Каковы основные параметры диода КД208А. Где применяется этот полупроводниковый прибор. Какие существуют отечественные и зарубежные аналоги КД208А. Как правильно выбрать и использовать данный диод в электронных схемах.
Основные характеристики диода КД208А
Диод КД208А — это кремниевый выпрямительный диод средней мощности. Он относится к классу полупроводниковых приборов и предназначен для преобразования переменного тока в постоянный. Рассмотрим основные электрические параметры КД208А:
- Максимальное постоянное обратное напряжение: 100 В
- Максимальный прямой ток: 1,5 А
- Максимальное импульсное обратное напряжение: 100 В
- Максимальный импульсный прямой ток: 30 А
- Прямое падение напряжения при Iпр = 1 А: не более 1 В
- Емкость перехода при Uобр = 4 В: не более 30 пФ
- Рабочая температура перехода: от -60°C до +130°C
Как мы видим, КД208А способен выдерживать достаточно высокие напряжения и токи, что делает его универсальным решением для многих схем выпрямления.

Области применения диода КД208А
Благодаря своим характеристикам, диод КД208А нашел широкое применение в различных электронных устройствах:
- Однополупериодные и двухполупериодные выпрямители
- Источники питания радиоаппаратуры
- Зарядные устройства для аккумуляторов
- Сварочные аппараты
- Электронные регуляторы мощности
- Схемы защиты от перенапряжений
- Импульсные источники питания
Диод КД208А особенно эффективен в маломощных выпрямительных схемах с током до 1,5 А. При необходимости работы с большими токами, диоды КД208А можно включать параллельно.
Аналоги диода КД208А
При отсутствии КД208А его можно заменить аналогичными по характеристикам диодами. Рассмотрим некоторые отечественные и зарубежные аналоги:
Отечественные аналоги:
- КД209А
- КД243А
- Д226А
- КД202А
Зарубежные аналоги:
- 1N4001
- 1N4002
- 1N5391
- FR101
- S2A
При выборе аналога важно сравнивать не только основные параметры (максимальный прямой ток и обратное напряжение), но и другие характеристики — прямое падение напряжения, обратный ток, емкость перехода, температурный диапазон.

Особенности выбора и применения диода КД208А
При использовании КД208А в электронных схемах необходимо учитывать следующие факторы:
- Рабочее напряжение схемы не должно превышать 100 В.
- Средний выпрямленный ток через диод не должен быть больше 1,5 А.
- Для повышения надежности рекомендуется выбирать диод с запасом по току и напряжению 20-30%.
- При параллельном включении диодов необходимо применять выравнивающие резисторы.
- Для защиты от перенапряжений рекомендуется шунтировать диод варистором или супрессором.
Расчет параметров выпрямителя на диоде КД208А
Рассмотрим пример расчета однополупериодного выпрямителя на основе диода КД208А:
Исходные данные: Напряжение вторичной обмотки трансформатора U2 = 30 В Сопротивление нагрузки Rн = 20 Ом
Расчет основных параметров:
- Среднее значение выпрямленного напряжения: Uср = 0.45 * U2 = 0.45 * 30 = 13.5 В
- Среднее значение выпрямленного тока: Iср = Uср / Rн = 13.5 / 20 = 0.675 А
- Максимальное обратное напряжение на диоде: Uобр.макс = 1.41 * U2 = 1.41 * 30 = 42.3 В
- Максимальный прямой ток через диод: Iпр.макс = π * Iср = 3.14 * 0.675 = 2.12 А
Как видим, рассчитанные параметры не превышают предельно допустимых значений для диода КД208А, поэтому он подходит для данной схемы.

Преимущества и недостатки диода КД208А
Рассмотрим основные достоинства и ограничения КД208А:
Преимущества:
- Высокая надежность и долговечность
- Широкий диапазон рабочих температур
- Низкое прямое падение напряжения
- Доступность и невысокая стоимость
- Совместимость со многими схемами
Недостатки:
- Ограниченный максимальный ток (1,5 А)
- Относительно высокая емкость перехода
- Не подходит для высокочастотных применений
- Уступает современным диодам Шоттки по ряду параметров
Несмотря на некоторые ограничения, диод КД208А остается популярным выбором для многих радиолюбителей и инженеров благодаря своей надежности и универсальности.
Рекомендации по монтажу и эксплуатации КД208А
Для обеспечения долгой и надежной работы диода КД208А следует соблюдать следующие правила:
- При монтаже соблюдать полярность диода. Катод обычно помечен полосой на корпусе.
- Использовать теплоотвод при работе с токами, близкими к максимальным.
- Не превышать максимальную температуру перехода 130°C.
