Какие микросхемы УНЧ обеспечивают высокую выходную мощность. Как выбрать подходящую микросхему УНЧ для автомобиля или домашней аудиосистемы. На какие параметры обращать внимание при выборе микросхем УНЧ.
Популярные микросхемы УНЧ для мощных усилителей звука
Усилители низкой частоты (УНЧ) на интегральных микросхемах получили широкое распространение благодаря ряду преимуществ:
- Компактные размеры
- Простота схемотехнических решений
- Минимум внешних компонентов
- Высокая надежность
- Хорошие электрические параметры
- Доступность и низкая стоимость
Рассмотрим наиболее популярные микросхемы УНЧ, способные обеспечить высокую выходную мощность для автомобильных и бытовых аудиосистем.
TDA7294 — мощный УНЧ для Hi-Fi систем
Микросхема TDA7294 от STMicroelectronics является одной из самых популярных для построения мощных Hi-Fi усилителей звука. Ее основные характеристики:
- Выходная мощность до 100 Вт на нагрузке 8 Ом
- Напряжение питания ±10…±40 В
- Коэффициент нелинейных искажений менее 0.1%
- Полоса пропускания 20 Гц — 20 кГц
- Отношение сигнал/шум 100 дБ
TDA7294 обеспечивает отличное качество звучания при высокой выходной мощности, что делает ее отличным выбором для домашних Hi-Fi систем.

TDA1562Q — мощный автомобильный УНЧ
Микросхема TDA1562Q от NXP Semiconductors специально разработана для применения в автомобильных аудиосистемах. Ее ключевые особенности:
- Выходная мощность до 70 Вт на нагрузке 4 Ом
- Однополярное питание 12-18 В
- Встроенная защита от перегрева и КЗ
- Низкий уровень искажений
- Возможность мостового включения
TDA1562Q позволяет создать мощный автомобильный усилитель с минимумом внешних компонентов.
TDA7293 — высокомощный УНЧ с защитой
TDA7293 от STMicroelectronics обеспечивает высокую выходную мощность при низком уровне искажений:
- Выходная мощность до 120 Вт на 8 Ом
- Напряжение питания ±12…±50 В
- КНИ 0.005% при 100 Вт
- Встроенная тепловая защита
- Защита от КЗ выхода
Благодаря встроенным схемам защиты, TDA7293 отлично подходит для создания надежных мощных усилителей.
Какие параметры важны при выборе микросхемы УНЧ?
При выборе микросхемы УНЧ для конкретного применения следует обратить внимание на следующие ключевые параметры:
- Выходная мощность — должна соответствовать требуемой мощности акустических систем
- Напряжение питания — важно для выбора блока питания
- Коэффициент нелинейных искажений — влияет на качество звучания
- Отношение сигнал/шум — определяет уровень фоновых шумов
- Полоса пропускания — должна перекрывать слышимый диапазон частот
- Наличие встроенных схем защиты — повышает надежность
- Тип корпуса и способ отвода тепла — влияет на конструкцию усилителя
Правильный выбор микросхемы УНЧ с учетом этих параметров позволит создать качественный и надежный усилитель звука.

Рекомендации по применению микросхем УНЧ
При разработке усилителей на микросхемах УНЧ следует придерживаться следующих рекомендаций:
- Использовать качественные компоненты обвязки микросхемы
- Обеспечить эффективный теплоотвод от микросхемы
- Применять сглаживающие фильтры по цепям питания
- Разделять силовые и сигнальные цепи на печатной плате
- Использовать рекомендованные производителем схемы включения
Соблюдение этих рекомендаций позволит реализовать все преимущества микросхем УНЧ и создать качественный усилитель звука.
Многоканальные микросхемы УНЧ
Для создания многоканальных аудиосистем удобно использовать специализированные многоканальные микросхемы УНЧ. Рассмотрим некоторые популярные варианты:
TDA7850 — четырехканальный автомобильный УНЧ
Микросхема TDA7850 содержит четыре независимых канала усиления и обладает следующими характеристиками:
- Выходная мощность 4×50 Вт на 4 Ом
- Напряжение питания 8-18 В
- Встроенная защита от перегрева и КЗ
- Возможность мостового включения каналов
TDA7265 — двухканальный УНЧ для домашней техники
TDA7265 представляет собой двухканальный УНЧ для бытовой аудиотехники:

- Выходная мощность 2×25 Вт на 8 Ом
- Напряжение питания ±12…±22 В
- Низкий уровень искажений
- Встроенная защита от перегрузки
Использование многоканальных микросхем позволяет упростить схемотехнику и уменьшить габариты многоканальных усилителей.
Заключение
Современные микросхемы УНЧ предоставляют широкие возможности для создания высококачественных усилителей звука различного назначения. Правильный выбор микросхемы с учетом требуемых параметров и соблюдение рекомендаций по применению позволят реализовать все преимущества интегральных УНЧ и получить отличные результаты.
Усилители мощности звука (УНЧ) на микросхемах, схемы самодельных УМЗЧ
Усилитель мощности низкой частоты — это электронное устройство, которое предназначено для усиления низкочастотного (НЧ) сигнала с последующей его подачей на акустические системы. Часто самодельные интегральные усилители мощности низкой частоты собирают на мощных микросхемах, поскольку они требуют минимум внешних компонентов и очень просты в наладке.
В разделе собраны принципиальные схемы усилителей мощности НЧ на мощных микросхемах, а также на основе интегральных микросхем — драйверов для выходных транзисторов. Используя специализированные интегральные микросхемы можно собрать усилитель мощности разной конфигурации:
- Стерео — два канала усиления мощности;
- Квадро — четыре канала усиления мощности;
- 2+1 — сабвуфер и два сателлита;
- 5+1 — сабвуфер и пять сателлитов;
- и другие.
Если нужна большая выходная мощность усилителя НЧ (например для канала сабвуфера — 200Втт) то зачастую применяются мостовые схемы включения микросхем или же в параллель.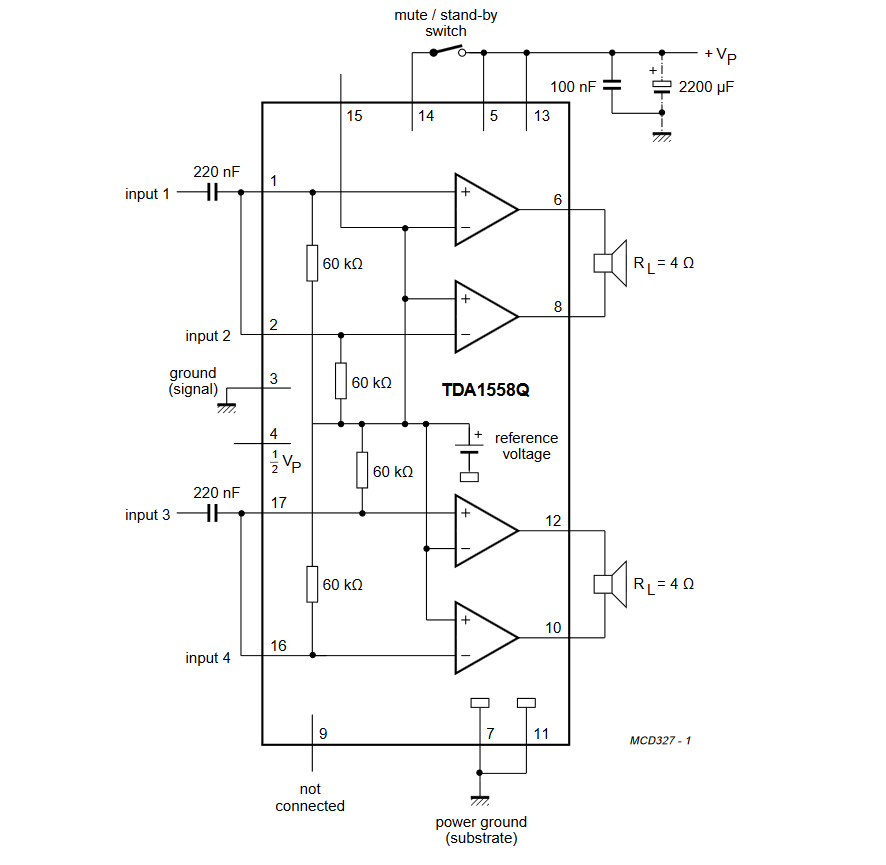
Здесь вы найдете схемы самодельных УМЗЧ разной сложности для внешних и интегрированных акустических систем, схемы простых усилителей для наушников и миниатюрной бытовой техники (плееры, MP3, диктофоны, игрушки и т.д).
Схема усилителя мощности звука на микросхеме OPA541 (60W)Можно сказать, уже сложилась такая традиция, если нужен мощный УМЗЧ с минимальным набором обвязки и хорошими параметрами, его делают на микросхеме TDA или LM. Традиция традицией, но есть и другие варианты, хотя и не такие проверенные и отработанные… Опыт с УМЗЧ на микросхеме OPA541 еще интересен …
1 320 0
Усилитель мощности на микросхеме LM3876, LM3886 (40-100 Вт)Схема и описание самодельного усилителя мощности на микросхеме LM3876, LM3886 фирмы NS (National Semiconductor). Параметры усилителя: 1. Номинальный уровень входного сигнала … 1V. 2. Выходная мощность на нагрузке 8 Ом при КНИ не более 0,1%….. 40W …
0 357 0
Мощный УМЗЧ на микросхеме TDA7294, печатная платаТем кто занялся конструированием усилителя для аудиосистемы или DVD-плеера конечно же хочется достигнуть наилучших результатов с минимальными трудовыми затратами.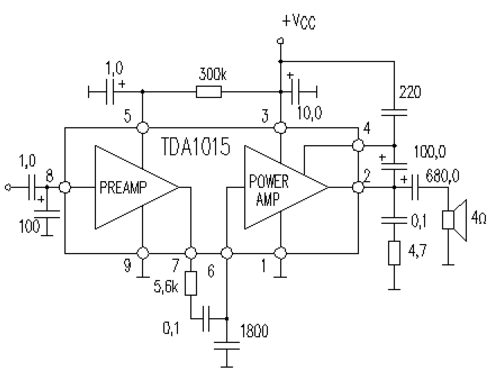 УМЗЧ на микросхеме TDA7294 в этом смысле как раз то что нужно. Вот девять доводов в пользу УМЗЧ на TDA7294: 1 Выходная мощность …
УМЗЧ на микросхеме TDA7294 в этом смысле как раз то что нужно. Вот девять доводов в пользу УМЗЧ на TDA7294: 1 Выходная мощность …
0 199 0
Самодельный усилитель звука для планшета или смартфона на микросхеме TDA1554QСейчас многие автолюбители в машине пользуются планшетными компьютерами. Это очень удобно, потому что планшет -это и средство мобильной аудио и видео связи, это навигатор, с его помощью можно оперативно найти нужную информацию в интернете. Кроме того, планшет может работать как радиоприемник, как …
2 766 0
Двухканальный усилитель звука на микросхеме TDA7496L (2 Вт на 8 Ом)Интегральная микросхема типа TDA7496L производства фирмы SGT-Thomson Microelectronics представляет собой двухканальный усилитель звуковой частоты с выходной мощностью в каждом канале до 2 Вт на нагрузке сопротивлением 8 Ом. Максимальная рассеиваемая мощность 6 Вт. напряжение питания однополярное …
0 941 0
Простой стерео усилитель НЧ для компьютера на микросхеме К174УН20Усилитель стереофонический, выполнен на микросхеме К174УН20 советского производства. Микросхема содержит два УНЧ, по схемотехнике, аналогичных двум микросхемам типа К174УН14, но меньшей мощности и в корпусе типа DIP16, но с двумя радиаторными пластинами, вместо выводов …
Микросхема содержит два УНЧ, по схемотехнике, аналогичных двум микросхемам типа К174УН14, но меньшей мощности и в корпусе типа DIP16, но с двумя радиаторными пластинами, вместо выводов …
0 739 0
Усилитель мощности звука с регулятором тембра (LM741, LM1875)1 2176 0
Простой стереоусилитель на микросхеме TDA2005 с регулятором тембраМикросхема TDA2005 устаревшая, и уже давно не выпускается, однако она все еще остается одной из самых недорогих и широкодоступных, интегральных УМЗЧ. Относительно небольшое число навесных элементов, в сочетании с вполне хорошими электрическими характеристиками, наличие защиты выхода от перегрузки …
1 919 0
Стерео усилитель звука на микросхемах TDA2050 с регулятором ВЧ и НЧЭто несложный полный УНЧ на двух микросхемах TDA2050, питающийся отимпульсного блока питания для галогеновых светильников (выходное переменное напряжение 12V, мощность 75W).
1 2377 0
Простой усилитель звукового сигнала на микросхеме TDA1010A (7 Ватт)Микросхема TDA1010A представляет собой ИМС УНЧ для телевизоров и другой электронной техники. Особенность этой микросхемы в том, что в ней есть как усилитель мощности ЗЧ, так и предварительный усилитель. Причем, выход предварительного усилителя и вход усилителя мощности выведены на разные …
1 882 0
1 2 3 4 5 … 35Радиодетали, электронные блоки и игрушки из китая:
93 Схем УНЧ на микросхемах TDA, LA, HA, KA, AN и другие
Коллекция усилителей НЧ и мощности на «буржуйских» микросхемах, более или менее часто встречаются при «аутопсии» очередного дохлого музцентра (или ещё чего-нибудь)! В архивах куча схем формата PCX от ТМП «Ассоциация» (респект им если они еще «живы»). С краткими данными: максимальное, рабочее напряжение; входное и выходное сопративление; сила тока и т.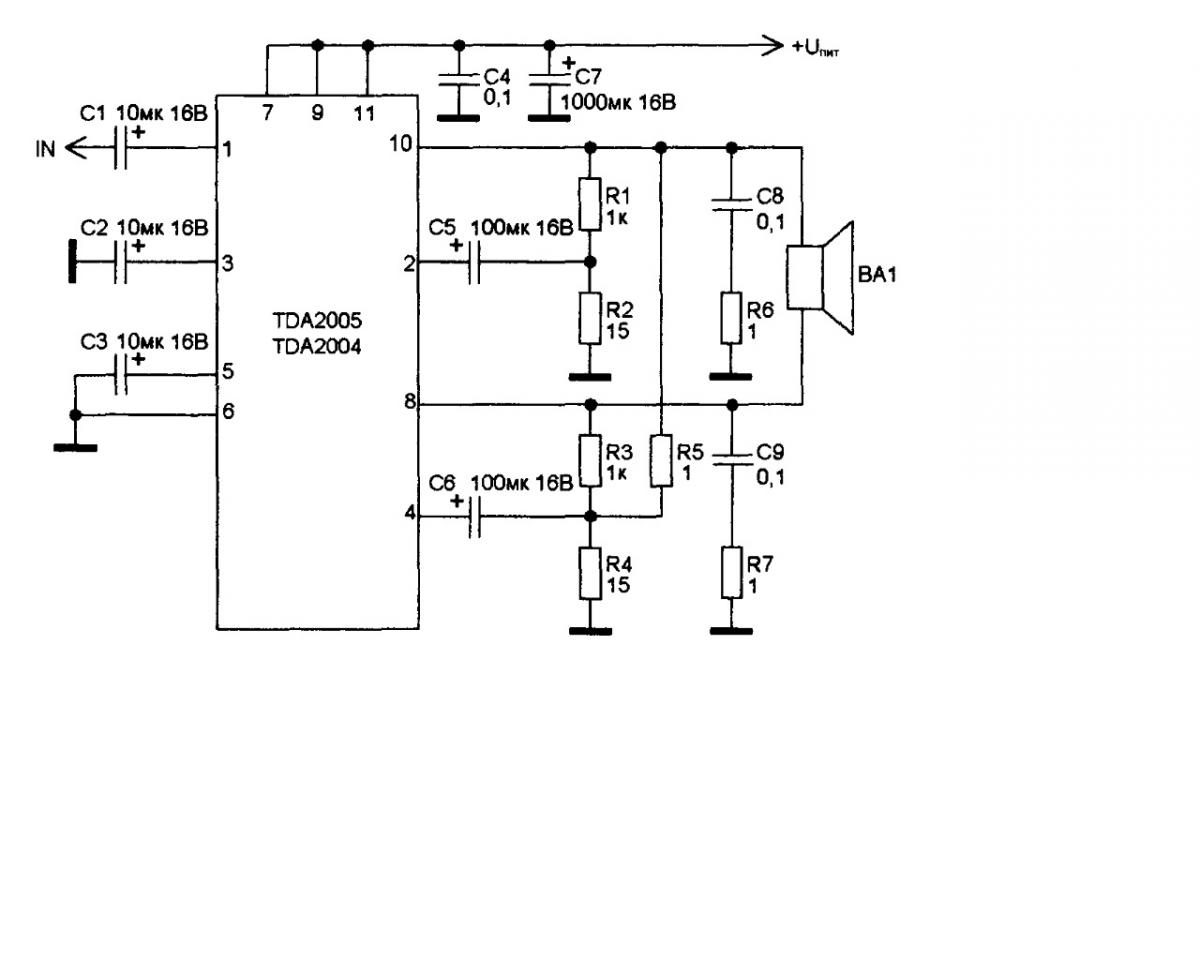
Учитывая универсальность этих микросхем, можно собрать неплохой усилитель «на коленках» минут за 15. Рекомендую скачать все архивы, пока есть =).
микросхемы TDA.
к Примеру 4 конденсатора, выключатель и 2 динамика — получаете стерео усилитель на 2х22Вт.
и другие: TDA1551Q, TDA1552Q, TDA1553Q, TDA1554Q, TDA1555Q, TDA1904, TDA1905, TDA2003, TDA2004, TDA2005, TDA2006, TDA2007, TDA2008, TDA2009, TDA2020, TDA2030, TDA2030А, TDA2040, TDA2611A, TDA2613, 2822(d,m), 7050(t), TDA7052A, TDA7056, TDA7056A, TDA7057Q, TDA7230A, TDA7231, TDA7233D, TDA7233S, TDA7240A, TDA7245, TDA7285, TDA7350, TDA7241.
Скачать архив с TDA (265 кб)
Микросхемы AN.
AN7112, AN7116, AN7117, AN7147, AN7149N, AN7116N, AN7168, AN7171NK, AN7173NK, AN7177, AN7178.
Скачать архив с AN (89 кб)
Микросхемы HA.
HA13001, HA1377, HA1384, HA1388.
Скачать архив с HA (32 кб)
Микросхемы KA.

KA2211, KA2213, KA2214.
Скачать архив с KA (25 кб)
Микросхемы LA.
LA4265, LA4101, LA4145, LA4182, LA4182, LA4183, LA4185, LA4190, LA4191, LA4261, LA4440, LA4445, LA4446, LA4460N, LA4461N, LA4465, LA4475, LA4476, LA4480, LA4497, LA4498, LA4500, LA4505, LA4507, LA4510, LA4520, LA4550, LA4555, LA4557, LA4558, LA4570, LA4575, LA4700, LA4422.
Скачать архив с LA (299 кб)
Микросхемы LM386, MB3722, MB3730, MB3731, MDA2020, STK0050, STK0050 II.
Скачать архив других (56 кб)
Art!P. 2004.
УНЧ на микросхеме
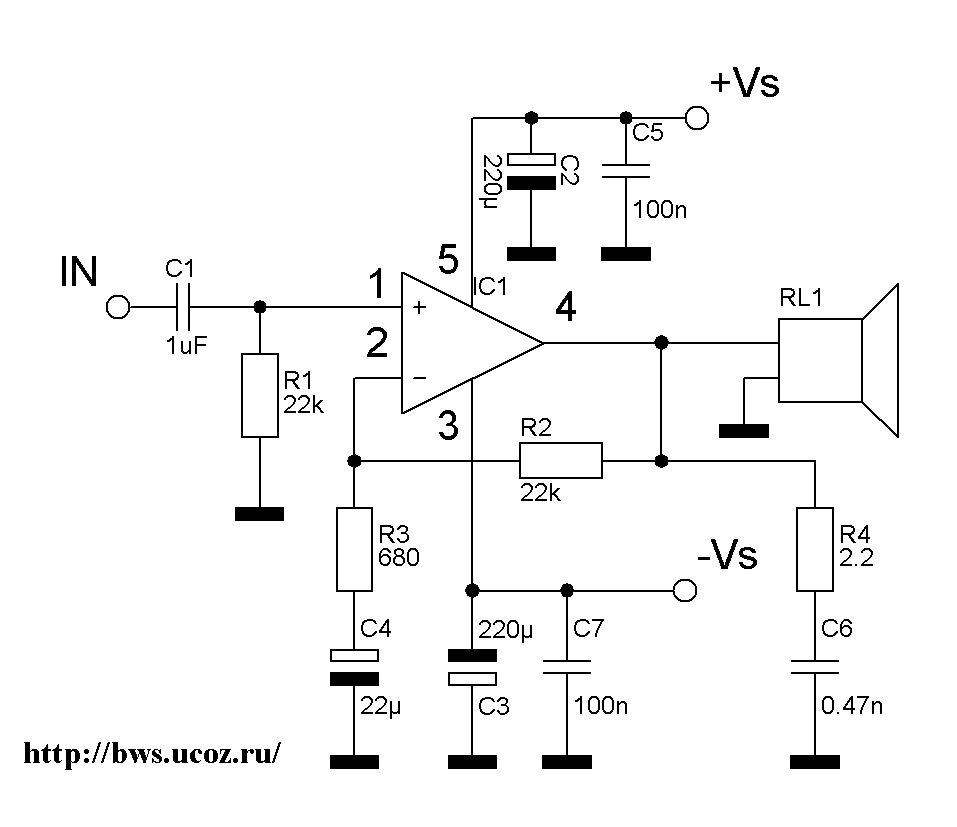
С помощью микросхемы мы построим качественный и мощный, как для домашних условий, усилитель звука на 100 ватт. Мы используем для построения микросхему LM3886 которая является усовершенствованной версией её предшественника LM3875. Отличие от стандартного включения микросхемы будет за счет добавления обратной связи и басс-компенсации. Между прочим, данные этой схемы гораздо лучше, чем многие из промышленных HI-FI усилителей, которые продаются на рынке. Особенно трудно найти приличное отношение сигнал / шум, которое тут составляет 110 дБ. А при отсутствии входного сигнала, усилитель совсем как мертвый — почти невозможно услышать шум, когда вы прижимаетесь ухом к динамикам.
Между прочим, данные этой схемы гораздо лучше, чем многие из промышленных HI-FI усилителей, которые продаются на рынке. Особенно трудно найти приличное отношение сигнал / шум, которое тут составляет 110 дБ. А при отсутствии входного сигнала, усилитель совсем как мертвый — почти невозможно услышать шум, когда вы прижимаетесь ухом к динамикам.
Параметры усилителя на LM3886
- Максимальная Выходная мощность: 68W RMS — 108 ВТ пиковая.
- THD: 0.03% @ 60W.
- SNR: 110 дБ @ 60W — 92.5 дБ @ 1 Вт.
- Широкий диапазон: 120dB.
- Схемы защиты: DC / AC защита от короткого замыкания, тепловая защита.
Электросхема УНЧ на микросхеме
Как вы видите, схема является достаточно простой и выполнимой при небольшом опыте паяния. Улучшения стандартной даташитовской схемы включения заключаются в том, что номиналы отрицательной обратной линии связи были радикально пересмотрены.
 Таким образом, в диапазоне низких частот 30 Гц — 70 Гц, был получен прирост 7dB. УНЧ даёт настоящий глубокий бас (на реальных 20 Гц), и будет очень мощным дополнением к активному сабвуферу, хотя можно при таких параметрах обойтись и без него. Мощности и так достаточно, ведь 26В постоянного тока и 4 Ома динамики выдают в среднем 65 Вт номинальной мощности (RMS).
Таким образом, в диапазоне низких частот 30 Гц — 70 Гц, был получен прирост 7dB. УНЧ даёт настоящий глубокий бас (на реальных 20 Гц), и будет очень мощным дополнением к активному сабвуферу, хотя можно при таких параметрах обойтись и без него. Мощности и так достаточно, ведь 26В постоянного тока и 4 Ома динамики выдают в среднем 65 Вт номинальной мощности (RMS).Электросхема блока питания УНЧ на микросхеме
Схема питания также очень простая. Как мы видим, используется трансформатор с двумя независимыми обмотками, диодными мостами и конденсаторами на 10.000 микрофарад. Как правило такого стабилизатора достаточно для стабилизации HI-FI устройств.
Список деталей для УНЧ
Полупроводники:
LM3886TA 2 шт.
KBU608 2 шт.
Конденсаторы:
10.000 35 (50) v 2 шт.
10 мкФ 35 (50) v Тантал — 4 шт.
2.2 35 (50) v Тантал — 2 шт.
470nF 63v полиэстер 1 Шт
100 нФ керамические 63v 2 шт.
150nF 63v полиэстер 2 шт.
Резисторы:
20K 0,6 Вт 1% металопленочные 2 шт.
20K 1/8W 5% углеродный резистор 2 шт.
15K 0,6 Вт 1% металопленочные 2 шт.
10K 0,6 Вт 1% металопленочные 2 шт.
0.6 Вт 1% металопленочные 1K 4 шт.
2.2 ом 1/4W 5% углеродный резистор 1 шт.
Другие детали:
10K x 2 логарифмический потенциометр
Радиаторы на микросхемы
RCA-типа гнёзда
Клеммы для подключения АС
Разъем для сетевого питания.
Выбор трансформатора питания
Лучше поставить тороидальный трансформатор, часто используемый в схемах HI-FI. Этот тип трансформаторов дорогой, но его преимущества в малом рассеивании магнитного потока. Трансформатор имеет следующие данные:
— 8 ом — 220 / 2 x 24В,
— 4 ом — 220 / 2 x 18В,
— Мощность надо по крайней мере, 300W.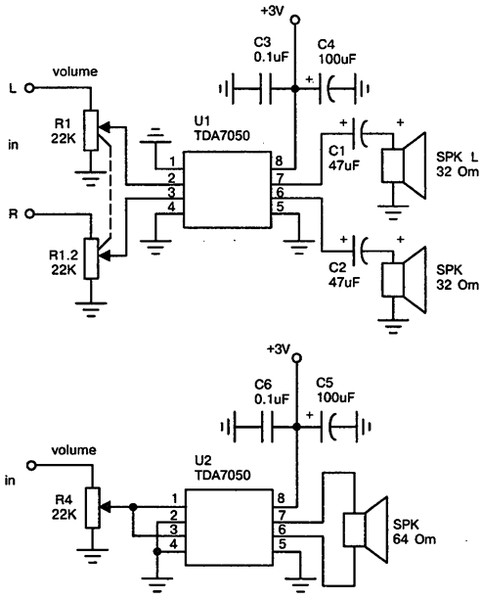
Зачем нам нужен такой на трансформатор, если на канал всего 68W выходная мощности RMS? Потому что с учётом КПД и второго канала, а также броска тока на НЧ пиках, потребуется не менее 250 ватт.
Корпус для УНЧ на микросхемах
Это устройство можно собрать в подходящем красивом ящики, из дерева, куска металла или пластика… безусловно, конструкция должна иметь вентиляционные отверстия. В противном случае, система долгое время не проработает — всё таки 200 ватт общей мощности! Используйте стандартные клеммы для выходов АС. Для аудиовходов можно использовать стандартные RCA гнёзда. Входной сигнал внутри коробки подключите к RCA гнёздам по кратчайшему пути от разъема до платы с микросхемой.Понравилась схема — лайкни!
ПРИНЦИПИАЛЬНЫЕ СХЕМЫ УНЧ
Смотреть ещё схемы усилителей
УСИЛИТЕЛИ НА ЛАМПАХ УСИЛИТЕЛИ НА ТРАНЗИСТОРАХ
УСИЛИТЕЛИ НА МИКРОСХЕМАХ СТАТЬИ ОБ УСИЛИТЕЛЯХ
УНЧ и Звукотехника | Усилители мощности низкой частоты | Микросхема
Как много в этой аббревиатуре для сердца радиолюбителя слилось. Каждый, кто когда-нибудь занимался радиотехникой и электроникой, собирал различные усилители низкой частоты. Простые и сложные, маломощные и мощные. Сейчас, с развитием интегральных микросхем, стало вообще всё намного проще. Усилители не содержат каких-то уникальных радиодеталей. Одна микросхема, которая, собственно, и представляет собой уже готовый усилитель мощности низкой частоты, и схема, практически, собрана. Как правило, выходная мощность таких усилителей и качество воспроизведения на высоте. А если прикупить головку динамическую прямого излучения Ватт так на 1500 — 2000 и встроить в корпус с фазоинвертором, выполненный по рассчитанным размерам, то вообще замечательно. Получится сабвуфер не хуже покупного. В большинстве случаев даже лучше.
Каждый, кто когда-нибудь занимался радиотехникой и электроникой, собирал различные усилители низкой частоты. Простые и сложные, маломощные и мощные. Сейчас, с развитием интегральных микросхем, стало вообще всё намного проще. Усилители не содержат каких-то уникальных радиодеталей. Одна микросхема, которая, собственно, и представляет собой уже готовый усилитель мощности низкой частоты, и схема, практически, собрана. Как правило, выходная мощность таких усилителей и качество воспроизведения на высоте. А если прикупить головку динамическую прямого излучения Ватт так на 1500 — 2000 и встроить в корпус с фазоинвертором, выполненный по рассчитанным размерам, то вообще замечательно. Получится сабвуфер не хуже покупного. В большинстве случаев даже лучше.
Чистота и качество воспроизведения постоянно совершенствуются. Основные термины в данном разделе:
Бел (Б) — логарифмическая единица, соответствующая (при частоте 1000 Гц) десятикратному изменению силы звука.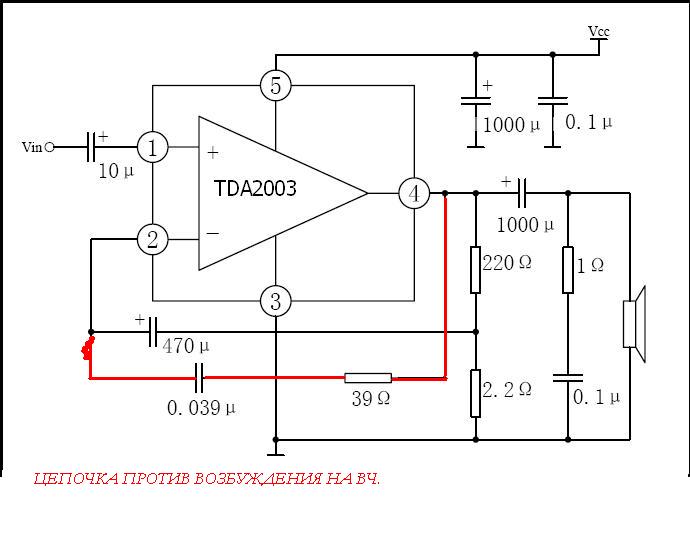 Логарифмическая единица, соответствующая 1/10 бела, называется децибелом (дБ). Одному дБ соответствует изменение звукового давления в 1,12 раза.
Логарифмическая единица, соответствующая 1/10 бела, называется децибелом (дБ). Одному дБ соответствует изменение звукового давления в 1,12 раза.
Частота звуковых колебаний воспринимается на слух как высота тона. Самый низкий предел, воспринимаемый человеком, 20 Гц, а самый высокий — 20000 Гц.
Тембр — окраска звука, определяемая количеством, частотой и интенсивностью обертонов.
Уровень звукового давления — отношение данного звукового давления p к нулевому уровню p0, выраженное в дБ. Вычисляется как N=20 lg(p/p0).
Болевой порог — звуковое давление, которое вызывает болевое ощущение на коже. Уровень равен 120 дБ.
В радиолюбительской практике принято делить УНЧ на обычные и высокого качества (Hi-Fi класса). Максимальная выходная мощность всех звуковых усилителей определяется по простой формуле: Pвых=U2/Rн. Т.е. замеряете напряжение на выходе УНЧ (обязательно под нагрузкой), возводите в квадрат и делите на сопротивление нагрузки (обычно сопротивление динамика 4-8 Ом). Можно ещё упомянуть о предварительном усилении. К усилителям мощности обязательно нужны такие каскады, чтобы напряжение на его входе было достаточным.
Можно ещё упомянуть о предварительном усилении. К усилителям мощности обязательно нужны такие каскады, чтобы напряжение на его входе было достаточным.
Бывают ещё различные по сложности усилительные каскады. Однотактные, двухтактные, трансформаторные и бестрансформаторные, мостовые схемы включения усилительных элементов. Одна из возможных схем двухтактного трансформаторного каскада усилителя звуковой частоты приведена ниже. Номинальная выходная мощность 4 Вт, максимальная — 6 Вт.
Но такие, я думаю, уже никто не будет собирать. Слишком трудоёмко наматывать трансформатор, плюс ко всему нужно найти подходящий магнитопровод.
Приведу ещё пример двухтактного бестрансформаторного каскада УНЧ. Выходная мощность порядка 10 Вт.
У нас в наличии имеется более 850 схем УНЧ на интегральных микросхемах. По мере необходимости будем выкладывать их на сайт, особенно самые лучшие, на наш взгляд. Если Вам нужен какой-то усилитель и Вы не можете найти его схему, то пишите, пожалуйста, в комментариях или в форме обратной связи.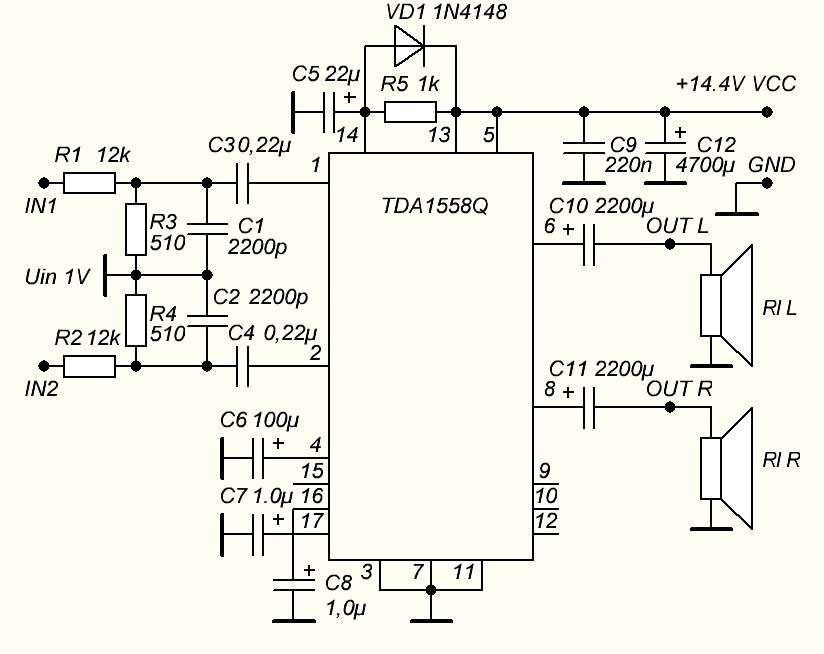 Мы обязательно поможем.
Мы обязательно поможем.
Ниже приведены ссылки на различные материалы по данной теме. Особо отметим, что среди них есть полностью опубликованные с полным описанием схемы, входящих радиоэлементов, различных настроек и замеров основных параметров (например, силы тока и напряжения) на разных участках цепи и между элементами. Также есть с кратким описанием, содержащие ссылку на скачивание всего документа в одном архиве, где, в свою очередь, содержится уже полное описание конструкции, печатной платы и прочее. Архивы имеют расширение *.rar (распаковать можно, например, программой WinRAR версии 2.9 и выше) и доступны для скачивания. Примечание: эта мера введена из-за того, что многие запакованные материалы являются целыми пособиями. Подразумевается, что Вам будет удобнее скачать на жесткий диск и просматривать уже локально, нежели листать страницу за страницей, расходуя трафик и время.
Радиосхемы. — Схемы усилителей на микросхемах
В данной категории нашего сайта Вы найдете схемы усилителей, собранных на микросхемах— как специализированных, так и тех, где микросхемы используются совместно с транзисторами.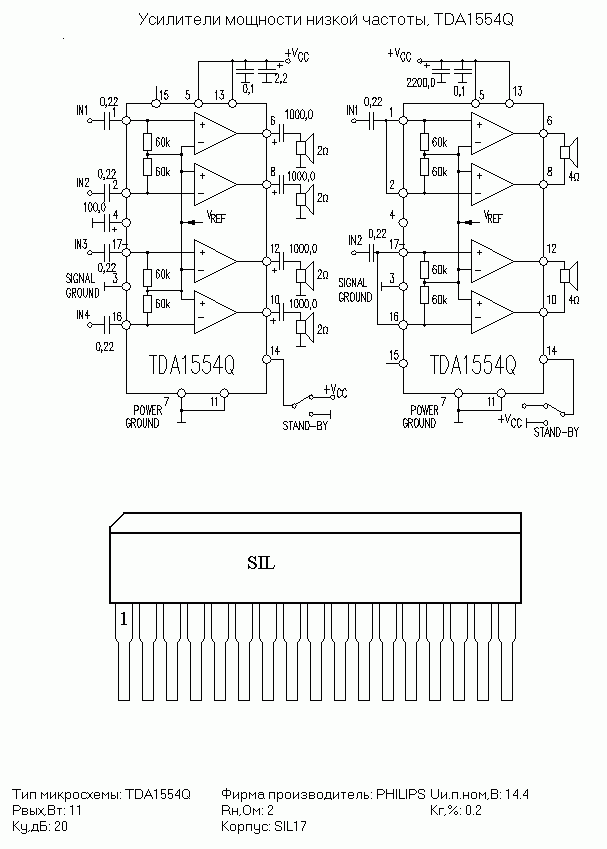
Так как микросхем-усилителей очень много, то рассказать о всех в пределах одной категории очень трудно, то возможно Вас заинтересует справочник по микросхема-усилителям НЧ. Вы можете найти его здесь, и при желании скачать (конечно-же бесплатно!).
Если у Вас возникли вдруг какие-то вопросы по сборке, настройке или ремонту микросхемных усилителей, нашли неточность в схеме, либо Вы сами захотите поделиться опытом- то заходите к нам на ФОРУМ
Материалы категории Схемы усилителей на микросхемах
Усилитель на 600 Ватт
Усилитель на 150 Ватт с эквалайзером
Усилитель на TDA2822 c темброблоком
Микросхема TDA1010
Цифровая микросхема в роли усилителя
Микросхема TEA2025
Микросхема TDA2003
Микросхема TDA2004(05)
TDA 2030
Радиолюбительские конструкции на микросхеме TDA2030
TDA 1517 (1519)
TDA 1557
TA 8205 (10,15)
TDA2822
TA8227
KIA6283
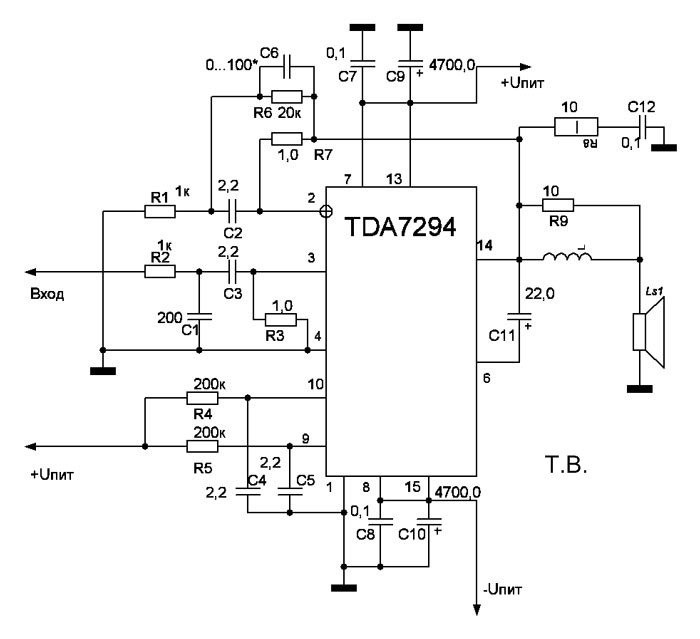
TDA7294
КА2206- простой стереоусилитель 3 Ватта
Микросхема AN17803- трехканальный УНЧ 2Х10 + 18 Вт
AN17810- двухканльный УНЧ 2Х6,5 Вт
AN17820- двухканальный УНЧ 2 х 7,5 Вт
AN17821 двухканальный УНЧ 2Х5 Вт
AN17823- одноканальный УНЧ 4Вт
STK4122II двухканальный УНЧ 2Х15 Вт
усилитель на операционной микросхеме К140УД6 с выходным каскадом на транзисторах
Усилитель на операционном усилителе и транзисторах с выходной мощностью до 50 Ватт.
Экономичный усилитель на К140УД1Б и КТ808, КТ806 (30 Ватт)
Экономичный усилитель на К140УД1Б и шести транзисторах
Усилитель 20 Ватт на КР544УД2А и выходными транзисторами КТ818, КТ819
Автомобильный усилитель на TDA7376B (12V, 2×35 W)
TDA1514A высококачественный усилитель 50 Ватт
TA3020 усилитель 2х300 Ватт
Усилитель 120 Ватт на NE5534 и полевых транзисторах
Усилитель 60 Вт на КР1408УД1 и выходном каскаде на КТ972, КТ908
Усилитель 100 Ватт для сабвуфера
Миниатюрный усилитель для переносной аппаратуры
Усилитель 60 Вт класса D
Трехполосный усилитель на 574УД1А и транзисторах КП904
Усилитель 90 Ватт на К574УД1А, КТ827,КТ825
Усилитель 80 Ватт на К574УД1Б, КТ818ГМ, КТ819ГМ
Усилитель 50 Вт на К140УД11 с выходным каскадом на КТ827
Высококачественный УНЧ 42 ВТ на К544УД2, КТ818Б, КТ819Б
Малогабаритный низковольтный усилитель класса D
Простой усилитель на К548УН1 и выходном каскаде на транзисторах
Простой усилитель с эквалайзером
Нестандартные включения микросхем TDA2003, TDA2030
Усилитель мощности для автомобильной аппаратуры
УНЧ с нестандартным включением ОУ
Автомобильный УНЧ 2 х 70 Ватт
Усилитель для кассетного проигрывателя
УНЧ с регулируемым выходным сопротивлением
Стереоусилитель на микросхеме TDA7294
Улучшение качества звучания для переносных магнитол
Трехканальный усилитель на двухканальной микросхеме
Стереоусилитель для аудиокомплекса на микросхемах
HI-FI стереоусилитель на с эквалайзером и LM3886
Система 2. 1 для автомагнитолы
1 для автомагнитолы
УНЧ для аудиоплеера
Стереоусилитель на КР544УД2А и КТ972(73)
Микросхемы серии TDA. Усилители низкой частоты.
В настоящее время стала доступна широкая номенклатура импортных интегральных усилителей низкой частоты. Их достоинствами являются удовлетворительные электрические параметры, возможность выбора микросхем с заданной выходной мощностью и напряжением питания, стереофоническое или квадрафоническое исполнение с возможностью мостового включения.Для изготовления конструкции на основе интегрального УНЧ требуется минимум навесных деталей. Применение заведомо исправных компонентов обеспечивает высокую повторяемость и, как правило, дополнительной настройки не требуется.
Приводимые типовые схемы включения и основные параметры интегральных УНЧ призваны облегчить ориентацию и выбор наиболее подходящей микросхемы.
Для квадрафонических УНЧ не
указаны параметры в мостовом стереофоническом включении.
