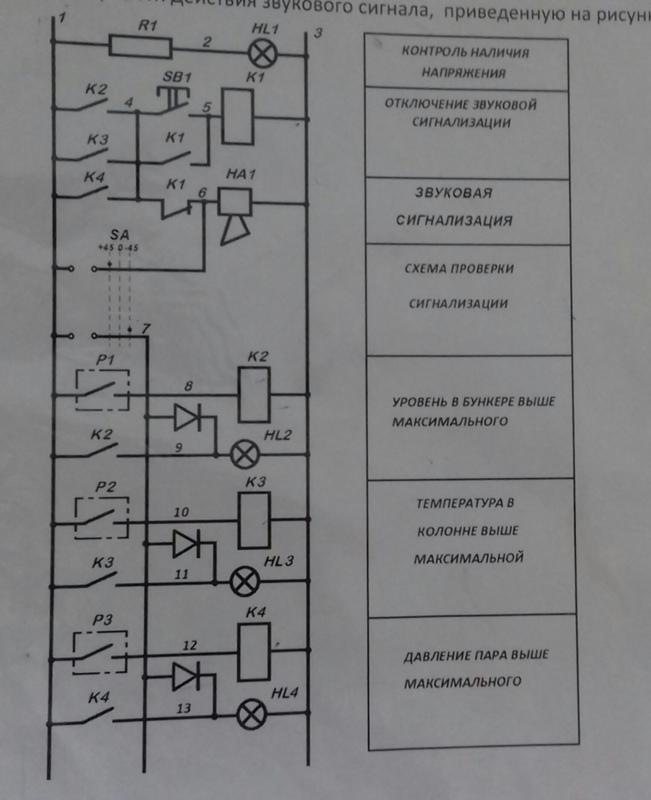
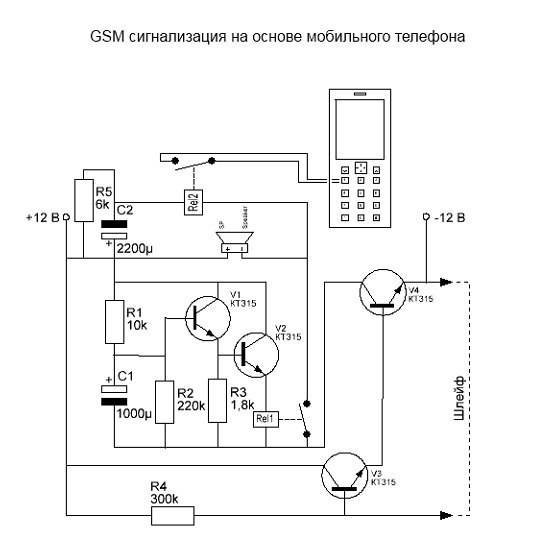
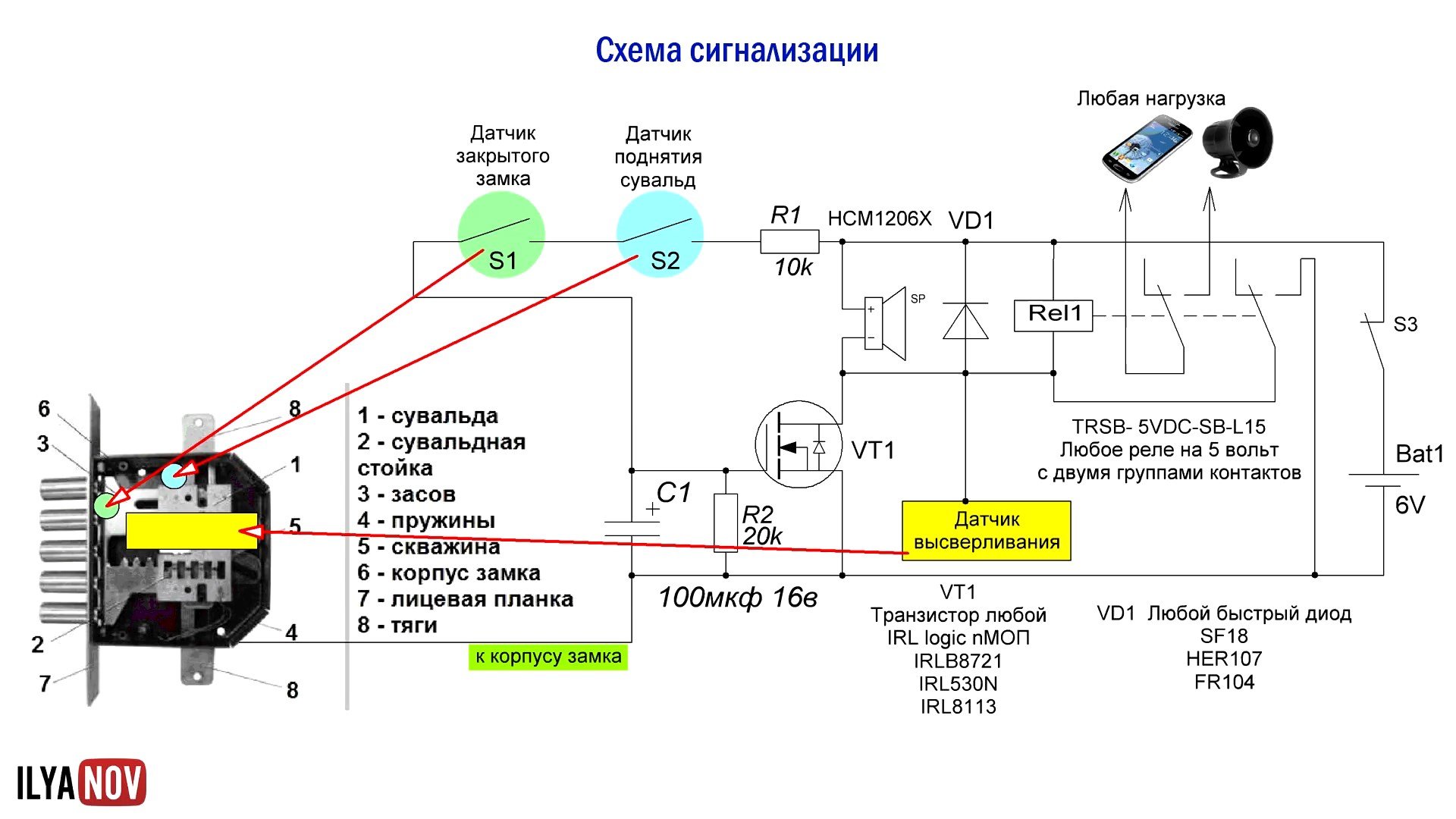
Как устроены современные автосигнализации. Какие основные компоненты входят в состав охранных систем. Как правильно подключить сигнализацию в автомобиле. На каких принципах основана работа противоугонных комплексов. Какие схемы используются в популярных моделях автосигнализаций.
Устройство и компоненты современных автомобильных сигнализаций
Современная автомобильная сигнализация представляет собой сложный электронный комплекс, состоящий из нескольких основных компонентов:
- Центральный блок управления — «мозг» всей системы
- Датчики (удара, объема, наклона и др.)
- Сирена
- Брелоки-передатчики
- Антенный модуль
- Реле блокировки двигателя
- Светодиодный индикатор
- Сервисная кнопка
Центральный блок управления обрабатывает сигналы от датчиков, управляет всеми функциями системы и обменивается данными с брелоками. Датчики фиксируют различные воздействия на автомобиль. Сирена служит для звуковой сигнализации. Брелоки позволяют дистанционно управлять системой. Антенный модуль обеспечивает радиосвязь между центральным блоком и брелоками.
Принцип работы автомобильной сигнализации
Работа автомобильной сигнализации основана на следующем принципе:
- При постановке на охрану активируются все датчики системы
- При срабатывании любого датчика сигнал поступает в центральный блок
- Блок управления включает сирену и отправляет оповещение на брелок
- Одновременно активируются блокировки двигателя
- Система переходит в режим тревоги до снятия с охраны
Современные сигнализации имеют множество дополнительных функций — автозапуск двигателя, GSM-оповещение, управление через смартфон и др. Но базовый принцип работы остается неизменным.
Схема подключения автосигнализации в автомобиле
Правильное подключение автосигнализации — ответственный процесс, требующий знания электрооборудования конкретного автомобиля. Типовая схема подключения включает следующие этапы:
- Установка центрального блока в салоне автомобиля
- Подключение питания от аккумулятора
- Подсоединение к штатной проводке автомобиля (двери, капот, багажник)
- Установка и подключение датчиков
- Монтаж сирены в подкапотном пространстве
- Подключение цепей блокировки двигателя
- Установка сервисной кнопки и светодиода
- Монтаж антенного модуля
Важно использовать качественные материалы и надежно изолировать все соединения. Неправильное подключение может привести к выходу из строя как самой сигнализации, так и штатного электрооборудования автомобиля.
Популярные схемы автомобильных сигнализаций
Рассмотрим схемы некоторых популярных моделей автосигнализаций:
Схема сигнализации Tomahawk TW-9030
Tomahawk TW-9030 — современная автосигнализация с автозапуском двигателя. Ее схема включает следующие основные компоненты:
- 6-контактный разъем для подключения питания и цепей запуска
- 14-контактный разъем для подключения датчиков и управляющих цепей
- 4-контактный разъем для подключения GSM-модуля
- Разъемы для подключения датчиков температуры и удара
Особенность данной схемы — наличие встроенного CAN-модуля для работы с цифровыми шинами современных автомобилей.