- При пайке не перегревать выводы диода. Рекомендуемая температура пайки 260°C в течение не более 10 секунд.
- В импульсных режимах учитывать ограничения по максимальному импульсному току.
Соблюдение этих рекомендаций поможет избежать преждевременного выхода диода из строя и обеспечит его стабильную работу в течение длительного времени.
Аналоги для кд208а — Аналоги
КД208А 1.5E1Отечественный и зарубежный аналоги
КД208А 1.5J1Отечественный и зарубежный аналоги
КД208А 1500SitОтечественный и зарубежный аналоги
КД208А 1N1053 Отечественный и зарубежный аналоги КД208А 1N1085Отечественный и зарубежный аналоги
КД208А 1N1115Отечественный и зарубежный аналоги
КД208А 1N1446Отечественный и зарубежный аналоги
КД208А 1N1450Отечественный и зарубежный аналоги
КД208А 1N1563Отечественный и зарубежный аналоги
КД208А 1N1617Отечественный и зарубежный аналоги
КД208А 1N2289AОтечественный и зарубежный аналоги
КД208А 1N2290AОтечественный и зарубежный аналоги
КД208А 1N2391Отечественный и зарубежный аналоги
КД208А 1N2400Отечественный и зарубежный аналоги
КД208А 1N2409Отечественный и зарубежный аналоги
КД208А 1N2418Отечественный и зарубежный аналоги
КД208А 1N2638Отечественный и зарубежный аналоги
КД208А 1N2847Отечественный и зарубежный аналоги
КД208А 1N4817Отечественный и зарубежный аналоги
КД208А 1N5392Отечественный и зарубежный аналоги
КД208А 1S020Отечественный и зарубежный аналоги
КД208А 1WS1Отечественный и зарубежный аналоги
КД208А A121-1tОтечественный и зарубежный аналоги
КД208А A132-1tОтечественный и зарубежный аналоги
КД208А A168-1tОтечественный и зарубежный аналоги
КД208А A7B1 Отечественный и зарубежный аналоги КД208А A7B5Отечественный и зарубежный аналоги
КД208А A7B9Отечественный и зарубежный аналоги
КД208А B7B1Отечественный и зарубежный аналоги
КД208А B7B5Отечественный и зарубежный аналоги
КД208А B7B9Отечественный и зарубежный аналоги
Отечественный и зарубежный аналоги
КД208А COD15314Отечественный и зарубежный аналоги
КД208А CTN100Отечественный и зарубежный аналоги
КД208А CTP100Отечественный и зарубежный аналоги
КД208А IS1071Отечественный и зарубежный аналоги
КД208А IS1849Отечественный и зарубежный аналоги
КД208А IS444Отечественный и зарубежный аналоги
КД208А M1B1Отечественный и зарубежный аналоги
КД208А M1B5Отечественный и зарубежный аналоги
КД208А M1B9Отечественный и зарубежный аналоги
КД208А MB236Отечественный и зарубежный аналоги
КД208А ZL103MОтечественный и зарубежный аналоги
КД208А ZS271Отечественный и зарубежный аналоги
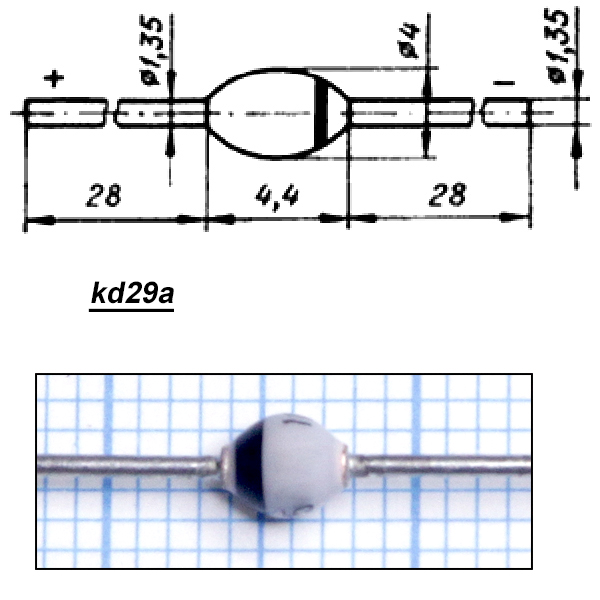
Диод КД208А
Справочник количества содержания ценных металлов в диоде КД208А согласно паспорта на изделие и информационной литературы. Указано точное значение драгоценных металлов в граммах (Золото, серебро, платина, палладий и другие) на единицу изделия.
Указано точное значение драгоценных металлов в граммах (Золото, серебро, платина, палладий и другие) на единицу изделия.
