Суммирование может быть пространственным или временным, что означает, что оно может быть результатом нескольких градуированных потенциалов в разных местах нейрона или всех в одном месте, но разделенных во времени. Пространственное суммирование связано с связыванием активности нескольких входов нейрона друг с другом. Суммирование во времени — это соотношение множественных потенциалов действия одной клетки, приводящее к значительному изменению мембранного потенциала.Пространственное и временное суммирование также может действовать вместе.
Посмотрите это видео, чтобы узнать о суммировании.
Процесс преобразования электрических сигналов в химические и обратно требует тонких изменений, которые могут привести к кратковременному увеличению или уменьшению мембранного напряжения. Чтобы вызвать длительное изменение в целевой ячейке, несколько сигналов обычно складываются или суммируются. Должно ли происходить пространственное суммирование сразу, или отдельные сигналы могут поступать на постсинаптический нейрон в несколько разное время? Поясните свой ответ.
Синапсы
Есть два типа соединений между электрически активными клетками: химические синапсы и электрические синапсы. В химическом синапсе химический сигнал, а именно нейротрансмиттер, высвобождается из одной клетки и воздействует на другую клетку. В электрическом синапсе существует прямая связь между двумя ячейками, так что ионы могут проходить напрямую от одной ячейки к другой. Если одна клетка деполяризуется в электрическом синапсе, присоединенная клетка также деполяризуется, потому что ионы проходят между клетками.Химические синапсы включают передачу химической информации от одной клетки к другой. В этом разделе мы сосредоточимся на химическом типе синапсов.
Примером химического синапса является нервно-мышечное соединение (НМС), описанное в главе о мышечной ткани. В нервной системе есть намного больше синапсов, которые по сути такие же, как NMJ. Все синапсы имеют общие характеристики, которые можно кратко изложить в этом списке:
- пресинаптический элемент
- Нейромедиатор (в пузырьках)
- синаптическая щель
- рецепторные белки
- постсинаптический элемент
- Удаление или повторное поглощение нейромедиаторов
Для NMJ эти характеристики следующие: пресинаптический элемент — это окончания аксона моторного нейрона, нейротрансмиттер — ацетилхолин, синаптическая щель — это пространство между клетками, в котором диффундирует нейротрансмиттер, рецепторный белок — это никотиновый рецептор ацетилхолина, постсинаптическим элементом является сарколемма мышечной клетки, а нейромедиатор удаляется ацетилхолинэстеразой.Другие синапсы похожи на этот, и их особенности различны, но все они содержат одинаковые характеристики.
Выпуск нейротрансмиттера
Когда потенциал действия достигает терминалов аксона, управляемые по напряжению каналы Ca 2+ в мембране синаптической конечной луковицы открываются. Концентрация Ca 2+ увеличивается внутри концевой луковицы, и ион Ca 2+ связывается с белками на внешней поверхности везикул нейромедиатора. Ca 2+ способствует слиянию везикулы с пресинаптической мембраной, так что нейромедиатор высвобождается посредством экзоцитоза в небольшой промежуток между клетками, известный как синаптическая щель .
Попав в синаптическую щель, нейромедиатор диффундирует на короткое расстояние к постсинаптической мембране и может взаимодействовать с рецепторами нейромедиатора. Рецепторы специфичны для нейротрансмиттера, и они подходят друг другу, как ключ и замок. Один нейротрансмиттер связывается со своим рецептором и не связывается с рецепторами других нейротрансмиттеров, что делает связывание специфическим химическим событием (рис. 3).
Рисунок 3. Синапс. Синапс — это соединение между нейроном и его клеткой-мишенью (которая не обязательно является нейроном).Пресинаптический элемент — это синаптическая концевая луковица аксона, где Ca 2+ входит в луковицу, чтобы вызвать слияние пузырьков и высвобождение нейромедиатора. Нейромедиатор диффундирует через синаптическую щель, чтобы связываться со своим рецептором. Нейротрансмиттер выводится из синапса либо ферментативной деградацией, нейрональным обратным захватом, либо глиальным повторным захватом.
Нейротрансмиттерные системы
Есть несколько систем нейромедиаторов, которые находятся в различных синапсах нервной системы.Эти группы относятся к химическим веществам, которые являются нейротрансмиттерами, и внутри групп представляют собой определенные системы.
Первая группа, которая представляет собой собственную нейромедиаторную систему, — это холинергическая система . Это система на основе ацетилхолина. Это включает в себя NMJ как пример холинергического синапса, но холинергические синапсы обнаруживаются в других частях нервной системы. Они находятся в вегетативной нервной системе, а также распределяются по всему мозгу.
Холинергическая система имеет два типа рецепторов, никотиновый рецептор находится в НМС, а также в других синапсах.Существует также рецептор ацетилхолина, известный как мускариновый рецептор . Оба эти рецептора названы в честь лекарств, которые взаимодействуют с рецептором в дополнение к ацетилхолину. Никотин связывается с никотиновым рецептором и активирует его подобно ацетилхолину. Мускарин, продукт некоторых грибов, связывается с мускариновыми рецепторами. Однако никотин не будет связываться с мускариновым рецептором, а мускарин не будет связываться с никотиновым рецептором.
Другая группа нейромедиаторов — аминокислоты.Сюда входят глутамат (Glu), ГАМК (гамма-аминомасляная кислота, производное глутамата) и глицин (Gly). Эти аминокислоты имеют в своей химической структуре аминогруппу и карбоксильную группу. Глутамат — одна из 20 аминокислот, которые используются для производства белков. Каждый аминокислотный нейромедиатор будет частью своей собственной системы, а именно глутаматергической, ГАМКергической и глицинергической систем. У каждого из них есть свои рецепторы, и они не взаимодействуют друг с другом. Аминокислотные нейротрансмиттеры удаляются из синапсов путем обратного захвата.Насос в клеточной мембране пресинаптического элемента или иногда соседней глиальной клетки очищает аминокислоту от синаптической щели, чтобы ее можно было повторно использовать, переупаковывать в пузырьки и снова высвобождать.
Другой класс нейротрансмиттеров — это биогенный амин , группа нейротрансмиттеров, которые ферментативно производятся из аминокислот. В них есть аминогруппы, но больше нет карбоксильных групп, и поэтому они больше не классифицируются как аминокислоты. Серотонин производится из триптофана.Это основа серотонинергической системы, которая имеет свои специфические рецепторы. Серотонин транспортируется обратно в пресинаптическую клетку для переупаковки.
Другие биогенные амины производятся из тирозина и включают дофамин, норадреналин и адреналин. Дофамин является частью собственной системы, дофаминергической системы, которая имеет дофаминовые рецепторы. Дофамин удаляется из синапса транспортными белками пресинаптической клеточной мембраны. Норадреналин и адреналин относятся к адренергической нейромедиаторной системе.Две молекулы очень похожи и связываются с одними и теми же рецепторами, которые называются альфа- и бета-рецепторами. Норадреналин и адреналин также транспортируются обратно в пресинаптическую клетку. Химический адреналин ( epi — = «включен»; «- нефрин, » = почка) также известен как адреналин ( почечный, = «почка»), а норадреналин иногда называют норадреналином. Надпочечники вырабатывают адреналин и норадреналин, которые попадают в кровоток в виде гормонов.
Нейропептид представляет собой молекулу нейромедиатора, состоящую из цепочек аминокислот, соединенных пептидными связями. Вот что такое белок, но термин «белок» подразумевает определенную длину молекулы. Некоторые нейропептиды довольно короткие, например мет-энкефалин, который состоит из пяти аминокислот. Другие длинные, например бета-эндорфин, который состоит из 31 аминокислоты. Нейропептиды часто высвобождаются в синапсах в сочетании с другим нейротрансмиттером, и они часто действуют как гормоны в других системах организма, таких как вазоактивный кишечный пептид (VIP) или вещество P.
Действие нейротрансмиттера на постсинаптический элемент полностью зависит от рецепторного белка. Во-первых, если в мембране постсинаптического элемента нет рецепторного белка, то нейромедиатор не действует. Деполяризующий или гиперполяризационный эффект также зависит от рецептора. Когда ацетилхолин связывается с никотиновым рецептором, постсинаптическая клетка деполяризуется. Это связано с тем, что рецептор представляет собой катионный канал, и положительно заряженный Na + устремляется в клетку.Однако, когда ацетилхолин связывается с мускариновым рецептором, который существует в нескольких вариантах, он может вызвать деполяризацию или гиперполяризацию клетки-мишени.
Аминокислотные нейротрансмиттеры, глутамат, глицин и ГАМК, почти всегда связаны только с одним эффектом. Глутамат считается возбуждающей аминокислотой, но только потому, что рецепторы Glu у взрослых вызывают деполяризацию постсинаптической клетки. Глицин и ГАМК считаются ингибирующими аминокислотами, опять же потому, что их рецепторы вызывают гиперполяризацию.
Биогенные амины обладают смешанным действием. Например, рецепторы дофамина, которые классифицируются как рецепторы D1, являются возбуждающими, тогда как рецепторы типа D2 являются ингибирующими. Биогенные аминовые рецепторы и рецепторы нейропептидов могут иметь еще более сложные эффекты, поскольку некоторые из них могут не напрямую влиять на мембранный потенциал, а скорее влияют на транскрипцию генов или другие метаболические процессы в нейроне. Характеристики различных нейротрансмиттерных систем, представленные в этом разделе, представлены в таблице 1.
| Таблица 1. Характеристики нейротрансмиттерных систем | ||||
|---|---|---|---|---|
| Система | Холинергический | Аминокислоты | Биогенные амины | Нейропептиды |
| Нейротрансмиттеры | Ацетилхлоний | Глутамат, глицин, ГАМК | Серотонин (5-HT) | мет-энкафалин, бета-эндорфин, VIP, вещество P и т. Д. |
| Рецепторы | Никотонические и мускариновые рецепторы | Рецепторы Glu, рецепторы gly, рецепторы GABA | 5-HT рецепторы, D1 и D2 рецепторы, a-адренорецепторы и B-адренергические рецепторы | Рецепторовслишком много, чтобы их перечислить, но они специфичны для пептидов |
| Исключение | Дегрегация ацетилхолинэстеразой | Обратный захват нейронами глии | Обратный захват нейронами | Дегрегация ферментами, называемыми пептидазами |
| Постсинаптический эффект | Никотонический рецептор вызывает деполяризацию.Мускариновые рецепторы могут вызывать как деполяризацию, так и гиперполяризацию в зависимости от подтипа | РецепторыGlu вызывают деполяризацию. Рецепторы Gly и GABA вызывают гиперполяризацию | Деполяризация или гиперполяризация зависит от конкретного рецептора. Например, рецепторы D1 вызывают деполяризацию, а рецепторы D2 вызывают гиперполяризацию | Деполяризация или гиперполяризация зависит от конкретного рецептора |
Важно помнить о нейротрансмиттерах и сигнальных химических веществах в целом, что эффект полностью зависит от рецептора.Нейротрансмиттеры связываются с одним из двух классов рецепторов на поверхности клетки, ионотропными или метаботропными (рис. 4). Ионотропные рецепторы представляют собой ионные каналы, управляемые лигандами, такие как никотиновый рецептор для ацетилхолина или рецептор глицина. Метаболический рецептор включает комплекс белков, которые приводят к метаболическим изменениям внутри клетки. Рецепторный комплекс включает трансмембранный рецепторный белок, G-белок и эффекторный белок. Нейромедиатор, называемый первым мессенджером, связывается с рецепторным белком на внеклеточной поверхности клетки, а внутриклеточная сторона белка инициирует активность G-белка.Белок G представляет собой гидролазу гуанозинтрифосфата (GTP), которая физически перемещается от рецепторного белка к эффекторному белку для активации последнего. Эффекторный белок представляет собой фермент, который катализирует образование новой молекулы, которая действует как внутриклеточный медиатор сигнала, который связывается с рецептором. Этот внутриклеточный посредник называется вторым мессенджером.