Схема сигнализации Starline A93
Starline A93 — популярная модель с диалоговым кодом и автозапуском. Ее схема отличается следующими особенностями:
- Основной 18-контактный разъем для подключения большинства цепей
- Отдельный 6-контактный разъем для цепей автозапуска
- Встроенный 2CAN+2LIN интерфейс для цифровых подключений
- Разъем для подключения GPS/ГЛОНАСС антенны
Схема Starline A93 позволяет реализовать сложные алгоритмы охраны и дистанционного управления автомобилем.
Типовые неисправности автомобильных сигнализаций
При эксплуатации автосигнализаций могут возникать различные неисправности. Наиболее распространенные из них:
- Ложные срабатывания датчиков
- Уменьшение дальности действия брелоков
- Отказ функции автозапуска двигателя
- Некорректная работа центрального замка
- Полный отказ системы
Причины неисправностей могут быть различными — от разрядки элемента питания в брелоке до выхода из строя центрального блока управления. Важно своевременно обнаруживать и устранять неполадки, чтобы обеспечить надежную защиту автомобиля.
Перспективы развития автомобильных охранных систем
Современные технологии открывают новые возможности для совершенствования автомобильных сигнализаций. Основные тенденции развития включают:
- Интеграцию с мобильными устройствами и «умным домом»
- Использование технологий машинного обучения для анализа окружения
- Применение биометрической идентификации владельца
- Расширение функций телеметрии и удаленной диагностики
- Повышение защищенности от электронного взлома
Будущие автосигнализации смогут не только защищать от угона, но и предоставлять владельцу расширенные возможности контроля и управления автомобилем.
Выбор и установка автосигнализации: на что обратить внимание
При выборе и установке автосигнализации следует учитывать несколько важных факторов:
- Совместимость с конкретной моделью автомобиля
- Набор необходимых функций (автозапуск, GSM-модуль и т.д.)
- Качество и надежность производителя
- Уровень защиты от взлома и электронного перехвата
- Удобство использования и информативность брелока
- Возможность дальнейшего расширения системы
- Наличие квалифицированных установщиков в вашем регионе
Правильно подобранная и грамотно установленная сигнализация обеспечит надежную защиту автомобиля и комфорт для владельца на долгие годы.
типовая установка сигнализации своими руками
Противоугонный комплекс Томагавк используется во многих странах мира и считается одним из самых надёжных. При эксплуатации системы не возникает непредвиденных ситуаций. Пользоваться сигнализацией несложно, так как она комплектуется руководством по эксплуатации и инструкцией, в которой есть разделы.
- Что в наборе охранной системы.
- Основные функции сигнализации.
- Установка и настройка противоугонного комплекса.
- Схема подключения Томагавк 9030.
Кроме инструкций, в руководстве размещены рисунки, чертежи, которые помогут разобраться в подсоединении и эксплуатации сигнализации.
Содержание
- Главные правила подключения
- Порядок размещения основных блоков
- Установка Tomahawk TW 9030 своими руками
- Описание и фото схем Томагавк 9030
- Технические сведения
- Комплектация системы
- Возможности сигнализации
- Видео
Вы можете скачать официальную инструкцию Tomahawk TW 9030.
- легко
- характеристики
- клаксон
- управление
- негативное
- подсветка
- схема
- брелок
- принципы
Узнайте как привязать новый брелок Томагавк 9030.
Главные правила подключения
Инструкция по установке Томагавк 9030 является основным помощником при монтаже и установке противоугонной системы. Сигнализация подсоединяется по стандартной схеме. Предварительно снимите минусовую клемму аккумулятора. При монтаже охранной системы соблюдайте следующие правила расположения компонентов.
- Место установки должно быть скрытным и труднодоступным для взломщиков.

- Не размещайте блоки сигнализации рядом с источниками тепла и влаги.
- Электрооборудование автомобиля не должно оказывать никакого влияние на компоненты противоугонного комплекса.
- Антенну установите подальше от модуля управления сигнализацией.
Порядок размещения основных блоков
Используя руководство по подключению, выполните следующие шаги.
- Установите блок управления в салоне автомобиля в скрытом и защищённом месте. Устройство закрепляется на ровной поверхности с помощью саморезов или пластиковых хомутов. Для дополнительной защиты от влаги обмотайте модуль полиэтиленовой плёнкой. Можно закрепить блок за панелью приборов, тем самым обеспечив минимальную длину проводов при подключении.
- Сирену разместите под капотом, предварительно вмонтировав кронштейн. Поставьте сирену подальше от блока цилиндров. Электрические провода протяните через отверстие в перегородке, разделяющей салон автомобиля и моторный отсек. Раструб устройства направьте вниз, для избегания скопления влаги в нём.

- Трансивер с антенной установите в салоне машины на поверхности лобового или заднего стекла. Перед монтажом обезжирьте место установки. Кабель от устройства к блоку управления проложите под декоративной пластиковой панелью. Не размещайте рядом с трансивером электронные устройства, которые могут ухудшить качество сигнала.
- Светодиод установите в салоне автомобиля на самом видном месте. Его работа должна хорошо просматриваться с улицы. Если диод устанавливается на панель, то заранее сделайте в ней отверстие соответствующего диаметра.
- Кнопку сервисного режима также установите в салоне в скрытном месте. Доступ водителя к устройству должен быть всегда свободным. Дополнительно замаскируйте кнопку, обмотав изолентой штанного цвета.
- Триггеры капота и дверей разместите в местах без водостоков. В запертом положении элементов ход штока датчика должен быть не менее пяти миллиметров.
- Датчик удара поставьте внутри салона автомобиля. Наиболее подходящее расположение – центральная часть кузова.
 Установите регулятор на ровную поверхность, закрепив пластиковыми хомутами или саморезами. Устройство должно быть доступным для последующей регулировки.
Установите регулятор на ровную поверхность, закрепив пластиковыми хомутами или саморезами. Устройство должно быть доступным для последующей регулировки.
Узнайте как найти кнопку Override на сигнализации Томагавк.
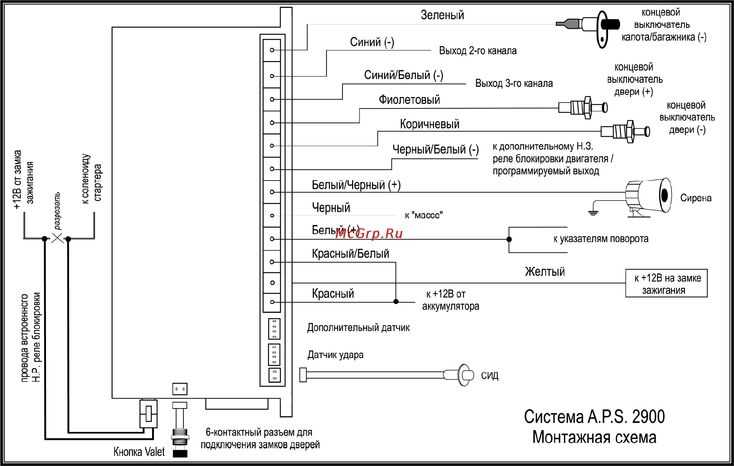
Установка Tomahawk TW 9030 своими руками: распиновка
Как подключить автосигнализацию самостоятельно? Для того чтобы подсоединить 6-ти пиновый разъём следуйте следующим пунктам.
- Красный провод – питание 12 вольт. Соедините его напрямую с аккумулятором автомобиля. Электрическую цепь защитите предохранителем на 30 ампер.
- Чёрно-жёлтым толстым проводом соедините стартёрный механизм.
- Жёлтый провод включите в соответствующий вход на замке зажигания. Напряжение 12 вольт появляется при переводе ключа в положение «АСС» и не пропадает при прокрутке в положение «Стартер».
- Синий провод — положительный контакт IGN 3 организует дополнительную линию при автоматическом пуске двигателя автомобиля.
- Зелёный провод — плюсовой выход IGN 2.
 Соедините с контактом замка зажигания, на котором образуется 12 вольт при переводе ключа в положение «АСС». В режиме «Стартер» напряжение должно исчезнуть.
Соедините с контактом замка зажигания, на котором образуется 12 вольт при переводе ключа в положение «АСС». В режиме «Стартер» напряжение должно исчезнуть. - Чёрно-жёлтый тонкий провод – положительный контакт блокировки стартера при автоматическом запуске двигателя. Соедините его с соответствующим контактом на замке зажигания.
Подключение 14-ти пинового разъёма выполните по следующей инструкции.
- Чёрно-серый провод соедините с тахометром или с датчиком давления масла.
- Чёрно-зелёный провод – положительный выход реле поворотных или габаритных огней. Установите в цепь предохранитель на 7,5 ампер.
- Чёрный – это минус или масса. Подключите провод к заземлению. Используйте любой штатный болт, вкрученный в кузов.
- Жёлто-зелёный провод – положительный выход реле «поворотников» или «габаритов». Установите предохранитель на 7,5 ампер.
- Синий подключите для управления стеклоподъёмниками.
- Чёрно-красный провод – минусовой выход служит для обхода штанного блокиратора при удалённом запуске двигателя.

- Чёрно-жёлтый провод – отрицательный контакт на реле блокировки.
- Серый плюсовой используйте для подключения рулевого сигнала и сирены.
- Оранжево-фиолетовый соедините с отрицательным сигналом ручного тормоза, стоповыми огнями (автомобиль с автоматической коробкой).
- Сине-красный провод служит для управления триггеров дверей.
- Оранжево-белый и оранжево-серый – минусовые выходы концевых выключателей дверей. Багажника и капота.