Содержание драгоценных металлов в диоде КД208А
Золото: 3,30E-04 грамм.
Серебро: 0 грамм.
Платина: 0 грамм.
Палладий: 0 грамм.
Источник информации: Из справочника Связь-Инвест.
Фото диода КД208А:
Панель ламповая виды
Диод — электронный элемент, обладающий различной проводимостью в зависимости от направления электрического поля. Электрод диода, подключаемый к положительному полюсу источника тока, когда диод открыт (то есть имеет маленькое сопротивление), называют анодом, подключаемый к отрицательному полюсу — катодом.
О комплектующем изделии – Диод
Диод – видео.
Диод это полупроводниковый прибор основанный на PN-переходе. А если без теории, то диод в одном направлении пропускает ток, а в другом нет. Вот и все.
Как работает диод – видео.
В этом выпуске вы узнаете: что такое диод, принцип действия диода, как работает диод, что такое p – n переход; что такое прямой ток диода, что такое обратный ток диода; каково внутреннее сопротивление диода; что такое вольт- амперная характеристика диода; что такое пропускное и не пропускное напряжение диода; как работает диод в цепи постоянного тока, как работает диод в цепи переменного тока; как устроен плоскостной диод; какие существуют виды диодов; как устроен выпрямительный диод.
Характеристики диодов КД208А:
Купить или продать а также цены на Диод КД208А:
Оставьте отзыв о КД208А:
Диод кд208а.
|
Диод (цоколевка) |
Uоб/Uимп В/В |
Iпр/Iимп А/А |
Uпр/Iпр В/А |
Cд/Uд пф/В (T нс) |
Io(25)Ioм мА/мА |
Fmax кГц |
P/Pт Вт/Вт |
|
КД208А |
100/100 |
1. |
1.0/1 |
|
0.05/0.2 |
1 |
|
 5/
5/
Диод д2б.
Приложение 4
Сердечник: ШЛ12х25
Мощность: 9 Вт
Ток первичной обмотки: 0,071/0,041 А
Масса: 0,49 кг
|
Электрические параметры, габаритные и установочные размеры, а также масса трансформаторов ТПП236 на 220 В такие же, как у соответствующих трансформаторов ТПП236 на 127/220 В.
Напряжение на отводах первичных обмоток трансформаторов ТПП236 на 127/220 В:
-
между выводами 1 и 2, 6 и 7 — 7 В;
-
между выводами 2 и 3, 7 и 8 — 100 В;
-
между выводами 3 и 4, 8 и 9 — 20 В;
-
между выводами 4 и 5, 9 и 10 — 11 В.
При использовании трансформаторов ТПП236-127/220 на 127 В необходимо:
-
соединить выводы 1 и 6, 4 и 9, при этом первичные обмотки 1-6 и 4-9 соединяются последовательно;
-
подать напряжение 127 В на выводы 1 и 4 (6 и 9).
При использовании трансформаторов ТПП236-127/220 на 220 В необходимо:
-
соединить выводы 3 и 7;
-
подать напряжение 220 В на выводы 2 и 9.
-
или соединить выводы 4 и 7;
-
подать напряжение 220 В на выводы 2 и 8.
В трансформаторах ТПП236 возможно последовательное и параллельное согласное соединение вторичных обмоток. Последовательное включение различных вторичных обмоток позволяет подобрать необходимое выходное напряжение, параллельное — повысить мощность на выходных обмотках. При последовательном включении обмоток с разными допустимыми токами ток через обмотки не должен превышать минимально допустимого. Параллельное соединение допускается только для тех обмоток, напряжение на зажимах которых одинаковы.
Диод in5399 характеристики аналоги
Выпрямительный кремниевый диод. Диапазон напряжения от 50 до 1000 вольт ток 1.5 ампера.
Особенности:
- Номинальное напряжение до 1000В
- Низкое прямое падение напряжения
- Высокая допустимая нагрузка по току
- Высокая надежность
- Высокая импульсная перегрузочная способность
Механические данные
- Корпус: литой пластиковый корпус D0-15
- Пластиковые материалы UL классификация воспламеняемости 94 V-0
- Вывода: аксиальные вывода, пайка в MIL-STD-202, методика 208
- Полярность: цветовое кольцо обозначает катод
- Высокая температура пайки, гарантированно: 250°С в течение 10 секунд
- Монтажное положение: любое
- Вес: 0.40 грамма
Максимальные технические и электрические характеристики диодов 1N5391 -1N5399
Значения параметров при 25°С температуре окружающей среды, если не указано иное.
