Какие микросхемы используются для организации связи ПК и микроконтроллера по RS-232. Как работают гальванические развязки в таких схемах. Какие современные решения предлагают производители для создания компактных и надежных интерфейсов.
Современные микросхемы для реализации интерфейса RS-232
Для организации связи между компьютером и микроконтроллером по интерфейсу RS-232 используются специализированные микросхемы-преобразователи уровней. Они выполняют следующие основные функции:
- Преобразование уровней сигналов RS-232 в TTL-уровни и обратно
- Защита входов/выходов от статического электричества
- Формирование необходимых напряжений для работы интерфейса
Наиболее распространенными микросхемами для этих целей являются:
- MAX232 — классическая микросхема от Maxim Integrated
- ADM3202 — современный аналог от Analog Devices
- SP3232 — микросхема от Sipex
- ICL3232 — вариант от Intersil
Эти микросхемы позволяют реализовать базовый интерфейс RS-232 с минимумом внешних компонентов. Однако для промышленного применения часто требуется гальваническая развязка между устройствами.
Гальванические развязки для интерфейса RS-232
Гальваническая развязка позволяет защитить устройства от повреждения из-за разности потенциалов и исключить образование паразитных контуров. Для реализации гальванической развязки RS-232 используются специальные микросхемы, например:
- ADUM1201 — двухканальная цифровая изоляция от Analog Devices
- ISO7221 — изолятор от Texas Instruments
- Si8440 — изолятор от Silicon Labs
Принцип работы таких микросхем основан на передаче данных через изолирующий барьер с помощью трансформаторов или оптопар. Это позволяет обеспечить изоляцию до нескольких киловольт.
Компактные решения для изолированного RS-232
Современные производители предлагают интегрированные решения, объединяющие в одном корпусе преобразователь уровней и гальваническую развязку. Примеры таких микросхем:
- ADM3251E — изолированный RS-232 трансивер от Analog Devices
- ISO1050 — изолированный RS-232 от Texas Instruments
- Si86xx — семейство изолированных интерфейсов от Silicon Labs
Использование таких решений позволяет существенно уменьшить размеры устройства и повысить надежность.
Особенности применения изолированного RS-232
При разработке устройств с изолированным интерфейсом RS-232 необходимо учитывать следующие аспекты:
- Требуется изолированное питание для части схемы со стороны RS-232
- Необходимо обеспечить согласование импедансов линии связи
- Следует учитывать ограничения по скорости передачи данных через изолирующий барьер
- Важно правильно организовать заземление изолированных частей схемы
При соблюдении этих требований можно создать надежный изолированный интерфейс RS-232 для связи компьютера с микроконтроллером.
Примеры схем изолированного RS-232
Рассмотрим несколько вариантов схем для реализации изолированного интерфейса RS-232:
Схема на основе ADUM1201 и ADM3202
Эта схема использует цифровой изолятор ADUM1201 и преобразователь уровней ADM3202:
«` «`В данной схеме ADUM1201 обеспечивает гальваническую развязку сигналов TxD и RxD, а ADM3202 выполняет преобразование уровней. Для питания изолированной части используется DC/DC преобразователь.
Интегрированное решение на ADM3251E
Более компактный вариант можно реализовать с помощью микросхемы ADM3251E:
«` «`ADM3251E объединяет в одном корпусе изолятор и преобразователь уровней, что позволяет минимизировать количество внешних компонентов.
Выбор оптимального решения для изолированного RS-232
При выборе компонентов для реализации изолированного интерфейса RS-232 следует учитывать следующие факторы:
- Требуемая скорость передачи данных
- Необходимый уровень изоляции
- Доступное пространство на печатной плате
- Стоимость решения
- Энергопотребление
Для большинства применений оптимальным выбором будут интегрированные решения вроде ADM3251E или ISO1050. Они обеспечивают хороший баланс между функциональностью, компактностью и стоимостью.
Перспективы развития изолированных интерфейсов
Развитие технологий изоляции и интеграции компонентов ведет к появлению все более совершенных решений для изолированных интерфейсов. Основные тенденции включают:

- Повышение скорости передачи данных через изолирующий барьер
- Увеличение уровня изоляции
- Снижение энергопотребления
- Уменьшение размеров корпусов
- Интеграция дополнительной функциональности (например, защиты от ESD)
Это позволит создавать еще более компактные и надежные устройства с изолированным интерфейсом RS-232 для связи компьютеров с микроконтроллерами.
Современные аппаратные средства связи микроконтроллера с компьютером по интерфейсу RS-232. Часть 3 — Компоненты и технологии
Все статьи цикла:
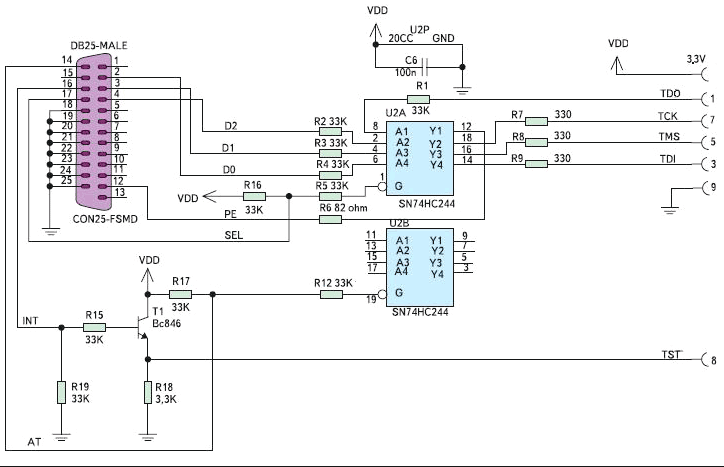
1.4.1. Простой гальванически изолированный интерфейс RS-232 на базе микросхем развязки ADUM1201, преобразователя SN75155 и оптронов
На рис. 18 показана схема сопряжения компьютера с микроконтроллером (например, из семейства MSC12XX) с помощью гальванически изолированного интерфейса RS-232. Сам микроконтроллер показан условно в правой нижней части схемы.
Рис. 18. Схема гальванически изолированного интерфейса RS-232 на базе развязки ADUM1201 и преобразователя интерфейса SN75155 для микроконтроллеров семейств MSC12XX
В схеме в качестве гальванических развязок помимо двунаправленной развязки ADUM1201 (DD2) используются широко известные, доступные и дешевые оптроны h21L1 (DD3) и TLP521 (DD4). В схеме также применена известная высокоскоростная микросхема одноканального приемопередатчика SN75155 (DD1) от Texas Instruments, которая редко используется из-за повышенного энергопотребления. Ток потребления части схемы со стороны компьютера составляет около 12 мА; из них 11 мА потребляет SN75155. Для питания части схемы со стороны компьютера используется DC/DC-конвертор RQD-0512 (DD5) компании Recom мощностью 1 Вт, преобразующий входное напряжение +5 В в два напряжения +12 В и –12 В. Он выдает ток по 42 мА на каждое напряжение питания (с лихвой перекрывающий энергию, требующуюся для питания развязки). Микроконтроллер показан схематично; к нему подходят сигналы данных RS-232 — TxDM и RxDM и сигналы управления — RST и PSEN.
В прямоугольнике, очерченном пунктирной линией, показана схема кабеля связи с компьютером. Линия RxD соединена с линией DSR непосредственно в разъеме DB9M, который подключается к COM-порту компьютера. Таким образом, драйвер SN75155 (2-й и 7-й вывод) работает сразу на два входа (на два приемника RS-232). Для получения напряжения питания +5 В (питание ADUM1201) используется малопотребляющий и достаточно прецизионный стабилизатор LP2950CZ5.0 (DD6). Схема идеально работает на скорости обмена 115 200 бод на расстоянии между компьютером и микроконтроллером до 15 м.
Микросхема ADUM1201 выпускается в корпусе SOIC8, который неудобен для ручного макетирования. Для удобства работы с этой микросхемой при ручном макетировании автор рекомендует изготовить платупереходник SOIC8→DIP8, вариант разводки которой приведен на рис. 19.
Рис. 19. Вариант разводки платы-переходника SOIC8→DIP8 для развязки ADUM1201 (слева — верхний слой монтажа микросхемы, справа — нижний слой платы)
В плате-переходнике в качестве «ножек» используются штырьки, которые легко «отламываются» от линейки из 40 двусторонних штырьков круглого сечения диаметром около 0,5–0,7 мм. Верхняя часть штырька (меньшего диаметра) впаивается в плату-переходник (лишняя его часть откусывается), а нижняя служит ножкой. Штырек, связанный с 1 выводом микросхемы, можно не откусывать; по нему удобно ориентировать плату-переходник при установке ее в панельку.
Необходимо отметить, что расстояние между центрами ножек (штырьков), находящихся на одной линии, но с противоположных сторон платы-переходника (например, между центрами 1-й и 8-й ножек), составляет по стандарту корпуса DIP 0,3 дюйма или 7,62 мм. Учитывая толщину штырька, а также погрешность монтажа, расстояние между штырьками может составить чуть менее 6 мм. Максимальное же расстояние между концами ножек, находящихся на одной линии, но с противоположных сторон микросхемы в корпусе SOIC8 (например, между концами 1-й и 8-й ножек), составляет 6,2 мм. Поэтому микросхема в корпусе SOIC8 «не влезет» на плату-переходник с топологией, показанной на рис. 19 (в левой нижней части рисунка).
Рис. 20. Процедура «подгибания» ножек микросхемы под ее «дно»
Если же «подогнуть» ножки микросхемы под ее «дно», как показано на рис. 20, то она свободно умещается на плату-переходник, и еще даже остается свободное пространство, что намного упрощает пайку. Для того чтобы более прояснить ситуацию, напомним, что, как известно, аналогичное расположение выводов имеют все конденсаторы, резисторы и диоды, предназначенные для поверхностного монтажа, а также микросхемы в корпусах QFN. Следует также отметить, что процедуру «подгибания» можно производить только
Фотография платы-переходника SOIC8→DIP8 для развязки ADUM1201 показана на рис. 21. Как можно убедиться, микросхема ADUM1201 (с подогнутыми ножками) очень удачно «вписывается» в топологию платы. На фотографии можно заметить, что штырек, связанный с 1 выводом микросхемы, не откусан и служит для удобства ориентации (как уже говорилось ранее).
Рис. 21. Фотография платы-переходника SOIC8→DIP8 для развязки ADUM1201
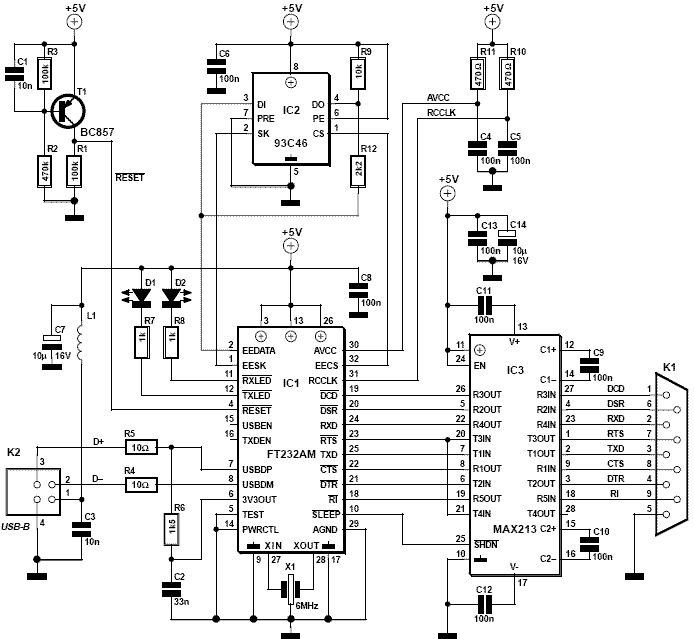
1.4.2. Гальванически изолированный интерфейс RS-232 на базе микросхем развязок ADUM1201, ADUM1200 и преобразователей ADM3202 и MAX3181
На рис. 22 приведена схема гальванически изолированного интерфейса RS-232 на базе двух микросхем развязок ADUM1201(DD2), ADUM1200(DD3) и преобразователей ADM3202(DD1), MAX3181(DD5). Схема, как и предыдущая (рис. 18), формирует четыре сигнала — TxDM, RxDM, RST и PSEN для сопряжения компьютера с микроконтроллерами семейства MSC12XX, поэтому условное изображение микроконтроллера не нужно.
Рис 22. Схема гальванической развязки на базе ADUM1201, ADUM1200 и преобразователя интерфейса ADM3202 для микроконтроллеров семейства MSC12XX
В связи с тем, что преобразователь ADM3202 имеет два передатчика (10–7 и 11–14 выводы DD1), линии RxD и DSR разнесены. Это значительно повышает надежность передатчиков, так как каждый из них работает на один вход. Схема кабеля сопряжения с компьютером показана в прямоугольнике, очерченном пунктиром. Как можно увидеть, для каждой из линий RxD и DSR предусмотрен отдельный провод, хотя эти два сигнала формируются преобразователем ADM3202 из одного и того же сигнала (RxDa), для генерации которого используется единственный канал развязки ADUM1201 (2–7 выводы DD2). Часть схемы, непосредственно контактирующая с интерфейсом компьютера (слева от пунктирной линии, условно показывающей изоляционный барьер), питается от единственного источника питания напряжением +5 В (сигнал «+5A»), формируемым DC/DC-преобразователем RQS-0505 мощностью всего 0,25 Вт. Размер корпуса RQS-0505 (и соответственно стоимость) меньше, чем у RQD-0512, использованном в предыдущей схеме (рис. 18). Кроме того, отпадает необходимость в дополнительном стабилизаторе +5 В (DD6 на рис. 18).
Для более удобного макетирования устройств, использующих обе развязки ADUM1201 и ADUM1200, автор рекомендует изготовить плату-переходник 2×SOIC8→DIP12, схема и вариант разводки которой приведен на рис. 23. Для того чтобы микросхемы «вписались» в топологию платы-переходника, их ножки следует подогнуть так же, как и для переходника SOIC8→DIP8 (см. рис. 20, 21).
Рис. 23. Вариант разводки платы-переходника 2×SOIC8→DIP12 для двух развязок ADUM1201 и ADUM1200
Микросхема приемника RS-232 MAX3181 (DD5 на рис. 22) выпускается в планарном корпусе SOT23-5, который, как и корпус SOIC8 неудобен для макетирования, поэтому автор рекомендует изготовить переходник SOT23-6→DIP6 аналогично переходнику SOIC8→DIP8, показанному на рис. 19 (схема и вариант разводки не приводятся ввиду их простоты). Благодаря малым размерам микросхемы MAX3181 она свободно умещается на плате-переходнике без процедуры «подгибания» ножек.
На рис. 24 приведены фотографии вышеуказанных двух плат переходников: 2×SOIC8→DIP12 и SOT-23-6→DIP6.
Рис. 24. Фотографии плат-переходников 2×SOIC8→DIP12 для двух развязок ADUM1201 и ADUM1200 (а) и SOT-23-6→DIP6 (б) для приемника MAX3181
Кроме того, следует отметить, что микросхема приемника MAX3181 труднодоступна (в малых количествах), поэтому вместо нее можно применить КМОП-транзистор 2N7000. Схема замены микросхемы MAX3181 на транзистор 2N7000 приведена в прямоугольнике, очерченном пунктиром, в левой нижней части рис. 22.
Несколько слов о достоинствах схемы рис. 22. Схема достаточно проста, в ней применены относительно доступные (и дешевые) компоненты, и она не содержит ни одного резистора (если использовать приемник MAX3181). Кроме того, схема идеально работает на скорости 115 200 бод, так как максимальные скорости работы ADM3202 и MAX3181 значительно выше и составляют 460 кбод и 1,5 Мбод соответственно. Длина кабеля связи с компьютером может достигать 20 м. Автор рекомендует эту схему гальванических развязок для большинства применений (не только для микроконтроллеров семейств MSC12XX).
1.4.3. Гальванически изолированный интерфейс RS-232 на базе микросхем развязок ADUM1201, ADUM1200 и преобразователя MAX1406
Рис. 25. Схема гальванической развязки на базе ADUM1201, ADUM1200 и преобразователя интерфейса MAX1406 для микроконтроллеров семейства MSC12XX
На рис. 25 приведена еще одна схема гальванически изолированного интерфейса RS-232 на базе развязок ADUM1201 и ADUM1200. Схема несколько проще, чем предыдущая (рис. 22), так как в ней применена единственная микросхема преобразователя интерфейса MAX1406 (DD1), включающая три приемника и три передатчика. В связи с этим отпадает необходимость в дополнительном приемнике (MAX3181, как в схеме на рис. 22). Правда, возникает необходимость в дополнительном стабилизаторе +5 В (DD5 — LP2950Z5.0) и, кроме того, требуется DC/DC-преобразователь с двумя (+12 В и –12 В) выходными напряжениями питания (DD4 — RQD-0512). Схема достаточно скрупулезно тестировалась автором и отлично работала на скорости 115 200 бод (как было упомянуто ранее, максимальная скорость работы MAX1406 составляет 230 кбод). Размах сигнала драйвера повышен и составляет около ±11,5 В на нагрузке в 5 кОм, что является несомненным преимуществом микросхемы MAX1406 перед упоминавшейся ранее ADM3202 (размах сигнала составляет ±10 В при питании +5 В). Длина кабеля связи (на такой скорости работы схемы) может достигать 20 м. Еще одно достоинство схемы в том, что она более удобна для макетирования, чем предыдущая (рис. 22), если использовать микросхему MAX1406 в корпусе DIP16 (MAX1406CPE). Правда, стоимость такой микросхемы (при малом количестве) довольно высока — почти $5.
И последнее, что необходимо отметить по поводу MAX1406. Эта микросхема была выпущена компанией MAXIM для прямой замены более старых микросхем MC145406 производства Motorola и SN75C1406 от Texas Instruments в устройствах, где требуется высокая скорость обмена в 115 200 бод, поскольку ни MC145406, ни SN75C1406 такую скорость обмена не поддерживают. Автор настоятельно рекомендует ни в коем случае не применять две последние микросхемы в качестве преобразователей интерфейса RS-232 (в схеме рис. 25). Эксперименты, проведенные автором с этими микросхемами, показали, что скорость обмена свыше 38 400 бод от них получить невозможно; кроме того, даже на такой скорости обмена они работают нестабильно, что приводит к частым «зависаниям» компьютера.
1.4.4. Гальванически изолированный интерфейс RS-232 на базе микросхем развязок ADUM1201, ADUM1200 и преобразователей MAX3190 и MAX3181
Малый размер микросхем гальванических развязок ADUM1201 и ADUM1200 и крошечный размер преобразователей MAX3190, MAX3181 и MAX3183 позволяют конструировать очень компактные устройства гальванически развязанного интерфейса RS-232, которые могут свободно умещаться на плате размером с обычную микросхему в DIP-корпусе. Такая плата, оснащенная «ножками», подобными ножкам микросхем в DIP-корпусах, может представлять собой что-то типа гибридной микросхемы в DIP-корпусе, которую, с одной стороны, можно легко сконфигурировать под ту или иную задачу, и с другой — очень удобно использовать для макетирования. Впоследствии такая «микросхема» после тестирования может быть установлена уже в готовое изделие. Ниже приводится два варианта подобных «гибридных микросхем» гальванически изолированного интерфейса, предназначенного для программирования и штатного режима работы микроконтроллеров семейства MSC12XX (1-й вариант) и семейств P89LPC9XX (2-й вариант).
1.4.4.1. Плата гальванически изолированного интерфейса на базе развязок ADUM1201, ADUM1200, MAX3181 иMAX3190 для микроконтроллеров семейства MSC12XX
Рис. 26. Схема гальванически изолированного преобразователя интерфейса ADUMAX на базе ADUM1201, ADUM1200, MAX3181 и MAX3190
На рис. 26 приведена схема гальванически изолированного преобразователя интерфейса на базе ADUM1201, ADUM1200, MAX3181 и MAX3190. Как видно из рисунка, схема состоит из трех инвертирующих приемников RS-232 — MAX3181 (DD4, DD5 и DD6), одного инвертирующего передатчика RS-232—MAX3190 (DD3), одной двухканальной двунаправленной развязки ADUM1201 (DD1) и одной двухканальной однонаправленной развязки ADUM1200 (DD2). Назовем условно такое устройство ADUMAX.
Рис 27. Вариант разводки платы гальванически изолированного преобразователя интерфейса ADUMAX на базе ADUM1201, ADUM1200, MAX3181 и MAX3190
На рис. 27 приведен вариант разводки платы с размещением компонентов с двух сторон. На одной стороне платы (верхняя часть) находится двунаправленная развязка ADUM1201 (DD1) совместно с инвертирующим передатчиком MAX3190 (DD3) и инвертирующим приемником MAX3181 (DD4), на другой (нижняя часть) — двухканальная однонаправленная развязка ADUM1200 (DD2) и два инвертирующих приемника MAX3181 (DD5 и DD6).
На рис. 28 показаны фотографии платы «микросхемы» ADUMAX.
Рис. 28. Фотографии платы гальванически изолированного преобразователя интерфейса ADUMAX на базе ADUM1201, ADUM1200, MAX3181 и MAX3190 а) верхняя часть платы, б) — нижняя
Здесь можно увидеть, что такая топология разводки компонентов позволила расположить 6 микросхем, 5 конденсаторов и 14 штырьков, которые являются «ножками» «микросхемы», на довольно ограниченном пространстве, соответствующем площади стандартной микросхемы в корпусе DIP16. На фотографии (рис. 28а) можно заметить явно выступающий (не откусанный) штырек, соответствующий 1 выводу «микросхемы» ADUMAX, по которому легко ориентировать плату при установке в панельку.
На рис. 29 приведена схема гальванической развязки на базе ADUMAX для микроконтроллеров семейства MSC12XX.
Рис. 29. Схема гальванической развязки на базе ADUMAX для микроконтроллеров семейства MSC12XX
Как видно из рис. 29, схема формирует сигналы для микроконтроллера RxDM, RST и PSEN из сигналов интерфейса RS-232 соответственно TxD, DTR и RTS, а сигнал интерфейса RxD формируется сигналом микроконтроллера TxDM. Пунктиром показан изоляционный барьер. Схема тестировалась на скорости обмена 115 200 бод и показала отличные результаты. Длина кабеля связи с компьютером может достигать 20 м. В прямоугольнике, очерченном пунктиром, показана схема кабеля связи с компьютером.
Схема развязки достаточно проста и занимает мало места на плате.
1.4.4.2. Плата гальванически изолированного интерфейса на базе развязок ADUM1201, ADUM1200, MAX3181, MAX3183 и MAX3190 для микроконтроллеров семейства P89LPC9XX
Рис. 30. Схема гальванически изолированного преобразователя интерфейса ADUMAX1 на базе ADUM1201, ADUM1200, MAX3181, MAX3183 и MAX3190
На рис. 30 приведена схема платы гальванически изолированного интерфейса на базе развязок ADUM1201, ADUM1200, MAX3181, MAX3183 и MAX3190. Назовем такую плату «микросхемой» ADUMAX1. Из рис. 30 можно заметить, что ADUMAX1 отличается от ADUMAX (рис. 26) только тем, что два инвертирующих приемника ADUMAX — MAX3181(DD5 и DD6)—заменены на два неинвертирующих приемника MAX3183 (тe же DD5 и DD6). В остальном схемы и разводка плат идентичны.
Рис. 31. Схема гальванической развязки на базе ADUMAX1 для микроконтроллеров семейств P89LPC9XX
На рис. 31 приведена схема гальванической развязки на базе ADUMAX1 для микроконтроллеров семейства P89LPC9XX. Как видно из рисунка, схема формирует сигналы для микроконтроллера RxDM, RST и Vcc из сигналов интерфейса RS-232 соответственно TxD, DTR и RTS, а сигнал интерфейса RxD формируется сигналом микроконтроллера TxDM. Сигнал Vcc формируется на стоке КМОП-транзистора BS250 (T1), исток которого соединен с источником питания +3,3 В, а на затвор подается сигнал ТТЛ-уровня с выхода приемника ADUMAX1 (выводы 10 и 16). На вход этого же приемника подается сигнал RTS с интерфейса RS-232. Пунктиром показан изоляционный барьер. Схема тестировалась на скорости обмена в 115 200 бод и показала отличные результаты. Длина кабеля связи с компьютером может достигать 20 м. В прямоугольнике, очерченном пунктиром, показана схема кабеля связи с компьютером. Схема достаточно проста и занимает мало места на плате.
1.4.5. Новейшие микросхемы гальванических развязок
В марте 2006 года компания Silicon Laboratories анонсировала выпуск новых гальванических развязок Si8440/1/2, работающих по тому же принципу, что и развязки iCoupler от Analog Devices (например, ADUM1200/1), о которых рассказывалось в предыдущей главе. Внутренняя структура развязок приведена на рис. 32, а принцип работы и цоколевка корпуса SOIC16 — на рис. 33.
Рис. 32. Внутренняя логическая структура микросхем Si8440/1/2
Рис. 33. Микросхема Si8440/1/2 а) принцип работы и б) цоколевка
Развязки Si8440/1/2 имеют три градации скоростей: A — 0–1 Мбод, B — 0–10 Мбод и C— 0–150 Мбод. Так, например, микросхема Si8441-A-IS работает на скоростях 0–1 Мбод. Потребление тока этой развязки составляет чуть менее 5 мА на канал. Для сравнения укажем, что потребление тока развязки ADUM1200/1AR (0÷1 Мбод) составляет около 1 мА, то есть примерно в 5 раз меньше. Однако максимальная скорость работы развязок ADUM1200/1 составляет только 25 Мбод (ADUM1200/1CR). Поскольку интерфейс RS-232 работает на скоростях до 0,1 Мбод (115 200 бод), максимальная скорость развязок Si8440/1/2-C (150 Мбод) не представляет особого интереса. Кроме того, корпус у Si8440/1/2 — «широкий» SOIC (16-Pin Wide Body SOIC) шириной 7,5 мм, тогда как корпус у ADUM1200/1 — «узкий» SOIC (ширина — 4 мм). Тем не менее, развязки Si8440/1/2, по мнению автора, заслуживают внимания, особенно если их стоимость (на канал передачи) будет ниже, чем у ADUM1200/1.
В том же марте 2006 года Analog Devices анонсировала еще одно семейство своих гальванических развязок ADUM5240/1/2, которое в буквальном смысле перевернуло общее представление (в том числе и автора) о применении DC/DC-конверторов в таких развязках. Развязки ADUM5240/1/2 имеют встроенный DC/DC-конвертор! Это уникальное свойство позволяет вообще отказаться от использования DC/DC-конверторов в гальванических развязках и таким образом сэкономить, с одной стороны, $6–12, с другой — дополнительное место на плате.