Описание и фото схем Томагавк 9030
Ниже представлена схема блока Tomahawk TW 9030, распиновка.
- управление
- схема
- обозначения
- электропривод
- негативное
- подсветка
- организация
- блокировка
- легко
- клаксон
Узнайте почему сигнализация Томагавк не открывает двери с брелка.
Технические сведения
Автосигнализация Томагавк является универсальной и может устанавливаться на все марки автомобилей: на ВАЗ 2114, Раум 97 года, Форд, Шевроле и многие другие. Противоугонный комплекс отличается следующими техническими характеристиками:
- напряжение питания составляет 12 вольт;
- при выключенном зажигании система потребляет не более 16 мАч;
- передача данных осуществляется при помощи радиоканала на 434 МГц;
- коммутируемая связь – FM;
- питание брелока сигнализации поступает от батареек класса ААА на 1,5 В.
Комплектация системы
Противоугонка Tomahawk 9030 состоит из:
- микропроцессорного модуля управления, размещённого в прямоугольном чёрном корпусе;
- сервисного руководства по монтажу и использованию;
- основного пульта управления с жидкокристаллическим дисплеем. Запасного брелока без экрана;
- двухзонного датчика чувствительности;
- светодиодной лампочки состояния охранной системы;
- набора проводов для монтажа и соединения всех элементов;
- концевого переключателя, устанавливаемого на капот или багажник;
- гарантийного талона;
- упаковки.

Возможности сигнализации
Tomahawk 9030 обладает следующими функциями.
- Автоматическим запуском двигателя по температуре или по команде.
- Установкой дополнительных триггеров на двери автомобиля и багажное отделение.
- Наличием иммобилайзера, производящим блокировку двигателя машины при несанкционированном запуске.
- Охраной автомобиля с заведённым мотором.
- Дистанционная настройка датчика удара и чувствительности.
- Наличием встроенного температурного датчика в блоке управления. Что позволяет владельцу контролировать температуру салона.
- Настройки сигналов центрального замка.
- Режим «Комфорт» — двери закрываются по истечении тридцати секунд после отправки команды с брелока.
- «Паника», привлекающего внимание людей и отпугивающего злоумышленников.
- Наличием режима «Валет».
- Тихой охраной автомобиля. При взломе не срабатывает сигнализация и световые приборы, информация отправляется не брелок.

- Программированием до четырёх пейджеров.
- Автоматическим запиранием дверных замков при начале движения автомобиля.
- Наличием режима «Турботаймер». Турбина работает определённое время после выключения зажигания.
- Подсветка экрана пульта дистанционного управления.
- Встроенными часами, отображающими текущее время.
- Блокировкой кнопок управления коммутатора, предотвращающее случайное нажатие.
Видео
Новинки авто > Автосигнализации >
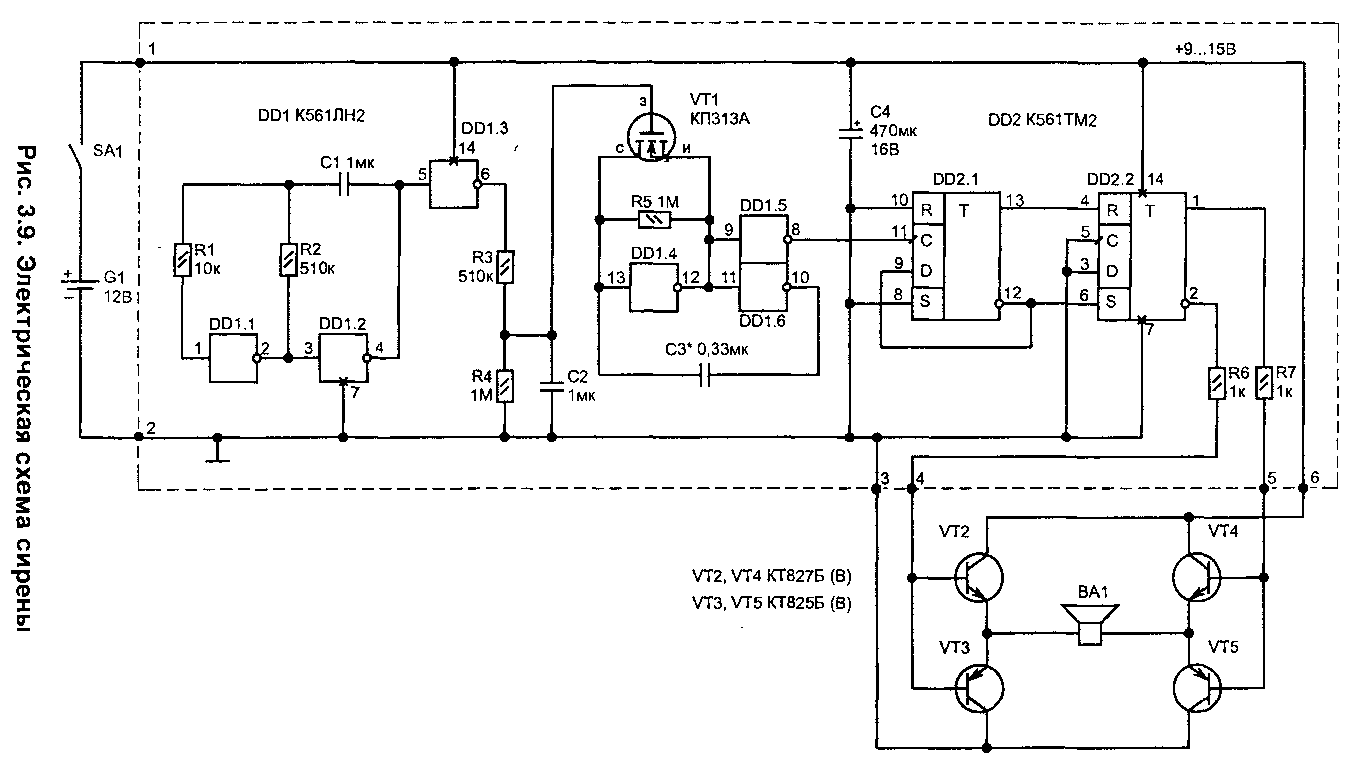
Схема сигнализации на микросхеме К176ИЕ12 » Паятель.Ру
Категория: Автомобильные устройства / Сигнализации
Данная сигнализация реагирует на контактные датчики, в качестве которых используются имеющиеся в автомобиле дверные выключатели внутрисалонного освещения. При открывании двери звуковой сигнал автомобиля начинает издавать короткие гудки. Это продолжается 19,5 секунд, плюс то время, в течение которого дверь автомобиля остается открытой.
Это продолжается 19,5 секунд, плюс то время, в течение которого дверь автомобиля остается открытой.
Чтобы поставить машину на охрану нужно перед выходом из салона включить питание сигнализации с помощью потайного выключателя, спрятанного в салоне. При этом, индикаторный светодиод загорается зеленым светом. Пока светодиод зеленый сигнализация не реагирует на датчики.
Время в течение которого сигнализация находится в таком состоянии составляет не менее 19,5 секунд, но может быть очень долгим, если вы откроете одну из дверей. В этом случае отсчет 19,5 секунд начнется только после закрывания всех дверей.
Таким образом, вы можете неспешно выйти из машины, выгрузить весь багаж, и т.д. И только спустя 19,5 секунд после закрывания всех дверей система станет на охрану. Это будет подтверждено покраснением индикаторного светодиода.
Чтобы снять машину с охраны нужно иметь магнитный ключ, представляющий собой цилиндрический брелок для ключей с расположенным в нем постоянным магнитом. В известном только вам месте остекления автомобиля нужно установить геркон, со стороны из салона. Этот брелок нужно поднести к тому месту стекла, за которым расположен геркон.
В известном только вам месте остекления автомобиля нужно установить геркон, со стороны из салона. Этот брелок нужно поднести к тому месту стекла, за которым расположен геркон.
При поднесении магнита к геркону светодиод меняет цвет на зеленый. Это значит, что вы можете открыть дверь машины не вызывая срабатывания сигнала. Открыв дверь, вы, уже находясь внутри, отключаете питание сигнализации потайным выключателем. Светодиод гаснет. Можно ехать.
Принципиальная схема показана на рисунке. Микросхема D1 — К176ИЕ12 обычно используется в электронных часах или таймерах. Она содержит кварцевый генератор и два счетчика. Частота кварцевого генератора 32768 Гц (соответственно частоте стандартного часового резонатора).
Первый счетчик, в данной схеме, делит эту частоту на 16384, и на его выходе (выв. 6) образуются импульсы с частотой 2 Гц. Второй счетчик имеет коэффициент деления 60. Но, единица на его выходе (вывод 10) появляется через 39 входных импульсов, то есть, при входной частоте 2 Гц, через 19,5 секунд.
На элементах D2.1 и D2.2 сделан RS-триггер, с помощью которого сигнализацию снимают с охраны и ставят на охрану.
Рассмотрим работу схемы. Потайной выключатель — это S1. В момент его включения на схему подается питание от автомобильного аккумулятора. В этот момент С4 устанавливает RS-триггер D2.1-D2.2 в состояние когда на выходе D2.1 ноль, а на выходе D2.2 — единица. Горит зеленая половина светодиода HL1.
Единица поступает на один из входов элемента D2.4 и транзисторный ключ VT4-VT5 фиксируется закрытым. В этот же момент С1 устанавливает второй счетчик D1 в нулевое состояние. Импульсы частотой 2 Гц с выхода первого счетчика поступают на вход второго счетчика (выв. 7).