Однофазный, напряжение (В) половина волны, частота – 60 Гц, для резистивных и индуктивных нагрузок. Для емкостной нагрузки уменьшайте ток на 20%
ДИОДЫ, АНАЛОГИ
Здесь представлена самая большая таблица взаимозаменяемости импортных и отечественных диодов собраных в интернете. Часть 2. Полные и функциональные аналоги диодов. Даташит на каждый диод можно посмотреть введя её название в поисковую форму datasheet вправой части сайта. Цены на радиодетали можно посмотреть в любом интернет магазине.
1N3064 КД521А
1N3064M КД521А
1N3065 КД521А
1N3067 КД521Г
1N3082 КД205Г
1N3083 КД205Б
1N3121 Д220
1N3184 КД205А
1N3193 КД205Л
1N3194 Д229Л
1N3228 КД105Г
1N3229 КД205А
1N3238 Д229Ж
1N3239 КД205Л
1N3253 КД205Л
1N3254 Д229Л
1N3270 Д246Б
1N3277 КД205Л
1N3278 Д229Л
1N3282 МД218
1N3545 КД205Г
1N3547 Д229Л
1N3600 КД209А
1N3604 КД521А
1N3606 КД521А
1N3607 КД521А
1M3639 КД205Л
1N3640 Д229Л
1N3657 Д246Б
1N3659 КД205Л
1N3748 КД205Г
1N3749 КД205Б
1N3750 КД205Ж
1N3827 КС456А
1N3827A КС456А
1N3873 КД509А
1N3873H КД509А
1N3954 КД509А
1N4001 КД208, КД209, КД226А, КД243А
1N4002 КД243Б
1N4003 КД243В
1N4004 КД243Г
1N4005 КД243Д
1N4006 КД243Е
1N4007 КД243Ж
1N4008 МД3Б
1N4099 КС168А
1N4147 КД503А
1N4148 КД510, КД521А, КД522Б, КД106А
1N4149 КД521А
1N4150 КД522Б, КД106А
1N4153 КД521А
1N4305 КД521А
1N4364 Д229Ж
1N4365 КД205Л
1N4366 Д229К
1N4367 Д229Л
1N4437 Д246
1N4438 КД206В
1M4439 КД210Б
1N4446 КД521А, КД522Б
1N4447 КД521А
1N4448 КД521А
1N4449 КД521А
1N4454 КД521А
1N4531 КД521А
1N4622 КС139А
1N4624 КС147А
1N4655 КС456А
1N4686 КС139А
1N4688 КС147А
1N4734 КС456А
1N4817 КД208А
1N5151 КД521А
1N5209 Д223Б
1N5216 КД205Б
1N5217 КД205Ж
1N5318 КД521А
1N5392 КД208А
1N5393 КД258А
1N5395 КД258Б
1N5397 КД258В
1N5398 КД258Г
1N5399 КД258Д
1N5400 КД280А
1N5401 КД227А, КД280Б
1N5402 КД280В
1N5404 КД280Г
1N5406 КД280Д
1N5407 КД280Е
1N5408 КД280Ж
1N5624 КД257А
1N5720 КД503А
1N5819 КДШ2105В
1P644 Д229В
1P647 Д229Е
1S032 КД205Л
1S034 Д229Л
1S41 КД205Л
1S43 Д229Л
1S101 КД205Л
1S103 Д229Л
1S113 Д229Е
1S148 Д229К
1S162 Д243
1S163 Д245
1S164 Д246
1S165 КД206Б
1S307 Д18
1S313 КД205В
1S314 КД205Б
1S315 КД205А
1S421 Д243
1S423 Д246
1S427 КД210Б
1S473 Д811
1S544 КД210Б
1S558 КД205А
1S559 КД205В
1S1219 КД521Г
1S1220 КД521Г
1S1230 КД205Б
1S1231 КД205А
1S1232 КД205Ж
1S1473 КД521Г
1S1763 КД205Б
1S1943 КД205Б
1S1944 КД205Ж
1T502 КД205Г
1T504 КД205Б
1T505 КД205А
1T506 КД205Ж
20S5 КД205Г
20TQ045 КДШ2965Б
20TQ060 КДШ2965А
24J2 Д223Б
2A04 КД411ЕМ
2A05 КД411ВМ-ДМ
2A06 КД411АМ, БМ, НМ
2T502 КД205Г
2T504 КД205Б
2T505 КД205А
2T506 КД205Ж
3C15 Д303
3T502 КД205Г
4T502 КД205Г
7,00E+01 Д229Ж
7J1 Д229Ж
7J2 КД205Л
75R2B КД205Л
BAS32 КД811А
BAV682 КД811Б
BY296P КД266А
BY297P КД226Б
BY298P КД226В
BY299P КД226Д
DL4148 КД521А, 522Б-SMD
ESP5300 Д245Б
F0100 КД509А
F1E3 Д245Б
F1K3 Д248Б
F2B3 Л242
F2h4 КД206Б
F2M3 КД203Г
F2N3 КД210Б
FD600 КД521А
FDN600 КД521А
FPZ5V6 КС456А
FR101 КД247Е
FR102 КД247А
FR103 КД247Б