TDA1010
Напряжение питания — 6…24 B
Максимальный потребляемый ток — 3 A
Выходная мощность (Un =14,4 В,.КНИ=10%):
RL=2 Ом — 6,4 Вт
RL=4 Ом — 6,2 Вт
RL=8 Ом — 3,4 Вт
КНИ (Р=1 Вт, RL=4 Ом) — 0,2 %
Ток покоя — 31 мА
Схема включения
TDA1011
Напряжение питания — 5,4…20 B
Максимальный потребляемый ток — 3 A
Выходная мощность (RL=4 Ом, КНИ=10%):
Un=16B — 6,5 Вт
Un=12В — 4,2 Вт
Un=9В — 2,3 Вт
Un=6B — 1,0 Вт
КНИ (Р=1 Вт, RL=4 Ом) — 0,2 %
Ток покоя — 14 мА
Схема включения
TDA1013
Напряжение питания — 10.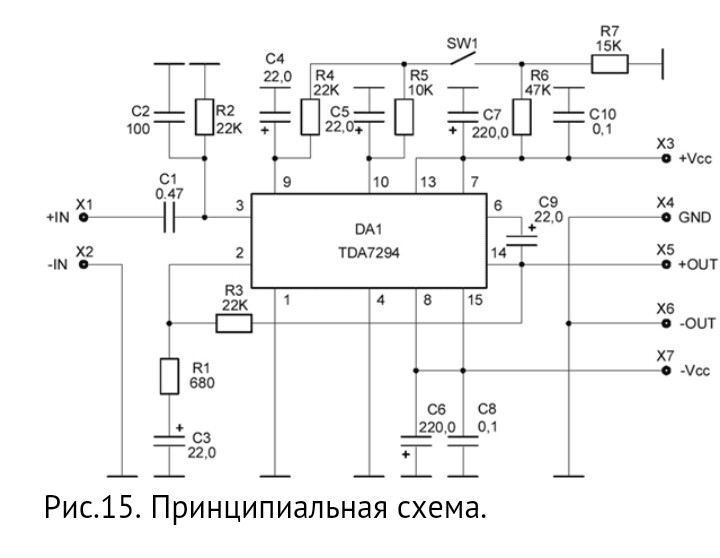 ..40 B
..40 B
Максимальный потребляемый ток — 1,5 A
Выходная мощность (КНИ=10%) — 4,2 Вт
КНИ (Р=2,5 Вт, RL=8 Ом) — 0,15 %
Схема включения
TDA1015
Напряжение питания — 3,6…18 В
Максимальный потребляемый ток — 2,5 А
Выходная мощность (RL=4 Ом, КНИ=10%):
Un=12В — 4,2 Вт
Un=9В — 2,3 Вт
Un=6B — 1,0 Вт
КНИ (Р=1 Вт, RL=4 Ом) — 0,3 %
Ток покоя — 14 мА
Схема включения
TDA1020
Напряжение питания — 6…18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Un =14,4 В, КНИ=10%):
RL=2 Oм — 12 Вт
RL=4 Ом — 7 Вт
RL=8 Ом — 3,5 Вт
Ток покоя — 30 мА
Схема включения
TDA1510
Напряжение питания — 6. ..18 В
..18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Un=14,4B RL=4 Oм):
КНИ=0,5% — 5,5 Вт
КНИ=10% — 7,0 Вт
Ток покоя — 120 мА
Схема включения
TDA1514
Напряжение питания - ±10…±30 В
Максимальный потребляемый ток — 6,4 А
Выходная мощность:
Un =±27,5 В, R=8 Ом — 40 Вт
Un =±23 В, R=4 Ом — 48 Вт
Ток покоя — 56 мА
Схема включения
TDA1515
Напряжение питания — 6…18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Un =14,4 В, КНИ=0,5%):
RL=2 Ом — 9 Вт
RL=4 Ом — 5,5 Вт
Выходная мощность (Un=14,4 В, КНИ=10%):
RL=2 Oм — 12 Вт
RL4 Ом — 7 Вт
Ток покоя — 75 мА
Схема включения
TDA1516
Напряжение питания — 6. ..18 В
..18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Un =14,4 В, КНИ=0,5%):
RL=2 Ом — 7,5 Вт
RL=4 Ом — 5 Вт
Выходная мощность (Un =14,4 В, КНИ=10%):
RL=2 Oм — 11 Вт
RL=4 Ом — 6 Вт
Ток покоя — 30 мА
Схема включения
TDA1517
Напряжение питания — 6…18 В
Максимальный потребляемый ток — 2,5 А
Выходная мощность (Un=14,4B RL=4 Oм):
КНИ=0,5% — 5 Вт
КНИ=10% — 6 Вт
Ток покоя — 80 мА
Схема включения
TDA1518
Напряжение питания — 6.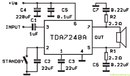 ..18 В
..18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Un =14,4 В, КНИ=0,5%):
RL=2 Ом — 8,5 Вт
RL=4 Ом — 5 Вт
Выходная мощность (Un =14,4 В, КНИ=10%):
RL=2 Oм — 11 Вт
RL=4 Ом — 6 Вт
Ток покоя — 30 мА
Схема включения
TDA1519
Напряжение питания — 6…17,5 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Uп=14,4 В, КНИ=0,5%):
RL=2 Ом — 6 Вт
RL=4 Ом — 5 Вт
Выходная мощность (Un =14,4 В, КНИ=10%):
RL=2 Ом — 11 Вт
RL=4 Ом — 8,5 Вт
Ток покоя — 80 мА
Схема включения
TDA1551
Напряжение питания -6. ..18 В
..18 В
Выходная мощность (Un =14,4 В, RL=4 Ом):
КНИ=0,5% — 5 Вт
КНИ=10% — 6 Вт
Ток покоя — 160 мА
Схема включения
TDA1521
Напряжение питания - ±7,5…±21 В
Максимальный потребляемый ток — 2,2 А
Выходная мощность (Un=±12 В, RL=8 Ом):
КНИ=0,5% — 6 Вт
КНИ=10% — 8 Вт
Ток покоя — 70 мА
Схема включения
TDA1552
Напряжение питания — 6…18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Un =14,4 В, RL=4 Ом):
КНИ=0,5% — 17 Вт
КНИ=10% — 22 Вт
Ток покоя — 160 мА
Схема включения
TDA1553
Напряжение питания — 6. ..18 В
..18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Uп=4,4 В, RL=4 Ом):
КНИ=0,5% — 17 Вт
КНИ=10% — 22 Вт
Ток покоя — 160 мА
Схема включения
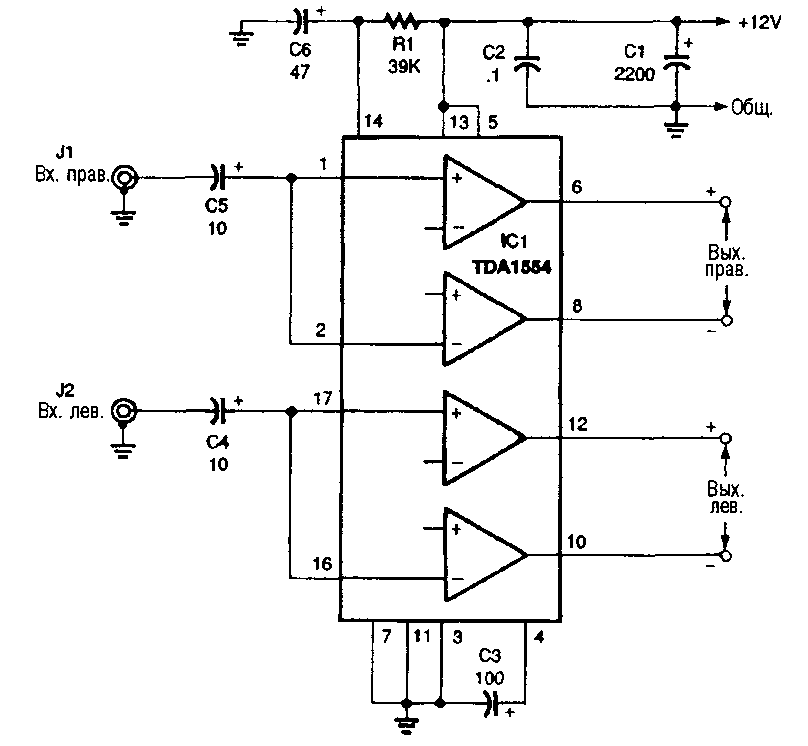
TDA1554
Напряжение питания — 6…18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Uп =14,4 В, RL=4 Ом):
КНИ=0,5% — 5 Вт
КНИ=10% — 6 Вт
Ток покоя — 160 мА
Схема включения
TDA2004
Сдвоенный интегральный УНЧ, разработанный
специально для применения в автомобиле и допускающий работу на
низкоомную нагрузку (до 1,6 Ом).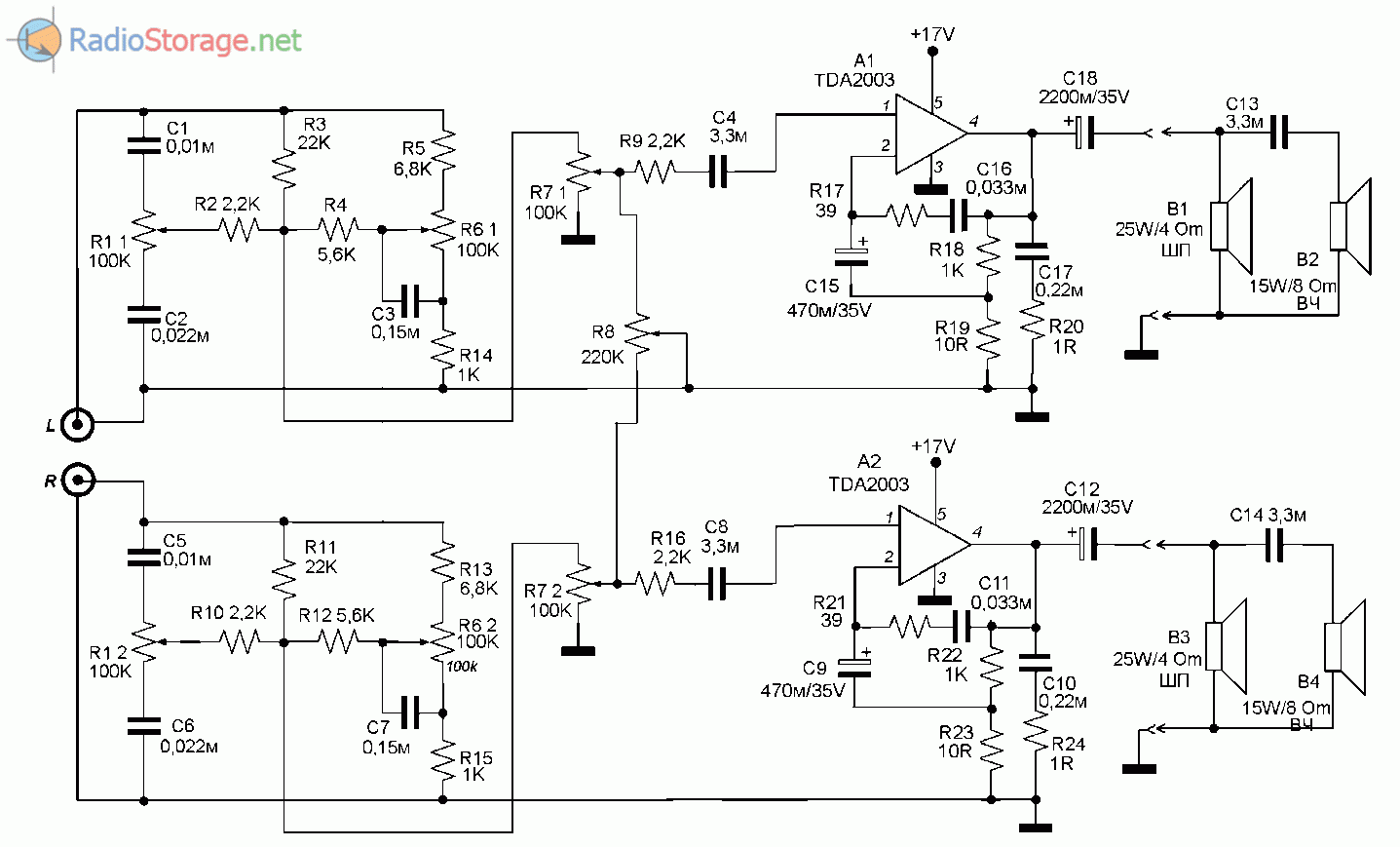
Напряжение питания — 8…18 В
Максимальный потребляемый ток — 3,5 А
Выходная мощность (Un=14,4 В, КНИ=10%):
RL=4 Ом — 6,5 Вт
RL=3,2 Ом — 8,0 Вт
RL=2 Ом — 10 Вт
RL=1,6 Ом — 11 Вт
KHИ (Un=14,4B, Р=4,0 Вт, RL=4 Ом)- 0,2%;
Полоса пропускания (по уровню -3 дБ) — 35…15000 Гц
Ток покоя — <120 мА
Схема включения
TDA2005
Сдвоенный интегральный УНЧ, разработанный специально для применения в автомобиле и допускающий работу на низкоомную нагрузку (до 1,6 Ом).
Напряжение питания — 8…18 В
Максимальный потребляемый ток — 3,5 А
Выходная мощность (Uп =14,4 В, КНИ=10%):
RL=4 Ом — 20 Вт
RL=3,2 Ом — 22 Вт
КНИ (Uп =14,4 В, Р=15 Вт, RL=4 Ом) — 10 %
Полоса пропускания (по уровню -3 дБ) — 40. ..20000
Гц
..20000
Гц
Ток покоя — <160 мА
Схема включения
TDA2006
Интегральный УНЧ, обеспечивающий большой выходной ток, низкое содержание гармоник и интермодуляционных искажений.Расположение выводов совпадает с расположением выводов микросхемы TDA2030.
Напряжение питания - ±6,0…±15 В
Максимальный потребляемый ток — 3 А
Выходная мощность (Еп=±12В,КНИ=10%):
при RL=4 Oм — 12 Вт
при RL=8 Ом — 6…8 Вт КНИ (Еп=±12В):
при Р=8 Вт, RL= 4 Ом — 0,2 %
при Р=4 Вт, RL= 8 Ом — 0,1 %
Полоса пропускания (по уровню -3 дБ) — 20…100000 Гц
Ток потребления:
при Р=12 Вт, RL=4 Ом — 850 мА
при Р=8 Вт, RL=8 Ом — 500 мА
Схема включения
TDA2007
Сдвоенный интегральный УНЧ с однорядным
расположением выводов, специально разработанный для применения в
телевизионных и портативных радиоприемниках.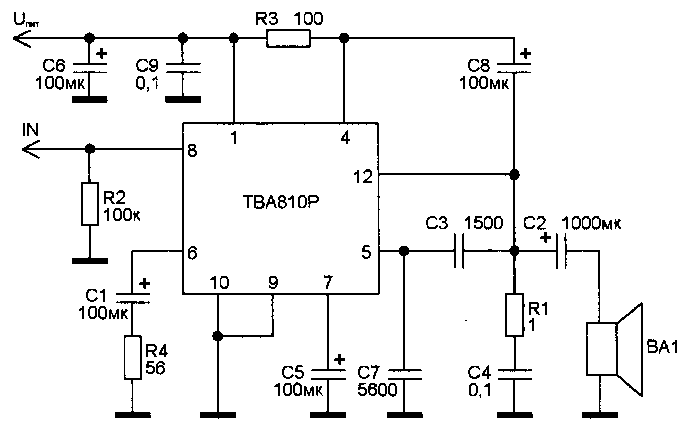
Напряжение питания — +6…+26 В
Ток покоя (Eп=+18 В) — 50…90 мА
Выходная мощность (КНИ=0,5 %):
при Еп=+18 В, RL=4 Ом — 6 Вт
при Еп=+22 В, RL=8 Ом — 8 Вт
КНИ:
при Еп=+18 В Р=3 Вт, RL=4 Ом — 0,1 %
при Еп=+22 В, Р=3 Вт, RL=8 Ом — 0,05 %
Полоса пропускания (по уровню -3 дБ) — 40…80000 Гц
Максимальный ток потребления — 3 А
Схема включения
TDA2008
Интегральный УНЧ, предназначенный для работы на низкоомную нагрузку, обеспечивающий большой выходной ток, очень низкое содержание гармоник и интермодуляционных искажений.
Напряжение питания — +10…+28 В
Ток покоя (Еп=+18 В) — 65. ..115 мА
..115 мА
Выходная мощность (Еп=+18В, КНИ= 10%):
при RL=4 Oм — 10…12 Вт
при RL=8 Ом — 8 Вт
КНИ (Еп= +18 В):
при Р=6 Вт, RL=4 Ом — 1 %
при Р=4 Вт, RL=8 Ом — 1 %
Максимальный ток потребления — 3 А
Схема включения
TDA2009
Сдвоенный интегральный УНЧ, предназначенный для применения в высококачественных музыкальных центрах.
Напряжение питания — +8…+28 В
Ток покоя (Еп=+18 В) — 60…120 мА
Выходная мощность (Еп=+24 В, КНИ=1 %):
при RL=4 Oм — 12,5 Вт
при RL=8 Ом — 7 Вт
Выходная мощность (Еп=+18 В, КНИ=1 %):
при RL=4 Oм — 7 Вт
при RL=8 Ом — 4 Вт
КНИ:
при Еп= +24 В, Р=7 Вт, RL=4 Oм — 0,2 %
при Еп= +24 В, Р=3,5 Вт, RL=8 Oм — 0,1 %
при Еп= +18 В, Р=5 Вт, RL=4 Oм — 0,2 %
при Еп= +18 В, Р=2,5 Вт, RL=8 Ом — 0,1 %
Полоса пропускания (по уровню -3 дБ) — 20.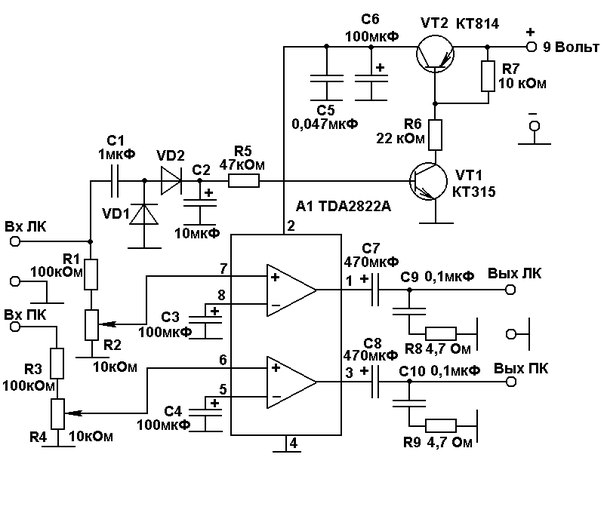 ..80000
Гц
..80000
Гц
Максимальный ток потребления — 3,5 А
Схема включения
TDA2030
Интегральный УНЧ, обеспечивающий большой выходной ток, низкое содержание гармоник и интермодуляционных искажений.
Напряжение питания - ±6…±18 В
Ток покоя (Еп=±14 В) — 40…60 мА
Выходная мощность (Еп=±14 В, КНИ = 0,5
%):
при RL=4 Oм — 12…14 Вт
при RL=8 Ом — 8…9 Вт
КНИ (Еп=±12В):
при Р=12 Вт, RL=4 Ом — 0,5 %
при Р=8 Вт, RL=8 Ом — 0,5 %
Полоса пропускания (по уровню -3 дБ) — 10…140000 Гц
Ток потребления:
при Р=14 Вт, RL=4 Ом — 900 мА
при Р=8 Вт, RL=8 Ом — 500 мА
Схема включения
TDA2040
Интегральный УНЧ, обеспечивающий большой выходной
ток, низкое содержание гармоник и интермодуляционных искажений.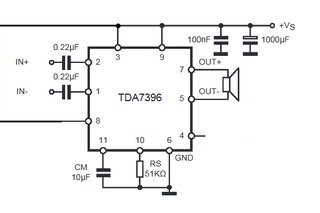
Напряжение питания - ±2,5…±20 В
Ток покоя (Еп=±4,5…±14 В) — мА 30…100 мА
Выходная мощность (Еп=±16 В, КНИ = 0,5
%):
при RL=4 Oм — 20…22 Вт
при RL=8 Ом — 12 Вт
КНИ(Еп=±12В, Р=10 Вт, RL = 4 Ом) — 0,08 %
Максимальный ток потребления — 4 А
Схема включения
TDA2050
Интегральный УНЧ, обеспечивающий большую выходную мощность, низкое содержание гармоник и интермодуляционных искажений. Предназначен для работы в Hi-Fi-стереокомплексах и телевизорах высокого класса.
Напряжение питания - ±4,5…±25 В
Ток покоя (Еп=±4,5…±25 В) — 30…90 мА
Выходная мощность (Еп=±18, RL = 4 Ом,
КНИ = 0,5 %) — 24.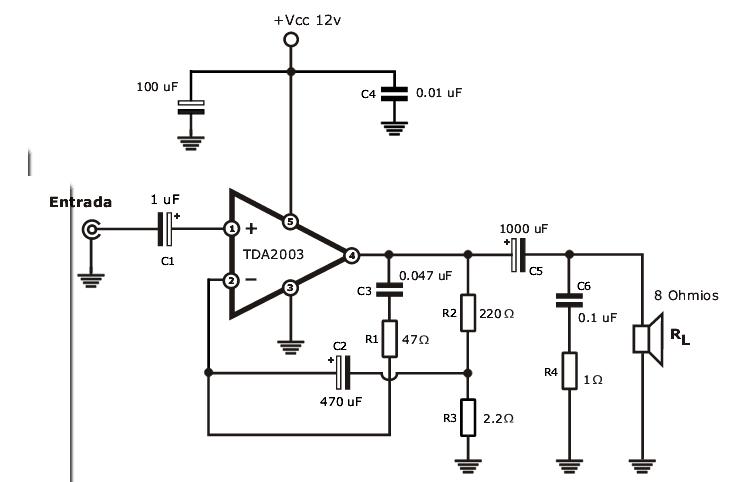 ..28 Вт
..28 Вт
КНИ (Еп=±18В, P=24Bт, RL=4 Ом) - 0,03…0,5 %
Полоса пропускания (по уровню -3 дБ) — 20…80000 Гц
Максимальный ток потребления — 5 А
Схема включения
TDA2051
Интегральный УНЧ, имеющий малое число внешних элементов и обеспечивающий низкое содержание гармоник и интермодуляционных искажений. Выходной каскад работает в классе АВ, что позволяет получить большую выходную мощность.
Выходная мощность:
при Еп=±18 В, RL=4 Ом, КНИ=10% — 40 Вт
при Еп=±22 В, RL=8 Ом, КНИ=10% — 33 Вт
Схема включения
TDA2052
Интегральный УНЧ, выходной каскад которого
работает в классе АВ. Допускает широкий диапазон напряжений питания и
имеет большой выходной ток. Предназначен для работы в телевизионных и
радиоприемниках.
Допускает широкий диапазон напряжений питания и
имеет большой выходной ток. Предназначен для работы в телевизионных и
радиоприемниках.
Напряжение питания - ±6…±25 В
Ток покоя (En = ±22 В) — 70 мА
Выходная мощность (Еп = ±22 В, КНИ =
10%):
при RL=8 Ом — 22 Вт
при RL=4 Ом — 40 Вт
Выходная мощность (En = 22 В, КНИ = 1%):
при RL=8 Ом — 17 Вт
при RL=4 Ом — 32 Вт
КНИ (при полосе пропускания по уровню -3 дБ 100…
15000 Гц и Рвых=0,1…20 Вт):
при RL=4 Ом — <0,7 %
при RL=8 Ом — <0,5 %
Схема включения
TDA2611
Интегральный УНЧ, предназначенный для работы в
бытовой аппаратуре.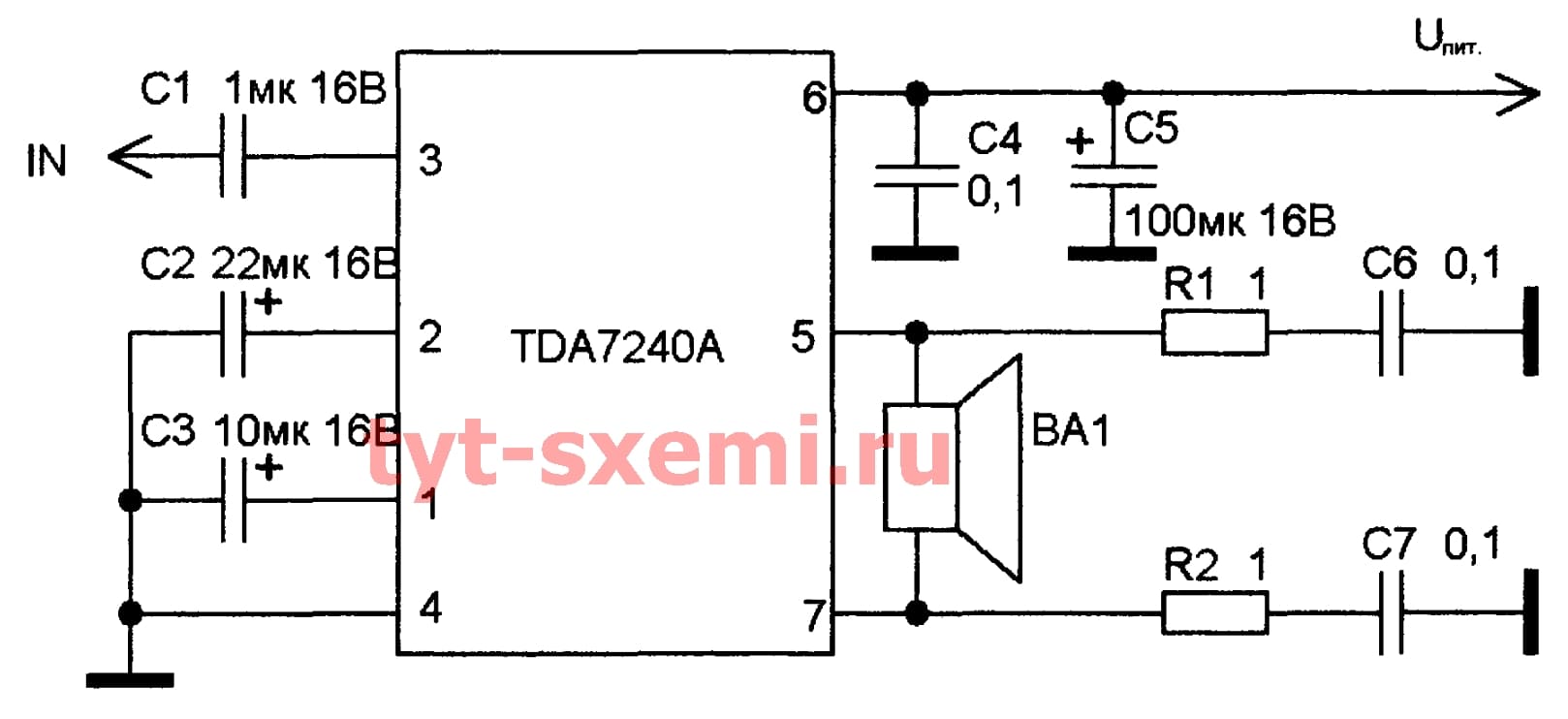
Напряжение питания — 6…35 В
Ток покоя (Еп=18 В) — 25 мА
Максимальный ток потребления — 1,5 А
Выходная мощность (КНИ=10%): при Еп=18 В, RL=8 Ом
— 4 Вт
при Еп=12В, RL=8 0м — 1,7 Вт
при Еп=8,3 В, RL=8 Ом — 0,65 Вт
при Еп=20 В, RL=8 Ом — 6 Вт
при Еп=25 В, RL=15 Ом — 5 Вт
КНИ (при Рвых=2 Вт) — 1 %
Полоса пропускания — >15 кГц
Схема включения
TDA2613
Интегральный УНЧ, предназначенный для работы в бытовой аппаратуре (телевизионных и радиоприемниках).
Напряжение питания — 15…42 В
КНИ:
(Еп=24 В, RL=8 Ом, Рвых=6 Вт) — 0,5 %
(Еп=24 В, RL=8 Ом, Рвых=8 Вт) — 10 %
Ток покоя (Еп=24 В) — 35 мА
Максимальный ток потребления — 2,2 А
Схема включения
TDA2614
Интегральный УНЧ, предназначенный для работы в
бытовой аппаратуре (телевизионных и радиоприемниках).
Напряжение питания — 15…42 В
Максимальный ток потребления — 2,2 А
Ток покоя (Еп=24 В) — 35 мА
КНИ:
(Еп=24 В, RL=8 Ом, Рвых=6,5 Вт) — 0.5 %
(Еп=24 В, RL=8 Ом, Рвых=8,5 Вт) — 10 %
Полоса пропускания (по уровню -3 дБ) — 30…20000 Гц
Схема включения
TDA2615
Сдвоенный УНЧ, предназначенный для работы в стереофонических радиоприемниках или телевизорах.
Напряжение питания — ±7,5…21 В
Максимальный потребляемый ток — 2,2 А
Ток покоя (Еп=7,5…21 В) — 18…70 мА
Выходная мощность (Еп=±12 В, RL=8 Ом):
КНИ=0,5% — 6 Вт
КНИ=10% — 8 Вт
Полоса пропускания (по уровню-3 дБ и Рвых=4 Вт) -
20. ..20000 Гц
..20000 Гц
Схема включения
TDA2822
Сдвоенный УНЧ, предназначенный для работы в носимых радио и телеприемниках.
Напряжение питания — 3…15 В
Максимальный потребляемый ток — 1,5 А
Ток покоя (Еп=6 В) — 12 мА
Выходная мощность (КНИ=10%, RL=4 Ом):
Еп=9В — 1,7 Вт
Еп=6В — 0,65 Вт
Еп=4.5В — 0,32 Вт
Схема включения
TDA7052
Схема включения
TDA7053
Схема включения
TDA2824
Сдвоенный УНЧ, предназначенный для работы в носимых радио- и телеприемниках
Напряжение питания — 3.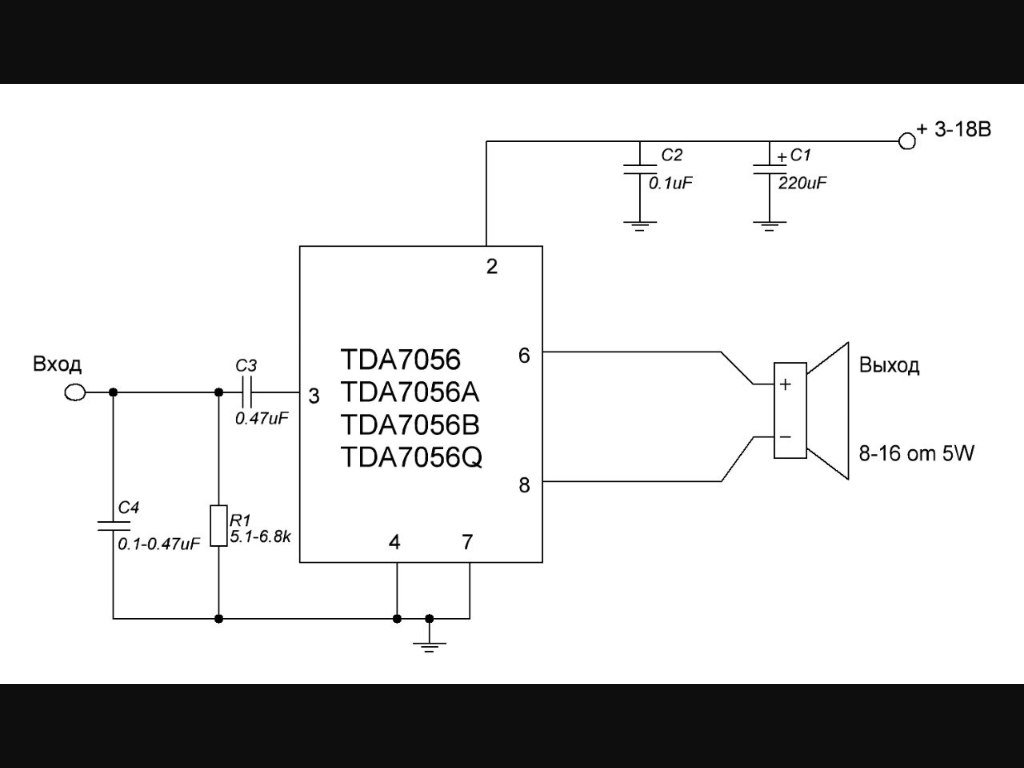 ..15 В
..15 В
Максимальный потребляемый ток — 1,5 А
Ток покоя (Еп=6 В) — 12 мА
Выходная мощность (КНИ=10%, RL=4 Oм)
Еп=9 В — 1,7 Вт
Еп=6 В — 0,65 Вт
Еп=4,5 В — 0,32 Вт
КНИ (Еп=9 В, RL=8 Ом, Рвых=0,5 Вт) — 0,2 %
Схема включения
TDA7231
УНЧ с широким диапазоном напряжений питания, предназначенный для работы в носимых радиоприемниках, кассетных магнитофонах и т.д.
Напряжение питания — 1,8…16 В
Максимальный потребляемый ток — 1,0 А
Ток покоя (Еп=6 В) — 9 мА
Выходная мощность (КНИ=10%):
En=12B, RL=6 Oм — 1,8 Вт
En=9B, RL=4 Ом — 1,6 Вт
Еп=6 В, RL=8 Ом — 0,4 Вт
Еп=6 В, RL=4 Ом — 0,7 Вт
Еп=З В, RL=4 Oм — 0,11 Вт
Еп=3 В, RL=8 Ом — 0,07 Вт
КНИ (Еп=6 В, RL=8 Ом, Рвых=0.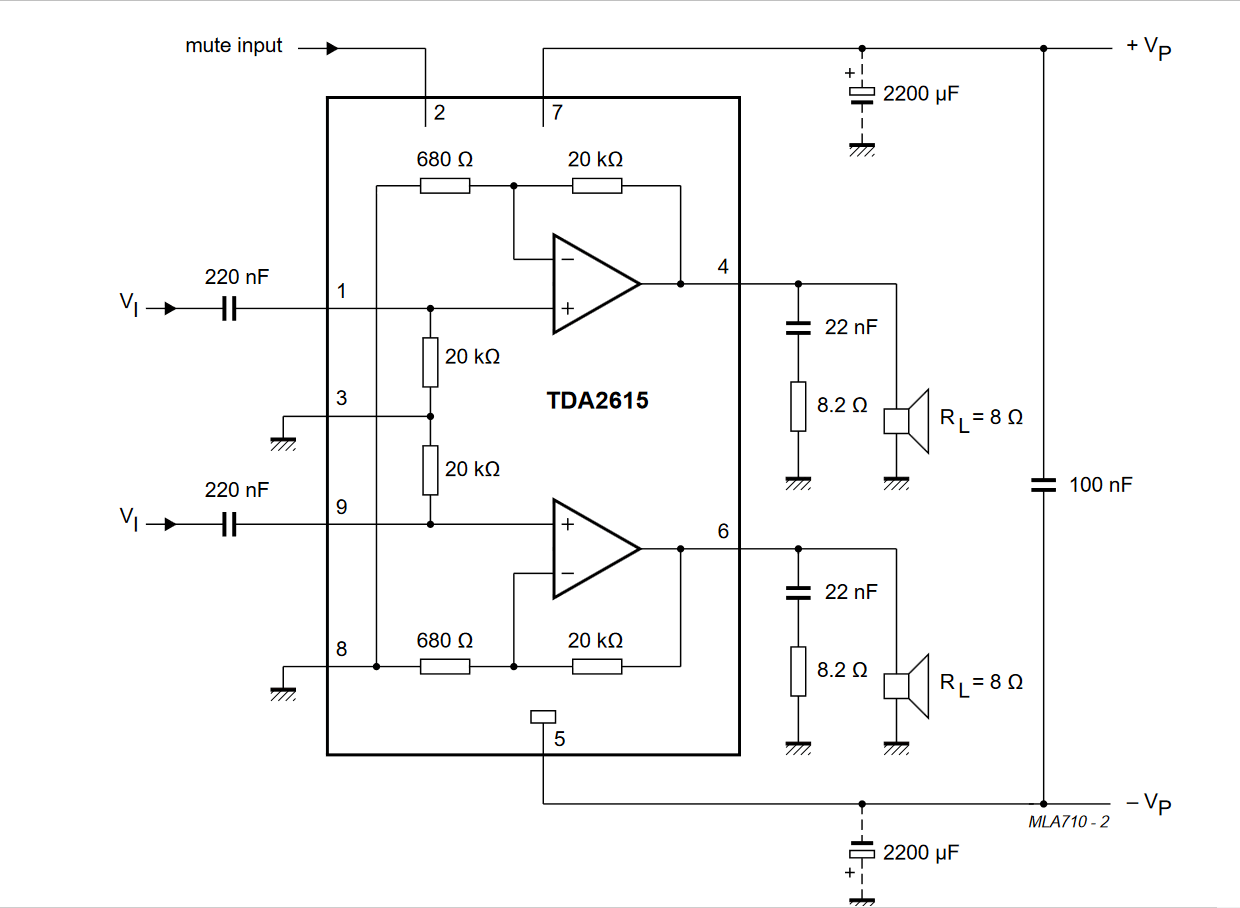 2 Вт) — 0,3 %
2 Вт) — 0,3 %
Схема включения
TDA7235
УНЧ с широким диапазоном напряжений питания, предназначенный для работы в носимых радио- и телеприемниках, кассетных магнитофонах и т.д.
Напряжение питания — 1,8…24 В
Максимальный потребляемый ток — 1,0 А
Ток покоя (Еп=12 В) — 10 мА
Выходная мощность (КНИ=10%):
Еп=9 В, RL=4 Oм — 1,6 Вт
Еп=12 В, RL=8 Oм — 1,8 Вт
Еп=15 В, RL=16 Ом — 1,8 Вт
Eп=20 B, RL=32 Oм — 1,6 Вт
КНИ (Еп=12В, RL=8 Oм, Рвых=0,5 Вт) — 1,0 %
Схема включения
TDA7240
Мостовой УНЧ, разработанный для применения в
автомобильных магнитолах. Имеет защиту от короткого замыкания в
нагрузке, а также от перегрева.
Имеет защиту от короткого замыкания в
нагрузке, а также от перегрева.
Максимальное напряжение питания — 18 В
Максимальный потребляемый ток — 4,5 А
Ток покоя (Еп=14,4 В) — 120 мА
Выходная мощность (Еп=14,4 В, КНИ=10%):
RL=4 Ом — 20 Вт
RL=8 Ом — 12 Вт
КНИ:
(Еп=14,4 В, RL=4 Ом, Рвых=12 Вт) — 0,1 %
(Еп=14,4 В, RL=8 Ом, Рвых=12Вт) — 0,05 %
Полоса пропускания по уровню -3 дБ (RL=4 Ом, Рвых=15 Вт) — 30…25000 Гц
Схема включения
TDA7241
Мостовой УНЧ, разработанный для применения в
автомобильных магнитолах. Имеет защиту от короткого замыкания в
нагрузке, а также от перегрева.
Максимальное напряжение питания — 18 В
Максимальный потребляемый ток — 4,5 А
Ток покоя (Еп=14,4 В) — 80 мА
Выходная мощность (Еп=14,4 В, КНИ=10%):
RL=2 Ом — 26 Вт
RL=4 Ом — 20 Вт
RL=8 Ом — 12 Вт
КНИ:
(Еп=14,4 В, RL=4 Ом, Рвых=12 Вт) — 0,1 %
(Еп=14,4 В, RL=8 Ом, Рвых=6 Вт) — 0.05 %
Полоса пропускания по уровню -3 дБ (RL=4 Ом, Рвых=15 Вт) — 30…25000 Гц
Схема включения
TDA1555Q
Напряжение питания — 6…18 B
Максимальный потребляемый ток — 4 А
Выходная мощность (Uп =14,4 В. RL=4 Ом):
— КНИ=0,5% — 5 Вт
— КНИ=10% — 6 Вт Ток покоя — 160 мА
Схема включения
TDA1557Q
Напряжение питания — 6. ..18 В
..18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Uп =14,4 В, RL=4 Ом):
— КНИ=0,5% — 17 Вт
— КНИ=10% — 22 Вт
Ток покоя, мА 80
Схема включения
TDA1556Q
Напряжение питания -6…18 В
Максимальный потребляемый ток -4 А
Выходная мощность: (Uп=14.4 В, RL=4 Ом):
— КНИ=0,5%, — 17 Вт
— КНИ=10% — 22 Вт
Ток покоя — 160 мА
Схема включения
TDA1558Q
Напряжение питания — 6..18 В
Максимальный потребляемый ток — 4 А
Выходная мощность (Uп=14 В, RL=4 Ом):
— КНИ=0. 6% — 5 Вт
6% — 5 Вт
— КНИ=10% — 6 Вт
Ток покоя — 80 мА
Схема включения
TDA1561
Напряжение питания — 6…18 В
Максимальный потребляемы ток — 4 А
Выходная мощность (Uп=14В, RL=4 Ом):
— КНИ=0.5% — 18 Вт
— КНИ=10% — 23 Вт
Ток покоя — 150 мА
Схема включения
TDA1904
Напряжение питания — 4…20 В
Максимальный потребляемы ток — 2 А
Выходная мощность (RL=4 Ом, КНИ=10%):
— Uп=14 В — 4 Вт
— Uп=12В — 3,1 Вт
— Uп=9 В — 1,8 Вт
— Uп=6 В — 0,7 Вт
КНИ (Uп=9 В, P<1,2 Вт, RL=4 Ом) — 0,3 %
Ток покоя — 8. ..18 мА
..18 мА
Схема включения
TDA1905
Напряжение питания — 4…30 В
Максимальный потребляемы ток — 2,5 А
Выходная мощность (КНИ=10%)
— Uп=24 В (RL=16 Ом) — 5,3 Вт
— Uп=18В (RL=8 Ом) — 5,5 Вт
— Uп=14 В (RL=4 Ом) — 5,5 Вт
— Uп=9 В (RL=4 Ом) — 2,5 Вт
КНИ (Uп=14 В, P<3,0 Вт, RL=4 Ом) — 0,1 %
Ток покоя — <35 мА
Схема включения
TDA1910
Напряжение питания — 8…30 В
Максимальный потребляемы ток — 3 А
Выходная мощность (КНИ=10%):
— Uп=24 В (RL=8 Ом) — 10 Вт
— Uп=24 В (RL=4 Ом) — 17,5 Вт
— Uп=18 В (RL=4 Ом) — 9,5 Вт
КНИ (Uп=24 В, P<10,0 Вт, RL=4 Ом) — 0,2 %
Ток покоя — <35 мА
Схема включения
TDA2003
Напряжение питания — 8.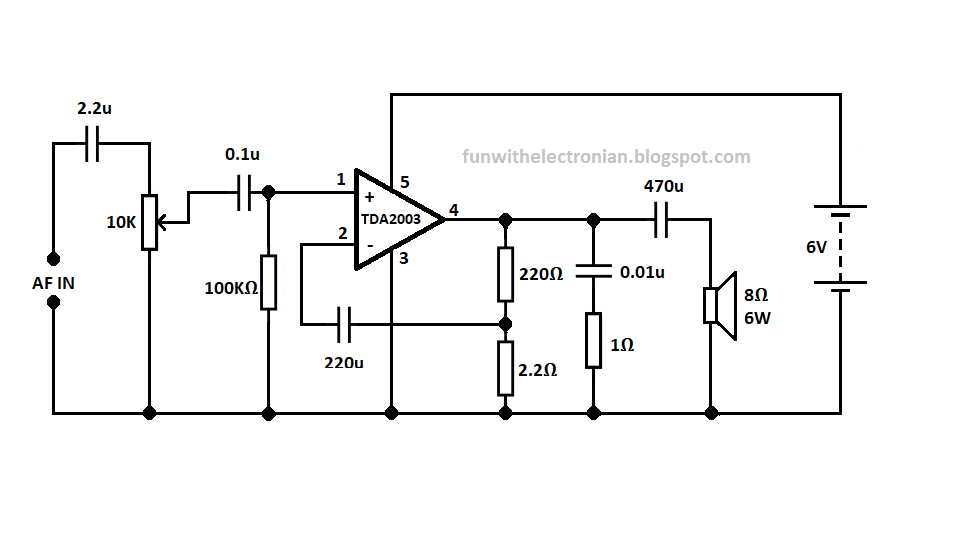 ..18 В
..18 В
Максимальный потребляемы ток — 3,5 А
Выходная мощность (Uп=14В, КНИ=10%):
— RL=4,0 Ом — 6 Вт
— RL=3,2 Ом — 7,5 Вт
— RL=2,0 Ом — 10 Вт
— RL=1,6 Ом — 12 Вт
КНИ (Uп=14,4 В, P<4,5 Вт, RL=4 Ом) — 0,15 %
Ток покоя — <50 мА
Схема включения
| STK021, STK024, STK031, STK035 Интегральные микросхемы STK021, STKO24, STK031 и STK035 фирмы Sanyo выполнены в корпусах SIP10 с 10 выводами и представляют собой усилители мощности низкой частоты в гибридном исполнении с идентичными схемами (цоколевками) и различными параметрами. Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса.
Интегральные микросхемы STK030, STK058, STK075, STK077, STK078, STK080, STK082, STK083, STK084 и STK086 фирмы Sanyo выполнены в корпусах SIP10 с 10 выводами и представляют собой усилители мощности низкой частоты в гибридном исполнении с идентичными схемами (цоколевками) и различными параметрами.
Интегральные микросхемы STK050 и STK070 фирмы Sanyo выполнены в корпусах SIP 10 с 16 выводами и представляют собой усилители мощности низкой частоты в гибридном исполнении с идентичными схемами (цоколевками) и различными параметрами.
Интегральные микросхемы фирмы STK075G, STK077G, STK078G, STK080G,STK082G, STK084G, STK085, STK086G фирмы Sanyo выполнены в корпусах SIP 10 с 10 выводами и представляют собой усилители мощности низкой частоты в гибридном исполнении с идентичными схемами (цоколевками) и различными параметрами.
Интегральные микросхемы STK0292, STK0352 и STK0452 фирмы Sanyo выполнены в корпусах SIP10 с 10 выводами и представляют собой выходные модули усилителей мощности низкой частоты в гибридном исполнении с идентичными схемами (цоколевками) и различными параметрами.
Перечисленные микросхемы фирмы Sanyo выполнены в корпусах SIP10 с 16 выводами и представляют собой двухканальные усилители мощности низкой частоты в гибридном исполнении с идентичными схемами (цоколевками) и различными параметрами.
Перечисленные микросхемы фирмы Sanyo выполнены в корпусах SIP10 с 16 выводами и представляют собой двухканальные (стереофонические) усилители мощности низкой частоты в гибридном исполнении с идентичными схемами (цоколевками) и различными параметрами. Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем (выходные параметры для одного канала) следующие:
Интегральные микросхемы STK1030, STK1040, STK1050, STK1050II, STK1060, STK1060II, STK1070, STK1070II, STK1080II и STK1100II фирмы Sanyo выполнены в корпусах SIP10 с 10 выводами и представляют собой выходные модули усилителей мощности низкой частоты в гибридном исполнении с идентичными схемами (цоколевками) и различными параметрами.
Источник: radvs.  boom.ru boom.ru |
 В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие:
В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие: Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса с двухполярным питанием. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие:
Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса с двухполярным питанием. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие: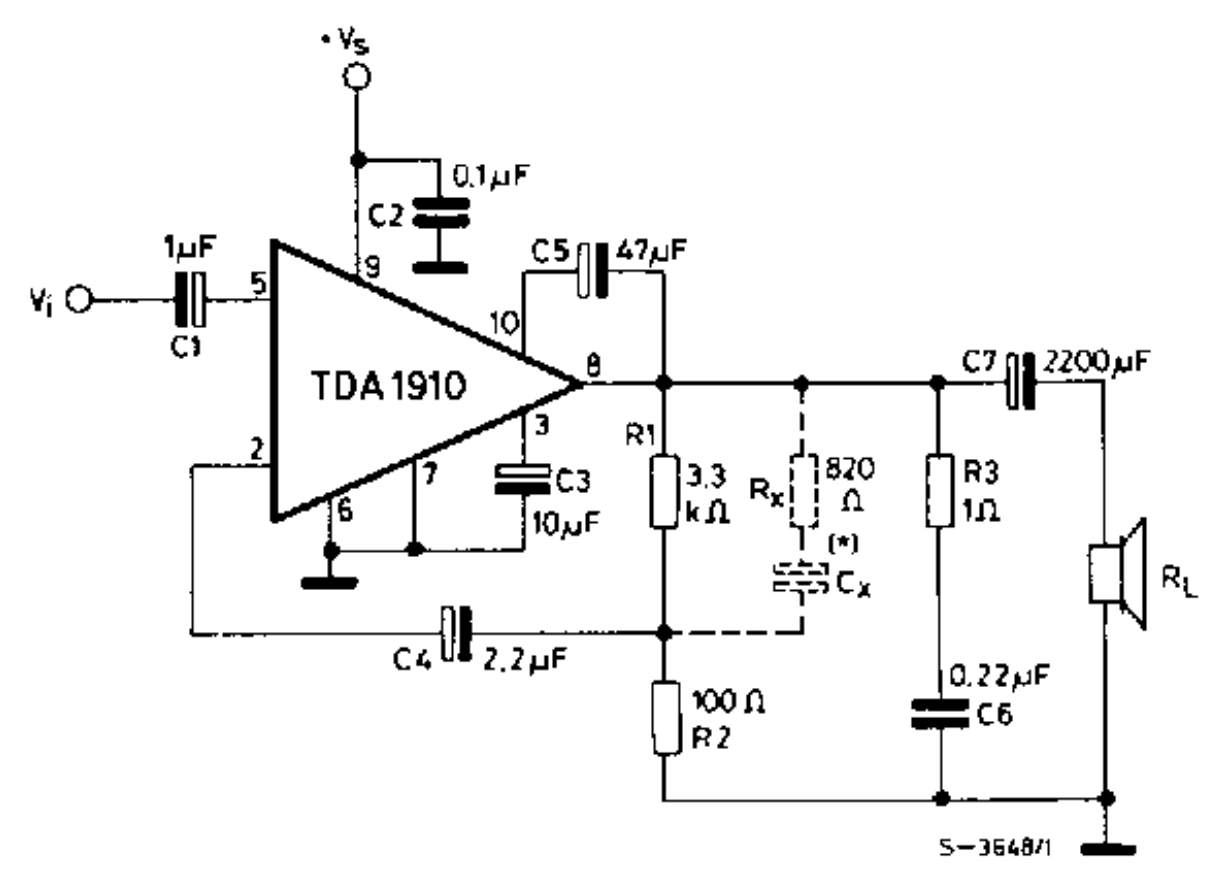 Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса с двухполярным питанием. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие:
Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса с двухполярным питанием. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие: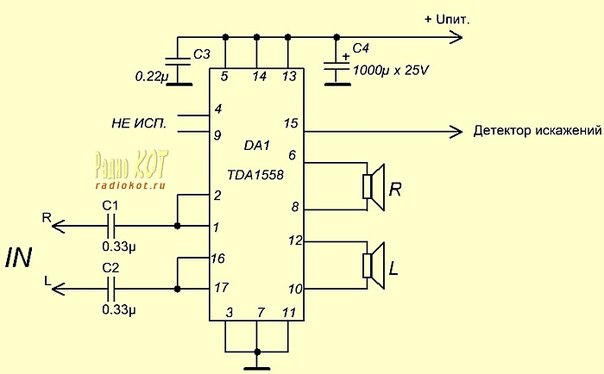 Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса с двухполярным питанием. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие:
Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса с двухполярным питанием. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие: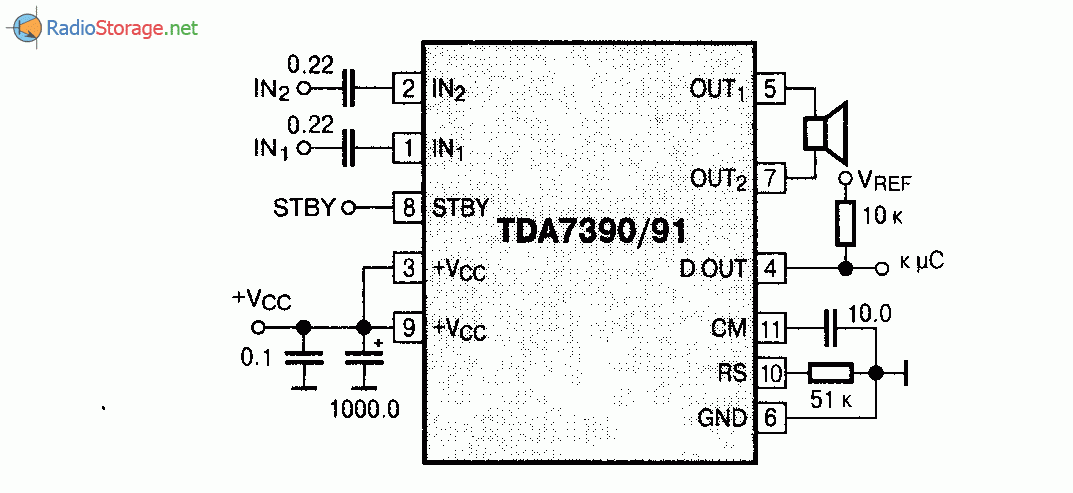 Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса с двухполярным питанием. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие:
Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса с двухполярным питанием. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем следующие: Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем (выходные параметры для одного канала) следующие:
Предназначены для использования в магнитофонах, электрофонах, телевизионных и радиоприемниках, другой аудиоаппаратуре высокого класса. В микросхемах отсутствует защита выхода от короткого замыкания в нагрузке. Для получения максимальной выходной мощности микросхемы необходимо установить на теплоотвод (радиатор). Некоторые из основных параметров микросхем (выходные параметры для одного канала) следующие: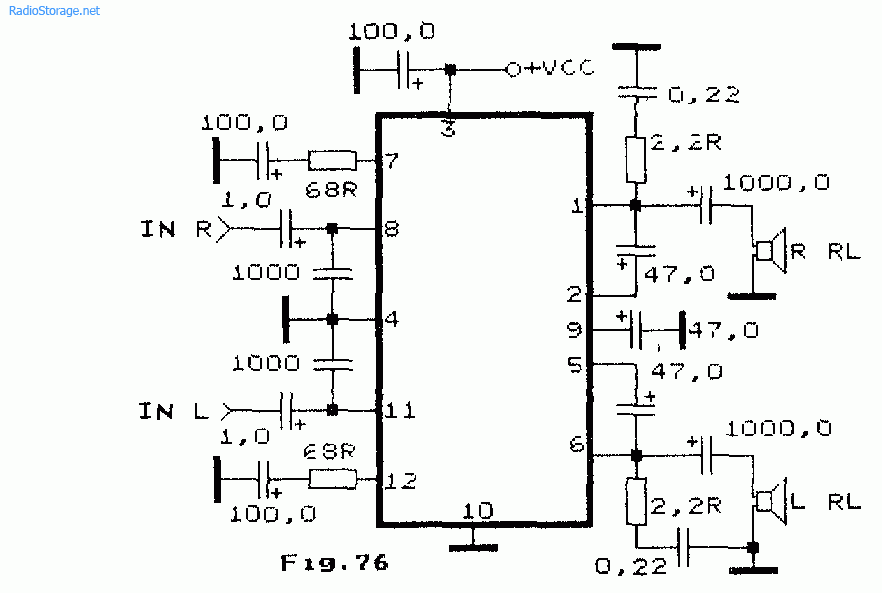 5W
5W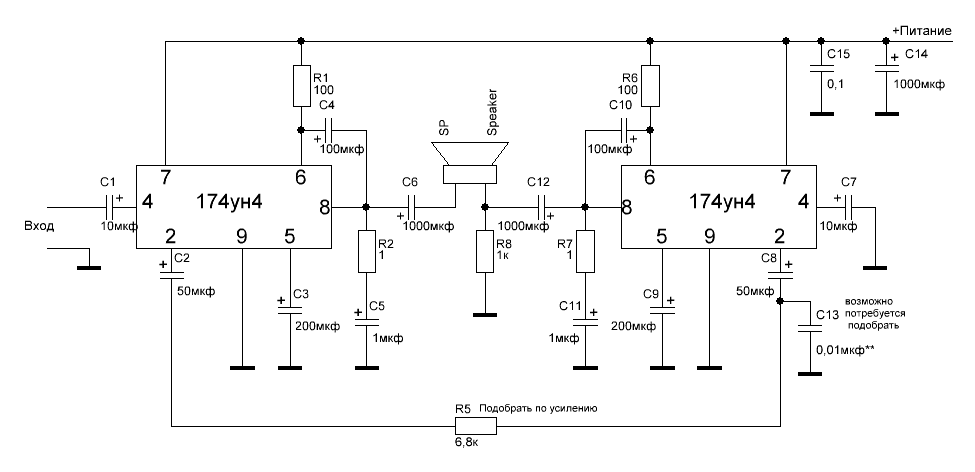 02%
02%Расшифровка схем сознания: от локальных микросхем до сетей масштаба мозга
Обзор
. 2020 1 апреля; 4 (2): 315-337. DOI: 10.1162 / netn_a_00119. Электронная коллекция 2020.Принадлежности Расширять
Принадлежность
- 1 Университет Ренна, INSERM, LTSI-U1099, Ренн, Франция.