Рисунок 4. Типы рецепторов. (a) Ионотропный рецептор — это канал, который открывается, когда с ним связывается нейромедиатор.(б) метаботропный рецептор — это комплекс, который вызывает метаболические изменения в клетке, когда с ней связывается нейромедиатор (1). После связывания G-белок гидролизует GTP и переходит к эффекторному белку (2). Когда белок G контактирует с эффекторным белком, генерируется второй мессенджер, такой как цАМФ (3). Затем второй мессенджер может вызывать изменения в нейроне, такие как открытие или закрытие ионных каналов, метаболические изменения и изменения в транскрипции генов.
Различные рецепторы используют разные вторичные мессенджеры.Двумя распространенными примерами вторичных мессенджеров являются циклический аденозинмонофосфат (цАМФ) и инозитолтрифосфат (IP 3 ). Фермент аденилатциклаза (пример эффекторного белка) производит цАМФ, а фосфолипаза C является ферментом, который образует IP 3 . Вторичные мессенджеры после того, как они вырабатываются эффекторным белком, вызывают метаболические изменения внутри клетки. Эти изменения, скорее всего, являются активацией других ферментов в клетке. В нейронах они часто изменяют ионные каналы, открывая или закрывая их.Эти ферменты также могут вызывать изменения в клетке, такие как активация генов в ядре и, следовательно, усиление синтеза белков. В нейронах подобные изменения часто являются основой более прочных связей между клетками в синапсе и могут быть основой обучения и памяти.
Посмотрите это видео, чтобы узнать о выбросе нейромедиатора.
Потенциал действия достигает конца аксона, называемого концом аксона, и высвобождается химический сигнал, чтобы сказать клетке-мишени что-то сделать — либо инициировать новый потенциал действия, либо подавить эту активность.За очень короткое время электрический сигнал потенциала действия преобразуется в химический сигнал нейромедиатора, а затем обратно в электрические изменения в мембране клетки-мишени. Какое значение имеют потенциал-управляемые кальциевые каналы в высвобождении нейромедиаторов?
Заболевания нервной системы
Основная причина некоторых нейродегенеративных заболеваний, таких как болезни Альцгеймера и Паркинсона, по-видимому, связана с белками, в частности с белками, которые плохо себя ведут.Одна из самых сильных теорий о том, что вызывает болезнь Альцгеймера, основана на накоплении бета-амилоидных бляшек, плотных скоплений белка, который не функционирует должным образом. Болезнь Паркинсона связана с увеличением белка, известного как альфа-синуклеин, который токсичен для клеток ядра черной субстанции в среднем мозге.
Правильное функционирование белков зависит от их трехмерной формы. Линейная последовательность аминокислот складывается в трехмерную форму, которая основана на взаимодействиях между этими аминокислотами.Когда сворачивание нарушается и белки принимают другую форму, они перестают правильно функционировать. Но болезнь не обязательно является результатом функциональной потери этих белков; скорее, эти измененные белки начинают накапливаться и могут стать токсичными. Например, при болезни Альцгеймера признаком болезни является накопление этих амилоидных бляшек в коре головного мозга. Термин «протеопатия», придуманный для описания этого вида заболеваний, включает и другие заболевания. Болезнь Крейтцфельда-Якоба, человеческий вариант прионной болезни, известной как коровье бешенство у крупного рогатого скота, также включает накопление амилоидных бляшек, аналогичных болезни Альцгеймера.В эту группу также могут попадать заболевания других систем органов, например муковисцидоз или диабет 2 типа. Признание взаимосвязи между этими заболеваниями открыло новые терапевтические возможности. Вмешательство в накопление белков и, возможно, уже на их первоначальном производстве в клетке, может открыть новые способы облегчения этих разрушительных заболеваний.
Вопросы для самопроверки
Пройдите тест ниже, чтобы проверить свое понимание коммуникации между нейронами:
Возможности действия | Анатомия и физиология I
Цели обучения
- Опишите компоненты мембраны, которые определяют мембранный потенциал покоя
- Опишите изменения, происходящие с мембраной, которые приводят к потенциалу действия
Функции нервной системы — ощущение, интеграция и реакция — зависят от функций нейронов, лежащих в основе этих путей.Чтобы понять, как нейроны могут общаться, необходимо описать роль возбудимой мембраны в генерации этих сигналов. В основе этой коммуникации лежит потенциал действия, который демонстрирует, как изменения в мембране могут составлять сигнал. Рассмотрение того, как эти сигналы работают в более изменчивых обстоятельствах, предполагает рассмотрение градуированных потенциалов, которые будут рассмотрены в следующем разделе.
Электрически активные клеточные мембраны
Большинство клеток тела используют заряженные частицы, ионы, для накопления заряда на клеточной мембране.Ранее было показано, что это часть работы мышечных клеток. Для сокращения скелетных мышц, основанного на взаимодействии возбуждения и сокращения, требуется входной сигнал от нейрона. Обе клетки используют клеточную мембрану для регулирования движения ионов между внеклеточной жидкостью и цитозолем.
Как вы узнали из главы о клетках, клеточная мембрана в первую очередь отвечает за регулирование того, что может пересекать мембрану, а что остается только с одной стороны. Клеточная мембрана представляет собой бислой фосфолипидов, поэтому только вещества, которые могут проходить непосредственно через гидрофобное ядро, могут диффундировать без посторонней помощи.Заряженные частицы, которые по определению являются гидрофильными, не могут проходить через клеточную мембрану без посторонней помощи (рис. 1). Трансмембранные белки, в частности канальные белки, делают это возможным. Несколько каналов, а также специализированные энергозависимые «ионные насосы» необходимы для генерации трансмембранного потенциала и для генерации потенциала действия. Особый интерес представляет белок-носитель, называемый натриево-калиевым насосом, который перемещает ионы натрия (Na + ) из клетки, а ионы калия (K + ) в клетку, тем самым регулируя концентрацию ионов с обеих сторон. клеточная мембрана.
Рис. 1. Клеточная мембрана и трансмембранные белки Клеточная мембрана состоит из фосфолипидного бислоя и имеет множество трансмембранных белков, включая различные типы канальных белков, которые служат ионными каналами.
Натриево-калиевый насос требует энергии в виде аденозинтрифосфата (АТФ), поэтому его также называют АТФазой. Как было объяснено в главе о ячейке, концентрация Na + выше вне ячейки, чем внутри, а концентрация K + выше внутри ячейки выше, чем снаружи.Это означает, что этот насос перемещает ионы против градиентов концентрации натрия и калия, поэтому он требует энергии. Фактически, насос в основном поддерживает эти градиенты концентрации.
Ионные каналы — это поры, которые позволяют определенным заряженным частицам пересекать мембрану в ответ на существующий градиент концентрации. Белки способны проникать через клеточную мембрану, включая ее гидрофобное ядро, и могут взаимодействовать с зарядом ионов из-за различных свойств аминокислот, обнаруженных в определенных доменах или областях белкового канала.Гидрофобные аминокислоты находятся в доменах, которые примыкают к углеводородным хвостам фосфолипидов. Гидрофильные аминокислоты подвергаются воздействию жидкой среды внеклеточной жидкости и цитозоля. Кроме того, ионы будут взаимодействовать с гидрофильными аминокислотами, которые будут селективными в отношении заряда иона. Каналы для катионов (положительных ионов) будут иметь отрицательно заряженные боковые цепи в порах. Каналы для анионов (отрицательных ионов) будут иметь в порах положительно заряженные боковые цепи.Это называется электрохимическим исключением , что означает, что поры канала зависят от заряда.
Ионов также можно указать по диаметру поры. Расстояние между аминокислотами зависит от диаметра иона, когда он отделяется от окружающих его молекул воды. Из-за окружающих молекул воды более крупные поры не идеальны для более мелких ионов, потому что молекулы воды будут взаимодействовать посредством водородных связей более легко, чем боковые цепи аминокислот.Это называется исключением размера . Некоторые ионные каналы избирательны по заряду, но не обязательно по размеру, и поэтому называются неспецифическим каналом . Эти неспецифические каналы позволяют катионам, особенно Na + , K + и Ca 2+ , проходить через мембрану, но исключают анионы.
Ионные каналы не всегда позволяют ионам свободно диффундировать через мембрану. Они открываются определенными событиями, то есть каналы закрыты .Таким образом, еще один способ классификации каналов — это их стробирование. Хотя эти классы ионных каналов находятся в основном в клетках нервной или мышечной ткани, их также можно найти в клетках эпителиальной и соединительной ткани.
Управляемый лигандом канал открывается, потому что сигнальная молекула, лиганд, связывается с внеклеточной областью канала. Этот тип канала также известен как ионотропный рецептор , потому что, когда лиганд, известный как нейротрансмиттер в нервной системе, связывается с белком, ионы пересекают мембрану, изменяя ее заряд (рис. 2).
Рис. 2. Каналы, управляемые лигандами Когда лиганд, в данном случае нейротрансмиттер ацетилхолин, связывается с определенным местом на внеклеточной поверхности белка канала, пора открывается, позволяя отобрать ионы. Ионы в данном случае представляют собой катионы натрия, кальция и калия.
Канал с механическим затвором, открывается из-за физического искажения клеточной мембраны. Многие каналы, связанные с осязанием (соматосенсорное ощущение), закрываются механически.Например, когда на кожу оказывается давление, эти каналы открываются и позволяют ионам проникать в клетку. Подобным каналу этого типа может быть канал, который открывается при изменении температуры, как при тестировании воды в душе (рис. 3).
Рис. 3. Механически закрытые каналы Когда в окружающей ткани происходит механическое изменение, такое как давление или прикосновение, канал физически открывается. Терморецепторы работают по аналогичному принципу. Когда местная температура ткани изменяется, белок реагирует, физически открывая канал.
Управляемый по напряжению канал — это канал, который реагирует на изменения электрических свойств мембраны, в которую он встроен. Обычно внутренняя часть мембраны находится под отрицательным напряжением. Когда это напряжение становится менее отрицательным, канал начинает позволять ионам пересекать мембрану (рис. 4).
Рис. 4. Каналы с ограничением по напряжению Каналы с ограничением по напряжению открываются, когда вокруг них изменяется трансмембранное напряжение. Аминокислоты в структуре белка чувствительны к заряду и заставляют поры открываться для выбранного иона.
Канал утечки имеет случайное закрытие, что означает, что он открывается и закрывается случайным образом, отсюда и ссылка на утечку. Нет фактического события, открывающего канал; вместо этого он имеет внутреннюю скорость переключения между открытым и закрытым состояниями. Каналы утечки вносят вклад в трансмембранное напряжение покоя возбудимой мембраны (рис. 5).