FR104 КД247В
FR105 КД247Г
FR106 КД247Д
FR153 КД258А
FR154 КД258Б
FR155 КД258В
FR156 КД258Г
FR157 КД258Д
FR202 КД226А
FR203 КД226Б
FR204 КД226В
FR205 КД226Г
FR206 КД226Д
FR303 КД257А
FR304 КД257Б
FR305 КД257В
FR306 КД257Г
FR307 КД257Д
G65HZ Д248Б
G1010 Д242
G3010 Д245
G4010 Д246
GP15d КД258А
GP15g КД258Б
GP15j КД258В
GP15k КД258Г
GP15m КД258Д
HDS901 КД521Г
HDS9003 КД509А
HMG626A Д220
HMG662 Д220Б
HMG662A Д220Б
HMG663 Д220Б
HMG844 Д220Б
HMG904 КД521Г
HMG904A КД521Г
HMG907 КД521Г
HMG907A КД521Г
HMG2873 КД509А
HMG3064 КД521А
HMG3596 КД521Г
HMG3598 КД521А
HMG3600 КД509А
HMG4150 КД509А
HMG4319 КД521А
HMG4322 КД509А
HR9 Д818А
HS033A КС133А
HS033B КС133А
HS2039 КС139А
HS7033 КС133А
HS9010 КД521Г
HS9501 КД521А
HS9504 КД521А
HS9507 КД521А
JE2 КД205Л
LAC2002 КС147А
LD57C АЛ336В
LDD5 КД521Б
LDD10 КД521Б
LDD15 КД521Б
LDD50 КД521Б
LR33H КС133А
M1B1 КД208А
M1B5 КД208А
M1B9 КД208А
M4HZ Д229Е
M14 Д229В
M68 Д229Ж
M69B КД205Л
M69C КД205Г
M500B КД205Е
M500C КД205А
R604 Д246
R606 КД206В
R612 Д243
R614 Д246
R616 КД206В
RGP10a КД247Е
RGP10b КД247А
RGP10d КД247Б
RGP10g КД247В
RGP10j КД247Г
RGP10k КД247Д
RGP15d КД258А
RGP15g КД258Б
RGP15j КД258В
RHP15k КД258Г
RGP15m КД258Д
RGP30d КД257А
RGP30g КД257Б
RGP20j КД257В
RGP30k КД257Г
RGP30m КД257Д
RL204 КД411ЕМ
RL205 КД411ВМ-ДМ
RL206 КД411АМ, БМ, НМ
RZ18 КС218Ж
RZ22 КС222Ж
RZZ11 КС211Ж
S1,5-0,1 КД208А
S2A-12 Д243
S2E20 КД205Г
S2E60 КД205Ж
S5A1 Д304
S5A2 Д243Б
S5A3 Д245Б
S5A6 Д248Б
S5AN12 КД206Б
S6AN12 КД206В
S7AN12 КД203Г
S8AN12 КД210Б
S15 КД205А
S17 КД205Г
S18 КД205А
S18A КД205А
S19 Д7Ж
S20-06 Д248Б
S23A КД205Ж
S26 Д229К
S28 КД105Г
S30 КД205Ж
S31 КД205В
S83 Д229К
S92A КД205Л
S101 КД205Г
S106 Д7Ж
S205 Д210
S206 Д211
S208 МД217
S210 МД218
S219 Д7Ж
S222 КД205Г
S223 КД205В
S234 КД105Г
S252 КД205Г
S253 КД205В
S256 КД105Ж
S425 КД206В
S427 КД210Б
S65250 КД509А
SD1A КД205Ж
SD11 Д101
SD17Z КД205Г
SD91A Д229Ж
SD92A КД205Л
SD93 Д229К
SE05B КД205Ж
SE05S КД205Г
SE1,5SS КД208А
SFD43 КД521Г
SFD83 КД521Г
SG203E, K Д243Б
SG5200 КД521А
SG5260 КД521А
SJ103E, K Д304
SJ104E, K Д242
SJ204E, K Д243
SL3 Д245Б
SM20 КД205Л
SM230 Д229К
SV131 Д818А
SV134 Д811
SVM91 Д818А
SVM905 Д818А
SVM9010 Д818А
SVM9011 Д818А
SVM9020 Д818А
SVM9021 Д818А
SW05B КД205Ж
SW05S КД205Г
SW1S Д229Ж
SW1SS КД205Л
SZ9 Д818А
SZ11 Д811
TIC106 КУ223И
TF24 Д226В
TK20 КД205Л
TK40 Д229Л
TMD45 Д207
TS1 Д229Ж
TS2 КД205Л
TS4 Д229Л
UR215 Д303
UP12069 КД205Л
UP12070 Д229Л
UP12070A Д229Л
URE100X Д304
URF100X Д304
URG100X Д304
UT112 Д229Ж
UT113 КД205Л
UT114 Д229К
UT115 Д229Л
UT212 Д229К
UT213 Д229Л
XS10 Д229Ж
XS17 КД205Л
Z1550 КС156А
Z1555 КС156А
Z1560 КС156А
Z1565 КС156А
Z1570 КС156А
Z1A5,6 КС156А
Z1A6,8 КС168А
Z1A11 Д811
Z1B5,6 КС156А
Z1B6,8 КС168А
Z1B11 Д811
Z1C5,6 КС156А
Z1C11 Д811
Z1D6,8 КС168А