Элемент в буфере обмена
Обзор
Жюльен Модоло и др. Netw Neurosci. .
Netw Neurosci. .
Показать варианты
Формат АннотацияPubMedPMID
.2020 1 апреля; 4 (2): 315-337. DOI: 10.1162 / netn_a_00119. Электронная коллекция 2020.Принадлежность
- 1 Университет Ренна, INSERM, LTSI-U1099, Ренн, Франция.

Элемент в буфере обмена
Полнотекстовые ссылки Опции CiteDisplayПоказать варианты
Формат АннотацияPubMedPMID
Абстрактный
Выявление физиологических процессов, лежащих в основе возникновения и поддержания сознания, является одной из самых фундаментальных проблем нейробиологии, с последствиями, варьирующимися от фундаментальной нейробиологии до лечения пациентов с расстройствами сознания (DOC).Одна из основных проблем — понять, как корковые цепи в совершенно разных пространственных масштабах, от локальных сетей до сетей в масштабе мозга, работают согласованно, чтобы обеспечить сознание, и как эти процессы нарушаются у пациентов с DOC. В этом обзоре мы пытаемся связать имеющиеся нейрофизиологические и клинические данные с существующими теоретическими моделями сознания, связывая при этом уровни микро- и макроцепей. Во-первых, мы обращаемся к отношениям между осознаванием и бодрствованием, с одной стороны, и кортико-корковой и таламо-корковой связью, с другой стороны.Во-вторых, мы обсуждаем роль трех основных типов ГАМКергических интернейронов в конкретных цепях, ответственных за динамическую реорганизацию функциональных сетей. В-третьих, мы исследуем достижения в функциональной роли вложенных колебаний для нейронной синхронизации и коммуникации, подчеркивая важность баланса между локальной (высокочастотной) и удаленной (низкочастотной) активностью для эффективной обработки информации. Представлены клинические последствия этих теоретических соображений.Мы предполагаем, что такие механизмы клеточного масштаба могли бы расширить существующие теории сознания.
В этом обзоре мы пытаемся связать имеющиеся нейрофизиологические и клинические данные с существующими теоретическими моделями сознания, связывая при этом уровни микро- и макроцепей. Во-первых, мы обращаемся к отношениям между осознаванием и бодрствованием, с одной стороны, и кортико-корковой и таламо-корковой связью, с другой стороны.Во-вторых, мы обсуждаем роль трех основных типов ГАМКергических интернейронов в конкретных цепях, ответственных за динамическую реорганизацию функциональных сетей. В-третьих, мы исследуем достижения в функциональной роли вложенных колебаний для нейронной синхронизации и коммуникации, подчеркивая важность баланса между локальной (высокочастотной) и удаленной (низкочастотной) активностью для эффективной обработки информации. Представлены клинические последствия этих теоретических соображений.Мы предполагаем, что такие механизмы клеточного масштаба могли бы расширить существующие теории сознания.
Ключевые слова: Коммуникация через согласованность; Расстройства сознания; Электроэнцефалография; Функциональная связность; Стробирование по запрещению; Микросхемотехника.
© 2019 Массачусетский технологический институт.
Цифры
Рисунок 1.Бодрствование и осведомленность — два…
Рисунок 1.Бодрствование и осознанность — два основных измерения сознания.На этой диаграмме несколько…
Рисунок 1. Бодрствование и осознанность — два основных измерения сознания. На этой диаграмме несколько качественно различных состояний сознания расположены на двумерной матрице в зависимости от связанных осей «содержание сознания» (осведомленность) и «уровень сознания» (бодрствование). По материалам Laureys (2005).
По материалам Laureys (2005).
Базовые (принципиальные) схемы, участвующие в…
Рисунок 2.Базовые (принципиальные) схемы, участвующие в генерации локальных и дальних колебаний. В…
Рисунок 2. Базовые (принципиальные) схемы, участвующие в генерации локальных и дальних колебаний.В локальной корковой сети (кортикальном столбе), щелевом соединении, взаимное ингибирование «быстрых» ГАМКергических интернейронов является одним из основных механизмов генерации локальной гамма-активности, наряду с ПИНГ (пирамида-интернейрон гамма). Напротив, петля обратной связи между проецирующими дендриты «медленными» ГАМКергическими интернейронами и пирамидными клетками может генерировать низкочастотную активность. Важно отметить, что дистанционное общение посредством дисинаптического пути позволяет кратковременно генерировать гамма-колебания в отдаленных популяциях.Пирамидные клетки в исходной популяции (левая цепь) проецируются на пирамидные клетки отдаленной популяции (справа), но также и на интернейроны VIP, которые проецируются на нейроны SST, проецирующие дендриты. Временная активация VIP-нейронов из исходной популяции временно подавляет SST-нейроны в целевой популяции, позволяя генерировать гамма-колебания через PV-PV и пирамидно-PV-цепь. Как только вход на удаленные VIP-нейроны уменьшается, SST-нейроны возобновляют свой тормозной вход, который может прекратить генерацию гамма-колебаний.
Важно отметить, что дистанционное общение посредством дисинаптического пути позволяет кратковременно генерировать гамма-колебания в отдаленных популяциях.Пирамидные клетки в исходной популяции (левая цепь) проецируются на пирамидные клетки отдаленной популяции (справа), но также и на интернейроны VIP, которые проецируются на нейроны SST, проецирующие дендриты. Временная активация VIP-нейронов из исходной популяции временно подавляет SST-нейроны в целевой популяции, позволяя генерировать гамма-колебания через PV-PV и пирамидно-PV-цепь. Как только вход на удаленные VIP-нейроны уменьшается, SST-нейроны возобновляют свой тормозной вход, который может прекратить генерацию гамма-колебаний.
Предлагаемый синтез того, как переходный процесс…
Рисунок 3.
Предлагаемый синтез того, как происходит временное избирательное связывание между корковыми сетями через…
Рисунок 3. Предлагаемый синтез того, как временное избирательное связывание между корковыми сетями происходит через механизмы клеточного масштаба. (A) Схематическая диаграмма анатомической сети с основными проекциями между областями в масштабе мозга. (B) На верхней панели селективное связывание (коммуникация через когерентность; Fries, 2015) в подмножестве областей коры происходит посредством генерации гамма-колебаний (в основном через микросхемы с участием корзинчатых клеток), в то время как удаленное растормаживание определенных областей мозга происходит через дисинаптические растормаживание (активация удаленных VIP-нейронов ингибирует SST-нейроны, которые, в свою очередь, уменьшают их тормозную проекцию на пирамидные нейроны).Это способствует преобразованию анатомической сети в функциональную сеть. Альфа-ритм действует как импульсное торможение, подавляя «нерелевантные» сети, дополнительно увеличивая отношение сигнал / шум (SNR). На нижней панели снижение интеграции (например, после повреждения головного мозга) приводит к нарушению синхронизации удаленных областей мозга (что отражается снижением низкочастотного ритма на иллюстративном колебании), и, таким образом, уменьшение связывания, что в совокупности приводит к снижению GBI. (Jensen & Mazaheri, 2010) и сильно снижает общее SNR, что приводит к дисфункции сети с точки зрения интеграции и связывания, необходимых для сознания.Отметим, что помимо уменьшения амплитуды низкочастотного ритма, соотношение фаз вложенного гамма-колебания могло быть нарушено (т. Е. Более случайным) по сравнению с физиологическим случаем. Такую потенциальную взаимосвязь вложенных тета / гамма-колебаний еще предстоит изучить в DOC.
Альфа-ритм действует как импульсное торможение, подавляя «нерелевантные» сети, дополнительно увеличивая отношение сигнал / шум (SNR). На нижней панели снижение интеграции (например, после повреждения головного мозга) приводит к нарушению синхронизации удаленных областей мозга (что отражается снижением низкочастотного ритма на иллюстративном колебании), и, таким образом, уменьшение связывания, что в совокупности приводит к снижению GBI. (Jensen & Mazaheri, 2010) и сильно снижает общее SNR, что приводит к дисфункции сети с точки зрения интеграции и связывания, необходимых для сознания.Отметим, что помимо уменьшения амплитуды низкочастотного ритма, соотношение фаз вложенного гамма-колебания могло быть нарушено (т. Е. Более случайным) по сравнению с физиологическим случаем. Такую потенциальную взаимосвязь вложенных тета / гамма-колебаний еще предстоит изучить в DOC.

Синтез механизмов сетевого уровня…
Рисунок 4.Синтез механизмов сетевого уровня, лежащих в основе сложности, связанной с сознательными процессами. The…
Рисунок 4.Синтез механизмов сетевого уровня, лежащих в основе сложности, связанной с сознательными процессами. Структурные характеристики мозговых цепей на разных уровнях упоминаются с некоторыми ключевыми колебательными ритмами и связанными с ними функциями.
Похожие статьи
- COALIA: вычислительная модель человеческой ЭЭГ для исследования сознания.
Бенсаид С.
 , Модоло Дж., Мерле I, Вендлинг Ф., Бенке П.
Bensaid S, et al.
Front Syst Neurosci. 2019 Ноябрь 13; 13:59. DOI: 10.3389 / fnsys.2019.00059. Электронная коллекция 2019.
Front Syst Neurosci. 2019.
PMID: 31798421
Бесплатная статья PMC.
, Модоло Дж., Мерле I, Вендлинг Ф., Бенке П.
Bensaid S, et al.
Front Syst Neurosci. 2019 Ноябрь 13; 13:59. DOI: 10.3389 / fnsys.2019.00059. Электронная коллекция 2019.
Front Syst Neurosci. 2019.
PMID: 31798421
Бесплатная статья PMC. - Нейронные корреляты сознания и родственные расстройства: от фенотипических дескрипторов поведенческого и относительного сознания до кортико-подкорковых цепей.
Лемэр Дж. Дж., Понтье Б., Ша Р., Эль-Уадих И., Халил Т., Синардет Д., Ахим В., Постельнику А., Косте Дж., Жермен В., Саррет К., Сонтхаймер А. Lemaire JJ и др. Нейрохирургия. 2021, 26 мая: S0028-3770 (21) 00156-9.DOI: 10.1016 / j.neuchi.2021.05.003. Интернет впереди печати. Нейрохирургия. 2021 г. PMID: 34051246 Рассмотрение.
- Новый свет на диагностику расстройств сознания: динамическая функциональная связь.
Наро А, Браманти А, Лео А, Каччола А, Манули А, Браманти П., Калабро Р.С. Наро А. и др. Cortex. 2018 июн; 103: 316-328. DOI: 10.1016 / j.cortex.2018.03.029. Epub 2018 4 апр. Cortex. 2018. PMID: 29684751
- Снижение интеграции пространственно-источниковых сетей ЭЭГ при расстройствах сознания.
Ризкаллах Дж., Аннен Дж., Модоло Дж., Госсерис О, Бенке П, Мортахеб С., Амуд Х, Кассол Х, Мейх А, Тибо А, Шатель С, Хассан М, Панда Р, Вендлинг Ф, Лори С. Ризкаллах Дж. И др. Neuroimage Clin. 2019; 23: 101841.DOI: 10.1016 / j.nicl.2019.101841. Epub 2019 29 апреля. Neuroimage Clin. 2019. PMID: 31063944 Бесплатная статья PMC.
- Пробуждение и восстановление сознания при расстройствах сознания: есть ли роль наркотики?
Пистойя Ф.
 , Мура Э., Говони С., Фини М., Сара М.
Пистойя Ф. и др.
Препараты ЦНС. 2010 августа; 24 (8): 625-38. DOI: 10.2165 / 11535940-000000000-00000.
Препараты ЦНС.2010 г.
PMID: 20658796
Рассмотрение.
, Мура Э., Говони С., Фини М., Сара М.
Пистойя Ф. и др.
Препараты ЦНС. 2010 августа; 24 (8): 625-38. DOI: 10.2165 / 11535940-000000000-00000.
Препараты ЦНС.2010 г.
PMID: 20658796
Рассмотрение.
Процитировано
2 статей- Крупномасштабные функциональные сети мозга для сознания.
Хан МЭ, Пак SY, О, ТАК. Хан МЭ и др. Anat Cell Biol. 2021, 30 июня; 54 (2): 152-164. DOI: 10.5115 / acb.20.305. Anat Cell Biol. 2021 г. PMID: 33967030 Бесплатная статья PMC. Рассмотрение.
- Состояние сегрегированного мозга во время гипноза.
Туоминен Дж., Каллио С., Каасинен В., Раило Х. Туоминен Дж.
 И др.
Neurosci Conscious. 2021 10 марта; 2021 (1): niab002. DOI: 10,1093 / NC / Niab002. Электронная коллекция 2021 г.
Neurosci Conscious. 2021 г.
PMID: 33747546
Бесплатная статья PMC.
И др.
Neurosci Conscious. 2021 10 марта; 2021 (1): niab002. DOI: 10,1093 / NC / Niab002. Электронная коллекция 2021 г.
Neurosci Conscious. 2021 г.
PMID: 33747546
Бесплатная статья PMC.
Рекомендации
- Адамс Дж.Х., Грэм Д. И. и Дженнет Б. (2000). Невропатология вегетативного состояния после острого инсульта головного мозга. Мозг, 123, 1327–1338. — PubMed
- Аллитт Б. Дж., Ива П., Ян Э. Б. и Раджан Р. (2016). Гиповозбуждение по всем кортикальным пластинкам определяет промежуточные стадии дисфункции корковых нейронов при диффузной черепно-мозговой травме.
 Неврология, 334, 290–308. 10.1016 / j.neuroscience.2016.08.018
—
DOI
—
PubMed
Неврология, 334, 290–308. 10.1016 / j.neuroscience.2016.08.018
—
DOI
—
PubMed
- Аллитт Б. Дж., Ива П., Ян Э. Б. и Раджан Р. (2016). Гиповозбуждение по всем кортикальным пластинкам определяет промежуточные стадии дисфункции корковых нейронов при диффузной черепно-мозговой травме.
- Алвис Д.С., Ян Э. Б., Джонстон В., Каррон С., Хеллуэлл С., Морганти-Косманн М. К. и Раджан Р. (2016). Обогащение окружающей среды уменьшает черепно-мозговые травмы: индуцированная гипервозбудимость нейронов в надгранулярных слоях сенсорной коры. Журнал нейротравмы, 33 (11), 1084–1101. 10.1089 / neu.2014.3774 — DOI — PubMed
- Алвис Д.
 С., Ян Э. Б., Морганти-Косманн М. К. и Раджан Р. (2012). Сенсорная кора головного мозга, лежащая в основе дефицита черепно-мозговой травмы. PLoS One, 7 (12), e52169 10.1371 / journal.pone.0052169
—
DOI
—
ЧВК
—
PubMed
С., Ян Э. Б., Морганти-Косманн М. К. и Раджан Р. (2012). Сенсорная кора головного мозга, лежащая в основе дефицита черепно-мозговой травмы. PLoS One, 7 (12), e52169 10.1371 / journal.pone.0052169
—
DOI
—
ЧВК
—
PubMed
- Алвис Д.
- Арсивалла X.D., & Verschure P. (2018). Измерение сложности сознания. Границы неврологии, 12, 424 10.3389 / fnins.2018.00424 — DOI — ЧВК — PubMed
Показать все 133 ссылки
[Икс]цитировать
КопироватьФормат: AMA APA ГНД NLM
Гипотеза о контроле частоты тета-ритма в микросхемах CA1
РЕЗЮМЕ
Вычислительные модели нейронных цепей с различными уровнями биофизической детализации были созданы в поисках основного механизма, объясняющего повсеместный тета-ритм в гиппокампе. Однако внутри тета-ритма есть по крайней мере два типа с различными частотами, связанными с разными поведенческими состояниями, аспект, который необходимо учитывать в поисках этих механистических объяснений. Здесь, используя наши ранее разработанные модели возбуждающих-тормозных сетей, которые генерируют тета-ритмы, мы исследуем устойчивость тета-генерации к внутренней изменчивости нейронов, создавая базу данных гетерогенных возбуждающих клеток и внедряя их в нашу модель микросхемы.Мы специально исследуем влияние трех ключевых особенностей модели возбуждающих клеток, лежащих в основе нашей модели: реобазы этих клеток, их способности к постингибиторному отскоку и их адаптации к частоте всплесков. Мы показываем, что тета-ритмы на различных частотах могут возникать в зависимости от комбинации этих свойств строительных блоков, и мы обнаруживаем, что скорость этих колебаний зависит от реакции возбуждающих клеток на тормозное возбуждение, что отражено в их кривых фазовой реакции.Взятые вместе, эти результаты подтверждают гипотезу о контроле тета-частоты, которая включает два аспекта: (i) внутренний механизм, который проистекает из особенностей строительных блоков динамики возбуждающих клеток; (ii) внешний механизм, который мы описываем как «основанную на ингибировании настройку» возбуждения возбуждающих клеток. Мы предполагаем, что эти механизмы контролируют частоты тета-ритма и лежат в основе их устойчивости.
Однако внутри тета-ритма есть по крайней мере два типа с различными частотами, связанными с разными поведенческими состояниями, аспект, который необходимо учитывать в поисках этих механистических объяснений. Здесь, используя наши ранее разработанные модели возбуждающих-тормозных сетей, которые генерируют тета-ритмы, мы исследуем устойчивость тета-генерации к внутренней изменчивости нейронов, создавая базу данных гетерогенных возбуждающих клеток и внедряя их в нашу модель микросхемы.Мы специально исследуем влияние трех ключевых особенностей модели возбуждающих клеток, лежащих в основе нашей модели: реобазы этих клеток, их способности к постингибиторному отскоку и их адаптации к частоте всплесков. Мы показываем, что тета-ритмы на различных частотах могут возникать в зависимости от комбинации этих свойств строительных блоков, и мы обнаруживаем, что скорость этих колебаний зависит от реакции возбуждающих клеток на тормозное возбуждение, что отражено в их кривых фазовой реакции.Взятые вместе, эти результаты подтверждают гипотезу о контроле тета-частоты, которая включает два аспекта: (i) внутренний механизм, который проистекает из особенностей строительных блоков динамики возбуждающих клеток; (ii) внешний механизм, который мы описываем как «основанную на ингибировании настройку» возбуждения возбуждающих клеток. Мы предполагаем, что эти механизмы контролируют частоты тета-ритма и лежат в основе их устойчивости.
1 ВВЕДЕНИЕ
Тета-ритмы гиппокампа (≈ 3-12 Гц), наблюдаемые в записях потенциала локального поля (LFP), связаны с когнитивными процессами формирования памяти и пространственной навигации (Colgin, 2013, 2016; Hinman et al., 2018). Как именно возникают тета-ритмы — сложная и многослойная проблема, но известно, что существует два типа, обозначенные типом 1 и типом 2, которые имеют высокие (7-12 Гц) или низкие (4-7 Гц) частоты соответственно. . Ритмы 2-го, но не 1-го типа зависят от холинергического влечения (Bland, 1986; Buzsáki, 2002; Kramis et al., 1975). У грызунов было показано, что социальные стимулы вызывают высокий тета, а пугающие стимулы вызывают низкий тета (Tendler and Wagner, 2015), а тета-осцилляции 2 типа связаны с повышенным рискованным поведением (Mikulovic et al., 2018). У людей тета-частоты в целом ниже (Jacobs, 2014), но все еще можно различать высокие и низкие тета-частоты, при этом низкие тета поддерживают кодирование и извлечение воспоминаний (Kota et al., 2020). Ясно, что регулировка тета-частоты функционально важна.
В настоящее время хорошо задокументировано, что тета-ритмы могут генерироваться внутри гиппокампа, возникая спонтанно из изолированного цельного препарата гиппокампа in vitro (Goutagny et al., 2009). Одновременный доступ к клеточной и популяционной продукции дает возможность распутать клеточную и популяционную динамику генерации тета-ритмов.В предыдущей работе мы воспользовались этим и построили модели клеток и микросхем, которые могут генерировать тета-ритмы с параметрами, напрямую ограниченными экспериментальными данными из всего препарата гиппокампа и экспериментальной литературой (Ferguson et al., 2013, 2015a, 2017). На основе точки зрения, представленной Gjorgjieva et al. (2016), мы рассмотрели подход к анализу «строительных блоков для динамики схемы» при проектировании нашей модели микросхемы (Ferguson et al., 2017). С этой точки зрения, биологически известные клеточные, синаптические и связные характеристики рассматриваются как строительные блоки для динамики схемы.Например, одним из таких клеточных «строительных блоков» является постингибиторный отскок (PIR), который ранее рассматривался как участник генерации кортикальных колебаний (McCormick et al., 2015).
В этой статье мы используем нашу модель тета-генерирующей микросхемы, чтобы разработать гипотезу о том, как можно управлять тета-частотами. Сначала мы описываем модель микросхемы, а затем оцениваем надежность генерации тета в модели, рассматривая гетерогенные пирамидальные (PYR) популяции клеток.Исходя из этого, мы используем кривые фазовой характеристики (PRC) и показываем, что тормозящие входы влияют на тета-частоту. Таким образом, мы предлагаем гипотезу для контроля тета-частоты в микросхемах CA1, которая зависит от внутренних характеристик клеток PYR и «основанной на ингибировании настройки» активации клеток PYR. Мы резюмируем наше исследование в схематической форме в Рис. 1.
Рис. 1. Схема, показывающая аспекты, участвующие в гипотезе, разработанной в этом исследовании.Тета-ритмы генерируются в целом в препарате гиппокампа Goutagny et al.(2009) («Эксперимент»). Их поколение отражено в модели микросхем, разработанной Фергюсоном и др. (2017) («Модельные сети»). В настоящей статье мы оцениваем надежность этой модели и разрабатываем гипотезу для контроля тета-частоты («Развитие гипотезы»).
2 КОНСТРУКЦИЯ МОДЕЛЕЙ МИКРОСХЕМЫ, ПРОИЗВОДИВАЮЩЕЙ ТЕТА-РИТМЫ
Мы построили клеточные модели возбуждающе-ингибирующей (EI) сети (Ferguson et al., 2017), чтобы понять, как внутренние тета-ритмы наблюдаются во всем препарате гиппокампа. Goutagny et al.(2009) могут быть созданы. Модельные сети (см. Схему Рис. 1) предназначены для представления «кусочка» области CA1 гиппокампа — приблизительно один мм 3 , которого было определено достаточно для самогенерирования тета-ритмов. Он включает только два различных типа клеток, пирамидные (PYR) клетки и быстровоспламеняющиеся парвальбумин-положительные (PV +) клетки, что представлено моделью одного отсека со структурой математической модели Ижикевича (Ижикевич, 2006). Модельная сеть состоит из 10 500 ячеек (10 000 ячеек PYR и 500 быстродействующих ячеек PV +) (Ferguson et al., 2013, 2015б). Мы отмечаем, что мы воспользовались масштабным соотношением между числом клеток, вероятностью соединения и возбуждающим синаптическим весом, что позволило нам использовать 10 000 клеток PYR, а не размер числа 30 000 клеток, рассчитанный для «кусочка» ткани.
Мы изучили наши модели с точки зрения «строительного блока для динамики контуров» (Gjorgjieva et al., 2016), чтобы определить, могут ли тета-ритмы (то есть всплески популяции тета-частоты) генерироваться в соответствии с экспериментальными ограничениями.Сначала мы обнаружили, что экспериментально ограниченные модели сотовых сетей PYR (только сети E-cell) могут генерировать всплески тета-частоты в популяции (Ferguson et al., 2015b), предполагая, что присутствует функция клеточного «строительного блока» адаптации частоты всплесков (SFA). в ограниченных моделях клеток PYR может вносить важный вклад в генерацию тета-ритма. Однако мы также обнаружили, что в этих сетях только E-клеток клетки PYR не срабатывают редко, как это наблюдалось экспериментально (Huh et al., 2016). Когда мы включили клетки PV + для создания сетей моделей E-I, всплески тета-частоты популяции все еще были возможны и теперь были связаны с разреженным срабатыванием клеток PYR в соответствии с экспериментальными данными. Поскольку добавление PV + клеток позволяет PIR быть возможным в PYR-клетках, мы рассматриваем PIR как еще одну особенность строительных блоков, важную для генерации этих внутренних тета-ритмов. Наряду с характеристиками SFA и PIR нейроны PYR обладают свойственной реобазой (Rheo) характеристикой, которая представляет собой величину тока, необходимого для создания пика клеток PYR (полученного в результате подгонки к экспериментальным данным в Ferguson et al.(2015a)). Мы считаем, что это третий строительный элемент для генерации тета-ритма. Кроме того, для того, чтобы выходные данные модели согласовывались с экспериментальными наблюдениями соотношений амплитуд возбуждающего постсинаптического тока (EPSC) и тормозного постсинаптического тока (IPSC), мы обнаружили, что вероятность соединения от PV + к клеткам PYR должна быть больше, чем от PYR к PV +. клетки — конкретное предсказание, которое было исследовано и признано соответствующим эмпирически полученным связям (Chatzikalymniou et al., 2020).
3 ОЦЕНКА КОНСТРУКЦИИ МОДЕЛИ НАДЕЖНЫХ ТЕТА-РИТМОВ
В нашей предыдущей работе мы специально не исследовали чувствительность тета-ритмов к функциям SFA, PIR или Rheo. Чтобы решить эту проблему, мы создаем базу данных моделей из 10 000 моделей ячеек PYR. Хотя существуют различные способы создания базы данных моделей, мы делаем это, просто изменяя значения конкретных параметров модели ячеек PYR обычным образом. Значения параметров модели ячейки PYR, определенные из подгонки к экспериментальным данным (Ferguson et al., 2015a) считаются значениями по умолчанию. Подробности создания модельной базы данных приведены в Приложении дополнительных материалов.
Из созданной базы данных моделей клеток PYR мы получаем различные характеристики SFA, PIR и Rheo. Мы определяем количественные характеристики характеристик SFA, PIR и Rheo следующим образом: чем больше количественное значение SFA, тем сильнее степень адаптации ячеек PYR, т. Е. Мы получаем большее снижение частоты всплесков ячеек PYR для фиксированного количества входной ток; чем отрицательнее количественное значение PIR, тем больше шаг гиперполяризации, необходимый для генерации всплеска в конце шага; чем больше количественное значение Рео, тем больше требуется ввода, чтобы вызвать всплеск клетки.Подробности приведены в Приложении к дополнительным материалам. Для модели ячейки PYR со значениями параметров по умолчанию, используемыми в Ferguson et al. (2017) количественные значения характеристик строительных блоков следующие: SFA = 0,46 Гц / пА, Rheo = 4,0 пА и PIR = −5,0 пА. Мы называем их «базовыми» значениями. Здесь, с созданной базой данных моделей ячеек PYR, мы получаем диапазон значений характеристик строительных блоков, распределенных, как показано на рис. Рис. 2. Более подробная информация представлена в Приложении к дополнительным материалам.
Рисунок 2. Распределение характеристик ячеек PYR из созданной базы данных модели.Был создан гетерогенный набор клеток PYR и количественно определены их характеристики «строительный блок» SFA, Rheo и PIR. Подробная информация об этой количественной оценке представлена в Приложении к дополнительным материалам. Гистограммы показывают количество появлений значений SFA [=] Hz / pA, Rheo [=] pA, PIR [=] pA, а вертикальные черные стрелки указывают базовые значения [SFA, Rheo, PIR]. Также показаны узкие (N) и широкие (B) подмножества гетерогенных популяций клеток PYR и низкие (L), средние (M) или высокие (H) подмножества гетерогенных популяций клеток PYR, которые включают или не включают значения базовых строительных блоков.Гистограмма SFA имеет разрешение ячейки 0,05, а гистограммы Rheo, PIR имеют разрешение ячейки 0,5.
В обширном моделировании сети E-I, проведенном Ferguson et al. (2017), использованные модели ячеек PYR были однородными, и все они имели значения параметров модели по умолчанию. Однако сами сети не были однородными из-за шумных внешних приводов моделей ячеек PYR. Чтобы проверить устойчивость механизма генерации тета в моделях сети EI к изменчивости функций SFA, PIR и Rheo, мы создаем гетерогенные популяции клеток PYR из базы данных модели и исследуем, влияет ли присутствие тета-ритмов в сетях EI. варьируя эти функции строительных блоков.
Мы проводим наше исследование таким образом, что гетерогенная популяция клеток PYR в сетях E-I либо включает, либо не включает клетки PYR, которые имеют базовые значения. Вкратце отметим, что когда мы исследуем сети EI, которые имеют однородные модели ячеек PYR со значениями параметров, отличными от значений по умолчанию, но которые имеют аналогичные базовые значения SFA, PIR и Rheo, результирующие сети создают явные всплески популяции, но немного вариации по частоте и мощности. Конкретные примеры приведены в Приложении к дополнительным материалам.
Для сетей E-I с гетерогенными популяциями клеток PYR, которые имеют клетки PYR от до , включая базовые значения SFA, Rheo и PIR, тета-ритмы продолжают экспрессироваться. Мы также обнаружили, что тета-мощность сети больше, когда существует узкий, а не широкий диапазон значений, включающий базовые. Рис. 2 показывает узкий и широкий диапазоны значений в нашей созданной базе данных. Более подробная информация представлена в Приложении к дополнительным материалам.Это наблюдение разницы тета-мощности предполагает, что определенные количественно определенные значения характеристик влияют на устойчивость тета-ритмов, поскольку мощность больше, когда она более узко охватывает базовые значения.
Для гетерогенных сетей EI, которые имеют ячейки PYR, которые делают , а не , включают базовые значения для всех функций, мы создаем сети EI, которые имеют низкий (L), средний (M) или высокий (H) диапазон значений для SFA, Rheo. и функции PIR в различных комбинациях. Таким образом, данная гетерогенная сеть E-I имеет тройку функций [SFA, Rheo, PIR], которые имеют диапазон значений L, M или H.Эти значения показаны на Рис. 2. На Рис. 3 мы показываем частоту (слева) и мощность (справа) выходных сигналов этих гетерогенных сетей E-I, обозначенных точками заданного цвета. Красная точка в кружке — это единственная сеть E-I, в которой действительно имеет базовые значения для всех функций строительных блоков, то есть [SFA, Rheo, PIR] = HML. Мы наблюдаем следующее для сетевой частоты: сети с Rheo = L не производят тета-ритмы, когда PIR и SFA = M или H; Нет тета-ритмов, когда Rheo = M значения, а SFA и PIR = H; По мере увеличения Rheo частота сети увеличивается, и, похоже, более сильный контроль частоты с помощью функции Rheo по сравнению с функциями SFA и PIR.Для тета-мощности мы обнаруживаем, что она самая низкая, когда Rheo = L, и увеличивается с увеличением Rheo, но уменьшается с увеличением SFA или PIR. Однако, когда Rheo = M, мощность увеличивается по мере увеличения SFA и уменьшения PIR. Исходя из этих тенденций, может показаться, что функция Rheo контролирует тета-частоту и мощность больше, чем SFA или PIR. Поскольку большие значения Rheo относятся к большим токам деполяризации, необходимым для срабатывания ячейки PYR, наши наблюдения подразумевают, что величина тока, необходимая для срабатывания ячейки PYR, является важным контроллером тета-частоты и мощности, предполагая, что другие функции допускают ритмы. существовать в первую очередь.Более подробная информация об этом исследовании представлена в Приложении к дополнительным материалам.
Рисунок 3. Частота и мощность тета-ритмов в гетерогенных сетях E-I.Каждая точка представляет частоту (слева) или мощность (справа) выходного сигнала сети, которая имеет функции [SFA, Rheo, PIR] с диапазоном значений L, M или H, как показано на графике, причем цвет точки представляет конкретный значение частоты или мощности, указанное на цветной шкале. Точка в красном кружке — это сеть, значения объектов которой включают базовые значения для всех функций, т.е.е., [SFA, Rheo, PIR] = HML. Темно-синие круги не дают ритмичного вывода, а вершины, не имеющие точек, находятся там, где не было отдельных моделей ячеек PYR для генерации конкретной гетерогенной сети. Более подробная информация представлена в Приложении к дополнительным материалам.
Таким образом, исследование нашей микросхемной модели генерации тета-ритма во всем препарате гиппокампа приводит нас к следующим выводам относительно влияния трех «строительных блоков» на эту динамику: (i) большая тета-мощность возникает в EI сети с гетерогенными клетками PYR, которые включают свои базовые значения и узко распределены вокруг них, и (ii) определенные значения тока реобазы контролируют частоту и мощность сетевых ритмов в большей степени, чем способность клетки PYR к всплеску при тормозящем отскоке или конкретном количестве адаптации частоты всплесков.Таким образом, эти симуляции сетей E-I с гетерогенными популяциями клеток PYR позволили нам оценить вклад различных функций и помогли нам подтвердить устойчивость к клеточной гетерогенности механизма тета-генерирующего ритма в нашей модели микросхем.
4 ИСПОЛЬЗОВАНИЕ ОЦЕНКИ И РАЗРАБОТКИ ДЛЯ РАЗРАБОТКИ ГИПОТЕЗЫ ДЛЯ КОНТРОЛЯ ТЕТАЧАСТОТЫ
Как описано выше, мы обнаруживаем, что большие, минимально связанные повторяющиеся сети с быстродействующими PV + клетками и PYR-клетками могут создавать популяционные ритмы с тета-частотой, согласующиеся с экспериментом, частично управляется и контролируется структурными элементами SFA, PIR и Rheo в ячейках PYR.В наших предыдущих сетевых моделях PV + ячеек только для I-cell когерентный сетевой вывод был возможен с экспериментально ограниченными моделями PV + клеток и синаптическими связностями (Ferguson et al., 2013). При создании сетевой модели E-I сеть ячеек PV + была «спроектирована» так, чтобы находиться в когерентном состоянии — функция соответствующего принимаемого возбуждающего импульса и возможности соединения ячеек PV +. В частности, мы выбрали синаптический вес (между PV + клетками) таким, чтобы он мог быть на «границе» когерентного срабатывания (высокая частота) или нет (см. Рис.3 в Ferguson et al. (2013)), и, как таковой, при соответствующем возбуждающем импульсе от клеток PYR, сеть PV + клеток может находиться в высокочастотном когерентном режиме и считается производящей тормозной «болюс» для клеток PYR. Это важное соображение при рассмотрении приведенной ниже кривой фазовой характеристики (PRC).
Из нескольких наборов моделей гетерогенных сетевых выходных данных модели E-I, описанных в предыдущем разделе, мы выбираем три, которые демонстрируют сильные популяционные ритмы разной частоты.Подробности об этих трех выбранных сетях (в частности, о гетерогенной популяции PYR, а также о классификации их ритмов как «сильных») можно найти в Приложении к дополнительным материалам. Выходные данные растровых графиков ячеек PYR в этих выбранных гетерогенных сетях E-I показаны на рис. Рис. 4, где различные ритмы называются «медленными», «средними» и «быстрыми». Учитывая минимальный характер модели микросхемы, частоты этих ритмов немного выходят за пределы тета-диапазонов (выше) для некоторых сетей, хотя лежащий в основе механизм генерации тета и конструкция модели одинаковы.
Рис. 4. Выходные данные ячеек PYR на растровом графике из трех разнородных моделей E-I.Эти три набора моделей, генерирующие ритмический выходной сигнал всплеска популяции, демонстрируют три разные частоты, которые мы называем «медленными» (9,6 Гц), «средними» (13 Гц) и «быстрыми» (15 Гц) из соответствующих наборов моделей. Для всех трех наборов гетерогенные ячейки PYR включают ячейки с базовыми значениями Rheo, тогда как только набор моделей, дающий «средний» выходной сигнал, имеет ячейки PYR с базовыми значениями SFA. За исключением набора моделей, производящего «медленный» вывод, ячейки PYR имеют базовые значения PIR.То есть триплет [SFA, Rheo, PIR] для медленных, средних и быстрых сетей — это MMH, HML и LML соответственно.
Давайте теперь воспользуемся преимуществами нашей конструкции микросхемы, чтобы изучить, как эти частоты контролируются, обратившись к соображениям PRC (Schultheiss et al., 2011). Мы отмечаем, что хотя PRC обычно рассчитываются с использованием короткого, сильного возбуждающего импульса тока в качестве возмущения, мы немного модифицируем эту парадигму здесь и предполагаем использовать отрицательный импульс, амплитуда и продолжительность которого мотивированы типом синаптических входов, генерируемых во время « тормозящего » bolus ‘в нашей сетевой модели (см. рис. 5).Мы знаем, что сотовая сеть PYR может генерировать всплески тета-популяции сама по себе, учитывая ее характеристики клеточной адаптации (функция SFA) (Ferguson et al., 2015b). Хотя сами по себе клетки PYR не срабатывают редко, как в эксперименте, они делают это при включении популяции PV + клеток (Ferguson et al., 2017). Мы считаем, что результирующая частота всплесков популяции сети E-I обусловлена комбинацией частоты срабатывания отдельной ячейки PYR и того, насколько тормозящий вход может опережать или задерживать всплеск ячейки PYR (как количественно оценивается с помощью PRC).Схема, которая учитывает это, схематически представлена на рис. Рис. 5 и состоит из следующего: Каждая ячейка PYR в гетерогенной популяции получает возбуждающий вход от других ячеек PYR, а также шумный привод (другой вход). Величина входного сигнала, который получает элемент PYR, конечно, будет колебаться со временем, но при разумном приближении элемент PYR получает средний возбуждающий вход примерно от 20 до 30 пА. Это приближение основано на том факте, что в наших моделях сети E-I (см. Рис. 1) всплески тета-популяции происходят, когда клетки PYR получают возбуждающий импульс с нулевым средним значением с колебаниями ≈ 10-30 пА (Ferguson et al., 2017). Затем мы вычисляем PRC, как описано выше. Тормозной импульс может опережать или задерживать последующий всплеск PYR-клетки, что определяется количественно PRC, что, в свою очередь, зависит от внутренних свойств PYR-клетки. Все эти аспекты схематизированы на Рис. 5.
Рис. 5. Схема установки для расчета кривой фазовой характеристики (PRC).Предполагая, что механизм генерации тета основан на дизайне модели, PRC генерируются на основе тормозящего входа (’болюса’), поступающего из сети ячеек PV + в ячейку PYR в сети ячеек PYR.Каждая ячейка PYR получает привод с шумом, обозначенный как «Другой вход», и для одной из ячеек PYR показана иллюстративная кривая f-I. Также показана иллюстрация вычисленного PRC, основанного на ингибирующем входе в конкретную ячейку PYR. Это будет зависеть от значений параметров модели конкретной ячейки PYR, которые определяют ее кривую f-I.
Мы рассматриваем три случая гетерогенных сетей E-I, демонстрирующих разные частоты всплесков совокупности, показанные на рис. Рис. 4 и описанные как имеющие «медленный», «средний» или «быстрый» выходной частотный всплеск.Мы генерируем PRC для нескольких моделей ячеек PYR в популяции для каждого из этих наборов моделей, которые производят выходы импульсов популяции с разной частотой. Каждая модель ячейки PYR в гетерогенной популяции имеет определенные характеристики PRC из-за заданных значений параметров модели и, таким образом, демонстрирует определенную внутреннюю частоту для данного входа.
4.1 Расчеты PRC
Они выполняются следующим образом: установленный входной ток (20: 2: 30 пА) тонически прикладывается к модельной ячейке, и период (определенный λ ) срабатывания ячейки рассчитывается как время между девятым и десятым спайком клеток.Обратный период представляет собой частоту срабатывания ячейки, выраженную в виде средних значений и стандартных отклонений для всех наборов моделей. Мы вычисляем фазовый отклик модельного нейрона на возмущение на 100 эквидистантных временах в его нормальном цикле активации, где возмущение представляет собой импульс тока длительностью 1 мс с амплитудой -500 пА (как упоминалось ранее, считается приближением синаптического входа, полученного эти клетки после «тормозящего болюса»). Для 1 ≤ i ≤ 100 мы определяем и доставляем возмущение на i * Δ p мс после 10-го всплеска ячейки.Затем мы измеряем время между 10-м и 11-м всплеском ячеек как «период возмущения» (определяемый λ p ). Мы вычисляем разницу между этим и ранее рассчитанным периодом (при отсутствии каких-либо возмущений) и нормализуем это на нормальный период срабатывания, что означает, что на графиках PRC ось y равна. Это означает, что отрицательные значения, нанесенные на график PRC, соответствуют фазовой задержке, то есть период возмущения был длиннее периода невозмущения, и наоборот.По оси абсцисс на графиках PRC отложено нормированное время, в которое возникло возмущение, вычисленное просто как. Мы отмечаем, что мы выполняем этот расчет отдельно для каждого i , т.е. мы повторно инициализируем ячейку и позволяем ей естественным образом реагировать на тонический ввод до 10-го всплеска для каждого значения i , вместо того, чтобы выполнять эти возмущения последовательно и рисковать смешивая ответы.
В Fig 6 B и C мы количественно оцениваем аспекты кривых PRC.В Рис. 6 B мы просто извлекаем значение нормализованной разности фаз из средней кривой PRC для возмущения, доставленного с нормализованной фазой 0,3 (обозначено стрелками, наложенными на Рис. 6, A ). В Рис. 6 C , мы количественно оцениваем один аспект скорости изменения средней кривой PRC, в частности, изменчивость коэффициента разности, вычисляемого на каждом этапе фазы, следующим простым способом: во-первых, этот коэффициент разности вычисляется для всех но последнее значение нормализованной фазы; во-вторых, дисперсия этих данных вычисляется просто с использованием функции var в MATLAB.
Рис. 6. На частоту тета-ритма влияет подавляющее влечение, что определяется количественно через PRC и частоты срабатывания отдельных клеток PYR.(A) Среднее значение PRC (сплошная линия) для гетерогенной популяции клеток PYR, участвующих в «медленных» (слева), «средних» (в центре) и «быстрых» (справа) тета-колебаниях, рассчитанное с входным током 20 пА, а затемнение представляет ± стандартное отклонение. Существует 25, 556 и 74 различных моделей клеток PYR в 10 000 популяциях клеток PYR для медленных, средних и быстрых случаев соответственно.Более подробная информация представлена в Приложении к дополнительным материалам. Среднее и стандартное отклонение частот срабатывания ячеек PYR на этом входном уровне включены во вставку каждой панели. (BC) После расчета как среднего PRC, так и средней собственной частоты возбуждения для популяций клеток PYR, связанных с нашими «медленными» (красный), «средними» (синий) и «быстрыми» (зеленый) тета-колебаниями для шести входных сигналов. токах (20: 2: 30 пА), мы выделяем особую особенность среднего PRC (средний фазовый сдвиг, вызванный возмущением, возникающим на фазе 0.3 на панели B и дисперсию средней производной PRC на панели C ) и нанесите ее на график в зависимости от средней собственной частоты возбуждения. Ни в том, ни в другом случае нет линейной зависимости между любой осью и частотой тета-ритма, что указывает на то, что это более сложная комбинация, которая определяет частоту популяции. Обратите внимание, что, учитывая монотонное соотношение между входным током и частотой зажигания в этом диапазоне, крайняя левая точка для каждого цвета представляет входной ток 20 пА, причем каждая последующая точка, перемещающаяся вправо, представляет следующий шаг входного тока.
Код для создания и построения этих PRC можно найти по адресу https://github.com/sbrich/Theta_PRCs. PRC для входных токов, отличных от 20 пА, которые показаны на Рис. 6 A можно найти на https://osf.io/yrkfv/).
4.2 Наблюдения
В Рис. 6 мы сначала показываем пример PRC, рассчитанный для входного тока 20 пА ( Рис. 6 A ). PRC рассчитываются для каждой модели в конкретном наборе моделей разнородных моделей ячеек PYR, при этом усредненная кривая представлена вместе с диапазоном ± одно стандартное отклонение (показано штриховкой вокруг кривой на каждом графике Рис. 6 A ).Эти PRC демонстрируют отличительные особенности: например, ячейки PYR в среднем случае однозначно демонстрируют область опережения фазы, в то время как ячейки PYR в быстром случае имеют наибольшую фазовую задержку для возмущений, доставленных на всех, кроме последних фаз. Четкие различия между PRC для каждого набора моделей сохраняются для всех используемых входных токов.
Чтобы лучше визуализировать влияние внутренних свойств нейронов PYR на частоту тета-ритма, мы наносим извлеченный признак среднего PRC против средней частоты срабатывания этих наборов моделей для каждого из наших вычисленных входных токов в Рис. B и C , с соответствующими частотами тета-ритма, связанными с каждым модельным набором, обозначенным цветом точки данных, с извлеченными характеристиками PRC в каждом случае, описанном в предыдущем разделе.Эти визуализации ясно показывают, что и PRC, и средняя собственная частота возбуждения нейронов PYR в данном наборе моделей вносят вклад в общую частоту тета-ритма; в противном случае эти точки были бы «плоскими» относительно оси x или y. Кроме того, взаимосвязь между извлеченной интересующей характеристикой PRC и средней внутренней частотой возбуждения заметно варьируется в зависимости от выходной частоты тета-ритма: например, в Рис. 6 B и «медленный», и «средний» наборы моделей показывают монотонно убывающая взаимосвязь между извлеченным значением PRC и средней собственной частотой срабатывания, в то время как набор «быстрых» моделей показывает монотонно возрастающую взаимосвязь.Взятые вместе, эти результаты показывают, что именно комбинация тормозящего влечения и возбудимости PYR-клетки вносит вклад в общую частоту тета-ритма.
Внутренние свойства, определяемые количественно с помощью PRC, помогают сформулировать потенциальные механизмы, с помощью которых возникают эти различные частоты тета-ритма. Например, в то время как клетки PYR в быстром случае имеют самую быструю индивидуальную частоту срабатывания (заметно быстрее, чем то, что видно в популяционных моделях), их PRC могут быть иллюстрацией того, как ингибирующий «болюс» снижает эту частоту срабатывания в направлении тета-диапазона.Между тем, клетки PYR в среднем случае имеют самые медленные индивидуальные частоты срабатывания, хотя они участвуют в «средних» частотах тета-ритма. PRC в этом случае, особенно область смещения фазы, может прояснить, как ингибирующий синаптический вход на самом деле ускоряет активность PYR-клеток. В этих конкретных примерах используется функция PRC, извлеченная и нанесенная на график Рис. 6, B .
Этот анализ характеристик PRC наших наборов моделей поддерживает нашу гипотезу о том, что частота всплесков популяции сети обусловлена комбинацией входных данных, которые получают ячейки PYR, и внутренних свойств этих ячеек, определяющих их ответы на указанные входные данные.Реакция клеток на возбуждающее побуждение частично оценивается средней внутренней частотой срабатывания наборов моделей, в то время как их реакция на тормозное побуждение количественно оценивается свойствами вычисленных PRC. Тем не менее, это все в контексте возможности иметь стабильный всплеск популяции в первую очередь, как это предусмотрено нашим дизайном модели с функциями SFA, PIR и Rheo: наши модели включают популяцию клеток PYR, которая может генерировать всплески популяции тета-частоты. сама по себе, при этом популяция клеток PV + служит для облегчения активации разреженных клеток PYR.Расчеты PRC здесь показывают, что соответствующий тормозящий вход вносит вклад в результирующую частоту всплеска популяции.