Через 19,5 секунд на выходе второго счетчика (выв. 10) появляется логическая единица, которая делает две вещи, — останавливает первый счетчик (единица на выв. 5) и перекидывает RS-триггер D2.1-D2.2 в противоположное состояние. Загорается красная половина светодиода HL1, и на вывод 12 D2.4 поступает логический ноль.
При срабатывании происходит замыкание цепи дверных выключателей, подключенных к клемме «1». Практически это выглядит как замыкание данной клеммы на общий минус. Транзистор VT1 в этот момент открывается и подает на вывод 9 D1 напряжение логической единицы. Второй счетчик обнуляется и на выводе 5 D1 устанавливается логический ноль.
Теперь первый счетчик работает и на его выходе (выв. 6) возникают логические импульсы, которые проходят через D2.3-D2.4 на транзисторный ключ VT4-VT5. Ключ периодически открывается, и периодически включает реле звукового сигнала автомобиля, к которому он подключен. Раздаются гудки звукового сигнала.
Эти гудки будут повторяться до тех пор, пока на выводе 10 D1 не появится логическая единица. Для того чтобы она появилась второй счетчик микросхемы D1 должен считать импульсы, поступающие на его вход. Но для счета нужно чтобы на выводе 9 был логический ноль. То есть, двери должны быть закрытыми. Поэтому, гудки повторяются все время пока дверь открыта, плюс, 19,5 секунд, пока счетчик будет считать импульсы.
Чтобы заблокировать сигнализацию используется геркон SG1. Поднесите к нему магнит, и его контакты замыкаются. Это равносильно зарядному току С4, который в момент включения предустанавливает RS-триггер.
Таким образом, после замыкания геркона схема переходит в такое же состояние, как и после включения питания. То есть, есть время, чтобы открыть дверь и спокойно выключить сигнализацию потайным выключателем.
Диод VD1 вместе с транзистором VT1 не только инвертирует сигнал от датчика, но и предохраняет схему от выхода из строя от выбросов напряжения, которые могут быть в автомобильной бортовой сети. Диод VD2 исключает выход схемы из строя из-за неправильного подключения по полярности питания.
Диод VD3 — защищает транзисторы VT4-VT5 от выхода из строя от выбросов ЭДС самоиндукции обмотки реле звукового сигнала. Микросхемы питаются напряжением 9V от интегрального стабилизатора А1.
Пути передачи сигнала Wnt — PMC
1. Wodarz A, Nusse R. Механизмы передачи сигналов Wnt в развитии. Annu Rev Cell Dev Biol. 1998; 14:59–88. [PubMed] [Google Scholar]
Annu Rev Cell Dev Biol. 1998; 14:59–88. [PubMed] [Google Scholar]
2. Хабас Р., Давид И.Б. Disheveled и передача сигналов Wnt: является ли ядро последней границей? Дж. Биол. 2005; 4:2. [Бесплатная статья PMC] [PubMed] [Google Scholar]
3. Yamaguchi TP. Головы или хвосты: Wnts и передне-задний рисунок. Карр Биол. 2001; 11: 713–724. [PubMed] [Академия Google]
4. Logan CY, Nusse R. Сигнальный путь Wnt в развитии и заболевании. Annu Rev Cell Dev Biol. 2004 [PubMed] [Google Scholar]
5. He X, Semenov M, Tamai K, Zeng X. Белки 5 и 6, связанные с рецептором ЛПНП, в передаче сигналов Wnt/бета-катенина: стрелки указывают путь. Разработка. 2004; 131:1663–1677. [PubMed] [Google Scholar]
6. Schulte G, Bryja V. Семейство Frizzled нетрадиционных рецепторов, связанных с G-белком. Trends Pharmacol Sci. 2007; 28: 518–525. [PubMed] [Академия Google]
7. Кадиган К.М., Лю Ю.И. Сигнализация Wnt: сложность на поверхности. Дж. Клеточные науки. 2006; 119: 395–402. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
8. Уоллингфорд Дж. Б., Хабас Р. Биология развития растрепанных: загадочный белок, определяющий клеточную судьбу и клеточную полярность. Разработка. 2005; 132:4421–4436. [PubMed] [Google Scholar]
9. Смолич Б.Д., МакМахон Дж.А., МакМахон А.П., Папкофф Дж. Белки семейства Wnt секретируются и связываются с клеточной поверхностью. Мол Биол Селл. 1993; 4: 1267–1275. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Willert K, Brown JD, Danenberg E, Duncan AW, Weissman IL, Reya T, Yates JR, 3rd, Nusse R. Белки Wnt модифицированы липидами и могут действовать как факторы роста стволовых клеток. Природа. 2003; 423:448–452. [PubMed] [Google Scholar]
11. Banziger C, Soldini D, Schutt C, Zipperlen P, Hausmann G, Basler K. Wntless, консервативный мембранный белок, предназначенный для секреции белков Wnt из сигнальных клеток. Клетка. 2006; 125: 509–522. [PubMed] [Google Scholar]
12. Bartscherer K, Pelte N, Ingelfinger D, Boutros M. Для секреции лигандов Wnt требуется Evi, консервативный трансмембранный белок. Клетка. 2006; 125: 523–533. [PubMed] [Академия Google]
Для секреции лигандов Wnt требуется Evi, консервативный трансмембранный белок. Клетка. 2006; 125: 523–533. [PubMed] [Академия Google]
13. Hausmann G, Banziger C, Basler K. Помощь Wingless в полете: как секретируются белки WNT. Nat Rev Mol Cell Biol. 2007; 8: 331–336. [PubMed] [Google Scholar]
14. Хофманн К. Надсемейство мембраносвязанных O-ацилтрансфераз с последствиями для передачи сигналов wnt. Тенденции биохимических наук. 2000; 25:111–112. [PubMed] [Google Scholar]
15. Lin X. Функции протеогликанов сульфата гепарана в передаче сигналов клетки во время развития. Разработка. 2004; 131:6009–6021. [PubMed] [Академия Google]
16. Glinka A, Wu W, Delius H, Monaghan AP, Blumenstock C, Niehrs C. Dickkopf-1 является членом нового семейства секретируемых белков и участвует в индукции головы. Природа. 1998; 391:357–362. [PubMed] [Google Scholar]
17. Hsieh JC, Kodjabachian L, Rebbert ML, Rattner A, Smallwood PM, Samos CH, Nusse R, Dawid IB, Nathans J. Новый секретируемый белок, который связывается с белками Wnt и ингибирует их деятельность. Природа. 1999; 398:431–436. [PubMed] [Google Scholar]
Новый секретируемый белок, который связывается с белками Wnt и ингибирует их деятельность. Природа. 1999; 398:431–436. [PubMed] [Google Scholar]
18. Хоанг Б.Х., Томас Дж.Т., Абдул-Карим Ф.В., Коррейя К.М., Конлон Р.А., Луйтен Ф.П., Баллок Р.Т. Паттерн экспрессии двух родственных Frizzled генов, Frzb-1 и Sfrp-1, во время эмбриогенеза мышей указывает на роль модулирующего действия членов семейства Wnt. Дев Дин. 1998;212:364–372. [PubMed] [Google Scholar]
19. Боумистер Т., Ким С., Сасаи Ю., Лу Б., Де Робертис Э.М. Cerberus представляет собой головной индуцирующий секретируемый фактор, экспрессируемый в передней энтодерме организатора Шпемана. Природа. 1996; 382: 595–601. [PubMed] [Google Scholar]
20. Wang S, Krinks M, Lin K, Luyten FP, Moos M., Jr Frzb, секретируемый белок, экспрессируемый в организаторе Шпемана, связывает и ингибирует Wnt-8. Клетка. 1997; 88: 757–766. [PubMed] [Google Scholar]
21. Itasaki N, Jones CM, Mercurio S, Rowe A, Domingos PM, Smith JC, Krumlauf R. Wise, контекстно-зависимый активатор и ингибитор передачи сигналов Wnt. Разработка. 2003;130:4295–4305. [PubMed] [Google Scholar]
Wise, контекстно-зависимый активатор и ингибитор передачи сигналов Wnt. Разработка. 2003;130:4295–4305. [PubMed] [Google Scholar]
22. Kawano Y, Kypta R. Секретируемые антагонисты сигнального пути Wnt. Дж. Клеточные науки. 2003; 116: 2627–2634. [PubMed] [Google Scholar]
23. Xu Q, Wang Y, Dabdoub A, Smallwood PM, Williams J, Woods C, Kelley MW, Jiang L, Tasman W, Zhang K, Nathans J. Развитие сосудов сетчатки и внутреннее ухо: контроль с помощью Norrin и Frizzled-4, пары лиганд-рецептор с высоким сродством. Клетка. 2004; 116: 883–895. [PubMed] [Google Scholar]
24. Казанская О., Глинка А., дель Барко Баррантес И., Станнек П., Нихрс С., Ву В. R-Spondin2 является секретируемым активатором передачи сигналов Wnt/бета-катенин и необходим для Xenopus. миогенез. Ячейка Дев. 2004; 7: 525–534. [PubMed] [Академия Google]