Рис. 34. Внутренняя логическая структура микросхем ADUM5240/1/2
На рис. 34 приведены структурные схемы развязок ADUM5240/1/2, а на рис. 35 — цоколевка корпусов SOIC8 (узких).
Рис. 35. Цоколевка корпусов SOIC8 (узких)
Развязки ADUM5240/1/2 имеют максимальную скорость работы 10 Мбод (ADUM5240BR), а на скорости до 1 Мбод потребляют около 5 мА (на оба канала). Встроенный DC/DC-конвертор имеет КПД около 10%, то есть при выходном токе в 10 мА (5 В) входной ток составляет около 100 мА. Это, однако, не является таким уж большим недостатком, поскольку блоки питания современных компьютеров способны выдавать ток до 20 А, и лишние 100 мА совершенно не являются проблемой.
Остается только ждать, когда такие развязки появятся в продаже и у нас.
1.5. Что делать, если длина линии связи интерфейса RS-232 превышает 20 м?
Иногда возникает задача «удлинить» линию связи для интерфейса RS-232; при этом задача поставлена так, что нельзя изменять ни схему устройства на базе микроконтроллера, ни его программное обеспечение, ни программное обеспечение компьютера, с которым этот микроконтроллер связан по интерфейсу RS-232. С подобными задачами автор сталкивался достаточно часто.
Ниже приведен пример подобной ситуации и, на взгляд автора, достаточно простое решение возникшей проблемы, которая некоторых разработчиков может поставить в тупик.
Задача была поставлена следующим образом.
Имеется прибор (на базе микроконтроллера), который сопрягается с компьютером всего двумя линиями интерфейса RS-232— TxD и RxD и, естественно, «землей» — сигналом SG. Причем, связь между компьютером и прибором по интерфейсу RS-232 должна быть дуплексной (как это обычно делается в интерфейсе RS-232), то есть передатчик и приемник должны работать независимо друг от друга (так было написано программное обеспечение для компьютера и микроконтроллера в приборе).
Прибор располагался непосредственно на месте снятия показаний, а компьютер был расположен на расстоянии около одного километра от прибора (≈900м). Программное обеспечение, имеющееся на компьютере, по снятию показаний прибора было давно куплено и оплачено, поэтому его изменить уже было невозможно; кроме того, отсутствовала возможность как-то изменить программное обеспечение и в приборе, так как прибор был разработан достаточно давно. Попытки заказчика применить стандартный преобразователь интерфейса RS-232 в интерфейсе RS-485 ни к чему не привели ввиду того, что в преобразователе интерфейса RS-485 была предусмотрена только полудуплексная связь, которая обычно осуществляется в этом интерфейсе.
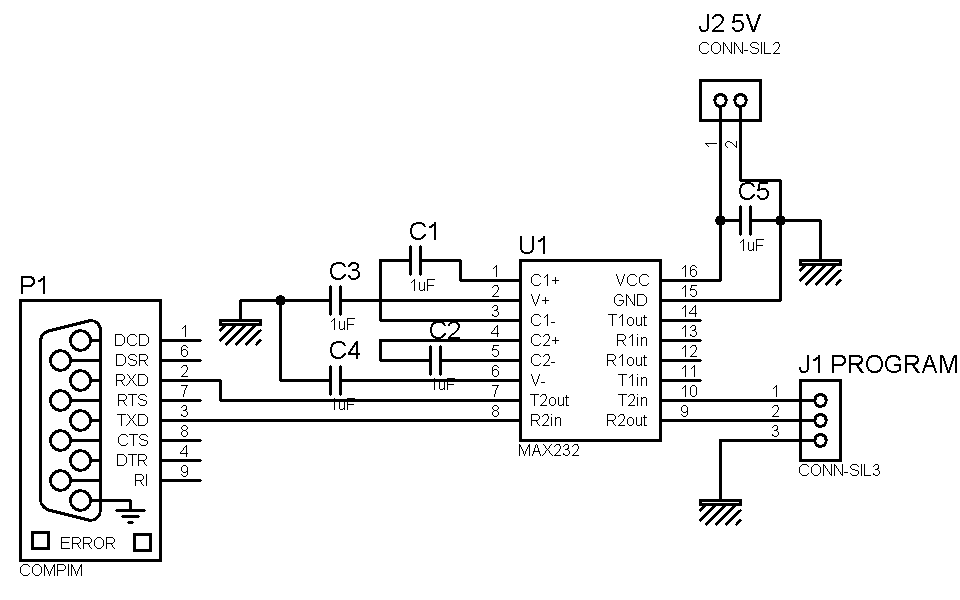
Для решения задачи автор использовал достаточно известные преобразователи RS-485/RS-422↔TTL (ADM488), работающие в дуплексном режиме, и уже упоминавшийся преобразователь интерфейса RS-232 ADM3202. Схема устройства приведена на рис. 36. Как видно из рис. 36, все сигналы интерфейсов и питание схемы выведены на клеммы (K01–K08). Для индикации включения питания используется светодиод VD1. Для питания устройства применяется достаточно дешевый стабилизированный источник питания +5 В, 200 мА.
Рис. 36. Схема двунаправленного преобразователя RS-232↔RS-485/RS-422
На рис. 37 приведены расположение элементов (а) и вариант разводки платы (б, в) устройства преобразователя интерфейсов. Использование микросхем в корпусах DIP существенно облегчает производство устройства. Для распайки компонентов не требуется высокой квалификации монтажника (вся работа по монтажу плат была довольно ловко проделана школьником 9-го класса).
Рис. 37. Вариант разводки платы двунаправленного преобразователя RS-232↔RS-485/RS-422
На рис. 38 показана фотография устройства. Как видно из рисунка, размеры платы выбраны таким образом, что она легко умещается в обыкновенную телефонную розетку и прикручивается к ней 8 саморезами, служащими одновременно крепежом клемм к плате. В крышке розетки просверлено отверстие, через которое наружу выведена верхняя часть светодиода.
Рис. 38. Фотография устройства преобразователя RS-232↔RS-485/RS-422
На рис. 39 приведена фотография общего вида двух устройств — преобразователей RS-232↔RS-485/RS-422. Слева показано устройство в собранном виде, предназначенное для сопряжения с компьютером (на фотографии можно увидеть кабель с разъемом DB9M, который подключается к COM-порту компьютера), справа — устройство со снятой крышкой.
Рис. 39. Фотография общего вида двух устройств — преобразователей RS-232↔RS-485/RS-422
На рис. 40 приведена схема сопряжения прибора с компьютером с помощью двух устройств — преобразователей RS-232↔RS-485/RS-422.
Рис. 40. Схема сопряжения прибора с компьютером с помощью двунаправленного преобразователя RS-232↔RS-485/RS-422
Для сопряжения устройств с компьютером и прибором используется обыкновенный (неэкранированный) 4-жильный телефонный провод длиной 1–1,5 м. Линия связи между двумя устройствами представляет собой достаточно дешевый провод, состоящий из двух неэкранированных витых пар в одной общей оплетке.
Тестирование устройств показало отличную их работу в дуплексном режиме на скорости в 115 200 бод на линии связи длиной в 1 км.
Литература
- Кузьминов А. Ю. Интерфейс RS-232. Связь между компьютером и микроконтроллером. М.: Радио и связь. 2004.
- Кузьминов А.Ю. Интерфейс RS-232. Связь между компьютером и микроконтроллером. От DOS к Windows98/XP. М.: ДМК-ПРЕСС. 2006 (в печати).
- Кузьминов А. Ю. Универсальная система сбора и обработки данных АСИР-3. // Мир ПК. 1996. № 6.
- Кузьминов А. Ю.Удаленные системы сбора информации с датчиков на базе однокристальных микро-ЭВМ // Автоматизация и производство. 1996. № 3.
- Кузьминов А. Ю. Однокристальные микроЭВМ—основа удаленных систем сбора и обработки сигналов, поступающих с датчиков // Электроника и компоненты. 1998. № 2.
- Кузьминов А. Ю. Новые MCS51-совместимые микроконтроллеры и их применение в системах сбора информации с датчиков // Контрольно-измерительные приборы и системы. 1997. № 6. 1998. № 7.
- www.analog.com
- www.atmel.com
- www.maxim-ic.com
- www.semiconductor-philips.com
- www.silabs.com
- www.ti.com
- www.msdn.microsoft.com/library
- www.gapdev.com
Компоненты материнской платы: назначение основных элементов
Материнская плата–это главная монтажная деталь, к которой подключаются внутренние компоненты и внешние, периферийные устройства. Она является базовым элементом любого настольного, переносного, планшетного или карманного компьютера. В этой публикации будет рассмотрено устройство материнской платы, а также назначение ее основных компонентов.
Материнская плата обеспечивает взаимодействие различных устройств ПК. Обязательными ее элементами являются разъем или socket процессора (ЦП), чип-сет, коннекторы для модулей памяти. Кроме этого, обязательными элементами ПК являются: микросхема BIOS, контролеры, различные разъемы для подачи питания, коннекторы подключения элементов и устройств компьютера.
Микросхема BIOS
BIOS представляет собой микросхему, с определенными тестовыми и загрузочными программами для ЦП, ведь на начальном этапе процессор еще не знает, как работать со всеми устройствами, расположенными в ПК. Находится эта микросхема в непосредственной близости от ЦП и запитана от отдельной батарейки — «таблетки», расположенной на плате, рядом со слотами расширения.
- Первоначальное тестирование происходит без участия процессора. Только после проверки электронных элементов компьютера на предмет отсутствия короткого замыкания, и соответствия токов в БП, BIOS «разрешает» подавать блоку питания необходимый ток на плату и процессор.
- После, ЦП считывает с БИОС определенный алгоритм действий, благодаря которым происходит тестирование работоспособности всех компонентов ПК. Если все в порядке, то производится запуск операционной системы, расположенной на «винчестере». После этого, все управление ПК передается исключительно операционной системе.
Благодаря своей работе, BIOS незаметно для пользователя выполняет одну из важнейших функций в компьютере.
Разъем центрального процессора
Разъем или socket процессора, на материнской плате самый большой. Модели разъемов ЦП различаются по внешнему виду, расположению и числу контактов. В зависимости от модели процессора, они бывают двух типов:
- Сокетный, от анг. «гнездо». Он представляет собой прямоугольный или квадратный коннектор с множеством отверстий-контактов, расположенным по его периметру. Монтируется процессор в такой разъем – горизонтально.
- Слотовый разъем, (от анг. «щель») представляет собой длинный ряд контактов, расположенных в пластмассовом корпусе. Он предназначен для вертикального монтажа ЦП. На сегодняшний день, такой тип разъемов практически не используется.
Назначение и конструктивные особенности чипсета
Чипсет материнской платы обычно представляет собой две разные микросхемы, именуемые «северным» и «южным» мостом. Свои названия они получили исключительно из расположения: северный мост находится возле ЦП, а южный – ниже, возле разъема видеокарты и слотов расширения. Они являются важнейшим связующим звеном между аппаратной частью и процессором ПК. В некоторых современных моделях системных плат может использоваться чипсет, состоящий из одной микросхемы. Это связано с конструктивными особенностями процессора, взявшего на себя функции южного моста.
Северный мост осуществляет связь ЦП с наиболее быстрыми компонентами ПК и с южным мостом. Из-за высокой нагрузки, мост достаточно сильно нагревается, поэтому, чаще всего, его оснащают радиатором для лучшего охлаждения.
- Северный мост связан с ЦП высокоскоростной шиной. Наиболее современной, на сегодняшний день, считается шина QPI, от компании Intel.
- Для связи моста с видеоадаптером, может использоваться шина AGP 3.0, или PCI Еxpress 3.0, массовый выпуск которой начался с 2012 г.
- Микросхемы мостов между собой по одному из представленных интерфейсов: PCI, Hub Link, DMI или HyperTransport.
Южная часть чипсета, согласовывает работу более медленных компонентов ПК, а также обеспечивает передачу данных с этих компонентов на северный чип.
Южный мост состоит из контроллеров: для связи с северной частью чипсета, платами расширения, периферийными устройствами и компонентами, жесткими дисками и другими медленными устройствами
Наиболее распространенными интерфейсами, в современных ПК считаются: USB 2.0 — 3.0; WI-FI и Ethernet. Для связи южного моста с жесткими дисками используются шины PCI, а также более современные шины с SATA и SCSI и SAS. Для согласования работы моста с BIOS чаще всего используется шина LPС с тактовой частотой 33,3 МГц.
Коннекторы модулей памяти и других плат расширения
В любом компьютере существует временное хранилище информации, именуемое оперативной памятью. Она представляет собой модуль с несколькими микросхемами на борту, который монтируется в специальные разъемы, так называемые слоты. Как правило, они расположены рядом с процессором и северной частью чипсета. Слоты модулей памяти оснащены замками, которые изготовлены с защитой от неправильного монтажа. Количество их варьируется от 2-х до 6,в зависимости от стоимости «материнки» и ее форм-фактора.
Важными компонентом материнской платы является слот PCI-Express, для подключения дискретного видеоадаптера, без которого невозможен вывод изображения на монитор (если только в компьютере нет интегрированной видеокарты).
Кроме вышеперечисленных слотов, на плате расположены штыревые разъемы для подключения шин передачи данных жестких дисков. Раньше, для этого использовался интерфейс IDE. На современных ПК, чаще всего используется интерфейс SATA.
На любой «материнке» в обязательном порядке есть порты, для подключения дополнительных периферийных устройств. Как правило, основной набор включает в себя: разъемы питания вентиляторов и кнопок, расположенных на панели системного блока; светодиодов, сигнализирующих о работе «винчестера» и наличии питания. Кроме того, на задней части платы находится: несколько портов USB; разъем LAN для подключения кабеля к сетевой карте; коннекторы аудиовхода и выхода; разъем HDMI и пр. В большинстве моделей можно встретить разъемы PS/2для подключения клавиатуры и манипулятора.
Для питания материнской платы, а также всех электронных и механических компонентов, на ней установлен 20 или 24 контактный разъем, на который приходит напряжение от блока питания, расположенного в корпусе ПК.
Не уверенны в выборе модели МП — довертесь нашей компании! Подберем оптимальную модель.
Типы системной платы
Форм фактор материнской платы – это определенный стандарт, благодаря которому определяется размер системной платы, ее способ крепления в корпусе ПК, наличие и расположение портов и интерфейсов шин, а также разъемов ЦП и оперативной памяти.
В настоящее время существует несколько форм-факторов системных плат:
- ATX – самая большая по величине материнская плата, которая имеет много слотов и удобное расположение портов вода — вывода.
- EATX – это системная плата ничем не отличающаяся от АТХ, кроме размеров, которые составляют 30,5 см. х33 см.
- MicroATX – это уменьшенный вариант материнской платы АТХ. Она была разработана для ПК, которым не требуется изменение конфигурации, поэтому она имеет 4 слота расширения. Как правило, такие платы оснащены 24 пиновым разъемом питания и имеют размеры, 24, 5 см. х 24,5 см.
- BTX – материнская плата, разработанная для создания небольших системных блоков. Тем не менее, она имеет 7 слотов расширения при размерах 26.7 х 32.5 см.
- Мicro BTX является уменьшенной копией «материнки» ВТХ, которая имеет 4 слота расширения и размеры 26.7 х 26.4 см.
- MiniITX – самая маленькая, из современных системных плат для компьютера. Ее размеры составляют 17 см на 17 см.
- SSIEEB и SSIСEB. Материнские платы этих форм-факторов применяются для создания серверов. Размеры составляют: 30.5 x 33.0 см. и соответственно 30.5 x 25.9 см.
Если вы решили собрать ПК, то мы рекомендуем использовать для этого материнскую плату, форм-фактора АТХ. Много свободного пространства и возможностей для изменения конфигурации, позволит в полной мере получить удовольствие от сборки и использования домашнего ПК.
Основные устройства компьютера, их назначение и взаимосвязь
По своему назначению компьютер — это универсальный прибор для работы с информацией. По принципам своего устройства компьютер — это модель человека, работающего с информацией.
Персональный компьютер (ПК) — это компьютер, предназначенный для обслуживания одного рабочего места. По своим характеристикам он может отличаться от больших ЭВМ, но функционально способен выполнять аналогичные операции. По способу эксплуатации различают настольные (desktop), портативные (laptop и notebook) и карманные (palmtop) модели ПК.
Аппаратное обеспечение. Поскольку компьютер предоставляет все три класса информационных методов для работы с данными (аппаратные, программные и естественные), принято говорить о компьютерной системе как о состоящей из аппаратных и программных средств, работающих совместно. Узлы, составляющие аппаратные средства компьютера, называют аппаратным обеспечением. Они выполняют всю физическую работу с данными: регистрацию, хранение, транспортировку и преобразование как по форме, так и по содержанию, а также представляют их в виде, удобном для взаимодействия с естественными информационными методами человека.
Совокупность аппаратных средств компьютера называют его аппаратной конфигурацией.
Видео YouTube
Программное обеспечение. Программы могут находиться в двух состояниях: активном и пассивном. В пассивном состоянии программа не работает и выглядит как данные, содержательная часть которых — сведения. В этом состоянии содержимое программы можно «читать» с помощью других программ, как читают книги, и изменять. Из него можно узнать назначение программы и принцип ее работы. В пассивном состоянии программы создаются, редактируются, хранятся и транспортируются. Процесс создания и редактирования программ называется программированием.
Когда программа находится в активном состоянии, содержательная часть ее данных рассматривается как команды, согласно которым работают аппаратные средства компьютера. Чтобы изменить порядок их работы, достаточно прервать исполнение одной программы и начать исполнение другой, содержащей иной набор команд.
Совокупность программ, хранящихся на компьютере, образует его программное обеспечение. Совокупность программ, подготовленных к работе, называют установленным программным обеспечением. Совокупность программ, работающих в тот или иной момент времени, называют программной конфигурацией.
Устройство компьютера. Любой компьютер (даже самый большой)состоит из четырех частей:
устройства ввода информации
устройства обработки информации
устройства хранения
устройства вывода информации.
Конструктивно эти части могут быть объединены в одном корпусе размером с книгу или же каждая часть может состоять из нескольких достаточно громоздких устройств
Базовая аппаратная конфигурация ПК. Базовой аппаратной конфигурацией персонального компьютера называют минимальный комплект аппаратных средств, достаточный для начала работы с компьютером. С течением времени понятие базовой конфигурации постепенно меняется.
Чаще всего персональный компьютер состоит из следующих устройств:
Системный блок
Монитор
Клавиатура
Мышь
Дополнительно могут подключатся другие устройства ввода и вывода информации, например звуковые колонки, принтер, сканер…
Системный блок — основной блок компьютерной системы. В
нем располагаются устройства, считающиеся внутренними. Устройства, подключаемые
к системному блоку снаружи, считаются внешними. Для внешних устройств
используют также термин периферийное оборудование.
Монитор — устройство для визуального воспроизведения символьной и
графической информации. Служит в качестве устройства вывода. Для настольных ПК
в настоящее время наиболее распространены мониторы, основанные на
электронно-лучевых трубках. Они отдаленно напоминают бытовые телевизоры.
Клавиатура — клавишное устройство, предназначенное для управления работой
компьютера и ввода в него информации. Информация вводится в виде
алфавитно-цифровых символьных данных.
Мышь — устройство «графического» управления.
Внутренние устройства персонального компьютера.
Внутренними считаются устройства, располагающиеся в системном блоке. Доступ к
некоторым из них имеется на лицевой панели, что удобно для быстрой смены
информационных носителей, например гибких магнитных дисков. Разъемы некоторых
устройств выведены на заднюю стенку — они служат для подключения периферийного
оборудования. К некоторым устройствам системного блока доступ не предусмотрен —
для обычной работы он не требуется.
Процессор. Микропроцессор — основная микросхема персонального компьютера.
Все вычисления выполняются в ней. Основная характеристика процессора — тактовая
частота (измеряется в мегагерцах, МГц). Чем выше тактовая частота, тем выше
производительность процессора. Так, например, при тактовой частоте 500 МГц
процессор может за одну секунду изменить свое
состояние 500 миллионов раз. Для большинства операций одного такта
недостаточно, поэтому количество операций, которые процессор может выполнить в
секунду, зависит не только от тактовой частоты, но и от сложности операций.
Единственное устройство, о существовании которого процессор «знает от рождения», — оперативная память — с нею он работает совместно. Оттуда поступают данные и команды. Данные копируются в ячейки процессора (они называются регистрами), а потом преобразуются в соответствии с содержанием команд. Более полную картину того, как процессор взаимодействует с оперативной памятью, вы получите в главах, посвященных основам программирования.
Оперативная память. Оперативную память можно представить как обширный массив ячеек, в которых хранятся числовые данные и команды в то время, когда компьютер включен. Объем оперативной памяти измеряется в миллионах байтов — мегабайтах (Мбайт).
Процессор может обратиться к любой ячейке оперативной памяти (байту), поскольку она имеет неповторимый числовой адрес. Обратиться к индивидуальному биту оперативной памяти процессор не может, так как у бита нет адреса. В то же время, процессор может изменить состояние любого бита, но для этого требуется несколько действий.
Материнская плата. Материнская плата — это самая большая плата персонального компьютера. На ней располагаются магистрали, связывающие процессор с оперативной памятью, — так называемые шины. Различают шину данных, по которой процессор копирует данные из ячеек памяти, адресную шину, по которой он подключается к конкретным ячейкам памяти, и шину команд, по которой в процессор поступают команды из программ. К шинам материнской платы подключаются также все прочие внутренние устройства компьютера. Управляет работой материнской платы микропроцессорный набор микросхем — так называемый чипсет.
Видеоадаптер. Видеоадаптер — внутреннее устройство, устанавливаемое в один из разъемов материнской платы. В первых персональных компьютерах видеоадаптеров не было. Вместо них в оперативной памяти отводилась небольшая область для хранения видеоданных. Специальная микросхема (видеоконтроллер) считывала данные из ячеек видеопамяти и в соответствии с ними управляла монитором.
По мере улучшения графических возможностей компьютеров область видеопамяти отделили от основной оперативной памяти и вместе с видеоконтроллером выделили в отдельный прибор, который назвали видеоадаптером. Современные видеоадаптеры имеют собственный вычислительный процессор (видеопроцессор), который снизил нагрузку на основной процессор при построении сложных изображений. Особенно большую роль видеопроцессор играет при построении на плоском экране трехмерных изображений. В ходе таких операций ему приходится выполнять особенно много математических расчетов.
В некоторых моделях материнских плат функции видеоадаптера выполняют микросхемы чипсета — в этом случае говорят, что видеоадаптер интегрирован с материнской платой. Если же видеоадаптер выполнен в виде отдельного устройства, его называют видеокартой. Разъем видеокарты выведен на заднюю стенку. К нему подключается монитор.
Звуковой адаптер. Для компьютеров IBM PC работа со
звуком изначально не была предусмотрена. Первые десять лет существования
компьютеры этой платформы считались офисной техникой и обходились без звуковых
устройств. В настоящее время средства для работы со звуком считаются
стандартными. Для этого на материнской плате устанавливается звуковой адаптер.
Он может быть интегрирован в чипсете материнской платы или выполнен как
отдельная подключаемая плата, которая называется звуковой картой.
Разъемы звуковой карты выведены на заднюю стенку компьютера. Для
воспроизведения звука к ним подключают звуковые колонки или наушники. Отдельный
разъем предназначен для подключения микрофона. При наличии специальной
программы это позволяет записывать звук. Имеется также разъем (линейный выход)
для подключения к внешней звукозаписывающей или звуковоспроизводящей аппаратуре
(магнитофонам, усилителям и т.п.).
Жесткий диск. Поскольку оперативная память компьютера очищается при
отключении питания, необходимо устройство для длительного хранения данных и
программ. В настоящее время для этих целей широко применяют так называемые
жесткие диски.
Принцип действия жесткого диска основан на регистрации изменений магнитного
поля вблизи записывающей головки.
Основным параметром жесткого диска является емкость, измеряемая в гигабайтах (миллиардах байтов), Гбайт. Средний размер современного жесткого диска составляет 80 — 160 Гбайт, причем этот параметр неуклонно растет.
Дисковод гибких дисков. Для транспортировки данных между удаленными компьютерами используют так называемые гибкие диски. Стандартный гибкий диск (дискета) имеет сравнительно небольшую емкость 1,44 Мбайт. По современным меркам этого совершенно недостаточно для большинства задач хранения и транспортировки данных, но низкая стоимость носителей и высокая степень готовности к работе сделали гибкие диски самыми распространенными носителями данных.
Для записи и чтения данных, размещенных на гибких дисках, служит специальное устройство — дисковод. Приемное отверстие дисковода выведено на лицевую панель системного блока.
Дисковод CD-ROM. Для транспортировки больших объемов данных удобно использовать компакт-диски CD-ROM. Эти диски позволяют только читать ранее записанные данные — производить запись на них нельзя. Емкость одного диска составляет порядка 650-700 Мбайт.
Для чтения компакт-дисков служат дисководы CD-ROM. Основной
параметр дисковода CD-ROM— скорость чтения. Она измеряется в кратных единицах.
За единицу принята скорость чтения, утвержденная в середине 80-х гг. для
музыкальных компакт-дисков (аудиодисков). Современные дисководы CD-ROM
обеспечивают скорость чтения 40х — 52х.
Основной недостаток дисководов CD-ROM — невозможность записи дисков — преодолен
в современных устройствах однократной записи — CD-R. Существуют также
устройства CD-RW, позволяющие осуществлять многократную запись.
Принцип хранения данных на компакт-дисках не магнитный, как у гибких дисков, а оптический.
Коммуникационные порты. Для связи с другими устройствами, например принтером, сканером, клавиатурой, мышью и т. п., компьютер оснащается так называемыми портами. Порт — это не просто разъем для подключения внешнего оборудования, хотя порт и заканчивается разъемом. Порт — более сложное устройство, чем просто разъем, имеющее свои микросхемы и управляемое программно.
Сетевой адаптер. Сетевые адаптеры необходимы компьютерам, чтобы они могли обмениваться данными между собой. Этот прибор следит за тем, чтобы процессор не подал новую порцию данных на внешний порт, пока сетевой адаптер соседнего компьютера не скопировал к себе предыдущую порцию. После этого процессору дается сигнал о том, что данные забраны и можно подавать новые. Так осуществляется передача.
Когда сетевой адаптер «узнает» от соседнего адаптера, что у
того есть порция данных, он копирует их к себе, а потом проверяет, ему ли они
адресованы. Если да, он передает их процессору. Если нет, он выставляет их на
выходной порт, откуда их заберет сетевой адаптер очередного соседнего
компьютера. Так данные перемещаются между компьютерами до тех пор, пока не
попадут к адресату.