Отличная новость! Если вы решили приобрести товар по запросу diode in5399, то вы находитесь в нужном месте. AliExpress — это платформа для онлайн-шопинга, где для вас представлены тысячи товаров самых разных категорий. С AliExpress вы всегда можете быть уверены, что найдете нужные вам вещи, будь то дорогостоящие изделия или небольшие покупки. Наша база пополняется каждый день, поэтому мы предлагаем широкий выбор продукции самых разных категорий. Наши поставщики — как известные торговые марки, так и независимые продавцы — гарантируют быструю доставку и надежность, а также удобные и безопасные способы оплаты.
Удобный поиск помогает находить не только нужные товары, но и похожие продукты и возможные комплектующие. Вместе с этим вы получаете самые лучшие цены в сети, выгодную доставку и возможность забрать товар в ближайшей удобной для вас точке.
Иногда выбрать товар среди всех возможных предложений оказывается непросто. Мы позаботились о вашем удобстве и создали удобную систему сравнения. С AliExpress вы легко можете сопоставить цены и воспользоваться наиболее выгодным предложением. Мы также будем рады сообщить вам о начале специальных акций, а также о купонах на скидку. Если у вас возникнут сомнения, вы всегда можете ознакомиться с отзывами покупателей и сравнить рейтинг магазинов. Мы высоко ценим мнение клиентов, поэтому под каждым товаром вы найдете комментарии тех, кто уже сделал покупку. Одним словом, вам больше не нужно слепо доверять — вы можете просто положиться на опыт других покупателей.
Для новичков в AliExpress мы раскроем секрет, как получить самые выгодные предложения на нашем сайте. Перед тем как нажать кнопку «Купить сейчас», проверьте наличие купонов на скидку. Это могут быть купоны AliExpress или же купоны магазинов-сотрудников. Также вы можете получить купоны, выигрывая в нашей игре в приложении AliExpress app. Вместе с бесплатной доставкой, которую предлагают большинство продавцов на нашем сайте, вы сможете приобрести diode in5399 по самой выгодной цене.
AliExpress — это передовые технологии, новейшие тенденции и самые обсуждаемые бренды, а также отличное качество, цена и сервис. Онлайн-покупки стало делать еще проще и надежней. Экономьте время и деньги без ущерба качеству.
Расчет однополупериодного выпрямителя — КиберПедия
Цель: Формирование у студента компетенций ПК-18, ПК-38.
Теоретическая часть
Однополупериодный выпрямитель. Однополупериодный выпрямитель (рисунок 2.1 а) состоит из трансформатора, ко вторичной обмотке которого последовательно присоединены диод VDи нагрузочный резистор .
Для упрощения анализа работы выпрямителей трансформатор и диод считают идеальными, т.е. принимают следующие допущения: у трансформатора активное сопротивление обмоток, а у диода прямое сопротивление равны нулю; обратное сопротивление диода равно бесконечности; в трансформаторе отсутствуют потоки рассеяния. При таких допущениях с подключением первичной обмотки трансформатора к сети переменного синусоидального напряжения во вторичной обмотке будет наводиться синусоидальная ЭДС (рисунок 2.1 б).