25. Semenov M, Tamai K, He X. SOST является лигандом для LRP5/LRP6 и ингибитором передачи сигналов Wnt. Дж. Биол. Хим. 2005; 280:26770–26775. [PubMed] [Google Scholar]
26. Gordon MD, Nusse R. Передача сигналов Wnt: несколько путей, несколько рецепторов и несколько факторов транскрипции. Дж. Биол. Хим. 2006; 281:22429–22433. [PubMed] [Google Scholar]
Gordon MD, Nusse R. Передача сигналов Wnt: несколько путей, несколько рецепторов и несколько факторов транскрипции. Дж. Биол. Хим. 2006; 281:22429–22433. [PubMed] [Google Scholar]
27. Mao J, Wang J, Liu B, Pan W, Farr GH, 3rd, Flynn C, Yuan H, Takada S, Kimelman D, Li L, Wu D. Липопротеины низкой плотности родственный рецептору белок-5 связывается с Axin и регулирует канонический сигнальный путь Wnt. Мол Ячейка. 2001; 7: 801–809.. [PubMed] [Google Scholar]
28. Zeng X, Huang H, Tamai K, Zhang X, Harada Y, Yokota C, Almeida K, Wang J, Doble B, Woodgett J, Wynshaw-Boris A, Hsieh JC, He X. Инициация передачи сигналов Wnt: контроль фосфорилирования/активации корецептора Wnt Lrp6 посредством функций взъерошенных, растрепанных и аксинов. Разработка. 2008; 135: 367–375. [PMC free article] [PubMed] [Google Scholar]
29. Tolwinski NS, Wehrli M, Rives A, Erdeniz N, DiNardo S, Wieschaus E. Сигнал Wg/Wnt может передаваться через arrow/LRP5,6 и Axin независимо активности Zw3/Gsk3beta. Ячейка Дев. 2003; 4: 407–418. [PubMed] [Академия Google]
Ячейка Дев. 2003; 4: 407–418. [PubMed] [Академия Google]
30. Willert K, Logan CY, Arora A, Fish M, Nusse R. Гомолог Axin дрозофилы, Daxin, ингибирует передачу сигналов Wnt. Разработка. 1999; 126:4165–4173. [PubMed] [Google Scholar]
31. Yamamoto H, Kishida S, Kishida M, Ikeda S, Takada S, Kikuchi A. Фосфорилирование аксина, негативного регулятора сигнала Wnt, киназой-3бета гликогенсинтазы регулирует его стабильность. Дж. Биол. Хим. 1999; 274:10681–10684. [PubMed] [Google Scholar]
32. Chen HJ, Lin CM, Lin CS, Perez-Olle R, Leung CL, Liem RK. Роль актинового сшивающего фактора 1 микротрубочек (MACF1) в сигнальном пути Wnt. Гены Дев. 2006;20:1933–1945. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Хабас Р. Каноническая сигнализация Wnt: неожиданный новый игрок. Ячейка Дев. 2006; 11: 138–139. [PubMed] [Google Scholar]
34. Kishida S, Yamamoto H, Hino S, Ikeda S, Kishida M, Kikuchi A. Домены DIX Dvl и аксина необходимы для белковых взаимодействий и их способности регулировать стабильность бета-катенина. Мол Селл Биол. 1999;19:4414–4422. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мол Селл Биол. 1999;19:4414–4422. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Wharton KA., Jr Runnin ‘с Dvl: белки, которые связаны с Dsh/Dvl, и их значение для передачи сигнала Wnt. Дев биол. 2003; 253:1–17. [PubMed] [Академия Google]
36. Wong HC, Bourdelas A, Krauss A, Lee HJ, Shao Y, Wu D, Mlodzik M, Shi DL, Zheng J. Прямое связывание домена PDZ Disheveled с консервативной внутренней последовательностью в C-концевой области из Фризлд. Мол Ячейка. 2003; 12:1251–1260. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Peters JM, McKay RM, McKay JP, Graff JM. Казеинкиназа I передает сигналы Wnt. Природа. 1999; 401:345–350. [PubMed] [Google Scholar]
38. Willert K, Brink M, Wodarz A, Varmus H, Nusse R. Казеинкиназа 2 связывается с растрепанными и фосфорилирует их. EMBO J. 1997;16:3089–3096. [бесплатная статья PMC] [PubMed] [Google Scholar]
39. Кибардин А., Осипова О., Сокол С.Ю. Киназа, ассоциированная с метастазами, модулирует передачу сигналов Wnt, чтобы регулировать формирование паттерна и морфогенез мозга. Разработка. 2006; 133: 2845–2854. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Разработка. 2006; 133: 2845–2854. [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Chen W, ten Berge D, Brown J, Ahn S, Hu LA, Miller WE, Caron MG, Barak LS, Nusse R, Lefkowitz RJ. Disheveled 2 рекрутирует бета-аррестин 2 для опосредования Wnt5A-стимулированного эндоцитоза Frizzled 4. Наука. 2003;301:1391–1394. [PubMed] [Google Scholar]
41. Осипова О., Дхаван С., Сокол С., Грин Дж. Б. Отдельные белки PAR-1 функционируют в разных ветвях передачи сигналов Wnt во время развития позвоночных. Ячейка Дев. 2005; 8: 829–841. [PubMed] [Google Scholar]
42. Hatsell S, Rowlands T, Hiremath M, Cowin P. Бета-катенин и Tcfs в развитии молочной железы и раке. J Биол. неоплазия молочной железы. 2003; 8: 145–158. [PubMed] [Google Scholar]
43. Fagotto F, Gluck U, Gumbiner BM. Независимый от сигнала ядерной локализации и независимый от импортина/кариоферина ядерный импорт бета-катенина. Карр Биол. 1998;8:181–190. [PubMed] [Google Scholar]
44. Конг Ф., Вармус Х. Ядерно-цитоплазматический перенос аксина регулирует субклеточную локализацию бета-катенина. Proc Natl Acad Sci USA. 2004; 101: 2882–2887. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ядерно-цитоплазматический перенос аксина регулирует субклеточную локализацию бета-катенина. Proc Natl Acad Sci USA. 2004; 101: 2882–2887. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Schwarz-Romond T., Metcalfe C., Bienz M. Динамическое рекрутирование аксина с помощью белковых сборок Disheveled. Дж. Клеточные науки. 2007; 120:2402–2412. [PubMed] [Google Scholar]
46. Hendriksen J, Fagotto F, van der Velde H, van Schie M, Noordermeer J, Fornerod M. RanBP3 увеличивает ядерный экспорт активного (бета)-катенина независимо от CRM1. Джей Селл Биол. 2005; 171: 785–79.7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Clevers H. Передача сигналов Wnt/бета-катенина в развитии и заболевании. Клетка. 2006; 127: 469–480. [PubMed] [Google Scholar]
48. Reya T, Clevers H. Передача сигналов Wnt в стволовых клетках и раке. Природа. 2005; 434: 843–850. [PubMed] [Google Scholar]
49. Харланд Р., Герхарт Дж. Формирование и функция организатора Шпемана. Annu Rev Cell Dev Biol. 1997; 13: 611–667. [PubMed] [Google Scholar]
Annu Rev Cell Dev Biol. 1997; 13: 611–667. [PubMed] [Google Scholar]
50. Kramps T, Peter O, Brunner E, Nellen D, Froesch B, Chatterjee S, Murone M, Zullig S, Basler K. Для передачи сигналов Wnt/wingless требуется BCL9/legless-опосредованное рекрутирование pygopus к ядерному комплексу бета-катенин-TCF. Клетка. 2002; 109:47–60. [PubMed] [Google Scholar]
51. Паркер Д.С., Джемисон Дж., Кэдиган К.М. Pygopus, ядерный белок PHD-finger, необходимый для передачи сигналов Wingless у дрозофилы. Разработка. 2002; 129: 2565–2576. [PubMed] [Google Scholar]
52. Thompson B, Townsley F, Rosin-Arbesfeld R, Musisi H, Bienz M. Новый ядерный компонент сигнального пути Wnt. Nat Cell Biol. 2002; 4: 367–373. [PubMed] [Академия Google]
53. Townsley FM, Cliffe A, Bienz M. Pygopus и Legless нацеливают Armadillo/бета-катенин на ядро, чтобы обеспечить его транскрипционную функцию коактиватора. Nat Cell Biol. 2004; 6: 626–633. [PubMed] [Google Scholar]
54. Jessen JR, Solnica-Krezel L. Формирование оси — бета-катенин захватывает Wnt. Клетка. 2005; 120:736–737. [PubMed] [Google Scholar]
Формирование оси — бета-катенин захватывает Wnt. Клетка. 2005; 120:736–737. [PubMed] [Google Scholar]
55. Tao Q, Yokota C, Puck H, Kofron M, Birsoy B, Yan D, Asashima M, Wylie CC, Lin X, Heasman J. Материнский wnt11 активирует требуемый канонический сигнальный путь wnt для формирования оси у эмбрионов Xenopus. Клетка. 2005; 120:857–871. [PubMed] [Академия Google]
56. De Robertis EM, Larrain J, Oelgeschlager M, Wessely O. Установление организатора Шпемана и формирование паттерна эмбриона позвоночного. Нат Рев Жене. 2000; 1: 171–181. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. De Robertis EM. Организатор Шпемана и саморегуляция у зародышей амфибий. Nat Rev Mol Cell Biol. 2006; 7: 296–302. [Бесплатная статья PMC] [PubMed] [Google Scholar]