5 ОБСУЖДЕНИЕ
Было разработано несколько моделей тета-ритмов (Ferguson and Skinner, 2018; Kopell et al., 2010), но они специально не рассматривали контроль частоты тета в сочетании с его генерацией в экспериментальном контексте. Здесь мы использовали модель микросхемы, разработанную для генерации тета-ритмов, представляющих те, которые наблюдаются во всем препарате гиппокампа, чтобы разработать гипотезу для контроля тета-частоты.Наша работа позволила нам предложить гипотезу для контроля ритма тета-частоты, которая охватывает два аспекта: (i) внутренний механизм, который проистекает из свойств строительных блоков SFA, PIR и Rheo клеток PYR; (ii) внешний механизм, который включает «основанную на ингибировании настройку» активации клеток PYR. Из нашей предыдущей работы мы уже знали, что минимально связанные сети ячеек PYR сами по себе производили тета-частотные всплески популяции (Ferguson et al., 2015b), но большинство PYR-клеток срабатывали во время популяционных тета-всплесков, что в отличие от экспериментальных наблюдений за тета-всплесками. разреженное срабатывание клеток PYR.С включением клеток PV + для создания сетей E-I популяция клеток PYR активировалась редко в соответствии с экспериментом. Имеет смысл, что добавление ингибирующих клеток приводит к меньшему возбуждению клеток PYR из-за потенциального молчания из-за ингибирования. То, что сильные тета-ритмы все еще могут возникать, несмотря на участие в ритме меньшего количества клеток PYR, вероятно, связано с тем, что клетки PV + настраивают различные в остальном частоты клеток PYR на аналогичные частоты, позволяя этой меньшей группе клеток производить сильные ритмы.Это составляет основную часть предлагаемой нами гипотезы. Соответственно, было показано, что ингибирование с прямой связью играет роль в поддержании низких уровней коррелированной изменчивости пиковой активности (Middleton et al., 2012).
Важно выделить два ключевых аспекта, лежащих в основе предлагаемой нами гипотезы. Во-первых, популяция клеток PYR должна быть достаточно большой, чтобы она могла коллективно генерировать сильный возбуждающий импульс к ингибирующим PV + клеткам, и, в свою очередь, популяция PV + клеток должна быть способна активировать достаточно (и последовательно), чтобы создать сильное тормозящее действие. bolus ‘, чтобы настроить выход популяции клеток PYR.Во-вторых, чистый ввод (повторяющееся возбуждение, возбуждающее побуждение, входящее торможение), полученный клетками PYR, приводит к генерации тета-ритмов и их результирующей частоты. Интересно отметить сходство между этими ключевыми аспектами и «механизмом PING», лежащим в основе генерации гамма-ритмов в сетях EI (Kopell et al., 2010; ter Wal and Tiesinga, 2013), особенно с учетом недавних исследований, показывающих, что ритмы с частотами, приближающимися к тета-диапазону, могут возникать в сетях, мотивированных PING (Rich et al., 2017).
Мы не знаем, существует ли на самом деле четкая взаимосвязь между входами ячеек PYR и частотой сети, как описано во втором ключевом аспекте выше, и было бы очень сложно напрямую проверить это экспериментально. Тем не менее, можно использовать подробные биофизические сетевые модели для изучения этого и получения биологической информации. Мы сделали это, объединив описанную модель микросхемы, используемую здесь, и подробную полномасштабную модель микросхемы CA1 (Bezaire et al., 2016) и изучение того, как частота тета-сети, создаваемая подробной моделью, зависит от чистого ввода, полученного ячейками PYR (Chatzikalymniou et al., 2020). Мы обнаружили, что детализированные с биологической точки зрения модели убедительно подтверждают эту зависимость и, таким образом, предложенную нами гипотезу для контроля частоты тета-ритма. Таким образом, это указывает на то, что тета-частоты в биологической системе можно контролировать таким образом.
В предыдущей работе Ferguson et al. (2015a), мы создали модели клеток PYR, которые либо сильно адаптируются на основе соответствия экспериментальным данным, либо слабо адаптируются на основе другого экспериментального набора данных.В Ferguson et al. (2015b), когда любая из моделей ячеек PYR использовалась в сетях только с E-ячейками, это могло производить всплески популяции тета-частоты. Как обсуждалось в Ferguson et al. (2015a), маловероятно, что существуют отдельные типы биологических клеток PYR, которые сильно или слабо адаптируются, а скорее континуум степени адаптации, зависящей от лежащих в основе балансов токов биофизических ионных каналов. Наши исследования устойчивости механизма генерации тета в модели микросхемы здесь показали, что частота и мощность тета-ритмов не сильно контролировались значениями характеристик SFA по сравнению со значениями характеристик Rheo.Таким образом, хотя мы создали базу данных моделей, исходя из сильно адаптирующейся базы параметров модели клеток PYR, это, вероятно, не имело бы значения, если бы вместо этого проводилось исследование устойчивости генерации тета-ритма с использованием слабо адаптируемых моделей клеток PYR.
Возможно, неудивительно, что значения характеристик Rheo являются основным контроллером существования тета-ритмов, их частоты и мощности, поскольку конкретное значение Rheo определяет, будет ли PYR-ячейка всплывать или нет.Отметим, что экспериментальные данные Goutagny et al. (2009) уже предположили важность PIR в генерации тета-ритмов. В реальных клетках CA1 PYR было показано, что пик PIR действительно происходит, опосредуется h-каналами и локально контролируется балансом биофизических ионных каналов (Ascoli et al., 2010). То, действительно ли клетки PYR срабатывают из-за PIR во время продолжающихся тета-ритмов, может иметь или не иметь место, и это потенциально можно распутать в моделях. Тем не менее, гипотеза, развитая в этой работе, указывает на слияние функций, которые приводят к тому, что чистый ток к отдельным клеткам PYR является центром контроля частоты тета-ритма.Таким образом, можно ожидать, что изменения в чистом побуждении к клеткам PYR или изменения внутренних свойств клеток PYR, таких как h-токи, которые будут влиять на PIR, повлияют на результирующую частоту тета-ритма.
Теория PRC использовалась множеством способов в области нейробиологии (Schultheiss et al., 2011), в частности, при рассмотрении сетевой динамики. Например, Hansel et al. (1995) использовали PRC для объяснения дифференциальной способности возбуждающего драйва для синхронизации сетей нейронов типа I или типа II (эти типы различаются по типу бифуркации (Izhikevich, 2006)), Rich et al.(2016) проанализировали особенности синхронизации в чисто тормозящих сетях с использованием PRC, а Achuthan и Canavier (2009) использовали PRC для понимания кластеризации в сетях. Мы воспользовались теорией PRC, рассмотрев сброс фазы ячеек PYR из-за входящего тормозящего входа. Таким образом, мы смогли выдвинуть гипотезу о механизме настройки, основанном на ингибировании, для управления частотой тета-ритма на основе формы PRC (величины опережения или задержки) и собственной частоты срабатывания клетки PYR. Наше использование PRC основывалось на наших наблюдениях за влиянием различных форм PRC на результирующий тета-ритм.Например, такое рассмотрение использовали Rich et al. (2016) для объяснения паттернов дифференциальной синхронизации в тормозных сетях нейронов типа 1 и типа II.
В заключение, мы разработали гипотезу о том, как частоты тета-ритма контролируются в гиппокампе CA1. Эта гипотеза основана на тета-генерирующем механизме конструкции модели микросхемы. Несмотря на то, что он не включает все известные типы тормозных клеток, он, возможно, улавливает важные элементы, задействованные в биологических цепях, и может более широко применяться в мозге в отношении генерации и контроля частот тета-ритма.
ЗАЯВЛЕНИЕ О КОНФЛИКТЕ ИНТЕРЕСОВ
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
ВКЛАД АВТОРА
FS, JL, AC внесли свой вклад в разработку концепции и руководство исследованием. SR, AL выполнили расчеты и анализ. Ф.С. написал первый черновик рукописи. SR написал разделы рукописи. Все авторы внесли свой вклад в доработку рукописи, прочитали и одобрили представленную версию.
ФИНАНСИРОВАНИЕ
Эта работа была поддержана Канадским советом по естественным наукам и инженерным исследованиям (NSERC), грант на открытие RGPIN-2016-06182 (FKS).
ДОПОЛНИТЕЛЬНЫЕ ДАННЫЕ
Существует дополнительный файл, который в основном тексте называется Приложением.
ЗАЯВЛЕНИЕ О ДОСТУПНОСТИ ДАННЫХ
Дополнительные наборы данных, созданные в ходе этого исследования, которых нет здесь или в Приложении (Дополнительные материалы), можно найти по адресу https://osf.io/yrkfv/.
ССЫЛКИ
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Chatzikalymniou, A.П., Гумус М., Лунёв А. Р., Рич С., Лефевр Дж. И Скиннер Ф. К. (2020). Связывание минимальных и подробных моделей микросхем CA1 показывает, как возникают тета-ритмы и как контролируются их частоты. bioRxiv, 2020.07.28.225557 doi: 10.1101 / 2020.07.28.225557. Издатель: Лаборатория Колд-Спринг-Харбор. Раздел: новые результаты. ↵
Ижикевич, Э.М. (2006). Динамические системы в нейробиологии: геометрия возбудимости и разрыва (MIT Press), 1-е изд.
- ↵
- ↵
- ↵
Кота, С., Рагг, М. Д., и Лега, Б. К. (2020). Тета-колебания гиппокампа поддерживают успешное формирование ассоциативной памяти. Журнал неврологии 40, 9507–9518. DOI: 10.1523 / JNEUROSCI.0767-20.2020. Издатель: Общество нейробиологии Раздел: Исследовательские статьи
- ↵
- ↵
- ↵
Миддлтон, Дж.В., Омар, К., Дуарон, Б., и Саймонс, Д. Дж. (2012). Нейронная корреляция — это стимул, модулируемый блокирующими цепями прямой связи. Журнал неврологии 32, 506–518. DOI: 10.1523 / JNEUROSCI.3474-11.2012. Издатель: Общество неврологии Раздел: Статьи
- ↵
- ↵
- ↵
- ↵
Schultheiss, N., Butera, R., and Prinz, A. (2011). Кривые фазового отклика в нейробиологии: теория, эксперимент и анализ (Springer)
- ↵
- ↵
CMX902QT8 CML Microcircuits (USA) Inc.Усилитель | RFMW
НАЖИМАЯ КНОПКУ «ПРИНЯТЬ», «ВЫ» (ОТНОСИТЕЛЬНО ВАС ЛИЧНО ИЛИ КОМПАНИЯ, которую вы представляете, И ОТ ЧЬЮ ВЫ ПОЛНОСТЬЮ УПОЛНОМОЧЕННЫ ПОДАТЬ ЗАЯВЛЕНИЕ НАСТОЯЩЕГО СОГЛАШЕНИЯ) СОГЛАШАЕТЕСЬ С НАСТОЯЩИМ СОГЛАШЕНИЕМ С ДАННОЙ ЛИЦЕНЗИЕЙ И ЯВЛЯЕТСЯ СОГЛАШЕНИЕМ «СОГЛАШЕНИЕ»). ЕСЛИ ВЫ НЕ СОГЛАСНЫ СО ВСЕМИ УСЛОВИЯМИ НАСТОЯЩЕГО СОГЛАШЕНИЯ, НАЖМИТЕ КНОПКУ «ОТМЕНА», И ПРОЦЕСС ЗАГРУЗКИ / УСТАНОВКИ НЕ ПРОДОЛЖИТСЯ. ЕСЛИ ЭТИ УСЛОВИЯ ЯВЛЯЮТСЯ ПРЕДЛОЖЕНИЕМ, ПРИНЯТИЕ ЯВНО ОГРАНИЧИВАЕТСЯ ЭТИМИ УСЛОВИЯМИ.1. ГРАНТ. В соответствии с условиями настоящего Соглашения («Компания») настоящим предоставляет вам (и только вам) ограниченную личную, не подлежащую сублицензированию, непередаваемую, бесплатную, неисключительную лицензию на использование программного обеспечения, которое вы собираетесь использовать для внутренних целей. загружать / устанавливать («Программное обеспечение») только в соответствии с настоящим Соглашением и документацией Компании, прилагаемой к Программному обеспечению, и без каких-либо модификаций, кроме модификаций, предоставленных непосредственно Компанией. 2. ОГРАНИЧЕНИЯ. Вы не можете (и соглашаетесь не делать, не разрешать или не позволять другим) прямо или косвенно: (а) копировать, распространять или иным образом использовать Программное обеспечение в интересах третьей стороны; (б) дизассемблировать или иным образом реконструировать Программное обеспечение; или (c) удалить из Программного обеспечения любые уведомления о правах собственности.Вы понимаете, что Компания может изменить или прекратить предложение Программного обеспечения в любое время. 3. ПОДДЕРЖКА И ОБНОВЛЕНИЯ. Настоящее Соглашение не дает вам права на какую-либо поддержку, обновления, исправления, улучшения или исправления для Программного обеспечения (совместно именуемые «Поддержка»). Любая такая Поддержка Программного обеспечения, которая может быть предоставлена Компанией, становится частью Программного обеспечения и регулируется настоящим Соглашением. 4. ОТКАЗ ОТ ГАРАНТИЙ. КОМПАНИЯ ПРЕДОСТАВЛЯЕТ ПРОГРАММНОЕ ОБЕСПЕЧЕНИЕ «КАК ЕСТЬ» И БЕЗ КАКИХ-ЛИБО ГАРАНТИЙ, И НАСТОЯЩИМ ОТКАЗЫВАЕТСЯ ОТ ВСЕХ ЯВНЫХ ИЛИ ПОДРАЗУМЕВАЕМЫХ ГАРАНТИЙ, ВКЛЮЧАЯ БЕЗ ОГРАНИЧЕНИЙ ГАРАНТИИ ТОВАРНОСТИ, ПРИГОДНОСТИ, СООТВЕТСТВИЯ ИДЕНТИФИКАЦИОННЫМ ХАРАКТЕРИСТИКАМ, СООТВЕТСТВУЮЩИМ ХАРАКТЕРИСТИКАМ.НАСТОЯЩИЙ ОТКАЗ ОТ ГАРАНТИЙ ЯВЛЯЕТСЯ НЕОБХОДИМОЙ ЧАСТЬЮ НАСТОЯЩЕГО СОГЛАШЕНИЯ. 5. ОГРАНИЧЕНИЕ ОТВЕТСТВЕННОСТИ. НИ ПРИ КАКИХ ОБСТОЯТЕЛЬСТВАХ И НИ ПРИ КАКИХ ОБСТОЯТЕЛЬСТВАХ, ВКЛЮЧАЯ, БЕЗ ОГРАНИЧЕНИЙ, ПРАВО, ДОГОВОР, СТРОГОУЮ ОТВЕТСТВЕННОСТЬ ИЛИ ИНЫМ ОБРАЗОМ, КОМПАНИЯ ИЛИ ЕЕ ЛИЦЕНЗИАРЫ, ПОСТАВЩИКИ ИЛИ ТОРГОВЫЕ ПРОДАВЦЫ НЕСЕТ ОТВЕТСТВЕННОСТЬ ПЕРЕД ВАМИ ИЛИ ЛЮБЫМ ЛИЦОМ, ЗА (A) СЛУЧАЙНЫЕ ИЛИ КОСВЕННЫЕ УБЫТКИ ЛЮБОГО ХАРАКТЕРА, ВКЛЮЧАЯ, БЕЗ ОГРАНИЧЕНИЙ, УБЫТКИ, УБЫТАННЫЕ ПРИБЫЛИ, ПОТЕРЮ ДОЛЖНОСТИ, ОСТАНОВКА РАБОТЫ, ТОЧНОСТЬ РЕЗУЛЬТАТОВ, СБОЙ ИЛИ НЕИСПРАВНОСТЬ КОМПЬЮТЕРА, ПОВРЕЖДЕНИЯ ИЛИ БЕСПЛАТНОГО ИСПОЛЬЗОВАНИЯ ПО 100 долларов США.6. ПРЕКРАЩЕНИЕ. Вы можете прекратить действие настоящего Соглашения и предоставленной в нем лицензии в любое время, уничтожив или удалив со всех компьютеров, сетей и носителей информации все копии Программного обеспечения. Компания может немедленно прекратить действие настоящего Соглашения и предоставленной в нем лицензии, если вы нарушите какое-либо положение настоящего Соглашения. Получив уведомление о прекращении действия от Компании, вы уничтожите или удалите со всех компьютеров, сетей и носителей все копии Программного обеспечения. Разделы 2–7 остаются в силе после прекращения действия настоящего Соглашения.7. РАЗНОЕ. Вы обязуетесь соблюдать все применимые экспортные законы, ограничения и постановления в связи с использованием вами Программного обеспечения и не будете экспортировать или реэкспортировать Программное обеспечение в нарушение этих правил. Настоящее Соглашение является личным для вас, и вы не имеете права переуступать или передавать Соглашение или Программное обеспечение третьим лицам ни при каких обстоятельствах; Компания может переуступать или передавать настоящее Соглашение без согласия. Настоящее Соглашение представляет собой полное соглашение относительно данной лицензии между сторонами и заменяет собой все предыдущие соглашения и заявления между ними.В него могут быть внесены изменения только в письменной форме, оформленной обеими сторонами. Если какое-либо положение настоящего Соглашения будет признано не имеющим исковой силы по какой-либо причине, такое положение должно быть изменено только в той мере, в какой это необходимо для обеспечения его исполнения. Настоящее Соглашение регулируется и толкуется в Калифорнии без учета каких-либо положений коллизионного права.
Дендритная возбудимость и контроль усиления в рекуррентных кортикальных микросхемах | Кора головного мозга
Аннотация
Слой 5 толстых пирамидных клеток (TTC) в неокортексе особенно электрически сложен из-за их очень возбудимых дендритов.Взаимодействие между дендритной нелинейностью и повторяющейся активностью кортикальной микросхемы в формировании сетевой реакции в значительной степени неизвестно. Мы смоделировали подробные основанные на проводимости модели TTC, образующие повторяющиеся микросхемы, которые были связаны между собой экспериментально; сеть была встроена в реалистичную фоновую синаптическую активность. Микросхемы ТТС значительно усиливали кратковременные таламокортикальные входы; это усиление коры головного мозга было опосредовано активированной обратным распространением деполяризации N -метил-d-аспартата и активированным обратным распространением дендритов спайком Ca 2+ , вызванным совпадением активируемого таламом соматического спайка и локальных дендритных синаптических входов , происходящие из корковой микросхемы.Удивительно, но дендритная нелинейность в микросхемах TTC линейно умножала таламические входы — усиливая их, сохраняя при этом входную селективность. Наши результаты показывают, что дендритные нелинейности имеют решающее значение в управлении усилением и вычислительными функциями микросхем TTCs, которые служат в качестве основного выходного источника для неокортекса.
Введение
На реакцию нейронов в неповрежденном мозге динамически влияет как активность локальной рекуррентной цепи, так и нелинейные дендритные свойства (Ben-Yishai et al.1995; Лю и др. 2007; Такахаши и Маги 2009; Buesing et al. 2011; Larkum 2012; Xu et al. 2012; Ли, Ли и др. 2013; Ли, Ибрагим и др. 2013; Reimann et al. 2013; Вайдья и Джонстон 2013). Это особенно верно для важного класса корковых нейронов, слоя 5 (L5) толстых пирамидных клеток (TTC), поскольку их дендритное дерево очень возбудимо (см. Ниже). Тем не менее, взаимодействие между сетевой активностью из-за повторяющихся соединений и дендритной нелинейности, а также их комбинированное влияние на динамику сети, изучено недостаточно.
Предыдущие исследования модуляции ответа на сенсорный стимул из-за повторяющейся сетевой активности рассматривали сеть клеток L4, которые считались основными мишенями таламокортикальных аксонов (Gilbert and Wiesel 1979; Douglas and Martin 2004). Однако в последние годы эксперименты in vivo продемонстрировали существенное прямое воздействие таламуса на перисоматическую область TTC, что привело к появлению постсинаптических потенциалов (PSP) и спайковых латентностей, которые были такими же ранними, как и зарегистрированные в клетках L4 (Heimel et al.2005; de Kock et al. 2007; Meyer et al. 2010; Константинополь и Бруно 2013). Таким образом, исследование реакции сети TTC на прямое таламическое воздействие очень актуально и в настоящее время отсутствует. Несколько недавних экспериментальных исследований возбуждающих клеток в L4 in vivo показывают, что во время слуховых или зрительных ощущений рекуррентная корковая сеть усиливает ответ этих клеток на таламический вход без изменения их селективности стимулов (Liu et al.2007; Li, Li et al. 2013; Ли, Ибрагим и др.2013; Lien and Scanziani 2013). Предыдущие теоретические исследования показали, что повторяющаяся активность среди клеток, которые аналогичным образом настроены на свойство стимула, такое как зрительная ориентация, может увеличить усиление ответа клеток, тем самым усиливая ответ на вход таламуса, а также повышая избирательность ответа или усиливая сигнал. отношение шума к шуму (Ben-Yishai et al. 1995; Douglas et al. 1995; Suarez et al. 1995; Chance et al. 1999). Как в экспериментальных, так и в теоретических исследованиях, упомянутых выше, вклад дендритной возбудимости в модуляцию отклика не учитывался.
Основные дендритные нелинейности в TTC связаны с шипами Ca 2+ на главном бифуркации апикальных дендритов (Helmchen et al. 1999; Larkum and Zhu 2002; Xu et al. 2012) и N -метил-d- шипы аспартата (NMDA) в тонких ветвях апикальных и базальных дендритных деревьев (Schiller et al. 2000; Rhodes 2006; Larkum et al. 2009; Polsky et al. 2009). Предыдущие исследования показали, что спайки NMDA могут усиливать реакцию клетки, а также повышать ее избирательность (Lavzin et al.2012; Smith et al. 2013; Palmer et al. 2014). Проводимость NMDA, в целом, может действовать более локально, чтобы снизить порог генерации спайков Ca 2+ (Schiller and Schiller 2001; Polsky et al. 2004; Larkum et al. 2009). Спайки Ca 2+ длиннее и имеют большую амплитуду, чем спайки NMDA. Недавние экспериментальные исследования in vivo показали, что растормаживание апикальных дендритов в TTC резко увеличивает усиление их ответа на соматосенсорную стимуляцию в корреляции с дендритным сигналом Ca 2+ (Murayama et al.2009; Palmer et al. 2012). Эти находки дополняют ранее полученные in vitro доказательства влияния дендритных спайков Ca 2+ на усиление частота-ток (Larkum et al. 2004). Влияние дендритных шипов Ca 2+ на выходное (аксональное) возбуждение клетки особенно очевидно при совпадающих входах в сому и дистальный апикальный дендрит, которые соответствуют, соответственно, таламическому (снизу вверх) входу и кортикальному ( сверху вниз или локально), которые эти клетки получают в неповрежденном мозге (Felleman and Van Essen 1991; Constantinople and Bruno 2013).Совпадение этих двух входных источников запускает активированный обратным распространением спайк Ca 2+ (BAC) in vitro (Larkum et al. 1999), который, как предполагалось, используется для нейронных вычислений, таких как корковые ассоциации (Larkum 2012). .
Участие дендритных спайков Ca 2+ при совпадении разных потоков входов было экспериментально продемонстрировано и в других областях мозга. В гиппокампе исследования in vitro показали, что совпадающие стимулы, поступающие в разные области дендритов, запускают потенциал плато Ca 2+ , продолжительность которого коррелирует с синаптической потенциацией (Takahashi and Magee 2009).Во время активного взбивания совпадение моторных и сенсорных входных потоков в пирамидных клетках L5 запускает дендритные сигналы Ca 2+ , которые коррелируют с потенциалами дендритных плато (Xu et al. 2012).
Несколько недавних исследований показали, что корковые сети можно рассматривать как состоящие из относительно небольших функциональных микросхем, от десятков до сотен клеток (Perin et al. 2011). Связность в коре головного мозга не случайна, а скорее связана с более высоким числом взаимосвязанных микросхем (или мотивов), чем ожидалось случайно (Song et al.2005; Perin et al. 2011), а также более высокая вероятность соединения между клетками, которые имеют общие входы (Yoshimura et al. 2005; Kampa et al. 2006) или имеют сходные предпочтения стимулов (Ko et al. 2011). Более того, исследования показали, что корковые связи также зависят от типа клеток (Yoshimura and Callaway 2005; Anderson et al. 2010). Эти исследования показали существование отдельных микросхем, которые могут действовать как функциональные модули. Понимание того, как работают такие локальные корковые модули, способствует пониманию вычислительных возможностей и функций более крупных сетей, состоящих из таких модулей (Traub et al.2005; Дуглас и Мартин 2007; Heinzle et al. 2007, 2010; Буономано и Маасс 2009; Литвак и Ульман 2009; Папуци и др. 2013).
Чтобы исследовать совокупный вклад как рекуррентной активности корковой сети, так и дендритной возбудимости в модуляцию таламокортикальных входов, мы смоделировали корковые микросхемы, состоящие из детальных TTC, используя новую детальную модель для TTC, которую мы недавно разработали (Hay и др. 2011). Эти локальные корковые сети были погружены в контекст фоновой активности, подобной in vivo.Мы показываем, что модуляция ответа в микросхемах TTC за счет повторяющейся активности после соматического («снизу вверх») ввода в значительной степени зависит от повышения потенциала обратного распространения (BAP) дендритных возбуждающих постсинаптических потенциалов (EPSP), запуска токов NMDA и активации BAC. . А именно, дендритные свойства TTC играют важную роль в формировании динамического ответа локальной возбуждающей сети. Используя настройку ориентации в качестве примера, мы проиллюстрировали возможное функциональное значение этих двух механизмов для модуляции реакции возбуждающих корковых сетей.
Материалы и методы
Модель с одной ячейкой
Каждый TTC в микросхеме был представлен с использованием детальной модели на основе проводимости, которую мы недавно разработали (Hay et al. 2011). Эта модель продемонстрировала соматический спайк Na + , дендритный спайк Ca 2+ , БАТ из-за дендритных каналов Na + и всплеск соматических спайков Na + в ответ на совпадающую деполяризацию в соме и апикальной части. пучок (обжиг BAC, см. Larkum et al.1999). Смоделированная микросхема в настоящем исследовании состояла из N = 1–150 смоделированных ячеек, все они идентичны по своей морфологии и биофизическим свойствам.
Фоновый синаптический вход
Чтобы смоделировать схему в контексте активности in vivo, мы случайным образом распределили 10 000 возбуждающих синапсов и 2500 тормозных синапсов на каждой модели TTC, как было предложено предыдущими тщательными оценками в нейронах кошек (Binzegger et al. 2004).Отметим, что возможное различие в оценках нейронов крысы из-за разницы в размере клеток не было решающим для нашего исследования, так как оно могло быть компенсировано изменением соотношения тормозной и возбуждающей проводимости. Каждый возбуждающий синапс включает комбинацию NMDA- и AMPA-зависимой проводимости, а также единую динамику краткосрочной пластичности (Ramaswamy et al. 2012) с равной максимальной проводимостью (Sarid et al. 2007). Постоянная времени нарастания проводимости NMDA составляла 2 мс, а постоянная времени спада составляла 65 мс (Rhodes 2006).Постоянная времени нарастания проводимости AMPA составляла 0,3 мс, а постоянная времени спада составляла 3 мс (Hestrin 1992). Тормозящие синапсы были типа GABA A . Постоянная времени нарастания проводимости GABA A составляла 1 мс, а постоянная времени спада составляла 20 мс (Salin and Prince 1996; Gidon and Segev 2012). Реверсивные потенциалы возбуждающих и тормозных синапсов составляли 0 и -80 мВ соответственно (Destexhe et al. 1994), а потенциал покоя смоделированных клеток составлял -80 мВ.И тормозные, и возбуждающие синапсы обладают вероятностным высвобождением с кратковременной пластичностью (Ramaswamy et al. 2012). Механизм, учитывающий комбинированные токи AMPA и NMDA, имел кратковременную депрессию с вероятностью высвобождения ( U ) 0,6, постоянной времени выхода из депрессии ( D ) 800 мс и постоянной времени восстановления после облегчения. ( F ) 0 мс (Цодыкс и Маркрам, 1997; Фурманн и др., 2002). Тормозящие синапсы тоже угнетали, U = 0.25, D = 800 мс и F = 0 мс (Гупта и др., 2000).
Мы установили максимальную тормозную проводимость синапсов равной 1 нСм, что дает среднюю амплитуду соматического ингибирующего постсинаптического потенциала (IPSP) -0,6 мВ при удерживающем мембранном потенциале -70 мВ для тормозного соединения, состоящего из 12 синаптических контактов (Silberberg and Markram 2007 ). Максимальная проводимость для NMDA- и AMPA-рецепторов составила 0,4 нСм, что дает среднюю амплитуду ВПСП 0,6 мВ при потенциале покоя (Song et al.2005; Томсон и Лами 2007; Perin et al. 2011) для возбуждающего соединения (одиночный аксон TTC), создающего 5 синаптических контактов на единственном целевом дендрите TTC (Markram et al. 1997). Фоновые пресинаптические последовательности потенциалов действия (AP) следовали распределению Пуассона со средней частотой возбуждения 0,72 Гц для возбуждающих синапсов (Heimel et al. 2005) и 7 Гц для тормозных синапсов (Gentet et al. 2010). Эти настройки дали частоту самопроизвольного срабатывания 3,1 ± 0,5 Гц в неподключенных TTC (размер микросхемы N = 1) и 4.3 ± 1,5 Гц в ТТС, составляющих микросхемы размером N = 150, оба порядка величины, обнаруженной in vivo (Heimel et al. 2005; Olsen et al. 2012). Кроме того, эта настройка дала нормальное распределение подпорогового мембранного потенциала со средним значением -63 мВ и стандартным отклонением 3 мВ, что согласуется с экспериментальными измерениями на бодрствующих животных (Constantinople and Bruno 2011).
Подключение микросхем
Между собой было подключеноТТС с вероятностью P = 0.13 для однонаправленных соединений и вероятность P rcp = 0,06 для взаимных соединений (Song et al. 2005). Реципрокные связи были в 1,5 раза сильнее (большая синаптическая проводимость), чем однонаправленные (Song et al. 2005). Задержка между началом пресинаптического AP в соме и началом постсинаптического EPSP составляла 1 мс (Markram et al. 1997). Каждое соединение состояло из 5 синаптических контактов типа NMDA / AMPA (смоделированных, как для соединений возбуждающего фона, описанных выше), за исключением того, что эти синапсы были вспомогательными, с U = 0.25, D = 0 мс, F = 0 мс (Рейес и Сакманн 1999; Фрик и др. 2007; Уильямс и Аткинсон 2007). Хотя исследования показали, что синаптические местоположения между TTCs имеют тенденцию контактировать с базальными и наклонными дендритами, они могут встречаться по всему дендритному дереву постсинаптической клетки (Markram et al. 1997). Таким образом, мы случайным образом распределили различные синаптические контакты из пресинаптических ячеек микросхемы на каждой постсинаптической ячейке. При сборе статистики срабатывания микросхем типоразмера N мы моделировали 4 разные случайные микросхемы для каждого случая.Микросхемы различались связностью между TTC, синаптическим расположением соединений между TTC и пресинаптическими цепочками фонового входа, воздействующего на каждую TTC. Микросхемы размером больше N = 150 ячеек не анализировали, так как в условиях настоящего исследования спонтанная частота в таких микросхемах существенно отклонялась от экспериментально измеренной спонтанной скорости.
Гистограмма времени после стимула
При создании временной гистограммы после стимула (PSTH) мы разбили смоделированные кривые напряжения всех ячеек в сети на интервалы по 5 мс.Каждый бункер получил значение 1, если он содержал начало всплеска, или значение 0 в противном случае.
Избирательность визуальной ориентации
Мы имитировали стимулы с разной зрительной ориентацией, используя разные амплитуды короткого (5 мс) деполяризующего импульса, вводимого в сому всех смоделированных клеток, от 0 нА (что соответствует отсутствию восходящего таламического входа или «нулевой ориентации»). до 1,4 нА (наша амплитуда по умолчанию в этом исследовании, которая вызвала ровно 1 дополнительный всплеск во всех ячейках микросхемы и соответствовала «предпочтительной ориентации»).Частота реакции спайков в нулевой и предпочтительной ориентациях была порядка величины, наблюдаемой экспериментально (Van Hooser 2007). Промежуточные амплитуды импульсов составляли 0,2, 0,4 и 0,6 нА, что давало скорости отклика в цепи, которые охватывали диапазон между откликами на нулевую и предпочтительную ориентации. Затем мы вывели соответствующие углы ориентации стимула, используя гауссову кривую отклика, которая имела полуширину на полувысоте 29 °, в соответствии с экспериментальной статистикой (Van Hooser 2007).Центр кривой отклика соответствовал предпочтительной ориентации, которая была произвольно назначена равной 90 °. Для количественной оценки селективности ориентации мы использовали 2 показателя: полуширину на полувысоте и индекс селективности, который представляет собой отношение между второй и первой составляющими Фурье кривой ориентации (Ringach et al. 2002). Мы отмечаем, что выбор имитации стимулов с короткими токами, а не с подробным синаптическим вводом, служил для (i) надежного запуска соматического всплеска и (ii) сохранения анализа на фундаментальном уровне тока ответа ( f – I ), тем самым позволяя нашим выводам быть общими и не привязанными к конкретным предположениям относительно синаптического входа.
Все моделирование проводилось с использованием NEURON (Carnevale and Hines, 2006), работающего в сетке из 60 Sun X4100 AMD 64-битных двухъядерных процессоров Opteron (всего 240 ядер), под управлением Linux 2.6. Среднее время выполнения моделирования каждой микросхемы (из N нейронов с использованием N ядер) составило 30 мин. Код моделирования доступен на ModelDB (Hines et al. 2004).
Результаты
Чтобы изучить динамику сети в реалистичной повторяющейся сети TTC, мы подключили микросхемы, состоящие из 1–150 детальных моделей TTC (см. Материалы и методы).На рис. 1 A показан пример микросхемы из 50 ячеек с вероятностями однонаправленного (черные линии) и обратного (красные линии) соединения, как обнаружено экспериментально (см. Материалы и методы). В дополнение к соединениям от микросхемы, каждая ячейка получала случайные пуассоновские фоновые тормозные и возбуждающие синаптические входы, которые заставляли ее спонтанно срабатывать с приемлемым диапазоном частот для всех размеров микросхем, которые мы исследовали (рис. 1 B ). Несоединенные элементы (размер микросхемы N = 1) спонтанно сработали на 3.1 ± 0,5 Гц, а клетки в микросхемах размером N = 150 спонтанно запускались с частотой 4,3 ± 1,5 Гц, как в экспериментальном диапазоне, так и в неокортексе (см. Материалы и методы). Пример самовозгорания показан на рисунке 1 C .
Рисунок 1.
Имитация микросхем ТТС. ( A ) Образец микросхемы на 50 ячеек. Вероятность соединения между ячейками составляла 0,13 для однонаправленных соединений (черные края) и 0.06 для ответных соединений (красные края). Реципрокные возбуждающие синаптические связи в 1,5 раза сильнее однонаправленных. ( B ) Пример смоделированной TTC с ее подробной морфологией, получающей фоновый пуассоновский ввод от 10 000 возбуждающих синапсов (красный) и 2500 тормозных (синий) синапсов (см. Материалы и методы). В дополнение к этому фоновому входу все клетки в микросхеме одновременно стимулировались кратковременным вводом соматического тока 1,4 нА и длительностью 5 мс (черный электрод), имитируя кратковременный перисоматический ввод таламуса.В дополнение к запуску соматических AP Na + , модель TTC была способна генерировать дендритные шипы Ca 2+ в апикальной «горячей зоне» (заштрихованный красный эллипс). ( C ) Самопроизвольное срабатывание типичного одиночного (изолированного) TTC в ответ на фоновый синаптический ввод. ( D ) Следы напряжения другой примерной ячейки, от микросхемы из 150 ячеек, спонтанно срабатывающих (2 пика, слева) и в ответ на одновременный соматический ввод во все ячейки сети (4 пика, справа).Стимул обозначается черной полосой под кривой напряжения.
Рисунок 1.
Имитация микросхем ТТС. ( A ) Образец микросхемы на 50 ячеек. Вероятность соединения между ячейками составляла 0,13 для однонаправленных соединений (черные края) и 0,06 для взаимных соединений (красные края). Реципрокные возбуждающие синаптические связи в 1,5 раза сильнее однонаправленных. ( B ) Пример смоделированной TTC с ее подробной морфологией, получающей фоновый пуассоновский ввод от 10 000 возбуждающих синапсов (красный) и 2500 тормозных (синий) синапсов (см. Материалы и методы).В дополнение к этому фоновому входу все клетки в микросхеме одновременно стимулировались кратковременным вводом соматического тока 1,4 нА и длительностью 5 мс (черный электрод), имитируя кратковременный перисоматический ввод таламуса. В дополнение к запуску соматических AP Na + , модель TTC была способна генерировать дендритные шипы Ca 2+ в апикальной «горячей зоне» (заштрихованный красный эллипс). ( C ) Самопроизвольное срабатывание типичного одиночного (изолированного) TTC в ответ на фоновый синаптический ввод.( D ) Следы напряжения другой примерной ячейки, от микросхемы из 150 ячеек, спонтанно срабатывающих (2 пика, слева) и в ответ на одновременный соматический ввод во все ячейки сети (4 пика, справа). Стимул обозначается черной полосой под кривой напряжения.
Нас сначала интересовала активность клеток после единственного соматического спайка, индуцированного синхронно во всех клетках. Поэтому мы применили кратковременный (5 мс) надпороговый соматический ввод 1,4 нА ко всем клеткам в цепи (пример кривой показан на рис.1 D ). Одиночные (несвязанные) клетки ответили на этот соматический ввод одиночным спайком, привязанным к времени стимула, t = 0, без последующего увеличения частоты активации сверх базовой частоты (рис. 2 A ). Напротив, в микросхемах из 150 клеток соматические стимулы запускали повышенную активность в цепи на продолжительность ~ 70 мс, после чего активность возвращалась к исходному уровню (рис. 2 B ). Мы сосредоточили анализ на временном окне 50 мс после начала стимула ( t = 0–50 мс) и рассчитали, что средняя частота срабатывания в микросхемах из N = 150 ячеек равна 47.1 ± 3,9 Гц для этого временного окна. Средняя частота срабатывания ответа в одиночных неподключенных ячейках в течение окна 50 мс составляла 22,1 ± 0,7 Гц.
Рисунок 2.
Модуляция скорости ответа за счет повторяющейся активности и дендритной возбудимости. ( A ) Пример растрового графика (вверху) и PSTH (внизу) активации одиночных (несвязанных) клеток в ответ на кратковременный надпороговый соматический / таламический вход при t = 0 (стимул, как на рис. 1 B ).( B ) Пример растрового графика (вверху) и PSTH (внизу) для ответа на соматический ввод в микросхемах из N = 150 клеток, все из которых получают кратковременный надпороговый соматический ввод при t = 0. ( C ) То же, что и в ( B ), за исключением того, что были удалены апикальные дендритные каналы Na + в смоделированных клетках (обозначены: -a.Na). ( D ) Частота ответа клеток в микросхемах размером N = 1–150 клеток, в контрольных условиях, как в ( B ) (черная кривая), и при удалении определенных апикальных дендритных каналов (высоковольтно-управляемые Ca 2+ каналов, зеленая кривая; Ca 2+ каналов, управляемых высоким и низким напряжением, красная кривая; каналы Na + , фиолетовая кривая) или рецепторов NMDA на базальных и апикальных дендритах (синяя кривая) .Мгновенную скорость измеряли во временном окне 50 мс после появления стимула.
Рисунок 2.
Модуляция скорости ответа за счет повторяющейся активности и дендритной возбудимости. ( A ) Пример растрового графика (вверху) и PSTH (внизу) активации одиночных (несвязанных) клеток в ответ на кратковременный надпороговый соматический / таламический вход при t = 0 (стимул, как на рис. 1 B ). ( B ) Примерный растровый график (вверху) и PSTH (внизу) для ответа на соматический ввод в микросхемах из N = 150 клеток, все получают кратковременный надпороговый соматический ввод при t = 0.( C ) То же, что и в ( B ), за исключением того, что были удалены апикальные дендритные каналы Na + в смоделированных клетках (обозначены: -a.Na). ( D ) Частота ответа клеток в микросхемах размером N = 1–150 клеток, в контрольных условиях, как в ( B ) (черная кривая), и при удалении определенных апикальных дендритных каналов (высоковольтно-управляемые Ca 2+ каналов, зеленая кривая; Ca 2+ каналов, управляемых высоким и низким напряжением, красная кривая; каналы Na + , фиолетовая кривая) или рецепторов NMDA на базальных и апикальных дендритах (синяя кривая) .Мгновенную скорость измеряли во временном окне 50 мс после появления стимула.
Чтобы определить степень влияния дендритной возбудимости на активность микросхемы, мы исследовали аналогичные микросхемы, в которых были удалены определенные дендритные каналы или рецепторы NMDA. Мы отмечаем, что удаление рецепторов NMDA было сделано путем установки постоянных времени нарастания и затухания компонента NMDA в возбуждающем синапсовом механизме как бесконечно малые. Кроме того, когда мы удалили рецепторы NMDA, мы увеличили максимальную возбуждающую проводимость, чтобы спонтанные скорости оставались неизменными.Удаление дендритных каналов Na + существенно снизило длительное увеличение сетевой активности после первого индуцированного спайка, так что средний ответ в микросхемах из N = 150 клеток составил 24,7 ± 1 Гц (рис. 2 C ), только немного выше, чем отклик неподключенных ячеек (22,1 ± 0,7 Гц). Удаление дендритных каналов Ca 2+ уменьшило ответ микросхемы до 41,1 ± 2,8 Гц (рис. 2 D , красная кривая), что на 13% меньше скорости по сравнению с контрольными условиями (см. Выше).Удаление только высоковольтно-активированных дендритных каналов Ca 2+ привело к аналогичному эффекту (рис.2 D , зеленая кривая), тогда как удаление низковольтно-активированных дендритных каналов Ca 2+ не имело значительного эффекта ( не показано). Блокирование каналов Na + или рецепторов NMDA имело аналогичный эффект, отменяя длительное увеличение сетевой активности после второго спайка (рис. 2 D , фиолетовый и синий, соответственно). Эта ограниченная повторяющаяся активность в микросхемах с невозбужденными дендритами ожидается из-за их небольшого размера и низкой вероятности соединения между клетками, а также поскольку все TTC в подключенной сети получают одновременный соматический ввод (как один дополнительный всплеск), который не может эффективно распространяться через сеть из-за рефрактерности во всех клетках после этого всплеска.Отметим, что удаление дендритных каналов Ca 2+ и Na + уменьшало спонтанное возбуждение (2,6 ± 0,5 Гц по сравнению с 4,3 ± 1,5 Гц в контрольных условиях в микросхемах из 150 клеток). Однако обе эти нормы были в пределах физиологического диапазона (см. Материалы и методы). Мы также отмечаем, что вариабельность реакции или скорости спонтанного возбуждения наблюдалась in vivo (Heimel et al. 2005).
Повторяющаяся активность и дендритная возбудимость в микросхемах N = 150 клеток с полностью возбудимыми дендритами привели к частоте ответа, равной 2.В 3 раза больше, чем скорость отклика одиночных (несвязанных) клеток без дендритных каналов Na + (47,1 ± 3,9 Гц против 20,3 ± 0,4 Гц, рис. 2 D ). Скорость отклика увеличивалась сверхлинейно в зависимости от количества ячеек в микросхеме (рис. 2 D , черная кривая), но лишь немного увеличивалась с увеличением количества ячеек в микросхемах с невозбужденными дендритами (рис. 2 D , рис. фиолетовые или синие кривые). Взятые вместе, эти результаты демонстрируют, что как возбудимость дендритов, так и рекуррентная активность микросхем способствовали наблюдаемой модуляции скорости ответа.Из Рисунка 2 D очевидно, что дендритная нелинейность играет главную роль в усилении сетевой активности после одновременной надпороговой соматической стимуляции (имитирующей кратковременный перисоматический таламический сигнал).
Мы дополнительно исследовали участие дендритных потенциал-зависимых ионных каналов и NMDA-рецепторов в формировании ответной реакции сети, исследуя следы дендритного и соматического напряжения в микросхемах N = 150 клеток. Для образцовой ячейки (рис.3 A ) в контрольных условиях после кратковременного соматического / таламического воздействия наблюдалась большая деполяризация на главной апикальной бифуркации, которая длилась ~ 70 мс после окончания соматического стимула. Эта дендритная деполяризация обычно сопровождалась всплеском 2 дополнительных спайков в соме. Дендритная деполяризация была порядка величины и продолжительности спайков Ca 2+ во время срабатывания BAC, наблюдаемых in vitro (Larkum et al. 1999, 2004; Hay et al. 2011). Добавление 2 соматических спайков с коротким интервалом между спайками (<30 мс) также было типичным для активации ВАС.Когда дендритные каналы Ca 2+ были удалены, дендритная деполяризация значительно уменьшилась, как и один из дополнительных соматических всплесков Na + (рис. 3 B ), тем самым подтверждая, что дендритная деполяризация и увеличение соматического ответа частично опосредуются потенциалозависимыми дендритными каналами Ca 2+ . Когда NMDA-рецепторы также были удалены, дендритная деполяризация исчезла, а вместе с ней и дополнительный соматический спайк Na + (рис.3 С ). Оставшаяся деполяризация дендритов произошла из-за ВПСП, исходящих от микросхемы. Удаление только дендритных каналов Na + было достаточным для устранения дендритной деполяризации и соматических шипов (рис. 3 D ). Взятые вместе, эти результаты показывают, что в контрольных условиях BAP (опосредованный дендритными Na + каналами) запускает деполяризацию NMDA и срабатывание BAC, что согласуется с недавними исследованиями (Larkum et al. 2009).
Рисунок 3.
Отклик микросхемы связан с сильной дендритной нелинейностью. ( A ) Следы напряжения на соме (черный) и на апикальной главной бифуркации (650 мкм от сомы, красный цвет, см. Схему на рис. 1 B ) образцовой ячейки. Ячейка была частью микросхемы размером N = 150 ячеек, получая кратковременный надпороговый соматический ввод на все ячейки схемы в контрольных условиях. Соматический стимул при t = 0 обозначен черной полосой под кривой. Область под ответом дендритного напряжения, для t = 0–50 мс, заштрихована красным.( B — D ) То же, что ( A ), но для случаев, когда были удалены дендритные каналы Ca 2+ ( B ), а также были удалены рецепторы NMDA ( C ), или были удалены только дендритные каналы Na + ( D ). ( E ) Распределение интеграла деполяризации мембраны в апикальной главной бифуркации во временном окне 50 мс во всех ячейках из микросхем из 150 ячеек из 4 различных рандомизированных симуляций ( n = 600 ячеек для каждой кривой, см. Материалы и методы), получая одновременный соматический ввод, в контрольных условиях (сплошная черная кривая), когда были удалены дендритные каналы Ca 2+ (красная кривая), когда также были удалены каналы NMDA (синяя кривая) или когда был удален только дендритный Na + каналов удалены (фиолетовая кривая).Распределение неподключенных ячеек в контрольных условиях показано пунктирной черной кривой ( n = 400 ячеек). Бимодальное распределение для сплошной черной кривой указывает на нелинейные дендритные события.
Рисунок 3.
Отклик микросхемы включает сильные дендритные нелинейности. ( A ) Следы напряжения на соме (черный) и на апикальной главной бифуркации (650 мкм от сомы, красный цвет, см. Схему на рис. 1 B ) образцовой ячейки. Ячейка была частью микросхемы размером N = 150 ячеек, получая кратковременный надпороговый соматический ввод на все ячейки схемы в контрольных условиях.Соматический стимул при t = 0 обозначен черной полосой под кривой. Область под ответом дендритного напряжения, для t = 0–50 мс, заштрихована красным. ( B — D ) То же, что ( A ), но для случаев, когда были удалены дендритные каналы Ca 2+ ( B ), а также были удалены рецепторы NMDA ( C ), или были удалены только дендритные каналы Na + ( D ). ( E ) Распределение интеграла деполяризации мембраны в апикальной главной бифуркации во временном окне 50 мс во всех ячейках из микросхем из 150 ячеек из 4 различных рандомизированных симуляций ( n = 600 ячеек для каждой кривой, см. Материалы и методы), получая одновременный соматический ввод, в контрольных условиях (сплошная черная кривая), когда были удалены дендритные каналы Ca 2+ (красная кривая), когда также были удалены каналы NMDA (синяя кривая) или когда был удален только дендритный Na + каналов удалены (фиолетовая кривая).Распределение неподключенных ячеек в контрольных условиях показано пунктирной черной кривой ( n = 400 ячеек). Бимодальное распределение для сплошной черной кривой указывает на нелинейные дендритные события.
Характеризовать вклад и нелинейную природу дендритных явлений труднее в условиях синаптического шума, чем в условиях покоя. Тем не менее, распределение интеграла дендритного напряжения по t = 0–50 мс в микросхемах из 150 клеток, получающих одновременный соматический ввод, иллюстрирует дендритные события во время сетевой активности (рис.3 E ). В контрольных условиях наблюдалась большая дендритная деполяризация, величина которой в разных клетках имела бимодальное распределение с одним пиком ∼500 мВ мс и другим пиком ∼1300 мВ мс, что указывает на сильное надлинейное дендритное событие в некоторых клетках (рис. 3). E , черная кривая). Особенно большие интегральные значения (> 800 мВ мс) наблюдались в 55% клеток и отражали дендритную деполяризацию с продолжительностью и величиной, типичными для всплесков Ca 2+ , таких как показано на рисунке 3 A .По-видимому, для этих клеток входные условия (как пространственные, так и временные) оказались благоприятными для запуска сильных дендритных нелинейностей. Когда дендритные каналы Ca 2+ были удалены, распределение дендритного интеграла напряжения было унимодальным и в среднем 700 мВ мс (рис. 3 E , красная кривая). Когда NMDA-рецепторы также были удалены или когда были удалены только дендритные ионные каналы Na + , среднее значение деполяризации дендритов снизилось до 100 мВ мс и никогда не было> 700 мВ мс, что указывает на слабое и по существу линейное электрическое поведение дендритов (рис.3 E , синие и фиолетовые кривые соответственно). Для несвязанных клеток, где не было значительной активности за пределами индуцированного соматического спайка (рис. 2 A ), большинство клеток демонстрировали одномодальное распределение интеграла дендритного напряжения с центром ~ 100 мВ мс. Большая дендритная деполяризация происходила только в редких случаях в несвязанных клетках (в <3% клеток), когда самого фонового ввода оказывалось достаточно для запуска активации ВАС (рис. 3 E , пунктирная черная кривая).