Рис. 5. Каналы утечки В определенных ситуациях ионы должны беспорядочно перемещаться через мембрану.Конкретные электрические свойства определенных клеток изменяются наличием этого типа канала.
Мембранный потенциал
Электрическое состояние клеточной мембраны может иметь несколько вариаций. Все это вариации мембранного потенциала . Потенциал — это распределение заряда через клеточную мембрану, измеряемое в милливольтах (мВ). Стандарт заключается в сравнении внутренней части клетки с внешней, поэтому мембранный потенциал — это величина, представляющая заряд на внутриклеточной стороне мембраны, исходя из того, что внешняя сторона равна нулю, условно говоря (рис. 6).
Рис. 6. Измерение заряда на мембране с помощью вольтметра Регистрирующий электрод вставлен в ячейку, а электрод сравнения находится вне ячейки. Путем сравнения заряда, измеренного этими двумя электродами, определяется трансмембранное напряжение. Это значение обычно выражается для цитозоля относительно внешней среды.
Концентрация ионов во внеклеточной и внутриклеточной жидкости в значительной степени сбалансирована с чистым нейтральным зарядом. Однако небольшая разница в заряде возникает прямо на поверхности мембраны как внутри, так и снаружи.Разница в этой очень ограниченной области, которая имеет всю способность нейронов (и мышечных клеток) генерировать электрические сигналы, включая потенциалы действия.
Прежде чем описывать эти электрические сигналы, необходимо объяснить состояние покоя мембраны. Когда ячейка находится в состоянии покоя и ионные каналы закрыты (за исключением каналов утечки, которые открываются случайным образом), ионы распределяются по мембране очень предсказуемым образом. Концентрация Na + вне клетки в 10 раз больше, чем концентрация внутри.Кроме того, концентрация K + внутри ячейки больше, чем снаружи. Цитозоль содержит высокую концентрацию анионов в форме фосфат-ионов и отрицательно заряженных белков. Крупные анионы являются компонентом внутренней клеточной мембраны, включая специализированные фосфолипиды и белки, связанные с внутренним листком мембраны (листок — это термин, используемый для обозначения одной стороны двухслойной липидной мембраны). Отрицательный заряд локализован в больших анионах.
Когда ионы распределены по мембране при этих концентрациях, разница в заряде измеряется при -70 мВ, значение, описываемое как потенциал мембраны в состоянии покоя .Точное значение, измеренное для мембранного потенциала покоя, варьируется от клетки к клетке, но чаще всего используется значение -70 мВ. Это напряжение на самом деле было бы намного ниже, если бы не вклад некоторых важных белков в мембрану. Каналы утечки позволяют Na + медленно перемещаться в ячейку или K + медленно перемещаться наружу, а насос Na + / K + восстанавливает их. Это может показаться пустой тратой энергии, но каждый из них играет определенную роль в поддержании мембранного потенциала.
Возможности действия
Мембранный потенциал покоя описывает устойчивое состояние клетки, которое представляет собой динамический процесс, который уравновешивается утечкой и накачкой ионов. Без какого-либо внешнего влияния это не изменится. Чтобы запустить электрический сигнал, мембранный потенциал должен измениться.
Это начинается с открытия канала для Na + в мембране. Поскольку концентрация Na + выше вне ячейки, чем внутри ячейки в 10 раз, ионы будут устремляться в ячейку, что в значительной степени обусловлено градиентом концентрации.Поскольку натрий является положительно заряженным ионом, он изменяет относительное напряжение непосредственно внутри ячейки по сравнению с непосредственно снаружи. Потенциал покоя — это состояние мембраны при напряжении -70 мВ, поэтому катион натрия, попадающий в ячейку, сделает ее менее отрицательной. Это известно как деполяризация , что означает, что мембранный потенциал приближается к нулю.
Градиент концентрации Na + настолько велик, что он будет продолжать поступать в клетку даже после того, как мембранный потенциал станет нулевым, так что напряжение непосредственно вокруг поры начинает становиться положительным.Электрический градиент также играет роль, поскольку отрицательные белки под мембраной притягивают ион натрия. Мембранный потенциал достигнет +30 мВ к тому моменту, когда натрий попадет в ячейку.
Когда мембранный потенциал достигает +30 мВ, в мембране открываются другие потенциалозависимые каналы. Эти каналы специфичны для иона калия. Градиент концентрации действует и на K + . Когда K + начинает покидать ячейку, унося с собой положительный заряд, мембранный потенциал начинает возвращаться к своему напряжению покоя.Это называется реполяризацией , что означает, что мембранное напряжение возвращается к значению -70 мВ потенциала покоя мембраны.
Реполяризация возвращает мембранный потенциал к значению -70 мВ, которое указывает потенциал покоя, но на самом деле оно превышает это значение. Ионы калия достигают равновесия, когда напряжение на мембране ниже -70 мВ, поэтому возникает период гиперполяризации, когда каналы K + открыты. Эти каналы K + закрываются с небольшой задержкой из-за этого короткого выброса.
Рис. 7. График потенциала действия График зависимости напряжения, измеренного на клеточной мембране от времени, потенциал действия начинается с деполяризации, за которой следует реполяризация, которая переходит за потенциал покоя в гиперполяризацию, и, наконец, мембрана возвращается в состояние покоя.
Здесь был описан потенциал действия, который представлен в виде графика зависимости напряжения от времени на рисунке 7. Это электрический сигнал, который нервная ткань генерирует для связи.Изменение мембранного напряжения от -70 мВ в состоянии покоя до +30 мВ в конце деполяризации представляет собой изменение на 100 мВ. Это также можно записать как изменение на 0,1 В.
Чтобы оценить эту ценность в перспективе, подумайте о батарее. Батарея AA, которую вы можете найти в пульте дистанционного управления телевизором, имеет напряжение 1,5 В, или батарея 9 В (прямоугольная батарея с двумя штырями на одном конце), очевидно, составляет 9 В. Изменение потенциала действия составляет на один-два порядка меньше заряда в этих аккумуляторах.Фактически, мембранный потенциал можно описать как батарею. Через мембрану накапливается заряд, который может высвобождаться при правильных условиях. Батарея вашего пульта ДУ накопила заряд, который «высвобождается», когда вы нажимаете кнопку.
То, что происходит через мембрану электрически активной ячейки, представляет собой динамический процесс, который трудно визуализировать с помощью статических изображений или текстовых описаний. Просмотрите эту анимацию, чтобы узнать больше об этом процессе.
В чем разница между движущей силой Na + и K + ? А что общего в движении этих двух ионов?
Теперь вопрос в том, что инициирует потенциал действия? Приведенное выше описание удобно замалчивает этот момент.Но очень важно понимать, что происходит. Мембранный потенциал будет оставаться на уровне напряжения покоя, пока что-то не изменится. В приведенном выше описании просто говорится, что открывается канал Na + . Сказать «канал открывается» не означает, что изменяется отдельный трансмембранный белок. Вместо этого это означает, что открывается один вид канала. Есть несколько различных типов каналов, которые позволяют Na + проходить через мембрану. Управляемый лигандом канал Na + откроется, когда с ним свяжется нейромедиатор, и канал Na + , управляемый механически, откроется, когда физический стимул воздействует на сенсорный рецептор (например, давление, приложенное к коже, сжимает рецептор прикосновения).Будь то связывание нейротрансмиттера с его рецепторным белком или сенсорный стимул, активирующий сенсорную рецепторную клетку, некоторый стимул запускает процесс. Натрий начинает поступать в клетку, и мембрана становится менее отрицательной.
Третий тип канала, который играет важную роль в деполяризации потенциала действия, — это потенциалзависимый канал Na + . Каналы, которые начинают деполяризовать мембрану из-за раздражителя, помогают клетке деполяризоваться от -70 мВ до -55 мВ.Как только мембрана достигает этого напряжения, открываются управляемые по напряжению каналы Na + . Это то, что называется порогом. Любая деполяризация, которая не изменяет мембранный потенциал до -55 мВ или выше, не достигнет порога и, следовательно, не приведет к потенциалу действия. Кроме того, любой стимул, который деполяризует мембрану до -55 мВ или выше, вызовет открытие большого количества каналов и инициирование потенциала действия.
Из-за порога потенциал действия можно сравнить с цифровым событием — оно либо происходит, либо нет.Если порог не достигнут, потенциал действия не возникает. Если деполяризация достигает -55 мВ, тогда потенциал действия продолжается и достигает +30 мВ, при котором K + вызывает реполяризацию, включая гиперполяризационный выброс. Кроме того, эти изменения одинаковы для каждого потенциала действия, а это означает, что при достижении порога происходит то же самое. Более сильный стимул, который может деполяризовать мембрану далеко за порог, не приведет к «большему» потенциалу действия.Потенциалы действия бывают «все или ничего». Либо мембрана достигает порога, и все происходит, как описано выше, либо мембрана не достигает порога, и больше ничего не происходит. Все потенциалы действия достигают пика при одном и том же напряжении (+30 мВ), поэтому один потенциал действия не больше другого. Более сильные стимулы быстрее инициируют множественные потенциалы действия, но отдельные сигналы не сильнее. Таким образом, например, вы не почувствуете более сильного болевого ощущения или не почувствуете более сильного сокращения мышц из-за величины потенциала действия, поскольку они не различаются по размеру.
Как мы видели, деполяризация и реполяризация потенциала действия зависят от двух типов каналов (потенциал-зависимый канал Na + и потенциал-управляемый канал K + ). Управляемый по напряжению канал Na + фактически имеет два затвора. Одним из них является активационный вентиль , который открывается, когда мембранный потенциал пересекает -55 мВ. Другой вентиль — вентиль инактивации , который закрывается через определенный период времени — порядка долей миллисекунды.Когда ячейка находится в состоянии покоя, ворота активации закрыты, а ворота дезактивации открыты. Однако при достижении порога активирующие ворота открываются, позволяя Na + ворваться в ячейку. В момент пика деполяризации ворота инактивации закрываются. Во время реполяризации в клетку больше не может попасть натрий. Когда мембранный потенциал снова превышает -55 мВ, активирующий вентиль закрывается. После этого ворота инактивации снова открываются, делая канал готовым к повторному запуску всего процесса.
Управляемый по напряжению канал K + имеет только один затвор, чувствительный к мембранному напряжению -50 мВ. Однако он не открывается так быстро, как закрытый по напряжению канал Na + . Открытие канала после достижения этого напряжения может занять доли миллисекунды. Время этого точно совпадает с пиками потока Na + , так что управляемые по напряжению каналы K + открываются, как только управляемые по напряжению каналы Na + деактивируются.Когда мембранный потенциал переполяризуется и напряжение снова становится равным -50 мВ, канал закрывается — снова с небольшой задержкой. Калий продолжает покидать клетку в течение короткого времени, и мембранный потенциал становится более отрицательным, что приводит к гиперполяризационному выбросу. Затем канал снова закрывается, и мембрана может вернуться в состояние покоя из-за продолжающейся активности не закрытых каналов и насоса Na + / K + .