Сетевые адаптеры могут быть встроены в материнскую плату, но чаще устанавливаются отдельно, в виде дополнительных плат, называемых сетевыми картами.
2 Архитектура системных плат — СтудИзба
Архитектура системных плат (Лекция 2)
Системная плата, иногда называемая «материнской платой» (motherboard) – основная плата микро-ЭВМ. Именно посредством элементов системной платы происходит взаимодействие между различенными устройствами, составляющими ПК.
Архитектура системной платы определяется набором микросхем (chipset) – это одна или несколько микросхем, таймеры, система управления специально разработанная для «обвязки» микропроцессора. Они содержат в себе контроллеры прерываний, прямого доступа к памяти, памятью и шиной – все те компоненты, которые в оригинальной IBM PC были собраны на отдельных микросхемах. Обычно в одну из микросхем набора входят также часы реального времени с CMOS-памятью и иногда – клавиатурный контроллер, однако эти блоки могут присутствовать и в виде отдельных чипов. В последних разработках в состав микросхем наборов для интегрированных плат стали включаться и контроллеры внешних устройств.
Внешне микросхемы Chipset’а выглядят, как самые большие после процессора, с количеством выводов от нескольких десятков до двух сотен.
Тип набора микросхем определяет основные функциональные возможности платы: типы поддерживаемых процессоров, структура/объем кэша, возможные сочетания типов и объемов модулей памяти, поддержка режимов энергосбережения, возможность программной настройки параметров и т.п. На одном и том же наборе может выпускаться несколько моделей системных плат, от простейших до довольно сложных с интегрированными контроллерами портов, дисков, видео и т.п.
Главными структурными элементами системной платы являются северный и южный мосты.
Северный мост служит для скоростной связи между процессором, оперативной памятью и видеоадаптером, подключенным к высокоскоростной шине (PCI-E или AGP).
Южный мост предназначен для связи с контроллерами портов и периферийных устройств. Также на южном мосте находится BIOS и контроллеры устройств ввода-вывода (I/O, Input/Output devices).
Мосты соединены между собой внутренней шиной, которая обеспечивает связь процессора с периферийными устройствами.
Рекомендуемые файлы
Структурная схема современной системной платы представлена на рис. 1.
Рис. 1. Структурная схема современной системной платы.
Шины ввода-вывода (XT, ISA, EISA, MCA, VESA, PCI).
Любая вычислительная машина обязательно включает в себя такие базовые компоненты как процессор – для проведения вычислительных операций или выполнения программ, запоминающие устройства – как для хранения промежуточной информации в процессе вычислений, так и для длительного хранения и перемещения информации в пространстве. Кроме того, каждая вычислительная машина конфигурируется периферийными устройствами для ввода и вывода информации. Несмотря на всю значимость указанных узлов вычислительной машины, не менее важным устройством является канал передачи данных от одного узла ЭВМ к другому – шина ввода/вывода.
На заре компьютерной техники ПК оснащались 8-битовой XT-шиной, которая устарела уже с появлением процессора 80286 (РС АТ). ПК данного класса стали оснащаться шиной ISA, которую иногда еще называют АТ по названию класса. Новая шина стала 16-битовой, но сохраняла при этом полную физическую и электронную совместимость с существующими 8-битовыми картами, предназначенными для шины оригинального РС.
Шина EISA также построена по принципу полной совместимости со старыми конструкциями. Она является 32-битовым расширением шины ISA и так же совместима с 8- и 16-битовыми картами. В отличие от шины МСА – старой 32-битовой шины, разработанной фирмой IBM для PS/2, EISA совместима с шинами ISA и РС.
Шины EISA и МСА являются интеллектуальными. В шинах и совместимых картах были предусмотрены аппаратные средства, позволяющие системе идентифицировать новые карты дополнительных устройств непосредственно при их подключении к шине. Аппаратные средства данных шин конфигурировались автоматически, обеспечивая, фактически, всеми любимый Plug-and-Play.
Несмотря на это, в мире ПК доминировала шина ISA. Это можно объяснить значительной дешевизной как самой шины, так и совместимых с ней 8- и 16-битовых карт. Кроме того, впоследствии появились карты ISA, не требующие ручной конфигурации (переключателей и джамперов), что сделало шину еще привлекательнее.
Усовершенствованием технологии шин ПК стало использование локальных шин для обмена данными с периферийными устройствами. Локальные шины, по сути, соединяют процессор с памятью. Они «быстрые и широкие», 32- 64-битовые. Скорость передачи данных по ним зависит от скорости работы микропроцессора. Связь между процессором и памятью настолько важна, что традиционно к локальной шине больше ничего не подключали. Однако производители видеоконтроллеров изменили эту традицию, что привело к большим изменениям в технологии компьютерных шин. Эти изменения вызваны тем, что шина ISA недостаточно быстра, чтобы поддерживать режим SVGA и высокое разрешение. Один образ экрана SVGA требует от шины передачи сотен килобайт и даже мегабайт данных. Передача такого количества данных по медленной шине, которая к тому же передает данные и другим периферийным устройствам, снижает эффективность работы видеоадаптера. Проблему решили созданием стандарта, который позволил видеоконтроллеру соединяться прямо с локальной шиной ПК – локальная шина VESA.
Шина VESA имеет несколько существенных ограничений, но исправно выполняет свои функции – высокоскоростная передача данных видеоконтроллеру. Но VESA использовалась только для этого и не являлась шиной ввода/вывода общего назначения. Следующая версия данного стандарта стала более надежной и универсальной, но тоже не получила широкого применения. Наиболее широко используемой стала шина PCI.
Шина PCI (Peripheral Component Interconnect) не имеет ограничений, присущих более ранним стандартам. К основным преимуществам шины можно отнести:
· Широкая сфера применения. Крупнейшие производители аппаратного обеспечения ПК стали развивать и продвигать стандарт PCI. Шина универсальна и используется не только для видеокарт, но и для других устройств.
· Стандартная тактовая частота. Локальная шина работает на тактовой частоте внешнего генератора процессора. В разных системах значение частоты может отличаться, поэтому при подключению адаптеров к локальной шине производителям приходилось предусматривать поддержку различных частот. Интерфейс PCI решает эту проблему, стабильно поддерживая тактовую частоту 33 МГц, и облегчает задачи производителей контроллеров.
· Стандартный разъем.
· Интеллектуальность. Шина защищает жизненно важную связь между процессором и памятью и позволяет множеству устройств использовать локальную шину. Полная поддержка Plug-and-Play.
· Низкая стоимость по сравнению с другими шинами такого уровня.
Максимальная скорость передачи данных через шину PCI – 132 Мбит/с. Это позволяет эффективно использовать этот стандарт при работе в сети. В течение достаточно долгого времени производители обеспечивали совместимость своих ПК со старыми адаптерами за счет включения второй шины – обычно EISA или ISA. Наиболее распространенной конфигурацей были несколько слотов PCI для быстрого ввода/вывода и ISA для совместимости со старыми адаптерами. Однако в настоящее время стандарт ISA является сильно устаревшим и более не используется и не поддерживается производителями.
Сравнение и характеристики шин.
XT-Bus – шина архитектуры XT – первая в семействе IBM PC. Относительно проста, поддерживает обмен 8-разрядными данными внутри 20-разрядного (1 Мб) адресного пространства (обозначается как «разрядность 8/20»), работает на частоте 4.77 МГц. Совместное использование линий IRQ в общем случае невозможно. Конструктивно оформлена в 62-контактних разъемах.
ISA (Industry Standard Architecture, архитектура промышленного стандарта) – основная шина на компьютерах типа PC AT (другое название – AT-Bus). Является расширением XT-Bus, разрядность – 16/24 (16 Мб), тактовая частота – 8 МГц, предельная пропускная способность – 5.55 Мб/с. Разделение IRQ также невозможно. Возможна нестандартная организация Bus Mastering, но для этого нужен запрограммированный 16-разрядный канал DMA. Конструктив – 62-контактный разъем XT-Bus с прилегающим к нему 36-контактным разъемом расширения.
EISA (Enhanced ISA, расширенная ISA) – функциональное и конструктивное расширение ISA. Внешне разъемы имеют такой же вид, как и ISA, и в них могут вставляться платы ISA, но в глубине разъема находятся дополнительные ряды контактов EISA, а платы EISA имеют более высокую ножевую часть разъема с дополнительными рядами контактов. Разрядность – 32/32 (адресное пространство – 4 Гб), работает также на частоте 8 МГц. Предельная пропускная способность – 32 Мб/с. Поддерживает Bus Mastering — режим управления шиной со стороны любого из устройств на шине, имеет систему арбитража для управления доступом устройств к шине, позволяет автоматически настраивать параметры устройств, возможно разделение каналов IRQ и DMA.
MCA (Micro Channel Architecture, микроканальная архитектура) – шина компьютеров PS/2 фирмы IBM. Не совместима ни с одной другой, разрядность – 32/32, (базовая — 8/24, остальные – в качестве расширений). Поддерживает Bus Mastering, имеет арбитраж и автоматическую конфигурацию, синхронная (жестко фиксирована длительность цикла обмена), предельная пропускная способность – 40 Мб/с. Конструктив – одно-трехсекционный разъем (такой же, как у VLB). Первая, основная, секция – 8-разрядная (90 контактов), вторая – 16-разрядное расширение (22 контакта), третья – 32-разрядное расширение (52 контакта). В основной секции предусмотрены линии для передачи звуковых сигналов. Дополнительно рядом с одним из разъемов может устанавливаться разъем видеорасширения (20 контактов). EISA и MCA во многом параллельны, появление EISA было обусловлено собственностью IBM на архитектуру MCA.
VLB (VESA Local Bus, локальная шина стандарта VESA) – 32-разрядное дополнение к шине ISA. Конструктивно представляет собой дополнительный разъем (116-контактный, как у MCA) при разъеме ISA. Разрядность – 32/32, тактовая частота – 25..50 МГц, предельная скорость обмена – 130 Мб/с. Электрически выполнена в виде расширения локальной шины процессора – большинство входных и выходных сигналов процессора передаются непосредственно VLB-платам без промежуточной буферизации. Из-за этого возрастает нагрузка на выходные каскады процессора, ухудшается качество сигналов на локальной шине и снижается надежность обмена по ней. Поэтому VLB имеет жесткое ограничение на количество устанавливаемых устройств: при 33 МГц – три, 40 МГц – два, и при 50 МГц – одно, причем желательно интегрированное в системную плату.
PCI (Peripheral Component Interconnect, соединение внешних компонент) – развитие VLB в сторону EISA/MCA. Не совместима ни с какими другими, разрядность – 32/32 (расширенный вариант – 64/64), тактовая частота – до 33 МГц (PCI 2.1 – до 66 МГц), пропускная способность – до 132 Мб/с (264 Мб/с для 32/32 на 66 МГц и 528 Мб/с для 64/64 на 66 МГц), поддержка Bus Mastering и автоконфигурации. Количество разъемов шины на одном сегменте ограничего четырьмя. Сегментов может быть несколько, они соединяются друг с другом посредством мостов (bridge). Сегменты могут объединяться в различные топологии (дерево, звезда и т.п.). Самая популярная шина в настоящее время; используется также на компьютерах, отличных от IBM-совместимых. Разъем похож на MCA/VLB, но чуть длиннее (124 контакта). 64-разрядный разъем имеет дополнительную 64-контактную секцию с собственным ключом. Все разъемы и карты к ним делятся на поддерживающие уровни сигналов 5 В, 3.3 В и универсальные; первые два типа должны соответствовать друг другу, универсальные карты ставятся в любой разъем.
Существует также расширение MediaBus, введенное фирмой ASUSTek – дополнительный разъем содержит сигналы шины ISA.
PCMCIA (Personal Computer Memory Card International Association, ассоциация производителей плат памяти персональных компьютеров) – внешняя шина компьютеров класса NoteBook. Другое название модуля PCMCIA – PC Card. Предельно проста, разрядность – 16/26 (адресное пространство – 64 Мб), поддерживает автоконфигурацию, возможно подключение и отключение устройств в процессе работы компьютера. Конструктив – миниатюрный 68-контактный разъем. Контакты питания сделаны более длинными, что позволяет вставлять и вынимать карту при включенном питании компьютера.
Основные микросхемы IBM PC
Все интегральные микросхемы состоят из огромного множества микроскопических полупроводниковых транзисторов и других элементов радио цепей, которые составляют сложные схемы внутри корпуса микросхемы.
Базовой схемой всех упраляющих микросхем явлеятся схема RS – триггера, реализованного с помощью логических элементов И — НЕ или ИЛИ — НЕ с соответствующими обратными связями и позволяет хранить один бит цифровой информации.
При подаче на вход R уровня лог. «1» триггер принимает состояние логического «0», а при подаче управляющего сигнала «1» на вход S — состояние «1». Следует отметить также, что если до подачи управляющего сигнала, например, на вход R, триггер находился в состоянии логического «0», его состояние не изменится и после подачи сигнала «1» на вход R. Если на обоих входах триггера имеются уровни логического «0» – это состояние соответствует режиму хранения, и триггер сохраняет предыдущее состояние. При подаче на входы R и S одновременно уровня «1» триггер будет находиться в неопределенном состоянии, поэтому такое сочетание сигналов R и S называется запрещенной комбинацией управляющих.
Главной микросхемой PC является микропроцессор. Рядом с микропроцессором предусмотрено место для микросхемы 80X87, числового сопроцессора, или процессора числовых данных, с его специальными возможностями по выполнению очень быстрых (и с повышенной точностью) вычислений над числами с плавающей точкой.
Среди специализированных микросхем можно выделить генератор тактовых (или синхронизирующих) сигналов. В IBM PC АТ это микросхема 88248. В микросхеме генератора тактовых сигналов используется кварцевый кристалл в качестве точной основы для синхронизации. Генератор тактовых сигналов понижает частоту колебаний кристалла до частоты, требующейся компьютеру, и преобразует их в форму, приемлемую для использования другими компонентами схемы.
С генератором тактовых сигналов близко связана микросхема программируемого таймера, идентифицируемая номером 8253. Программируемый таймер может порождать другие сигналы синхронизации.
Некоторые компоненты компьютера могут обмениваться данными непосредственно через компьютерную память, без участия микропроцессора. Такой процесс называется прямым доступом к памяти (Bus Mastering). Имеется специальная микросхема, предназначенная для управления этим процессом, которая называется контроллером прямого доступа к памяти (номер микросхемы – 8237).
Прерывания контролируются специальной микросхемой 8259. В компьютерах прерывания поступают с различной степенью важности, и одной из задач контроллера прерываний является отслеживания их в порядке приоритетов.
К другим основным микросхемам относится микросхема программируемого интерфейса периферийных устройств (номер – 8255). Эта микросхема следит за работой некоторых из более простых периферийных устройств компьютера. Однако большинство периферийных устройств компьютера являются слишком сложными для того, чтобы они могли регулироваться простой обычной схемой. Для управления такими устройствами (дисковые накопители, монитор и т.д.) используются специальные контроллеры.
BIOS
В лекции «Способы управления обменом данными» также много полезной информации.
Одной из главных микросхем системной платы является микросхема BIOS – это основная система ввода/вывода (Basic Input/Output System), «зашитая» в ПЗУ (отсюда название ROM – read only memory BIOS). Она представляет собой набор программ проверки и обслуживания аппаратуры компьютера, и исполняет роль посредника между операционной системой и аппаратурой. BIOS получает управление при включении и «сбросе» системной платы, тестирует саму плату и основные блоки компьютера (видеоадаптер, клавиатуру, контроллеры дисков и портов ввода/вывода), настраивает Chipset платы и загружает внешнюю операционную систему. При работе под DOS/Windows BIOS управляет основными устройствами, при работе под OS/2, UNIX, WinNT BIOS практически не используется, выполняя лишь начальную проверку и настройку.
Обычно на системной плате установлено только ПЗУ с системным (Main, System) BIOS, отвечающим за саму плату и контроллеры FDD, HDD, портов и клавиатуры; в системный BIOS практически всегда входит System Setup – программа настройки системы. Видеоадаптеры и контроллеры HDD с интерфейсом ST-506 (MFM) и SCSI имеют собственные BIOS в отдельных ПЗУ; их также могут иметь другие платы – интеллектуальные контроллеры дисков и портов, сетевые карты и т.п.
Обычно BIOS для современных системных плат разрабатывается одной из специализирующихся на этом фирм, однако некоторые производители плат сами разрабатывают BIOS’ы для них. Иногда для одной и той же платы имеются версии BIOS от разных производителей – в этом случае допускается копировать «прошивки» или заменять микросхемы ПЗУ; в общем же случае каждая версия BIOS привязана к конкретной модели платы.
Раньше BIOS «зашивался» в однократно программируемые ПЗУ либо в ПЗУ с ультрафиолетовым стиранием; сейчас в основном выпускаются платы с электрически перепрограммируемыми ПЗУ (Flash ROM), которые допускают перешивку BIOS средствами самой платы. Это позволяет исправлять заводские ошибки в BIOS, изменять заводские умолчания, программировать собственные экранные заставки и т.п.
Тип микросхемы ПЗУ обычно можно определить по маркировке: 27xxxx – обычное ПЗУ, 28xxxx или 29xxxx – flash. Если на корпусе микросхемы 27xxxx есть прозрачное окно – это ПЗУ с ультрафиолетовым стиранием, которое можно «перешить» программатором; если окна нет – это однократно программируемое ПЗУ, которое в общем случае можно лишь заменить на другое.
Радио86РК — советский самодельный компьютер / Хабр
Вы когда-нибуть собирали компьютер? Ну хотя бы один раз в жизни. Хотя бы не полностью.
— «Ну конечно!» — скажете вы.
И будете не совсем правы.
Нет, я ни в коем разе не умаляю ваших достижений, однако, в далеком 1986 году в СССР собрать компьютер могли лишь опытные радиолюбители. Это сейчас достаточно купить материнскую плату, процессор, достать пару планок оперативной памяти и жесткий диск на побольше, и водрузить это все в красивый корпус с блоком питания на 500W. Все это, заметьте, делается без помощи паяльника, осциллографа и кучи принципиальных схем.
Лишь немногие читатели Хабра могут похвастаться, что в советское время действительно собирали бытовые компьютеры. Доставали честными и нечестными путями дефицитные микросхемы, вытравливали печатные платы, до ночи воевали с УФ ПЗУ, вооружившись тумблерным программатором или наматывали трансформаторы для блоков питания. Спустя несколько месяцев кропотливой сборки и наладки наконец-то получалось то, что уже можно гордо называть бытовым компьютером. А если удавалось найти или сделать подходящий и красивый корпус для своего творения, то такой компьютер вызывал зависть у всех, кто еще не прокачал свои навыки крафта до 80 уровня.
Одним из наиболее популярных самодельных бытовых компьютеров стал Радио86РК, описание и схема которого были опубликованы в цикле статей в журнале «Радио» № 4-6/1986 г. Два автора этих статей, Г. Зеленко и С. Попов, в соавторстве с В.В. Пановым, впоследствии выпустили книгу «Домашний компьютер» (Издательство «Радио и Связь»), целиком и полностью посвященную компьютеру Радио86РК.
Радио86РК — не первый бытовой компьютер, который предлагался для сборки радиолюбителями. Его предшественником был «Микро-80», состоявший из большого количества модулей, и насчитывавший порядка 200 микросхем. Он был сложен в сборке и наладке, поэтому особого распространения не получил.
Один из авторов компьютера «Микро-80». Изображение взято с сайта http://mrsaing.my1.ru
А вот Радио86РК состоял всего лишь из 29 микросхем и небольшого количества других радиодеталей, был прост в сборке и, если не было допущено ошибок в монтаже, начинал работать сразу после включения питания. Однако, из-за попадавшихся бракованных радиодеталей, Радио86РК мог и не запуститься, и вот тогда приходилось брать в руки щупы осциллографа, и искать неисправность.
Лишь потом, когда государство приняло решение выпускать радиолюбительские конструкторы (и даже производить клоны радиолюбительских бытовых компьютеров) серийно, жить радиолюбителям стало немного проще.
Об одном из клонов Радио86РК, серийно выпускавшемся на производстве, уже было написано на Хабре в обзоре ПК «Микроша» — один из клонов «Радио-86РК», однако у автора этот бытовой компьютер корректно работать отказался из-за выкушенных конденсаторов КМ, которые, из-за содержащихся в них драгметаллов, в свое время, видимо, сдали в цветмет.
Микроша. Изображение взято с сайта wikipedia.org
К тому же, «Микроша» лишь частично совместим со своим прародителем, поэтому многие программы и игры для Радио86РК, не смогут запуститься на нем без соответствующей адаптации.
Помимо «Микроши», на страницах Хабра был обзор радиолюбительского компьютера Орион-128, за основу которого, судя по всему, был взят Радио86РК. Орион-128 имел гораздо больше оперативной памяти, чем Радио86РК, к нему подключался ROM-диск и дисковод, также была возможность подключить через специальный контроллер жесткий диск до 10Mb.
Но до сих пор никто не написал обзор про родителя всех этих многочисленных клонов, а именно про Радио86РК. Сегодня я хотел бы исправить это досадное недоразумение. Итак, начнем.
Технические характеристики Радио86РК.
Процессор: К580ИК80А (КР580ВМ80А) — советский аналог процессора I8080А, работающий на частоте 1.78 МГц. На самом деле, номинальная тактовая частота этого процессора составляла 2.5 МГц, однако для упрощения схемы бытового компьютера Радио86РК было решено отказаться от тактового генератора для видеоконтроллера, и использовать в качестве него генератор тактовой частоты процессора. В связи с тем, что видеоконтроллер переваривал только частоту 1.78 МГц, приходилось довольствоваться заниженной частотой процессора.
ПЗУ: 2 КБ на микросхеме с ультрафиолетовым стиранием К573РФ2 или К573РФ5 (буржуйский аналог — 2716) в которой размещена управляющая программа «Монитор». Во время отладки компьютера, вместо «Монитора», в ПЗУ записывали диагностический тест, проверяющий корректную работу памяти, знакогенератора и клавиатуры. Поэтому данную микросхему обычно устанавливали на панельку.
Оперативная память: в оригинальном варианте — 16 КБ на восьми микросхемах К565РУ3. В доработанном варианте — 32КБ на шестнадцати микросхемах К565РУ6 (напаенных друг на друга вторым этажом). Для нормальной работы компьютера Радио86РК с 32 КБ оперативной памяти было необходимо изменить прошивку ПЗУ «Монитора» в связи с изменением конечных адресов ОЗУ.
Видеоподсистема: построена на связке микросхем КР580ВГ75 + КР580ВТ57. Символы для знакогенератора прошиты в микросхеме ПЗУ К573РФ1. Данная микросхема требовала для своего питания аж целых три напряжения: +5, +12 и -5 вольт, и часто выходила из строя в случае пропадания одного из напряжений питания. Многие радиолюбители заменяли ее на микросхему К573РФ5, благо изменения в схему вносились небольшие. Доступен только текстовый черно-белый режим 25 строк на 64 символа, не смотря на то, что микросхема КР580ВГ75 позволяла реализовать цветное изображение. Для простоты конструкции от цветного изображения решили отказаться.
В связи с дефицитностью микросхемы КР580ВГ75, некоторые радиолюбители заменяли ее на отдельную плату, содержащую около двадцати более распространенных микросхем. Правда после такой замены на компьютере некорректно работали программы, обращающиеся к видеопамяти напрямую, в частности — игры. Без адаптации работали только те программы, которые обращались к видеопамяти через функции ПЗУ.
В качестве монитора использовался обычный телевизор или видеомонитор Электроника МС 6105.
Клавиатура представляла собой матрицу с контактами на замыкание, которая полностью обслуживалась ЦПУ через порт ввода-вывода, выполненного на одной из микросхем КР580ВВ55 (всего таких микросхем в компьютере две). В клавиатуре чаще всего использовались герконовые кнопки от калькуляторов, которые не отличались плавностью хода, зато имели высокую надежность.
В качестве источника звука использовался вывод процессора INTE (на котором формировался сигнал «разрешение прерываний»), переключаемый командами EI/DI. Поскольку прерывания в компьютере не использовались, этот вывод был свободен. Не пропадать же добру, ведь правда?
Блок питания: один из наиболее важных узлов компьютера, проектировавшийся с максимально возможной надежностью, ведь из-за его неисправности могли выйти из строя довольно дефицитные и дорогостоящие микросхемы. Дело в том, что для своей работы компьютер Радио86РК использовал три напряжения питания (-5В,+5В и +12В), причем первым должно подаваться напряжение -5В, а затем +5В и +12В. Сниматься же они должны в обратном порядке. Также допускается одновременная подача и снятие всех напряжении, а при пропадании одного из них должны сниматься и все остальные.
В качестве внешнего устройства для хранения программ и данных применялся бытовой кассетный магнитофон.
Периферийные устройства (например, принтер) подключались свободному порту на второй микросхеме КР580ВВ55.
Внешний вид Радио86РК.
Говорить о внешнем виде самодельного компьютера не совсем корректно.
Каждый старался как мог, и делал корпус под свой РК86 из того, что под руку попадется. Некоторые вообще не заморачивались, и оставляли компьютер совсем без корпуса, а потом холили и лелеяли его — не дай бог пролить на него пиво чай, или убить статикой.
Радиолюбители, постигшие Дзен, старались спрятать свое детище в корпуса от советских калькуляторов соответствующих габаритов (например, Искра 111М), а те люди, которые умели не только держать в руках паяльник, но и работать с оргстеклом делали свои собственные корпуса.
Радио86РК в корпусе от калькулятора Искра 111М. Изображение взято с сайта http://sfrolov.livejournal.com
А теперь, давайте посмотрим на корпусное исполнение моего экземпляра Радио86РК.
Он сделан из оргстекла, и покрашен в желтый цвет. В верхней крышке корпуса вырезаны отверстия для кнопок и двух светодиодов.
Клавиатура набрана из кнопок от советских калькуляторов. Буквы и стрелки приходилось рисовать на кнопках вручную — сверху таких кнопок имеются прозрачные колпачки, защищающие надпись от стирания. Цифры на кнопках рисовать было не нужно, так как они уже были нанесены заводским способом на кнопки каждого уважающего себя калькулятора.
По бокам корпуса нет ничего интересного, а вот на задней стенке расположились все необходимые разъемы — разъем для подключения магнитофона, телевизора, кнопка «Сброс», и разъем для блока питания.
Корпус блока питания также сделан из оргстекла и покрашен в желтый цвет.
Помимо разъема питания Радио86РК и кнопки включения, на передней стенке корпуса БП имеется розетка +5В, которая, в принципе, не нужна. Но это не баг, это фича (с).