Поскольку не все клетки в корковой цепи, вероятно, будут получать таламический вход в одно и то же время, мы исследовали влияние временного шума во входных данных на наши результаты, применив случайный джиттер в 10 мс к началу соматического стимула. (т.е. начало может быть в любой момент времени между t = 0 и t = 10 мс). Рис. 4 A показывает растровый график и PSTH в микросхемах из 150 ячеек в этом зашумленном состоянии. Явное отличие от состояния отсутствия шума (рис.2 B ) было отсутствие равномерного пика по всем клеткам в момент времени t = 0 мс, поскольку соматический стимул мог произойти где угодно между 0 и 10 мс. Скорость ответа в микросхемах из 150 клеток была в 2,1 раза больше, чем скорость ответа изолированных клеток без дендритных каналов Na + (43,2 ± 2,8 Гц против 20,5 ± 0,4 Гц, рис. 4 B ). Следовательно, даже при временном шуме на входе таламуса модуляция скорости отклика из-за дендритных нелинейностей и повторяющейся сетевой активности оставалась устойчивой и аналогичной по величине усилению в контрольных условиях (без джиттера).Как и раньше, вклад как дендритной возбудимости, так и рекуррентной связности был необходим для наблюдаемой модуляции, поскольку ответ был низким в отдельных клетках или в микросхемах клеток с невозбужденными дендритами (сравните Фиг.2 D и 4 B ). Дендритные нелинейности и срабатывание BAC также происходили в условиях зашумленного входа, что продемонстрировано бимодальным распределением интеграла дендритной деполяризации (рис. 4 C , сплошная черная кривая). Доля клеток, демонстрирующих большую дендритную деполяризацию (дендритный интеграл> 800 мВ мс), была немного уменьшена, 44% по сравнению с 55% в случае отсутствия дрожания (см. Выше).Как и в случае отсутствия дрожания, удаление дендритных каналов Ca 2+ устраняет срабатывание BAC в связанных клетках (рис. 4 C , красная кривая), а удаление дендритных каналов Na + устраняет дендритную деполяризацию (рис. 4 C). , пурпурная кривая). Точно так же для нестабильного входа неподключенные ячейки (размер микросхемы N = 1) показали лишь незначительную модуляцию скорости ответа из-за дендритной возбудимости (рис.4 B ) и обычно не проявляли срабатывания BAC или деполяризации NMDA (рис.4 C , пунктирная черная кривая).
Рисунок 4.
Модуляция отклика сети за счет повторяющейся активности и возбудимости дендритов устойчива к временному шуму на входе таламуса. ( A ) Растровый график (вверху) и PSTH (внизу) для микросхем из 150 клеток, получающих короткий надпороговый соматический ввод, со случайным джиттером 0–10 мс в начале соматического стимула. ( B ) Ответ клеток в микросхемах размером N = 1–150, получающих соматический ввод с джиттером 0–10 мс, в контрольных условиях (черная кривая) и при дендритных каналах Ca 2+ или Na + были удалены (красная и пурпурная кривые).( C ) Распределение интеграла напряжения на главной апикальной бифуркации во всех ячейках от микросхем из 150 ячеек ( n = 600), получающих дрожащий соматический вход, в контрольных условиях (сплошная черная кривая) и при дендритном Ca 2+ Каналы или Na + были удалены (красные и фиолетовые кривые). Распределение дендритного интеграла для несвязанных ячеек представлено пунктирной черной кривой ( n = 400).
Рисунок 4.
Модуляция отклика сети за счет повторяющейся активности и дендритной возбудимости устойчива к временному шуму на входе таламуса.( A ) Растровый график (вверху) и PSTH (внизу) для микросхем из 150 клеток, получающих короткий надпороговый соматический ввод, со случайным джиттером 0–10 мс в начале соматического стимула. ( B ) Ответ клеток в микросхемах размером N = 1–150, получающих соматический ввод с джиттером 0–10 мс, в контрольных условиях (черная кривая) и при дендритных каналах Ca 2+ или Na + были удалены (красная и пурпурная кривые). ( C ) Распределение интеграла напряжения на главной апикальной бифуркации во всех ячейках от микросхем из 150 ячеек ( n = 600), получающих дрожащий соматический вход, в контрольных условиях (сплошная черная кривая) и при дендритном Ca 2+ Каналы или Na + были удалены (красные и фиолетовые кривые).Распределение дендритного интеграла для несвязанных ячеек представлено пунктирной черной кривой ( n = 400).
Мы также исследовали случай, когда только часть микросхем из 150 клеток получила таламический ввод (рис. 5). В полностью возбудимой сети скорость ответа линейно увеличивалась с количеством активированных клеток (рис. 5 B ). Рисунок 5 B также показывает, что дендритная нелинейность проявляется даже тогда, когда небольшой процент клеток активируется через таламический вход (сравните черные и цветные линии).
Рисунок 5.
Ответ микросхемы на частичный таламический ввод. ( A ) Пример растрового графика (вверху) и PSTH (внизу) для ответа на соматический / таламический вход, заданный 60% клеток в микросхемах, состоящих из 150 клеток. ( B ) Скорость ответа в микросхемах из 150 клеток в зависимости от процента стимулированных клеток в контрольных условиях (черная кривая) и при удалении дендритных каналов Ca 2+ или Na + (красный и фиолетовый кривые).
Рисунок 5.
Ответ микросхемы на частичный таламический ввод. ( A ) Пример растрового графика (вверху) и PSTH (внизу) для ответа на соматический / таламический вход, заданный 60% клеток в микросхемах, состоящих из 150 клеток. ( B ) Скорость ответа в микросхемах из 150 клеток в зависимости от процента стимулированных клеток в контрольных условиях (черная кривая) и при удалении дендритных каналов Ca 2+ или Na + (красный и фиолетовый кривые).
Вычислительные последствия модуляции ответа из-за дендритной возбудимости и повторяющейся активности, изображенные выше, могут быть использованы для усиления и / или формирования селективности ответа кортикальных клеток на вход таламуса (Douglas et al. 1995; Liu et al. 2007; Ли, Ли и др., 2013 г .; Ли, Ибрагим и др., 2013 г .; Лиен, Сканциани, 2013 г.). Мы исследовали эти возможности, используя нашу сеть TTCs, имитируя визуальный ввод различных ориентаций, поступающих из таламуса. Различные визуальные ориентации эмулировались путем изменения амплитуды короткого (5 мс) импульса, представляющего таламический вход (см. Материалы и методы).В контрольных условиях с возбудимыми дендритами ответ в микросхемах из 150 клеток был усилен в 2,3 раза по сравнению с ответом одиночных несвязанных клеток (фиг. 6 A , сплошные кривые против пунктирных черных кривых). Однако не было изменений в селективности (в форме кривой настройки) ячеек (фиг. 6 B , сплошные и пунктирные черные кривые). А именно, сеть имела мультипликативный эффект, который масштабировал кривую настройки одинаково для всех входных углов.
Рисунок 6.
Дендритная возбудимость и повторяющаяся активность усиливают ответ на таламический вход, сохраняя при этом избирательность. ( A ) Средний ответ в микросхемах из 150 клеток (сплошные кривые) или в несвязанных клетках (пунктирная черная кривая) на различные амплитуды соматических импульсов, отражающих различные ориентации стимулов (см. Материалы и методы). Контрольные условия изображены черными кривыми, тогда как цветные кривые соответствуют случаям, когда дендритные каналы Ca 2+ или Na + , а также рецепторы NMDA были удалены.( B ) Кривые нормализованного отклика взяты из ( A ). ( C ) Вероятность больших дендритных деполяризаций в микросхемах из 150 клеток увеличивалась с ориентацией стимула в сторону предпочтительной ориентации (90 °). Большая дендритная деполяризация была определена как случай, когда интеграл дендритного напряжения на главной апикальной бифуркации составлял> 800 мВ мс. ( D ) Прогнозируемая скорость ответа в микросхемах из 150 клеток, основанная на вероятности получения большой дендритной деполяризации, как функция ориентации (приведенная в C ) показана зеленой линией.Пурпурная и черная кривые такие же, как у ( A ).
Рисунок 6.
Дендритная возбудимость и рекуррентная активность усиливают ответ на таламический вход, сохраняя при этом избирательность. ( A ) Средний ответ в микросхемах из 150 клеток (сплошные кривые) или в несвязанных клетках (пунктирная черная кривая) на различные амплитуды соматических импульсов, отражающих различные ориентации стимулов (см. Материалы и методы). Контрольные условия изображены черными кривыми, тогда как цветные кривые соответствуют случаям, когда дендритные каналы Ca 2+ или Na + , а также рецепторы NMDA были удалены.( B ) Кривые нормализованного отклика взяты из ( A ). ( C ) Вероятность больших дендритных деполяризаций в микросхемах из 150 клеток увеличивалась с ориентацией стимула в сторону предпочтительной ориентации (90 °). Большая дендритная деполяризация была определена как случай, когда интеграл дендритного напряжения на главной апикальной бифуркации составлял> 800 мВ мс. ( D ) Прогнозируемая скорость ответа в микросхемах из 150 клеток, основанная на вероятности получения большой дендритной деполяризации, как функция ориентации (приведенная в C ) показана зеленой линией.Пурпурная и черная кривые такие же, как у ( A ).
Мы количественно оценили селективность в каждом случае, используя полуширину на полувысоте, которая характеризует селективность вокруг предпочтительной ориентации; а также использовали индекс селективности, который является более глобальным показателем селективности (см. Материалы и методы). В обоих случаях ячейки имели среднюю полуширину настройки на полувысоте 29 °, вокруг предпочтительного угла (произвольно назначенного равным 90 °). Индекс селективности был несколько меньше в неподключенных ячейках по сравнению с микросхемами из 150 ячеек (0.45 против 0,53 соответственно) из-за уменьшения разницы между нормализованными ответами на предпочтительную и нулевую ориентации.
Интересно, что когда дендритные каналы Na + были удалены, усиление было отменено, а средняя кривая настройки в ячейках микросхем 150 была расширена неоднородным образом (рис. 6 A , B , фиолетовая кривая). Центральная часть кривой (вокруг предпочтительной ориентации) выровнялась, в результате чего микросхема реагировала одинаково на все углы от 70 ° до 110 °.Полуширина перестройки на полувысоте увеличилась с 29 ° до 39 °. Общий индекс селективности в этом случае был лишь незначительно снижен (0,48 по сравнению с 0,53 в контроле) из-за резкости кривой. Следовательно, удаление возбудимых каналов из дендритов в микросхемах из 150 ячеек привело к общему снижению селективности на 35%, а также к полному устранению селективности для углов в пределах 20 ° от исходной предпочтительной ориентации. Напротив, удаление дендритных каналов в отдельных несвязанных клетках не изменило их селективности (не показано).Небольшое и широкое усиление из-за повторяющейся активности микросхемы в случае удаленных дендритных каналов Na + выравнивается на ∼22 Гц (рис. 6 A , фиолетовая кривая). Таким образом, повторяющаяся активность привела к пороговому значению всех клеток, в которых нежелательные углы (более слабые таламические стимулы) были недостаточны для запуска возбуждения, в результате все клетки микросхемы запускали один AP (что эквивалентно средней скорости ответа ~ 20 Гц в Временное окно 50 мс) для широкого диапазона углов от предпочтительной ориентации.Как отмечалось выше, одна рекуррентная активность (без дендритной возбудимости) не была эффективной для запуска более одного спайка на клетку из-за рефрактерности спайков и низкой вероятности соединений между клетками в микросхеме. Следовательно, при усилении ответа кортикальными микросхемами дендритная возбудимость имела решающее значение для увеличения скорости ответа таким образом, чтобы поддерживать селективность стимула, индуцированную в отдельных клетках. Удаление дендритных каналов Ca 2+ частично уменьшило усиление и увеличило ширину перестройки (рис.6 A , B , красная кривая), тогда как удаление рецепторов NMDA еще больше расширило ширину настройки до такой же степени, что и при удалении дендритных каналов Na + (Рис.6 A , B , синяя кривая ). В этом случае верхняя часть кривой не была плоской, что указывает на то, что дендритные каналы Na + вносят непосредственный вклад в усиление соматического ответа вблизи предпочтительных ориентаций.
Участие дендритной возбудимости в противодействии широкой амплификации из-за рекуррентной активности также было очевидно из связи между вероятностью достижения большой дендритной деполяризации (интеграл дендритного напряжения> 800 мВ мс, либо через возбуждение NMDA или BAC) и ориентацией стимула в микросхемы из 150 ячеек (рис.6 С ). При предпочтительной ориентации (90 °) вероятность достижения большой дендритной деполяризации составляла 55% и почти линейно снижалась до низкой вероятности 1% при ориентациях дальше 45 ° от предпочтительной ориентации. Это указывало на то, что возбудимость дендритов усиливала ответ в большей степени, поскольку ориентация стимула была ближе к предпочтительной ориентации. Действительно, если предположить, что во время срабатывания BAC произойдет выброс из 3 AP, прогнозируемая скорость реакции из-за этого увеличения вероятности большой дендритной деполяризации хорошо учитывала резкую селективность по сравнению с таковой в случае невозбудимых дендритов (рис.6 D , зеленая линия против фиолетовой).
Моделирование таламического входа с использованием соматического тока в форме ВПСП (а не короткого импульса тока) с постоянной времени затухания 5 мс (Константинополь и Бруно, 2013) или замена соматического стимула комбинацией соматического стимула и дендритного стимула в основная бифуркация существенно не изменила результаты (не показано). Мы отмечаем, что, поскольку эффект усиления примерно пропорционален изменению коэффициента усиления между максимальным и минимальным откликом, величина усиления должна изменяться в зависимости от размера микросхемы, так как скорости отклика показаны на рисунке 2 D .
Обсуждение
Путем анализа повторяющихся микросхем подробных взаимосвязанных клеток с толстыми пучками L5, получающих таламический вход, мы предоставили первое указание на то, что дендритная возбудимость вносит значительный вклад в сетевую модуляцию ответа на восходящий (таламический) вход. Мы показали, что значительная часть этой модуляции опосредована BAP, запускающим нелинейности NMDA, а также активацией BAC (Larkum et al., 1999; Stuart and Hausser, 2001) из-за совпадения спайка, вызванного таламическим входом и интракортикальным (дендритным) вход, возникающий из локальной корковой микросхемы клеток с одинаковым предпочтением стимулов.Мы показываем, что этот усиленный ответ из-за дендритной нелинейности может служить механизмом для линейного умножения таламокортикальных сигналов — усиления ответа на восходящий вход при сохранении селективности клеток (формы их настраиваемой кривой).
Кортикальная амплификация была оценена экспериментально только в возбуждающих клетках из L4. Эти клетки, как известно, не генерируют феномен активации BAC, однако они демонстрируют дендритные шипы NMDA и рекуррентно связаны аналогично L5 TTC (Thomson and Lamy 2007; Lavzin et al.2012). Недавние экспериментальные исследования показали, что повторяющаяся корковая активность увеличивает скорость ответа клеток L4 на таламический вход в 2-3 раза без изменения селективности ответа (Li, Li et al. 2013; Li, Ibrahim et al. 2013; Lien and Scanziani 2013). Вовлечет ли это усиление в L4 всплески NMDA и / или точно настроенное ингибирование (см. Ниже) в дополнение к повторяющейся возбуждающей активности микросхемы, еще предстоит определить. Наше моделирование показывает, что повторяющаяся активность в микросхемах 150 TTC привела к аналогичному увеличению скорости отклика (коэффициент 2.3, неизменная селективность стимула), и что режим по умолчанию этого усиления основан на дендритной нелинейности (срабатывание NMDA и BAC). Микросхемы такого размера были способны обеспечивать дендритный вход, необходимый для запуска BAC-импульса после одиночного соматического всплеска, вызванного таламическим входом. Для усиления были необходимы каналы NMDA и дендритный Na + . Таким образом, соматический спайк Na + распространялся обратно в дендриты и суммировался с дендритным входом, поступающим от других клеток микросхемы, вызывая опосредованную NMDA деполяризацию и локальный дендритный спайк Ca 2+ , который, в свою очередь, приводил к возникновению высокочастотный всплеск дополнительных всплесков Na + в соме, как ранее было охарактеризовано in vitro (Larkum et al.1999, 2004). Без дендритной возбудимости небольшие сети из нескольких сотен кортикальных клеток были способны лишь к незначительному и неспецифическому усилению одноклеточного ответа в изолированном состоянии, что увеличивало селективность ответа почти на 40% (рис. 6). Таким образом, наши результаты показывают, что дендритная возбудимость увеличивает вычислительную мощность корковых сетей и, в частности, имеет решающее значение для повышения активности микросхем с аналогичным предпочтением стимулов при сохранении избирательности ответа.
Участие активной дендритной проводимости в корковой динамике также предсказано недавними исследованиями потенциала локального поля в смоделированном крупномасштабном кортикальном контуре (Reimann et al. 2013). Будущие достижения в экспериментальных методах должны позволить избирательно блокировать дендритную возбудимость в неповрежденном мозге и, таким образом, проверить наши прогнозы относительно мультипликативной роли дендритной возбудимости. Мы отмечаем, что этот режим корковой модуляции может быть эффективно настроен с помощью целевого дендритного ингибирования, например, ингибирования, которое предпочтительно блокирует дендритный спайк Ca 2+ (Gidon and Segev 2012).
Важная роль токов NMDA в модуляции клеточного ответа согласуется с предыдущими исследованиями (Schiller et al. 2000; Losonczy and Magee 2006; Lavzin et al. 2012; Smith et al. 2013). Хотя подавление с прямой связью было достаточным для ограничения микросхем, возможно, что повторяющееся подавление в смоделированной микросхеме может ограничить активность схемы в достаточной степени, что потребует еще большего вовлечения нелинейностей NMDA для усиления срабатывания BAC (Larkum et al.2009 г.). В будущих исследованиях следует изучить эту возможность путем включения повторяющегося ингибирования, такого как интернейроны Мартинотти, которые образуют дисинаптическую петлю с пирамидными клетками (Silberberg and Markram 2007). Мы ожидаем, что основной эффект такого включения потребует активации микросхем большего размера для достижения того же усиления, которое мы наблюдали здесь. В самом деле, большие микросхемы при наших упрощенных симуляциях имели аномально большую частоту спонтанных действий, которая, следовательно, лучше сдерживалась бы повторяющимся торможением.Вдобавок, клетки Мартинотти, нацеленные на пучки, позволят четко модулировать вклад NMDA по сравнению с активацией BAC в вычисления микросхемы.
Детальное понимание сетевой динамики в реальных биологических цепях в значительной степени отсутствует. По этой причине в настоящем исследовании мы построили более простой сетевой модуль, состоящий из локальной возбуждающей микросхемы L5, реально подключенной, при этом упростив вклад более крупной сети, допуская стохастический фоновый вход с разумными возбуждающими и тормозящими фоновыми синапсами, который воспроизводит экспериментально наблюдали самовозгорание в ТТК.В нашем моделировании, как и в предыдущих исследованиях повторяющихся микросхем (Дуглас и др., 1995), фоновое прямое ингибирование было достаточным, чтобы ограничить уровень активности в микросхемах, включающих до 150 клеток. Анатомические и функциональные исследования показывают, что корковая сеть действительно содержит кластеры клеток такого размера (Perin et al. 2011). Поскольку наши микросхемы из 150 клеток произвели амплификацию, аналогичную наблюдаемой экспериментально в L4, возможно, что эффективная локальная кортикальная активация включает такое же количество клеток.Следует изучить участие точно настроенного целевого ингибирования, растормаживания или даже нейромодуляции (Palmer et al. 2012; Haider et al. 2013; Pfeffer et al. 2013; Polack et al. 2013). Будущие теоретические исследования, следовательно, должны увеличить сложность сети, чтобы включить ингибирование обратной связи с помощью смоделированных тормозных клеток (Druckmann et al. 2007). Особый интерес представляют ингибирующие клетки, которые нацелены на дистальные дендриты в TTC и, таким образом, влияют на сцепление между соматическими и дендритными шипами (Murayama et al.2009; Гидон и Сегев 2012). Кроме того, включение рецепторов GABA B , которые, как было показано, модулируют управляемые по напряжению каналы Ca 2+ (Chalifoux and Carter 2011), позволит провести дальнейшее исследование средств точной настройки усиления и вычислений микросхем для Например, переключаясь с полной амплификации на частичную только с помощью NMDA, аналогично тому, что мы показали, блокируя каналы Ca 2+ и рецепторы NMDA.
Модуляция ответа была устойчивой к временному дрожанию (10 мс) таламического стимула.Этот таламический джиттер по порядку величины наблюдается в большинстве корковых слоев во время ответа in vivo (Heimel et al. 2005). Действительно, предыдущие исследования показывают, что высокосинхронный вход таламуса (Bruno and Sakmann, 2006) приводит к синхронному внутрикортикальному входу (Bruno 2011) и не должен вызывать большого дрожания. Устойчивость наших результатов в реалистичных шумных условиях указывает на устойчивость усиления ВПСП с помощью BAP, а также срабатывания BAC в таких ожидаемых нестабильных условиях в неповрежденном мозге и дополняет экспериментальные оценки временного окна 20-30 мс, в котором совпадающие ( соматические плюс дендритные) стимулы снижают порог генерации дендритного шипа Ca 2+ (Larkum et al.1999, 2004).
Было высказано предположение, что совпадение восходящего (таламического) входа и дендритного входа (происходящего как из локального контура, так и из других областей коры головного мозга, Феллеман и Ван Эссен, 1991) в TTCs лежит в основе когнитивных ассоциаций и перцептивного связывания внешних сенсорных сигналов. входы и внутренние корковые представления через дендритные нелинейности (Felleman and Van Essen 1991; Larkum 2012). Наше моделирование показывает, что повторяющаяся активность в локальных кортикальных микросхемах, состоящих из 50–150 TTCs, обеспечивает достаточный совпадающий дендритный вход для запуска дендритных нелинейностей и, таким образом, значительно модулирует восходящий сигнал.Более того, замена соматического стимула комбинацией соматического стимула и дендритного стимула на главной бифуркации не изменила наши результаты (не показаны). В будущих исследованиях следует изучить, как нисходящий дендритный вход из других областей коры головного мозга может способствовать этой модуляции, возможно, за счет снижения порога срабатывания дендритной нелинейности (Larkum et al. 2009) и тем самым увеличения мощности усиления для данного размера микросхемы.
Финансирование
Это исследование финансировалось Израильским научным фондом, проектным фондом EPFL для проекта Blue Brain Project, внутренней стипендией Э.Хэй из Междисциплинарного центра нейронных вычислений и Благотворительного фонда Гэтсби. Средства на оплату публикации в открытом доступе для этой статьи были предоставлены из вышеуказанных источников.
Банкноты
Конфликт интересов : Не объявлен.
Список литературы
.
2010
.Подслойные микросхемы кортикоспинальных и кортикостриатных нейронов моторной коры
.Nat Neurosci
.13
:739
—744
..
1995
.Теория настройки ориентации зрительной коры
.Proc Natl Acad Sci USA
.92
:3844
—3848
..
2004
.Количественная карта цепи первичной зрительной коры кошки
.Дж. Neurosci
.24
:8441
—8453
..
2011
.Синхронность в ощущении
.Curr Opin Neurobiol
.21
:701
—708
..
2006
.Кора головного мозга управляется слабыми, но синхронно активными таламокортикальными синапсами
.Наука
.312
:1622
—1627
..
2011
.Нейронная динамика как выборка: модель для стохастических вычислений в рекуррентных сетях импульсных нейронов
.PLoS Comput Biol
.7
:e1002211
..
2009
.Вычисления, зависящие от состояния: пространственно-временная обработка в корковых сетях
.Нат Рев Neurosci
.10
:113
—125
..
2006
.Книга НЕЙРОН
.Кембридж, Великобритания
/
Нью-Йорк
:Cambridge University Press
..
2011
.Модуляция рецептором GABAB чувствительных к напряжению кальциевых каналов в шипах и дендритах
.Дж. Neurosci
.31
:4221
—4232
..
1999
.Сложные клетки как простые клетки с кортикальной амплификацией
.Nat Neurosci
.2
:277
—282
..
2013
.Глубокие корковые слои активируются непосредственно таламусом
.Наука
.340
:1591
—1594
..
2011
.Эффекты и механизмы бодрствования на локальные корковые сети
.Нейрон
.69
:1061
—1068
..
2007
.Слоистая и специфичная для клеточного типа надпороговая репрезентация стимула в первичной соматосенсорной коре крыс
.Дж. Физиол
.581
:139
—154
..
1994
.Синтез моделей возбудимых мембран, синаптической передачи и нейромодуляции с использованием общего кинетического формализма
.Дж. Comput Neurosci
.1
:195
—230
..
1995
.Рекуррентное возбуждение в контурах неокортекса
.Наука
.269
:981
—985
..
2007
.Картирование матрицы: пути неокортекса
.Нейрон
.56
:226
—238
..
2004
.Нейронные цепи неокортекса
.Анну Рев Neurosci
.27
:419
—451
..
2007
.Новая структура многоцелевой оптимизации для ограничения моделей нейронов на основе проводимости экспериментальными данными
.Фронт Neurosci
.1
:7
—18
..
1991
.Распределенная иерархическая обработка в коре головного мозга приматов
.Cereb Cortex
.1
:1
—47
..
2007
.Постнатальное развитие синаптической передачи в локальных сетях пирамидных нейронов L5A соматосенсорной коры головного мозга крыс
.Дж. Физиол
.585
:103
—116
..
2002
.Кодирование временной информации синапсами, зависящими от активности
.Дж Нейрофизиол
.87
:140
—148
..
2010
.Динамика мембранного потенциала ГАМКергических нейронов в бочкообразной коре головного мозга мышей
.Нейрон
.65
:422
—435
..
2012
.Принципы действия синаптического торможения в дендритах
.Нейрон
.75
:330
—341
..
1979
.Морфология и интракортикальные проекции функционально охарактеризованных нейронов зрительной коры головного мозга кошки
.Природа
.280
:120
—125
..
2000
.Принципы организации разнообразных ГАМКергических интернейронов и синапсов в неокортексе
.Наука
.287
:273
—278
..
2013
.Торможение доминирует над сенсорными реакциями в бодрствующей коре головного мозга
.Природа
.493
:97
—100
..
2011
.Модели пирамидных клеток неокортикального слоя 5b, обладающие широким спектром дендритных и перисоматических активных свойств
.PLoS Comput Biol
.7
:e1002107
..
2005
.Ламинарная организация свойств ответа в первичной зрительной коре серого белка ( Sciurus carolinensis )
.Дж Нейрофизиол
.94
:3538
—3554
..
2010
.Биологически реалистичная корковая модель управления движением глаз при чтении
.Psychol Rev
.117
:808
—830
..
2007
.Модель микросхемы лобных полей глаза
.Дж. Neurosci
.27
:9341
—9353
..
1999
.Динамика дендритного кальция in vivo в пирамидных нейронах глубокого слоя коры
.Nat Neurosci
.2
:989
—996
..
1992
.Активация и десенсибилизация активированных глутаматом каналов, обеспечивающих быстрые возбуждающие синаптические токи в зрительной коре
.Нейрон
.9
:991
—999
..
2004
.ModelDB: база данных для поддержки вычислительной нейробиологии
.Дж. Comput Neurosci
.17
:7
—11
..
2006
.Кортикальные сети прямой связи для связывания различных потоков сенсорной информации
.Nat Neurosci
.9
:1472
—1473
..
2011
.Функциональная специфика локальных синаптических связей в неокортикальных сетях
.Природа
.473
:87
—91
..
2012
.Клеточный механизм корковых ассоциаций: принцип организации коры головного мозга
.Trends Neurosci
.36
:141
—151
..
2009
.Синаптическая интеграция в пучковые дендриты пирамидных нейронов слоя 5: новый объединяющий принцип
.Наука
.325
:756
—760
..
2004
.Нисходящий дендритный вход увеличивает усиление пирамидных нейронов слоя 5
.Cereb Cortex
.14
:1059
—1070
..
2002
.Передача сигналов слоя 1 и вызванных усами потенциалов действия Ca 2+ и Na + в дистальных и концевых дендритах пирамидных нейронов неокортекса крысы in vitro и in vivo
.Дж. Neurosci
.22
:6991
—7005
..
1999
.Новый клеточный механизм для связывания входов, поступающих в разные слои коры
.Природа
.398
:338
—341
..
2012
.Нелинейная дендритная обработка определяет угловую настройку нейронов бочкообразной коры in vivo
.Природа
.490
:397
—401
..
2013
.Линейное преобразование таламокортикального входа путем внутрикортикального возбуждения
.Nat Neurosci
.16
:1324
—1330
..
2013
.Интракортикальное умножение таламокортикальных сигналов в слуховой коре мышей
.Nat Neurosci
.16
:1179
—1181
..
2013
.Настроенное таламическое возбуждение усиливается зрительными кортикальными контурами
.Nat Neurosci
.16
:1315
—1323
..
2009
.Кортикальная схема, реализующая графические модели
.Нейронные вычисления
.21
:3010
—3056
..
2007
.Определение настройки корковой частоты с помощью схемы повторяющегося возбуждения
.Nat Neurosci
.10
:1594
—1600
..
2006
.Интегративные свойства радиальных косых дендритов в пирамидных нейронах СА1 гиппокампа
.Нейрон
.50
:291
—307
..
1997
.Физиология и анатомия синаптических связей между толстыми пучками пирамидных нейронов в развивающемся неокортексе крысы
.Дж. Физиол
.500
(Pt 2)
:409
—440
..
2010
.Тип клеток таламической иннервации в колонке вибриссальной коры крыс
.Cereb Cortex
.20
:2287
—2303
..
2009
.Дендритное кодирование сенсорных стимулов, контролируемых глубокими кортикальными интернейронами
.Природа
.457
:1137
—1141
..
2012
.Усиление контроля на шестом уровне в корковых цепях зрения
.Природа
.483
:47
—52
..
2012
.Клеточная основа межполушарного ингибирования, опосредованного ГАМК (В)
.Наука
.335
:989
—993
..
2014
.Шипы NMDA усиливают генерирование потенциала действия во время сенсорного ввода
.Nat Neurosci
.17
:383
—390
..
2013
.Индукция и модуляция постоянной активности в слое V микросхемы PFC модели
.Передние нейронные цепи
.7
:161
..
2011
.Принцип синаптической организации групп корковых нейронов
.Proc Natl Acad Sci USA
.108
:5419
—5424
..
2013
.Ингибирование торможения в зрительной коре: логика связей между молекулярно различными интернейронами
.Nat Neurosci
.16
:1068
—1076
..
2013
.Клеточные механизмы модуляции усиления в зависимости от состояния мозга в зрительной коре
.Nat Neurosci
.16
:1331
—1339
..
2004
.Вычислительные субъединицы в тонких дендритах пирамидных клеток
.Nat Neurosci
.7
:621
—627
..
2009
.Кодирование и декодирование пакетов с помощью пиков NMDA в базальных дендритах пирамидных нейронов 5-го слоя
.Дж. Neurosci
.29
:11891
—11903
..
2012
.Внутреннее морфологическое разнообразие пирамидных нейронов слоя 5 с толстыми пучками обеспечивает устойчивые и инвариантные свойства in silico синаптических связей
.Дж. Физиол
.590
:737
—752
..
2013
.Биофизически подробная модель неокортикальных локальных потенциалов поля предсказывает критическую роль активных мембранных токов
.Нейрон
.79
:375
—390
..
1999
.Переключатель развития при краткосрочной модификации унитарных ВПСП, вызванных в пирамидных нейронах 2/3 и 5 слоя неокортекса крысы
.Дж. Neurosci
.19
:3827
—3835
..
2006
.Свойства и значение шипов NMDA в пирамидных клетках неокортекса
.Дж. Neurosci
.26
:6704
—6715
..
2002
.Ориентационная селективность у макаки V1: разнообразие и ламинарная зависимость
.Дж. Neurosci
.22
:5639
—5651
..
1996
.Электрофизиологическое картирование опосредованного рецепторами GABAA ингибирования в соматосенсорной коре взрослых крыс
.Дж Нейрофизиол
.75
:1589
—1600
..
2007
.Моделирование модуля слой 4-слой 2/3 одного столбца в неокортексе крысы: переплетение экспериментальных наблюдений in vitro и in vivo
.Proc Natl Acad Sci USA
.104
:16353
—16358
..
2000
.NMDA-шипы в базальных дендритах кортикальных пирамидных нейронов
.Природа
.404
:285
—289
..
2001
.Опосредованные рецептором NMDA дендритные шипы и совпадающее усиление сигнала
.Curr Opin Neurobiol
.11
:343
—348
..
2007
.Дисинаптическое ингибирование пирамидных клеток коры неокортекса, опосредованное клетками Мартинотти
.Нейрон
.53
:735
—746
..
2013
.Дендритные шипы повышают избирательность стимулов в корковых нейронах in vivo
.Природа
.503
:115
—120
..
2005
.Сильно неслучайные особенности синаптической связности в локальных корковых цепях
.ПЛоС Биол
.3
:e68
..
2001
.Обнаружение дендритных совпадений ВПСП и потенциалов действия
.Nat Neurosci
.4
:63
—71
..
1995
.Моделирование направленной селективности простых клеток полосатой зрительной коры в рамках канонической микросхемы
.Дж. Neurosci
.15
:6700
—6719
..
2009
.Взаимодействия путей и синаптическая пластичность в дендритных областях пучков пирамидных нейронов CA1
.Нейрон
.62
:102
—111
..
2007
.Функциональные карты локальной схемотехники неокортекса
.Фронт Neurosci
.1
:19
—42
..
2005
.Модель таламокортикальной сети с одной колонкой, демонстрирующая гамма-колебания, сонное веретено и эпилептогенные всплески
.Дж Нейрофизиол
.93
:2194
—2232
..
1997
.Нейронный код между пирамидными нейронами неокортекса зависит от вероятности высвобождения нейромедиатора
.Proc Natl Acad Sci USA
.94
:719
—723
..
2013
.Временная синхронность и преобразование мощности гамма-тета в дендритах пирамидных нейронов CA1
.Nat Neurosci
.16
:1812
—1820
..
2007
.Сходство и разнообразие зрительной коры: существует ли объединяющая теория корковых вычислений?
Невролог
.13
:639
—656
..
2007
.Зависящая от пути динамика возбуждающей синаптической передачи в интракорковых цепях крыс, зависящая от использования
.Дж. Физиол
.585
:759
—777
..
2012
.Нелинейная дендритная интеграция сенсорной и моторной информации во время задачи активного зондирования
.Природа
.492
:247
—251
..
2005
.Мелкомасштабная специфичность корковых сетей зависит от типа тормозных клеток и связности
.Nat Neurosci
.8
:1552
—1559
..
2005
.Возбуждающие корковые нейроны образуют мелкомасштабные функциональные сети
.Природа
.433
:868
—873
.© Автор, 2014. Опубликовано Oxford University Press
Это статья в открытом доступе, распространяемая в соответствии с условиями некоммерческой лицензии Creative Commons Attribution (http://creativecommons.org/licenses/by-nc/4.0/) , который разрешает некоммерческое повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы. По вопросам коммерческого повторного использования обращайтесь в журналы[email protected]
Расшифровка схем сознания: от локальных микросхем до сетей масштаба мозга | Сеть Neuroscience
Понимание того, как сознание возникает в результате коммуникации между областями мозга, является вопросом первостепенной важности в области нейробиологии в целом и для диагностики и лечения пациентов, страдающих расстройствами сознания (DOC) в частности.Проблему сознания можно рассматривать как фундаментальную (например, «Что такое сознание?» И «Почему у нас есть субъективные, сознательные переживания?»; Такие вопросы называются «жесткой» проблемой сознания [Harnad, 1998]). или более эмпирический (например, «Какие процессы связаны с возникновением и поддержанием сознания?»; это формирует «мягкую» проблему сознания [Harnad, 1998]). В этом обзоре мы стремимся понять (1) как сети мозга на разных уровнях участвуют в обеспечении и поддержании сознательных процессов передачи и обработки информации, связанных с осознанием и бодрствованием, и (2) как эти механизмы связаны с нарушениями сознания. у пациентов с DOC.
Было предложено множество теорий, объясняющих происхождение сознания, от абстрактных и информационных концепций до теорий, основанных на нейрофизиологии. Наиболее распространенные теории сознания имеют общее фундаментальное предположение: обработка информации в сетях человеческого мозга неразрывно связана с сознанием. В недавней статье Дехаена и его коллег этот принцип резюмируется следующим образом (Dehaene, Lau, & Kouider, 2017): «То, что мы называем« сознанием », является результатом определенных типов вычислений по обработке информации, физически реализуемых аппаратными средствами мозга.”
В этом обзоре мы упоминаем три теории, которые кажутся наиболее известными в области исследования сознания. Однако читатель должен знать, что было предложено множество других теорий с разной степенью успеха. В рамках данного обзора, посвященного микро- и макросхемам, задействованным в сознании, мы сосредоточимся на этих трех основных теориях, которые можно связать с такими соображениями о свойствах сети мозга.Три основные теории сознания — это теория интегрированной информации (IIT; Tononi, 2004), гипотеза динамического ядра (DCH; Tononi & Edelman, 1998) и теория глобального рабочего пространства (GWT; Baars, 1988; Dehaene, Kerszberg, & Changeux, 1998; Dehaene, Sergent, & Changeux, 2003). Исторически теория DCH была первой, кто ссылался на понятие обработки информации, связанной с сознанием (Tononi & Edelman, 1998). Эта теория основана на центральной роли функциональных кластеров в таламо-корковой системе и реентерабельных взаимодействиях, при этом высокая интеграция и дифференциация нейрональной активности имеют решающее значение в возникновении сознательных явлений.ИИТ, который представляет собой эволюцию и обобщение DCH, основан на наборе аксиом, из которых вытекают постулаты. IIT также предоставляет вычислимую величину Φ, также называемую интегрированной информацией , которая количественно определяет уровень сознания. В случае ИИТ не было предложено никакого конкретного физиологического субстрата, поскольку эта теория в основном сосредоточена на понимании того, как сознание возникает в результате интеграции информации между несколькими системами. В этой структуре, если объединение подэлементов увеличивает возможности обработки информации больше, чем линейное добавление этих элементов, тогда увеличивается объем интегрированной информации.GWT — это теория теории сознания, которая более напрямую связана с нейрофизиологией и нейроанатомией. Основная гипотеза GWT состоит в том, что информация, связанная с сознательными процессами, доступна в глобальном масштабе в мозгу и что сосуществуют две принципиально разные вычислительные системы: (1) сеть распределенных «локальных» процессоров, работающих параллельно в мозгу («бессознательное »), И (2)« глобальное »рабочее пространство, образованное сетью распределенных взаимосвязанных областей коры головного мозга, участвующих в сознательном восприятии (Baars, 1988).В оставшейся части этого обзора следует проводить различие между понятиями «сознание» и «сознательное восприятие» с точки зрения субъективного опыта. Концепция сознания намного шире, чем сознательное восприятие, и включает, среди прочего, способность запоминать, воспринимать и сообщать, а также чувство самости (Seth, 2018). Сознательное восприятие относится, в частности, к возможности отчетности о фокусе нашего восприятия (Naccache, 2005). Ключевой концепцией здесь является то, что глобальное рабочее пространство состоит из удаленных регионов, плотно связанных глутаматергическими кортико-кортикальными связями, в отличие от сети локальных процессоров, работающих «изолированно» (параллельно).Стоит отметить, что это различие между бессознательными и сознательными процессами недавно было поставлено под сомнение и может быть чрезмерным упрощением (Melnikoff & Bargh, 2018). В GWT сознательное восприятие связано с «зажиганием», широкомасштабным паттерном активации мозга, вызванным воздействием стимула (Dehaene et al., 2003). Если стимул не вызывает воспламенения, и если индуцированная реакция мозга остается пространственно ограниченной и краткой, то восприятие не достигает сознания.Другими словами, стимул должен быть достаточно длинным и сильным, чтобы достичь сознания, что предполагает форму механизма фильтрации, согласующуюся с представлением о том, что только ограниченный объем информации эффективно входит в глобальное рабочее пространство. Несмотря на эти успехи в учете экспериментальных данных у людей в отношении подсознательного (бессознательного) и сознательного восприятия (King, Pescetelli, & Dehaene, 2016; Sergent & Dahaene, 2004), одним из недостатков GWT является то, что он явно не связывает крупномасштабные рекрутирование областей мозга во время сознательного доступа с клеточными механизмами.Точнее, что мешает воспламенению при коротких, не относящихся к делу раздражителях? И наоборот, что дает возможность зажигания при сильных раздражителях? Нейроанатомические, нейрофизиологические и динамические механизмы, лежащие в основе воспламенения, имеют фундаментальное значение для понимания того, как мы осознаем стимул или как изменения сетей мозга могут привести к нарушению сознания у пациентов с DOC. Поскольку GWT и DCH — это две теории, которые больше всего фокусируются на нейрофизиологических субстратах сознания и фокусируются соответственно на кортико-корковой связности и таламо-корковой связности, которые являются двумя ключевыми проблемами, подробно описанными в этом обзоре, мы обсуждаем только эти две теории. в оставшейся части этого обзора.
Если принять, что сознание связано с достаточно сложной (в алгоритмическом смысле «менее сжимаемой») обработкой информации, то возникновение сознания критически зависит от трех факторов: (1) физической сети, обеспечивающей взаимодействие между ее компонентами; (2) гибкость для временной реорганизации подсетей для достижения больших вычислительных возможностей за счет увеличения числа возможных конфигураций и функций ввода-вывода за счет функциональной связности; и (3) динамическое взаимодействие между его компонентами.Эти три критических компонента могут быть изменены, например, при поражениях после черепно-мозговой травмы. Хотя физическая крупномасштабная сеть, связывающая области мозга, хорошо определена и известна как коннектом, все еще остаются нерешенными вопросы, касающиеся временной организации кластеров, выполняющих определенные вычисления (функциональные сети), связанных средств коммуникации (нейронное кодирование) и того, как крупномасштабные функциональные сети мозга и маршрутизация информации могут быстро реконфигурироваться в зависимости от регулирования микросхемы.
В то время как основные теории сознания, представленные в этом обзоре, были сосредоточены на крупномасштабных процессах, связанных с сознанием, было приложено мало усилий, чтобы понять, как масштаб микросхемы может помочь понять, как возникает крупномасштабная скоординированная деятельность. Соединение уровней микро- и макроцепей кажется необходимым для получения целостного представления о физиологических процессах, лежащих в основе поддержания сознания (Changeux, 2017).Поэтому в этом обзоре мы формулируем гипотезу о том, что учет механизмов клеточного масштаба может обеспечить возможные направления для расширения существующих теорий сознания, охватывая несколько пространственных масштабов описания. Такое многомасштабное понимание является предпосылкой для понимания того, как сети мозга становятся дисфункциональными в DOC, и может способствовать согласованию GWT и DCH в единую структуру. Более конкретно, мы предполагаем, что совместное возникновение низкочастотных и высокочастотных нейрональных колебаний может обеспечить отдаленную коактивацию (низкочастотное, глутаматергическое происхождение), которая затем сделает возможным связывание (высокочастотное, ГАМКергическое происхождение).Это будет включать формирование функциональной сети из нескольких областей мозга посредством низкочастотного ритма, прежде чем информация будет передаваться и обрабатываться посредством гамма-активности, в частности.
Обзор организован следующим образом. Во-первых, мы исследуем взаимосвязь между двумя измерениями сознания, а именно бодрствованием и осведомленностью, с функциональной связью между корковыми областями и таламусом.Во-вторых, мы рассматриваем «средства коммуникации», обеспечивающие комплексную обработку информации, связанную с сознанием, которые регулируют кортико-корковые коммуникации, среди которых коммуникация посредством когерентности (CTC) и стробирование посредством торможения (GBI). Изменение этих механизмов представлено в клинической литературе. В-третьих, мы пытаемся связать эти результаты с недавно появившимися концепциями, основанными на взаимодействии между областями мозга, основанном на перекрестных частотных связях между колебаниями с определенными функциональными ролями.Наконец, мы предлагаем возможные клинические последствия для этой структуры с точки зрения новых протоколов нейромодуляции в DOC.
Сознательное восприятие является результатом взаимодействия двух взаимодействующих друг с другом процессов: осознания и бодрствования . Глубокий сон отключает осознанность, тогда как способность к сознательному восприятию (осознанию) стимулов окружающей среды обычно подразумевает состояние бодрствования, как показано на рисунке 1.Например, во время общей анестезии отсутствует сознательное восприятие, то есть осознание и бодрствование. Однако в некоторых особых случаях эти два компонента могут не быть связаны. Состояние безответного бодрствования (UWS) является примером DOC, в котором бодрствование присутствует без каких-либо обнаруживаемых признаков осведомленности (Laureys & Boly, 2012). Кроме того, во время осознанных сновидений существует форма осознания в отсутствие бодрствования (во время сна) (Voss, Schermelleh-Engel, Windt, Frenzel, & Hobson, 2013).Другой пример — синдром пространственного пренебрежения, при котором пациенты не осознают визуальные стимулы, находясь в состоянии бодрствования на противоположной стороне коркового поражения (Le, Stojanoski, Khan, Keough, & Niemeier, 2015). Отметим, что человеческое сознание — это многокомпонентное понятие, которое чрезвычайно сложно определить, поскольку оно включает в себя объективные состояния и субъективный опыт, включающий различные «содержания» и «уровни» сознания. Каждый автор предлагает собственное определение сознания, поскольку эти компоненты все еще обсуждаются и не всегда точно определены (Storm et al., 2017). Поскольку основная цель этой статьи — обсудить нейронные корреляты сознания (Crick & Koch, 1998), мы ограничили здесь понятие сознания базовым компонентом бодрствования , (состояние бодрствования / сна, бдительность, бдительность) и осведомленность (содержание осознанного восприятия).
Рисунок 1.
Бодрствование и осознанность — два основных измерения сознания.На этой диаграмме несколько качественно различных состояний сознания расположены на двумерной матрице в зависимости от связанных осей «содержание сознания» (осведомленность) и «уровень сознания» (бодрствование). По материалам Laureys (2005).
Рис. 1.
Бодрствование и осведомленность — два основных измерения сознания. На этой диаграмме несколько качественно различных состояний сознания расположены на двумерной матрице в зависимости от связанных осей «содержание сознания» (осведомленность) и «уровень сознания» (бодрствование).По материалам Laureys (2005).
Осведомленность поддерживается внимательными, лобно-теменными сетями, которые усиливают синаптические связи в определенных корковых путях (Tallon-Baudry, 2011). Это усиление соответствующих стимулов усиливает активированную сеть, связанную с представлением стимула. Параллельно, сопутствующее подавление нерелевантных окружающих сетей (1) оптимизирует кортико-корковую маршрутизацию информации, ограничивая возможное распространение нейронной активности по всем возможным кортикальным «маршрутам», что (2) ограничивает распространение ограниченным числом управляемых стимулами возможности, и (3) увеличивает отношение сигнал / шум.Такие механизмы связаны с концепцией функциональной связности и подробно описаны в этом обзоре. Важно отметить, что такие механизмы активного торможения, вероятно, включают корковое торможение с активной модуляцией таламо-кортикальными входами (Gabernet, Jadhav, Feldman, Carandini, & Scanziani, 2005), подразумевая, что паттерн таламо-корковой активности влияет на обработку информации в кортикальных слоях. корковые сети.
Бодрствование критически зависит от таламкортикальной связи и критических нейромодулирующих сигналов ствола мозга к таламусу (включая проекции норадреналина из голубого пятна [Monti, 2011] и проекции системы ретикулярной формации), заднего гипоталамуса и самого таламуса (Lin, 2000).Например, во время медленноволнового сна низкочастотная синхронизированная активность между корой и таламусом (так называемый ритм «вверх-вниз» [Neske, 2015]) препятствует передаче подкорковых сигналов к коре во время сна. . Это пример того, как таламо-корковая активность резко снижает обработку информации кортико-корковыми сетями, что приводит к потере сознания. Во время бодрствования таламо-корковая активность слабо синхронизирована (Gent, Bandarabadi, Herrera, & Adamantidis, 2018), что является необходимым, но не достаточным условием для включения сознания.Например, как уже упоминалось, бодрствование присутствует у пациентов с UWS, но кортико-корковая коммуникация серьезно нарушена (Noirhomme et al., 2010), что мешает регистрируемому «осознанному» компоненту сознания. Еще одно необходимое условие для сознания — это эффективная крупномасштабная кортико-корковая коммуникация, которая может поддерживать осознание за счет активации лобно-теменных сетей внимания (Luckmann, Jacobs, & Sack, 2014; Ptak, Schnider, & Fellrath, 2017). Следовательно, с точки зрения нейроанатомии, можно связать бодрствование с таламо-кортикальной, «вертикальной» связностью, в то время как осознание зависит от кортико-корковой, «горизонтальной» связи.Это согласуется с недавним взглядом Наккэша (2018) о том, что пациенты с минимальным сознанием (MCS), которые до некоторой степени сознательны, демонстрируют «кортикально опосредованные состояния», тогда как пациенты с UWS не проявляют такой активности, предположительно из-за кортико-корковой связи (критично для осознания) слишком сильно нарушена. В частности, для возникновения и поддержания сознания требуется «критическая масса» обработки информации, которая жестко регулируется таламо-корковой и кортико-корковой связями.Рисунок 1 представляет в двухмерной плоскости континуум состояний сознания как функцию осознания и бодрствования.