Рисунок 8. Этапы потенциала действия
Все это происходит примерно за 2 миллисекунды (Рисунок 8).Пока существует потенциал действия, другой не может быть инициирован. Этот эффект называется рефрактерным периодом . Есть две фазы огнеупорного периода: абсолютный огнеупорный период и относительный огнеупорный период . Во время абсолютной фазы другой потенциал действия не запускается. Это происходит из-за затвора инактивации управляемого по напряжению канала Na + . Как только этот канал вернется в состояние покоя (менее -55 мВ), новый потенциал действия может быть запущен, но только более сильным стимулом, чем тот, который инициировал текущий потенциал действия.Это происходит из-за вытекания K + из ячейки. Поскольку этот ион устремляется наружу, любой Na + , который пытается войти, не будет деполяризовать клетку, а только будет удерживать клетку от гиперполяризации.
График зависимости напряжения, измеренного на клеточной мембране, от времени (как показано на рисунке 8), события потенциала действия могут быть связаны с конкретными изменениями мембранного напряжения.
- В состоянии покоя напряжение на мембране -70 мВ.
- Мембрана начинает деполяризоваться при воздействии внешнего раздражителя.
- Напряжение на мембране начинает быстро расти до +30 мВ.
- Напряжение мембраны начинает возвращаться к отрицательному значению.
- Реполяризация продолжается после напряжения покоя мембраны, что приводит к гиперполяризации.
- Напряжение мембраны возвращается к исходному значению вскоре после гиперполяризации.
Распространение потенциала действия
Потенциал действия инициируется в начале аксона, в так называемом начальном сегменте.Существует высокая плотность управляемых по напряжению каналов Na + , так что здесь может иметь место быстрая деполяризация. Спускаясь вниз по длине аксона, потенциал действия распространяется, потому что по мере распространения деполяризации открывается больше потенциал-управляемых каналов Na + . Это распространение происходит из-за того, что Na + проникает через канал и движется по внутренней части клеточной мембраны. Когда Na + перемещается или проходит небольшое расстояние вдоль клеточной мембраны, его положительный заряд деполяризует немного больше клеточной мембраны.По мере того, как эта деполяризация распространяется, открываются новые потенциалозависимые каналы Na + , и все больше ионов устремляется в клетку, распространяя деполяризацию немного дальше.
Поскольку управляемые по напряжению каналы Na + инактивируются на пике деполяризации, они не могут быть открыты снова на короткое время — период абсолютной рефрактерности. Из-за этого деполяризация, распространяющаяся обратно на ранее открытые каналы, не имеет никакого эффекта. Потенциал действия должен распространяться к окончанию аксона; в результате полярность нейрона сохраняется, как упоминалось выше.
Распространение, как описано выше, применимо к немиелинизированным аксонам. Когда присутствует миелинизация, потенциал действия распространяется иначе. Ионы натрия, которые входят в клетку в начальном сегменте, начинают распространяться по длине сегмента аксона, но до первого узла Ранвье нет управляемых по напряжению каналов Na + . Поскольку не существует постоянного открытия этих каналов вдоль сегмента аксона, деполяризация распространяется с оптимальной скоростью. Расстояние между узлами — это оптимальное расстояние, чтобы мембрана оставалась деполяризованной выше порога в следующем узле.Когда Na + распространяется по внутренней части мембраны сегмента аксона, заряд начинает рассеиваться. Если бы узел находился дальше по аксону, эта деполяризация упала бы слишком сильно для того, чтобы управляемые по напряжению каналы Na + были активированы в следующем узле Ранвье. Если бы узлы были ближе друг к другу, скорость распространения была бы ниже.
Распространение по немиелинизированному аксону обозначается как с непрерывной проводимостью ; По длине миелинизированного аксона она составляет скачкообразной проводимости .Непрерывная проводимость происходит медленно, потому что всегда открываются управляемые по напряжению каналы Na + , и все больше и больше Na + устремляется в ячейку. Солевое проведение происходит быстрее, потому что потенциал действия в основном перескакивает от одного узла к другому ( saltare = «прыгать»), а новый приток Na + обновляет деполяризованную мембрану. Наряду с миелинизацией аксона диаметр аксона может влиять на скорость проводимости. Подобно тому, как вода течет быстрее в широкой реке, чем в узком ручье, деполяризация на основе Na + распространяется быстрее по широкому аксону, чем по узкому.Эта концепция известна как сопротивление и в целом верна для электрических проводов или водопровода, так же как и для аксонов, хотя конкретные условия различаются в масштабах электронов или ионов по сравнению с водой в реке.
Гомеостатический дисбаланс: концентрация калия
Глиальные клетки, особенно астроциты, отвечают за поддержание химической среды ткани ЦНС. Концентрация ионов во внеклеточной жидкости является основой того, как устанавливается мембранный потенциал и изменяется электрохимическая сигнализация.Если баланс ионов нарушен, возможны тяжелые исходы.
Обычно концентрация K + выше внутри нейрона, чем снаружи. После фазы реполяризации потенциала действия каналы утечки K + и насос Na + / K + гарантируют, что ионы возвращаются в свои исходные положения. После инсульта или другого ишемического события уровни внеклеточного K + повышаются. Астроциты в этой области оборудованы для удаления излишков K + , чтобы помочь помпе.Но когда уровень далеко не сбалансирован, последствия могут быть необратимыми.
Астроциты могут стать реактивными в подобных случаях, что снижает их способность поддерживать локальную химическую среду. Глиальные клетки увеличиваются, и их отростки набухают. Они теряют свою буферную способность K + , и это влияет на работу насоса или даже отменяет его. Если градиент Na + нарушается, это имеет более важный эффект, чем прерывание потенциала действия. Транспорт глюкозы в клетки сочетается с ко-транспортом Na + .Когда это потеряно, клетка не может получить необходимую ей энергию. В центральной нервной системе метаболизм углеводов — единственный способ производства АТФ. В других частях тела клетки полагаются на углеводы, липиды или аминокислоты для выработки митохондриального АТФ. Но ЦНС не хранит липиды в адипоцитах (жировых клетках) в качестве энергетического резерва. Липиды в ЦНС находятся в клеточных мембранах нейронов и глиальных клеток, в частности, как неотъемлемый компонент миелина. Белки в ЦНС имеют решающее значение для функции нейронов в таких ролях, как каналы для передачи электрических сигналов или как часть цитоскелета.Эти макромолекулы не используются для выработки митохондриального АТФ в нейронах.
Посетите этот сайт, чтобы увидеть виртуальную лабораторию нейрофизиологии и понаблюдать за электрофизиологическими процессами в нервной системе, где ученые непосредственно измеряют электрические сигналы, производимые нейронами. Часто потенциалы действия возникают так быстро, что смотреть на экран, чтобы увидеть, как они возникают, бесполезно. Динамик приводится в действие сигналами, записанными от нейрона, и он «выскакивает» каждый раз, когда нейрон запускает потенциал действия.Эти потенциалы действия срабатывают так быстро, что по радио это звучит как статика. Электрофизиологи могут распознать закономерности в этой статике, чтобы понять, что происходит. Почему модель пиявки используется для измерения электрической активности нейронов, а не людей?Потенциал действия — определение, шаги, фазы
Автор:
Яна Васькович
•
Рецензент:
Франческа Сальвадор, магистр наук
Последняя редакция: 31 мая 2021 г.
Время чтения: 10 минут.
Долгое время процесс коммуникации между нервами и их тканями-мишенями был большим неизвестным для физиологов.С развитием электрофизиологии и открытием электрической активности нейронов было обнаружено, что передача сигналов от нейронов к тканям-мишеням опосредуется потенциалами действия.
Потенциал действия определяется как внезапное, быстрое, временное и распространяющееся изменение мембранного потенциала покоя. Только нейроны и мышечные клетки способны генерировать потенциал действия; это свойство называется возбудимостью .
| Определение | Внезапное, быстрое, преходящее и распространяющееся изменение мембранного потенциала покоя |
| Стимулы | Дополнительный порог Порог Suprathreshold |
| Фазы | Гипополяризация Деполяризация Overshoot Реполяризация |
| Огнеупорность | Абсолют — деполяризация, 2/3 реполяризации Относительно — последняя 1/3 реполяризации |
| Синапс | Пресинаптическая мембрана Синаптическая щель Постсинаптическая мембрана |
В этой статье мы обсудим определение, шаги и фазы потенциала действия.
Определение
Потенциалы действия — это нервные сигналы. Нейроны генерируют и проводят эти сигналы в своих процессах, чтобы передать их в ткани-мишени. При стимуляции они будут либо стимулироваться, либо подавляться, либо каким-либо образом модулироваться. структура и все типы нейронов со следующим блоком исследования.
Ступеньки
Но что вызывает потенциал действия? С электрической точки зрения, это вызвано стимулом с определенной величиной, выраженной в милливольтах [мВ].Не все стимулы могут вызывать потенциал действия. Адекватный стимул должен иметь достаточную электрическую ценность, которая снизит негативность нервной клетки до порога потенциала действия. Таким образом, существуют подпороговые, пороговые и надпороговые стимулы. Подпороговые стимулы не могут вызывать потенциал действия. Порог стимулы обладают достаточной энергией или потенциалом для создания потенциала действия (нервного импульса). Suprathreshold Стимулы также производят потенциал действия, но их сила выше, чем пороговые стимулы.
Итак, потенциал действия генерируется, когда стимул изменяет мембранный потенциал до значений порогового потенциала . Пороговый потенциал обычно составляет от -50 до -55 мВ. Важно знать, что потенциал действия ведет себя по закону «все или ничего» . Это означает, что любой подпороговый стимул ничего не вызовет, в то время как пороговые и надпороговые стимулы вызывают полную реакцию возбудимой клетки.
Отличается ли потенциал действия в зависимости от того, вызван ли он пороговым или надпороговым потенциалом? Ответ — нет.Длина и амплитуда потенциала действия всегда одинаковы. Однако увеличение силы стимула вызывает увеличение частоты потенциала действия. Потенциал действия распространяется по нервному волокну без уменьшения или ослабления амплитуды и длины. Кроме того, после того, как генерируется один потенциал действия, нейроны становятся невосприимчивыми к стимулам в течение определенного периода времени, в течение которого они не могут генерировать другой потенциал действия.
Фазы
С точки зрения ионов, потенциал действия вызывается временными изменениями проницаемости мембраны для диффундирующих ионов.Эти изменения вызывают открытие ионных каналов и уменьшение градиентов концентрации ионов. Величина порогового потенциала зависит от проницаемости мембраны, внутри- и внеклеточной концентрации ионов и свойств клеточной мембраны.
Потенциал действия имеет несколько фаз ; гипополяризация, деполяризация, перерегулирование, реполяризация и гиперполяризация.
Кривая и фазы потенциала действия (диаграмма)Гипополяризация — это начальное увеличение мембранного потенциала до значения порогового потенциала.Пороговый потенциал открывает управляемые по напряжению натриевые каналы и вызывает большой приток ионов натрия. Эта фаза называется деполяризацией . Во время деполяризации внутренняя часть ячейки становится все более и более электроположительной, пока потенциал не приблизится к электрохимическому равновесию для натрия +61 мВ. Эта крайняя положительная фаза — это фаза выброса .