Боковая стенка блока питания — гигантский радиатор. Вот таким образом были решены проблемы с охлаждением.
Что же у него внутри?
Снимаем верхнюю часть корпуса — никаких винтов, просто тянем вверх.
Нашему взору предстает клавиатура, соединенная шлейфом с основной платой компьютера.
Открутив 4 винтика и отсоединив шлейф от клавиатуры, можно добраться до основной платы компьютера Радио86РК. Печатная плата — двухсторонняя, довольно качественно вытравленная. Однако из-за ошибок разводки некоторые дорожки перерезаны и проложены с помощью обычного провода.
Очень многие микросхемы установлены на панельку. Например, процессор.
Оперативная память также гнездится в панельках. Да еще и вторым этажом.
Знакогенератор. Вместо отечественной микросхемы К573РФ1, он выполнен на немецком аналоге U555. Так надежней.
Микросхема К573РФ5 (ПЗУ «Монитора») также заменена на аналогичную ей микросхему 2716.
Генератор тактовой частоты КР580ГФ24. Тоже в панельке.
Клавиатурный порт ввода-вывода. Рядом пустая панелька для такой же микросхемы, предназначенной для подключения 8-ми битных периферийных устройств.
Видеоконтроллер.
Напоминаю, что всю эту красоту радиолюбитель должен собрать сам, руководствуясь описанием и схемами из журнала «Радио». А помочь в кропотливой сборке и наладке должны были справочные таблицы, опубликованные там же.
Это выглядело примерно так.
Делу время, потехе — час.
Многим наверняка будет интересно, во что же играли суровые дядьки, сумевшие собрать бытовой компьютер Радио86РК. Оказывается, для РК86 было написано довольно приличное количество программ, в том числе и развлекательного характера.
Хотите на них взглянуть? Пожалуйста!
Сперва подключим Радио86РК к монитору и магнитофону. Теперь вставляем кассету с любимой игрой, жмем на компьютере кнопку I (начальная буква слова «Input»), после нее не забываем нажать клавишу ВК (возврат каретки, аналог клавиши Enter), и включаем магнитофон на воспроизведение. Монитор волшебным образом перестает выводить что-либо на экран (это особенность работы компьютера), и лишь после загрузки программы изображение снова появляется. Загрузка завершена.
Для запуска программы необходимо нажать кнопку G (начальная буква слова «GO»), и подтвердить команду нажатием клавиши ВК. Иногда необходимо указывать начальный адрес памяти, где располагается программа, но это бывает очень редко.
Давайте же посмотрим, во что можно было поиграть на компьютере Радио86РК.
Червяк.
Крестики-нолики
Цирк
Lode Runner
Pacman
Тетрис
Заключение.
Вот, в принципе, и все, что я хотел рассказать о советском самодельном бытовом компьютере Радио86РК. Возможно кто-то из читателей Хабра в свое время занимался сборкой такого компьютера, или работал на нем. Если вы можете поделиться своими впечатлениями, замечаниями и просто интересными фактами о Радио86РК, я буду вам признателен.
РадиоКот :: USB 1.1 хаб. Light
РадиоКот >Схемы >Цифровые устройства >Примочки к ПК >USB 1.1 хаб. Light — версия
В последнее время ощущается повышенный интерес радиолюбительской общественности к шине USB. Интерес этот проявляется во все возрастающем количестве разработок, использующих этот интерфейс для связи с ПК, причем интерфейс может быть реализован как с помощью специализированных микросхем (типа FT232BM или TUSB3410), так и полностью программными способами. Однако у нас речь пойдет о том, что при таком изобилии USB устройств пользователи не слишком современных ПК сталкиваются с нехваткой портов для подключения этих устройств. Одним из способов решение этой проблемы является использование USB хабов, причем, как оказалось, их вполне возможно изготовить самостоятельно.
Для создания нашего собственного хаба мы будем использовать специализированные чипы TUSB2046 и TUSB2077 от Texas Instr., являющиеся, соответственно, 4- и 7-ми портовыми хабами с поддержкой USB версии 1.1. Конечно, сейчас версия стандарта 1.1 постепенно уходит в прошлое, однако часто в радиолюбительской практике ее более чем достаточно, да и обратная совместимость устройств не слишком сильно снижает функциональность.
Итак, познакомимся с этими микросхемами поподробнее. Выпускаются они в 32- и 48-ми выводном LQFP корпусе соответственно, шаг выводов 0.8 и 0.5 мм. Микросхемы имеют интерфейс для подключения внешней EEPROM памяти, в которой могут храниться уникальные идентификационные коды производителя и типа устройства, однако, поскольку мы такими кодами не располагаем, да и вообще собираем лайт-версию устройства, использовать эту возможность мы не будем. Оба чипа требуют для своего питания напряжения 3.3В и могут работать в режимах питания либо от шины USB, либо от внешнего источника питания. В качестве генератора тактовой частоты мы будем использовать подключенный к микросхеме кварц на 6MHz.
Рассмотрим более подробно назначение некоторых выводов микросхем.
Выводы DP0 и DM0 служат для подключения микросхемы к хосту, т.е. к ПК. DP0 – это Data+, DM0 – Data-. DP и DM выводы с другими индексами служат для подключения дочерних устройств, это так называемые downstream порты. Выводы PWRON служат для индикации активности данного порта и могут использоваться для подключения светодиодов, активный уровень низкий. OVRCUR – входы датчиков перегрузки по току для каждого порта: если в процессе работы на таком входе появляется низкий уровень, соответствующий порт отключается. Эти входы, как и выходы PWRON служат, в основном, для работы со специализированными микросхемами-мониторами питания шины USB и мы их использовать не будем. Низкий уровень на входе EXTMEM активирует интерфейс внешней EEPROM памяти, поэтому нам надо будет подать туда высокий уровень. При отключенной памяти вход EEDATA/GANGED заведует типом мониторинга перегрузки портов: индивидуальный или групповой. Подаем туда 0 для TUSB2077 (групповой мониторинг: при детектировании 0 на любом из входов OVRCUR все порты отключаются), соединяем все входы OVRCUR между собой и подцепляем к Vcс – перегрузки у нас никогда не будет (для TUSB2046 сигнал GANGED имеет обратную полярность, однако поскольку отключения портов не происходит, что туда подавать, 0 или 1, не так важно, главное объединить и OVRCUR и подать на них 1. Для TUSB2046 выводы TSTMODE и TSTPLL/48MCLK определяют частоту тактирования и должны быть заземлены для работы от 6MHz кварца, для TUSB2077 ту же операцию надо проделать с выводом MODE. Вывод BUSPOWER определяет источник питания для хаба и дочерних устройств: шина USB или внешний БП. Например, в режиме питания от шины TUSB2077 отключает 3 последних порта (потому как считается, что больше 4 устройств один канал шины USB по питанию выдержать не может). А оно нам надо? Заземляем этот вывод и говорим хабу, что у него есть внешний источник питания. Вот, в принципе, все то основное, что нам надо знать. У хабов есть еще некоторые выводы, предназначенные для подключения светодиодов или других устройств диагностики и отображения текущего состояния микросхемы, которые могут быть весьма полезны в ряде случаев. Все они описаны в документации на эти чипы.
Таким образом, получаем следующую конфигурацию: 4- или 7-ми портовый хаб, без EEPROM (определяется системой как Generic USB Hub), с питанием от внешнего источника, без защиты от перегрузки портов. Поскольку защиты нет, все же задумывайтесь над тем, что и в каком количестве вы подключаете к хабу и используйте соответствующий блок питания.
В принципе, с некоторыми оговорками, наше включение соответствует следующим типовым схемам:
У нас, соответственно, не используются мониторы питания TPS2044 и микросхемы защиты от перенапряжения SN75240.
В качестве системы сброса можно использовать RC-цепь, подходящий супервизор или просто подтянуть этот вход к плюсу питания – все работает.
Для получени необходимых для питания хаба 3.3В я использовал LDO-стабилизатор REG102NA-3.3/250 в корпусе SO-8 от того же Texas»a, но здесь также подойдет параметрический стабилизатор, LM317 (только не забываем, что это регулируемый стабилизатор, и для получения искомых 3,3В потребуется два резистора и вдумчивое чтение даташита!) или просто два последовательных кремниевых диода к +5В (хотя последний вариант все же не желателен).
Несколько удивили резисторы 15к на линиях данных USB, однако без них при подключении хаба к ПК Windows находит на каждом из портов «Неизвестное устройство» и переубедить его не удается. Во всех линиях данных также стоят последовательные резисторы, но здесь как раз все понятно – согласование и защита – они нужны обязательно и номинал их 22-24 ома.
На печатной плате сделана разводка сразу под оба чипа, однако установлены индивидуальные разъемы USB и питания для каждого из них. Это позволит при необходимости разделить плату на две части и использовать только нужную. Если оба чипа установлены на единой плате, то для их питания достаточно одного 3.3В стабилизатора (хотя посадочное место для него есть возле каждого чипа) и питание на второй чип подается посредством перемычки, земля и питание 5В на плате общие.
В принципе, можно 5В с USB разъемов через диоды Шоттки подать на общую шину питания, тогда для работы хаба не потребуется дополнительного внешнего источника питания, но здесь следует очень осторожно подходить к вопросу подключения устройств: все же 7 устройств один USB порт компьютера может по питанию и не вытянуть.
Печатная плата разведена в SL5 и сделана двухсторонней, так что внимательно смотрите, что там куда зеркалить, если дело дойдет до утюга. Дорожки под TUSB2077 0.2мм, шаг выводов 0.5мм. Я паял обычной паяльной станцией. При некоторой сноровке и достаточном количестве флюса пайка весьма качественная, нет ни залипаний, ни непропаев.
В качестве эксперимента, можно соединить вход одного хаба с одним из downstream портов другого – получим 10-ти портовый хаб.
Готовое устройство выглядит так:
Я планирую встраивать такие хабы в устройства или комплексы, содержащие несколько компонентов, подключаемых по USB (например, в проектируемом ресивере управляющий МК должен быть соединен с ПК и имеется USB аудио интерфейс PCM2702 – ставим хаб и подключаем все это к ПК одним проводом; проектируется домашний измерительный комплекс, компоненты которого (5 или 6 устройств) также будут соединяться с ПК – хаб будет очень кстати)
Файлы:
Плата в формате SL5
Вопросы, как обычно, складываем тут.
|
Как вам эта статья? |
Заработало ли это устройство у вас? |
Введение в мир полупроводников | AMD
Полупроводники: мозг современной электроники
Щелкая мышкой, проводя пальцем по экрану, печатая на клавиатуре или говоря в динамик электронного устройства, мы ожидаем мгновенной реакции на свои команды. Но что осуществляет поиск, расчеты, оптимизацию и представление требуемых результатов? В большинстве случаев это полупроводники.
Под «полупроводниками» подразумевают критически важные компоненты миллионов электронных устройств, используемых в сферах образования, здравоохранения, транспорта, энергетики, связи, в научных исследованиях и многих других отраслях. Современные персональные компьютеры, смартфоны, автомобили, серверы для центров обработки данных, игровые приставки — во всех этих устройствах полупроводниковые компоненты отвечают как за основные, так и за расширенные функции. Например, когда мы бронируем отель, ищем хороший ресторан, ведем потоковую трансляцию или читаем электронную почту на своем компьютере, построенные из полупроводниковых элементов центральный и графический процессоры компьютера осуществляют вычислительные функции, которые мгновенно преобразуют вопросы в ответы.
Что же такое полупроводники?
Микросхемы (или интегральные схемы), изготовленные из полупроводникового материала (например, кремния), являются важнейшей частью современных электронных устройств, предназначенных для личного и коммерческого применения. Чтобы иметь возможность осуществлять базовые логические вычисления в компьютере, эти микросхемы (так называемые транзисторы) должны быть способны действовать подобно выключателям с электронным управлением. Чтобы добиться практически мгновенного переключения, эти схемы должны быть изготовлены из полупроводникового материала, значение электрического сопротивления которого находится между проводником и изолятором.
Изготавливают полупроводниковые устройства в несколько этапов на заводах-изготовителях полупроводниковых элементов. Отраслевая практика показывает, что на проектирование, разработку, изготовление, выпуск и обслуживание одного семейства полупроводниковой продукции уходит несколько лет. Типичная компания-производитель полупроводниковых элементов должна одновременно работать над несколькими семействами продуктов, причем на разных этапах их жизненного цикла. Когда продукт выпущен в продажу, некоторые контракты предусматривают продолжительность его поставки до десяти лет.
Ингибирующие микросхемы для нисходящей пластичности сенсорных репрезентаций
Goltstein, PM, Coffey, EBJ, Roelfsema, PR & Pennartz, CMA Двухфотонная визуализация Ca 2+ in vivo выявляет избирательные эффекты вознаграждения на специфические стимулы сборки в зрительной коре головного мозга мыши. J. Neurosci. 33 , 11540–11555 (2013).
CAS PubMed PubMed Central Статья Google ученый
Хан, А. Г. и др. Отчетливые изменения в селективности стимулов и взаимодействия классов ГАМКергических интернейронов в зрительной коре, вызванные обучением. Нат. Neurosci. 21 , 851–859 (2018).
CAS PubMed Статья PubMed Central Google ученый
Poort, J. et al. Обучение усиливает сенсорные и множественные несенсорные представления в первичной зрительной коре. Нейрон 86 , 1478–1490 (2015).
CAS PubMed PubMed Central Статья Google ученый
Letzkus, J. J. et al. Растормаживающая микросхема для обучения ассоциативному страху в слуховой коре. Природа 480 , 331–335 (2011).
ADS CAS PubMed Статья PubMed Central Google ученый
Чубыкин А.А., Плотва Е.Б., Медведь М.Ф. и Шулер, М. Г. Х. Холинергический механизм для определения времени вознаграждения в первичной зрительной коре. Нейрон 77 , 723–735 (2013).
CAS PubMed PubMed Central Статья Google ученый
Fu, Y. et al. Корковый контур для управления усилением поведенческим состоянием. Cell 156 , 1139–1152 (2014).
CAS PubMed PubMed Central Статья Google ученый
Петро, Л., Визиоли, Л. и Макли, Л. Вклад корковой обратной связи в сенсорную обработку в первичной зрительной коре. Фронт. Psychol. 5 , 1223 (2014).
PubMed PubMed Central Статья Google ученый
Zhang, S. et al. Дальнодействующие и локальные схемы для модуляции обработки зрительной коры сверху вниз. Наука 345 , 660–665 (2014).
ADS CAS PubMed PubMed Central Статья Google ученый
Пи, Х.-Дж. и другие. Корковые интернейроны, специализирующиеся на растормаживающем контроле. Природа 503 , 521–524 (2013).
ADS CAS PubMed PubMed Central Статья Google ученый
Ли, С., Кругликов, И., Хуанг, З. Дж., Фишелл, Г. и Руди, Б. Растормаживающая цепь опосредует моторную интеграцию в соматосенсорной коре. Нат. Neurosci. 16 , 1662–1670 (2013).
CAS PubMed PubMed Central Статья Google ученый
Аталлах, Б. В., Брунс, В., Карандини, М. и Сканциани, М. Интернейроны, экспрессирующие парвальбумин, линейно трансформируют корковые ответы на зрительные стимулы. Нейрон 73 , 159–170 (2012).
CAS PubMed PubMed Central Статья Google ученый
Vogels, T. et al. Тормозная синаптическая пластичность: временная зависимость спайков и предполагаемая сетевая функция. Фронт. Нейронные схемы 7 , 119 (2013).
CAS PubMed PubMed Central Статья Google ученый
Адлер А., Чжао Р., Шин М. Э., Ясуда Р. и Ган В.-Б. Интернейроны, экспрессирующие соматостатин, обеспечивают и поддерживают зависимую от обучения последовательную активацию пирамидных нейронов. Нейрон 102 , 202–216 (2019). e7.
CAS PubMed Статья PubMed Central Google ученый
Лукас, Э. К. и Клем, Р. Л. ГАМКергические интернейроны: оркестр или дирижер в изучении страха и памяти? Brain Res. Бык. 141 , 13–19 (2018).
CAS PubMed Статья PubMed Central Google ученый
Пакан, Дж. М., Франсиони, В. и Рошфор, Н. Л. Действие и обучение формируют активность нейронных цепей в зрительной коре. Curr. Opin. Neurobiol. 52 , 88–97 (2018).
CAS PubMed PubMed Central Статья Google ученый
Хан, А. Г. и Хофер, С. Б. Контекстные сигналы в зрительной коре. Curr. Opin. Neurobiol. 52 , 131–138 (2018).
CAS PubMed Статья PubMed Central Google ученый
Cossell, L. et al. Функциональная организация возбуждающей синаптической силы в первичной зрительной коре. Природа 518 , 399–403 (2015).
ADS CAS PubMed PubMed Central Статья Google ученый
Коттам, Дж. К. Х., Смит, С. Л. и Хойссер, М. Целевые эффекты интернейронов, экспрессирующих соматостатин, на неокортикальную визуальную обработку. J. Neurosci. 33 , 19567–19578 (2013).
CAS PubMed PubMed Central Статья Google ученый
млн лет назад, W.-p. и другие. Визуальные репрезентации кортикальных нейронов, ингибирующих соматостатин, избирательны, но со слабыми и отсроченными ответами. J. Neurosci. 30 , 14371–14379 (2010).
CAS PubMed PubMed Central Статья Google ученый
Hofer, S.B. et al. Дифференциальная связность и динамика ответа возбуждающих и тормозных нейронов зрительной коры. Нат. Neurosci. 14 , 1045 EP (2011).
Артикул CAS Google ученый
Пфеффер, К. К., Сюэ, М., Хе, М., Хуанг, З. Дж. И Сканциани, М. Ингибирование торможения в зрительной коре: логика связей между молекулярно различными интернейронами. Нат. Neurosci. 16 , 1068–1076 (2013).
CAS PubMed PubMed Central Статья Google ученый
Jiang, X. et al. Принципы связи между морфологически определенными типами клеток в неокортексе взрослого человека. Наука 350 , aac9462 (2015).
PubMed PubMed Central Статья CAS Google ученый
Пала, А. и Петерсен, К. С. Измерение in vivo синаптических связей и синаптической передачи, специфичных для клеточного типа, в бочкообразной коре 2/3 мышей. Нейрон 85 , 68–75 (2015).
CAS PubMed PubMed Central Статья Google ученый
Jouhanneau, J.-S., Kremkow, J. & Poulet, J. F. A. Одиночные синаптические входы управляют высокоточными потенциалами действия в парвальбумине, экспрессирующем ГАМКергические кортикальные нейроны in vivo. Нат. Commun. 9 , 1540 (2018).
ADS PubMed PubMed Central Статья CAS Google ученый
Керлин, А.М., Андерманн, М.Л., Березовский, В.К. и Рейд, Р.С. Широко настроенные свойства реакции различных подтипов тормозных нейронов в зрительной коре головного мозга мышей. Нейрон 67 , 858–871 (2010).
CAS PubMed PubMed Central Статья Google ученый
Galarreta, M. & Hestrin, S. Сеть быстрорастущих клеток в неокортексе, соединенных электрическими синапсами. Nature 402 , 72–75 (1999).
ADS CAS PubMed Статья PubMed Central Google ученый
Clopath, C., Büsing, L., Vasilaki, E. & Gerstner, W. Связность отражает кодирование: модель основанного на напряжении STDP с гомеостазом. Нат. Neurosci. 13 , 344 (2010). Статья.
CAS PubMed Статья PubMed Central Google ученый
Ko, H. et al. Возникновение функциональных микросхем в зрительной коре. Природа 496 , 96 (2013).
ADS CAS PubMed PubMed Central Статья Google ученый
Guan, W. et al. Открытие глаза по-разному модулирует тормозную синаптическую передачу в развивающейся зрительной коре. eLife 6 , e32337 (2017).
PubMed PubMed Central Статья Google ученый
Niell, C. M. & Stryker, M. P. Высокоселективные рецептивные поля в зрительной коре головного мозга мышей. J. Neurosci. 28 , 7520–7536 (2008).
CAS PubMed PubMed Central Статья Google ученый
Liu, B.-h. и другие. Структура зрительного рецептивного поля кортикальных тормозных нейронов, выявленная с помощью записи под контролем двухфотонной визуализации. J. Neurosci. 29 , 10520–10532 (2009).
CAS PubMed PubMed Central Статья Google ученый
Шулер М. и Медведь М. Ф. Время вознаграждения в первичной зрительной коре. Наука 311 , 1606–1609 (2006).
ADS CAS PubMed Статья PubMed Central Google ученый
Килгард, М. П. и Мерзенич, М. М. Реорганизация кортикальной карты возможна благодаря активности базального ядра. Наука 279 , 1714–1718 (1998).
ADS CAS PubMed Статья PubMed Central Google ученый
Батт, А. Э. и Ходж, Г. К. Приобретение, сохранение и исчезновение оперантных различений у крыс с поражениями ядра Basalis magnocellularis. Behav. Neurosci 109 , 699–713 (1995).
Metherato, R. & Weinberger, N. M.Ацетилхолин вызывает специфические для стимула изменения рецептивного поля в слуховой коре головного мозга кошки. Brain Res. 480 , 372–377 (1989).
Артикул Google ученый
Метерат, Р. и Эш, Дж. Х. Базальная стимуляция переднего мозга изменяет чувствительность слуховой коры за счет воздействия на мускариновые рецепторы. Brain Res. 559 , 163–167 (1991).
CAS PubMed Статья PubMed Central Google ученый
Медведь М. Ф. и Сингер В. Модуляция зрительной корковой пластичности ацетилхолином и норадреналином. Природа 320 , 172 (1986).
ADS CAS PubMed Статья PubMed Central Google ученый
Muñoz, W., Tremblay, R., Levenstein, D. & Rudy, B. Слоистая модуляция неокортикального дендритного торможения во время активного бодрствования. Наука 355 , 954–959 (2017).
ADS PubMed Статья CAS PubMed Central Google ученый
Yaeger, C.E., Ringach, D. L. & Trachtenberg, J. T. Нейромодуляторный контроль локализованного дендритного шипа в коре головного мозга в критический период. Природа 567 , 100–104 (2019).
ADS CAS PubMed PubMed Central Статья Google ученый
Prönneke, A. et al. Характеристика VIP-нейронов в стволовой коре головного мозга мышей VIPcre / Tdtomato выявляет межслойные различия. Cereb. Cortex 25 , 4854–4868 (2015).
PubMed PubMed Central Статья Google ученый
Williams, L.E. & Holtmaat, A. Таламокортикальные входы более высокого порядка закрывают синаптическую долгосрочную потенциацию через растормаживание. Neuron 101 , 91–102 (2019).
CAS PubMed Статья PubMed Central Google ученый
Sohya, K., Kameyama, K., Yanagawa, Y., Obata, K. & Tsumoto, T. ГАМКергические нейроны менее избирательны к ориентации стимулов, чем возбуждающие нейроны в слое II / III зрительной коры. как было выявлено с помощью функциональной визуализации Ca 2+ in vivo у трансгенных мышей. J. Neurosci. 27 , 2145–2149 (2007).
CAS PubMed PubMed Central Статья Google ученый
Runyan, C.A. et al. Характеристики ответа интернейронов, экспрессирующих парвальбумин, предполагают точную роль подтипов торможения в зрительной коре. Нейрон 67 , 847–857 (2010).
CAS PubMed PubMed Central Статья Google ученый
Runyan, C. A. & Sur, M. Избирательность ответа коррелирует с дендритной структурой в нейронах-ингибиторах, экспрессирующих парвальбумин, в зрительной коре головного мозга. J. Neurosci. 33 , 11724–11733 (2013).
CAS PubMed PubMed Central Статья Google ученый
Li, L.-Y. и другие. Дифференциальные свойства рецептивного поля парвальбумина и нейронов, ингибирующих соматостатин, в слуховой коре мышей. Cereb. Cortex 25 , 1782–1791 (2015).
PubMed Статья PubMed Central Google ученый
Goltstein, P.M., Meijer, G.T. и Pennartz, C.M. Кондиционирование обостряет пространственное представление стимулируемых стимулов в первичной зрительной коре головного мозга мыши. eLife 7 , e37683 (2018).
PubMed PubMed Central Статья Google ученый
Макино, Х. и Комияма, Т. Обучение усиливает относительное влияние нисходящей обработки в зрительной коре головного мозга. Нат. Neurosci. 18 , 1116–1122 (2015).
CAS PubMed PubMed Central Статья Google ученый
Одетт, Н. Дж., Бернхард, С. М., Рэй, А., Стюарт, Л. Т. и Барт, А. Л. Быстрая пластичность таламокортикальных входов высшего порядка во время сенсорного обучения. Нейрон 103 , 277–291 (2019).
CAS PubMed Статья PubMed Central Google ученый
Бещад, К. М. и Вайнбергер, Н. М. Исчезновение показывает, что первичная сенсорная кора предсказывает результат подкрепления. евро. J. Neurosci. 35 , 598–613 (2012).
PubMed PubMed Central Статья Google ученый
Grewe, B. F. et al. Динамика нейронного ансамбля, лежащая в основе долговременной ассоциативной памяти. Nature 543 , 670 EP (2017).
ADS Статья CAS Google ученый
Бутон М. Э. Контекст и поведенческие процессы на грани исчезновения. Учиться. Mem. 11 , 485–494 (2004).
PubMed Статья PubMed Central Google ученый
Бэррон, Х. К., Фогельс, Т. П., Беренс, Т. Э. и Рамасвами, М. Тормозящие инграммы в восприятии и памяти. Proc. Natl Acad. Sci. США 114 , 6666–6674 (2017).
CAS PubMed PubMed Central Google ученый
Mongillo, G., Rumpel, S. & Loewenstein, Y. Тормозная связность определяет область возбуждающей пластичности. Нат. Neurosci. 21 , 1463–1470 (2018).
CAS PubMed Статья PubMed Central Google ученый
Чумаченко Т. и Клопат К. Колебания, возникающие из управляемого шумом устойчивого состояния в сетях с электрическими синапсами и подпороговым резонансом. Нат.Commun. 5 , 5512 EP (2014).
ADS Статья Google ученый
Би, Г. и Пу, М. Синаптические модификации в культивируемых нейронах гиппокампа: зависимость от времени спайков, синаптической силы и типа постсинаптических клеток. J. Neurosci. 18 , 10464–10472 (1998).
CAS PubMed PubMed Central Статья Google ученый
Маркрам, Х., Любке, Дж., Фротшер, М. и Сакманн, Б. Регулирование синаптической эффективности путем совпадения постсинаптических ПД и ВПСП. Наука 275 , 213–215 (1997).