Одной из популярных парадигм в исследовании сознания, которая используется для исследования этих двух компонентов, является использование возможности отчетности, то есть инструктирование субъектов, когда они сознательно воспринимают стимул, что позволяет сопоставить активность мозга с / без сознательного отчета, предположительно указывая в ключевых структурах сознательного восприятия.Эта парадигма привела к ключевому пониманию того, как информация становится осознанной, что подтверждается работами, демонстрирующими, как «зажигание» происходит в мозгу, как это формализовано в GWT. Кратковременно представленный стимул приведет к кратковременной локальной активации, например, в визуальных областях, и субъект не будет сообщать о восприятии. Если стимул предъявляется в течение достаточно длительного времени, в течение длительного периода в несколько сотен миллисекунд произойдет крупномасштабная активация мозговых сетей, что приведет к глобальной доступности информации через воспламенение, и субъект сможет сообщить о восприятии.Информация, которая обрабатывается мозгом, но остается ограниченной и не передается глобально другим областям мозга, тогда называется предсознательной (а не бессознательной, что означало бы полное отсутствие обработки мозгом, связанной со стимулами). Наконец, отметим, что возможность отчетности подвергалась сомнению (Tsuchiya, Wilke, Frassle, & Lamme, 2015), поскольку указание испытуемым сообщать о своем восприятии вызывает, например, большую префронтальную активацию, которая напрямую не связана с сознанием как таковым.Следовательно, использование показателей сознания, не связанных с отчетностью, должно помочь уменьшить систематические ошибки при оценке минимальной сети областей мозга, необходимой для поддержки сознания (так называемые NCC, или нейронные корреляты сознания).
Основным недостатком статических сетевых архитектур является их ограниченность с точки зрения объема и разнообразия (сложности) обработки информации, которая может иметь место.Понятие сложности занимает центральное место в изучении сознания, поэтому необходимо подчеркнуть, что были предложены многочисленные меры сложности и что ни одно универсальное определение не является удовлетворительным. Например, сложность Лэмпеля-Зива (LZ), используемая для вычисления метрик сознания Perturbational Complexity Index (Casali et al., 2013), не совсем подходит: Perturbational Complexity Index вычисляется путем применения алгоритма LZ к ответам, вызванным TMS. ; однако полностью случайный набор вызванных TMS ответов приведет к максимальной сложности LZ, что, очевидно, нереально.Также были предложены другие меры сложности для оценки состояния сознания по сигналам ЭЭГ, такие как многомасштабная энтропия (Eagleman et al., 2018) и сложность Колмогорова (Ruffini, 2017), среди других (см. Arsiwalla & Verschure, 2018, для подробный обзор по теме). Несмотря на это концептуальное ограничение сложности LZ, полностью случайная активность, вероятно, нереальна в человеческом мозгу, поскольку всегда существует некоторая степень временной или пространственной организации нейронной активности из-за лежащих в основе микро- и макроподключений.Таким образом, в рамках этих теоретических ограничений LZ, по-видимому, приспособлен для количественной оценки сложности, связанной с наличием сознательных процессов. Обширное изучение метрик сложности, применяемых к изучению нейронных сигналов в различных состояниях сознания, несомненно, продвинет область вперед в этом отношении. Отметим, что все меры сложности не страдают ограничением максимальности для случайного сигнала, что является проблемой, подробно обсуждаемой Wang et al. (2017).Можно выделить два различных типа мер сложности: так называемые меры сложности типа 1 линейно возрастают с уровнем случайности, тогда как меры типа 2 увеличиваются со случайностью, затем достигают плато, прежде чем уменьшатся и станут нулевыми для максимальной случайности (колокол -подобная кривая). По предположению Ванга и др. (2017), измерения сложности типа 2 (например, сложность флуктуаций) могут лучше подходить для количественной оценки процессов, связанных с сознанием.
Мозг использует преимущества различных механизмов, которые преодолевают это ограничение, обеспечивая динамическую, временную реконфигурацию сетей мозга, увеличивая репертуар возможных ответов на входные данные (т.е., сложность функций ввода-вывода) (Sporns, 2013). Такие временные сети, включающие только несколько областей мозга, скоординированных для достижения определенной функции, ограниченной по продолжительности, образуют то, что называется функциональной связностью. Растет интерес к функциональным сетям, связанным с конкретными когнитивными задачами (Hassan et al., 2015), и недавно появились новые структуры (Avena-Koenigsberger, Misic, & Sporns, 2017), чтобы объяснить, как связаны анатомические связи в масштабе мозга. к функциональной связи.Функциональные сети организуются с помощью сетевых «средств коммуникации», также называемых коммуникационной динамикой, которые управляют маршрутизацией информации через определенные сети, вместо распространения информации по всей мозговой сети (Avena-Koenigsberger et al., 2017). Если бы не было таких механизмов маршрутизации, любая информация, генерируемая одной областью мозга, могла бы вызвать фазовую синхронизацию во всех анатомически связанных областях (коннектоме), что, в свою очередь, привело бы к синхронизированной активности в масштабе мозга.Такая синхронизированная деятельность в больших масштабах имеет низкую информативность и сложность (например, в случае потери сознания, вызванной судорогами), и поэтому несовместима с поддержанием сознательных процессов. Таким образом, сетевые средства коммуникации критически связаны с динамическим репертуаром нейронных состояний сети, что является ключевым, поскольку сознательные процессы связаны с богатой и разнообразной обработкой информации в крупномасштабных сетях мозга. Отметим, что таких ключевых элементов функционирования мозговых сетей, вероятно, недостаточно для возникновения сознания, а скорее они являются необходимой предпосылкой.Например, рассмотрение периферических функций тела, вероятно, важно для понимания возникновения сознания, как подчеркивается в Azzalini, Rebollo, & Tallon-Baudry (2019).
Более конкретно, один фундаментальный вопрос: какие механизмы регулируют динамику коммуникации и позволяют функциональным сетям возникать в сетях масштаба мозга? Этот вопрос является центральным для понимания того, как мозг оптимизирует свои возможности обработки информации, которые тесно связаны с сознанием.Мы предполагаем, что фундаментальные механизмы, лежащие в основе динамики коммуникации, на самом деле являются механизмами клеточного масштаба, которые (1) предотвращают нейронную синхронизацию в масштабе мозга после стимула и (2) обеспечивают временное соединение определенных отдаленных областей мозга. Большой интерес вызывают крупномасштабные паттерны активности мозга, связанные с сознанием, поскольку их можно измерить с помощью различных методов нейровизуализации (например, электроэнцефалографии, функциональной магнитно-резонансной томографии).Тем не менее, механизмы на клеточном уровне по-прежнему более сложны для изучения у людей по очевидным причинам инвазивности, связанной с необходимыми методами записи у людей. В этом разделе мы рассмотрим доказательства таких механизмов, которые могут соединить уровни микросхемы и масштаба мозга.
С точки зрения крупномасштабных нейроанатомических путей, обеспечивающих сознание, глутаматергические проекции на большие расстояния между пирамидными нейронами через волокна белого вещества, вероятно, имеют решающую роль (Dehaene & Changeux, 2011), поскольку они обеспечивают быструю (из-за миелина) связь между удаленными регионами.На уровне мозга эти волокна белого вещества, вероятно, имеют решающее значение для обеспечения сознательного доступа, который включает в себя временную стабилизацию нейронной активности, кодирующей определенный информационный паттерн, в сети высокоуровневых областей мозга, связанных между собой дальнодействующими связями. префронтальная кора, действующая как ключевой узел (Berkovitch, Dehaene, & Gaillard, 2017; Dehaene, Changeux, Naccache, Sackur, & Sergent, 2006). На сознательных испытаниях распределенная активность в гамма-диапазоне отражает стабилизацию локальной информации, транслируемой в другие области.Во время сознательного доступа к определенной информации другие нейроны окружающего глобального рабочего пространства будут заблокированы и недоступны для обработки других стимулов, поэтому остаются предсознательными (не достигая сознания). В дополнение к характеристикам связности в масштабе микросхемы, другим ключевым фактором в формировании нейронных колебаний является наличие временных задержек, в частности, из-за конечной скорости проводимости потенциалов действия по волокнам. Задержки по времени могут привести к возникновению и стабильности колебаний на определенных частотах (Petkoski & Jirsa, 2019; Petkoski, Palva, & Jirsa, 2018).Следовательно, повреждение кортико-кортикальных волокон не только ухудшает передачу активности между областями, но также может изменить частотный состав и стабильность нервных колебаний, тем самым влияя на их функцию.
В локальном масштабе микросхема также была идентифицирована как участвующая в коммуникации между удаленными областями мозга: проекция пирамидных нейронов в одной области мозга на VIP-положительные (вазоактивный кишечный пептид) нейроны в другой области.Активируя VIP-положительные нейроны в отдаленной области, это вызывает ингибирование соматостатин-положительных (SST) нейронов, которые нацелены на дендриты пирамидных клеток, что приводит к дисинаптическому растормаживанию (Karnani et al., 2016). Благодаря этому дисинаптическому растормаживанию генерация гамма-активности может происходить посредством взаимного ингибирования парвальбумин-парвальбумин (PV-PV), и может иметь место связывание между двумя вовлеченными областями (Munoz, Tremblay, Levenstein, & Rudy, 2017), временно обеспечивая передачу информации и обработка.Эти механизмы в клеточном масштабе представлены на Рисунке 2.
Рисунок 2.
Базовые (схематические) схемы, участвующие в генерации локальных и далеких колебаний. В локальной корковой сети (кортикальном столбе), щелевом соединении, взаимное ингибирование «быстрых» ГАМКергических интернейронов является одним из основных механизмов генерации локальной гамма-активности, наряду с ПИНГ (пирамида-интернейрон гамма).Напротив, петля обратной связи между проецирующими дендриты «медленными» ГАМКергическими интернейронами и пирамидными клетками может генерировать низкочастотную активность. Важно отметить, что дистанционное общение посредством дисинаптического пути позволяет кратковременно генерировать гамма-колебания в отдаленных популяциях. Пирамидные клетки в исходной популяции (левая цепь) проецируются на пирамидные клетки отдаленной популяции (справа), но также и на интернейроны VIP, которые проецируются на нейроны SST, проецирующие дендриты. Временная активация VIP-нейронов из исходной популяции временно подавляет SST-нейроны в целевой популяции, позволяя генерировать гамма-колебания через PV-PV и пирамидно-PV-цепь.Как только вход на удаленные VIP-нейроны уменьшается, SST-нейроны возобновляют свой тормозной вход, который может прекратить генерацию гамма-колебаний.
Рисунок 2.
Базовые (схематические) схемы, участвующие в генерации локальных и далеких колебаний. В локальной корковой сети (кортикальном столбе), щелевом соединении, взаимное ингибирование «быстрых» ГАМКергических интернейронов является одним из основных механизмов генерации локальной гамма-активности, наряду с ПИНГ (пирамида-интернейрон гамма).Напротив, петля обратной связи между проецирующими дендриты «медленными» ГАМКергическими интернейронами и пирамидными клетками может генерировать низкочастотную активность. Важно отметить, что дистанционное общение посредством дисинаптического пути позволяет кратковременно генерировать гамма-колебания в отдаленных популяциях. Пирамидные клетки в исходной популяции (левая цепь) проецируются на пирамидные клетки отдаленной популяции (справа), но также и на интернейроны VIP, которые проецируются на нейроны SST, проецирующие дендриты. Временная активация VIP-нейронов из исходной популяции временно подавляет SST-нейроны в целевой популяции, позволяя генерировать гамма-колебания через PV-PV и пирамидно-PV-цепь.Как только вход на удаленные VIP-нейроны уменьшается, SST-нейроны возобновляют свой тормозной вход, который может прекратить генерацию гамма-колебаний.
В нескольких исследованиях изучается, как черепно-мозговая травма (ЧМТ) влияет на определенные типы клеток и как это может повлиять на крупномасштабную обработку информации в головном мозге. В моделях ЧМТ на животных гипервозбудимость, индуцированная в долгосрочной перспективе (8–10 недель после лечения) (Allitt, Iva, Yan, & Rajan, 2016; Alwis et al., 2016; Alwis, Yan, Morganti-Kossmann, & Rajan, 2012) была связана с избирательным сокращением интернейронов, нацеленных на дендриты, в этой коре головного мозга (Carron, Yan, Allitt, & Rajan, 2018). Следовательно, можно предположить, что TBI-индуцированная потеря функциональных нацеленных на дендриты интернейронов будет, следовательно, влиять как на генерацию низкочастотных колебаний, так и на растормаживание через VIP интернейроны. Более того, на животных моделях также была показана прогрессирующая потеря фазового ингибирования с соответствующей потерей соматических нацеленных на PV + ГАМКергических интернейронов после ЧМТ (Cantu et al., 2015; Guerriero, Giza, & Rotenberg, 2015), которые могут быть, по крайней мере, частично ответственны за нарушение гамма-колебаний и синхронность (C. Wang et al., 2017). Потеря интернейронов PV + была обнаружена не только в моделях ЧМТ на грызунах, но и в иммунохимическом исследовании, проведенном в посмертных образцах человеческого мозга (Schiavone, Neri, Trabace, & Turillazzi, 2017). В целом, помимо вызванного ЧМТ повреждения ГАМКергических интернейронов, участвующих в генерации медленных и быстрых колебаний, наличие аномальных возбуждающих синапсов и повреждение аксонов также может участвовать в изменении нейронной связи (Harris et al., 2016).
Рассматривая эти механизмы клеточного масштаба, одним из важнейших вопросов является понимание того, как они участвуют в кратковременном возникновении и поддержании функциональных сетей, таких как поддерживающие сознание. Установлено, что состояние сознания критически зависит от функциональной кортико-корковой связи мозга (Marino, Bonanno, & Giorgio, 2016; Naro et al., 2018). Напомним, функциональная связность относится к статистически значимым связям между временными курсами нейронной активности в разных регионах, тогда как анатомическая связь обозначает физические связи между областями мозга.Таким образом, функциональные сети могут отражать косвенные связи между областями мозга и быть временными в зависимости от того, какие задачи выполняются или какие стимулы воспринимаются. Было показано, что даже в отсутствие какой-либо конкретной задачи или стимулов сети в состоянии покоя (например, сеть режима по умолчанию) также являются временными (Kabbara, El Falou, Khalil, Wendling, & Hassan, 2017). Экспериментальные данные подтверждают идею о том, что функциональная связь может пролить свет на сети, участвующие в различных состояниях сознания (Jin & Chung, 2012).Например, во время потери сознания, вызванной общей анестезией, происходит нарушение кортико-корковой функциональной связи (Ferrarelli et al., 2010; Gomez et al., 2013; Hudetz, 2012), что серьезно ухудшает способность корковых сетей к интегрировать информацию и сделать ее доступной в большом масштабе, как требуется для сознательного восприятия в IIT или GWT. Точно так же при переходе от бодрствования к медленноволновому сну скорость возбуждения в коре остается относительно неизменной во время деполяризующих фаз медленных колебаний сна (Steriade, Timofeev, & Grenier, 2001), в то время как эффективная связь мозга (эффективная связь определяется как быстрое причинное взаимодействие между областями мозга; Rosanova et al., 2012) резко изменилась (Esser, Hill, & Tononi, 2009; Tononi & Sporns, 2003). После засыпания без быстрого сна корковые активации также становятся более локальными и стереотипными, нарушая эффективную корковую связь (Massimini et al., 2010), как показано с помощью вызванных TMS ответов ЭЭГ, которые остаются очень близкими к месту стимуляции, в то время как эти ответы вовлекают сеть отдаленных областей мозга, претерпевающих сложные динамические паттерны последовательной активации во время бодрствования (Casali et al., 2013; Casarotto et al., 2016; Guillery & Sherman, 2002). Эти результаты также подчеркивают решающую роль таламо-кортикального пути в кортико-корковой функциональной связности. Стоит отметить, что существенная роль таламо-кортикальной петли, а также так называемые «реентерабельные взаимодействия» ранее считались ключевыми в DCH (Tononi & Edelman, 1998).
В соответствии с этими результатами, полученными во время сна, это нарушение кортико-корковой связи также наблюдалось во время общей анестезии и у пациентов с DOC и даже исследовалось с помощью компьютерного моделирования (Casali et al., 2013; Esser et al., 2009). В рамках GWT это объясняет, почему сознание нарушается в таких состояниях: крупномасштабная связь между удаленными областями мозга нарушается из-за таламо-кортикальной модуляции, предотвращающей возникновение воспламенения. У пациентов с DOC с повреждением головного мозга крупномасштабная кортико-корковая коммуникация может быть нарушена из-за частичного разрушения волокон дальнего действия, что физически затрудняет синхронизацию мозга на больших расстояниях. Что касается эффектов на клеточном уровне, разрушение волокон дальнего действия может препятствовать синхронизации удаленных VIP-интернейронов, что имеет решающее значение для индукции дисинаптического растормаживания и связанной с ним гамма-активности, необходимой для CTC.
Патологические изменения функциональной связности были исследованы с использованием различных методов: (1) функциональная связность во время «состояния покоя» с использованием фМРТ или ЭЭГ, (2) импульсная стимуляция с использованием транскраниальной магнитной стимуляции (ТМС) во время записи ЭЭГ и (3) другие подходы, основанные на пертурбации, исследующие реакцию мозга на сенсорные стимулы (Boly et al., 2017). Преимущество функциональной связи заключается в том, что ее можно использовать для улучшения оценки и классификации DOC (Holler et al., 2014; Наро и др., 2018; Росси Себастьяно и др., 2015; Сандерс, Тонони, Лорис и Сани, 2012). Например, у пациентов с легкими когнитивными нарушениями было показано, что нарушение сознания связано с измененной эффективной связью (Crone et al., 2015; Varotto et al., 2014). Отказ крупномасштабного подключения, наряду с гиперсинхронией локальной ближней дельта- и альфа-активности, были обнаружены в DMN и коррелировали с уровнем осведомленности у пациентов с DOC (Fingelkurts, Fingelkurts, Bagnato, Boccagni, & Galardi, 2013 ; Каббара и др., 2017; Маки-Марттунен, Диез, Кортес, Кьялво и Вильярреал, 2013 г .; Наро и др., 2018; Vanhaudenhuyse et al., 2010; Varotto et al., 2014). Кроме того, паттерн функциональной связности нескольких областей мозга, таких как задняя поясная извилина коры и предклинье, может даже предсказывать состояние улучшения сознания пациентов с UWS с точностью, превышающей 80% (Wu et al., 2015). Отметим, что фМРТ внесла значительный вклад в определение изменений функциональной связности в различных сетях состояния покоя (RSN) в различных состояниях сознания (для обзора см. Heine et al., 2012. Более конкретно, с точки зрения фМРТ в состоянии покоя, Demertzi et al. (2014) использовали машинное обучение для автоматического извлечения RSN и определения их нейронального происхождения (в отличие от артефактов). Результаты показали, что для большинства RSN количество компонентов нейронального происхождения снижается с увеличением уровня сознания. В другом исследовании (Demertzi et al., 2015), также основанном на использовании фМРТ в состоянии покоя, авторы указали на снижение функциональной связи между различными RSN (например, DMN, визуальными и т. Д.)), что хорошо коррелировало с оценкой CRS-R. Другое недавнее исследование фМРТ (Demertzi et al., 2019) продемонстрировало, что динамика функциональных сетей фМРТ может различать различные состояния сознания (контроль, MCS, UWS). В частности, вероятность присутствия различных модульных состояний с течением времени вместе с их временем пребывания была надежным индикатором сознания. Сложность карты переходов между различными модульными состояниями также уменьшалась с увеличением уровня сознания.Тот факт, что исследования ЭЭГ и фМРТ сходятся, хотя и в разных временных масштабах, в снижении сложности динамических реакций, дополнительно подчеркивает фундаментальную взаимосвязь между уровнем сознания и очевидной сложностью динамики мозга. Тесная взаимосвязь между BOLD-сигналом, который имеет низкую частоту (<0,5 Гц, также называется сверхмедленными колебаниями), и низкочастотной составляющей локальных потенциалов поля (LFP) и электрокортикографических (ECOG) сигналов (He, Snyder, Zempel, Smyth, & Raichle, 2008), может обеспечить связь между исследованиями фМРТ и ЭЭГ в области исследования сознания (обзор см. В He & Raichle, 2009).Действительно, было высказано предположение, что сверхмедленные колебания в BOLD, которые связаны с низкочастотной областью записей LFP / ECOG (для тщательного исследования корреляции между BOLD-сигналом и электрофизиологической активностью в разных частотных диапазонах см. Scheeringa, Koopmans, van Mourik, Jensen, & Norris, 2016) вносят ключевой вклад в интеграцию информации между удаленными областями коры головного мозга. Интересно, что это предложение соответствует нашему предложению о функциональной роли низкочастотных колебаний в интеграции информации, в то время как более высокочастотные колебания будут участвовать в связывании.
Помимо «пассивного» исследования функциональной связности в состоянии покоя, использование ответов ЭЭГ, вызванных TMS, позволяет проводить активное «зондирование» эффективной связи. Например, резкое нарушение функциональной связи было выявлено у пациентов с UWS при использовании определенного протокола TMS, вызывающего у этих пациентов стереотипный местный ответ ЭЭГ, аналогичный спящим без сознания или анестезированным субъектам (Rosanova et al., 2012). Восстановление крупномасштабной эффективной связи коры головного мозга с транскраниальной стимуляцией мозга, такой как транскраниальная стимуляция переменным током (tACS), в DOCs, следовательно, может быть полезным подходом для облегчения частичного восстановления за счет усиления колебаний и пластичности. Одним из клинических результатов, подтверждающих эту идею, является недавняя демонстрация того, что DLPFC (дорсолатеральная префронтальная кора) -tACS смогла временно восстановить нарушение связи у людей с DOC (Naro, Bramanti, Leo, Russo, & Calabro, 2016).
В целом, мы утверждаем, что для обеспечения оптимального взаимодействия требуется тонкий баланс фазовой синхронизации между областями мозга. Фазовая синхронизация может происходить через прямые (или косвенные, через общий вход) связи между двумя регионами, когда какая-то форма активности в одном меняет активность в другом. Такое увеличение фазовой синхронизации лежит в основе так называемой функциональной связности. Очевидно, что при низкой фазовой синхронизации (функциональной связности) эффективная передача информации между регионами невозможна.С другой стороны, чрезмерная синхронизация приводит к потере сложности из-за того, что активность становится слишком похожей в задействованных регионах. Ярким примером является сложный эпилептический припадок, при котором большое количество структур мозга тесно функционально связаны и имеют сходный судорожный ритм и при котором происходит потеря сознания (Blumenfeld & Taylor, 2003). Поэтому мы утверждаем, что существует «золотая середина» синхронизации на определенных частотах, которая позволяет надлежащим образом поддерживать обработку информации, связанной с сознанием, сетью областей мозга.
GBI (Jensen & Mazaheri, 2010) — один фундаментальный механизм микроскопического масштаба, участвующий в маршрутизации информации в головном мозге и способствующий формированию функциональных сетей внутри анатомической сети. GBI включает в себя тормозные процессы, приводящие к избирательной активации подсетей и инактивации других подсетей. Предотвращая активацию в масштабе мозга в ответ на стимул и ограничивая количество областей мозга, участвующих в выполнении задач, GBI также предотвращает состояния низкой сложности (например,g., все области мозга демонстрируют одинаковую активность) и, следовательно, неэффективная обработка информации. Таким образом, процессы GBI предполагают, что роль торможения более сложна, чем предотвращение чрезмерной активации сетей мозга, вместо этого способствуя преобразованию анатомических сетей мозга в функциональные сети (Avena-Koenigsberger et al., 2017). О возможных изменениях GBI сообщалось в исследованиях, показывающих, что альфа-мощность ЭЭГ ниже у пациентов с UWS, чем у пациентов с MCS (Lehembre et al., 2012; Stefan et al., 2018), намекая на то, что нейробиологические механизмы, лежащие в основе генерации альфа-осцилляций и связанного с ними GBI, глубоко изменены у неотзывчивых пациентов. Более того, альфа-активность была высоко синхронизирована и сгруппирована в центральных и задних областях коры у пациентов с UWS (Lehembre et al., 2012; Stefan et al., 2018), что указывает на возможную неэффективность GBI при наиболее тяжелых DOC. Действительно, во время общей анестезии пропофол и кетамин изменяют фронтальные альфа-колебания (Feshchenko, Veselis, & Reinsel, 2004; Purdon et al., 2013), предполагая, что снижение лобно-теменной связи в альфа-диапазоне может играть важную роль в потере сознания (Blain-Moraes, Lee, Ku, Noh, & Mashour, 2014). Когерентные медленные колебания, такие как альфа (8–12 Гц) и дельта (1–4 Гц) ритмы, появляются в медиальной префронтальной коре и таламусе во время потери сознания, вызванной пропофолом, и, как полагают, вносят свой вклад в их функциональное нарушение. площадей (Flores et al., 2017; Lewis et al., 2018). В частности, дельта-колебания, вызванные анестезией пропофолом у людей, по-видимому, нарушают крупномасштабную коммуникацию между областями мозга, сохраняя при этом активность локальных нервных сборок (Lewis et al., 2018). Недавнее исследование (Todorova & Zugaro, 2019) пришло к аналогичному выводу относительно функциональной роли дельта-колебаний во время сна, которые изолируют небольшие сети в коре головного мозга от остальной части мозга (тем самым нарушая интеграцию), что, по-видимому, играет определенную роль. в усилении памяти во время сна.
Другой ключевой установленный процесс, с помощью которого отдаленные области мозга участвуют вместе в паттерне активности, связанной с выполнением данной задачи, — это CTC (Bonnefond, Kastner, & Jensen, 2017; Deco & Kringelbach, 2016; Fries, 2005, 2009, 2015) .CTC действительно включает в себя фазовую гамма-активность между удаленными областями мозга, что позволяет обрабатывать информацию. Было высказано предположение, что ЦКО является субстратом «связывания», то есть слияния различных характеристик стимула в единое, объединенное сознательное восприятие (Singer, 2001). Точнее, флуктуация возбудимости в группе нейронов обеспечивает особую сигнатуру, характеризующуюся определенной полосой частот и характером разряда (Womelsdorf, Valiante, Sahin, Miller, & Tiesinga, 2014), распространяясь через крупномасштабную сеть, состоящую из анатомически взаимосвязанные области мозга и впоследствии запускающие активность в связанных областях.Обработка информации в мозге тесно связана с синхронизированными по фазе, скоординированными во времени флуктуациями возбудимости (Fries, 2005) в сетях распределенных популяций нейронов. Результирующие колебания генерируют определенный нейронный код, а когерентность позволяет связывать информацию и общение. Кроме того, ЦОК включает гамма-активность, генерируемую в основном ГАМКергическими интернейронами (PV-положительные корзиночные клетки). В совокупности тормозящие процессы оказываются ключевыми для маршрутизации и обработки информации в сетях масштаба мозга, задействованных в сознании: GBI пространственно формирует мозговые сети (какие области мозга задействованы, а какие заблокированы), а CTC контролирует их во времени (поток информации).Однако это поднимает интригующий вопрос: если гамма-ритмы генерируются локально межнейронными ГАМКергическими сетями, как могут удаленные области мозга, связанные через глутаматергические волокна дальнего действия, эффективно общаться и достигать ЦОК? Наша гипотеза может дать возможное объяснение этому парадоксу, если предположить, что удаленные области мозга «блокируют» свою активность посредством низкочастотных колебаний (что было бы предпосылкой), а затем позволяют связывание посредством высокочастотных колебаний (гамма, через CTC).
Подтверждая эту возможность, сознательное восприятие действительно характеризуется увеличением активности распределенного гамма-диапазона (Melloni et al., 2007; Wyart & Tallon-Baudry, 2009). Интересно, что эти быстрые колебания модулируются медленными колебаниями (Jensen, Gips, Bergmann, & Bonnefond, 2014; Osipova, Hermes, & Jensen, 2008). Недавно было предложено, что фазовая синхронизация низкочастотных колебаний, играющая роль временной системы отсчета для информации, несущей высокочастотную активность, является общим механизмом мозговой коммуникации (Bonnefond et al., 2017). Эти вложенные колебания могут быть ключевым механизмом не только для кортико-корковой коммуникации и обработки, но также между подкорковыми структурами. Эмоциональная память актуальна для рассмотрения для нашей цели, поскольку эмоциональная память включает как корковые, так и подкорковые структуры и задействует синхронизацию больших сетей через тета-гамма-колебания (Bocchio, Nabavi, & Capogna, 2017). Во время экспериментов in vivo, проведенных на грызунах, воспринимаемая угроза (стимул, объявляющий удар ногой) усиливает тета-силу и согласованность в миндалине, префронтальной коре и гиппокампе (Lesting et al., 2011; Likhtik, Stujenske, Topiwala, Harris, & Gordon, 2014), в то время как быстрые гамма-всплески синхронизированы по фазе с тета-колебаниями (Stujenske, Likhtik, Topiwala, & Gordon, 2014). В целом, эти результаты подтверждают идею о том, что вложенные колебания на частотах тета и гамма являются правдоподобным субстратом для открытия / маршрутизации информационных каналов (тета) и обработки / передачи (гамма) в головном мозге.
Внимание — еще один ключевой элемент для сознательной обработки, и оно участвует в синхронизации удаленных областей мозга (Niebur, Hsiao, & Johnson, 2002; Steinmetz et al., 2000). Основными ритмами мозга, участвующими в процессах внимания, являются альфа- и гамма-колебания: области мозга синхронизируют гамма-колебания (Womelsdorf et al., 2014) и модулируются медленными альфа-колебаниями. Медленные колебания позволяют блокировать нерелевантные сети и влияют на локальную обработку сигналов, широкий обмен информацией и восприятие (Sadaghiani & Kleinschmidt, 2016). Информационный поток устанавливается нейрональной синхронизацией в нижних частотных диапазонах, а именно в тета (4–7 Гц), альфа (8–13 Гц) и бета (14–25 Гц) диапазонах (Bonnefond et al., 2017). Одна из возможных причин заключается в том, что низкочастотная активность вызывает кратковременное изменение возбудимости в целевых структурах мозга, что обеспечивает оптимальное окно для связывания нейронных сигналов из разных регионов посредством высокочастотной активности (например, гамма-излучения) (Canolty et al., 2006) . Это дополнительно подтверждает идею о том, что низкочастотные нейронные колебания в основном участвуют в установлении кратковременной связи на большие расстояния через глутаматергические проекции, тогда как высокочастотные нейронные колебания участвуют в обработке / передаче информации.Мы предполагаем, что можно связать понятие «интеграция» с этой глутаматергической коактивацией дальнего действия, обеспечивающей связь в масштабе мозга между областями мозга; тогда как «дифференциация / сегрегация» будет зависеть от локально генерируемой гамма-активности (и частично от уровня низкочастотной активации, что приведет к массово синхронизированной активности и уменьшению дифференциации и сложности). Обзор вышеупомянутых механизмов представлен на Рисунке 3.
Рисунок 3.
Предлагаемый синтез того, как временное избирательное связывание между корковыми сетями происходит через механизмы клеточного масштаба. (A) Схематическая диаграмма анатомической сети с основными проекциями между областями в масштабе мозга. (B) На верхней панели селективное связывание (коммуникация через когерентность; Fries, 2015) в подмножестве областей коры происходит посредством генерации гамма-колебаний (в основном через микросхемы с участием корзинчатых клеток), в то время как удаленное растормаживание определенных областей мозга происходит через дисинаптические растормаживание (активация удаленных VIP-нейронов ингибирует SST-нейроны, которые, в свою очередь, уменьшают их тормозную проекцию на пирамидные нейроны).Это способствует преобразованию анатомической сети в функциональную сеть. Альфа-ритм действует как импульсное торможение, подавляя «нерелевантные» сети, дополнительно увеличивая отношение сигнал / шум (SNR). На нижней панели снижение интеграции (например, после повреждения головного мозга) приводит к нарушению синхронизации удаленных областей мозга (что отражается снижением низкочастотного ритма на иллюстративном колебании), и, таким образом, уменьшение связывания, что в совокупности приводит к снижению GBI. (Jensen & Mazaheri, 2010) и сильно снижает общее SNR, что приводит к дисфункции сети с точки зрения интеграции и связывания, необходимых для сознания.Отметим, что помимо уменьшения амплитуды низкочастотного ритма, соотношение фаз вложенного гамма-колебания могло быть нарушено (т. Е. Более случайным) по сравнению с физиологическим случаем. Такую потенциальную взаимосвязь вложенных тета / гамма-колебаний еще предстоит изучить в DOC.
Рисунок 3.
Предлагаемый синтез того, как временное избирательное связывание между корковыми сетями происходит через механизмы клеточного масштаба. (A) Схематическая диаграмма анатомической сети с основными проекциями между областями в масштабе мозга.(B) На верхней панели селективное связывание (коммуникация через когерентность; Fries, 2015) в подмножестве областей коры происходит посредством генерации гамма-колебаний (в основном через микросхемы с участием корзинчатых клеток), в то время как удаленное растормаживание определенных областей мозга происходит через дисинаптические растормаживание (активация удаленных VIP-нейронов ингибирует SST-нейроны, которые, в свою очередь, уменьшают их тормозную проекцию на пирамидные нейроны). Это способствует преобразованию анатомической сети в функциональную сеть.Альфа-ритм действует как импульсное торможение, подавляя «нерелевантные» сети, дополнительно увеличивая отношение сигнал / шум (SNR). На нижней панели снижение интеграции (например, после повреждения головного мозга) приводит к нарушению синхронизации удаленных областей мозга (что отражается снижением низкочастотного ритма на иллюстративном колебании), и, таким образом, уменьшение связывания, что в совокупности приводит к снижению GBI. (Jensen & Mazaheri, 2010) и сильно снижает общее SNR, что приводит к дисфункции сети с точки зрения интеграции и связывания, необходимых для сознания.Отметим, что помимо уменьшения амплитуды низкочастотного ритма, соотношение фаз вложенного гамма-колебания могло быть нарушено (т. Е. Более случайным) по сравнению с физиологическим случаем. Такую потенциальную взаимосвязь вложенных тета / гамма-колебаний еще предстоит изучить в DOC.
Новые данные показывают, что баланс обработки локальной и глобальной информации может быть нарушен при неврологических расстройствах.Как правило, недавнее исследование функциональных сетей у пациентов с болезнью Альцгеймера выявило снижение интеграции мозга, количественно определяемое коэффициентом участия (отражает связь между удаленными модулями мозга), в то время как сегрегация, определяемая количественно коэффициентом кластеризации (отражает локальную связь между отдельными областями мозга) был увеличен (Kabbara et al., 2018). Это согласуется с нейродегенеративными процессами, которые, вероятно, влияют на «блокировку» определенных областей мозга или ингибирование нерелевантных сетей, тем самым серьезно нарушая крупномасштабную интеграцию информации.В контексте DOC недавние клинические данные (Chennu et al., 2017; Rizkallah et al., 2019) подтверждают эту точку зрения. В исследовании Chennu et al. Сети на уровне кожи головы были оценены у пациентов с DOC и указали на снижение интеграции в альфа-диапазоне. Более конкретно, лобно-теменная сеть в альфа-диапазоне была дискриминационной между пациентами с MCS и UWS. В недавнем исследовании Rizkallah et al. (2019) количественно оценили уровень локальной и глобальной обработки информации в частотно-зависимых функциональных сетях (уровень источника) у пациентов с DOC и контрольной группы.Интеграция в функциональные сети тета-диапазона снижалась с увеличением уровня сознания, и между контрольной группой и любой группой пациентов систематически были задействованы две анатомические области: часть левой орбитофронтальной коры и левое предклинье. Одна из возможностей состоит в том, что физическое повреждение длинных волокон белого вещества нарушает крупномасштабную интеграцию и баланс обработки локальной / глобальной информации. Одним из возможных подходов к изучению такого анатомического повреждения волокон белого вещества является диффузионно-тензорная визуализация, выполняемая у пациентов с тяжелой травмой головного мозга DOC (Fernandez-Espejo et al., 2011; Galanaud et al., 2012; Luyt et al., 2012), которые выявили широко распространенные нарушения белого вещества. Более низкая фракционная анизотропия действительно была обнаружена в трактах подкорковых и кортикальных волокон у пациентов с DOC по сравнению с контрольной группой (Lant, Gonzalez-Lara, Owen, & Fernandez-Espejo, 2016; Weng et al., 2017), что позволяет предположить, что основной дефицит сознания у пациентов с DOC может быть связан с измененными связями белого вещества между базальными ганглиями, таламусом и лобной корой.Это также согласуется с эффектом поражения миелинизированных волоконных трактов, что может привести к нарушению связи между удаленными областями мозга (Adams, Graham, & Jennett, 2000). Следовательно, кажется разумным, что поражения белого вещества могут изменять, модифицировать или предотвращать как CTC, так и GBI между крупномасштабными сетями. Кроме того, мы предполагаем, что если конкретная фазовая синхронизация гамма-колебаний с тета-колебаниями будет нарушена, то могут появиться клинические проявления, связанные с DOC (потеря интеграции и снижение уровня сознания).
Позвольте нам также упомянуть, что, хотя и не связаны напрямую, другие патологии могут влиять на патофизиологию DOC, поскольку они имеют один важный компонент: изменения в функциональных сетях. Когда эти изменения влияют на ключевые структуры / сети мозга, сознание может измениться. Следовательно, понимание того, как нарушение регуляции функциональных сетей при неврологических расстройствах не связано с ухудшением уровня сознания (например,g., пациенты с легкими когнитивными нарушениями) все еще могут быть информативными с точки зрения фундаментальных механизмов, лежащих в основе DOC. В том же духе, поскольку функциональная связность зависит от уровня фазовой синхронизации между регионами, очень важно подчеркнуть, что повышение уровня фазовой синхронизации между структурами, участвующими в конкретной задаче (например, лежащей в основе фронто-временной сети рабочая память) также улучшает поведенческую производительность этой задачи. Следовательно, литература по tACS может предоставить новые направления относительно того, как наиболее критически регулировать функциональные сети в сознательных процессах, например, путем увеличения фазовой синхронизации ключевых структур в определенных частотных диапазонах.Этот момент обсуждается в следующем разделе.
В этом обзоре мы попытались согласовать нейроанатомические и нейрофизиологические знания на уровне микро- и макроскопических сетей в отношении процессов, лежащих в основе возникновения и поддержания сознания и его изменений у пациентов с DOC. Мультиплексирование нейронных ритмов через вложенные колебания представляется вероятным механизмом коактивации в сети определенных удаленных областей мозга (интеграция), что является предпосылкой для осознанного восприятия.Кроме того, ключевым механизмом, по-видимому, является тонкий баланс между низкочастотной активностью (связанной с «глобальной» обработкой) и высокочастотной активностью (связанной с более «локальной» обработкой), что может обеспечить динамику нейронов, лежащую в основе оптимальной маршрутизации информации. и обработка. Чрезмерная низкочастотная активность (например, дельта-активность) приводит к массово синхронизированной активности, что приводит к потере сложности с точки зрения обработки информации, параллельно в таких случаях с потерей сознания (например,г., сон, судороги). Что касается дельта-диапазона, этот ритм не связан с возникновением или поддержанием сознательных процессов. Это подтверждается тем фактом, что дельта-активность является наиболее преобладающей при UWS (Sitt et al., 2014), а также во время глубокого сна, где в обоих случаях кортико-корковая коммуникация резко снижена. UWS и глубокий сон действительно являются примерами психических состояний, в которых таламо-корковая активность и синхронизация высоки («вертикальная» связь), в то время как кортико-кортикальная («горизонтальная») синхронизация снижается.Поскольку дельта-ритм имеет таламо-кортикальное происхождение, и поскольку таламус так широко связан со всей корой, дельта-активность связана с высоким уровнем синхронизации всей коры, что снижает интеграцию и сложность и препятствует сознательному доступу. . Кроме того, дельта-активность, по-видимому, модулирует в основном «бодрствующее» (таламо-кортикальное) измерение сознания, тогда как более быстрые ритмы, по-видимому, модулируют «осознанное» (кортико-корковое) измерение сознания.
Аналогичным образом, отсутствие лобно-теменной функциональной связи (сети внимания) было обнаружено в недавнем исследовании, что было количественно определено с помощью ЭЭГ высокого разрешения у пациентов с DOC (Chennu et al., 2017), что свидетельствует о достаточном уровне лобно-теменной связи. теменная связь необходима для достижения достаточной нейрональной интеграции и воспламенения для сознательного восприятия. В более общем плане динамика мозга у пациентов с DOC обычно характеризуется потерей интеграции в крупном масштабе (Chennu et al., 2017; Rizkallah et al., 2019), предотвращая эффективную крупномасштабную координацию удаленных областей мозга для достижения сознательного восприятия. Это говорит о том, что низкочастотный ритм, необходимый для корковой коммуникации на больших расстояниях, снижен, предотвращая связывание в гамма-диапазоне и, следовательно, дальнейшую обработку информации и воспламенение для доступа сознания. При этом, каковы возможные последствия этого баланса медленной / быстрой активности в качестве механизма-кандидата для сложной обработки, связанной с сознанием?
Интересная перспектива, которая также могла бы быть формой подтверждения этого механизма, — это использование методов нейромодуляции у пациентов с DOC для повышения их уровня сознания.Целью таких методов нейромодуляции может быть «ребалансировка» локальной и глобальной обработки, например, за счет использования транскраниальной прямой или попеременной стимуляции (tDCS / tACS применяется как к лобному, так и к теменному участку одновременно, чтобы увеличить низкочастотную стимуляцию). синхронизация в тета-диапазоне. В настоящее время считается, что tACS может модулировать эндогенные ритмы мозга с помощью электрических полей относительно низкого уровня (<1 В / м, как обсуждалось в Modolo, Denoyer, Wendling, & Benquet, 2018), что могло бы предполагают использование частоты стимуляции в тета-диапазоне для увеличения остаточных колебаний в этом частотном диапазоне (т.е., предполагая, что остаточные анатомические связи все еще присутствуют). Двухсайтовый лобно-теменной tACS в тета-диапазоне может тогда предоставить неинвазивную возможность повышения уровня сознания у пациентов с DOC, в ожидании, что некоторая остаточная анатомическая связь остается в случае пациентов с повреждением головного мозга. Интересно, что недавнее исследование (Violante et al., 2017) использовало двухсайтовый tACS в тета-диапазоне у здоровых добровольцев и сообщило об улучшении рабочей памяти, функции, также зависящей от лобно-теменной сети.Другое исследование с использованием tACS, нацеленного на лобно-височную сеть, сообщило об увеличении производительности рабочей памяти у пожилых людей по сравнению с сопоставимыми уровнями у молодых участников (Reinhart & Nguyen, 2019). Эти недавние результаты, полученные на людях, предоставляют убедительные доказательства того, что изменение баланса обработки информации с помощью протоколов нейромодуляции может способствовать повышению уровня сознания у некоторых пациентов с DOC (например, когда повреждение длинных волокон белого вещества не слишком серьезно). Интересно, что в нескольких исследованиях tDCS с разной степенью успеха предпринимались попытки улучшить уровень сознания у пациентов с DOC.Например, рандомизированное клиническое исследование пациентов с MCS (всего 40 пациентов), направленное на применение tDCS над левой префронтальной дорсолатеральной корой, показало умеренное улучшение уровня сознания после 4-недельного протокола (Martens et al., 2018). Это подтвердило более ранние исследования, демонстрирующие положительное влияние повторных tDCS (хотя и в течение короткого времени) на уровень сознания у пациентов с DOC (Thibaut, Bruno, Ledoux, Demertzi, & Laureys, 2014; Thibaut et al., 2017). Однако один сеанс tDCS над моторной корой не привел к улучшению (Martens et al., 2019), предполагая, что повторяющиеся сеансы tDCS могут быть ключевыми. Отметим, что, несмотря на ограничение, заключающееся в том, что tDCS ограничена с точки зрения параметров стимуляции (т.е. отсутствие возможности регулировать частоту стимуляции), эти клинические результаты обнадеживают и должны способствовать дальнейшему использованию неинвазивной нейромодуляции у пациентов с DOC, наряду с исследованиями. о лежащих в основе механизмах.
Доктор Джон Д. Кресслер
Материалы конференции
2019 | 2018 | 2017 | 2016 | 2015 | 2014 | 2013 | 2012 | 2011 | 2010 | 2009 | 2008 | 2007 | 2006 — 1986
2019 Документы
И.Джу, М. Макпартлин, К.-В.П. Хуанг, М. Доэрти, Б. Вайланкур, К.Д. Чеон и Дж.Д. Кресслер, «Высоко линейный, высокомощный усилитель мощности SiGe HBT для WLAN 802.11ac / ax, использующий компактный четырехканальный трансформатор с коротким замыканием второй гармоники и встроенные термодатчики», Технический сборник IEEE International Solid-State Circuits Conference , стр. 80-81, 2019.
C.T. Коэн, В. Уильямс, Н.Э. Лоуренко, М. Фрунчи, К. Чеон и Дж. Д. Кресслер, «MicroNimbus: одночиповый SiGe-радиометр 60 ГГц для дистанционного зондирования из космоса», Труды конференции GOMAC-Tech 2019 — Государственные приложения микросхем и критических технологий , стр. .1-4, 2019.
П. Гоули и Дж. Д. Кресслер, «Электронно-фотонные интегральные схемы на основе кремния: отказоустойчивость в космической среде», Труды конференции GOMAC-Tech, 2019 г. — Государственная конференция по применению микросхем и критическим технологиям , стр. 1-4, 2019 г.
Н. Лоренко, Д. Денисон, А. Кардосо, К. Коэн, М.-К. Чо, и Дж.Д. Кресслер, «Интегральная схема с поперечным радиочастотным фильтром (TRAFFIC)», Труды конференции GOMAC-Tech 2019 года по правительственным приложениям микросхем и критическим технологиям, стр.1-4, 2019.
S. Zeinolabedinzadeh, A.C. Ulusoy, R.L. Schmid, F. Inanlou, T. Chi, J.S. Парк, Х. Ван и Дж. Д. Кресслер, «SiGe-трансивер W-диапазона с новой встроенной функцией самотестирования», Труды тематического совещания IEEE 2019 года по кремниевым монолитным интегральным схемам в радиочастотных системах стр. 1-4, 2019.
2018 Документы
A.A. Наваз, Дж. Qayyum, A.C. Ulusoy, N. Karaulac, S. Jung, D. Fleischhauer, S. Shankar, J.D. Cressler и W.Ван Норт, «Новый образец качества для оценки задержки затвора CML», Труды тематической конференции IEEE по датчикам и сенсорным сетям , стр. 73-76, 2018.
А. Ильдефонсо и Дж. Д. Кресслер, «Стратегии радиационной защиты для ВЧ схем и систем на основе SiGe», Труды 2018 GOMAC-Tech — Конференция по правительственным приложениям микросхем и критическим технологиям , стр. 1-4, 2018.
М.-К. Чо, И. Сонг и Дж. Д. Кресслер, «SiGe двунаправленный приемопередающий чип на основе истинной временной задержки для крупномасштабных широкополосных антенн с синхронизацией», Труды симпозиума по радиочастотным интегральным схемам IEEE MTT-S 2018, стр.1-4, 2018.
M. Frounchi, C. Coen, C.D.Y. Чеон, Н. Лоренко, У. Уильямс и Дж. Д. Кресслер, «Интерфейс приемника SiGe с подавлением изображения в V-диапазоне для дистанционного зондирования атмосферы», Труды симпозиума IEEE BiCMOS и сложных полупроводниковых интегральных схем и технологий , 2018 г., С. 223-226, 2018.
С. Рао, Т. Чанг, И. Сонг, М.-К. Чо и Дж.Д. Кресслер, «Распределенный, многослойный SiGe-усилитель мощности на 1–20 ГГц», Труды 2018 IEEE BiCMOS and Compound Semiconductor Integrated Circuits and Technology Symposium , стр.202-205, 2018.
AP Omprakash, A. Ildefonso, G. Tzintzarov, J. Babcock, R. Mukhopadhyay и JD Cressler, «Использование SiGe-on-SOI HBT для создания аналоговых схем 300C», Proceedings of the 2018 IEEE BiCMOS and Compound Semiconductor Симпозиум по интегральным схемам и технологиям , стр. 206-209, 2018 г.
U.S. Raghunathan, B.R. Виер, З. Флитвуд, М.А. Окли, В. Джейн и Дж. Д. Кресслер, «Оптимизация профиля базы эмиттера SiGe HBT для повышения термостабильности и радиочастотной линейности», Труды симпозиума IEEE BiCMOS и сложных полупроводниковых интегральных схем и технологий 2018 г. , стр.190-193, 2018.