58. Де Робертис Э.М., Курода Х. Формирование дорсально-вентрального паттерна и индукция нейронов у эмбрионов Xenopus. Annu Rev Cell Dev Biol. 2004; 20: 285–308. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Seifert JR, Mlodzik M. Передача сигналов Frizzled/PCP: законсервированный механизм, регулирующий клеточную полярность и направленную подвижность. Нат Рев Жене. 2007; 8: 126–138. [PubMed] [Google Scholar]
Seifert JR, Mlodzik M. Передача сигналов Frizzled/PCP: законсервированный механизм, регулирующий клеточную полярность и направленную подвижность. Нат Рев Жене. 2007; 8: 126–138. [PubMed] [Google Scholar]
60. Mlodzik M. Плоская поляризация клеток: одни и те же механизмы регулируют полярность тканей дрозофилы и гаструляцию позвоночных? Тенденции Жене. 2002; 18: 564–571. [PubMed] [Google Scholar]
61. Wang Y, Nathans J. Полярность тканей и плоскостных клеток у позвоночных: новые идеи и новые вопросы. Разработка. 2007; 134: 647–658. [PubMed] [Академия Google]
62. Уоллингфорд Дж. Б., Фрейзер С. Е., Харланд Р. М. Конвергентное расширение: молекулярный контроль движения поляризованных клеток во время эмбрионального развития. Ячейка Дев. 2002; 2: 695–706. [PubMed] [Google Scholar]
63. Келлер Р., Дэвидсон Л.А., Шук Д.Р. Как мы сформированы: биомеханика гаструляции. Дифференциация. 2003; 71: 171–205. [PubMed] [Google Scholar]
64. Veeman MT, Axelrod JD, Moon RT. Второй канон. Функции и механизмы независимой от бета-катенина передачи сигналов Wnt. Ячейка Дев. 2003; 5: 367–377. [PubMed] [Академия Google]
Второй канон. Функции и механизмы независимой от бета-катенина передачи сигналов Wnt. Ячейка Дев. 2003; 5: 367–377. [PubMed] [Академия Google]
65. Djiane A, Riou J, Umbhauer M, Boucaut J, Shi D. Роль frizzled 7 в регуляции конвергентных разгибательных движений во время гаструляции у Xenopus laevis. Разработка. 2000;127:3091–3100. [PubMed] [Google Scholar]
66. Habas R, Dawid IB, He X. Коактивация Rac и Rho посредством передачи сигналов Wnt/Frizzled необходима для гаструляции позвоночных. Гены Дев. 2003; 17: 295–309. [Бесплатная статья PMC] [PubMed] [Google Scholar]
67. Habas R, Kato Y, He X. Активация Rho Wnt/Frizzled регулирует гаструляцию позвоночных и требует нового белка Daam1, гомологичного формину. Клетка. 2001; 107: 843–854. [PubMed] [Академия Google]
68. Парк М, Луна РТ. Ген планарной клеточной полярности stbm регулирует клеточное поведение и клеточную судьбу у эмбрионов позвоночных. Nat Cell Biol. 2002; 4: 20–25. [PubMed] [Google Scholar]
69. Сокол С.Ю. Анализ сигнальных путей Disheveled во время развития Xenopus. Карр Биол. 1996; 6: 1456–1467. [PubMed] [Google Scholar]
Сокол С.Ю. Анализ сигнальных путей Disheveled во время развития Xenopus. Карр Биол. 1996; 6: 1456–1467. [PubMed] [Google Scholar]
70. Tahinci E, Symes K. Различные функции Rho и Rac необходимы для конвергентного расширения во время гаструляции Xenopus. Дев биол. 2003; 259: 318–335. [PubMed] [Академия Google]
71. Takeuchi M, Nakabayashi J, Sakaguchi T, Yamamoto TS, Takahashi H, Takeda H, Ueno N. Родственный колючкам ген у позвоночных необходим для движения клеток гаструляции. Карр Биол. 2003; 13: 674–679. [PubMed] [Google Scholar]
72. Veeman MT, Slusarski DC, Kaykas A, Louie SH, Moon RT. Колючка рыбок данио, модулятор неканонической передачи сигналов Wnt/Fz, регулирует гаструляционные движения. Карр Биол. 2003; 13: 680–685. [PubMed] [Google Scholar]
73. Jessen JR, Topczewski J, Bingham S, Sepich DS, Marlow F, Chandrasekhar A, Solnica-Krezel L. Трилобит данио определяет новые роли косоглазия в гаструляции и движениях нейронов. Nat Cell Biol. 2002; 4: 610–615. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
74. Carreira-Barbosa F, Concha ML, Takeuchi M, Ueno N, Wilson SW, Tada M. Prickle 1 регулирует движения клеток во время гаструляции и миграции нейронов у рыбок данио. Разработка. 2003; 130:4037–4046. [PubMed] [Google Scholar]
75. Park E, Kim GH, Choi SC, Han JK. Роль PKA как негативного регулятора сигнального пути PCP во время гаструляционных движений Xenopus. Дев биол. 2006; 292:344–457. [PubMed] [Google Scholar]
76. Tahinci E, Thorne CA, Franklin JL, Salic A, Christian KM, Lee LA, Coffey RJ, Lee E. Lrp6 требуется для конвергентного расширения во время гаструляции Xenopus. Разработка. 2007;134:4095–4106. [Бесплатная статья PMC] [PubMed] [Google Scholar]
77. Sasai N, Nakazawa Y, Haraguchi T, Sasai Y. Белок NRh2, связанный с нейротрофиновым рецептором, необходим для конвергентных разгибательных движений. Nat Cell Biol. 2004; 6: 741–748. [PubMed] [Google Scholar]
78. Lu W, Yamamoto V, Ortega B, Baltimore D. Ryk млекопитающих является корецептором Wnt, необходимым для стимуляции роста нейритов. Клетка. 2004; 119: 97–108. [PubMed] [Google Scholar]
Ryk млекопитающих является корецептором Wnt, необходимым для стимуляции роста нейритов. Клетка. 2004; 119: 97–108. [PubMed] [Google Scholar]
79. Lu X, Borchers AG, Jolicoeur C, Rayburn H, Baker JC, Tessier-Lavigne M. PTK7/CCK-4 — новый регулятор плоскостной клеточной полярности у позвоночных. Природа. 2004;430:93–98. [PubMed] [Google Scholar]
80. Nishita M, Yoo SK, Nomachi A, Kani S, Sougawa N, Ohta Y, Takada S, Kikuchi A, Minami Y. Образование филоподий, опосредованное рецепторной тирозинкиназой Ror2, необходимо для Wnt5a. индуцированная миграция клеток. Джей Селл Биол. 2006; 175: 555–562. [Бесплатная статья PMC] [PubMed] [Google Scholar]
81. Танегасима К., Чжао Х., Давид И.Б. WGEF активирует Rho в пути Wnt-PCP и контролирует конвергентное расширение при гаструляции Xenopus. EMBO J. 2008; 27: 606–617. [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Marlow F, Topczewski J, Sepich D, Solnica-Krezel L. Rho-киназа 2 рыбок данио действует ниже Wnt11, опосредуя клеточную полярность и эффективное движение конвергенции и растяжения. Карр Биол. 2002; 12: 876–884. [PubMed] [Google Scholar]
Карр Биол. 2002; 12: 876–884. [PubMed] [Google Scholar]
83. Weiser DC, Pyati UJ, Kimelman D. Gravin регулирует изменения поведения мезодермальных клеток, необходимые для удлинения оси во время гаструляции рыбок данио. Гены Дев. 2007; 21: 1559–1571. [Бесплатная статья PMC] [PubMed] [Google Scholar]
84. Li L, Yuan H, Xie W, Mao J, Caruso AM, McMahon A, Sussman DJ, Wu D. Растрепанные белки ведут к двум сигнальным путям. Регуляция LEF-1 и N-концевой киназы c-Jun в клетках млекопитающих. Дж. Биол. Хим. 1999;274:129–134. [PubMed] [Google Scholar]
85. Solnica-Krezel L. Законсервированные паттерны движения клеток во время гаструляции позвоночных. Карр Биол. 2005; 15: 213–228. [PubMed] [Google Scholar]
86. Wallingford JB, Harland RM. Передача сигналов Xenopus Disheveled регулирует как нейральное, так и мезодермальное конвергентное расширение: параллельные силы, удлиняющие ось тела. Разработка. 2001; 128: 2581–2592. [PubMed] [Google Scholar]
87. Itoh K, Brott BK, Bae GU, Ratcliffe MJ, Sokol SY. Ядерная локализация необходима для функции Disheveled в передаче сигналов Wnt/β-catenin. Дж. Биол. 2005;4:3. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ядерная локализация необходима для функции Disheveled в передаче сигналов Wnt/β-catenin. Дж. Биол. 2005;4:3. [Бесплатная статья PMC] [PubMed] [Google Scholar]
88. Парк Т.Дж., Грей Р.С., Сато А., Хабас Р., Уоллингфорд Д.Б. Субклеточная локализация и сигнальные свойства взъерошенных в развивающихся эмбрионах позвоночных. Карр Биол. 2005; 15:1039–1044. [PubMed] [Google Scholar]