CAS PubMed Статья PubMed Central Google ученый
Герстнер В., Кемптер Р., ван Хеммен Дж. Л. и Вагнер Х. Правило обучения нейронов для субмиллисекундного временного кодирования. Природа 383 , 76 (1996).
ADS CAS PubMed Статья PubMed Central Google ученый
Стимберг, М., Гудман, Д. Ф., Бенишу, В. и Бретт, Р. Брайан 2 — второе пришествие: усиление моделирования нейронных сетей на Python с генерацией кода. BMC Neurosci. 14 , P38 – P38 (2013).
PubMed Central Статья Google ученый
соединение, данные, деталь, устройство, цифровое, аппаратное обеспечение, информация, интегрированное, мегабайты, микросхема, микросхема
соединение, данные, деталь, устройство, цифровое, аппаратное обеспечение, информация, интегрированное, мегабайты, микросхема, микросхема | Пикист соединение, данные, деталь, устройство, цифровой, аппаратное обеспечение, информация, интегрированный, мегабайты, микрочип, микросхемаПохожие фотографии
- материнская плата, электроника, компьютер, плата, компоненты, чипы, технология, основная плата, цифровой, схема, оборудование Всеобщее достояние
- доска, Электроника, компьютер, Электротехника, ток, печатная плата, Данные, ЦП, Цепи, Чип, технологии Всеобщее достояние
- материнская плата, электрические, технологии, чип, компьютер, плата, цепь, электрические, аппаратное обеспечение, процессор, технология Всеобщее достояние
- Электроника, компоненты, микросхемы, компьютер, плата, технология, основная плата, материнская плата, цифровой, схема, оборудование Всеобщее достояние
- фон, зеленый, доска, бизнес, микросхема, цепь, крупный план, связь, компонент, компьютер, соединить Всеобщее достояние
- электроника, интегральная схема, технология, чип, компьютер, процессор, микрочип, компонент, полупроводник, макрос Всеобщее достояние
- микрочип, компьютер, схема, технология, плата, процессор, цифровой, микрочип, аппаратное обеспечение, электроника, компонент Всеобщее достояние
- ЦП, процессор, макрос, ручка, штифт, компьютер, электроника, обработка данных, чип, плата, схемы Всеобщее достояние
- ЦП, процессор, электроника, компьютер, обработка данных, плата, он, микросхема, центр управления, схемы, линия Всеобщее достояние
- доска, компьютер, цепь, фон, технология, материнская плата, цифровой, электронный, чип, текстура, информация Всеобщее достояние
- плата, печатная плата, компьютер, Электроника, Пайка, технологии, Цепи, Отследить, Электротехника, Информация, Данные Всеобщее достояние
- плата, проводники, схемы, калькулятор, компьютер, доступ, компьютерные вирусы, компьютерный вирус, центр коммутации данных, трассировка, процессор Всеобщее достояние
- контакты, ЦП, процессор, макрос, ручка, компьютер, электроника, обработка данных, чип, плата, схемы Всеобщее достояние
- зеленый, доска, бизнес, Чип, Цепь, Крупный план, Связь, Компонент, компьютер, Подключить, Соединение Всеобщее достояние
- основная плата, чипы, электроника, компьютер, доска, технология, материнская плата, оборудование, схема, цифровой, электронный Всеобщее достояние
- искусственный интеллект, мозг, думать, контроль, информатика, электротехника, технология, разработчик, компьютер, человек, умный Всеобщее достояние
- интернет, кибер, сеть, палец, сенсорный экран, бизнес, бизнесмен, технологии, компьютер, цифровой, данные Всеобщее достояние
- сеть, сеть, технологии, разработчик, касание, палец, искусственный интеллект, думать, контроль, информатика, электротехника Всеобщее достояние
- технологии, разработчик, континенты, касание, палец, искусственный интеллект, думать, контроль, информатика, электротехника, компьютер Всеобщее достояние
- цепочка блоков, личное, рукопожатие, рукопожатие, данные, записи, концепция, система, коммуникация, консенсус, механизм Всеобщее достояние
- основная плата, компьютер, микросхемы, электроника, плата, технология, оборудование, материнская плата, схема, полупроводник, ПК Всеобщее достояние
- amd, ЦП, процессор, микропроцессор, аппаратное обеспечение, чип, ПК, технологии, компьютер, компонент, схема Всеобщее достояние
- он, переключатель, сеть, обработка данных, соединение, данные, соединительный кабель, LAN, интернет, технологии, электроника Всеобщее достояние
- Ethernet, данные, Интернет, сеть, компьютер, соединение, технологии, связь, информация, телекоммуникации, цифровой Всеобщее достояние
- плата, компьютер, микросхема, обработка данных, пайка, макрос, материнская плата, электроника, схемы, аппаратное обеспечение Всеобщее достояние
- материнская плата, ЦП, компьютер, процессор, технология, оборудование, схема, система, плата, ПК, печатная плата Всеобщее достояние
- печатная плата, плата, процессор, компьютер, схемы, технологии, обработка данных, микросхема, пайка, макрос, проводники Всеобщее достояние
- usb, технологии, компьютер, микрочип, материнская плата, пк, raspberry, монтажная плата, чип, it, raspberry pi Всеобщее достояние
- плата, оцифровка, схемы, центр управления, трассировка, процессор, пк, пайка, процессор, микросхема, обработка данных Всеобщее достояние
- интернет, кибер, сеть, палец, сенсорный экран, бизнес, бизнесмен, технологии, компьютер, цифровой, данные Всеобщее достояние
- доска, компьютер, материнская плата компьютера, Чип, обработка данных, Пайка, Электроника, Линии, Макрос, линия, Схемы Всеобщее достояние
- плата, компьютер, микросхема, обработка данных, пайка, макрос, материнская плата, электроника, схемы, аппаратное обеспечение Всеобщее достояние
- технологии, разработчик, касание, палец, искусственный интеллект, думать, контроль, информатика, электротехника, компьютер, интеллектуальный Всеобщее достояние
- Ethernet, данные, Интернет, сеть, компьютер, соединение, технологии, связь, информация, телекоммуникации, цифровой Всеобщее достояние
- кулер, компьютер, вентилятор, технологии, пк, процессор, оборудование, аппаратное обеспечение, ЦП, мощность, электроника Всеобщее достояние
- искусственный интеллект, робот, искусственный интеллект, ки, программирование, компьютер, среда, синтаксис, программа, обработка данных, реклама Всеобщее достояние
- технологии, разработчик, касание, палец, искусственный интеллект, думать, контроль, информатика, электротехника, компьютер, интеллектуальный Всеобщее достояние
- интегральная схема, устройство, чип, технологии, электронные, компьютер, аппаратное обеспечение, компонент, инженерия Всеобщее достояние
- сеть, соединение, компьютер, веб-адрес, Интернет, компьютер, Интернет, http, www, электронная коммерция, маршрутизатор Всеобщее достояние
- сервер, монтажная плата, электроника, цепь, компьютер, плата, материнская плата, система, аппаратное обеспечение, микросхема, полупроводник Всеобщее достояние
- аннотация, фон, доска, карта, микросхема, цепь, связь, компьютер, проводник, деталь, цифровой Всеобщее достояние
- оптоволоконный, кабель, синий, сеть, технологии, связь, скорость, волокно, данные, интернет, соединение Всеобщее достояние
- плата, печатная плата, компьютер, Электроника, Пайка, технологии, Цепи, Отследить, Электротехника, Информация, Данные Всеобщее достояние
- компьютер, аппаратное обеспечение, процессор, технология, ЦП, материнская плата, вентиляторы, инструмент, БД, компонент, электрические Всеобщее достояние
- материнская плата, графический процессор, графика, обработка, блок, компьютер, технология, процессор, аппаратное обеспечение, микрочип, данные Всеобщее достояние
- доска, карта, чип, компьютер, данные, цифровой, электрические, электронные, зеленый, аппаратное обеспечение, макрос Всеобщее достояние
- жесткий диск, детали, считывающая головка, внутреннее устройство, макрос, микросхема памяти, вырез, оборудование, магнитные головки, стилус, хранилище данных Всеобщее достояние
- плата, компьютер, микросхема, обработка данных, пайка, макрос, материнская плата, электроника, схемы, аппаратное обеспечение Всеобщее достояние
- социальные сети, facebook, twitter, instagram, значки, маркетинг, linkedin, бизнес, мобильный, pinterest, экран Всеобщее достояние
- материнская плата, процессор, восстановление, микро, оборудование, чип, байт, схема, технология, микрочип, компьютер Всеобщее достояние
- сетевая карта, компонент, схема, микросхема, компьютер, переходная плата, ПК, электрически, плата, обработка данных, соединения Всеобщее достояние
- биткойн, криптовалюта, валюта, деньги, рука, держать, визитная карточка, доска, трассировка, схемы, чип Всеобщее достояние
- плата, компьютер, микросхема, обработка данных, пайка, макрос, материнская плата, электроника, схемы, аппаратное обеспечение Всеобщее достояние
- главный процессор, биткойн, Intel, полупроводник, технология, компьютер, устройство отображения, сервер, плата, электроника, схема Всеобщее достояние
- цепочка блоков, рукопожатие, рукопожатие, контракт, данные, записи, концепция, система, коммуникация, консенсус, механизм Всеобщее достояние
- ЦП, процессор, макрос, ручка, штифт, компьютер, электроника, обработка данных, чип, плата, схемы Всеобщее достояние
- технологии, информация, компьютерная часть, заголовок, баннер, фон, оборудование, данные, процессор, вентилятор, температура Всеобщее достояние
- компьютер, материнская плата, материнская плата, аппаратное обеспечение, процессор, микросхема, схема, технология, микропроцессор, возможность подключения, вычисления Всеобщее достояние
- облако, палец, смартфон, телефон, набор текста, мобильный телефон, экран, смс, отправка, вызов, контакт Всеобщее достояние
- планшет, ipad, чтение, экран, смахивание, касание, обозначение, точка, гаджет, современное, интернет Всеобщее достояние
Плотная, неспецифическая связь парвальбумин-положительных интернейронов неокортекса: каноническая микросхема для ингибирования?
Введение
Неокортекс млекопитающих — это чудо биологической инженерии, способное на впечатляющие вычислительные возможности.Эта структура, по-видимому, организована стереотипным и иерархическим образом, предположительно для достижения высокого уровня параллельной вычислительной мощности (Mountcastle, 1982). К сожалению, структура корковых микросхем кажется «непроходимыми джунглями» (Ramón y Cajal, 1923) из-за плотного смешения разных типов нейронов, что затрудняет определение того, как они работают. В самом деле, хотя корковые препараты изучались анатомически и физиологически на протяжении более века, удивительно мало было обнаружено о мелкомасштабной организации этих нейронов в цепи.Ясно, что кора головного мозга делится на слои и области, каждая из которых имеет различные паттерны связи с другими слоями и областями, что, по-видимому, приводит к определенным функциям для каждого элемента (Douglas and Martin, 2004). Также известно, что отдельные нейроны принадлежат к определенным классам, хотя до сих пор нет действительной классификации для многих нейронов (Ascoli et al., 2008) или теоретической основы, описывающей, как они действуют согласованным образом для обработки информации.
Частично причиной этого незнания является экспериментальная трудность выявления того, как эти нейроны связаны друг с другом, чтобы сформировать функциональные цепи (Crick, 1979).В последнее десятилетие было разработано несколько новых методов для выявления синаптически связанных нейронов, включая парные записи целых клеток (Thomson and Lamy, 2007), серийные ЭМ-реконструкции (Denk and Horstmann, 2004; Bock et al., 2011), вирусные отслеживание (Wickersham et al., 2007), визуализация кальция (Peterlin et al., 2000), визуализация с обратной корреляцией (Aaron and Yuste, 2006) и фотостимуляция глутаматом в клетке (Callaway and Katz, 1993). Недавно мы представили вариант фотостимуляции, который использует двухфотонное снятие каркаса глутамата для картирования связанных нейронов с одноклеточным разрешением (Николенко и др., 2007). В сочетании с извлечением из каркаса RuBi-глутамата, недавно разработанного соединения глутамата в клетках (Fino et al., 2009), двухфотонная фотостимуляция может применяться для картирования ингибирующих связей. Действительно, используя RuBi-Glutamate, мы недавно картировали синаптические цепи соматостатин-положительных интернейронов в верхних слоях фронтальной коры мышей, обнаружив очень плотную и неспецифическую связь этих интернейронов с соседними пирамидными клетками (Fino and Yuste, 2011). Об этой плотной связности ранее не сообщалось, и мы задались вопросом, было ли это особенностью соматостатин-положительных интернейронов или лобной коры.
Чтобы ответить на этот вопрос, мы составили карту взаимосвязи от парвальбумин-положительных (PV +) корзиночных клеток — самой большой популяции интернейронов в неокортексе — к пирамидным клеткам (ПК) в различных слоях и областях коры. Мы обнаружили локально плотные и неспецифические связи от интернейронов PV + к ПК в двух разных областях коры (соматосенсорный и лобный), а также в двух разных слоях (2/3 и 5). Хотя мы использовали срезы мозга, в которых разорвано множество соединений, мы часто обнаруживали, что каждая локальная ячейка PV + была подключена к каждому компьютеру, на котором проводилась выборка.Наши результаты подтверждают точку зрения, что плотная архитектура межнейронных связей, посредством которой «одеяло ингибирования» распространяется на локальные пирамидные клетки, является универсальной или канонической особенностью в дизайне неокортикальных микросхем.
Результаты
Двухфотонная фотостимуляция интернейронов парвальбумина
Чтобы изучить возможность соединения четко определенного подмножества интернейронов с соседними ПК, мы использовали линию трансгенных мышей G42, в которых интернейроны, экспрессирующие парвальбумин, помечены GFP (Chattopadhyaya et al., 2004). В корональных срезах головного мозга мы нацелены и записали 91 GFP-положительную клетку (69 в слое соматосенсорной коры 2/3, 15 в слое соматосенсорной коры 5 и 7 в слое 2/3 лобной коры) и выполнили анатомические и электрофизиологические характеристики эти нейроны. Анатомически все нацеленные интернейроны PV + имели особенности корзиночных клеток с беспорядочными и плотно ветвящимися аксонами ( n = 15 реконструкций; Рис. 1 A ). Электрофизиологически все GFP-положительные нейроны представляли собой клетки с быстрым выбросом импульсов, возбуждаемые с частотой> 50 Гц (97 ± 6 Гц, n = 42) (Ascoli et al., 2008). У них также были высокие реобазы (312 ± 102 пА, n = 42), сильные токи быстрой постгиперполяризации (fAHP) и выпрямляющие кривые ВАХ (рис.1 B , C ) (Woodruff et al., 2009 г.).
Рисунок 1.Характеристика и фотостимуляция парвальбумин-положительных корзиночных клеток с быстрым выбросом. A , Реконструкция GFP-положительного интернейрона в слое 2/3 соматосенсорной коры из линии трансгенных мышей G42 (аксоны синего цвета, дендриты красного цвета). B , Электрофизиологические записи выявили классические высокие значения реобазы и тока fAHP (вверху). Быстрый импульсный ответ был вызван после введения в интернейрон током 1 нА (внизу). C , Этот подтип интернейрона характеризуется выпрямляющей кривой IV (Woodruff et al., 2009). D , E , Двухфотонный лазерный луч мультиплексировался во времени путем последовательного стимулирования нескольких целей вокруг тела клетки ( D ) и в пространстве за счет включения дифракционного оптического элемента для разделения лазерный луч на пять бимлетов ( E ), чтобы освободить RuBi-глутамат вокруг тела клетки (лазерные пятна нарисованы в реальном размере, см. H – I ). F . Одинарные потенциалы действия были вызваны путем освещения образца 150 мВт светом 800 нм с использованием объектива с числовой апертурой 20 × 0,5 (черная полоса показывает продолжительность освещения). G , При более высокой мощности (300 мВт) множественные потенциалы действия были вызваны с характерной формой волны с быстрыми всплесками. H , Функция рассеяния точки дифракционных пучков оптических элементов была измерена с помощью объектива с числовой апертурой 20 × 0,5, который мы использовали для картирования, и 0.Флуоресцентные шарики 17 мкм. Поперечный размер луча каждого дифракционного оптического элемента составляет ~ 1 мкм (вставка, необработанное изображение). I , Осевое разрешение каждого бимлета составляет 6 мкм. J , Прямой тест пространственного разрешения двухфотонной фотостимуляции. Расположение 180 мишеней стимуляции, которые были стимулированы во время записи ПК (в центре, черный), обозначены черными кружками. Местоположение 1. Мишень, обозначенная синим цветом, непосредственно на соме ПК, была непосредственно фотостимулирована для создания 10-кратных всплесков потенциалов действия (один типичный пример показан справа вверху).Местоположение 2, эта цель (верхний красный кружок), которая была лишь немного смещена от центра исправленного ПК, вызвала срабатывание ячейки только один раз (справа, в центре). Местоположение 3, Фотостимуляция этой цели (нижний красный кружок) также заставляла исправленный ПК запускать одиночные потенциалы действия (справа, внизу).
Чтобы отобразить синаптические связи от интернейронов PV + на постсинаптические ПК, мы следовали протоколу, первоначально разработанному Николенко и соавт. (2007) и недавно уточнены для ингибирующих связей Фино и Юсте (2011).Этот протокол основан на способности оптически стимулировать отдельные нейроны с помощью двухфотонного освобождения RuBi-глутамата (Fino et al., 2009). Мы достигли одноклеточного разрешения фотоактивации путем мультиплексирования двухфотонного лазерного луча во времени, фокусируя его в восьми точках вокруг сомы клетки-мишени (Рис. 1 D – G ) и в пространстве с помощью дифракционного оптического элемента (Николенко et al., 2007), чтобы разделить лазерный луч на пять бимлетов (рис. 1 E ). Эта стратегия сохранила целостность функции рассеяния точки двухфотонного пучка (рис.1 H , I ), что обеспечивает точность до одной ячейки. При фотостимуляции записанного интернейрона PV + с помощью этого метода, от одного до нескольких потенциалов действия (AP) выявлялись при промежуточных мощностях лазера (150 мВт на образце; рис. 1 F ), в то время как всплески AP вызывались при большей мощности (300 мВт на образец; рис.1 G ). Несмотря на высокую реобазу этих клеток, эти мощности лазера (> 150 мВт) никогда не перестали производить AP (AP в 21 из 21 протестированной клетки). ТД, индуцированные освобождением каркаса, отображали форму волны fAHP и высокочастотные последовательности, характерные для клеток с быстрым выбросом (рис.1 G ). Кроме того, когда стимуляция интернейрона PV + повторялась каждые 5 с, каждый раз возникали идентичные ответы (задержка 14,22 ± 0,08 мс до первого всплеска, 11,6 ± 0,2 потенциала действия на стимуляцию, n = 10 фотостимуляций). Таким образом, использованная нами стратегия фотостимуляции была точной и надежной, активируя нейроны с одноклеточным разрешением.
Чтобы проверить, не активирует ли наша фотостимуляция непреднамеренно близлежащие нейроны, мы пропатчили ПК с низкой реобазой (40 пА) и фотостимулировали всю область, окружающую клетку, в виде сетки, используя те же параметры, что и для стимуляции интернейронов.Во-первых, мы подтвердили, что этот ПК может быть фотостимулирован путем возбуждения потенциалов действия посредством извлечения клеток из клетки непосредственно над сомой 10 из 10 раз (рис. 1 J , верхний правый график). Затем мы фотостимулировали с той же мощностью лазера сетку из 180 точек в поле зрения (рис. 1 J ) по три раза каждая и обнаружили, что ПК регистрировал только произведенные потенциалы действия, когда стимулировались два места: одно только немного смещен относительно сомы (рис.1 J , позиция 2, красный кружок) и один на дне сомы (рис.1 J , позиция 3, красный кружок). Мы пришли к выводу, что очень редко возникают потенциалы действия, если они не стимулируют клеточную сому напрямую. Это делает маловероятным то, что нецелевые нейроны могут быть приведены к созданию потенциалов действия посредством непреднамеренной фотостимуляции, особенно для интернейронов с более высокими реобазами, что еще раз подтверждает высокое разрешение этой техники.
Картирование моносинаптических связей от интернейронов PV + к пирамидным клеткам
Затем мы сопоставили входы от интернейронов на ПК, следуя стратегии, разработанной Fino и Yuste (2011).Вкратце, ПК накладывали, в то время как соседние интернейроны PV + последовательно фотостимулировали (фиг. 2 A , синяя стрелка). Каждый интернейрон PV + был активирован диапазоном не менее пяти лазерных мощностей, включая максимум 300 мВт на образце. Постсинаптические ответы в ПК, которые были привязаны по времени к стимулу, измерялись с помощью фиксации напряжения (рис. 2 B , ячейки 1–5). Первоначально ПК выдерживали при +40 мВ, чтобы увеличить движущую силу тормозных токов, а затем при -40 мВ, чтобы проверить, действительно ли эти постсинаптические токи были тормозящими (рис.2 B , нижний след).
Рисунок 2.Картирование ингибирующих входов пирамидных клеток. A , Отображение потенциальных связей от пяти интернейронов (пронумерованных от 1 до 5) на один ПК (синяя стрелка) в соматосенсорном слое 2/3. Серые круги, несвязанные интернейроны; красные круги, связанные. B , Электрофизиологические записи, полученные с ПК при удерживающих потенциалах +40 мВ (вверху) и -40 мВ (внизу) во время фотостимуляции интернейронов, показанных в A .Обратите внимание на синаптический отклик от одного интернейрона ( A , красная стрелка; большее увеличение в D ) в то время как ближайший интернейрон ( A , серая стрелка; большее увеличение в F ) не показал ответа. C , E , Красный нейрон интернейрона PV + был исправлен и подтвержден как синаптически связанный, в то время как не было зарегистрировано никакого ответа для серого нейрона ( F ), который был определен как не связанный с помощью фотостимуляции . G , Истинно положительный ответ. ПК, вставленный в поле зрения, показывает множество GFP-положительных интернейронов PV + поблизости, один из которых (красный кружок), как было установлено, предположительно связан с помощью оптической стимуляции. H . Удерживая ПК при +40 мВ (вверху) или -40 мВ (внизу), мы фотостимулировали клетку, обведенную кружком G , вызывая IPSC. Потенциал реверсирования для ГАМК был установлен на уровне -80 мВ во внутреннем растворе, чтобы EPSC, но не IPSC, меняли направление в диапазоне от +40 до -40 мВ. I , J , Фотостимулированная клетка была исправлена, что электрофизиологически подтвердило, что она действительно была синаптически связана. K , Отрицательные отзывы. Интернейрон PV + (серый круг) был нацелен на фотостимуляцию во время записи целых клеток двух ближайших ПК. L , Записи с ПК в K не показывают свидетельств синаптической связи во время фотостимуляции интернейрона PV +. M , N , Фотостимулированная клетка была исправлена, что электрофизиологически подтвердило отсутствие синаптических связей от интернейрона PV + на любом ПК. O , Ложноположительный ответ. PV + интернейрон (черный кружок) непосредственно рядом с записанным ПК был нацелен на фотостимуляцию. P , Медленный ответ, который был направлен наружу при +40 мВ, но перевернулся при -40 мВ, отличался от синаптического события, показанного в H , как прямая стимуляция патченного нейрона. Q , интернейрон PV + (черный кружок), расположенный дистальнее записанного ПК (синяя стрелка), был нацелен для фотостимуляции. R , Возбуждающая клетка, подключенная к записанному ПК, была стимулирована, что привело к ложноположительному результату, определяемому наличием EPSC.
Если фотостимуляция PV + клетки вызвала синаптический ответ в ПК с ГАМКергическим обратным потенциалом (установленным на -80 мВ), мы идентифицировали их как истинно положительные, то есть предположительно связанный нейрон, поскольку он был привязан ко времени оптическая стимуляция тормозной клетки (рис.2 B , красный ящик). Чтобы проверить, действительно ли интернейроны, генерирующие эти истинно положительные ответы, были связаны с постсинаптической ПК-клеткой, мы закрыли их вторым электродом (рис. 2 A – D , ячейка 4, красные стрелки), стимулировали их внутриклеточными импульсами тока до производят потенциалы действия и подтверждают генерацию IPSC (рис. 2 E ). В целом, мы обнаружили, что в девяти из 10 случаев истинно положительные PV + клетки генерировали IPSC с короткой задержкой в ПК, что указывает на моносинаптическую передачу (1.12 ± 0,19 мс, n = 9 пар; Рис. 2 G – J ). Эти пресинаптические PV + интернейроны всегда запускали AP при фотостимуляции (Fig. 1 G ) и генерировали тормозные ответы с большими выходными токами (Fig. 2 D , H ).
Чтобы исследовать, могут ли интернейроны, которые не генерируют ответ в ПК, быть потенциально связаны с ним, мы также исправили восемь фотостимулированных интернейронов PV +, для которых не наблюдалось ответа (отрицательные).Мы подтвердили, что ни в одном из этих случаев не было синаптической связи (рис. 2 K – N ), заключив, что негативы были истинно негативными. Другими словами, мы никогда не наблюдали ложноотрицательных результатов, в которых интернейрон PV +, который, казалось, не был связан оптически, был фактически подключен. Даже интернейроны PV +, которые были очень близки друг к другу, можно было устранить неоднозначность, о чем свидетельствует полное отсутствие ответа (рис.2 B , ячейка 5; F ) от клеток, непосредственно прилегающих к истинно положительно связанным клеткам (рис.2 A , серая стрелка против красной стрелки).
В дополнение к истинно положительным и отрицательным ответам мы иногда наблюдали неожиданные постсинаптические токи, которые меняли направление при потенциале реверсии глутамата 0 мВ (ложноположительные ответы). Большинство этих ложных срабатываний происходило при непосредственной стимуляции вблизи зарегистрированного ПК, сразу за его сомой (рис. 2 O ). В дополнение к другому потенциалу обращения эти ответы имели более медленную кинетику (рис.2 P ), чем истинные положительные ответы от связанных PV + интернейронов (рис. 2 H ). Мы также наблюдали случайные ложноположительные ответы, которые возникали из стимулирующих участков далеко от дендритного дерева зарегистрированного нейрона (рис. 2 Q , черный кружок). Мы предполагаем, что эти возбуждающие реакции (рис.2 R ) вызваны ПК, расположенными поблизости от целевого интернейрона PV +, которые были случайно стимулированы, что согласуется с нашей стимуляцией управления сеткой (рис.1 Дж ). Эти два типа ложноположительных ответов были редкими, и их легко отличить от истинно положительных ответов из-за их способности к глутаматергической реверсии.