B.R. Виер, Р.П. Мартинес, США Рагхунатан, Х. Ин, С. Зейнолабединзаде и Дж.Д. Кресслер, «Возвращение к безопасной рабочей области: модели старения SiGe HBT для проектирования схем с учетом надежности», Труды IEEE BiCMOS и интегральных схем на основе сложных полупроводников 2018 и технологический симпозиум , стр. 215-218, 2018.
Т. Ши, Ю. Гонг, С. Равичандран, В. Сундарам, Дж. Д. Кресслер и Р. Туммала, «Следующее поколение автомобильных радаров с передовыми достижениями в области устройств SiGe и встраивания в стеклянные панели (GPE)», Proceedings of 68-я Конференция по электронным компонентам и технологиям IEEE 2018 , стр.1245-1250, 2018. DOI: 10.1109 / ECTC.2018.00192
2017 Документы
З. Флитвуд и Дж. Д. Кресслер, «Радиационное упрочнение и корреляция тяжелых ионов с лазером в устройствах и схемах SiGe», Труды 2017 GOMAC-Tech — Конференция по правительственным приложениям микросхем и критическим технологиям , стр. 1-4, 2017
И. Джу, М.А. Окли и Дж.Д. Кресслер, «Каскодный усилитель мощности SiGe HBT с обратной связью класса F класса F с гармонически настроенным выходным трансформатором», Труды симпозиума по радиочастотным интегральным схемам IEEE MTT-S 2017 г. , стр. .1-4, 2017.
Я. Гонг, М.-К. Чо и Дж.Д. Кресслер, «Двунаправленный 6-разрядный фазовращатель X-диапазона для антенн с фазированной решеткой, использующий метод активного DPDT-переключателя», Труды симпозиума по радиочастотным интегральным схемам IEEE MTT-S 2017 г. , стр. 1-4, 2017.
W. Williams, C. Coen, M. Frounchi, N. Lourenco и JD Cressler, «MicroNimbus: температурный профилометр Cubesat для атмосферы Земли с использованием однокристального SiGe-радиометра 60 ГГц», Proceedings of the 2017 IEEE International Geoscience и симпозиум по дистанционному зондированию , стр.1-4, 2017.
C.T. Коэн, А. Ильдефонсо, З. Флитвуд и Дж. Д. Кресслер, «Квадратный детектор SiGe HBT 19–34 ГГц со сверхнизким 1 / f-шумом для атмосферных радиометров», Труды Европейской конференции по СВЧ-интегральным схемам 2017 г. , стр. 163–166, 2017
M.T. Wachter, A. Ildefonso, Z.E. Флитвуд, Н. Лоуренко, Г. Цинцаров, Н.Дж.-Х. Рош, А. Хачатрян, П. Макмарр, Х. Хьюз, Дж. Уорнер, Д. МакМорроу, Полин Паки и Дж. Д. Кресслер, «Влияние общей ионизирующей дозы на переходную реакцию технологии SiGe BiCMOS», Proceedings of the Конференция по излучению и его воздействию на компоненты и системы, 2016 г., , статья J4, стр.1-4, 2017.
А. Ильдефонсо, И. Сонг, З.Е. Флитвуд, Н. Лоуренко, М. Вахтер и Дж. Д. Кресслер, «Моделирование распространения единичных переходных процессов в приемнике прямого преобразования SiGe BiCMOS», Труды конференции 2016 года по излучению и его воздействию на компоненты и системы , документ J9, стр. 1-4, 2017
R. Perez-Martinez, U.S. Raghunathan, A.P. Omprakash, B.R. Wier, M.A. Oakley, Z.E. Флитвуд и Дж.Д. Кресслер, «Прогнозирование серьезных отказов и максимальный рабочий диапазон SiGe HBT», Труды конференции IEEE Bipolar / BiCMOS, 2017 г., , стр.122-125, 2017.
2016 Документы
I. Ju, M-K. Чо, Р. Шмид, М. Митчелл и Дж. Д. Кресслер, «Совместная разработка SiGe X-диапазона, асимметричного переключателя SPDT с низкими вносимыми потерями, мощного переключателя с управляемой нагрузкой и сверхмалошумящего МШУ для модулей T / R следующего поколения. , « Труды Международного симпозиума IEEE MTT-S 2016 г. , стр. 1–4, 2016 г.
И. Сонг, М.-К. Чо, Дж. Хопкинс, М. Митчелл и Дж. Д. Кресслер, «Широкополосный активный двунаправленный делитель / сумматор мощности 2–22 ГГц на основе 130-нм SiGe BiCMOS Technology», Труды Международного симпозиума IEEE MTT-S 2016 , стр.1-4, 2016.
К. Коэн, С. Зейнолабединзаде, М. Кайнак, Б. Тиллак и Дж. Д. Кресслер, «Высокоэффективный удвоитель частоты 138–170 ГГц SiGe HBT для приложений с ограничением мощности», Труды 2016 IEEE MTT-S Симпозиум по радиочастотным интегральным схемам , стр. 23–26, 2016 г.
США Рагхунатан, Б. Виер, Р.П. Мартинес, Х. Ин, З. Флитвуд, С. Зейнолабединзаде, А. Омпракаш и Дж. Д. Кресслер, «Моделирование сильноточного повреждения в SiGe HBT при импульсном напряжении», Труды 2016 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp.17-20, 2016.
М.-К. Чо, И. Сонг, З. Флитвуд и Дж. Д. Кресслер, «Широкополосный активный двунаправленный цифровой ступенчатый аттенюатор SiGe с использованием активного переключателя DPDT», Труды конференции IEEE Bipolar / BiCMOS 2016, , стр. 122 -125, 2016.
B.R. Wier, U.S. Raghunathan, Z.E. Флитвуд, М.А.Оукли, А.Дж. Джозеф, Вибхор Джайн и Дж.Д. Кресслер, «Об использовании вертикальных коллекторов сверхперехода для улучшения характеристик пробоя в SiGe HBT», Труды конференции IEEE Bipolar / BiCMOS, 2016 г., , стр.21-24, 2016.
B.R. Wier, U.S. Raghunathan, Z.E. Флитвуд, М.А.Оукли, А.Дж. Джозеф, Вибхор Джайн и Дж.Д. Кресслер, «Об использовании вертикальных коллекторов сверхперехода для улучшения характеристик пробоя в SiGe HBT», Труды конференции IEEE Bipolar / BiCMOS 2016 and Technology Meeting , pp. 21-24, 2016.
2015 Документы
A.C. Ulusoy, R.L. Schmid, M. Kaynak, B. Tillack, J.D. Cressler, High-Performance W-band LNA и SPDT Switch in 0.13 м SiGe HBT Technology, Материалы симпозиума IEEE Radio and Wireless 2015, стр. 162-164, 2105.
Z.E. Fleetwood, B. Wier, U. Raghunathan, N.E. Лоренко, М. Окли, А.Дж. Джозеф и Дж.Д. Кресслер, Оптимизация вертикального профиля SiGe HBT для смягчения радиационно-индуцированных нарушений, Труды 2015 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp. 72-75, 2015. Награда за лучшую студенческую работу.
А.П. Омпракаш, П.С. Чакраборти, Х. Инь, А. Кардозу, А.Ильдефонсо и Дж.Д. Кресслер, «О потенциале использования SiGe HBT на SOI для поддержки новых приложений до 300C», Труды конференции IEEE Bipolar / BiCMOS 2015, 2015, , стр. 27-30, 2015.
Р. Шмид, П. Сонг, К. Коэн, А. К. Улусой и Дж. Д. Кресслер, «Интегрированный кремний-германиевый петлевой и входной передаточно-приемный переключатель W-диапазона для встроенного самотестирования», Протоколы Международного симпозиума IEEE MTT-S 2015 г. , стр.1-4, 2015.
AC Ulusoy, RL Schmid, M. Kaynak, B. Tillack и JD Cressler, «High-Performance W-band LNA and SPDT Switch in 0.13 m SiGe HBT Technology», Proceedings of the 2015 IEEE Radio and Wireless Symposium , С. 162-164, 2015.
W. Khan, C. Ulusoy, R. Schmid, T. Chi, JD Cressler, H. Wang и J. Papapolymerou, «SPDT-переключатель диапазона D (110–170 ГГц) в 32 нм CMOS SOI», Труды Международного симпозиума IEEE MTT-S 2015 г., , стр.1-4, 2015.
2014 Документы
A.S. Кардосо, П. Саха, П.С. Чакраборти, Д. Флейшхауэр и Дж.Д. Кресслер, Широкополосные переключатели SPDT с низкими потерями и фазовращатель с коммутируемой линией в 180-нм RF CMOS на технологии SOI, Proceedings of the 2014 IEEE Radio and Wireless Symposium , pp. 199-201, 2014.
AC Ulusoy, RL Schmid, C. Coen и JD Cressler, Малошумящий усилитель SiGe HBT с переключаемым ядром для радиометров миллиметрового диапазона, Труды тематического совещания IEEE 2014 года по кремниевым монолитным интегральным схемам в радиочастотных системах , стр.22-24, 2014.
С. Юнг, П. Сонг, И. Сонг, Дж. Д. Кресслер, Дж. А. Бэбкок, Исследование температурно-зависимой линейности слабонасыщенных, электрически согласованных NPN и PNP SiGe HBT, Труды тематического совещания 2014 IEEE по кремниевым монолитным интегральным схемам в радиочастотных системах , стр. 86-88, 2014.
П. Сонг, А.К. Улусой, Р. Шмид и Дж. Д. Кресслер, SiGe LNA с высоким коэффициентом усиления, W-диапазон и коэффициент шума ниже 4,0 дБ, Труды Международного симпозиума IEEE MTT-S 2014 г. 17, 4 стр., 2014.
И. Сонг, Р. Шмид, Д. Ховард, С. Юнг и Дж.Д.Кресслер, Широкополосный асимметричный балун Маршана с широкополосной связью 34–110 ГГц в технологии 180 нм SiGe BiCMOS, Труды 2014 IEEE MTT- S International Microwave Symposium , статья WEP-20, 4 страницы, 2014.
С. Зейнолабединзаде, П. Сонг, М. Кайнак, М. Камарей, Б. Тиллак и Дж. Д. Кресслер, Низкий фазовый шум и высокая выходная мощность 368 ГГц и 156 ГГц Источники сигналов в 130-нм технологии SiGe HBT, Труды 2014 IEEE MTT-S International Microwave Symposium , статья THP-18, 4 страницы, 2014.
M.A. Oakley, B. Wier, U.S. Raghunathan, P.S. Чакраборти и Джон Д. Кресслер, О надежности каскодных усилителей SiGe HBT, Труды симпозиума по радиочастотным интегральным схемам IEEE MTT-S 2014 года , стр. 445-448, 2014.
П. Сонг, Р. Л. Шмид, А. Улусой и Дж. Д. Кресслер, Однополюсный однополюсный переключатель высокой мощности с низкими потерями, использующий SiGe PIN-диоды, Труды симпозиума по радиочастотным интегральным схемам IEEE MTT-S 2014 г. , стр.195-198, 2014.
С. Зейнолабединзаде, М. Кайнак, В. Хан, М. Камарей, Б. Тиллак, Дж. Папаполимеру и Дж. Д. Кресслер, 314 ГГц, полностью интегрированный SiGe-передатчик и приемник со встроенной антенной, Proceedings of the 2014 IEEE Симпозиум по радиочастотным интегральным схемам МТТ-С , стр. 361-364, 2014.
AC Ulusoy, RL Schmid, WT Khan, M. Kaynak, B. Tillack и John D. Cressler, Исследование деградации fT / fmax из-за межсоединений устройств в технологии SiGe HBT 0,5 ТГц, Proceedings of the 2014 IEEE Bipolar / Совещание по схемам и технологиям BiCMOS , стр.211-214, 2014.
Р.Л. Шмид и Дж.Д. Кресслер, SiGe-усилитель с переменным коэффициентом усиления с семью состояниями с цифровым управлением, малошумящий усилитель, Труды конференции IEEE Bipolar / BiCMOS, 2014 г. , стр. 187-190, 2014.
П. Песня, А .. Улусой, Р.Л. Шмид, С.Н. Зейнолабединзаде и Дж.Д. Кресслер, SiGe-усилители мощности W-диапазона, Труды 2014 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp. 151-154, 2014.
С.Коэн, Р. Шмид, Дж. Д. Кресслер, М. Кайнак и Б. Тиллак, МШУ с радиометром со сверхмалым шумом и низким энергопотреблением 18,7 ГГц в SiGe-технологии 0,5 ТГц с использованием индукторов с травлением на задней стороне, Труды 2014 IEEE Bipolar / Совещание по схемам и технологиям BiCMOS , стр. 191–194, 2014 г.
T. Chi, J.S. Парк, Р.Л. Шмид, А.С. Улусой, Дж. Д. Кресслер и Х. Ван, Маломощный и сверхкомпактный передний интерфейс передатчика W-диапазона на основе 90-нм технологии SiGe BiCMOS, Труды 2014 IEEE Bipolar / BiCMOS Circuits and Technology Встреча , стр.155-158, 2014.
S. Zeinolabedinzadeh, A.L. Vera Lpez, N.E. Lourenco, M. Kaynak, B. Tillack, M. Kamarei, J. Papapolymerou и JD Cressler, Масштабируемая матрица передатчиков SiGe 22, 316 ГГц с новым методом фазовой синхронизации и встроенными антеннами, маломощные и сверхкомпактные W -полосный передатчик с использованием технологии 90 нм SiGe BiCMOS, Труды конференции IEEE Bipolar / BiCMOS 2014 г., , стр. 60-63, 2014.
С. Юнг, Т. Инглэнд, И. Сонг, Б. Виер, Дж.Д. Кресслер, Д.А. Бэбкок, Дополнительный SiGe HBT на стабилизаторе напряжения с малым падением напряжения SOI с использованием обнуляющего резистора, Труды Международного симпозиума IEEE 2014 по схемам и системам , стр. 2453-2456, 2014.
С. Юнг, И. Сонг и Дж. Д. Кресслер, Систематическая методология применения графа потока сигналов масонов к анализу цепей обратной связи, Труды Международного симпозиума IEEE 2014 по схемам и системам , стр. 2421-2424, 2014.
Ф.Inanlou, W. Khan, S. Zeinolabedinzadeh, P. Song, RL Schmid, T. Chi, AC Ulusoy, J. Papapolymerou, H. Wang и JD Cressler, Компактный, маломощный, несимметричный и дифференциальный SiGe W- диапазонные LNA, Proceedings of the 2014 IEEE European Microwave Symposium , pp. 1396-1399, 2014.
2013 Документы
Р. Шмид, Т. Квач, А. Маттамана и Дж.Д. Кресслер, Анализ переключаемых транзисторных ядер SiGe HBT для малошумящих усилителей, Труды конференции GOMAC Tech 2013 — правительственные приложения и критические технологии , стр.115-118, 2013.
S. Jung, J.D. Cressler, J.A. Бэбкок, Дж. Сестра и А. Бухгольц, Метод достижения низкого входного импеданса и независимости произведения коэффициента усиления и полосы пропускания в дифференциальных TIA, Труды Международного симпозиума IEEE 2013 г. по схемам и системам , стр. 598-601, 2013
С. Шанкар, В. ван Ноорт и Дж.Д. Кресслер, Прогнозирование задержки затвора CML для больших сигналов с использованием Y-параметров для быстрой оптимизации процесса, Труды конференции IEEE Bipolar / BiCMOS 2013, , стр.207-210, 2103.
У. Рагхунатан, П.С. Чакраборти, Б. Вейр, Дж. Д. Кресслер, Х. Ясуда и П. Менц, TCAD-моделирование накопленных стрессовых повреждений во время зависимого от времени смешанного стресса, Труды конференции IEEE Bipolar / BiCMOS 2013, , стр.179 -183, 2013.
Д. Ховард, Т. Инглэнд, Н. Лоренко, А. Кардосо и Дж. Д. Кресслер, Схема определения характеристик SiGe HBT на кристалле для использования в радиочастотных системах с самовосстановлением, Proceedings of the 2013 IEEE Bipolar / BiCMOS Circuits and Technology Встреча , стр.203-206, 2013.
Р. Шмид, П. Сонг и Дж. Д. Кресслер, Компактный ВЧ-переключатель SPDT на базе трансформатора 60 ГГц, использующий диодные SiGe HBT, Труды IEEE Bipolar / BiCMOS Circuits and Technology Meeting 2013, pp.111- 114, 2013.
К. Коэн, Дж.Р. Пипмайер и Дж.Д. Кресслер, Интегрированная кремний-германиевая электроника для радиометров на базе CUBESAT, Труды Международного симпозиума IEEE 2013 года по геонауке и дистанционному зондированию , стр. 1286-1289, 2013.
B.R. Вир, США Рагхунатан, П.С. Чакраборти, Дж. Д. Кресслер, Х. Ясуда, П. Менц, Механизмы деградации базового тока в SiGe HBT NPN, подверженных сильноточной нагрузке, Труды Международного симпозиума по исследованиям твердотельных устройств IEEE 2013 г. , стр. 1-2, 2013.
2012 Документы
П.К. Саха, С. Шанкар, Р. Шмид, Р. Миллс и Дж. Д. Кресслер, «Анализ и разработка малошумящего усилителя 326 ГГц в технологии SiGe HBT», Труды симпозиума IEEE Radio and Wireless 2012 , стр.203-206, 2012.
С. Сет, Дж. Д. Кресслер, Дж. А. Бэбкок, Дж. Цестра, Т. Краковски, Дж. Танг и А. Бухгольц, «Сравнение характеристик интермодуляционных искажений компактных моделей HICUM и VBIC для pnp SiGe HBT на SOI», Труды тематического совещания 2012 IEEE по кремнию Монолитные интегральные схемы в радиочастотных системах , стр. 219-222, 2012.
C.H.J. Пох, Р.Л. Шмид, Дж.Д. Кресслер и Дж. Папаполимеру, «Переключатель SPDT с X-диапазона на Ka-диапазон с использованием 200 нм SiGe HBT», Труды тематического совещания IEEE 2012 года по кремниевым монолитным интегральным схемам в радиочастотных системах , стр.183-186, 2012.
А. Гоял, М. Сваминатан, А. Чаттерджи, Д. Ховард и Дж. Д. Кресслер, «Самопроверяемый малошумящий усилитель на основе SiGe и встроенная методология самотестирования для множественных характеристик характеристик РЧ-усилителей», Труды 2012 г. Международный симпозиум по качеству электронного дизайна , стр. 7-12, 2012.
T.D. England, R.M. Дистелхорст, С. Финн, Л. Наджафизаде, Дж.Д. Кресслер, «Проводные приемопередатчики SiGe BiCMOS с холодным подключением для распределенных электронных систем», Труды конференции IEEE Aerospace 2012 , стр.1-8, 2012 (на CDROM).
С. Шанкар, П. Саха, Д. К. Ховард, Р. Дистелхорст, Т. Д. Англия, А. Кардосо и Дж. Д. Кресслер, Самовосстановление на кристалле коэффициента подавления изображения смесителя для электронных систем со смешанными сигналами, Proceedings of the 2012 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp. 190-193, 2012.
R.L. Schmid, C.T. Коэн, С. Шанкар и Дж.Д. Крессл, Лучшие практики обеспечения стабильности каскодных усилителей SiGe HBT с низким уровнем шума, Труды конференции IEEE Bipolar / BiCMOS 2012, , стр.194-197, 2012.
С. Зейнолабединзаде, Н. Лоуренко, М. Камарей и Дж. Д. Кресслер, Дифференциальный SiGe VCO V-диапазона с безваракторной настройкой, Труды конференции IEEE Bipolar / BiCMOS, 2012 г., , стр. 198-201 , 2012.
Л. Наджафизаде, С. Аслам и Дж.Д. Кресслер, Эксплуатация опорных схем SiGe в условиях планетарной внешней среды, Труды Международного семинара 2012 года по контрольно-измерительным приборам для планетарных миссий , стр.1-2, 2012.
D. Howard, C.-S. Чо, и Дж.Д. Кресслер, Широкополосный, миллиметровый, асимметричный балун Маршана в 180 нм SiGe BiCMOS технологии, Труды симпозиума по радиочастотным интегральным схемам 2012 IEEE MTT-S , стр. 425-428, 2012.
K.C. Правин, Н. Пушпа, Дж. Д. Кресслер и А. П. Гнана Пракаш, Исследование радиационной реакции двух поколений SiGe HBT на гамма-излучение Co-60 и облучение тяжелыми ионами, Труды 19 -го Национального симпозиума по радиационной физике , стр.244-247, 2012.
N.E. Лоуренко, Р.Л.Шмид, К.А. Моэн, С. Phillips, TD England, JD Cressler, J. Pekarik, J. Adkisson, R. Camillo-Castillo, P. Cheng, J. Ellis-Monaghan, P. Gray, D. Harame, M. Khater, Q. Liu, A. Валлетт, Б. Зеттерлунд, В. Джайн и В. Каушал, «Полная доза и переходная характеристика SiGe HBT от нового 4-го поколения, 90 нм SiGe BiCMOS Technology», Труды конференции IEEE 2012 г. по ядерным и космическим радиационным эффектам Data Workshop , стр. 205-209, 2012 г.
2011 Документы
Р. Арора, С. Сет, C.H.J. Пох, Дж.Д. Кресслер, А.К. Саттон и Х. Найфех, «Влияние расстояния между контактами исток / сток на ВЧ-надежность 45-нм ВЧ-полевых МОП-транзисторов», Труды Международного симпозиума по физике надежности IEEE 2011 г., стр. 461-466, 2011.
C.A.D. Морсилло, К. Паттерсон, Т. Тривикраман, Б. Лакруа, К. Коэн, К.Х.Дж. По, Дж. Д. Кресслер и Дж. Папаполимеру, «Легкая 64-элементная органическая фазированная решетка с интегрированной схемой передачи-приема SiGe в X-диапазоне», Труды Международного симпозиума IEEE MTT-S 2011 г., , стр. .1-4, 2011.
С. Шанкар, С.Дж. Хорст, П. Саха, Д. К. Ховард, Р. Дистелхорст, TD England и JD Cressler, «Широкий диапазон настройки, источник тестового сигнала с синхронизацией по амплитуде для самовосстановления, электронные системы со смешанными сигналами», Труды 2011 г. IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp. 29-32, 2011.
С. Сет, С. Х. По, Т. Тривикраман, Р. Арора и Дж. Д. Кресслер, «Использование насыщенных SiGe HBT для реализации сверхнизких / мощных малошумящих усилителей X-диапазона», Труды конференции IEEE Bipolar / BiCMOS 2011 г. , стр.102-105, 2011.
DC Howard, P. Saha, S. Shankar, R. Diestelhorst, T. England и JD Cressler, «UWB SiGe LNA для многополосных приложений с восстановлением процесса на основе извлечения характеристик устройства постоянного тока», Proceedings of the 2011 IEEE Совещание по биполярным / биКМОП схемам и технологиям , стр. 110-113, 2011.
П.К. Саха, Д. Ховард, С. Шанкар и Дж.Д. Кресслер, «Адаптивный широкополосный микшер для подавления изображений SiGe для самовосстанавливающегося приемника», Труды конференции IEEE Bipolar / BiCMOS, 2011 г., , стр.98-101, 2011.
П. Чакраборти, К.А. Моен и Дж.Д. Кресслер, «Прогнозирующее моделирование в TCAD индуцированного масштабированием, обратносмещенного, туннельного тока эмиттер-база в SiGe HBT», Труды конференции IEEE Bipolar / BiCMOS, 2011 г., , стр. 13–16, 2011 г. .
X. Geng, D. Ma, Z. Chen, F. Dai, J.D. Cressler, J.A. Яегер, М. Mojarradi, A. Mantooth, B.J. Blalock и R.W. Berger, «Интерфейс резистивного датчика высокоскоростного канала с RHBD в 0.5 м SiGe BiCMOS для UWT от -180 ° C до 120 ° C, Труды конференции IEEE Bipolar / BiCMOS, 2011 г., , стр.79-82, 2011.
С.Э. Паттерсон, Т. Тривикраман, С. Бхаттачарья, C.T. Коэн, Дж. Д. Кресслер и Дж. Папаполимеру, «Разработка технологии многослойной органической упаковки для полностью встроенного модуля T / R», Труды Европейского симпозиума IEEE по микроволновому излучению 2011 г., стр. 261-264, 2011.
Документы 2010 г.
Т.К. Тривикраман, А. Мадан и Дж. Д. Кресслер, «Об устойчивости SiGe HBT LNA к большим сигналам для высокочастотных беспроводных приложений», Труды тематического совещания IEEE 2010 г. по кремниевым монолитным интегральным схемам в радиочастотных системах , стр. 156-159, 2010.
А. Мадан, Т. Тривикраман, С. Сет, Р. Верма, Дж. По и Дж. Д. Кресслер, «Новый и простой метод измерения для определения интермодуляционных искажений без использования анализатора спектра», Труды тематического совещания IEEE 2010 г. по кремниевым монолитным интегральным схемам в RF Systems , стр.88-91, 2010.
J.C.-H. Пох, П. Ченг, Т.К. Тривикраман и Дж.Д. Кресслер, «SiGe-усилитель с низким уровнем шума, L-диапазон, L-диапазон и полностью интегрированная согласующая сеть», Труды тематического совещания IEEE 2010 г. по кремниевым монолитным интегральным схемам в радиочастотных системах , стр. -71, 2010.
С.Э. Паттерсон, Т. Тривикраман, С. Бхаттачарья, Дж.Д. Кресслер и Дж. Папаполимеру, «Легкая органическая антенная решетка X-диапазона со встроенным усилителем SiGe», Труды симпозиума IEEE Radio and Wireless 2010 , стр.84-87, 2010.
R.M. Дистелхорст, С. Финн, Л. Наджафизаде, Д. Ма, П. Си, К. Улаганатан, Дж. Д. Кресслер, Б. Блалок, Ф. Дай, А. Мантуз, Л. Дель Кастильо, М. Мохарради и Р. Бергер , «Монолитный, широкотемпературный канал усиления заряда для пьезоэлектрических датчиков в экстремальных условиях», Труды аэрокосмической конференции IEEE 2010 , стр. 1-9, 2010.
К. Уэббер, Дж. Холмс, М. Фрэнсис, Р. Бергер, А. Мантуз, К.Корнетт, Б. Блэлок, Р. Гринвелл, Дж. Д. Кресслер, Р. Дистелхорст и А. Аутурс, «Управляемые событиями методы моделирования смешанных сигналов для функциональной проверки системы в пакете», Труды конференции IEEE Aerospace Conference 2010 , С. 1-15, 2010.
Д. Томас, Н. Лоренко, Дж. Д. Кресслер и С. Финн, «SiGe-усилитель и буферные схемы для высокотемпературных приложений», Труды Международной конференции по высокотемпературной электронике IMAPS 2010 г. , стр.379-385, 2010.
P.S. Чакраборти, К. Моен, Дж. Д. Кресслер, Х. Хо, Х. Ясуда, Б. Эклунд и Р. Уайз, «Прогнозирующее моделирование проводимости SiGe HBT с использованием TCAD», Труды Международного собрания по технологиям и устройствам SiGe, 2010 г., , стр. 1-2 августа 2010 г.
C.H.J. Пох, С. Бхаттачарья, Дж. Фергюсон, Дж. Д. Кресслер и Дж. Папаполимеру, «Извлечение сосредоточенного элемента, модель эквивалентной схемы для межсоединений в трехмерных корпусах с использованием структуры с одним переходом со встроенными конденсаторами», Труды журнала «Электронные компоненты и технологии» 2010 г. Конференция , стр.1783-1788, 2010.
Дж. Д. Кресслер, «Новый подход к образованию в области микроэлектроники и нанотехнологий для студентов всех дисциплин», Труды Ежегодной конференции Американской ассоциации инженерного образования 2010 г., статья AC-2010-89, стр. 1-12, 2010 г.
С. Хорст, П. Чакраборти, П. Саха, Дж.Д. Кресслер, Х. Густат, Б. Хайнеманн, Г.Г. Фишер, Д. Нолл и Б. Тиллак, «Сравнение характеристик фазового шума осциллятора SiGe HBT npn и pnp на дополнительной платформе SiGe», Труды конференции IEEE Bipolar / BiCMOS, 2010 г., , стр.13-16, 2010.
С. Сет, Т. Тривикраман, П. Ченг, Дж. Д. Кресслер, Дж. Бэбкок и А. Буххольц, «Исследование надежности РЧ-сигналов для больших сигналов дополнительных SiGe HBT на SOI, предназначенных для использования в беспроводных приложениях», 2010 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp. 133-136, 2010.
П. Ченг, С. Хорст, С. Филлипс, С. Сет, Р. Миллс, Дж. Д. Кресслер, Дж. Бэбкок и А. Буххольц, «Исследование низкочастотных и фазовых шумов в дополнительных SiGe HBT на КНИ», Протоколы конференции IEEE Bipolar / BiCMOS, 2010 г. , стр.165-168, 2010.
Дж. Юань, К.А. Моэн и Дж.Д. Кресслер, «Исследование транспорта между коллектором и базой в SiGe HBT, разработанных для полутерагерцовых скоростей», Труды конференции IEEE Bipolar / BiCMOS, 2010, , стр. 157-160, 2010.
П. Саха и Дж. Д. Кресслер, «Настраиваемый микшер для подавления изображения SiGe X-диапазона», Труды конференции IEEE Bipolar / BiCMOS, 2010 г., , стр. 196-199, 2010.
К.А. Моэн, Дж. Юань, П. Чакраборти, М. Беллини, Дж. Д. Кресслер, Х. Хо, Х. Ясуда и Р. Уайз, «Улучшенный двухмерный региональный анализ времени прохождения для оптимального масштабирования SiGe HBT», Материалы конференции IEEE Bipolar / BiCMOS, 2010 г., , стр. 257-260, 2010 г.
С. Шанкар, С. Хорст и Дж. Д. Кресслер, «SiGe-кольцевые генераторы X-to-Ku диапазона с перестройкой частоты и амплитуды для многополосных приложений BIST», Труды конференции IEEE Bipolar / BiCMOS, 2010 г., , стр. .9-12, 2010.
А. Мадан, Дж.Д. Кресслер, А.Дж. Джозеф, «Высоколинейный обратный режим SiGe BiCMOS RF-переключатель», Труды конференции IEEE Bipolar / BiCMOS, 2010 г., , стр. 61-64, 2010.
P.S. Чакраборти, С.Дж. Хорст, К. Моэн, М. Беллини и Дж.Д. Кресслер, «Исследование электротермической нестабильности SiGe HBT 150 ГГц, изготовленных на основе КНИ», Труды конференции IEEE Bipolar / BiCMOS, 2010 г., , стр.141-144, 2010.
Л. Луо, Г. Ню, Л. Наджафизаде и Дж. Д. Кресслер, «Влияние неидеальной температурной зависимости IC-VBE на сверхширокий температурный диапазон эталонных схем SiGe HBT с запрещенной зоной», Proceedings of the 2010 IEEE Bipolar / Совещание по схемам и технологиям BiCMOS , стр. 220-223, 2010 г.
С.Э. Паттерсон, Т. Тривикраман, А. Йепес, С. Бхаттачарья, Дж. Д. Кресслер и Дж. Папаполимеру, «Внедрение недорогой и легкой антенны X-диапазона со встроенной электроникой SiGe RF», Труды Международного симпозиума по геофизике и дистанционному зондированию 2010 IEEE , стр.681-684, 2010.
В ВИДЕ. Киз, Дж. Адамс, Л.М. Смит, Р.Э. Рэй и Дж.Д. Кресслер, «Продвинутая авионика и процессорные системы для гибкой архитектуры космических исследований», Труды конференции AIAA Space Conference , 2010, стр. 1-8, 2010.
WT Khan, S. Bathacharya, S. Horst, JD Cressler и J. Papapolymerou, «Осциллятор K-диапазона с низким уровнем фазового шума на органической жидкокристаллической полимерной подложке (LCP)», Труды Международного симпозиума IEEE MTT-S 2010 г. , стр.1186-1189, 2010.
Д.К. Ховард, К.Х.Дж. Пох и Дж. Д. Кресслер, «Сверхширокополосный малошумящий усилитель SiGe HBT 3-20 ГГц с контролем усиления и обратных потерь для многополосных беспроводных приложений», Труды симпозиума IEEE Midwest Symposium по схемам и системам 2010 г., , стр. 445-448, 2010 г.
З. Сю, Г. Ниу, Л. Луо и Дж. Д. Кресслер, «Физическая модель туннельного тока с использованием ловушек для криогенного температурного компактного моделирования SiGe HBT», Труды симпозиума Электрохимического общества 2010 г. по SiGe и Ge: материалы , Обработка и устройства , vol.33, с. 301-310, 2010.
M. Mudholkar, HA Mantooth, G. Niu, JD Cressler, «Прямое извлечение параметров сопротивлений основания и эмиттера для SiGe HBT с использованием только данных постоянного тока», Труды симпозиума Электрохимического общества 2010 года по SiGe и Ge: материалы, обработка и Приборы , т. 33, стр. 337-348, 2010.
C.H.J. Пох, Д.К. Ховард, П. Ченг, Дж. Папаполимеру и Дж. Д. Кресслер, «Деэмбедирование линий передачи с использованием двухполупериодной модели контактной площадки с ЭМ-моделированием», Технический сборник 2010 IEEE Азиатско-Тихоокеанский симпозиум по СВЧ , стр.1208-1211, 2010.
C.H.J. Пох, С. Бхаттачарья, Дж. Д. Кресслер и Дж. Папаполимеру, «Выделение модели с сосредоточенными параметрами для межсоединений в LCP с использованием структуры с одним переходом со встроенным конденсатором», Труды конференции по электронным компонентам и технологиям IEEE 2010 г. , стр. 1783-1788, 2010 г.
А. Мадан, С. Сет, Дж. Д. Кресслер, X. Би и К. Грин «Понимание и моделирование линейности в кремниево-германиевых биполярных транзисторах с гетеропереходом», Труды корпорации TECHCON , 2010 г., статья 4.7. С. 1-4, 2010.
Р. Арора, К.А. Моэн, А. Мадан, Дж.Д. Кресслер, Э. Чжан, Д. Fleetwood, R.D. Schrimpf A.K. Саттон и Х. Найфех, «Влияние стыковки корпуса и расстояния между контактами истока / стока на надежность 45-нм ВЧ-КМОП горячего носителя», Труды Международного семинара по комплексной надежности 2010 г., стр. 56-60, 2010.
2009 Документы
С. Хорст, С. Филлипс, Х. Лавасани, Ф. Аязи и Дж. Д. Кресслер, «Цифровые делители частоты SiGe с пониженным остаточным фазовым шумом», Труды конференции IEEE Custom Integrated Circuits Conference, 2009 , стр.251-254, 2009.
Д. Ма, Х. Генг, Ф. Дай и Дж. Д. Кресслер, «Фильтр низких частот Баттерворта SC 6-го порядка для криогенных приложений от -180 ° C до 120 ° C», Proceedings of the 2009 IEEE Aerospace Conference , pp. 1 -8, 2009.
П. Ченг, С. Сет, К. М. Гренс, Т. Тривикраман, М. Беллини, Дж. Д. Кресслер, Дж. Бэбкок, Т. Чен, Дж. Ким и А. Буххольц, «Понимание зоны безопасности высокочастотной эксплуатации SiGe HBT на SOI», Proceedings of the 2009 IEEE Bipolar / Совещание по схемам и технологиям BiCMOS , стр.17-20, 2009.
С. Сет, П. Ченг, К. М. Гренс, Дж. Д. Кресслер, Дж. Бэбкок, Т. Чен, Дж. Ким и А. Бухгольц, «Сравнение радиочастотной линейности npn и pnp SiGe HBT, изготовленных на толстопленочной КНИ», Proceedings of the 2009 IEEE Совещание по биполярным схемам и технологиям BiCMOS , стр. 29-32, 2009 г.
T.K. Тривикраман, А. Аппасвами, С.Д. Филлипс, А. Саттон и Дж.Д. Кресслер, «Новая структура устройства с использованием совместно используемого субколлектора, каскодированного обратного режима SiGe HBT для повышения радиационной стойкости», Proceedings of the 2009 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp.79-82, 2009.
Дж. Юань и Дж. Д. Кресслер, «Новая конструкция коллектора суперперехода для улучшения напряжения пробоя в высокоскоростных SiGe HBT», Труды конференции IEEE Bipolar / BiCMOS, 2009 г., , стр. 75-78, 2009.
T.K. Тривикраман, W.-M.L. Куо и Дж.Д. Кресслер, «Двухканальный, сверхмалопотребляющий входной каскад SiGe BiCMOS-приемника для радаров с фазированной решеткой X-диапазона», Труды конференции IEEE Bipolar / BiCMOS, 2009 г., , стр.43-46, 2009.
Д. С. Ховард, Х. Ли и Дж. Д. Кресслер, «Маломощный, коэффициент шума 1,8–2,6 дБ, широкополосный малошумящий усилитель SiGe HBT для многополосных беспроводных приложений», Труды конференции IEEE 2009 г. С. 55-58, 2009.
Д. Б. Томас, Дж. Д. Кресслер, Л. Наджафизаде, Р. У. Джонсон, Л. Пельц и С. Филлипс, «Производительность и надежность SiGe-устройств и схем для высокотемпературных приложений», Труды Международной конференции по сетям высокотемпературной электроники IMAPS 2009 г. , с.49-56, 2009.
К. Паттерсон, Т.К. Тривикраман, С. Бхаттачарья, К.Х.Дж. По, Дж. Д. Кресслер и Дж. Папаполимеру, «Органическая вафельная упаковка для SiGe-малошумящего усилителя X-диапазона», Труды 2009 IEEE European Microwave Symposium , стр. 141-144, 2009.
T.K. Тривикраман, К. Гренс, Дж. М. Эндрюс, В.-М. Куо, Дж. П. Комо, М. Мортон, Дж. Д. Кресслер, Дж. Папаполимеру и М. Митчелл, «Однокристальный 4-битный модуль SiGe BiCMOS T / R для диапазона X Радиолокационные системы с фазированной решеткой », Труды XXX собрания Союза радионауки (URSI) , стр.1-2, 2009.
C.M. Гренс, П. Ченг и Дж.Д. Кресслер, «Исследование зоны безопасной работы РЧ сигналов с большим сигналом на каскодных SiGe HBT с агрессивным смещением для приложений усилителя мощности», Труды тематического совещания IEEE 2009 г. по кремниевым монолитным интегральным схемам в Системах РФ , с. 237-240, 2009.
T.K. Тривикраман, К. Гренс, W.-M.L. Куо, Дж. Эндрюс и Дж. Д. Кресслер, «Высоколинейный, X-диапазон, SiGe малошумящий усилитель для улучшенного динамического диапазона в радиолокационных и беспроводных системах следующего поколения», Труды тематического совещания IEEE 2009 г. по кремниевым монолитным интегральным схемам в РФ. Системы , стр.221-224, 2009.
П.К. Саха, Дж. П. Комо и Дж. Д. Кресслер, «Переключатель nMOS SPDT диапазона K и 1-битный фазовращатель, реализованный в 130-нм технологии SiGe BiCMOS», Труды тематического совещания IEEE 2009 г. по кремниевым монолитным интегральным схемам в радиочастотных системах , стр 92-95, 2009.
З. Сюй, Г. Ню, Л. Луо, П.С. Чакраборти, Д. Томас, П. Ченг и Дж.Д. Кресслер, «Криогенное моделирование слабых сигналов РЧ и извлечение параметров SiGe HBT», Труды тематического совещания IEEE 2009 г. по кремниевым монолитным интегральным схемам в радиочастотных системах , стр.96-99, 2009.
С. Хорст и Дж. Д. Кресслер, «Нелинейность AM / PM в SiGe HBT», Труды тематического совещания IEEE 2009 г. по кремниевым монолитным интегральным схемам в радиочастотных системах , стр. 241-244, 2009.
С. Санатанамурти, В. Рамачандран, М.Л. Аллес, Р.А. Рид, Л. Massengill, A. Raman, M. Turowski, A. Mantooth, M. Barlow, K. Moen, M. Bellini, A. Sutton и JD Cressler, «Моделирование чувствительности SRAM SEU при пониженной эксплуатации», Труды 2009 г. GOMAC-Tech — Государственная конференция по применению микросхем и критическим технологиям , стр.551-554, 2009.
К. Климер, Т. Хеффнер, М.Л. Аллес, Л. Массенгилл, Р.А. Рид, Б. Блэлок и Дж. Д. Кресслер, «Исследование одиночных переходных процессов на основе моделирования в операционном усилителе малой мощности SiGe BiCMOS», Труды конференции GOMAC-Tech-Government, 2009 г., , стр. 539 -542, 2009.
А. Мадан, Т. Тривикраман и Дж. Д. Кресслер, «Механизмы отказа в КМОП-переключателях, подверженных высокочастотному напряжению», Труды Международного симпозиума по физике надежности 2009 IEEE , стр.741-744, 2009.
С. Филлипс, А. Саттон, М. Беллини, А. Аппасвами, Дж. Кресслер, А. Грилло, Дж. Визкелети, П. Маршалл, М. МакКарди, Р. Рид и П. Додд, «Влияние глубин Изоляция траншеи на надежности передовых SiGe HBT в радиационной среде », Труды Международного симпозиума по физике надежности 2009 IEEE , стр. 157-164, 2009.
П.С. Чакраборти, А. Аппасвами, П.К. Саха, Н. Джа, Дж.Д. Кресслер, Х. Ясуда, Б. Эклунд и Р. Уайз, «Смешанные механизмы деградации напряжений в pnp SiGe HBT», Труды Международного симпозиума IEEE по физике надежности , 2009 г., стр.83-88, 2009.
2008 Документы
Дж. Юань и Дж. Д. Кресслер, «Повышение скорости SiGe HBT с использованием методов fT-удвоителя», Труды тематического совещания IEEE 2008 г. по кремниевым монолитным интегральным схемам в радиочастотных системах , стр. 50-53, 2008 г.
П. Ченг, А. Аппасвами, М. Беллини и Дж. Д. Кресслер, «Исследование явлений горячих носителей в npn и pnp SiGe HBT», Труды тематического совещания IEEE 2008 г. по кремниевым монолитным интегральным схемам в радиочастотных системах. , стр.54-57, 2008.
Р. Бергер, Дж. Д. Кресслер, Р. Гарбос, М. Мохарради, Л. Пельц, Б. Блалок, В. Джонсон, Г. Ниу, Ф. Дай, А. Мантуз, Дж. Холмс, М. Аллес, П. Маккласки, «Миниатюрная система сбора данных для экстремальных температур», Труды конференции IEEE Aerospace 2008 г., стр. 1-7, 2008 г.
A.S. Киз, Дж. Адамс, Дж.Д. Кресслер, Р. Дарти, М.А.Джонсон и М.С. Патрик, «Высокопроизводительная радиационно-стойкая электроника для космических и лунных сред, Труды Международного форума космических технологий и приложений 2008 года (STAIF) , стр.749-756, 2008.
А. Мадан, Дж. Д. Кресслер и С. Кестер, «Низкочастотный шум в полупроводниковых транзисторах SiGe n-MODFET со скрытым каналом», Труды Международного совещания по кремний-германиевым технологиям и устройствам (ISTDM) 2008 г. , стр. 60- 61, 2008.
П. Ченг, К.М. Гренс, А. Аппасвами, П. Чакраборти и Дж.Д. Кресслер, «Моделирование смешанного режима постоянного и высокочастотного напряжения в усилителях мощности SiGe HBT», Труды конференции по биполярным схемам и технологиям BiCMOS в 2008 г. , стр.133-136, 2008.
C.M. Гренс, С. Сет и Дж.Д. Кресслер, «Интермодуляционные характеристики SiGe HBT с общей базой», Труды конференции IEEE Bipolar / BiCMOS, 2008 г., , стр. 244-247, 2008 г.
Дж. Юань, Дж. Д. Кресслер, Ю. Цуй, Г. Ню, С. Финн и А. Джозеф, «О дизайне профиля SiGe HBT для радиочастотных лунных приложений вплоть до 43 К», Труды 2008 IEEE Bipolar / Совещание по схемам и технологиям BiCMOS , стр. 25-28, 2008 г.
X. Li, W.-M.L. Куо и Дж.Д. Кресслер, «SiGe-усилитель с отслеживанием и удержанием на 40 Гвыб / с», Труды конференции IEEE Bipolar / BiCMOS, 2008 г., , стр. 1-4, 2008.
Л. Луо, Г. Ниу, Д. Томас, Дж. Юань и Дж. Д. Кресслер, «Предел максимального выходного напряжения принудительного зажима IE в SiGe HBT, работающих при криогенных температурах», Труды 2008 IEEE Bipolar / BiCMOS Совещание по схемам и технологиям , стр. 41-44, 2008 г.
Дж.Эндрюс, Дж.Д. Кресслер, W.-M.L. Куо, К. Гренс, Т. Тривикраман и С. Филлипс, «Усилитель мощности X-BandSiGe мощностью 850 мВт», Труды конференции IEEE Bipolar / BiCMOS, 2008 г., , стр. 109-112, 2008 г.
З. Сю, Х. Вэй, Г. Ниу, Л. Луо, Д. Томас и Дж. Д. Кресслер, «Моделирование температурно-зависимых характеристик IC-VBE SiGe HBT в диапазоне 43–400K», Труды 2008 IEEE Bipolar / Совещание по схемам и технологиям BiCMOS , стр. 81-84, 2008 г.
T.K. Тривикраман, W.-M.L. Куо, Дж. П. Комо и Дж. Д. Кресслер, «Влияние масштабирования технологических узлов на ВЧ-переключатели nMOS SPDT», Технический дайджест Европейского симпозиума IEEE по микроволновому излучению, 2008 г., стр. 374-377, 2008 г.
К. Паттерсон, С. Хорст, С. Бхаттачарья, Дж. Д. Кресслер и Дж. Папаполимеру, «Недорогая органическая упаковка для беспроводных систем миллиметрового диапазона на основе кремния», Технический сборник Европейского симпозиума по микроволновому излучению IEEE 2008 г. , стр. 1242-1245, 2008.
М. Беллини, Дж. Д. Кресслер, М. Туровски, Г. Авенье, А. Шантр и П. Шевалье, «Трехмерный региональный анализ времени прохождения SiGe HBT на тонкопленочных КНИ», Труды Электрохимического общества 2008 г. Симпозиум по SiGe и Ge: материалы, обработка и устройства , vol. 16 (10), стр. 1079-1088, 2008.
A.S. Киз, Дж. Адамс, М. Патрик, М.А.Джонсон и Дж.Д. Кресслер, «Обзор проекта НАСА по радиационно-стойкой электронике для космической среды», Proceedings of the AIAA Space Conference 2008 , pp.1-7, 2008.
К. Улаганатан, Н. Намбиар, Б. Протро, Р. Гринвелл, С. Чен, Х. Хоанг, Р. Бротон, К. Корнетт, Г. Фу, Б. Дж. Блэлок, К. Бриттон-младший, M.N. Эриксон, Х.А. Мантуз, Дж. Д. Кресслер и Р. В. Бергер, «Инструментальный канал SiGe BiCMOS для приложений в экстремальных условиях», Труды симпозиума IEEE Midwest по схемам и системам , стр. 217-220, 2008 г.
М. Уллан, Дж. Райс, Г. Бройманс, Дж.Д. Кресслер, Д. Дамиани, С. Диес, Т. Гадфорт, А.А. Грилло, Р. Хакенбург, Г. Хэйр, А. Джонс, Дж. Кирстед, В. Коннененко, И. Мандич, Ф. Мартинес-МакКинни, Дж. Меткалф, Ф. М. Ньюкомер, Дж. А. Парсонс, С. Филлипс, С. Решиа, HF-W. Садрозинский, А. Сейден, Э. Спенсер, Х. Шпилер, А. К. Саттон, Ю. Тазава, М. Уайлдер и Э. Вульф, «Оценка двух технологий SiGe HBT для обновления ATLAS sLHC», Proceedings of Тематический семинар 2008 г. по электронике для физики элементарных частиц , стр. 111-115, 2008 г.
J.S. Райс, М. Уллан, Г.Brooijmans, J. D. Cressler, D. Damiani1, S. Diez, T. Gadfort, A.A. Грилло, Р. Хакенбург, Дж. Хэйр, А. Джонс, Дж. Кирстед, В. Кононенко, И. Мандич, Ф. Мартинес-МакКинни, Дж. Меткалф, новичок в области FM, Дж. А. Парсонс, С. Филлипс, С. Решиа, HF FW. Садрозинский, А. Сейден, Н. Спенсер, Х. Шпилер, А.К. Саттон, Ю. Тазава, Э. Вульф, М. Уайлдер, Характеристики технологий SiGe HBT 8HP и 8WL после облучения высокой дозой / флюенсом », Proceedings of симпозиум IEEE по ядерной науке и конференция по медицинской визуализации , стр.2206-2210, 2008.
А. Мадан, С.Д. Филлипс, Дж.Д. Кресслер, П.В. Маршалл, К. Лян и Г. Фриман, «Влияние протонного облучения на радиочастотные характеристики 65-нм КМОП-технологии», Труды конференции IEEE Radiation Effects on Components and Systems (RADECS) 2008 г., , стр. 47- 52, 2008.
2007 Документы
W.-M.L. Куо, Дж. П. Комо, Дж. М. Эндрюс, Дж. Д. Кресслер и М. А. Митчелл, «Сравнение однополюсных двухпозиционных переключателей на nMOSFET шунтирующих и последовательных / шунтирующих nMOSFET для модулей фазированной решетки X-диапазона T / R», Труды 2007 IEEE Тематическое совещание по кремниевым монолитным интегральным схемам в радиочастотных системах , стр.249-252, 2007.