После перерегулирования проницаемость для натрия внезапно снижается из-за закрытия его каналов.Превышение потенциала ячейки открывает управляемые по напряжению калиевые каналы, что вызывает большой отток калия, снижая электроположительность ячейки. Эта фаза представляет собой фазу реполяризации , цель которой — восстановить мембранный потенциал покоя. Реполяризация всегда сначала приводит к гиперполяризации , состоянию, в котором мембранный потенциал более отрицательный, чем мембранный потенциал по умолчанию. Но вскоре после этого мембрана снова устанавливает значения мембранного потенциала.
Обмен ионами в потенциале действия (диаграмма)После рассмотрения роли ионов мы можем теперь определить пороговый потенциал более точно как значение мембранного потенциала, при котором открываются потенциалозависимые натриевые каналы. В возбудимых тканях пороговый потенциал примерно на 10-15 мВ меньше мембранного потенциала покоя.
Огнеупорный период
Рефрактерный период — это время после генерирования потенциала действия, в течение которого возбудимая клетка не может производить другой потенциал действия.Есть две субфазы этого периода: абсолютная и относительная тугоплавкость.
Абсолютная рефрактерность перекрывает деполяризацию и примерно 2/3 фазы реполяризации. Новый потенциал действия не может быть сгенерирован во время деполяризации, потому что все потенциалозависимые натриевые каналы уже открыты или открываются с максимальной скоростью. Во время ранней реполяризации новый потенциал действия невозможен, поскольку натриевые каналы неактивны и нуждаются в потенциале покоя, чтобы быть в закрытом состоянии, из которого они снова могут быть в открытом состоянии.Абсолютная рефрактерность заканчивается, когда достаточное количество натриевых каналов восстанавливается из своего неактивного состояния.
Относительная рефрактерность — это период, когда генерация нового потенциала действия возможна, но только при надпороговом стимуле. Этот период перекрывает последнюю 1/3 реполяризации.
Распространение потенциала действия
В теле нейрона генерируется потенциал действия, который распространяется через его аксон.Распространение никоим образом не снижает и не влияет на качество потенциала действия, поэтому ткани-мишени получают один и тот же импульс независимо от того, насколько далеко они находятся от тела нейрона.
Потенциал действия создается в одном месте клеточной мембраны. Он распространяется по мембране, причем каждая последующая часть мембраны последовательно деполяризуется. Это означает, что потенциал действия не перемещает , а скорее вызывает новый потенциал действия соседнего сегмента нейрональной мембраны.
Необходимо подчеркнуть, что потенциал действия всегда распространяется вперед, , но никогда назад. Это связано с рефрактерностью частей мембраны, которые уже были деполяризованы, так что единственное возможное направление распространения — вперед. Из-за этого потенциал действия всегда распространяется от тела нейрона через аксон к ткани-мишени.
Скорость распространения во многом зависит от толщины аксона и от того, является ли он миелинизированным или нет.Чем больше диаметр, тем выше скорость распространения. Распространение также происходит быстрее, если аксон миелинизирован. Миелин увеличивает скорость распространения, поскольку увеличивает толщину волокна. Кроме того, миелин обеспечивает скачкообразное проведение потенциала действия, поскольку деполяризуются только узлы Ранвье, а миелиновые узлы перепрыгивают.
В немиелинизированных волокнах каждая часть аксональной мембраны должна подвергаться деполяризации, что значительно замедляет распространение.
Вы хотите быстрее изучить все части и функции нервной системы? Перейдите к нашей статье викторины по нервной системе и успешно сдайте следующий экзамен.
Синапс
Синапс — это соединение между нервной клеткой и тканью-мишенью. У людей синапсы химические , что означает, что нервный импульс передается от конца аксона к ткани-мишени с помощью химических веществ, называемых нейротрансмиттерами (лигандами).Если нейротрансмиттер стимулирует клетку-мишень к действию, то это нейротрансмиттер возбуждения. С другой стороны, если он ингибирует клетку-мишень, он является тормозным нейромедиатором.
Типы нейронов и синапсов (диаграмма)В зависимости от типа ткани-мишени различают центральные и периферические синапсы. Центральные синапсы находятся между двумя нейронами в центральной нервной системе, в то время как периферические синапсы возникают между нейроном и мышечным волокном, периферическим нервом или железой.
Каждый синапс состоит из:
- Пресинаптическая мембрана — мембрана терминальной кнопки нервного волокна
- Постсинаптическая мембрана — мембрана клетки-мишени
- Синаптическая щель — разрыв между пресинаптической и постсинаптической мембранами
Внутри терминальной кнопки нервного волокна образуются и хранятся многочисленные пузырьки, содержащие нейротрансмиттеры. Когда пресинаптическая мембрана деполяризуется под действием потенциала действия, потенциал-зависимые каналы кальция открываются.Это приводит к притоку кальция, который изменяет состояние определенных мембранных белков в пресинаптической мембране и приводит к экзоцитозу нейромедиатора в синаптической щели.
Постсинаптическая мембрана содержит рецепторы для нейромедиаторов. Как только нейромедиатор связывается с рецептором, лиганд-зависимые каналы постсинаптической мембраны открываются или закрываются. Эти закрытые лигандами каналы являются ионными каналами, и их открытие или закрытие вызывает перераспределение ионов в постсинаптической клетке.В зависимости от того, является ли нейротрансмиттер возбуждающим или тормозящим, это будет иметь разные ответы.
Изучите типы нейронов с помощью следующей викторины.
Сводка
Потенциал действия вызывается пороговыми или надпороговыми стимулами нейрона. Он состоит из четырех фаз; гипополяризация, деполяризация, выброс и реполяризация.
Потенциал действия распространяется вдоль клеточной мембраны аксона, пока не достигнет конечной кнопки.Как только терминальная кнопка деполяризуется, она высвобождает нейромедиатор в синаптическую щель. Нейромедиатор связывается со своими рецепторами на постсинаптической мембране клетки-мишени, вызывая ее ответ либо в виде стимуляции, либо ингибирования.
Потенциалы действия распространяются быстрее через более толстые и миелинизированные аксоны, чем через тонкие и немиелинизированные аксоны. После того, как генерируется один потенциал действия, нейрон не может генерировать новый из-за своей рефрактерности к стимулам.
потенциалов покоя и потенциалов действия (Раздел 1, Глава 1) Нейронаука в Интернете: Электронный учебник для нейронаук | Кафедра нейробиологии и анатомии
| Видео лекции |
Несмотря на огромную сложность мозга, можно получить представление о его функциях, обратив внимание на две основные детали:
- Во-первых, способы, которыми отдельные нейроны, компоненты нервной системы, связаны друг с другом для формирования поведения.
- Во-вторых, биофизические, биохимические и электрофизиологические свойства отдельных нейронов.
Хорошее место для начала — это компоненты нервной системы и то, как электрические свойства нейронов наделяют нервные клетки способностью обрабатывать и передавать информацию.
1.1 Введение в потенциал действий
|
Рисунок 1.1 |
Теории кодирования и передачи информации в нервной системе восходят к греческому врачу Галену (129–210 гг. Н.э.), который предложил гидравлический механизм, с помощью которого мышцы сокращаются, потому что жидкость течет в них из полых нервов. Основная теория существовала веками и была далее развита Рене Декартом (1596 — 1650), который предположил, что духи животных текут из мозга через нервы, а затем в мышцы, чтобы производить движения (см. Эту анимацию для современной интерпретации такой гидравлической теории для нервов). функция).Главный сдвиг парадигмы произошел с новаторской работой Луиджи Гальвани, который в 1794 году обнаружил, что нервы и мышцы могут активироваться заряженными электродами, и предположил, что нервная система функционирует посредством передачи электрических сигналов. Однако среди ученых были споры о том, находится ли электричество в нервах и мышцах или нервы и мышцы просто реагируют на вредный электрический шок через некий внутренний неэлектрический механизм. Проблема не была решена до 1930-х годов, когда были разработаны современные электронные усилители и записывающие устройства, которые позволили записывать электрические сигналы.Одним из примеров является новаторская работа Х.К. Хартлайн 80 лет назад об электрических сигналах на подковообразном крабе Limulus. Электроды помещали на поверхность зрительного нерва. (Поместив электроды на поверхность нерва, можно получить индикацию изменений мембранного потенциала, которые происходят между внешней и внутренней частью нервной клетки.) Затем были вспышки света различной интенсивности длительностью 1 с. представлен глазу; сначала тусклый свет, затем более яркий свет. Очень тусклый свет не влиял на активность, но более яркий свет производил небольшие повторяющиеся всплески.Эти шиповидные события называются потенциалами действия, нервными импульсами или иногда просто всплесками. Потенциалы действия — это основные события, которые нервные клетки используют для передачи информации из одного места в другое.
1.2 Характеристики потенциалов действия
Записи на рисунке выше иллюстрируют три очень важных характеристики потенциалов нервного действия. Первый , нервный потенциал действия имеет короткую продолжительность (около 1 мс). Второй, , потенциалы нервного действия выявляются по принципу «все или ничего». Третий , нервные клетки кодируют интенсивность информации частотой потенциалов действия. Когда интенсивность стимула увеличивается, величина потенциала действия не увеличивается. Скорее частота или количество потенциалов действия увеличивается. В целом, чем выше интенсивность стимула (будь то световой стимул к фоторецептору, механический стимул к коже или растяжение мышечного рецептора), тем большее количество вызванных потенциалов действия.Точно так же для двигательной системы, чем больше количество потенциалов действия в двигательном нейроне, тем выше интенсивность сокращения мышцы, которая иннервируется этим двигательным нейроном.
Потенциалы действия имеют большое значение для функционирования мозга, поскольку они передают информацию из нервной системы в центральную нервную систему и передают команды, инициированные в центральной нервной системе, на периферию. Следовательно, необходимо досконально разбираться в их свойствах.Чтобы ответить на вопросы о том, как потенциалы действия инициируются и распространяются, нам необходимо записать потенциал между внутренней и внешней стороной нервных клеток, используя методы внутриклеточной записи.
1.3 Внутриклеточные записи нейронов
Разность потенциалов на мембране нервной клетки может быть измерена с помощью микроэлектрода , кончик которого настолько мал (около микрона), что он может проникать в клетку без каких-либо повреждений. Когда электрод находится в ванне (внеклеточная среда), потенциал не регистрируется, потому что ванна изопотенциальна.Если аккуратно ввести микроэлектрод в ячейку, происходит резкое изменение потенциала. Показания вольтметра мгновенно изменяются от 0 мВ до показания разности потенциалов -60 мВ внутри ячейки по отношению к внешней стороне. Потенциал, который регистрируется, когда живая клетка пронизана микроэлектродом, называется потенциалом покоя и варьируется от клетки к клетке. Здесь показано, что оно составляет -60 мВ, но может находиться в диапазоне от -80 до -40 мВ, в зависимости от конкретного типа нервной клетки.В отсутствие стимуляции потенциал покоя обычно постоянен.