Таким образом, мы классифицировали все стимулированные интернейроны на три типа на основе их ответов в клетке ПК: (1) истинно положительные клетки, подключенные к ПК с помощью моносинаптических IPSC; (2) истинно отрицательные ячейки, не подключенные; и (3) ложноположительные клетки, которые генерировали контаминирующие EPSC. В оставшейся части исследования мы использовали эту классификацию для анализа карт входной связности.
Плотная локальная связь интернейронов PV + с пирамидными клетками в разных слоях и областях коры
Мы собрали в общей сложности 82 входных карты из слоев 2/3 ( n = 38) и 5 ( n = 21) соматосенсорной коры и слоя 2/3 лобной коры ( n = 23), возникающих из записей с 13 одиночных ПК, 12 пар ПК, 11 троек ПК и трех квадруплетов ПК. В оптимальных случаях мы могли отображать входы на четыре ПК в разных фокальных плоскостях (рис.3) Например, в показанном эксперименте мы протестировали возможность подключения 22 интернейронов PV +, оценив состояние подключения в общей сложности 88 пар в этом единственном срезе. Этот подход позволял проводить крупномасштабные измерения связности в пределах одного среза, тогда как предыдущая двойная запись целых ячеек, которая поддерживала точность по одной ячейке, требовала сравнений по множеству разных срезов.
Рисунок 3.Одновременное отображение входов PV + на четыре ПК. Были зашиты четыре ПК в слое 2/3 соматосенсорной коры и оптически стимулировались окружающие ЛВ + интернейроны в четырех разных фокальных плоскостях.Каждый столбец показывает результат сопоставления меченных GFP интернейронов PV + для каждого из четырех ПК. Каждая строка показывает результат для другой фокальной плоскости с проекциями всех фокальных плоскостей в верхнем ряду. Связанные интернейроны PV + обведены красным, несвязанные интернейроны PV + обведены серым, а интернейроны PV +, на которых произошла прямая стимуляция исправленного ПК, обведены черным.
Используя эту трехмерную стратегию с одновременной записью целых клеток с нескольких ПК, мы смогли опробовать связь между парами ПК 2002 PV + интернейрон.В среднем 24 ± 1 интернейрон PV + были протестированы для каждой карты из 5 ± 0,2 фокальных плоскостей, и мы обнаружили связи на всех исследованных глубинах. Корреляция между вероятностью соединения и глубиной исправленного ПК была значительной, но небольшой ( R 2 = 0,08, p <0,05 ANOVA, n = 82 карты, данные не показаны).
Мы обнаружили, что ПК получали соединения от большинства локальных интернейронов PV +, но не от более удаленных (рис. 3, 4 A ).В целом мы протестировали в общей сложности 21 ± 1, 29 ± 3 и 25 ± 2 интернейронов PV + на ПК в S2 / 3, S5 и F2 / 3 соответственно ( p > 0,05, Краскал-Уоллис), наблюдая среднее значение 3 ± 0,4, 10 ± 1,4 и 7 ± 0,8 соединений на входных картах S2 / 3, S5 и F2 / 3 соответственно. Эти различия в связях между корковыми областями и слоями были статистически значимыми (односторонний дисперсионный анализ, p <0,0001; тест множественного сравнения Тьюки, S2 / 3 против S5, p <0,001; S2 / 3 против F2 / 3, p <0.01; S5 по сравнению с F2 / 3, p <0,05; Рис.4 B ).
Рисунок 4. Входы интернейроновDense PV + на ПК. A , Типичная карта из S2 / 3, показывающая входы от интернейронов PV + к ПК (красный — подключен; серый — не подключен; черный — ложное срабатывание). B , Количество PV + интернейронов, протестированных оптически на исправленный ПК (без значительных различий), и сколько было подключено в каждой области коры и в каждом отобранном слое. C , Схема куба ткани, взятого нами на наших картах. D , Гистограмма, показывающая все межсоматические расстояния интернейронов PV + от ПК, чья связь была проверена (зеленые столбцы) в различных слоях и областях коры. Ложноположительных ответов (черные полосы) было немного. E , Гистограмма вероятности соединения в зависимости от межнейронного расстояния между интернейронами и ПК. Вероятность подключения интернейронов PV + к ПК во всех областях и слоях была очень высокой, когда интернейроны PV + находились близко к ПК, и уменьшались с расстоянием. F , Вероятность обнаружения соединения от интернейрона PV + к ПК в пределах 200 мкм в S2 / 3, S5 и F2 / 3. G , Вероятность соединения для всех оптически стимулированных интернейронов в различных областях и слоях коры. Вероятность была ниже в соматосенсорном слое 2/3, чем в соматосенсорном слое 5 или во фронтальном слое 2/3. H , Вероятность наблюдения соединения интернейрона PV + с ПК в пределах 200 мкм у старых животных (статистических различий нет). I , Вероятность соединения для всех оптически стимулированных интернейронов у старых животных (без статистических различий). n.s., не имеет значения. * p <0,05, ** p <0,01, *** p <0,001.
Чтобы проанализировать зависимость связности от межсоматического расстояния, мы определили вероятность соединения от интернейронов PV + к ПК в процентах (соединенные интернейроны, разделенные на соединенные и неподключенные интернейроны) и измерили ее как функцию интерсоматического расстояния между интернейронами и ПК.Мы собрали входные карты из прямоугольных пластин коры головного мозга размером 800 на 600 на 52 ± 3 мкм (рис. 4 C ), поэтому мы смогли рассчитать межсоматическое расстояние в трех измерениях (рис. 4 D , зеленые столбцы). Ложноположительные результаты были редкими и исключались из последующих анализов (фиг. 4 D , черные полосы).
Во всех областях вероятность соединения была чрезвычайно высокой в локальном диапазоне и обратно пропорциональна межсоматическому расстоянию между интернейроном PV + и ПК (рис.4 E ). Экспоненциальная аппроксимация вероятности соединения в зависимости от межсоматического расстояния показала пространственные константы затухания 124, 183 и 180 мкм для S2 / 3, S5 и F2 / 3 соответственно ( R 2 = 0,92, 0,91 и 0,86, соответственно).
Для межсоматических расстояний <200 мкм вероятность соединения интернейрона PV + с ПК была высокой: от 43 ± 6% в соматосенсорном слое 2/3, до 67 ± 6% в соматосенсорном слое 5 и до 76 ± 4% во фронтальном слое 2/3 (односторонний дисперсионный анализ, p <0.0001; Посттест множественных сравнений Тьюки, S2 / 3 против S5, p <0,05; S2 / 3 по сравнению с F2 / 3, p <0,001; S5 против F2 / 3, p > 0,05; n = 38, 20 и 23 карты в S2 / 3, S5 и F2 / 3 соответственно; Рис.4 F ). Фактически, на таких близких расстояниях мы обнаружили 13 (из 82) случаев полностью связанных локальных карт ( n = 5 для S2 / 3; n = 4 для S5, n = 4 для F2 / 3; ранжирование от 1 до 7 интернейронов, в среднем 3,4 ± 0,6 интернейронов, n = 13 карт).
С учетом всех расстояний средняя вероятность соединения (подключенных вне связанных и несвязанных нейронов) была ниже в S2 / 3 (18 ± 3%), чем в S5 (36 ± 4%) или F2 / 3 (33 ± 3%). ) ( p = 0,001, Краскал – Уоллис; p <0,01 для S2 / 3 против S5 и S2 / 3 против F2 / 3; p > 0,05 для S5 против F2 / 3, тест множественного сравнения Данна; Рис.4 G ). Аналогичный результат был получен при сравнении гистограмм связи в зависимости от расстояния (рис.0001, тест Фридмана). Это различие не было связано с разницей в количестве целей, протестированных в разных областях и слоях, переменные, которые существенно не различались (рис. 4 B ).
Мы также получили 12 PV + входных карт для ПК в S2 / 3 у взрослых животных ( n = 8 P22 – P25, n = 4 P31 – P45; Рис. 4 H , I ). Вероятность соединения в пределах 200 мкм была неизменно высокой во всех возрастах (43 ± 6% P13 – P15, 41 ± 9% P22 – P25, 48 ± 19% P31 – P45; p > 0.05, Крускал – Уоллис). С учетом всех расстояний средняя вероятность подключения снизилась; этот эффект не был статистически значимым (18 ± 3% P13 – P15, 13 ± 2% P22 – P25, 9 ± 2% P31 – P45; p > 0,05, Kruskal – Wallis). Таким образом, мы не нашли убедительных доказательств отсечения соединений интернейрона с ПК, поскольку локально плотная архитектура остается неизменной у взрослых животных.
Подтверждение плотной локальной связи с помощью двойных записей целых клеток
Предыдущие результаты показали, что интернейроны PV + могут быть очень плотно подключены к своим локальным ПК, и что в некоторых случаях эта плотная связь может существовать на физическом пределе, когда каждый интернейрон PV + подключен к каждому выбранному ПК.
Учитывая, что этот результат был получен с помощью нового метода картирования, мы стремились подтвердить высокую локальную связь с двойными записями целых клеток в неоптических экспериментах путем записи случайно выбранных пар интернейронов PV + и ПК, чьи клеточные тела находились в пределах 160 мкм друг от друга. (Рис.5 A ). Эти контрольные эксперименты были выполнены на животных в возрасте P13 – P16. В 27 из 33 пар, зарегистрированных в S2 / 3, интернейрон PV + был подключен к ПК (рис. 5 B – D ). Эти моносинаптические соединения имели низкую частоту отказов и большую амплитуду, которая снижалась в течение восьмипотенциальной последовательности, передаваемой с частотой 50 Гц, но восстанавливалась через 1 секунду (рис.5 B ; Таблица 1). Ограничение анализа межсоматическими расстояниями <100 мкм показало, что 24 из 27 пар были синаптически связаны.
Рисунок 5.Подтверждение плотной ПВ + иннервации ПК. A , Анатомическая реконструкция связанной пары с интернейроном PV +, подключенным к ПК. Синий, PV + аксон; красный — PV + дендриты; зеленый — аксон ПК; черный, дендрит ПК. B , Электрофизиологическая запись этого соединения PV + интернейрона с ПК, показывающая IPSC в ПК в ответ на серию из восьми потенциалов действия, вызванных с частотой 50 Гц в интернейроне PV +.IPSC угнетаются в ходе поезда потенциала действия. C , Межсоматические расстояния тестируемых пар из двойных экспериментов по записи целых клеток (красный, подключенный; серый, несвязанный). D , Вероятность соединения наблюдается на разных межсоматических расстояниях. E , Вероятность соединения в пределах 100 мкм не различалась независимо от того, наблюдалась ли при картировании или исправлении во всех протестированных областях и слоях коры. В пределах этого ближайшего диапазона не было никакой разницы в вероятности соединения между слоями и областями.
Таблица 1.Характеристики синаптических связей от интернейронов PV + к ПК ( n = 10 от S2 / 3, S5 и F2 / 3)
Мы также подтвердили плотную локальную связь в S5 и F2 / 3 с двойной записью патч-зажима, которая показала, что 86% (6 из 7) пар в S5 и 80% (4 из 5) пар в F2 / 3 были подключены. В двух случаях в S5 один интернейрон PV + был связан с двумя соседними ПК (на расстояниях 60 и 84 мкм в одном эксперименте и 77 и 92 мкм в другом).
Эти двойные записи целых клеток подтвердили локальную плотную связь интернейронов PV + с ПК, наблюдаемую при оптической стимуляции.Для оптического картирования мы наблюдали высокие вероятности соединения в пределах 100 мкм от ПК: 71 ± 9% в S2 / 3 ( n = 21 карта), 92 ± 8% в S5 ( n = 12 карт), и 80 ± 7% в F2 / 3 ( n = 14 карт) (рис. 5 E ). Результаты, полученные при оптическом картировании и записи патч-кламп, существенно не различались (S2 / 3, p = 0,28; S5, p = 0,38; F2 / 3, p = 0,13; Манн – Уитни; рис. 5 E ). Вероятность соединения в пределах 100 мкм также не различалась между областями и слоями ( p = 0.15 для отображения, p = 0,86 для исправления ошибок, Kruskal – Wallis). Это означает, что локально плотная тормозящая связность одинакова во всех тестируемых корковых областях и слоях.
Отсутствие специфичности в связи между нейронами и пирамидными клетками
Следствием локальной плотной связи от интернейронов PV + к ПК является то, что эти интернейроны должны контактировать со всеми или большинством компьютеров поблизости, независимо от их идентичности или принадлежности к конкретной подсхеме. Мы проверили этот прогноз, сравнив входные карты, полученные одновременно, для нескольких ПК из одного и того же среза (рис.3, 6). Мы использовали эти одновременно полученные карты, чтобы проверить, имеют ли пары ПК, которые были связаны между собой, предпочтительную иннервацию соседними интернейронами по сравнению с парами ПК, которые не были связаны между собой (рис. 6 A ) (Yoshimura et al., 2005) . Для этого анализа использовались карты животных в возрасте P12 – P17.
Рисунок 6.Отсутствие специфичности в связности PV + интернейрон. A , Общая вероятность соединения, рассчитанная как количество интернейронов PV +, которые подключаются к двум ПК, из числа интернейронов PV +, которые подключаются к любому ПК, не отличается в зависимости от того, подключены ли постсинаптические ПК друг к другу. . B , Общая вероятность соединения обратно пропорциональна расстоянию между ПК ( r = -0,3898).
Мы обнаружили, что общая вероятность соединения (количество интернейронов, подключенных к двум ПК, деленное на количество интернейронов, подключенных к любому ПК) для пар ПК была одинаковой, независимо от того, были ли эти пары ПК подключены или нет (0,34 ± 0,09 и 0,28 ± 0,07 для подключенных и неподключенных пар ПК соответственно; p = 0.33, n = 8 подключенных ПК, n = 26 неподключенных ПК, S2 / 3, Манн – Уитни; Рис.6 B ). Общая вероятность подключения для всех пар ПК, подключенных или нет, падала по мере увеличения расстояния между ПК ( p <0,05, n = 34 пары ПК S2 / 3; Рис. 6 D ). Учитывая, что соединенные пары часто встречаются на близких расстояниях, мы были обеспокоены тем, что общие вероятности соединения для подключенных компьютеров могут быть искажены, если расстояния между этими компьютерами будут систематически меньше.Фактически, среднее расстояние между подключенными ПК было значительно ближе, чем для неподключенных ПК (44 ± 5 мкм против 65 ± 6 мкм, p <0,05, непарный тест t с поправкой Велча). Чтобы избежать этого эффекта расстояния, мы ограничили наш анализ парами ПК в пределах 70 мкм друг от друга. Это уменьшило среднее расстояние между неподключенными ПК до 45 ± 5 мкм, что незначительно отличалось от среднего расстояния между подключенными ПК ( p = 0,9284, n = 8 подключенных ПК, n = 16 неподключенных ПК, Манн. –Уитни).В этом сокращенном наборе данных общая вероятность подключения для неподключенных или подключенных ПК была статистически неразличима (0,35 ± 0,11 для неподключенных ПК против 0,34 ± 0,09 для подключенных ПК, p = 0,7286, Манн – Уитни). Этот анализ показал, что интернейроны PV + не различают ПК, которые образуют или не образуют подсхемы.
Пространственные паттерны связанных интернейронов PV + различаются между слоями
Предыдущий анализ связности был выполнен с учетом расстояния интернейронов до пирамидальной ячейки.Для дальнейшего изучения пространственной структуры соединений для трех исследованных корковых локаций мы построили график положения связанных и несвязанных интернейронов PV + в координатной плоскости кортикального контура и использовали эти графики, чтобы выяснить, соответствует ли угловое положение соединенных интернейронов PV + относительно ПК. отображается любая конкретная геометрия (рис. 7 A ). Чтобы визуализировать это угловое распределение, мы вычислили полярные графики, на которых ПК были в центре, а сектор был нанесен с радиусом, пропорциональным средней вероятности соединения интернейронов PV + из этой угловой области (рис.7 В ). На близких расстояниях (0–200 мкм) средняя вероятность соединения под любым заданным углом была однородно высокой по областям и слоям (рис. 7 B , верхний ряд). На дальнейших расстояниях (200–600 мкм) появлялась пространственная картина, при которой ПК преимущественно получали связи от интернейронов, расположенных в вертикально ориентированных позициях (рис. 7 B , средний ряд). Усреднение всех расстояний вместе также выявило предпочтение связей из вертикальных ориентаций и разницу между слоем 2/3 и слоем 5 соматосенсорной и лобной коры (рис.7 B , нижний ряд). В то время как ПК уровня 2/3 получали соединения из мест, более близких к пиальной поверхности, ПК уровня 5 получали соединения из мест в более глубоких слоях ( p = 0,0027, круговой дисперсионный анализ). Согласно условию 90 ° по направлению к пиальной поверхности, среднее направление, из которого ПК получали соединения от интернейронов PV +, было 86 ° в S2 / 3, 258 ° в S5 и 105 ° в F2 / 3; эти углы значительно различались ( p <0,01 между S2 / 3 и S5, p <0.01 между F2 / 3 и S5, p = 0,6494 между S2 / 3 и F2 / 3, тест Уотсона – Вильямса).
Рис. 7.Пространственная картина связанных интернейронов PV + варьируется между слоями. A , Положение подключенных (красный) и несвязанных (серый) интернейронов PV + нанесено на график относительно записанного ПК (центр каждого графика). B , Полярные графики показывают вероятность соединения из заданной угловой области с пиальной поверхностью в верхней части круга.Вероятность соединения равна 1 на внешнем краю круга и 0,5 на среднем пунктирном круге. Полярные графики в каждом столбце представляют разные области и слои коры. В строках показано угловое распределение соединений с учетом только интернейронов на определенных расстояниях (в пределах 200 мкм, от 200 до 600 мкм или на всех расстояниях).
Таким образом, мы пришли к выводу, что существуют значительные различия в пространственном паттерне связи между PV + клетками верхнего и нижнего слоев, хотя в обоих случаях и в обеих корковых областях имело место преимущественное вертикальное расположение пресинаптических нейронов.
Оценка сходимости и расхождения в схемах PV – PC
Высокая плотность связи, которую мы наблюдаем от интернейронов PV + к соседним ПК, имеет два значения: (1) каждый ПК должен получать конвергентные соединения от многих интернейронов PV + и (2) каждый интернейрон PV + должен контактировать со многими ПК. Используя функцию вероятности, описывающую связность, мы вычислили эти числа и выяснили, являются ли они самосогласованными. Для этого мы построили куб неокортекса, 500 мкм на сторону, с одним ПК, расположенным в центре (рис.8, черный треугольник) и средней плотности 40 000 нейронов / мм 3 (рис. 8) (Knox, 1982). Мы предположили, что 80% нейронов были возбуждающими, а 20% — тормозными (Beaulieu et al., 1992) и что одна четверть тормозных нейронов была парвальбумин-положительной в S2 / 3 (Xu et al., 2010). Затем мы использовали экспериментально определенную функцию, описывающую вероятность подключения интернейронов PV + к центрально расположенному ПК, которая является экспоненциально убывающей функцией расстояния с постоянной затухания 124 мкм без других настраиваемых параметров (рис.4 E ). Мы предположили, что вероятность соединения уменьшалась одинаково во всех направлениях, и выбрали центральный ПК в качестве представителя любого S2 / 3 ПК в неокортексе.
Рис. 8. Модель, показывающая конвергенцию интернейронов PV + на одном ПК. A , Куб неокортекса 500 мкм с каждой стороны от 2/3 соматосенсорного слоя с 5000 нейронами. Куб был построен из интернейронов PV + и ПК с использованием профиля связности, который мы наблюдали экспериментально в S2 / 3, с одним репрезентативным ПК, выделенным черным цветом в центре.Другие ПК отмечены синим цветом, соединенные PV + интернейроны — красным, а несвязанные PV + интернейроны — серым. Плотность соединенных интернейронов PV +, сходящихся на одном ПК, можно визуализировать в трех измерениях. B , Схема архитектуры плотно связанных схем от интернейронов PV + на локальные ПК.
Если ограничиться пластиной толщиной 50 мкм с центром на ПК, эта модель точно воспроизвела наши результаты, предсказывающие, что к ПК будет подключено шесть интернейронов PV +.Хотя на картах мы наблюдали только в среднем три связанных PV + интернейрона в S2 / 3, у этой трансгенной мыши помечена только половина парвальбумин-положительных интернейронов (Chattopadhyaya et al., 2004).
Затем мы расширили модель, чтобы проанализировать два значения, изложенные в начале этого раздела, анализируя весь куб 500 мкм. Наша модель предсказывала, что к ПК будет подключено 46 интернейронов PV +. С оценкой того, сколько PV + интернейронов контактирует с каждым ПК, и отношения количества ПК к PV + интернейронам, мы затем вычислили, сколько ПК в среднем контактирует каждый PV + интернейрон.Для этого расчета мы определили C как количество ПК, V как количество PV + интернейронов, r = C / V как отношение ПК к PV + интернейронам, N как количество PV + интернейронов, которые подключаются к каждому ПК, N · C как общее количество PV + интернейронов к ПК, и N · C / V = N · R как количество ПК, с которыми связался один ПВ + интернейрон.
Учитывая, что количество интернейронов PV +, которые подключаются к каждому ПК (переменная N ), равно 46 и что отношение ПК к интернейронам PV + (переменная R ) составляет 80% к 5% (см. Параграф выше), или 16 число ПК, с которыми контактирует один PV + интернейрон, составляет N · r = 46 · 16 = 736. Известно, что PV + интернейроны контактируют также с другими PV + интернейронами (Galarreta and Hestrin, 2002). Следовательно, общее количество исходящих соединений, сделанных интернейронами PV +, вероятно, даже больше.Если мы предположим ту же конвергенцию 46 интернейронов PV + в один PV +, как мы нашли для соединений PV + с ПК, количество исходящих соединений интернейронов PV + изменится только с N · C на N ( C + V). ), так что количество ПК и интернейронов PV +, с которыми контактирует один интернейрон PV +, алгебраически уменьшается до N ( R + 1), что составляет 46 · 17 = 782. Таким образом, мы оцениваем, что типичные контакты PV + интернейрон ∼ 800 постсинаптических нейронов, как ПК, так и другие ЛВ + интернейроны.
Достаточно ли бутонов у PV + интернейронов в соматосенсорном слое 2/3 для контакта с 800 нейронами? Клетки корзины с быстрой добавкой (FS) в слое 2/3 содержат ~ 4000 бутонов (Tamás et al., 1997; Wang et al., 2002; Karube et al., 2004). В среднем на каждую постсинаптическую сому в тестируемом здесь возрасте приходится четыре бутона из интернейронов PV + (Chattopadhyaya et al., 2004). Следовательно, можно ожидать ∼1000 исходящих соединений от интернейронов PV +.
Аналогичный расчет был выполнен для наборов данных S5 и F2 / 3.Мы построили аналогичный куб неокортекса для соматосенсорного слоя 5, в котором доля интернейронов, которые являются PV +, составляет ∼60% (Xu et al., 2010), а наша экспериментально определенная константа пространственного распада для связности составила 183 мкм, что дало 184 PV + интернейроны, контактирующие с каждым ПК. Поскольку в целом имеется больше PV + интернейронов, количество нейронов, с которыми контактирует каждый PV + интернейрон, все еще составляло ~ 1000 [ N ( R + 1) = 184 · (80/12 + 1) = 1410]. Для лобного слоя коры 2/3 доля интернейронов, которые являются PV +, составляет ~ 40% (Xu et al., 2010), а константа распада составила 180 мкм, в результате с каждым ПК контактировал 121 интернейрон PV +. Число нейронов, с которыми контактировал каждый ЛВ + интернейрон, составило 121 · (80/8 + 1) = 1331.
Итак, используя модель, мы оцениваем, что типичный интернейрон PV + в неокортексе контактирует с несколькими сотнями до 1000 постсинаптических мишеней локально. В то же время, каждый ПК контактирует с большим количеством интернейронов PV +, в диапазоне от 50 до почти 200. Следовательно, эта цепь PV + интернейрон-ПК имеет высокую степень локальной дивергенции и конвергенции.
Наши результаты указывают на архитектуру схемы, в которой ПК по всей неокортексе и развитию получают постоянно высокую плотность входов от PV + интернейронов поблизости (Рис. 8 B ).
Обсуждение
Методологические соображения
Мы использовали новый метод для отображения синаптических связей, поэтому уместно обсудить его преимущества и потенциальные ограничения. Этот метод основан на двухфотонной развязке глутамата для активации отдельных нейронов при записи синаптических токов в других нейронах, входные данные которых мы пытаемся сопоставить.Двухфотонная активация имеет решающее значение для пространственного разрешения метода, поскольку малая функция рассеяния точки, созданная двухфотонным процессом (рис. 1 H , I ), обеспечивает точность для одной ячейки.
Две потенциальные проблемы с этой оптической техникой заключаются в следующем: (1) неспособность активировать целевой нейрон и (2) непреднамеренная активация нейрона, который не был оптически нацелен. Чтобы гарантировать, что целевой нейрон действительно сработал, мы использовали разные уровни мощности лазера, в том числе мощность, которая всегда запускала клетки PV + (300 мВт на образце).Тем не менее, мы не можем гарантировать, что каждый нейрон с оптическим прицелом действительно сработал. Более того, эти эксперименты проводились на срезах мозга, которые имеют много разорванных входов, поэтому наша стратегия дает заниженную оценку истинной связи, подразумевая, что плотность связей может быть выше, чем мы наблюдаем.
Чтобы решить проблему непреднамеренной активации нейронов, мы проверили, срабатывают ли ПК, когда фотостимуляция не сосредоточена на их соматах. Мы редко наблюдали активацию нейрона, когда мы нацеливали лазер за пределы его сомы, даже если мы целенаправленно выбираем ПК с более низкой реобазой, чем интернейроны PV + для этого калибровочного эксперимента (рис.1 Дж ). Кроме того, мы использовали тест переключения (рис. 2 B ), чтобы подтвердить ингибирующую идентичность фотостимулированной клетки. Учитывая более низкую частоту тормозных интернейронов по сравнению с возбуждающими нейронами в неокортексе, кажется маловероятным, что другой интернейрон будет достаточно близко, чтобы вызвать наблюдаемое ингибирующее событие.
Два независимых эксперимента по записи целых клеток подтверждают правильность метода, а также подтверждают результаты. Во-первых, предсказанные связи были подтверждены путем исправления целевого пресинаптического интернейрона (рис.2). Девять из 10 предположительно соединенных пар были действительно соединены, а восемь из восьми предположительно несвязанных пар не были соединены. Во-вторых, оптические результаты совпадают с двойными записями целых клеток из случайно выбранных пар PV + и PC (рис. 5).