а) б)
в)
Рисунок 2.1
Работу выпрямителя рассматривают с помощью временных диаграмм (рисунок 2.1 б, в). В первый полупериод, т.e. в интервале времени 0 – Т/2, диод открыт и в нем появляется ток , а в нагрузочном резисторе – ток , причем . Падение напряжения на диоде . В интервале времени Т/2 – Т диод закрыт, ток , а к запертому диоду прикладывается обратное напряжение т.е. , тогда , где – действующее значение напряжения во вторичной обмотке.
Основными электрическими параметрами однополулериодного выпрямителя являются:
– средние значения выпрямленных тока и напряжения и ;
– мощность нагрузочного устройства ;
– амплитуда основной гармоники выпрямленного напряжения ;
– коэффициент пульсаций выпрямленного напряжения ;
– коэффициент полезного действия ,
где – мощность потерь в трансформаторе, а – мощность потерь в диодах.
В однополупериодном выпрямителе (рис. 2.1):
, (2.1)
.
Действующее значение тока вторичной обмотки трансформатора:
. (2.3)
Коэффициент пульсаций можно получить из разложения в ряд Фурье выходного напряжения однополупериодного выпрямителя:
,
.
Основным преимуществом однополупериодного выпрямителя является его простота. Недостатками этого выпрямителя являются: большой коэффициент пульсаций, малые значения выпрямленных тока и напряжения. Вместе с тем, следует обратить внимание еще на один недостаток однополупериодного выпрямителя. Ток имеет постоянную составляющую, которая вызывает подмагничивание сердечника трансформатора, из-за чего уменьшается магнитная проницаемость сердечника, что в свою очередь снижает индуктивность обмоток трансформатора. Это приводит к росту тока холостого хода трансформатора, а следовательно, к снижению КПД всего выпрямителя.
Диод в выпрямителе является основным элементом, поэтому диоды должны соответствовать основным электрическим параметрам выпрямителей. При выборе типа диодов необходимо знать:
– среднее значение прямого тока ;
– максимальное обратное напряжение на диоде . Для надежной работы диодов в выпрямителях требуется, чтобы каталожные параметры превышали рассчитанные значения примерно на 30%. Отметим, что при выпрямлении напряжения, амплитудное значение которого превышает для одного диода, можно включать последовательно два или несколько однотипных диодов. Однако эти диоды должны быть зашунтированы сопротивлением, примерно равным .
Промышленность выпускает полупроводниковые диодные столбы (например, КЦ 106, КЦ 201). Выпрямительный столб – это группа последовательно соединенных диодов, помешенных в общий корпус. Такие столбы выдерживают напряжения свыше 15 кВ.
Задачи
1. В однополупериодном выпрямителе напряжение на вторичной обмотке трансформатора В, частота сети = 50 Гц. Сопротивление диода в прямом направлении = 0. Для нагрузочного резистора сопротивлением 200 Ом определить средние значения выпрямленного напряжения и тока на нагрузочном резисторе, среднее значение тока в диоде , максимальное обратное напряжение диода . Выбрать параметры необходимого диода.
2. В схеме однополупериодного выпрямителя задан диод КД208А с параметрами = 1,5 А и = 100 В. Определить максимальное напряжение вторичной обмотки трансформатора и мощность, выделяемую в нагрузке = 200 Ом.
3. Определить среднее и максимальное значение прямого тока, а также максимальное обратное напряжение полупроводникового диода в однополупериодном выпрямителе (рисунок 2.1 а) и коэффициент трансформации трансформатора, если на нагрузочном резисторе сопротивлением = 3 кОм среднее значение выпрямленного напряжения =180 В. Напряжение сети В. Сопротивление диода в прямом направлении и обратный ток считать равными нулю.
4. Определить амплитуду тока в нагрузочном резисторе сопротивлением = 2,5 кОм однополупериодного выпрямителя (рисунок 2.1 а), если напряжение на первичной обмотке трансформатора В, коэффициент трансформации п = 0,4. Сопротивление диода в прямом направлении считать равными нулю.
5. Для питания постоянным током потребителя мощностью 250 Вт при напряжении Ud= 100 В необходимо собрать схему однополупериодного выпрямителя, подобрав диоды из справочника. Указать допустимые параметры для выбранного диода.
Вопросы к практическому занятию
1. Из чего состоит однополупериодный выпрямитель?
2. Приведите соотношения между входным напряжением выпрямителя и его средним значением.
3. Пояснить связь между действующим значением тока выпрямителя и его постоянной составляющей.
4. Что называется коэффициентом пульсаций выпрямленного напряжения?
5. Чему равен коэффициент пульсаций для однополупериодного выпрямителя?
6. Какое условие используется при выборе диодов после расчета однополупериодного выпрямителя?
7. Каковы различия между входным и выходным сигналами однополупериодного выпрямителя?