С. Венкатараман, Б. Банерджи, С.-Х. Ли, Дж. Ласкар, Дж. Папаполимеру и Дж. Д. Кресслер, «Криогенная работа с малым сигналом для полевых МОП-транзисторов 0,18 мкм», Труды тематического совещания IEEE 2007 г. по кремниевым монолитным интегральным схемам в радиочастотных системах , стр. 52-55, 2007.
Я. Цуй, Г. Ниу, Й. Ли, С. С. Тейлор, К. Лян и Дж. Д. Кресслер, «О избыточных шумовых факторах и уравнениях шумовых параметров для РЧ КМОП», Труды тематического совещания 2007 IEEE по кремниевым монолитам. Интегральные схемы в радиочастотных системах , стр.40-43, 2007.
M.L. Аллес, Р.А. Рид, А. Калавагунта, Б. Серавски, Я. Пеллиш, Э. Монтес, Б. Блалок, Л. Пельц, Дж. Д. Кресслер, П. У. Маршалл и Дж. Ниу, «Влияние холодных температурных сред на единичные радиационные эффекты», Труды 2007 GOMAC-Tech — Конференция по правительственным применениям микросхем и критическим технологиям , стр. 429-432, 2007.
П. Ченг, К. Чжу, Дж.Д. Кресслер и А.Дж. Джозеф, «Спектр повреждений SiGe HBT в смешанном режиме», Технический сборник Международного симпозиума IEEE по физике надежности, 2007 г., , стр.566-567, 2007.
Дж. П. Комо, Дж. Д. Кресслер, М. Митчелл, Н. Кинайман и Ж.-П. Лантери, «Методы проектирования и компоновки для оптимизации шунтирующих SPDT-переключателей серии nMOS в 130-нм технологии SiGe BiCMOS», Proceedings of the 2007 IEEE MTT-S RFIC Symposium , pp. 457-460, 2007.
T.K. Тривикраман, W.-M.L. Куо, Дж.П. Комо, А.К. Саттон, Дж.Д. Кресслер, П.В. Маршалл и М.А. Митчелл, «Коэффициент шума 2 мВт, менее 2 дБ, SiGe-малошумящий усилитель для высокогорных или космических радиолокационных приложений X-диапазона», Труды симпозиума 2007 IEEE MTT-S RFIC , стр.629-632, 2007.
Дж. Эндрюс, Дж. Д. Кресслер и М. Митчелл, «Двухкаскадный SiGe-усилитель мощности X-диапазона с высоким коэффициентом усиления», Труды Международного симпозиума IEEE MTT-S 2007 г., посвященного микроволновому излучению , стр. 817-820, 2007.
М. Беллини, П. Ченг, А. Аппасвами, Дж. Д. Кресслер и Дж. Кай, «1 / f-шум в SiGe HBT, изготовленных на CMOS-совместимой тонкой пленке SOI», Труды Четвертого Международного симпозиума SPIE по шуму и колебания в схемах, устройствах и материалах , vol.6600, с. 0х2-0Н9, 2007.
Ченг, Э. Чжао, Дж. Д. Кресслер и Дж. Прасад, «О механизмах низкочастотного шума в вертикальных кремниевых транзисторах pnp BJT», Труды четвертого Международного симпозиума SPIE по шуму и колебаниям в схемах, устройствах, и Материалы , т. 6600, стр. 0C1-0C9, 2007.
А. Аппасвами, Дж. Д. Кресслер и Г. Ниу, «Новый феномен базового тока в SiGe HBT, работающих в инверсном режиме», Труды Европейской конференции по исследованиям твердотельных устройств IEEE 2007 г. , стр.350-353, 2007.
C.M. Гренс, Дж.Д. Кресслер и А.Дж. Джозеф, «Характеристики больших сигналов, линейность и надежность каскодных SiGe HBT с агрессивным смещением для приложений усилителя мощности», Труды 2007 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp. 135-138, 2007.
Дж. П. Комо, М. А. Мортон, W.-M.L. Куо, Т. Тривикраман, Дж.М. Эндрюс, К. Гренс, Дж. Д. Кресслер, Дж. Папаполимеру и М. Митчелл, «Монолитный 5-битный SiGe BiCMOS-приемник для радиолокационных систем с фазированной решеткой X-диапазона», Труды 2007 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp. .172-175, 2007.
М. Беллини, Дж. Д. Кресслер и Дж. Кай, «Оценка высокотемпературных возможностей SiGe HBT, изготовленных на CMOS-совместимой тонкой пленке SOI», Proceedings of the 2007 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp. . 234-237, 2007.
.J. Yuan, R. Krithivasan, J.D. Cressler, M.H. Хатер, Д.К. Альгрен и А.Дж. Джозеф, «О частотных пределах SiGe HBT для приложений TeraHertz», Труды 2007 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , pp.22-25, 2007.
Л. Луо, Г. Ню и Дж. Д. Кресслер, «Моделирование сужения запрещенной зоны для согласованного моделирования SiGe HBT в широком диапазоне температур», Труды конференции IEEE Bipolar / BiCMOS, 2007 г., , стр. 123- 126, 2007.
С. Финн, Дж. Юань, Р. Критивасан, Л. Наджафизаде, П. Ченг и Дж. Д. Кресслер, «Приемопередатчик SiGe BiCMOS 10 Мбит / с для работы вплоть до криогенных температур», Труды 2007 IEEE Bipolar / BiCMOS Circuits and Technology Meeting , стр.115-118, 2007.
A. Sutton, K. Moen, J.D. Cressler, M.A. Carts, P.W. Маршалл, Дж. Пеллиш, Р.А. Рид, М. Аллес и Г. Ниу, «Протонно-индуцированный SEU в цифровой логике SiGe при криогенных температурах», Труды Международного симпозиума по исследованию полупроводниковых устройств IEEE 2007 г. , статья WP6-04, стр. 1-2, 2007 г.
Б. Джун, Н. Меррет, С. Филлипс, А.К. Саттон, Дж.Д. Кресслер, Дж. Уильямс и П.В. Маршалл, «Сравнение эффектов протонного излучения с энергией 63 МэВ и рентгеновского излучения с энергией 10 кэВ в вертикальных желобчатых полевых транзисторах с режимом истощения 4H-SiC», Труды Международного симпозиума по исследованиям полупроводников IEEE 2007 г. , статья WP6-01, стр.1-2, 2007.
А. Раман, М. Туровски, А. Федосеев и Дж. Д. Кресслер, «Решение проблем моделирования цепей устройств для экстремальных условий космоса», Труды Международного симпозиума по исследованию полупроводниковых устройств IEEE 2007 г. , документ WP2-02, С. 1-2, 2007.
B.O. Вудс, Х. Мантуз и Дж.Д. Кресслер, «Компактное моделирование SiGe HBT для экстремальных температур», Труды Международного симпозиума по исследованиям полупроводников IEEE 2007 г. , статья WP2-05, стр.1-2 , 2007.
М. Митчелл и Дж. Д. Кресслер, «Модуль передачи / приема X-Band SiGe Single-MMIC для радиолокационных приложений», Proceedings of the 2007 IEEE Radar Conference , pp. 664-669, 2007.
A.S. Киз, Дж. Адамс, Д. Фрейзер, М. Патрик, М.Д.Ватсон, М.А.Джонсон, Дж.Д. Кресслер и Э.А. Колава, «Разработки радиационно-стойкой электроники, применимые к видению для исследования космоса», Труды конференции AIAA Space 2007 , стр.1-10, 2007.
2006 — 1986 Документы
Введение в нейроны и нейронные сети | Раздел 1, Вступительная глава | Neuroscience Online: Электронный учебник для неврологии | Кафедра нейробиологии и анатомии
Три фунта желеобразного материала, обнаруженные в наших черепах, — это самая сложная машина на Земле и, возможно, во Вселенной. Его феноменальные свойства были бы невозможны без сотен миллиардов нейронов, из которых он состоит, и, что важно, без связей между этими нейронами.К счастью, многое известно о свойствах отдельных нейронов и простых нейронных сетей, и аспекты сложных нейронных сетей начинают разгадываться. Эта глава начнется с обсуждения нейрона, элементарного узла или элемента мозга, а затем перейдет к обсуждению способов, которыми отдельные нейроны общаются друг с другом. Что делает нервную систему таким фантастическим устройством и отличает мозг от других органов тела, так это не то, что она имеет 100 миллиардов нейронов, а то, что нервные клетки способны взаимодействовать друг с другом таким высокоструктурированным образом, что формируют нейронные сети. .Чтобы понять нейронные сети, необходимо понять способы, которыми один нейрон взаимодействует с другим через синаптические связи, и процесс, называемый синаптической передачей. Синаптическая передача бывает двух основных видов: возбуждение и торможение. Всего несколько взаимосвязанных нейронов (микросхема ) могут выполнять сложные задачи, такие как опосредовать рефлексы, обрабатывать сенсорную информацию, генерировать движения и опосредовать обучение и память. Более сложные сети ( макросхем, ) состоят из множества встроенных микросхем.Макросхемы опосредуют высшие функции мозга, такие как распознавание объектов и познание. Итак, в нервной системе повсеместно встречаются сети нескольких уровней. Сети также преобладают в пределах нейронов. Эти наноцепей 90–150 составляют базовый биохимический механизм для обеспечения ключевых свойств нейронов, таких как обучение и память, а также генезиса ритмичности нейронов. Нейрон
Основные морфологические особенности нейронов
100 миллиардов нейронов в головном мозге имеют ряд общих черт (рис. 1).Нейроны отличаются от большинства других клеток тела тем, что они поляризованы и имеют различные морфологические области, каждая из которых выполняет определенные функции. Дендриты — это область, где один нейрон получает связи от других нейронов. Тело клетки или сома содержит ядро и другие органеллы, необходимые для клеточной функции. Аксон является ключевым компонентом нервных клеток, по которому информация передается от одной части нейрона (например,g., тело клетки) к концевым участкам нейрона. Аксоны могут быть довольно длинными, достигая метра или около того в некоторых сенсорных и двигательных нервных клетках человека. Синапс является конечной областью аксона, и именно здесь один нейрон образует соединение с другим и передает информацию посредством процесса синаптической передачи . Нейрон цвета морской волны на рисунке 1 (щелкните «Нейрон, связанный с постсинаптическим нейроном») называется постсинаптическим нейроном.Терминал коричневого цвета слева, следовательно, называется пресинаптическим нейроном. Один нейрон может получать контакты от множества разных нейронов. На рисунке 1 (щелкните «Нейрон, получающий синаптический ввод») показан пример трех пресинаптических нейронов, контактирующих с одним постсинаптическим нейроном коричневого цвета, но было подсчитано, что один нейрон может получать контакты до 10 000 других клеток. Следовательно, потенциальная сложность сетей огромна. Точно так же любой нейрон может контактировать до 10 000 постсинаптических клеток.(Обратите внимание, что нейрон коричневого цвета, который был пресинаптическим по отношению к нейрону цвета морской волны, является постсинаптическим по отношению к розовым, зеленым и синим нейронам. Таким образом, большинство «пресинаптических» нейронов являются «постсинаптическими» по отношению к некоторым другим нейронам.
Рисунок 1 (щелкните «Синапс») также показывает расширенный вид синапса. Обратите внимание, что пресинаптическая клетка не связана напрямую с постсинаптической клеткой. Они разделены промежутком, известным как синаптическая щель . Следовательно, чтобы общаться с постсинаптической клеткой, пресинаптический нейрон должен выпускать химический мессенджер.Этот мессенджер находится в везикулах, содержащих нейротрансмиттер , (синие точки обозначают нейромедиатор). Потенциал действия, который проникает в пресинаптический терминал, заставляет эти везикулы сливаться с внутренней поверхностью пресинаптической мембраны и высвобождать их содержимое посредством процесса, называемого экзоцитозом . Освободившийся передатчик диффундирует через промежуток между пре- и постсинаптической клеткой и очень быстро достигает постсинаптической стороны синапса, где связывается со специализированными рецепторами, которые «распознают» передатчик.Связывание с рецепторами приводит к изменению проницаемости ионных каналов в мембране и, в свою очередь, к изменению мембранного потенциала постсинаптического нейрона, известного как постсинаптический синаптический потенциал (PSP) . Таким образом, передача сигналов между нейронами связана с изменениями электрических свойств нейронов. Чтобы понять нейроны и нейронные цепи, необходимо понимать электрические свойства нервных клеток.
Потенциал покоя и потенциал действия
Потенциалы покоя. На рисунке 2 показан пример идеализированной нервной клетки. Во внеклеточную среду помещен микроэлектрод. Микроэлектрод — это не что иное, как небольшой кусок стеклянной капиллярной трубки, который растягивается под действием тепла, образуя очень тонкий наконечник, порядка 1 микрона в диаметре. Микроэлектрод заполняется проводящим раствором и затем подключается к подходящему записывающему устройству, например, осциллографу или самописцу. Когда электрод находится вне клетки во внеклеточной среде, регистрируется нулевой потенциал, поскольку внеклеточная среда является изопотенциальной.Однако, если электрод проникает в ячейку так, что кончик электрода теперь находится внутри ячейки, на записывающем устройстве наблюдается резкое отклонение. Регистрируется потенциал около -60 милливольт внутри отрицательного по отношению к внешней стороне. Этот потенциал называется потенциалом покоя и остается постоянным в течение неопределенных периодов времени при отсутствии какой-либо стимуляции. Если электрод вынуть, снова регистрируется нулевой потенциал. Потенциалы покоя — это не просто характеристики нервных клеток; все клетки тела обладают потенциалом покоя.Что отличает нервные клетки и другие возбудимые мембраны (например, мышечные клетки), так это то, что они способны изменять свой потенциал покоя. В случае нервных клеток для интеграции информации и передачи информации, а в случае мышечных клеток — для производства мышечных сокращений.
Потенциалы действия. На рисунке 3 показан еще один эскиз идеализированного нейрона. Этот нейрон был пронзен одним электродом для измерения потенциала покоя и вторым электродом, называемым стимулирующим электродом.Стимулирующий электрод подключается к батарее через переключатель. Если батарея ориентирована так, что положительный полюс подсоединен к переключателю, замыкание переключателя сделает внутреннюю часть элемента несколько более положительной в зависимости от размера батареи. (Такое уменьшение поляризованного состояния мембраны называется деполяризацией 90–150 .) На рис. 3 показана анимация, в которой переключатель многократно размыкается и замыкается, и каждый раз, когда он замыкается, в схему подключается батарея большего размера.Первоначально замыкание переключателя вызывает только небольшие деполяризации. Однако потенциалы становятся больше, и в конечном итоге деполяризация становится достаточно большой, чтобы вызвать потенциал действия , также известный как спайк или импульс . Потенциал действия связан с очень быстрой деполяризацией, позволяющей достичь пикового значения около +40 мВ всего за 0,5 миллисекунды (мс). За пиком следует столь же быстрая фаза реполяризации .
Напряжение, при котором деполяризация становится достаточной для запуска потенциала действия, называется порогом .Если для генерации надпороговой деполяризации используется батарея большего размера, единичный потенциал действия по-прежнему генерируется, и амплитуда этого потенциала действия такая же, как потенциал действия, запускаемый только пороговым стимулом. Простая запись на рисунке 3 иллюстрирует две очень важные особенности потенциалов действия. Во-первых, они вызываются по схеме «все или ничего» . Либо потенциал действия вызывается стимулами на уровне порога или выше, либо потенциал действия не вызывается.Во-вторых, потенциалы действия — это очень короткие события продолжительностью всего несколько миллисекунд. Запуск потенциала действия в некоторой степени аналогичен подаче спички на предохранитель. Для воспламенения предохранителя необходима определенная температура (т.е. предохранитель имеет порог). Спичка, выделяющая большее количество тепла, чем пороговая температура, не приведет к тому, что предохранитель будет гореть ярче или быстрее. Так же, как потенциалы действия вызываются по принципу «все или ничего», они также распространяются по принципу «все или ничего».Как только потенциал действия инициируется в одной области нейрона, такой как тело клетки, этот потенциал действия будет распространяться по аксону (как горящий предохранитель) и в конечном итоге проникнет в синапс, где он может инициировать процесс синаптической передачи.
В примере на Рисунке 3 был сгенерирован только один потенциал действия, потому что продолжительность каждого из двух надпороговых стимулов была настолько короткой, что было достаточно времени, чтобы инициировать только один потенциал действия (т.е.е. стимул закончился до того, как потенциал действия завершил свой цикл деполяризации-реполяризации). Но, как показано в анимации на Рисунке 4, более продолжительные стимулы могут привести к инициированию множественных потенциалов действия, частота которых зависит от интенсивности стимула. Таким образом, очевидно, что нервная система кодирует информацию не с точки зрения изменения амплитуды потенциалов действия, а с точки зрения их частоты. Это очень универсальное свойство.Чем выше интенсивность механического стимула к сенсорному рецептору, тем больше количество потенциалов действия; чем сильнее растягивается рецептор растяжения мышцы, тем больше количество потенциалов действия; чем больше интенсивность света, тем большее количество потенциалов действия передается в центральную нервную систему. Точно так же в двигательной системе, чем больше количество потенциалов действия в двигательном нейроне, тем сильнее будет сокращение мышцы, которая получает синаптическую связь от этого двигательного нейрона.Инженеры называют этот тип кодирования информации частотно-импульсной модуляцией .
Синаптические потенциалы и синаптическая интеграция
На рисунке 5 показаны три нейрона. Зеленый цвет будет называться возбуждающим нейроном по причинам, которые вскоре станут ясны. Он соединяется с постсинаптическим нейроном синего цвета. Следы ниже (нажмите «Play») иллюстрируют последствия запуска потенциала действия в зеленом нейроне.Этот потенциал действия в пресинаптическом нейроне приводит к снижению мембранного потенциала постсинаптической клетки. Мембранный потенциал изменяется от значения покоя около -60 милливольт до более деполяризованного состояния. Этот потенциал называется возбуждающим постсинаптическим потенциалом (ВПСП) . Он является «возбуждающим», потому что перемещает мембранный потенциал к пороговому значению, и является «постсинаптическим», потому что это потенциал, зарегистрированный на постсинаптической стороне синапса.Обычно (и это важный момент) единичный потенциал действия в пресинаптической клетке не производит ВПСП, достаточно большого, чтобы достичь порога и запустить потенциал действия. Но если в пресинаптической клетке запускаются множественные потенциалы действия, соответствующие множественные возбуждающие потенциалы могут суммироваться посредством процесса, называемого временным суммированием , чтобы достичь порога и запустить потенциал действия. ВПСП можно рассматривать как «сигнал движения» постсинаптическому нейрону для передачи информации по сетевому пути.
Нейрон красного цвета на рисунке 5 называется тормозным нейроном. Как и зеленый нейрон, он также устанавливает синаптический контакт с синим постсинаптическим нейроном. Он также высвобождает химический передатчик-мессенджер, но последствия связывания передатчика от голубой клетки с рецепторами на постсинаптической клетке противоположны последствиям передатчика, выпущенного зеленым нейроном. Следствием потенциала действия красного пресинаптического нейрона является увеличение мембранного потенциала синего постсинаптического нейрона.Мембранный потенциал более отрицательный, чем был раньше (гиперполяризация ), и поэтому мембранный потенциал дальше от порогового значения. Этот тип потенциала называется ингибирующим постсинаптическим потенциалом (IPSP) , потому что он предотвращает запуск постсинаптического нейрона потенциала действия. Это «стоп-сигнал» для постсинаптической клетки. Таким образом, зеленый нейрон говорит «вперед», а красный нейрон говорит «стоп». Что же делать постсинаптическому нейрону?
Нейроны похожи на счетные машины.Они постоянно суммируют возбуждающий и тормозной синаптический вход во времени (временное суммирование) и по площади дендритов, получающих синаптические контакты ( пространственное суммирование) 90-150, и если это суммирование находится на пороговом уровне или выше, они запускают потенциал действия. Если сумма ниже порога, потенциал действия не инициируется. Этот процесс, называемый синаптической интеграцией , проиллюстрирован на рисунке 5. Первоначально два потенциала действия в зеленом нейроне производили суммирующие ВПСП, которые запускали потенциал действия в синем нейроне.Но если IPSP от тормозного нейрона возникает непосредственно перед двумя потенциалами действия в возбуждающем нейроне, сумма одного IPSP и двух EPSP ниже порогового значения, и в постсинаптической клетке не возникает никакого потенциала действия. Тормозящий нейрон (и ингибирование в целом) — это способ стробирования или регулирования способности возбуждающего сигнала запускать постсинаптическую клетку.
Нейронные сети
Мотивы микросетей
Как указывалось ранее в этой главе, нейрон может получать контакты до 10 000 пресинаптических нейронов, и, в свою очередь, любой нейрон может контактировать до 10 000 постсинаптических нейронов.Комбинаторная возможность может привести к чрезвычайно сложным нейронным цепям или сетевым топологиям , которые могут быть очень трудными для понимания. Но, несмотря на потенциально огромную сложность, многое можно узнать о функционировании нейронных цепей, исследуя свойства подмножества простых конфигураций цепей. На рисунке 6 показаны некоторые из этих мотивов микросхем или микросетей. Несмотря на простоту, они могут делать многое из того, что должна делать нервная система.
Возбуждение с прямой связью . Позволяет одному нейрону передавать информацию своему соседу. Длинные цепочки из них могут использоваться для передачи информации через нервную систему.
Ингибирование с прямой связью . Пресинаптическая клетка возбуждает тормозящий интернейрон (интернейрон — это нейрон, расположенный между двумя нейронами), и этот тормозной интернейрон затем подавляет следующую клетку-последователь.Это способ отключения или ограничения возбуждения в нейроне, расположенном ниже по течению в нейронной цепи.
Схождение / расхождение . Одна постсинаптическая клетка получает конвергентный вход от ряда различных пресинаптических клеток, и любой отдельный нейрон может создавать дивергентные связи со многими разными постсинаптическими клетками. Дивергенция позволяет одному нейрону общаться со многими другими нейронами в сети. Конвергенция позволяет нейрону получать входные данные от многих нейронов в сети.
Боковое ингибирование . Пресинаптическая клетка возбуждает тормозящие интернейроны, и они подавляют соседние клетки в сети. Как подробно описано ниже в этой главе, этот тип схемы может использоваться в сенсорных системах для улучшения контуров.
Запрет обратной связи / повторения . На панели E1 пресинаптическая клетка соединяется с постсинаптической клеткой, а постсинаптическая клетка, в свою очередь, соединяется с интернейроном, который затем подавляет пресинаптическую клетку.Этот контур может ограничивать возбуждение в пути. Некоторое начальное возбуждение будет отключено после того, как красный интернейрон станет активным. На панели E2 каждый нейрон в замкнутой цепи подавляет нейрон, с которым он связан. Казалось бы, эта схема ничего не делает, но, как будет показано далее в этой главе, она может привести к генерации сложных паттернов всплесковой активности.
Обратная связь / рекуррентное возбуждение . На панели F1 пресинаптический нейрон возбуждает постсинаптический нейрон, а этот постсинаптический нейрон возбуждает пресинаптический нейрон.Этот тип цепи может выполнять функцию переключателя, потому что, как только пресинаптическая клетка активируется, эта активация может сохраняться. Активация пресинаптического нейрона могла включить эту сеть, и она могла остаться. На панели F2 показаны варианты возбуждения обратной связи, при которых пресинаптический нейрон возбуждает постсинаптический нейрон, который может по обратной связи возбуждать сам себя (а, аутапс 90–150), или другие нейроны, которые в конечном итоге имеют обратную связь (b) сам с собой.
Эти простые мотивы — вездесущие компоненты многих нейронных цепей.Давайте рассмотрим несколько примеров того, что могут делать эти сети.
Возбуждение с прямой связью и подавление с прямой связью
Одна из наиболее изученных микросхем — это схема, которая обеспечивает простое рефлекторное поведение. На рисунке 7 показана схема так называемого рефлекса рефлекса коленного рефлекса или растяжения. Невролог ударяет резиновым молоточком по колену, в результате чего нога разгибается. Этот тест используется как простой способ проверить целостность некоторых сенсорных и моторных путей в спинном мозге.Удар молотка растягивает мышцу и приводит к инициированию потенциалов действия в сенсорных нейронах, 90–150 внутри мышцы, которые чувствительны к растяжению. (Потенциалы действия представлены маленькими яркими «огоньками» на анимации.) Потенциалы действия инициируются по принципу «все или ничего» и распространяются в спинной мозг, где аксон разделяется (раздваивается) на две ветви.
Давайте сначала обсудим ветвь слева, которая формирует синаптическую связь (зеленый треугольник) с Extensor (E) мотонейроном (окрашен в синий цвет).Потенциал действия сенсорного нейрона вторгается в синаптический терминал сенсорного нейрона, вызывая высвобождение медиатора и последующее возбуждение двигательного нейрона. Растяжение мышцы приводит к потенциалу действия в двигательном нейроне (МН), который затем распространяется по периферическому нерву и вторгается в синапс в мышце, вызывая высвобождение передатчика и потенциал действия в мышце. Потенциал действия в мышечной клетке приводит к сокращению мышцы и разгибанию конечности.Итак, здесь у нас есть простая схема возбуждения с прямой связью, которая опосредует поведение.
Теперь давайте рассмотрим правую ветвь аксона сенсорного нейрона на рисунке 7. Потенциал действия сенсорного нейрона вторгается в синаптический терминал сенсорного нейрона, вызывая высвобождение медиатора и последующее возбуждение постсинаптического интернейрона, окрашенного в черный цвет. Этот нейрон называется интернейроном , потому что он расположен между одним нейроном (здесь SN) и другим нейроном (здесь MN).Возбуждение интернейрона приводит к инициированию действия и последующему высвобождению передатчика из пресинаптического терминала интернейрона (черный треугольник), но для этой ветви цепи передатчик ведет к IPSP в постсинаптическом сгибателе (F ) мотонейрон (окрашен в красный цвет). Функциональные последствия этого подавления прямой связи заключаются в уменьшении вероятности того, что двигательный нейрон-сгибатель станет активным и вызовет несоответствующее сгибание ноги.
Сходимость и расхождение
Упрощенная схема, опосредующая рефлекс растяжения, представлена на рисунке 8. Однако правильное функционирование схемы рефлекса растяжения также зависит от конвергенции и расхождения. Один сенсор имеет несколько ветвей, которые расходятся и образуют синаптические связи со многими отдельными моторными нейронами (щелкните «Дивергенция»). Следовательно, когда мышца сокращается в результате нажатия невролога, это происходит потому, что несколько мышечных волокон активируются одновременно несколькими двигательными нейронами.Кроме того, когда мышца растягивается, активируется не один, а несколько сенсорных нейронов, и все эти сенсорные нейроны проецируются в спинной мозг, где они сходятся к отдельным моторным нейронам-разгибателям (нажмите «Конвергенция»). Итак, рефлекс растяжения возникает из-за комбинированных эффектов активации нескольких сенсорных нейронов и моторных нейронов-разгибателей.
Боковое торможение
Улучшение края.Боковое торможение очень важно для обработки сенсорной информации. Одним из примеров является явление в зрительной системе, называемое усилением краев. На рисунке 9 показаны две полосы: темно-серая полоса слева и светло-серая полоса справа. Хотя темная полоса и светлая полоса имеют одинаковую яркость по всему полю, при внимательном рассмотрении обнаруживается, что светло-серая полоса кажется несколько светлее на границе темно-серой полосы, чем в других областях поля.Напротив, темно-серая полоса на границе выглядит несколько темнее, чем в других областях темного поля. Это явление улучшения границ, которое помогает визуальной системе извлекать важную информацию из визуальных сцен. Усиление краев опосредуется, по крайней мере частично, боковым торможением в сетчатке.
Давайте сначала рассмотрим схему без бокового торможения (Рисунок 10, щелкните «Без бокового торможения»). Свет падает на сетчатку (Часть A), а его интенсивность можно описать ступенчатым градиентом (Часть B).Для упрощения предположим, что темно-серая область имеет интенсивность пяти единиц, а светло-серая область — десять единиц. Градиент света активирует фоторецепторы, и фоторецепторы создают синаптические связи с нейронами второго порядка. Предположим, что интенсивность света 5 единиц приводит к 5 всплескам / с, а интенсивность света 10 единиц приводит к 10 всплескам / с (Часть C) в фоторецепторах, и что синаптическая сила достаточна (здесь обозначена как +1) так что интенсивность света 5 единиц приводит к 5 импульсам / с, а интенсивность света 10 единиц приводит к 10 импульсам / с (Часть C) соответственно в нейронах второго порядка.Если никакой дальнейшей обработки информации не происходит, воспринимаемый градиент будет точно таким же, как градиент интенсивности света (часть B, красный след). Но это не то, что воспринимается, и латеральное торможение объясняет разницу.
Теперь рассмотрим расширенную схему с боковым торможением (нажмите «С боковым торможением»). Каждый из фоторецепторов устанавливает тормозные синаптические связи со своим соседним нейроном второго порядка. Сила подавления (обозначается -0.2) меньше силы возбуждения (пожертвовано +1). Прежде чем смотреть на границу, рассмотрите выход схемы в однородных областях каждого поля. Далеко справа от границы все клетки получают одинаковое возбуждение и одинаковое торможение. Без бокового торможения интенсивность света в 10 единиц произвела бы 10 импульсов в секунду в нейроне второго порядка. Но из-за тормозящей связи от соседних нейронов справа и слева выходная мощность снижается до 6 импульсов в секунду.То же самое верно для клеток, расположенных далеко слева от границы, но величина возбуждения меньше и, соответственно, величина торможения меньше. Ключевая обработка происходит на границе или краю. Обратите внимание, что нейрон справа от границы получает такое же подавление от нейрона справа от нее, но получает меньшее подавление от нейрона слева от него на другой стороне границы. Следовательно, он получает больше чистого возбуждения и имеет выходной сигнал 7 пиков / с, а не 6 пиков / с его соседа справа.Теперь посмотрите на нейрон слева от границы. Он получает слабое торможение от своего соседа слева, но более сильное торможение от своего соседа справа по другую сторону границы. Следовательно, он получает меньше чистого возбуждения и имеет выходную мощность 2 пика / с, а не 3 пика / с его соседа слева. Таким образом, в результате бокового торможения информация, передаваемая нервной системе, и воспринимаемый градиент будут версией исходной с усиленной границей или краем (рис. 10B)!
диапазонов Маха.Простая схема сетчатки с боковым торможением может объяснить феномен усиления краев. Этим также можно объяснить визуальную иллюзию, известную как полосы Маха. На рисунке 11 показан градиент светлых и темных вертикальных полос, поперек которых проходит тонкая горизонтальная линия. Похоже, что горизонтальная линия имеет неравномерное распределение интенсивности: она темнее в области светлых вертикальных градиентов и светлее в области темных вертикальных градиентов. Это визуальная иллюзия .Иллюзию можно раскрыть, поместив маску поверх вертикального градиента. (Нажмите «Play», чтобы добавить маску.) Теперь вы можете видеть, что горизонтальная полоса имеет равномерную интенсивность. В некоторых областях он воспринимается более темным, потому что клетки сетчатки, которые реагируют на более темную область горизонтальной полосы, сильно подавляются клетками, реагирующими на яркую область вертикальной полосы. Напротив, полоса воспринимается ярче в некоторых областях, потому что клетки сетчатки, которые реагируют на более светлую область горизонтальной полосы, только слабо ингибируются клетками, реагирующими на темную область вертикальной полосы.
Обратная связь / повторное запрещение
Запрет обратной связи в микросхемах. Подавление обратной связи играет общую роль в гашении возбуждения через нейронную цепь. Классическим примером является клетка Renshaw в спинном мозге. Аксон ветви спинномозгового мотонейрона. Одна ветвь иннервирует мышцу, как описано ранее (например, рис. 7), а другая ветвь создает возбуждающую синаптическую связь с интернейроном, называемым клеткой Реншоу.Интернейрон, в свою очередь, подавляет двигательный нейрон, замыкая петлю. Другой пример подавления обратной связи обнаружен в гиппокампе. Пирамидные клетки СА3-типа создают возбуждающие связи с клетками корзины и обратную связь с клетками корзины, чтобы ингибировать клетки СА3. Термин повторяющееся ингибирование применяется к простым схемам подавления обратной связи, таким как цепь Реншоу в спинном мозге и цепь корзиночных клеток в гиппокампе.
Подавление обратной связи в наноцепях.Подавление обратной связи распространено не только во многих нейронных цепях; он также широко распространен в биохимических цепях. Здесь он может служить подложкой для генерации колебаний. Они могут охватывать несколько временных масштабов от секунд до дней в зависимости от молекулярных компонентов схемы.
Рис. 12. Данные Byrne, Canavier, Lechner, Clark and Baxter, 1996. |
- Эндогенное взрывное поведение в нейронах.Идеализированный нейрон, описанный ранее в этой главе, молчал в отсутствие стимуляции (например, рисунок 3). Однако некоторые нейроны активируют потенциалы действия в отсутствие стимуляции, и в некоторых случаях паттерны возбуждения могут демонстрировать взрывной паттерн, в котором последовательные высокочастотные всплески активности сменяются периодами покоя. Такие нейронные свойства могут быть важны для создания ритмического поведения, такого как дыхание. На рисунке 12 показан пример записи нейрона беспозвоночного, имеющего эндогенный импульсный ритм.Этот конкретный нейрон называется параболическим всплеском, потому что характер интервалов между всплесками является длинным в начале и в конце цикла всплеска, но очень коротким в середине цикла. Клетка запускает всплеск потенциалов действия, а затем замолкает, но вскоре происходит еще один всплеск, и этот процесс продолжается бесконечно каждые десять-пятнадцать секунд. Взрыв происходит, даже если нейрон хирургическим путем удален из ганглия и помещен в культуру, чтобы не было синаптических связей с другими нейронами.Таким образом, для этого ритма не требуется нейронная сеть — она эндогенная. Но это действительно связано с наноконтролем внутри клетки. На рисунке 13A показана очень упрощенная версия этой сети, в которой подчеркивается ключевой принцип работы. Критическим для этой сетевой функции является канал в
мембрана (обозначена g SI ), проницаемая для Ca 2+ . Поскольку концентрация Ca 2+ относительно высока во внеклеточной среде и низка внутри клетки, Ca 2+ будет двигаться вниз по градиенту концентрации и тем самым деполяризует клетку.В конце концов, деполяризация достигает порога, и клетка начинает срабатывать. Обжиг приводит к дополнительному притоку Ca 2+ (зеленая стрелка) и накоплению Ca 2+ внутри клетки. Ключевым этапом является то, что накопление Ca 2+ ингибирует (красная стрелка) дальнейший приток кальция и прекращает всплеск. Всплеск остается прекращенным, пока уровни внутриклеточного Ca 2+ остаются повышенными. Но уровни Ca 2+ не остаются повышенными надолго.Они восстанавливаются внутриклеточными буферами и удаляются из клетки с помощью насосов (I NaCa и I CaP ). Когда внутриклеточные уровни Ca 2+ снижаются, ингибирование канала снимается (растормаживание , ), и нейрон снова начинает деполяризоваться, и начинается новый всплеск. По сути, то, что мы видим здесь на уровне наносети, является повторением сети, препятствующей обратной связи (рис. 13B). Начальный процесс возбуждения приводит к активации тормозного процесса, который отключается, чтобы отключить процесс возбуждения.В такой сети возникнут колебания, если возбуждающее побуждение будет непрерывным, но тормозящий процесс ослабевает по своей эффективности.
Циркадные ритмы.Второй пример подавления обратной связи — это наноцепь регуляции генов, лежащих в основе циркадных ритмов. Циркадный ритм позвоночных обусловлен работой группы нейронов в области мозга, называемой супрахиазматическим ядром, которая расположена чуть выше зрительного нерва. Эти нейроны оказывают сильное влияние как на высвобождение гормонов, таких как мелатонин, кортизол, так и на вегетативные функции, такие как температура тела (рис. 14). Несмотря на огромное влияние этого генератора, его работа сводится к очень простой схеме, и действительно, не к нейронной схеме, а к другой наноконтуре.Основной механизм, по-видимому, сохраняется у всех видов животных, включая человека. На рисунке 15 представлена упрощенная принципиальная схема основных компонентов. Участвует несколько генов, но основной механизм включает ген под названием на , где на — на период. Этот ген был впервые идентифицирован у плодовой мухи Drosophila , но также присутствует у позвоночных. Ген на приводит к продукции на матричной РНК. на мРНК покидает ядро и попадает в цитоплазму, где приводит к синтезу белка PER.PER диффундирует или транспортируется обратно в ядро, где он репрессирует дальнейшую транскрипцию гена на . Концептуально эта система очень похожа на механизм разрывного нейрона, рассмотренный выше. Ген активируется, он производит сообщение и белок, а белковая обратная связь подавляет экспрессию гена. Но как цикл повторяется? Ключевым механизмом является деградация PER. Белок PER разлагается и разлагается в течение 24 часов.Так как белок PER разрушается, подавление или репрессия снимается (растормаживание), позволяя этому гену снова начать создавать информационную РНК и белок. Итак, как только этот цикл начинается, он повторяется снова и снова через 24 часа. Это основной механизм, лежащий в основе циркадных ритмов и мощного воздействия, которое они оказывают на ряд различных физиологических систем. По сути, все наши циркадные ритмы начинаются с сети подавления молекулярной обратной связи.Рисунок 14. По материалам Hastings et al., Nature Rev. Neurosci. , 2003.
Запрет обратной связи в кольцевых цепях.Повторяющееся торможение может, по крайней мере в принципе, объяснить генерацию сложных двигательных паттернов, примером которых является передвижение на четвероногих ногах. Четвероногие животные интересны тем, что четвероногие способны не только двигать своими четырьмя ногами, но и генерировать различные типы циклов активности, называемые походками. На рисунке 16 показаны четыре аллюра. Первая панель — это прогулка (тап по собаке). Последовательность начинается с разгибания левой передней конечности. Затем следуют разгибания правой задней конечности, правой передней конечности и левой задней конечности.На рыси (вторая панель рисунка 16) (постукивание по собаке) левая передняя и правая задние конечности находятся в фазе друг с другом и на 180 градусов не совпадают по фазе с правой передней и левой задней конечностями. В связке (третья панель) (постукивание по собаке) левая передняя и правая передние конечности находятся в фазе, но на 180 градусов не совпадают по фазе с левой задней и задней задней конечностями. Галоп (четвертая панель) (постукивание по собаке) — вариант скачек, в котором есть небольшая разница фаз между правой и левой передними и задними конечностями.
Как нервная система генерирует эти походки? И нужны ли для каждой отдельные нейронные цепи? К сожалению, нейробиологи не знают ответов на эти вопросы, но поучительно изучить некоторые возможности. Это подход в области нейробиологии под названием Вычислительная и теоретическая нейробиология . Один из способов создания походки проиллюстрирован на рисунке 17. Возьмите четыре отдельных нейрона, каждый с эндогенной импульсной активностью, подобной тому, который показан ранее на рисунке 12, и назначьте активность в каждом из этих нейронов контролю конкретной конечности.Нейроны могут быть «запущены» так, чтобы у них были соответствующие фазовые соотношения для генерации походки, такие как граница, показанная на рисунке 16. Трудность будет заключаться в том, чтобы запустить нейроны точно в точное время. Другой проблемой могут быть небольшие «дрейфы» в колебательных периодах четырех независимых нейронов, которые со временем приведут к тому, что паттерн станет несогласованным (рис. 18). Эта собака не выиграет ни одной гонки и, вероятно, не сможет ходить.
Итак, очевидно, что нейроны должны быть связаны.Один из способов сделать это — использовать повторяющуюся цепь подавления, состоящую из четырех связанных нейронов, чтобы сформировать так называемую «кольцевую» схему, где каждый нейрон в цепи имеет эндогенную импульсную активность, и каждый нейрон связан с другим с помощью тормозной синаптической связи ( Рисунок 19A).
Рис. 19. По материалам Canavier, Butera, Dror, Baxter, Clark and Byrne, 1997. |
Чтобы получить правильные фазовые соотношения для походок, вместо того, чтобы назначать нейрон 3 правую переднюю конечность, он назначается для управления правой задней конечностью, а нейрон 4 назначается для управления правой передней конечностью (a простой поворот схемы) (Рисунок 19Б).При реализации с помощью компьютерного моделирования эта единственная схема способна генерировать четвероногие ворота. Более того, одна и та же схема с небольшими изменениями свойств отдельных нейронов может генерировать каждую из четырех походок, показанных на рисунке 17 (рисунок 20).
Этот результат указывает на важный момент в отношении нейронных сетей.Чтобы понять их, необходимо понимать не только топологию сети, но и природу связей между нейронами (являются ли они возбуждающими или тормозящими), а также свойства отдельных узлов (т. Е. нейроны). Кроме того, это моделирование иллюстрирует явление, называемое динамической реконфигурацией . Для генерации этих четырех разных походок не обязательно иметь четыре разные сети — все это можно сделать с помощью одной схемы.Фактическая схема, генерирующая четырехпедальную походку, более сложна, чем схема на рис. 19. Заинтересованный читатель может отсылать к недавнему обзору Оле Кейна (см. Дополнительную литературу).
Обратная связь / рекуррентное возбуждение
Периодическое возбуждение в наноцепях и микросхемах имеет решающее значение для процессов обучения и памяти. Обучение включает в себя изменения биофизических свойств нейронов и изменения синаптической силы. Накапливающиеся данные указывают на то, что положительная обратная связь внутри биохимических каскадов и генных сетей является важным компонентом для индукции и поддержания этих изменений.Более того, периодическое возбуждение обнаруживается по крайней мере в некоторых микросхемах, участвующих в процессах памяти. Яркий пример находится в области CA3 гиппокампа.
Рисунок 21. Изменено из Byrne and Roberts, 2009. |
На рисунке 21 показаны основные характеристики рекуррентной возбуждающей цепи CA3. Шесть различных пирамидных нейронов гиппокампа обозначены как U , V , W , X , Y и Z .Каждый из этих нейронов получает синаптическую связь от пресинаптических нейронов, обозначенных a , b , c , d , e и f. Эти пресинаптические нейроны могут быть активными или неактивными с 0 и черным цветом, представляющим неактивный нейрон, и 1 и зеленым цветом, указывающим активный. Важным аспектом этой схемы является то, что синаптические связи входного пути достаточно сильны, чтобы активировать (активировать) пирамидный нейрон, с которым они связаны.Например, если нейрон a активирован, будет активирован нейрон Z , который представлен как 1 на панели вывода. Эта топология — не что иное, как возбуждение с прямой связью. Взаимное возбуждение делает эту схему особенной. Например, нейрон Z и другие пирамидные нейроны имеют коллатерали аксонов, которые посредством обратной связи соединяются друг с другом. Но они не только устанавливают связь между собой. Каждый нейрон устанавливает связь с каждым из четырех других пирамидных нейронов в цепи.Таким образом, каждая пирамида получает сходящуюся информацию от всех других клеток в сети, и, в свою очередь, выход каждого пирамидального нейрона расходится, чтобы установить синаптические связи со всеми другими пирамидными нейронами в цепи. (Следовательно, этот повторяющийся мотив возбуждения вложил в себя мотивы конвергенции и расхождения.) Матрица связности состоит из 36 элементов.
Чтобы эта сеть могла чему-либо научиться, в схему необходимо встроить правило обучения синаптической пластичности.Один из широко распространенных правил известен как «Правило обучения Хебба » . По сути, в нем говорится, что синапс изменит свою силу, если этот синапс активен (то есть высвобождает передатчик), и в то же время активна постсинаптическая клетка. Комбинация этого правила обучения и повторяющегося возбуждающего контура приводит к некоторым интересным эмерджентным свойствам. Например, если нейрон Z активирован входом a, , сила его соединения с самим собой (синапс 1 ) изменится, как показано зеленым синапсом на рисунке 21 (анимация находится в 2-секундном цикле, чтобы иллюстрируют контрольные и модифицированные синапсы).Однако синапс 1 не будет единственным синапсом, который будет усилен. Например, синапс 13 также будет усилен, потому что нейрон Z был активен в то же время, когда нейрон X был активирован вводом c . Напротив, синапс 7 не усиливается, потому что нейрон Y не был активен в то же время, что и нейрон Z . Чистый эффект этой конвергенции и расхождения и правила обучения состоит в том, что исходный входной шаблон активности будет сохранен как изменения в элементах матрицы связности.Поэтому этот канал был назван сетью с автоматическим объединением . Важная концепция здесь заключается в том, что «память» находится не в каком-то одном синапсе; это распределенных в сети .
Сводка
Значительный прогресс был достигнут в понимании того, как разные простые нейронные сети участвуют в обработке информации и опосредованном поведении. Возбуждение с прямой связью и торможение с прямой связью опосредуют рефлекторное поведение.Боковое торможение важно для усиления краев. Повторяющееся возбуждение — важный механизм памяти. Повторяющееся торможение может иметь важное значение для формирования локомоторного поведения. В этих микросхемах заложены сходимость и расхождение. Одни и те же типы сетевых мотивов воспроизводятся в биохимических и генных сетях.
Следующий уровень понимания находится на уровне нейронных сетей, которые опосредуют более сложные, так называемые функции мозга высшего порядка.Их понимание становится возможным благодаря использованию электрофизиологических и оптических методов записи и современных методов визуализации, таких как функциональная магнитно-резонансная томография (фМРТ) и диффузионно-тензорная визуализация (DTI)
. ФМРТ позволяет исследователям идентифицировать области мозга, которые заняты когнитивными задачами, тогда как DTI позволяет визуализировать пути, связывающие одну область мозга с другой. Рисунок 22 (любезно предоставлено Тимом Элмором, доктором философии, отдел.из нейрохирургии, Медицинская школа Техасского университета в Хьюстоне) — вид сбоку человеческого мозга, показывающий пути, соединяющие корковые области, выявленные с помощью DTI !.Рисунок 22. Предоставлено Тимом Элмором, доктором философии. |
Распознавание объектов — это пример прогресса в понимании макросхем.Как показано на рисунке 23, обработка визуальной информации начинается в сетчатке, а затем затрагивает несколько областей коры, таких как затылочная кора и височная кора. Внутри этой макросхемы находятся модули, извлекающие информацию более высокого порядка. Предположительно, каждый модуль включает в себя сотни, если не тысячи отдельных микросхем. Задача на будущее — определить, как эти модули работают и как они взаимодействуют с другими модулями. Хотя соединения с прямой связью присутствуют, соединения с обратной связью и боковые соединения широко распространены.Задача огромна, но, возможно, достижению цели будет способствовать использование преимуществ того, что было изучено о принципах работы наносхем и микросхем. Чтобы понять макросхемы, необходимо знать больше, чем топологию сетевых соединений. Необходимо будет знать, как работает каждый модуль, и о динамике межмодульных соединений.
Рисунок 23.Из Феллемана и Ван Эссена, 1991. |
Проверьте свои знания
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Подавление обратной связи
D. Боковое ингибирование
E.Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Ингибирование с прямой связью
C. Подавление обратной связи
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
А.Возбуждение с прямой связью
B. Запрещение прямой связи. Этот ответ НЕПРАВИЛЬНЫЙ.
C. Подавление обратной связи
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
С.Подавление обратной связи. Этот ответ НЕПРАВИЛЬНЫЙ.
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Подавление обратной связи
D. Боковое торможение. Ответ ПРАВИЛЬНЫЙ!
E.Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Подавление обратной связи
D. Боковое ингибирование
E. Расхождение. Этот ответ НЕПРАВИЛЬНЫЙ.
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А.Пластичность
B. Интеграция
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Интеграция
C. Конвергенция
Д.Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность
B. Интеграция. Этот ответ ПРАВИЛЬНЫЙ!
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А.Пластичность
B. Интеграция
C. Конвергенция Этот ответ НЕПРАВИЛЬНЫЙ.
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность
B. Интеграция
C. Конвергенция
Д.Частотно-импульсная модуляция. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность
B. Интеграция
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание Этот ответ НЕПРАВИЛЬНЫЙ.
Сетевой мотив, лежащий в основе циркадных ритмов:
А.Временное суммирование
B. Пространственное суммирование
C. Подавление обратной связи
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Пространственное суммирование
C. Подавление обратной связи
Д.Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
B. Пространственное суммирование. Этот ответ НЕПРАВИЛЬНЫЙ.
C. Подавление обратной связи
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
А.Временное суммирование
B. Пространственное суммирование
C. Запрет обратной связи. Ответ ПРАВИЛЬНЫЙ!
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
B. Пространственное суммирование
C. Подавление обратной связи
Д.Возбуждение обратной связи. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
B. Пространственное суммирование
C. Подавление обратной связи
D. Возбуждение обратной связи
E. Боковое торможение. Ответ НЕПРАВИЛЬНЫЙ.
Сеть с автоматической ассоциацией важна для:
А.Рефлексы
B. Улучшение кромки
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
A. Рефлексы. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Улучшение кромки
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А.Рефлексы
B. Улучшение кромки Этот ответ НЕПРАВИЛЬНЫЙ.
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
A. Рефлексы
B. Улучшение кромки
C. Память Этот ответ ПРАВИЛЬНЫЙ!
D. Передвижение
E.Нейронные колебания
Сеть с автоматической ассоциацией важна для:
A. Рефлексы
B. Улучшение кромки
C. Память
D. Передвижение. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
A. Рефлексы
Б.Улучшение края
C.