89. Heisenberg CP, Tada M, Rauch GJ, Saude L, Concha ML, Geisler R, Stemple DL, Smith JC, Wilson SW. Silberblick/Wnt11 опосредует конвергентные движения растяжения во время гаструляции рыбок данио. Природа. 2000;405:76–81. [PubMed] [Google Scholar]
90. Kohn AD, Moon RT. Передача сигналов Wnt и кальция: пути, независимые от бета-катенина. Клеточный кальций. 2005;38:439–446. [PubMed] [Google Scholar]
91. Slusarski DC, Pelegri F. Передача сигналов кальция в формировании эмбрионального паттерна и морфогенезе позвоночных. Дев биол. 2007; 307:1–13. [Бесплатная статья PMC] [PubMed] [Google Scholar]
92. Gilland E, Miller AL, Karplus E, Baker R, Webb SE. Визуализация многоклеточных крупномасштабных ритмичных кальциевых волн во время гаструляции рыбок данио. Proc Natl Acad Sci USA. 1999; 96: 157–161. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Gilland E, Miller AL, Karplus E, Baker R, Webb SE. Визуализация многоклеточных крупномасштабных ритмичных кальциевых волн во время гаструляции рыбок данио. Proc Natl Acad Sci USA. 1999; 96: 157–161. [Бесплатная статья PMC] [PubMed] [Google Scholar]
93. Wallingford JB, Ewald AJ, Harland RM, Fraser SE. Передача сигналов кальция во время конвергентного расширения у Xenopus. Карр Биол. 2001; 11: 652–661. [PubMed] [Академия Google]
94. Слюсарский Д.С., Корцес В.Г., Мун Р.Т. Взаимодействие Wnt и гомолога Frizzled запускает передачу сигналов фосфатидилинозитола, связанную с G-белком. Природа. 1997; 390:410–413. [PubMed] [Google Scholar]
95. Slusarski DC, Yang-Snyder J, Busa WB, Moon RT. Модуляция эмбриональной внутриклеточной передачи сигналов Ca 2+ с помощью Wnt-5A. Дев биол. 1997; 182:114–120. [PubMed] [Google Scholar]
96. Sheldahl LC, Slusarski DC, Pandur P, Miller JR, Kuhl M, Moon RT. Взлохмаченный активирует Ca 2+ flux, PKC и CamKII у эмбрионов позвоночных. Джей Селл Биол. 2003; 161: 769–777. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Джей Селл Биол. 2003; 161: 769–777. [Бесплатная статья PMC] [PubMed] [Google Scholar]
97. Kuhl M, Sheldahl LC, Malbon CC, Moon RT. Ca(2+)/calmodulin-зависимая протеинкиназа II стимулируется гомологами Wnt и Frizzled и способствует судьбе вентральных клеток у Xenopus. Дж. Биол. Хим. 2000; 275:12701–12711. [PubMed] [Google Scholar]
98. Saneyoshi T, Kume S, Amasaki Y, Mikoshiba K. Путь Wnt/кальций активирует NF-AT и способствует судьбе вентральных клеток у эмбрионов Xenopus. Природа. 2002;417:295–299. [PubMed] [Google Scholar]
99. Ишитани Т., Ниномия-Цудзи Дж., Нагаи С., Нишита М., Менегини М., Баркер Н., Уотерман М., Бауэрман Б., Клеверс Х., Сибуя Х., Мацумото К. TAK1-NLK Связанный с -MAPK путь противодействует передаче сигналов между бета-катенином и фактором транскрипции TCF. Природа. 1999; 399: 798–802. [PubMed] [Google Scholar]
100. Winklbauer R, Medina A, Swain RK, Steinbeisser H. Передача сигналов Frizzled-7 контролирует разделение тканей во время гаструляции Xenopus. Природа. 2001; 413: 856–860. [PubMed] [Академия Google]
Природа. 2001; 413: 856–860. [PubMed] [Академия Google]
101. Вестфолл Т.А., Бримейер Р., Тведт Дж., Гладон Дж., Олбердинг А., Фурутани-Сейки М., Слюсарски Д.К. Wnt-5/pipettail функционирует при формировании оси позвоночных как негативный регулятор активности Wnt/beta-catenin. Джей Селл Биол. 2003; 162: 889–898. [Бесплатная статья PMC] [PubMed] [Google Scholar]
102. Salinas PC. Факторы Wnt в ремоделировании аксонов и синаптогенезе. Biochem Soc Symp. 1999; 65: 101–109. [PubMed] [Google Scholar]
103. Салинас, ПК. Модуляция цитоскелета микротрубочек: роль расходящегося канонического пути Wnt. Тенденции клеточной биологии. 2007; 17: 333–342. [PubMed] [Академия Google]
104. Schmitt AM, Shi J, Wolf AM, Lu CC, King LA, Zou Y. Передача сигналов Wnt-Ryk опосредует медиально-латеральное ретинотектальное топографическое картирование. Природа. 2006; 439:31–37. [PubMed] [Google Scholar]
105. Цзоу Ю. Передача сигналов Wnt в направлении аксонов. Тренды Нейроси. 2004; 27: 528–532. [PubMed] [Google Scholar]
106. Habas R, He X. Передача сигналов клеток: переход на Wnt-Rap. Карр Биол. 2007; 17: 474–477. [PubMed] [Google Scholar]
107. Tsai IC, Amack JD, Gao ZH, Band V, Yost HJ, Virshup DM. Путь Wnt-CKIvarepsilon-Rap1 регулирует гаструляцию путем модулирования SIPA1L1, белка, активирующего Rap GTPase. Ячейка Дев. 2007; 12: 335–347. [Бесплатная статья PMC] [PubMed] [Google Scholar]
108. Чу А.Я., Ру П.П., Бленис Дж. Помните о пробеле: Wnt садится в поезд mTORC1. Клетка. 2006; 126: 834–836. [PubMed] [Google Scholar]
109. Иноки К., Оуян Х., Чжу Т., Линдвалл С., Ван Ю., Чжан Х., Ян К., Беннетт С., Харада Ю., Станкунас К., Ван С.И., Хе Х., Макдугалд О.А., Ю М, Уильямс Б.О., Гуан К.Л. TSC2 интегрирует Wnt и энергетические сигналы посредством скоординированного фосфорилирования с помощью AMPK и GSK3, чтобы регулировать рост клеток. Клетка. 2006; 126: 955–968. [PubMed] [Google Scholar]
110. Доллар Г.Л., Вебер Ю., Млодзик М. , Сокол С.Ю. Регулирование смертельных гигантских личинок компанией Dishavelled. Природа. 2005; 437:1376–1380. [PubMed] [Академия Google]
, Сокол С.Ю. Регулирование смертельных гигантских личинок компанией Dishavelled. Природа. 2005; 437:1376–1380. [PubMed] [Академия Google]
111. Чен А.Э., Гинти Д.Д., Фан К.М. Передача сигналов протеинкиназы А через CREB контролирует миогенез, индуцированный белками Wnt. Природа. 2005; 433:317–322. [PubMed] [Google Scholar]
112. Шамбони А., Ведлих Д. Wnt-5A/Ror2 регулируют экспрессию XPAPC через альтернативный неканонический сигнальный путь. Ячейка Дев. 2007; 12: 779–792. [PubMed] [Google Scholar]
113. Endo Y, Rubin JS. Передача сигналов Wnt и рост нейритов: выводы и вопросы. Онкологические науки. 2007; 98: 1311–1317. [PubMed] [Академия Google]
114. Пандур П., Маурус Д., Куль М. Все сложнее: новые игроки входят в сигнальную сеть Wnt. Биоэссе. 2002; 24:881–884. [PubMed] [Google Scholar]
115. Huang HC, Klein PS. Семейство Frizzled: рецепторы для множественных путей передачи сигнала. Геном биол. 2004; 5:234. [Бесплатная статья PMC] [PubMed] [Google Scholar]
116. Бутрос М., Млодзик М. Растрепанные: на перекрестке расходящихся внутриклеточных сигнальных путей. Мех Дев. 1999; 83: 27–37. [PubMed] [Академия Google]
Бутрос М., Млодзик М. Растрепанные: на перекрестке расходящихся внутриклеточных сигнальных путей. Мех Дев. 1999; 83: 27–37. [PubMed] [Академия Google]
117. Ротбахер У., Лоран М.Н., Дирдорф М.А., Кляйн П.С., Чо К.В., Фрейзер С.Е. Беспорядочное фосфорилирование, субклеточная локализация и мультимеризация регулируют его роль в раннем эмбриогенезе. EMBO J. 2000;19:1010–1022. [Бесплатная статья PMC] [PubMed] [Google Scholar]
118. Boutros M, Paricio N, Strutt DI, Mlodzik M. Disheveled активирует JNK и различает пути JNK в плоской полярности и бескрылую передачу сигналов. Клетка. 1998; 94: 109–118. [PubMed] [Академия Google]
119. Rosin-Arbesfeld R, Townsley F, Bienz M. Опухолевой супрессор APC имеет функцию ядерного экспорта. Природа. 2000; 406:1009–1012. [PubMed] [Google Scholar]
120. Wallingford JB, Rowning BA, Vogeli KM, Rothbacher U, Fraser SE, Harland RM. Disheveled контролирует полярность клеток во время гаструляции Xenopus. Природа. 2000;405:81–85. [PubMed] [Google Scholar]
Сигнальный путь WNT и сигнальная сеть стволовых клеток
Обзор
. 2007 15 июля; 13 (14): 4042-5.
doi: 10.1158/1078-0432.CCR-06-2316.
Масуко Като 1 , Масару Като
принадлежность
- 1 M and M Medical BioInformatics, Хонго, Япония.
- PMID: 17634527
- DOI: 10.1158/1078-0432.ЦКР-06-2316
Обзор
Masuko Katoh et al. Клин Рак Рез. .
. 2007 15 июля; 13 (14): 4042-5.
doi: 10. 1158/1078-0432.CCR-06-2316.
1158/1078-0432.CCR-06-2316.
Авторы
Масуко Като 1 , Масару Като
принадлежность
- 1 M and M Medical BioInformatics, Хонго, Япония.