Также можно записывать и изучать потенциал действия. На рис. 1.3 показан пример, в котором нейрон уже пронизан одним микроэлектродом (регистрирующим электродом), подключенным к вольтметру. Электрод регистрирует потенциал покоя -60 мВ. В ячейку также насаживают второй электрод, называемый стимулирующим электродом. Этот электрод подключен к батарее и устройству, которое может контролировать величину тока (I), протекающего через электрод.Изменения мембранного потенциала вызываются замыканием переключателя и систематическим изменением размера и полярности батареи. Если отрицательный полюс батареи подключен к внутренней части ячейки, как показано на рисунке 1.3A, мгновенное изменение величины тока будет проходить через стимулирующий электрод, и мембранный потенциал временно станет более отрицательным. Такой результат не должен вызывать удивления. Отрицательный полюс батареи делает внутреннюю часть элемента более отрицательной, чем это было раньше.Изменение потенциала, которое увеличивает поляризованное состояние мембраны, называется гиперполяризацией . Клетка более поляризована, чем обычно. Используйте еще большую батарею, и потенциал станет еще больше. Результирующие гиперполяризации являются градуированными функциями величины стимулов, используемых для их создания.
Теперь рассмотрим случай, когда положительный полюс батареи подключен к электроду (рисунок 1.3B). Когда положительный полюс батареи подключен к электроду, потенциал ячейки становится более положительным, когда переключатель замкнут (Рисунок 1.3Б). Такие потенциалы называются деполяризациями . Поляризованное состояние мембраны уменьшается. Батареи большего размера вызывают еще большую деполяризацию. Опять же, величина ответов пропорциональна величине стимулов. Однако необычное событие происходит, когда величина деполяризации достигает уровня мембранного потенциала, называемого порогом . Инициируется совершенно новый тип сигнала; потенциал действия. Обратите внимание, что если размер батареи увеличить еще больше, амплитуда потенциала действия будет такой же, как и у предыдущего (Рисунок 1.3Б). Процесс выявления потенциала действия в нервной клетке аналогичен зажиганию предохранителя с помощью источника тепла. Необходима определенная минимальная температура (порог). Температуры ниже порогового значения не приводят к воспламенению предохранителя. Температура выше порога воспламеняет предохранитель так же, как и пороговая температура, и предохранитель не горит ни ярче, ни горячее.
Однако, если надпороговое воздействие на ток достаточно длительное, будет вызвана серия потенциалов действия.В общем, потенциалы действия будут продолжать действовать, пока действует стимул, при этом частота возбуждения пропорциональна величине стимула (рис. 1.4).
Потенциалы действия не только инициируются по принципу «все или ничего», но они также распространяются по принципу «все или ничего». Потенциал действия, инициированный в клеточном теле моторного нейрона в спинном мозге, будет неукрепленным образом распространяться до синаптических окончаний этого моторного нейрона.Опять же, ситуация аналогична горящему запалу. После воспламенения предохранителя пламя распространится до конца.
1.4 Составляющие потенциала действия
Потенциал действия состоит из нескольких компонентов (рис. 1.3B). Порог — это значение мембранного потенциала, при достижении которого происходит полное инициирование потенциала действия. Начальная или возрастающая фаза потенциала действия называется фазой деполяризации или ходом вверх .Область потенциала действия между уровнем 0 мВ и максимальной амплитудой составляет выброс . Возврат мембранного потенциала к потенциалу покоя называется фазой реполяризации . Существует также фаза потенциала действия, в течение которой мембранный потенциал может быть более отрицательным, чем потенциал покоя. Эта фаза потенциала действия называется понижением или гиперполяризационным потенциалом . На рисунке 1.4, отрицательные побеги потенциалов действия не становятся более отрицательными, чем потенциал покоя, потому что они «едут» на постоянном деполяризующем стимуле.
1.5 Ионные механизмы потенциалов покоя
Прежде чем исследовать ионные механизмы потенциалов действия, сначала необходимо понять ионные механизмы потенциала покоя. Эти два явления тесно связаны. История потенциала покоя восходит к началу 1900-х годов, когда Юлиус Бернштейн предположил, что потенциал покоя (V m ) равен потенциалу равновесия калия (E K ).Где
Ключом к пониманию потенциала покоя является тот факт, что ионы неравномерно распределены внутри и снаружи клеток и что клеточные мембраны избирательно проницаемы для различных ионов. K + особенно важен для потенциала покоя. Мембрана обладает высокой проницаемостью по K + . Кроме того, внутри ячейки имеется высокая концентрация K + ([K + ] i ), а снаружи ячейки — низкая концентрация K + ([K + ] или ).Таким образом, K + естественным образом будет перемещаться путем диффузии из области высокой концентрации в область низкой концентрации. Следовательно, положительные ионы K + , покидая внутреннюю поверхность мембраны, оставляют после себя некоторые отрицательно заряженные ионы. Этот отрицательный заряд притягивает положительный заряд покидающего иона K + и имеет тенденцию «тянуть его назад». Таким образом, будет электрическая сила, направленная внутрь, которая будет стремиться уравновесить диффузионную силу, направленную наружу.В конце концов, равновесие будет установлено; сила концентрации, перемещающая K + наружу, уравновешивает удерживающую его электрическую силу. Потенциал, при котором достигается этот баланс, называется равновесным потенциалом Нернста .
Слева показан эксперимент по проверке гипотезы Бернштейна о том, что мембранный потенциал равен равновесному потенциалу Нернста (т.е. V m = E K ).
Концентрация K + вне клетки систематически варьировалась при измерении мембранного потенциала.Также показана линия, предсказанная уравнением Нернста. Точки, измеренные экспериментально, очень близки к этой линии. Более того, из-за логарифмического соотношения в уравнении Нернста изменение концентрации K + в 10 раз приводит к изменению потенциала на 60 мВ.
Обратите внимание, однако, что есть некоторые отклонения на рисунке слева от того, что предсказывается уравнением Нернста. Таким образом, нельзя сделать вывод, что V m = E K .Такие отклонения указывают на то, что другой ион также участвует в создании потенциала покоя. Этот ион — Na + . Высокая концентрация Na + вне клетки и относительно низкая концентрация внутри клетки приводит к химической (диффузионной) движущей силе для притока Na + . Существует также электрическая движущая сила, потому что внутренняя часть клетки отрицательна, и эта отрицательность притягивает положительные ионы натрия. Следовательно, если клетка имеет небольшую проницаемость для натрия, Na + будет перемещаться через мембрану, и мембранный потенциал будет более деполяризованным, чем можно было бы ожидать из равновесного потенциала K + .
1.6 Уравнение Гольдмана-Ходжкина и Каца (GHK)
Когда мембрана проницаема для двух разных ионов, уравнение Нернста больше не может использоваться для точного определения мембранного потенциала. Однако можно применить уравнение GHK. Это уравнение описывает потенциал на мембране, проницаемой как для Na + , так и для K + .
Обратите внимание, что α — это отношение проницаемости Na + (P Na ) к проницаемости K + (P K ).Также обратите внимание, что если проницаемость мембраны для Na + равна 0, то альфа в GHK равна 0, и уравнение Гольдмана-Ходжкина-Каца сводится к равновесному потенциалу Нернста для K + . Если проницаемость мембраны для Na + очень высока, а проницаемость для калия очень низкая, члены [Na + ] становятся очень большими, доминируя в уравнении по сравнению с членами [K + ] и Уравнение GHK сводится к равновесному потенциалу Нернста для Na + .
Если уравнение GHK применяется к тем же данным на Рисунке 1.5, есть гораздо лучшее соответствие. Значение альфа, необходимое для получения такого точного соответствия, составляло 0,01. Это означает, что проницаемость для калия K + в 100 раз превышает проницаемость для Na + . Таким образом, потенциал покоя обусловлен не только тем фактом, что существует высокая проницаемость для K + . Существует также небольшая проницаемость для Na + , которая имеет тенденцию делать мембранный потенциал немного более положительным, чем он был бы, если бы мембрана была проницаемой только для K + .
1,7 Лаборатория мембранного потенциала
Щелкните здесь, чтобы перейти в интерактивную лабораторию мембранного потенциала, чтобы поэкспериментировать с эффектами изменения внешней или внутренней концентрации ионов калия и проницаемости мембраны для ионов натрия и калия. Прогнозы делаются с использованием уравнений Нернста и Гольдмана, Ходжкина, Каца.
| Лаборатория мембранного потенциала |
Проверьте свои знания
|
Если нервная мембрана внезапно станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если нервная мембрана внезапно станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если нервная мембрана внезапно станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если нервная мембрана внезапно станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если нервная мембрана внезапно станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если нервная мембрана внезапно станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если концентрация K + в цитоплазме аксона беспозвоночного изменяется на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрация K + в цитоплазме аксона беспозвоночного изменяется на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрация K + в цитоплазме аксона беспозвоночного изменяется на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрация K + в цитоплазме аксона беспозвоночного изменяется на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрация K + в цитоплазме аксона беспозвоночного изменяется на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрация K + в цитоплазме аксона беспозвоночного изменяется на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
|
12.5 Потенциал действия — анатомия и физиология
Функции нервной системы — ощущение, интеграция и реакция — зависят от функций нейронов, лежащих в основе этих путей. Чтобы понять, как нейроны могут общаться, необходимо описать роль возбудимой мембраны в генерации этих сигналов.В основе этого процесса лежит потенциал действия . Потенциал действия — это предсказуемое изменение мембранного потенциала, которое происходит из-за открытия и закрытия потенциалзависимых ионных каналов на клеточной мембране.
Большинство клеток тела используют заряженные частицы ( иона ) для создания электрохимического заряда через клеточную мембрану. В предыдущей главе мы описали, как сокращаются мышечные клетки на основе движения ионов через клеточную мембрану. Для сокращения скелетных мышц из-за связи между возбуждением и сокращением им требуется входной сигнал от нейрона.И мышечные, и нервные клетки используют клеточную мембрану, которая специализируется на передаче сигналов, чтобы регулировать движение ионов между внеклеточной жидкостью и цитозолем.
Как вы узнали из главы о клетках, клеточная мембрана в первую очередь отвечает за регулирование того, что может пересекать мембрану. Клеточная мембрана представляет собой бислой фосфолипидов, поэтому только вещества, которые могут проходить непосредственно через гидрофобное ядро, могут диффундировать без посторонней помощи. Заряженные частицы, которые являются гидрофильными, не могут проходить через клеточную мембрану без посторонней помощи (Рисунок 12.5.1). Определенные белки трансмембранного канала позволяют заряженным ионам перемещаться через мембрану. Несколько пассивных транспортных каналов, а также активные транспортные насосы необходимы для создания трансмембранного потенциала и потенциала действия. Особый интерес представляет белок-носитель, называемый натриево-калиевым насосом , который использует энергию для перемещения ионов натрия (Na + ) из клетки и ионов калия (K + ) в клетку, таким образом регулируя ионную концентрация на обеих сторонах клеточной мембраны.
Рисунок 12.5.1 — Клеточная мембрана и трансмембранные белки: Клеточная мембрана состоит из фосфолипидного бислоя и имеет множество трансмембранных белков, включая различные типы канальных белков, которые служат ионными каналами.Натриево-калиевый насос требует энергии в виде аденозинтрифосфата (АТФ), поэтому его также называют насосом АТФазы. Как объяснялось в главе о ячейках, концентрация Na + выше вне ячейки, чем внутри, а концентрация K + выше внутри ячейки, чем снаружи.Таким образом, этот насос работает против градиентов концентрации ионов натрия и калия, поэтому он требует энергии. Насос для АТФазы Na + / K + поддерживает эти важные градиенты концентрации ионов.