Плотная связность PV + интернейронов
Наш главный результат — поразительно высокая плотность соединения интернейронов PV + с соседними пирамидными клетками в соматосенсорных слоях коры 2/3 и 5 и лобном слое коры 2/3 (рис.4, 8). Это подтверждается двойными записями целых клеток (рис. 5) и в некоторых экспериментах может приближаться к пределу подключения каждого интернейрона к каждому локальному ПК. Хотя большинство данных было получено в раннем возрасте развития (2 недели после рождения), мы измерили аналогичные результаты у взрослых животных, поэтому такая плотная связь не является временной стадией развития (рис. 4 H ).
Сообщенные скорости связи между интернейронами, идентифицированными как PV +, FS или корзины, и соседними ПК сильно различаются (Thomson et al., 1996, 2002; Тамаш и др., 1997; Рейес и др., 1998; Wang et al., 2002; Холмгрен и др., 2003; Йошимура и Каллауэй, 2005; Капфер и др., 2007; Томсон и Лами, 2007; Освальд и др., 2009). В соответствии с высокой плотностью, которую мы наблюдали, сообщалось о степени связности 67% для клеток с быстрым выбросом ПК в пределах 50 мкм в слое 2/3 соматосенсорной коры крыс (Kapfer et al., 2007). Тем не менее, в слое 3 коры головного мозга крыс зарегистрировано всего 16% (Thomson et al., 2002). Также наблюдались промежуточные значения 50–60% (Holmgren et al., 2003). Кроме того, небольшое снижение связи ФС с ПК с межсоматическим расстоянием было зарегистрировано для слоя 2/3 зрительной или соматосенсорной (Holmgren et al., 2003) и слуховой (Oswald et al., 2009) коры. Кроме того, исследования с однофотонным лазерным сканированием показывают, что тормозящие входы, поступающие от всех подтипов интернейронов, происходят преимущественно из местных источников (Dantzker and Callaway, 2000; Yoshimura et al., 2005) и имеют схожие интраламинарные паттерны во всех областях (Kätzel et al., 2011). ).
Мы обнаружили 468 пар подключенных PV + интернейрон-ПК среди 1747 протестированных пар (не считая ложноположительных), что является значительным увеличением количества проверенных соединений по сравнению с предыдущими исследованиями.Такое большое количество протестированных пар позволило нам сделать выводы об основных скоростях подключения и напрямую оценить истинные скорости для каждого среза.
Используя наш большой набор данных, мы вычислили функцию, описывающую, как вероятность соединения зависит от межатомного расстояния. Мы обнаружили большое расхождение с выходами интернейронов PV +, которое идет рука об руку с конвергенцией многих интернейронов PV + на одном ПК, которые мы наблюдали на наших очень плотных картах. Мы оцениваем более высокую плотность сходящихся соединений на ПК, чем рассчитывали ранее, хотя наши данные могут фактически недооценивать истинную вероятность соединения.Предыдущие исследования в неокортексе (Tamás et al., 1997) и гиппокампе (Buhl et al., 1994) оценили, что около 25 PV + интернейронов сходятся на одном PC. Эта оценка ниже, чем полученная нами, но она также предсказывает только N ( R + 1) = 25 · 17 = 425 соединений от PV + интернейронов к ПК, что мало, если учесть большое количество бутонов, присутствующих в одиночный аксон PV + (∼4000) (Wang et al., 2002; Karube et al., 2004). Следовательно, высокая плотность соединений, которые мы оцениваем от интернейронов PV + к ПК, кажется более совместимой с наблюдаемым большим количеством бутонов.
Мы недавно описали, как соматостатин-положительные (SOM +) интернейроны также создают локально плотные связи с соседними ПК (Fino and Yuste, 2011). Средняя вероятность соединения интернейрона SOM + на ПК в слое 2/3 лобной коры составила 43%, что несколько выше, чем средняя вероятность соединения, которую мы наблюдали для интернейронов PV + в любом слое или области. Но вероятность соединения для интернейронов ПОВ + в пределах 200 мкм от ПК составляла 71%, что аналогично вероятностям соединения, которые мы наблюдали в пределах 100 мкм.Сходная плотная матрица локальных связей из этих двух различных субпопуляций интернейронов на ПК подразумевает, что ингибирующие связи могут быть построены согласно одному и тому же базовому плану, который для обеих популяций нейронов также остается неизменным у взрослых животных.
Неспецифическая иннервация клеток ПК
Наш второй основной результат — открытие, что вероятность того, что два ПК получат соединения от одних и тех же интернейронов PV +, не зависит от того, связаны ли эти два ПК синаптически (рис.6). Этот результат согласуется с высокой плотностью соединений, которую мы наблюдаем, поскольку нет места для специфичности с такой высокой вероятностью — другими словами, нет нейронов, которые бы специально избегали контакта.
В прошлом исследовании рассмотрение пары ПК, которые синаптически связаны как часть одной и той же подсхемы, не выявило различий в подавлении, получаемом парой ПК (Yoshimura et al., 2005). Однако FS клетки, которые были реципрокно связаны с ПК, имели более общий вход от источников возбуждения, чем те, которые не были реципрокно связаны (Yoshimura and Callaway, 2005).Кроме того, клетки FS преимущественно нацелены на ПК, которые обеспечивают реципрокные возбуждающие связи (Yoshimura and Callaway, 2005). Наш метод с разрешением одной ячейки позволил нам напрямую определить, были ли соединения PV + интернейрон с соседними ПК специфичными. Мы не нашли доказательств того, что интернейроны PV + преимущественно контактируют с парами ПК в одной и той же подсхеме (Рис. 6 A ). Таким образом, наши результаты отличаются от результатов Yoshimura and Callaway (2005), которые показали предпочтительные связи между FS-клетками и ПК, обеспечивающими реципрокное возбуждение.Но в то же время мы не проверяли ту же гипотезу, поскольку наша оптическая стратегия не позволяла нам обнаружить реакцию интернейронов PV + на стимуляцию ПК. Кроме того, в предыдущем исследовании оценивалась связность зрительной коры крыс в возрасте P21 – P26, то есть область коры, возраст и вид, отличные от используемых нами.
Каноническая микросхема для неокортикального торможения?
Наши совместные результаты, картирование интернейронов SOM + (Fino and Yuste, 2011) и PV + (это исследование) демонстрируют, что ингибиторные связи не обладают специфичностью в терминах контакта с ПК.Кроме того, широкое ингибирование, обеспечиваемое интернейронами PV + и интернейронами SOM +, согласуется с недавними открытиями, что нейроглиоформные клетки высвобождают GABA, чтобы опосредовать передачу объема к соседним нейронам неспецифическим образом (Oláh et al., 2009).
В связи с отсутствием специфичности в их выходных сигналах интернейроны в слое 2/3 зрительной коры в целом настроены на ориентацию стимула (Sohya et al., 2007; Niell and Stryker, 2008; Kerlin et al., 2010; Ma et al., 2010; Zariwala et al., 2011), хотя некоторые PV + интернейроны в более поверхностной части слоя 2/3, по-видимому, резко настроены (Runyan et al., 2010). Неокортикальные интернейроны также получают входные данные от нейронов, настроенных на разные ориентации (Bock et al., 2011), обеспечивая анатомическую основу для наблюдения за широкой настройкой интернейронов.
Учитывая, что интернейроны действуют как чужеродные захватчики во время развития (Anderson et al., 1997), скупо, что их программа соединения может быть относительно простой, без необходимости в механизмах, специально соединяющихся с существующими цепями пирамидных клеток.Распространение этого покрытия ингибирования неспецифическим образом на все локальные ПК (Fig. 8 B ), следовательно, может представлять собой каноническую схему для всех ингибирующих соединений в неокортексе.
PE0003 Технический паспорт продукта по CML Microcircuits
% CML Microcircuits COMMUN / CATI‘ON SEMICON OPS© 2014 CML Microsystems Plc 1
Предварительный просмотр продукта
PE0003
Универсальная интерфейсная карта
Введение
Интерфейсная карта PE0003 представляет собой универсальную интерфейсную систему для использования с оценочными наборами CML,
, упрощая процесс оценки продукта и помогая процесс разработки конечного продукта.
PE0003 заменяет PE0002 и обратно совместим с существующими сценариями.
Краткое описание
Основанная на микропроцессоре NXP ARM LPC4330 и с использованием графического интерфейса ПК, генерируемая информация форматируется,
синхронизируется и доставляется в целевую ИС через выбранные последовательные интерфейсы C-BUS или порт хоста. Поставляемое программное обеспечение управления
можно использовать для выполнения операций чтения и записи C-BUS, чтения или записи выборочных данных или выполнения функций сценариев
.
В случае оценки продукта на основе FirmASIC®, изображения функций могут быть сохранены на карте памяти Micro SD PE0003
(если она установлена).
Связь между ПК и PE0003 осуществляется через высокоскоростной USB 2.0.
PE0003 обеспечивает среду для разработки и оценки кода C с использованием цепочки инструментов NXP, LPCXpresso
и JTAG. Набор проверенных библиотек C можно загрузить в пакете поддержки PE0003 для помощи в разработке приложения
.
Основные характеристики
Глобальный интерфейс для оценочных комплектов ИС нового поколения
Целевые операции чтения и записи C-BUS ИС
32-битный двухъядерный микропроцессор ARM Cortex-M4 / Cortex-M0
Интерфейс сопряжения для широкий спектр оценочных комплектов CML
Карта Micro SD для хранения изображений функций и массивов данных
Графический интерфейс ПК и оборудование предоставляется
Обработка изображений функций для проектов на основе FirmASIC®
Высокоскоростное выполнение скриптов в реальном времени
Управление ПК / связь через USB
Программно конфигурируемые линии GPIO и банк светодиодов
Библиотека C для быстрой разработки на языке C
Порт JTAG для отладки приложений C
Справочное руководство по сценариям
Маленькая печатная плата 80 мм x 93 мм
PP / 0003/1 августа 2014
PE0003 Универсальная интерфейсная карта
Raspberry Pi, Zero, Linux, Board, raspberry pi, zero, compu тер, компонент, процессор
Raspberry Pi, Zero, Linux, Board, raspberry pi, zero, компьютер, компонент, процессор | Piqsels Raspberry Pi, Zero, Linux, Board, raspberry pi, zero, компьютер, компонент, процессор, микросхема, чип, сверхмалый, технология, печатная плата, компьютерный чип, электронная промышленность, сложность, связь, компьютерное оборудование, беспроводные технологии, компьютер часть, Соединение, портативный компьютер, компьютерная клавиатура, клавиатура, компьютерная сеть, Интернет, Крупным планом, нет людей, Финансирование, зеленый цвет, Компьютерный ключ, оборудование, 2KPublic DomainКлючевые слова для фото
Измените размер и загрузите это фотоПК и ноутбук (720P, 1080P, 2K, 4K):
iMac:
iMac 21.5 дюймов со светодиодной подсветкой:
1920×1080iMac 21,5 дюйма Retina 4K:
4096×2304iMac 27 дюймов Retina 5K:
5120×2880MacBook:
MacBook Air 11.6 «:
Полный размер — 1366×768MacBook Air 13 дюймов, MacBook Pro 15.4 «:
Полный размер — 1440×900MacBook Pro 13.3 «:
1280×800MacBook Pro с дисплеем Retina с диагональю 15,6 дюйма:
2880×1800MacBook Pro 17 дюймов:
1920×1200MacBook Pro 13,3 дюйма Retina, MacBook Air 13 дюймов Retina:
2560×1600iPhone:
iPhone 2G, iPhone 3G, iPhone 3GS:
320 x 480iPhone 4, iPhone 4s:
640 x 960iPhone 5, iPhone 5s, iPhone 5c, iPhone SE:
640×1136iPhone 6, iPhone 6s, iPhone 7, iPhone 8:
750×1334iPhone 6 plus, iPhone 6s plus, iPhone 7 plus, iPhone 8 plus:
1242×2208iPhone X, iPhone Xs:
1125×2436iPhone Xs Max:
1242×2688iPhone Xr:
828×1792Телефон Android:
iPad:
iPad, iPad 2, iPad Mini:
768×1024iPad 3, iPad 4, iPad Air, iPad Air 2, iPad 2017 г., iPad Mini 2, iPad Mini 3, iPad Mini 4, 9.IPad Pro, 7 дюймов:
2048×1536iPad Pro 10,5 дюйма:
2224×1668iPad Pro 11 дюймов:
2388×1668iPad Pro 12,9 дюйма:
2732×2048Surface и планшеты с Android:
Связанные фотографии
- 2000x1334px свет, портативный компьютер, технологии, размытость, черный, крупный план, компьютер, электроникаPublic Domain
- 5456x2872px зеленый, печатная плата, wth, ЖК-экран, Чип, Цепь, Крупный план, Электроника
- 5472x3648px серебряный ноутбук, различные, предметы, стол, серебро, ноутбук, журналы, компьютерPublic Domain
- 5202x3468px серебристый macbook pro, macbook, pro, фокус, фото, аналитика, графики, трафик
- 5472x3648px серебряный ноутбук, различные, предметы, стол, серебро, ноутбук, журналы, компьютерPublic Domain
- 4104x2736px серебристый ноутбук acer, белый, блокнот, деревянный, стол, серебристый, Acer, ноутбукPublic Domain
- 4104x2736px серебристый ноутбук acer, деревянный, стол, зеленый, растение, карандаши, серебро, AcerPublic Domain
- 1600x2401px ноутбук, стол, пульт, белый, Apple, компьютер, macbook, ноутбукPublic Domain
- 5472x3648px серебро apple macbook, серебро, Apple MacBook, Apple, компьютер, macbook, ноутбук, ноутбукPublic Domain
- 4608x3456px Клавиатура, Телефон, Желтый, Примечание, Белый, Стол, компьютер, iphone
- 5007x3338px ноутбук, смартфон, наушники, белый, ноутбук, мобильный телефон, сотовый телефон, acerPublic Domain
- 5321x3547px ноутбук, смартфон, наушники, белый, ноутбук, мобильный телефон, сотовый телефон, acerPublic Domain
- 5076x3384px рабочее пространство, визитки, записные книжки, Elegant, Macbook, iPhone, место для копирования, copyspacePublic Domain
- 4104x2736px серебристый ноутбук acer, деревянный, стол, зеленый, растение, карандаши, серебро, AcerPublic Domain
- 5396x3597px ноутбук, смартфон, наушники, белый, ноутбук, сотовый телефон, acer, карандашиPublic Domain
- 5035x3357px Детали, Apple iMac, компьютеры, iMac, Apple, технология, технологии, компьютер
- 3264x2448px темно, ноутбук, технологии, компьютер, алфавит, концептуальные, устройство, дисплейPublic Domain
- 5398x3599px серебряный ноутбук, различные, предметы, стол, серебро, ноутбук, журналы, компьютерPublic Domain
- 4019x2679px белый, компьютер Apple imac, коврик для мыши слона, белое яблоко, Apple iMac, компьютер, слон, коврик для мыши
- 3780x2520px серебристый ноутбук acer, белый, блокнот, деревянный, стол, серебристый, Acer, ноутбукPublic Domain
- 2250x1529px зеленая монтажная плата, плата, электроника, компьютер, обработка данных, печатная плата, данные, процессор
- 4922x3281px маленький, розовый, цветы, горшок, различные, предметы, флора, таблицаPublic Domain
- 5472x3648px macbook, компьютер, ноутбук, технологии, программирование, кодирование, код, бизнес
- 5227x3485px Детали, Apple iMac, компьютеры, iMac, Apple, технология, технологии, компьютер
- 4047x2698px ноутбук, стол, пульт, белый, Apple, компьютер, macbook, ноутбукPublic Domain
- 5472x3648px современный домашний офис, интерьер, дизайн интерьера, офис, стул, рабочее пространство, рабочее место, ApplePublic Domain
- 5025x3350px серебряный ноутбук, различные, предметы, стол, серебро, ноутбук, журналы, компьютерPublic Domain
- 5472x3648px белый, стол, портативный компьютер, интерьер, минимальный, рабочее пространство, рабочее место, технология
- 5210x3473px стол, домашний офис, розовый, женский пол, дом, офис, рабочее пространство, рабочее место
- 5371x3581px ноутбук macbook, синий, пуф, технология, macbook, технологии, ноутбук, пуфик
- 5220x3480px офисный стол стол, красный, розы, офис, стол, стол, красные розы, женский
- 4104x2736px открытки, цитаты, черно-белое, фото, серебряный ноутбук, Вдохновляющее, серебро, ноутбукPublic Domain
- 5472x3648px портативный компьютер, ноутбук, macbook, технология, код, кодирование, компьютер, разработчик
- 5000x3333px печатная плата компьютера, технологии, данные, технология, компьютерный чип, печатная плата, электронная промышленность, материнская плата
- 2432x3648px стильный, рабочее место, компьютер macbook, дом, студия, технология, компьютер, macbook
- 4500x3000px беспроводной, оранжевый, фон, Apple, технологии, компьютер, компьютерная клавиатура, ноутбук
- 1920x1080px обои компьютерной системы, аннотация, рабочий стол, узор, печатная плата, печатная плата, электронная, компьютер
- 4000x2667px с подсветкой, клавиатура, портативный компьютер, крупным планом, технологии, бизнес, компьютер, работа
- 5253x3502px ноутбук, смартфон, наушники, белый, ноутбук, сотовый телефон, acer, карандашиPublic Domain
- 3927x2618px серебристый ноутбук acer, белый, блокнот, деревянный, стол, серебристый, Acer, ноутбукPublic Domain
- 3424x3424px воздух, MacBook Air, кодирование, HTML, JavaScript, код, macBook, веб-дизайн, общественное достояние
- 5438x3625px рабочее пространство, визитки, записные книжки, Elegant, Macbook, iPhone, место для копирования, copyspacePublic Domain
- 3597x5396px ноутбук, смартфон, наушники, Белый, ноутбук, мобильный телефон, сотовый телефон, acerPublic Domain
- 2410x1635px искусственный, концепция интеллекта, двоичный, код, расширенный, искусственный интеллект, анатомия, тело
- 3500x2318px ноутбук, яблоко, бизнес, компьютер, учеба, технологии, беспроводные технологии, общение
- 5386x3591px ноутбук, смартфон, наушники, белый, ноутбук, мобильный телефон, сотовый телефон, acerPublic Domain
- 5261x3507px стол, домашний офис, розовый, женский пол, дом, офис, рабочее пространство, рабочее место
- 5217x3478px женщина работает, стол, ноутбук, чашка, кофе, macbook, компьютер, работа
- 5553x3702px вид сбоку macbook, usb, hdmi, sd card входы, входы., apple macbook, деревянный, стол
- 5378x3585px рабочее пространство, macbook, pro, стильный, Macbook Pro, интерьер, белый, офис
Принципы кодирования канонической кортикальной микросхемы в головном мозге птиц
Значение
Шестислойный неокортекс является отличительной чертой мозга млекопитающих. Связи между слоями и прогрессивные изменения в свойствах нейронного кодирования каждого слоя определяют микросхему, которая, как предполагается, выполняет вычисления, лежащие в основе сложного поведения.У птиц отсутствует шестислойная кора головного мозга. Тем не менее, они демонстрируют сложное познание и поведение. Недавние анатомические исследования предполагают, что прилегающие области птичьего паллиума являются гомологами неокортикальных слоев. Здесь мы показываем, что слуховой покров птиц демонстрирует те же принципы обработки информации, которые определяют неокортикальную микросхему млекопитающих. Результаты предполагают, что кортикальная микросхема возникла у общего предка млекопитающих и птиц, и обеспечивают физиологическое объяснение нервных процессов, которые приводят к сложному поведению в отсутствие кортикального ламинирования.
Abstract
Неокортекс млекопитающих характеризуется многоуровневой архитектурой и общей или «канонической» микросхемой, управляющей потоком информации между слоями. Считается, что эта микросхема лежит в основе вычислений, необходимых для сложного поведения. Несмотря на отсутствие шестиуровневой коры, птицы способны к сложному познанию и поведению. Кроме того, слуховой покров птиц состоит из смежных областей обработки информации с генетически идентифицированными типами нейронов и выступами среди областей, сопоставимых с таковыми в неокортексе.Здесь мы показываем, что слуховой покров птиц демонстрирует те же принципы обработки информации, которые определяют каноническую корковую микросхему, которая долгое время считалась развившейся только у млекопитающих. Эти результаты предполагают, что каноническая кортикальная микросхема эволюционировала у общего предка млекопитающих и птиц, и обеспечивают физиологическое объяснение эволюции нервных процессов, которые приводят к сложному поведению в отсутствие кортикального ламинирования.
Когнитивные способности птиц позволяют предположить, что птичий мозг содержит сложные схемы обработки информации.Недавние исследования показали, что врановые животные, такие как вороны, грачи и сойки, демонстрируют новаторское производство орудий (1), ссылочные жесты (2), причинно-следственные рассуждения (3, 4), зеркальное самопознание (5) и планирование на будущее. необходимо использовать недавний опыт (6). Другие птицы могут выполнять сложное распознавание образов (7), долгосрочное запоминание (8) и численное различение (9), параллельно с работой приматов. Певчие птицы, такие как зебровые зяблики ( Taeniopygia guttata , вид, изучаемый здесь), учатся производить и распознавать сложные вокализации для социальной коммуникации, с многочисленными параллелями с обучением и восприятием речи (10).
Хотя когнитивные навыки птиц и других млекопитающих часто сопоставимы, паллиумы птиц и млекопитающих имеют очень разную анатомическую организацию. Хотя организация восходящих сенсорных путей и субпаллиальных структур сходна в головном мозге птиц и млекопитающих (11), птичий паллий (называемый в дальнейшем корой головного мозга) лишен характерной шестислойной структуры неокортекса млекопитающих (12). Вместо ламинарной структуры кора головного мозга птиц состоит из взаимосвязанных ядер и полос нейронов, которые образуют отдельные области обработки.Первичная слуховая кора птиц (птичий А1) организована в смежные обрабатывающие области, в совокупности называемые комплексом поля L / CM (каудальный мезопаллий), которые укладываются друг на друга в дорсоростральной и вентрокаудальной ориентации (13, 14) (рис.1 и рис. S1). Области птичьего A1 разграничены различиями в цитоархитектуре и связности (13, 15) ( SI Materials and Methods ) и организованы на поверхностные (поле L1 и латеральный каудальный мезопаллий, CML), промежуточные (поля L2a и -b), и глубокие (поле L3) области, аналогичные слоям неокортекса.Задний отдел, каудальный нидопаллий (NC), здесь называется «вторичной» корой. Рис. 2 A показывает схематическую диаграмму основных соединений между регионами (изменено из ссылки 16).
Рис. 1.Области слуховой коры певчих птиц. ( Left ) Парасагиттальный срез, окрашенный по Нисслю, показывающий тела и пластинки клеток. ( Правый ) Рисунок того же сечения с окрашенными и помеченными областями. Пунктирными линиями обозначены границы, определяемые переходами в цитоархитектуре.Области L поля L1, L2, L3 и боковой каудальный мезопаллий (CM) образуют первичную слуховую кору у птиц (13, 14). Поле L2a и L2b — промежуточные (таламореципиентные) области. Поле L1 и CML — это поверхностные области, а поле L3 — глубокая область. Хвостовой нидопаллий (NC) — вторичная слуховая зона. Область L (13) имеет цитоархитектуру, аналогичную L2b, но, как известно, не получает вход таламуса. CStr, хвостовое полосатое тело; Hp, гиппокамп; HVC, (имя собственное) премоторное ядро песенной системы; ПМЖВ, lamina arcopallialis dorsalis; LaM, lamina mesopallialis.
Рис. 2.Иерархическая обработка информации в слуховой коре певчих птиц. ( A , Left ) Схема основных выступов в слуховой коре певчих птиц (изменено из ссылки 16). Заштрихованная область указывает схему, соответствующую первичной слуховой коре млекопитающих. Вход таламуса в основном заканчивается в поле L2 (промежуточная область) и редко — в поле L3 (глубокая область). Дорсальные выступы от L2 заканчиваются в L1, а L1 проецируется на CML (поверхностные области).Вентральные выступы из L2 заканчиваются в L3. Поле L3 проецируется на NC (вторичный), таламус и ствол мозга. ( Right ) Средние задержки ответа в различных областях слуховой микросхемы и вторичной области NC. Задержки самые короткие в промежуточной области и значительно дольше в поверхностных и вторичных областях. ( B , Left ) Гистологическое изображение, используемое для определения местоположения политрода при одном проникновении записи. Четыре оранжевых дорожки показывают расположение четырех стержней политрода с маркировкой DiI.Зеленая стрелка указывает предполагаемое расположение наконечника стержня электрода. ( Правый ) Схема политрода. ( C ) Примеры спайковых поездов и рецептивных полей (RF) одиночных нейронов в поверхностных, промежуточных, глубоких и вторичных областях. Спектрограмма стимула (2 с мл шума) показана над последовательностью спайков. ( D ) Интенсивность вызванного возбуждения (разреженность отдельных ячеек). ( E ) Редкость населения. ( F ) Избирательность. ( G ) Индекс разделимости RF.( H ) RF линейность. На гистограммах столбцы ошибок указывают SEM. Звездочки имеют цветовую кодировку в зависимости от региона и указывают на значимые сравнения ( P <0,05, критерий Краскела – Уоллиса с поправкой на сравнение нескольких групп).
Несмотря на крупномасштабные различия в организации коры головного мозга птиц и неокортекса млекопитающих, сходство в этих областях мозга недавно было обнаружено на круговом (14), клеточном (11) и молекулярном уровнях (17-19). Анатомические исследования предоставляют доказательства сходной взаимосвязи между поверхностными, промежуточными и глубокими процессинговыми областями в птичьем A1 и соответствующих слоях неокортекса (11, 14, 15, 20, 21).Например, птичий А1 организован в столбцы радиально расположенных внутренних связей, охватывающих все области, аналогично столбцам, соединяющим слои неокортекса (14). Кроме того, одни и те же генетически идентифицированные типы нейронов обнаруживаются в промежуточных, поверхностных и глубоких слоях неокортекса и в различных регионах птичьего A1 (17, 18).