- PMID: 17634527
- DOI: 10.1158/1078-0432.ЦКР-06-2316
Абстрактный
Сигналы WNT преобразуются в канонический путь для определения судьбы клеток и в неканонический путь для контроля движения клеток и тканевой полярности. Канонические сигналы WNT передаются через рецепторы семейства Frizzled и корецептор LRP5/LRP6 в сигнальный каскад бета-катенина. Киназы семейства киназ, регулирующих аффинность микротрубочек (PAR-1), казеинкиназа I эпсилон (CKI эпсилон) и FRAT являются положительными регуляторами канонического пути WNT, тогда как APC, AXIN1, AXIN2, CKI альфа, NKD1, NKD2, бета TRCP1, бета TRCP2, ANKRD6, Nemo-подобная киназа (NLK) и гамма-рецептор, активируемый пролифератором пероксисом (PPAR гамма), являются отрицательными регуляторами. Ядерный комплекс, состоящий из Т-клеточного фактора/лимфоидного энхансера, бета-катенина, BCL9/BCL9L и PYGO активируют транскрипцию канонических генов-мишеней WNT, таких как FGF20, DKK1, WISP1, MYC, CCND1 и Glucagon (GCG). Неканонические сигналы WNT передаются через рецепторы семейства Frizzled и корецепторы ROR2/RYK в зависимые от Dishavelled (GTPазы семейства Rho и c-jun NH(2)-терминальную киназу) или Ca(2+)-зависимые (NLK и ядерный фактор активированные Т-клетки) сигнальные каскады. Сигналы WNT в зависимости от контекста передаются обоим путям на основе профиля экспрессии WNT, SFRP, WIF, DKK, Frizzled рецепторов, корецепторов и активности внутриклеточных сигнальных регуляторов WNT.
Киназы семейства киназ, регулирующих аффинность микротрубочек (PAR-1), казеинкиназа I эпсилон (CKI эпсилон) и FRAT являются положительными регуляторами канонического пути WNT, тогда как APC, AXIN1, AXIN2, CKI альфа, NKD1, NKD2, бета TRCP1, бета TRCP2, ANKRD6, Nemo-подобная киназа (NLK) и гамма-рецептор, активируемый пролифератором пероксисом (PPAR гамма), являются отрицательными регуляторами. Ядерный комплекс, состоящий из Т-клеточного фактора/лимфоидного энхансера, бета-катенина, BCL9/BCL9L и PYGO активируют транскрипцию канонических генов-мишеней WNT, таких как FGF20, DKK1, WISP1, MYC, CCND1 и Glucagon (GCG). Неканонические сигналы WNT передаются через рецепторы семейства Frizzled и корецепторы ROR2/RYK в зависимые от Dishavelled (GTPазы семейства Rho и c-jun NH(2)-терминальную киназу) или Ca(2+)-зависимые (NLK и ядерный фактор активированные Т-клетки) сигнальные каскады. Сигналы WNT в зависимости от контекста передаются обоим путям на основе профиля экспрессии WNT, SFRP, WIF, DKK, Frizzled рецепторов, корецепторов и активности внутриклеточных сигнальных регуляторов WNT. Эпигенетическое замалчивание и мутация с потерей функции негативных регуляторов канонического пути WNT происходят при различных видах рака человека. WNT, фактор роста фибробластов (FGF), Notch, Hedgehog и сигнальная сеть трансформирующего фактора роста бета/костного морфогенетического белка участвуют в поддержании тканевого гомеостаза путем регуляции самообновления нормальных стволовых клеток, а также пролиферации или дифференцировки клеток-предшественников. транзитно-амплифицирующие) клетки. Разрыв сигнальной сети стволовых клеток приводит к канцерогенезу. Нестероидные противовоспалительные препараты и гамма-агонисты PPAR, способные ингибировать канонический сигнальный путь WNT, являются кандидатами на химиопрофилактику. ZTM000990 и PKF118-310 являются ведущими соединениями, нацеленными на канонический сигнальный каскад WNT. Моноклональные антитела анти-WNT1 и анти-WNT2 демонстрируют эффекты in vitro при лечении рака. После оптимизации производные низкомолекулярных соединений и моноклональных антител человека, направленные на сигнальный путь WNT, могут быть использованы в медицине рака.
Эпигенетическое замалчивание и мутация с потерей функции негативных регуляторов канонического пути WNT происходят при различных видах рака человека. WNT, фактор роста фибробластов (FGF), Notch, Hedgehog и сигнальная сеть трансформирующего фактора роста бета/костного морфогенетического белка участвуют в поддержании тканевого гомеостаза путем регуляции самообновления нормальных стволовых клеток, а также пролиферации или дифференцировки клеток-предшественников. транзитно-амплифицирующие) клетки. Разрыв сигнальной сети стволовых клеток приводит к канцерогенезу. Нестероидные противовоспалительные препараты и гамма-агонисты PPAR, способные ингибировать канонический сигнальный путь WNT, являются кандидатами на химиопрофилактику. ZTM000990 и PKF118-310 являются ведущими соединениями, нацеленными на канонический сигнальный каскад WNT. Моноклональные антитела анти-WNT1 и анти-WNT2 демонстрируют эффекты in vitro при лечении рака. После оптимизации производные низкомолекулярных соединений и моноклональных антител человека, направленные на сигнальный путь WNT, могут быть использованы в медицине рака.
Похожие статьи
-
STAT3-индуцированная сигнальная петля WNT5A в эмбриональных стволовых клетках, взрослых нормальных тканях, хроническом персистирующем воспалении, ревматоидном артрите и раке (обзор).
Като М., Като М. Като М. и соавт. Int J Mol Med. 2007 Февраль; 19 (2): 273-8. Int J Mol Med. 2007. PMID: 17203201 Обзор.
-
Сигнальный путь WNT/PCP и рак человека (обзор).
Като М. Като М. Представитель Oncol, 2005 г., декабрь 14(6):1583-8. Представитель Oncol, 2005 г. PMID: 16273260 Обзор.
-
Ингибитор передачи сигналов FGF, SPRY4, является эволюционно законсервированной мишенью пути передачи сигналов WNT в клетках-предшественниках.

Като Ю., Като М. Като И. и др. Int J Mol Med. 2006 март; 17(3):529-32. Int J Mol Med. 2006. PMID: 16465403
-
Избыточная экспрессия канонических лигандов Wnt в клеточных линиях рака молочной железы человека.
Бенхадж К., Аккали К.С., Озтюрк М. Бенхай К. и др. Представитель Oncol, 2006 г., март 15(3):701-7. Представитель Oncol, 2006 г. PMID: 16465433
-
Ген антагониста Wnt DICKKOPF-1 является нижележащей мишенью бета-катенина/TCF и подавляется при раке толстой кишки человека.
Гонсалес-Санчо Х.М., Агилера О., Гарсия Х.М., Пендас-Франко Н., Пенья С., Кэл С., Гарсия де Эррерос А., Бонилья Ф., Муньос А. Гонсалес-Санчо Х.М. и др. Онкоген. 2005 г., 3 февраля; 24(6):1098-103.
 doi: 10.1038/sj.onc.1208303.
Онкоген. 2005.
PMID: 15592505
doi: 10.1038/sj.onc.1208303.
Онкоген. 2005.
PMID: 15592505
Посмотреть все похожие статьи
Цитируется
-
Ориентация на раковые стволовые клетки как ключевой фактор канцерогенеза и терапевтической резистентности.
Ид Р.А., Алаа Эдин М., Шедид Э.М., Камаль АСС, Варда М.М., Мамдух Ф., Хедр С.А., Солтан М.А., Чон Х.В., Заки М.С.А., Ким Б. Ид Р.А. и др. Int J Mol Sci. 2023 16 января; 24 (2): 1786. дои: 10.3390/ijms24021786. Int J Mol Sci. 2023. PMID: 36675306 Бесплатная статья ЧВК. Обзор.
-
Моделирование патологии хряща при мукополисахаридозе VI с использованием ИПСК выявляет раннюю дисрегуляцию экспрессии хондрогенных и метаболических генов.
Бродерс М.
 , ван Ройдж Дж., Уссорен Э., ван Гестель Т., Смит С., Кимбер С., Вердейк Р., Вагенмакерс М., ван ден Хаут Дж., ван дер Плоэг А., Нарциси Р., Пийнаппель В.
Бродерс М. и соавт.
Фронт Биоэнг Биотехнолог. 2022 6 декабря; 10:949063. doi: 10.3389/fbioe.2022.949063. Электронная коллекция 2022.
Фронт Биоэнг Биотехнолог. 2022.
PMID: 36561048
Бесплатная статья ЧВК.
, ван Ройдж Дж., Уссорен Э., ван Гестель Т., Смит С., Кимбер С., Вердейк Р., Вагенмакерс М., ван ден Хаут Дж., ван дер Плоэг А., Нарциси Р., Пийнаппель В.
Бродерс М. и соавт.
Фронт Биоэнг Биотехнолог. 2022 6 декабря; 10:949063. doi: 10.3389/fbioe.2022.949063. Электронная коллекция 2022.
Фронт Биоэнг Биотехнолог. 2022.
PMID: 36561048
Бесплатная статья ЧВК. -
Амниотическая мембрана человека модулирует пути передачи сигналов Wnt/β-catenin и NF-κβ в суставных хондроцитах in vitro .
Чанг С., Масси М., Кооб Т.Дж. Чанг С. и др. Osteoarthr Cartil Open. 2021 4 сентября; 3(4):100211. doi: 10.1016/j.ocarto.2021.100211. электронная коллекция 2021 дек. Osteoarthr Cartil Open. 2021. PMID: 36474757 Бесплатная статья ЧВК.
-
Сетевое расположение и кластеризация генетических мутаций определяют хроничность в стилизованной модели генетических заболеваний.