Ионные каналы — это поры, которые позволяют определенным заряженным частицам пересекать мембрану в ответ на существующий электрохимический градиент. Белки способны проникать через клеточную мембрану, включая ее гидрофобное ядро, и могут взаимодействовать с заряженными ионами из-за различных свойств аминокислот, обнаруженных в определенных областях белкового канала.Гидрофобные аминокислоты находятся в областях, которые примыкают к углеводородным хвостам фосфолипидов, где гидрофильные аминокислоты подвергаются воздействию жидкой среды внеклеточной жидкости и цитозоля. Кроме того, ионы будут взаимодействовать с гидрофильными аминокислотами, которые будут селективными в отношении заряда иона. Каналы для катионов (положительных ионов) будут иметь отрицательно заряженные боковые цепи в порах. Каналы для анионов (отрицательных ионов) будут иметь в порах положительно заряженные боковые цепи.Диаметр поры канала также влияет на конкретные ионы, которые могут проходить через него. Некоторые ионные каналы избирательны по заряду, но не обязательно по размеру. Эти неспецифические каналы позволяют катионам, особенно Na + , K + и Ca 2+ , проходить через мембрану, но исключают анионы.
Некоторые ионные каналы не позволяют ионам свободно диффундировать через мембрану, но вместо этого закрыты . Управляемый лигандом канал открывается, потому что молекула или лиганд связывается с внеклеточной областью канала (Фиг.12.5.2).
Рис. 12.5.2. Каналы, управляемые лигандом: Когда лиганд, в данном случае нейромедиатор ацетилхолин, связывается с определенным местом на внеклеточной поверхности белка канала, поры открываются, позволяя отобрать ионы. Ионы в данном случае представляют собой катионы натрия, кальция и калия.Канал с механическим затвором, открывается из-за физического искажения клеточной мембраны. Многие каналы, связанные с осязанием, закрываются механически.Например, когда на кожу оказывается давление, открываются механически закрытые каналы подкожных рецепторов, позволяя ионам проникать (рис. 12.5.3).
Рисунок 12.5.3 — Каналы с механическим закрытием: Когда в окружающей ткани происходит механическое изменение (например, давление или растяжение), канал физически открывается, и ионы могут перемещаться по каналу вниз по градиенту их концентрации.Управляемый по напряжению канал — это канал, который реагирует на изменения электрических свойств мембраны, в которую он встроен.Обычно внутренняя часть мембраны находится под отрицательным напряжением. Когда это напряжение становится менее отрицательным и достигает значения, характерного для канала, он открывается и позволяет ионам пересекать мембрану (рисунок 12.5.4).
Рисунок 12.5.4 — Каналы с ограничением по напряжению: Каналы с ограничением по напряжению открываются, когда вокруг них изменяется трансмембранное напряжение. Аминокислоты в структуре белка чувствительны к заряду и заставляют поры открываться для выбранного иона.Канал утечки закрывается случайным образом, что означает, что он открывается и закрывается случайным образом, отсюда и ссылка на утечку.Нет фактического события, открывающего канал; вместо этого он имеет внутреннюю скорость переключения между открытым и закрытым состояниями. Каналы утечки вносят вклад в трансмембранное напряжение покоя возбудимой мембраны (рис. 12.5.5).
Рисунок 12.5.5 — Каналы утечки: Эти каналы открываются и закрываются случайным образом, позволяя ионам проходить через них, когда они открыты.Мембранный потенциал — это распределение заряда через клеточную мембрану, измеряемое в милливольтах (мВ).Стандарт заключается в сравнении внутренней части клетки и внешней среды, поэтому мембранный потенциал представляет собой величину, представляющую заряд на внутриклеточной стороне мембраны (относительно нуля на внешней стороне; рис. 12.5.6).
Рисунок 12.5.6 — Измерение заряда через мембрану с помощью вольтметра: Регистрирующий электрод вставлен в ячейку, а электрод сравнения находится вне ячейки. Путем сравнения заряда, измеренного этими двумя электродами, определяется трансмембранное напряжение.Это значение обычно выражается для цитозоля относительно внешней среды.Обычно существует общий чистый нейтральный заряд между внеклеточной и внутриклеточной средой нейрона. Однако небольшая разница в заряде возникает прямо на поверхности мембраны как внутри, так и снаружи. Это различие в этой очень ограниченной области, которая обладает способностью генерировать электрические сигналы, включая потенциалы действия, в нейронах и мышечных клетках.
Когда клетка находится в состоянии покоя, ионы распределяются по мембране очень предсказуемым образом.Концентрация Na + вне клетки в 10 раз больше, чем концентрация внутри. Кроме того, концентрация K + внутри ячейки больше, чем снаружи. Цитозоль содержит высокую концентрацию анионов в форме фосфат-ионов и отрицательно заряженных белков. С ионами, распределенными по мембране при этих концентрациях, разница в заряде описывается как потенциал покоя мембраны . Точное значение, измеренное для мембранного потенциала покоя, варьируется между клетками, но обычно сообщается о -70 мВ.Это напряжение на самом деле было бы намного ниже, если бы не вклад некоторых важных белков в мембрану. Каналы утечки позволяют Na + медленно перемещаться в ячейку или K + медленно перемещаться наружу, а насос Na + / K + восстанавливает их градиенты концентрации через мембрану. Это может показаться пустой тратой энергии, но каждый из них играет определенную роль в поддержании мембранного потенциала.
Потенциал действия
Мембранный потенциал покоя описывает устойчивое состояние клетки, которое представляет собой динамический процесс, уравновешивающий ионы, утекающие вниз по градиенту их концентрации, и ионы, перекачиваемые обратно вверх по градиенту их концентрации.Без какого-либо внешнего воздействия мембранный потенциал покоя будет поддерживаться. Чтобы запустить электрический сигнал, мембранный потенциал должен стать более положительным.
Это начинается с открытия потенциал-управляемых каналов Na + в мембране нейрона. Поскольку концентрация Na + выше вне ячейки, чем внутри ячейки в 10 раз, ионы будут устремляться в ячейку под действием как химических, так и электрических градиентов. Поскольку натрий — это положительно заряженный ион, при попадании в клетку он сразу же изменяет относительное напряжение внутри клеточной мембраны.Мембранный потенциал покоя составляет приблизительно -70 мВ, поэтому катион натрия, попадающий в клетку, заставляет мембрану становиться менее отрицательной. Это известно как деполяризация , что означает, что мембранный потенциал приближается к нулю (становится менее поляризованным). Градиент концентрации Na + настолько велик, что он будет продолжать поступать в клетку даже после того, как мембранный потенциал станет нулевым, так что напряжение непосредственно вокруг поры тогда начинает становиться положительным.
Когда мембранный потенциал достигает +30 мВ, в мембране открываются более медленные, управляемые по напряжению калиевые каналы.Электрохимический градиент также действует на K + . Когда K + начинает покидать ячейку, унося с собой положительный заряд, мембранный потенциал начинает возвращаться к своему напряжению покоя. Это называется реполяризацией , что означает, что мембранное напряжение возвращается к значению -70 мВ потенциала покоя мембраны.
Реполяризация возвращает мембранный потенциал к значению потенциала покоя -70 мВ, но превышает это значение. Ионы калия достигают равновесия, когда напряжение на мембране ниже -70 мВ, поэтому возникает период гиперполяризации, когда каналы K + открыты.Эти каналы K + закрываются с небольшой задержкой из-за этого короткого выброса.
Здесь был описан потенциал действия, который представлен в виде графика зависимости напряжения от времени на рисунке 12.5.7. Это электрический сигнал, который нервная ткань генерирует для общения. Изменение мембранного напряжения от -70 мВ в состоянии покоя до +30 мВ в конце деполяризации представляет собой изменение на 100 мВ.
Рисунок 12.5.7 — График потенциала действия: График зависимости напряжения, измеренного на клеточной мембране от времени, потенциал действия начинается с деполяризации, за которой следует реполяризация, которая переходит за потенциал покоя в гиперполяризацию, и, наконец, мембрана возвращается в состояние покоя.Внешний веб-сайт
То, что происходит через мембрану электрически активной ячейки, представляет собой динамический процесс, который трудно визуализировать с помощью статических изображений или текстовых описаний. Просмотрите эту анимацию, чтобы узнать больше об этом процессе. В чем разница между движущей силой Na + и K + ? А что общего в движении этих двух ионов?
Мембранный потенциал будет оставаться на уровне напряжения покоя, пока что-то не изменится.Чтобы запустить потенциал действия, мембранный потенциал должен измениться от потенциала покоя приблизительно -70 мВ до порогового напряжения -55 мВ. Как только клетка достигает порога, потенциал-управляемые натриевые каналы открываются, и предсказуемые изменения мембранного потенциала описываются выше как потенциал действия. Любая подпороговая деполяризация, которая не изменяет мембранный потенциал до -55 мВ или выше, не достигнет порога и, следовательно, не приведет к потенциалу действия. Кроме того, любой стимул, который деполяризует мембрану до -55 мВ или выше, вызовет открытие большого количества каналов и инициирование потенциала действия.
Из-за предсказуемых изменений, которые происходят при достижении порогового значения, потенциал действия обозначается как «все или ничего». Это означает, что либо возникает потенциал действия и повторяется по всей длине нейрона, либо потенциал действия не возникает. Более сильный стимул, который может деполяризовать мембрану далеко за порог, не приведет к «большему» потенциалу действия. Либо мембрана достигает порога, и все происходит, как описано выше, либо мембрана не достигает порога, и больше ничего не происходит.Все потенциалы действия достигают пика при одном и том же напряжении (+30 мВ), поэтому один потенциал действия не больше другого. Более сильные стимулы быстрее инициируют множественные потенциалы действия, но отдельные сигналы не сильнее.
Как мы видели, деполяризация и реполяризация потенциала действия зависят от двух типов каналов (потенциал-зависимый канал Na + и потенциал-управляемый канал K + ). Управляемый по напряжению канал Na + фактически имеет два затвора.Один из них — это ворота активации , которые открываются, когда мембранный потенциал превышает -55 мВ. Другой вентиль — вентиль инактивации , который закрывается через определенный период времени — порядка долей миллисекунды. Когда ячейка находится в состоянии покоя, ворота активации закрыты, а ворота дезактивации открыты. Однако при достижении порога активирующие ворота открываются, позволяя Na + ворваться в ячейку. В момент пика деполяризации ворота инактивации закрываются.Во время реполяризации в клетку больше не может попасть натрий. Когда мембранный потенциал снова превышает -55 мВ, активирующий вентиль закрывается. После этого ворота инактивации снова открываются, делая канал готовым к повторному запуску всего процесса.
Управляемый по напряжению канал K + имеет только один затвор, чувствительный к мембранному напряжению -50 мВ. Однако он не открывается так быстро, как закрытый по напряжению канал Na + . Для открытия канала K + после достижения этого напряжения требуется доли миллисекунды, что точно совпадает с моментом пика потока Na + .Таким образом, закрытые по напряжению каналы K + открываются так же, как закрытые по напряжению каналы Na + деактивируются. Когда мембранный потенциал переполяризуется и напряжение снова достигает -50 мВ, каналы K + начинают закрываться. Калий продолжает покидать клетку в течение короткого времени, и мембранный потенциал становится более отрицательным, что приводит к превышению гиперполяризации. Затем каналы K + закрываются, и мембрана возвращается в состояние покоя из-за продолжающейся активности каналов утечки и насоса АТФазы Na + / K + .