Слои неокортекса млекопитающих и характерные связи между слоями определяют «каноническую» кортикальную микросхему, которая, как считается, лежит в основе вычислений, необходимых для сложного познания и поведения (22).Основываясь на недавних демонстрациях значительных когнитивных навыков у птиц и параллелях в анатомических схемах между корой головного мозга птиц и млекопитающих, мы предположили, что птичий A1 демонстрирует принципы кодирования отдельных нейронов и популяции, которые характеризуют каноническую корковую микросхему в неокортексе млекопитающих. Путем одновременной регистрации активности 2–38 отдельных нейронов в каждой обрабатывающей области птичьего А1 и анализа возбуждающих свойств мы обнаружили, что кора головного мозга птиц демонстрирует характерные особенности обработки информации канонической кортикальной микросхемы, обеспечивая функциональные доказательства для общей корковой микросхемы. у млекопитающих и птиц.Сходства обработки информации, описанные здесь, предполагают, что птицы и млекопитающие имеют гомологичные корковые микросхемы, и обеспечивают физиологическое объяснение эволюции нейронных процессов, которые приводят к сложному поведению в отсутствие ламинирования.
Результаты
Иерархическая обработка в слуховой коре птиц.
Определяющей особенностью микросхемы неокортекса млекопитающих в основном являются соединения с прямой связью, которые образуют иерархию этапов обработки информации.Сенсорная информация от таламуса в основном нацелена на промежуточный слой 4 и редко нацелена на границу слоя 5/6 (23, 24). Из слоя 4 аксональные проекции переходят в поверхностные слои 2/3, а затем в глубокие слои 5/6, где информация распространяется на другие корковые и подкорковые мишени (23, 25). В первичной слуховой (A1), зрительной (V1) и соматосенсорной (S1) корках рецептивные поля простые и разделимые в слое 4 и более сложные и нелинейные в поверхностных и глубоких слоях (26-29).Сенсорные вызванные ответы обычно более разрежены и более избирательны в слоях ниже по потоку от слоя 4 (30–32), хотя частота возбуждения в слоях 4 и 5 в некоторых системах схожа (30, 33, 34).
Как и в сенсорной коре млекопитающих, таламические проекции на птичий A1 заканчиваются в основном в промежуточной области и редко в глубокой области (15, 20) (Fig. 2 A ). Информация из поля L2 промежуточной области распространяется на поверхностные области (поле L1 и CML) и поле L3 глубокой области (14, 15, 20).Чтобы проверить, является ли иерархическая обработка информации главной особенностью слуховой микросхемы птиц, мы использовали многоэлектродные массивы для одновременной записи слуховых ответов от отдельных нейронов в промежуточных ( n = 94), поверхностных ( n = 219) и глубоких. ( n = 237) областей птичьего A1 у взрослых самцов зебра-зябликов без анестезии ( n = 6) (рис. 2, рис. S1 и S2, и таблица S1). Мы также записали NC вторичной слуховой коры ( n = 273), которая получает входные данные от глубоких и поверхностных областей птичьего A1 (15, 20).
Мы обнаружили, что слуховая кора птиц демонстрирует иерархические особенности обработки информации, которые характеризуют неокортекс млекопитающих (рис. 2). Задержки отклика на представление ограниченного модуляцией (ml) шума (коррелированный гауссовский шум, соответствующий песне зебры в диапазоне частот и максимальных частотах спектральной и временной модуляции) (35) ( Материалы и методы, ) были самыми короткими в промежуточной области и значительно более длинными. в поверхностной и вторичной областях (рис.2 A ) ( P <0,05, критерий Краскела – Уоллиса с поправкой на множественные сравнения). На рис. 2 C показаны последовательности спайков, записанные от одиночных нейронов в каждой области до, во время и после представления шума ml, а также рецептивные поля ( SI Materials and Methods ) для этих нейронов. Редкость единичных клеток, измеренная как вызванная звуком интенсивность возбуждения, и разреженность популяции, рассчитанная как доля клеток, которые не смогли произвести значительный ответ на каждый предъявленный стимул ( SI Materials and Methods ), различались по регионам; активность была наиболее высокой в промежуточной области и значительно реже в поверхностной, глубокой и вторичной областях ( P <0.05) (Рис.2 D и E ). Селективность стимула (доля стимулов, которые не вызывали значимого ответа нейрона) была самой низкой в промежуточной области и значительно выше на последовательных этапах обработки ( P <0,05, рис. 2 F ). Разделимость и линейность рецептивного поля (RF), два показателя простоты RF ( SI Materials and Methods ), были самыми высокими в промежуточной области, ниже в поверхностной области, еще ниже в глубокой области и самыми низкими во вторичной коре (рис. .2 G и H ) ( P <0,05). Эти результаты согласуются с анализом сенсорной обработки в неокортексе млекопитающих (26–28, 31, 32) и предоставляют доказательства того, что иерархическая обработка информации также является определяющей особенностью слуховой микросхемы птиц. Мы обнаружили одно различие в сравнении обработки корковых микросхем птиц и млекопитающих. Исследования A1 и S1 показывают, что активность более редкая и более избирательная в поверхностных слоях, чем в глубоких (28, 30).Мы обнаружили, что разреженность и селективность были выше у глубокой птицы A1, чем у поверхностной A1 (рис. 2 C — E ). Это наблюдение согласуется с недавними открытиями (36) у европейского скворца и может быть различием микросхем между птицами и млекопитающими.
Два основных класса нейронов в слуховой коре птиц.
Определяющей характеристикой неокортекса млекопитающих является наличие двух основных классов нейронов. Возбуждающие основные клетки (ПК) активируют широкие (также называемые широкими или регулярными) потенциалами действия с относительно низкой скоростью, а тормозные интернейроны (ИН) активируют узкие (т.е., тонкие или быстрые) потенциалы действия с более высокими скоростями (23, 25, 37). PC производят гетерогенные, сравнительно редкие и избирательные сенсорные ответы, тогда как IN реагируют на те же стимулы с меньшей редкостью и избирательностью, что приводит к плотным сенсорным репрезентациям (30, 38-40). В A1 млекопитающих PC и IN также различаются по структуре RF; ПК имеют более сложные RF, чем IN (41). Чтобы проверить, группируются ли нейроны птичьего A1 в два основных класса, мы подвергли все зарегистрированные клетки ( n = 823) кластерному анализу на основе формы потенциала действия (AP) (30, 36, 37) (рис.3 A , рис. S3, и SI Материалы и методы ). Нейроны сгруппированы в два класса, и оба класса были обнаружены в поверхностных, промежуточных и глубоких областях, а также во вторичной слуховой коре (рис. 3 B и рис. S3 и S4). Потенциалы действия предполагаемых IN (pIN, n = 235/823) были значительно уже (0,33 ± 0,05 мс), чем потенциалы действия предполагаемых ПК (pPC, n = 588/823) (0,77 ± 0,13 мс) ( P <1e − 10, двухвыборочный критерий Колмогорова – Смирнова).Как и в неокортексе (38), частота стрельбы и ширина AP были отрицательно коррелированы; нейроны с более высокой частотой возбуждения обычно имели более узкие AP (коэффициент корреляции Пирсона (R) = -0,58) (рис. 3 A и рис. S3). На рис. 3 B показаны примерные последовательности импульсов и RF для pIN и pPC в каждой области. Как и в неокортексе млекопитающих (30, 38, 39), pPCs имеют более гетерогенные ответы и RF-структуры, чем pINs, внутри и между регионами (Fig. 3). На рис. S5 показаны цепочки импульсов pPC и RF, организованные по ширине AP.Подобно неокортикальным IN и PC внутри слоя, pPC птиц демонстрировали более низкую частоту вызванных импульсов и более высокую селективность стимулов, чем pIN в той же области (рис. 3 C — E , сравните синий и красный на каждой панели, P <0,01 двухвыборочный критерий Колмогорова – Смирнова). Также, как и в неокортексе, RFs pPCs были менее линейными и более сложными, чем RFs pINs (Fig. 3 F и G ). Свойства отклика и RF-структуры pIN и pPC также различались в зависимости от региона.В широком соответствии с иерархической организацией, показанной на рис.2, разреженность ответа (рис.3 C и D ), селективность (рис.3 E ) и сложность RF (рис.3 F и G ) увеличивается на последовательных стадиях обработки за пределами таламореципиентной области как в pPC, так и в pIN (сравните столбцы одного цвета на каждой панели). Результаты показывают, что различия в свойствах кодирования отдельных нейронов в разных областях обработки и между классами нейронов сходны в первичной коре головного мозга птиц и млекопитающих.
Рис. 3.Свойства ответа pPCs и pIN в слуховой коре певчих птиц. ( A ) Идентификация предполагаемых основных клеток (pPCs, n = 588) и предполагаемых интернейронов с быстрым выбросом (pIN, n = 235) по форме волны спайка. Единицы с сортировкой по пикам были классифицированы на основе средней ширины пикового сигнала и сгруппированы в два класса с использованием модели смеси Гаусса (рис. S3). Предполагаемые IN (синий) имеют узкую форму волны спайка, тогда как pPC (красный) имеют более широкую форму волны.Ширина потенциала действия отрицательно коррелирует с вызванной скоростью стрельбы; Частота возбуждения выше у нейронов с более узкими спайками ( n = 823 нейрона), коэффициент корреляции Пирсона R = -0,58. ( B ) Спайковые реакции поезда на 30 повторений ml-шума для восьми примеров нейронов. Нейроны 1, 3, 5 и 7 — контакты (синие). Нейроны 2, 4, 6 и 8 — pPC (красные). Заштрихованная область обозначает период предъявления стимула. Приемные поля (RF) для каждого примера нейрона находятся справа от растровых графиков.( C ) Ящичковые диаграммы частоты вызванных импульсов для pIN (синий) и pPC (красный), показанных по регионам. ( D ) Редкость населения. ( E ) Избирательность. ( F ) RF разделимость. ( G ) ВЧ линейность. Черные звездочки указывают на достоверные различия между pPC и pIN внутри региона (** P <0,01; * P <0,05; двухвыборочный тест Колмогорова-Смирнова). Цветные звездочки указывают на значительные различия между регионами для pIN и pPC отдельно ( P <0.05, критерий Краскала – Уоллиса с поправкой на сравнение нескольких групп).
Популяционные различия кодирования по регионам, типам нейронов и анатомическому расстоянию.
Популяции нейронов в неокортексе демонстрируют различные паттерны связности в разных слоях и между классами нейронов (например, PC или IN) в популяции. Корреляции во времени спайков, производимых разными нейронами, отражают силу и специфичность связей между нейронами в популяции и измеряются как корреляции количества спайков между одновременно зарегистрированными нейронами.Вызванные стимулами и спонтанные корреляции шума (связанные с общим входом или прямыми синаптическими связями) различаются по слоям и в популяциях ПК и ИН в зависимости от паттернов связности в каждом слое и классе нейронов (23, 31, 42, 43). Во-первых, связность в таламореципиентном слое 4 является локальной и однородной (44), а отдельные нейроны обладают различными возбуждающими свойствами (43, 45, 46). Эти характеристики согласуются с наблюдением слабых парных корреляций в слое 4 по сравнению с таковыми в поверхностных и глубоких слоях (31, 43).В отличие от слоя 4, нейроны в глубоких выходных слоях имеют длительные, повторяющиеся связи и сильные корреляции шума (43, 46). Во-вторых, попарные корреляции различаются в популяциях ПК и ИН; корреляции между IN с быстрым всплеском сильнее, чем между ПК как на поверхностном, так и на глубоком уровнях, что согласуется с более плотной и более широкой связью между IN, чем между ПК (30, 42, 47). Кроме того, разреженные ответы и локализованная связь в ПК на поверхностном уровне по сравнению с ПК на глубине лежат в основе более слабых корреляций между ПК в поверхностных слоях по сравнению с таковыми в глубоких слоях (30, 42, 48).В-третьих, плотность связей между нейронами в популяции влияет на то, насколько сенсорная стимуляция контролирует активность популяции; парные корреляции вызванной и спонтанной активности значительно более сходны в сильно взаимосвязанных популяциях ИН, чем в популяциях ПК с более редкой связностью (23, 42). В-четвертых, корреляция вызванной и спонтанной активности зависит от анатомического расстояния и зависит от пространственной степени связи. Связи между нейронами более локализованы в поверхностных слоях, чем в глубоких, что обеспечивает анатомическую основу для зависимости силы парной корреляции от пространственного расстояния между нейронами в поверхностных, но не глубоких популяциях (30).Возбуждающие связи в поверхностных слоях значительно разрушаются, когда расстояние между нейронами приближается к 150 мкм (49, 50), тогда как возбуждающие связи в глубоких слоях обычно широко распространены (47, 49–52). Эти структурные различия поддерживают локальную кластеризацию всплесковой активности в поверхностных слоях и более широкие спреды коррелированного воспламенения в глубоких слоях.
Чтобы проверить, показывает ли птичий A1 различия в кодировании популяции по регионам, классам нейронов и анатомическим расстояниям, которые отображаются на таковые в неокортексе, мы сравнили вызванные и спонтанные попарные корреляции в ( — ) промежуточных, поверхностных и глубоких областях ( Инжир.4 и рис. S6), ( ii ) pPCs и pIN (рис. 4 и 5 и рис. S6), ( iii ) поверхностные и глубокие pPC (рис. 4 B ) и ( iv ) ) пары с короткими и большими пространственными расстояниями между нейронами (рис. 5). Мы вычислили попарные корреляции количества спайков между всеми одновременно зарегистрированными нейронами в каждой области (рис. S1 и S2 и SI Материалы и методы ) и обнаружили, что различия в популяционном кодировании в регионах, в классах нейронов и на пространственном расстоянии были поразительно похожими. тем, кто находится в неокортексе.Во-первых, как и у млекопитающих, общие шумовые корреляции (все пары вместе) в вызванной активности были слабыми в промежуточной области и сильнее в поверхностных и глубоких областях (рис. 4 A ) ( P <0,05, критерий Краскала-Уоллиса с коррекция множественных сравнений). Во-вторых, вызванные и спонтанные корреляции между парами pPC были значительно слабее, чем между парами pIN, что подтверждает преобладающий вывод о том, что IN более плотно связаны, чем ПК (рис.4 B ) ( P <0.05, критерий Краскала – Уоллиса с поправкой на множественные сравнения; На рис. S6 показаны пары pPC / pIN). Корреляции в парах pPC / pPC также были сильнее в глубокой области, чем в поверхностной (рис. 4 B , P <0,05, сравните красные столбцы на каждой панели). Эти результаты являются физиологическим доказательством того, что, как и в неокортексе, глубокие pPC более тесно взаимосвязаны, чем поверхностные pPC. В-третьих, корреляции спонтанной активности были сильнее, чем корреляции вызванной активности во всех регионах (сравните шкалы на рис.4 B ), что указывает на то, что стимул-драйв декоррелирует активизацию популяции как у птиц, так и у млекопитающих (30, 53). Мы также обнаружили более сильную взаимосвязь между спонтанными и вызванными корреляциями в парах pIN, чем в парах pPC, как в поверхностных, так и в глубоких областях, что является дополнительным доказательством того, что связь между IN более плотна, чем между PC, у птиц, как и у млекопитающих (рис. 4 C сравните красные и синие точки на каждой панели; для поверхностных пар R IN , P = 3.55e − 10 и R PC , P = 8.1e − 06; для глубоких пар R IN , P = 1,86e − 38 и R PC , P = 1,35e − 20). На рис. S6 показаны корреляции промежуточных областей, а в таблице S2 показаны средние значения силы корреляции для каждой области и класса нейронов, измеренные отдельно по вызванной и спонтанной активности. В-четвертых, мы обнаружили, что сила корреляции значительно больше зависела от пространственного расстояния между зарегистрированными нейронами в поверхностных популяциях, чем в глубоких (рис.5 и SI Материалы и методы ). Растровые графики мультиединичной (не сортированной по спайкам) активности на Рис. 5 A демонстрируют сходную пиковую активность при регистрации контактов в глубокой области, но не в поверхностной области, что указывает на связь между нейронами глубокой области на более дальние расстояния. При спонтанной активности корреляции между поверхностными нейронами были значительно слабее в парах «дистальных» (≥300 мкм друг от друга) нейронов, чем в парах «локальных» (≤150 мкм) нейронов (рис. 5 B и C ). ( P <0.01). Корреляции в глубокой области не уменьшались с анатомическим расстоянием, что соответствует результатам у млекопитающего A1 (30). Сила корреляции вызванной активности также уменьшалась с увеличением расстояния в верхней, но не глубокой области (рис. 5 B , справа ) ( P <0,05). Корреляции, вычисленные для пар pPC и пар pIN по отдельности, показали, что корреляции не уменьшались с анатомическим расстоянием в парах pIN (рис. 5 C , слева ) и что корреляции в парах pPC уменьшались с анатомическим расстоянием больше на поверхности, чем на глубокая область (рис.5 C , Right ) ( P <0,05, критерий Краскела – Уоллиса с поправкой на множественные сравнения). Эти анализы демонстрируют, что свойства коррелированной активности в обрабатывающих регионах и классах нейронов слуховой коры птиц близко соответствуют тем, о которых сообщается для соответствующих слоев и классов нейронов в неокортексе млекопитающих.
Рис. 4.Корреляция шума по регионам и типам нейронов. ( A ) Попарные корреляции шума во время вызванной активности для поверхностных, промежуточных и глубоких областей.Звездочки указывают на достоверные различия в корреляциях между регионами ( P <0,05, критерий Краскела – Уоллиса с поправкой на сравнение нескольких групп). Столбцы представляют медианы населения, а полосы ошибок указывают 95% доверительные интервалы для медиан. ( B ) Попарные корреляции для pIN и pPC в поверхностных и глубоких областях для вызванной ( слева, ) и спонтанной ( справа, ) активности. Обратите внимание, что оси y находятся в разных масштабах на панелях Left и Right .( C ) Отношения между нормализованными (по шкале z) спонтанными и вызванными корреляциями в парах pIN (синий) и парах pPC (красный), в поверхностной ( слева, ) и глубокой ( справа, ) областях. Как в поверхностных, так и в глубоких областях вызванные корреляции масштабируются со спонтанными корреляциями для пар pIN, но не для пар pPC. Графики показывают спонтанные парные корреляции вызванными парными корреляциями. Каждая точка представляет одну пару. Коэффициенты корреляции Пирсона (R), определяющие взаимосвязь между спонтанными и вызванными корреляциями для пар pIN / pIN и pPC / pPC, показаны в правом нижнем углу каждого графика.Красные и синие линии соответствуют линейному соответствию данным.
Рис. 5.Связь между попарными корреляциями и анатомическим расстоянием по регионам и типам нейронов. ( A ) Пример одновременной записи многоэлементной (без сортировки спайков) активности от нескольких контактов электродов в поверхностной ( вверху справа ) и глубокой ( внизу справа ) областях. Растры организованы путем увеличения расстояния от контакта с черной записью. Цветные точки слева от растров соответствуют цветам, обозначающим контакты записи на политроде ( Left ).Черный «x» указывает на нефункциональный контакт. ( B ) Пространственная зависимость нормализованных корреляций шума в поверхностных и глубоких областях для спонтанной (, слева, ) и вызванной (, правой, ) активности. Локальные пары — это те, в которых зарегистрированные нейроны находились на расстоянии ≤150 мкм. Дистальными парами были те, в которых зарегистрированные нейроны находились на расстоянии ≥300 мкм друг от друга. Парные корреляции в каждом регионе были нормализованы путем деления на медианное значение локальных корреляций в этом регионе, чтобы поместить корреляции во всех регионах в одну шкалу, что позволяет сравнивать корреляции между регионами.Столбцы показывают медианы населения, а полосы ошибок показывают 95% доверительные интервалы. Звездочки указывают на достоверные сравнения (критерий Краскала – Уоллиса с поправкой на сравнение нескольких групп, * P <0,05, ** P <0,01). Корреляции в поверхностной области затухали с анатомическим расстоянием больше, чем корреляции в глубокой области. ( C ) То же, что и в B , но с корреляциями для пар pIN и пар pPC, показанных отдельно.
Обсуждение
Анализ активности отдельных нейронов и популяций во всех областях слуховой коры птиц и во всех областях слуховой коры птиц показывает, что свойства кодирования информации птичьего A1 очень похожи на те, которые определяют «каноническую кортикальную микросхему» у млекопитающих.Во-первых, смежные и связанные области слуховой коры птиц образуют иерархию обработки информации. Во-вторых, одни и те же два класса нейронов обнаруживаются в коре головного мозга млекопитающих и птиц. В-третьих, нейроны в неокортексе и коре головного мозга птиц демонстрируют сопоставимые стратегии одноклеточного и популяционного кодирования в областях / слоях коры, между типами нейронов и на анатомическом расстоянии. Сходство между этими данными и тем, что известно о сенсорной обработке в неокортексе млекопитающих, подтверждает гипотезу гомологии эволюции мозга (54) и предполагает, что каноническая кортикальная микросхема эволюционировала у общего предка млекопитающих и птиц (11, 14, 17).Если это так, то микросхема эволюционировала в стволовых амниотах и предшествовала кортикальному расслоению по крайней мере на 100 миллионов лет (12). Альтернативная возможность состоит в том, что эволюция паллия у птиц и млекопитающих сходилась в одной и той же организации цепи обработки информации (11). Независимо от этих альтернатив, это исследование дает физиологическое объяснение эволюции нервных процессов, которые приводят к сложному поведению в отсутствие ламинации.
Материалы и методы
Полные методы представлены в SI Материалы и методы .
Группы слуховых нейронов были зарегистрированы в поверхностных ( n = 219), промежуточных ( n = 94) и глубоких ( n = 237) областях первичной слуховой коры шести подголовников без анестезии. , самцы зебровых амадин (Таблица S1). Все процедуры с участием животных были рассмотрены и одобрены Комитетом по уходу и использованию животных Колумбийского университета. Записи были сделаны у тех же птиц в хвостовом нидопаллии (NC), вторичной слуховой области коры ( n = 273).Нейронные отклики вызывались представлением 10 различных образцов (каждый = длительностью 2 с) шума, ограниченного модуляцией (ml). Шум Ml — это коррелированный гауссов шум, отфильтрованный для соответствия частотному диапазону (250–8000 Гц) и максимальным частотам спектральной и временной модуляции песни зебрового вьюрка (35, 55). Каждый из 10 стимулов предъявлялся 30–40 раз в псевдослучайном порядке. Анализ данных проводился в MATLAB (Mathworks). Спайки были отсортированы в автономном режиме (56⇓⇓ – 59). Все зарегистрированные единичные единицы ( n = 823) были классифицированы как предполагаемые интернейроны (pIN) или предполагаемые основные клетки (pPC) на основе ширины потенциала действия (рис.3 и рис. S3). Для каждой области обработки и каждого класса нейронов разреженность отдельных клеток вычислялась как вызванная звуком частота срабатывания импульсов. Редкость популяции рассчитывалась как доля клеток, которые не давали значимого ответа на данный стимул. Селективность рассчитывалась как доля различных стимулов, которые не вызывали значительного ответа от данного нейрона. Спектрально-временные рецептивные поля (STRF) были рассчитаны путем подгонки обобщенной линейной модели к вызванным звуком ответам каждого нейрона (60).Латентность ответа для каждого нейрона вычислялась из STRF при наилучшей частоте возбуждения (61) ( SI Materials and Methods ). Разделимость рецептивного поля была вычислена путем выполнения разложения по сингулярным значениям на STRF. Линейность рецептивного поля измерялась с помощью STRF для прогнозирования ответов на новые стимулы. Спонтанные и вызванные корреляции числа спайков были получены путем вычисления коэффициента корреляции Пирсона между подсчетами спайков (скользящее окно 50 мс) из последовательности спайков всех одновременно записанных нейронов (за исключением пар, записанных с одного контакта электрода).Корреляции шума измерялись с использованием предиктора сдвига для корректировки корреляций, вызванных стимулом (53, 62). На рис. 5 B и C парные корреляции в одном регионе были нормализованы путем деления на медианное значение локальных корреляций в этом регионе, так что корреляции во всех регионах были в одном масштабе, что позволяет сравнивать корреляции на поверхностных уровнях. и глубокие регионы.
Благодарности
Мы благодарим Дж. Мура и Дж. Шумахера за комментарии к рукописи и за их вклад в дизайн экспериментов и анализ данных, а также Р.Бруно и С. Розис за комментарии к рукописи. A.C. был поддержан Международной преддипломной стипендией Медицинского института Говарда Хьюза и S.M.N.W. был поддержан грантом NIH R01-DC009810.
Сноски
-
Вклад авторов: A.C. and S.M.N.W. спланированное исследование; A.C. провел исследование; A.C. проанализировал данные; A.C. и S.M.N.W. написал статью; и S.M.N.W. курировал проект.
-
Авторы заявляют об отсутствии конфликта интересов.
-
Эта статья представляет собой прямое представление PNAS.H.K. Приглашенный редактор по приглашению редакционной коллегии.
-
См. Комментарий на стр. 3184.
-
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1408545112/-/DCSupplemental.
PragmatIC Semiconductor привлекает 80 миллионов долларов финансирования
© PragmatIC Бизнес | 19 октября 2021 г.PragmatIC Semiconductor, компания, занимающаяся гибкой электроникой, получила финансирование серии C.Компания заявляет, что использует средства для увеличения своих мощностей в Великобритании.
Эти инвестиции в масштабирование будут использованы для строительства второй фабрики FlexLogIC на северо-востоке Англии, чтобы удовлетворить растущий спрос на сверхдешевые гибкие интегральные схемы (FlexIC) для всего Интернета, говорится в пресс-релизе. «Наша фабрика FlexLogIC-002 будет обеспечивать значительно более высокую производительность, чем наша первая линия, при этом сохраняя при этом наши фирменные сверхнизкие капитальные затраты, быстрое время производственного цикла и минимальный углеродный след.Помимо поддержки нашего постоянного коммерческого роста, он предоставляет шаблон для развертывания распределенной глобальной сети систем FlexLogIC, предлагая Fab-as-a-Service (FaaS) для выделенного производства на основных объектах клиентов, чтобы обеспечить эффективную и безопасную поставку полупроводников. цепи », — говорит Скотт Уайт, генеральный директор PragmatIC Semiconductor. Развитие PragmatIC за последние несколько лет привело к запуску его флагманской линейки продуктов ConnectIC — сверхдешевых RFID FlexIC, а также услуги FlexIC Foundry.В этом году компания продемонстрировала повышение сложности и вычислительных возможностей некремниевых микросхем на порядок за счет производства гибких микропроцессоров. К ним относятся 6502 и PlasticArm, ультра-минималистичная система на кристалле Arm Cortex-M0. PlasticArm — это результат многолетнего сотрудничества между двумя компаниями, партнерства, которое будет продолжаться и в будущем с участием в этом раунде финансирования компании Arm, лидера в области интеллектуальной собственности в области полупроводников. «Миллиарды предметов повседневного обихода могут выиграть от того, что станут частью Интернета вещей, но для дальнейшего устойчивого масштабирования нам необходимо изучить новые подходы для встраивания интеллекта в повседневные предметы», — добавляет Дипеш Патель, технический директор Arm.