Все это происходит примерно за 2 миллисекунды (рисунок 12.5.8). Пока существует потенциал действия, другой не может быть инициирован. Этот эффект называется рефрактерным периодом . Существует две фазы огнеупорного периода: абсолютный огнеупорный период и относительный огнеупорный период . В течение периода абсолютной рефрактерности другой потенциал действия не запускается. Это происходит из-за затвора инактивации управляемого по напряжению канала Na + .Как только канал Na + возвращается к своей конформации покоя, новый потенциал действия может быть запущен во время фазы гиперполяризации, но только более сильным стимулом, чем тот, который инициировал текущий потенциал действия.
Рисунок 12.5.8 — Этапы потенциала действия: График зависимости напряжения, измеренного на клеточной мембране, от времени, события потенциала действия могут быть связаны с конкретными изменениями мембранного напряжения. (1) В состоянии покоя напряжение на мембране составляет -70 мВ. (2) Мембрана начинает деполяризоваться при приложении внешнего раздражителя.(3) Напряжение на мембране начинает быстро расти до +30 мВ. (4) Напряжение мембраны начинает возвращаться к отрицательному значению. (5) Реполяризация продолжается после напряжения покоя мембраны, что приводит к гиперполяризации. (6) Напряжение на мембране возвращается к исходному значению вскоре после гиперполяризации.Распространение потенциала действия
Потенциал действия инициируется в начале аксона, в так называемом начальном сегменте ( триггерная зона) . Здесь может происходить быстрая деполяризация из-за высокой плотности потенциалзависимых каналов Na + .Спускаясь вниз по длине аксона, потенциал действия распространяется, потому что по мере распространения деполяризации открывается больше потенциал-управляемых каналов Na + . Это распространение происходит из-за того, что Na + проникает через канал и движется по внутренней части клеточной мембраны. Когда Na + перемещается или проходит небольшое расстояние вдоль клеточной мембраны, его положительный заряд деполяризует немного больше клеточной мембраны. По мере того, как эта деполяризация распространяется, открываются новые потенциалозависимые каналы Na + , и все больше ионов устремляется в клетку, распространяя деполяризацию немного дальше.
Поскольку управляемые по напряжению каналы Na + инактивируются на пике деполяризации, они не могут быть снова открыты на короткое время (период абсолютной рефрактерности). Из-за этого положительные ионы, распространяющиеся обратно к ранее открытым каналам, не имеют никакого эффекта. Потенциал действия должен распространяться от триггерной зоны к окончанию аксона.
Распространение, как описано выше, применимо к немиелинизированным аксонам. Когда присутствует миелинизация, потенциал действия распространяется по-другому и оптимизирован для скорости прохождения сигнала.Ионы натрия, которые входят в клетку в триггерной зоне, начинают распространяться по длине сегмента аксона, но до первого узла Ранвье нет управляемых по напряжению каналов Na + . Поскольку не существует постоянного открытия этих каналов вдоль сегмента аксона, деполяризация распространяется с оптимальной скоростью. Расстояние между узлами — это оптимальное расстояние, чтобы мембрана оставалась деполяризованной выше порога в следующем узле. Когда Na + распространяется по внутренней части мембраны сегмента аксона, заряд начинает рассеиваться.Если бы узел находился дальше по аксону, эта деполяризация упала бы слишком сильно для того, чтобы управляемые по напряжению каналы Na + были активированы в следующем узле Ранвье. Если бы узлы были ближе друг к другу, скорость распространения была бы ниже.
Распространение по немиелинизированному аксону обозначается как непрерывная проводимость ; По длине миелинизированного аксона он обозначается как скачкообразной проводимости . Непрерывная проводимость является медленной, потому что всегда открываются управляемые по напряжению каналы Na + , и все больше и больше Na + устремляется в ячейку.Солевое проведение происходит быстрее, потому что потенциал действия «прыгает» от одного узла к другому (saltare = «прыгать»), а новый приток Na + обновляет деполяризованную мембрану. Наряду с миелинизацией аксона диаметр аксона может влиять на скорость проводимости. Подобно тому, как вода течет быстрее в широкой реке, чем в узком ручье, деполяризация на основе Na + распространяется быстрее по широкому аксону, чем по узкому. Эта концепция известна как сопротивление и обычно применима для электрических проводов или водопровода, так же как и для аксонов, хотя конкретные условия отличаются в масштабах электронов или ионов по сравнению с водой в реке.
Гомеостатический дисбаланс — Концентрация калия
Глиальные клетки, особенно астроциты, отвечают за поддержание химической среды ткани ЦНС. Концентрация ионов во внеклеточной жидкости является основой того, как устанавливается мембранный потенциал и изменяется электрохимическая сигнализация. Если баланс ионов нарушен, возможны тяжелые исходы.
Обычно концентрация K + выше внутри нейрона, чем снаружи.После фазы реполяризации потенциала действия каналы утечки K + и насосы Na + / K + гарантируют, что ионы возвращаются в свои исходные положения. После инсульта или другого ишемического события уровни внеклеточного K + повышаются. Астроциты в этой области оборудованы для удаления излишков K + , чтобы помочь помпе. Но когда уровень далеко не сбалансирован, последствия могут быть необратимыми.
Астроциты могут стать реактивными в подобных случаях, что снижает их способность поддерживать локальную химическую среду.Глиальные клетки увеличиваются, и их отростки набухают. Они теряют свою буферную способность K + , и это влияет на работу насоса или даже обращается вспять. Одним из первых признаков заболевания клеток является «утечка» ионов натрия в клетки организма. Этот дисбаланс натрия / калия отрицательно влияет на внутреннюю химию клеток, препятствуя их нормальному функционированию.
Внешний веб-сайт
Посетите этот сайт, чтобы увидеть виртуальную лабораторию нейрофизиологии и понаблюдать за электрофизиологическими процессами в нервной системе, где ученые непосредственно измеряют электрические сигналы, производимые нейронами.Часто потенциалы действия возникают так быстро, что смотреть на экран, чтобы увидеть, как они возникают, бесполезно. Динамик приводится в действие сигналами, записанными от нейрона, и он «выскакивает» каждый раз, когда нейрон запускает потенциал действия. Эти потенциалы действия срабатывают так быстро, что по радио это звучит как статика. Электрофизиологи могут распознать закономерности в этой статике, чтобы понять, что происходит. Почему модель пиявки используется для измерения электрической активности нейронов, а не людей?
Неврология для детей — потенциал действия
Свет, камера, потенциал действия
Эта страница описывает как работают нейроны.Надеюсь, это объяснение не станет слишком сложным, но важно понимать, как нейроны делают то, что они делают. Есть много деталей, но не торопитесь и смотрите на цифры.
Многое из того, что мы узнать о том, как работают нейроны, удалось из экспериментов на гигантском аксоне кальмар. Этот гигантский аксон простирается от головы до хвоста кальмара. и используется для движения хвоста кальмара. Насколько велик этот аксон? Это может быть диаметром до 1 мм — легко увидеть невооруженным глазом.
Нейроны отправляют сообщения электрохимически .Это означает что химические вещества вызывают электрический сигнал. Химические вещества в организме «электрически заряженные» — когда у них есть электрический заряд, они называется иона. Важными ионами нервной системы являются: натрий и калий (оба имеют 1 положительный заряд, +), кальций (имеют 2 положительные заряды, ++) и хлорид (имеет отрицательный заряд, -). Есть также некоторые отрицательно заряженные белковые молекулы. Также важно помните, что нервные клетки окружены мембраной, которая позволяет некоторым ионы проходят и блокируют прохождение других ионов.Этот тип Мембрана называется полупроницаемой .
Мембранный потенциал покоя
Когда нейрон не посылая сигнал, он находится в состоянии покоя. Когда нейрон находится в состоянии покоя, его внутренняя часть нейрона отрицательна по отношению к внешней стороне. Хотя концентрации различных ионов пытаются уравновесить с обеих сторон мембраны, они не могут, потому что клеточная мембрана позволяет только некоторые ионы проходят через каналы (ионные каналы). В состоянии покоя ионы калия (K + ) может легко проходить через мембрану.Также в покое, ионы хлорида (Cl — ) и ионы натрия (Na + ) имеют более трудный переход времени. Отрицательно заряженные белковые молекулы (A — ) внутри нейрона не может пересечь мембрану. В дополнение к этим селективным ионным каналов, есть насос , который использует энергию для перемещения трех ионы натрия из нейрона на каждые два иона калия, которые он вводит. Наконец, когда все эти силы уравновешиваются, и разница в напряжение между внутренней и внешней частью нейрона измеряется, у вас есть потенциал покоя .Покоящаяся мембрана потенциал нейрона составляет около -70 мВ (мВ = милливольт) — это означает, что внутри нейрона на 70 мВ меньше, чем снаружи. В покое есть относительно больше ионов натрия вне нейрона и больше ионов калия внутри этого нейрона.
Возможности действия
потенциал покоя говорит о том, что происходит, когда нейрон находится в состоянии покоя. Потенциал действия возникает, когда нейрон посылает информацию по аксону от тела клетки.Нейробиологи используют другие слова, такие как «спайк» или «импульс» для обозначения потенциал действия. Потенциал действия — это взрыв электрическая активность, которая создается деполяризующей текущий . Это означает, что какое-то событие (стимул) вызывает потенциал покоя приближается к 0 мВ. Когда деполяризация достигает около -55 мВ нейрон вызовет потенциал действия. Это порог . Если нейрон не достигает этого критического пороговый уровень, то потенциал действия не сработает.Кроме того, когда пороговый уровень достигнут, потенциал действия фиксированного размера будет всегда срабатывает … для любого данного нейрона величина потенциала действия равна всегда одно и то же. В одном нерве нет большого или малого потенциалов действия. ячейка — все потенциалы действия одинакового размера. Следовательно, нейрон либо не достигает порога, либо полный потенциал действия уволен — это принцип «ВСЕ ИЛИ НЕТ».
Потенциалы действия вызваны когда разные ионы пересекают мембрану нейрона.Стимул сначала вызывает натриевые каналы открыть. Потому что на поверхности гораздо больше ионов натрия. снаружи, а внутренняя часть нейрона отрицательна относительно внешней, ионы натрия устремляются в нейрон. Помните, у натрия есть положительный заряд, так нейрон становится более позитивным и становится деполяризованным. Занимает дольше для открытия калиевых каналов. Когда они открываются, калий вылетает из клетки, обращая деполяризацию вспять. Также об этом время, натриевые каналы начинают закрываться.Это приводит к тому, что потенциал действия вернитесь к -70 мВ (реполяризация). Фактически потенциал действия превышает -70 мВ (гиперполяризация), потому что калиевые каналы остаются открывать слишком долго. Постепенно концентрация ионов возвращается в состояние покоя. уровни, и ячейка возвращается к -70 мВ.
И вот он … потенциал действия
| Знаете ли вы? |
Гигантский аксон кальмара может быть от 100 до 1000 раз больше, чем аксон млекопитающих.Гигантский аксон иннервирует мантийная мышца кальмара. Эти мышцы используются для продвижения кальмаров через вода. |
Авторские права © 1996-2020, Эрик Х. Чудлер. Все права. Зарезервированный.
.