8. Поясните принцип действия однополупериодного выпрямителя.
Практическое занятие 3
Диод rl 5245 его аналоги
ДИОДЫ, АНАЛОГИ
Здесь представлена самая большая таблица взаимозаменяемости импортных и отечественных диодов собраных в интернете. Часть 2. Полные и функциональные аналоги диодов. Даташит на каждый диод можно посмотреть введя её название в поисковую форму datasheet вправой части сайта. Цены на радиодетали можно посмотреть в любом интернет магазине.
1N3064 КД521А
1N3064M КД521А
1N3065 КД521А
1N3067 КД521Г
1N3082 КД205Г
1N3083 КД205Б
1N3121 Д220
1N3184 КД205А
1N3193 КД205Л
1N3194 Д229Л
1N3228 КД105Г
1N3229 КД205А
1N3238 Д229Ж
1N3239 КД205Л
1N3253 КД205Л
1N3254 Д229Л
1N3270 Д246Б
1N3277 КД205Л
1N3278 Д229Л
1N3282 МД218
1N3545 КД205Г
1N3547 Д229Л
1N3600 КД209А
1N3604 КД521А
1N3606 КД521А
1N3607 КД521А
1M3639 КД205Л
1N3640 Д229Л
1N3657 Д246Б
1N3659 КД205Л
1N3748 КД205Г
1N3749 КД205Б
1N3750 КД205Ж
1N3827 КС456А
1N3827A КС456А
1N3873 КД509А
1N3873H КД509А
1N3954 КД509А
1N4001 КД208, КД209, КД226А, КД243А
1N4002 КД243Б
1N4003 КД243В
1N4004 КД243Г
1N4005 КД243Д
1N4006 КД243Е
1N4007 КД243Ж
1N4008 МД3Б
1N4099 КС168А
1N4147 КД503А
1N4148 КД510, КД521А, КД522Б, КД106А
1N4149 КД521А
1N4150 КД522Б, КД106А
1N4153 КД521А
1N4305 КД521А
1N4364 Д229Ж
1N4365 КД205Л
1N4366 Д229К
1N4367 Д229Л
1N4437 Д246
1N4438 КД206В
1M4439 КД210Б
1N4446 КД521А, КД522Б
1N4447 КД521А
1N4448 КД521А
1N4449 КД521А
1N4454 КД521А
1N4531 КД521А
1N4622 КС139А
1N4624 КС147А
1N4655 КС456А
1N4686 КС139А
1N4688 КС147А
1N4734 КС456А
1N4817 КД208А
1N5151 КД521А
1N5209 Д223Б
1N5216 КД205Б
1N5217 КД205Ж
1N5318 КД521А
1N5392 КД208А
1N5393 КД258А
1N5395 КД258Б
1N5397 КД258В
1N5398 КД258Г
1N5399 КД258Д
1N5400 КД280А
1N5401 КД227А, КД280Б
1N5402 КД280В
1N5404 КД280Г
1N5406 КД280Д
1N5407 КД280Е
1N5408 КД280Ж
1N5624 КД257А
1N5720 КД503А
1N5819 КДШ2105В
1P644 Д229В
1P647 Д229Е
1S032 КД205Л
1S034 Д229Л
1S41 КД205Л
1S43 Д229Л
1S101 КД205Л
1S103 Д229Л
1S113 Д229Е
1S148 Д229К
1S162 Д243
1S163 Д245
1S164 Д246
1S165 КД206Б
1S307 Д18
1S313 КД205В
1S314 КД205Б
1S315 КД205А
1S421 Д243
1S423 Д246
1S427 КД210Б
1S473 Д811
1S544 КД210Б
1S558 КД205А
1S559 КД205В
1S1219 КД521Г
1S1220 КД521Г
1S1230 КД205Б
1S1231 КД205А
1S1232 КД205Ж
1S1473 КД521Г
1S1763 КД205Б
1S1943 КД205Б
1S1944 КД205Ж
1T502 КД205Г
1T504 КД205Б
1T505 КД205А
1T506 КД205Ж
20S5 КД205Г
20TQ045 КДШ2965Б
20TQ060 КДШ2965А
24J2 Д223Б
2A04 КД411ЕМ
2A05 КД411ВМ-ДМ
2A06 КД411АМ, БМ, НМ
2T502 КД205Г
2T504 КД205Б
2T505 КД205А
2T506 КД205Ж
3C15 Д303
3T502 КД205Г
4T502 КД205Г
7,00E+01 Д229Ж
7J1 Д229Ж
7J2 КД205Л
75R2B КД205Л
BAS32 КД811А
BAV682 КД811Б
BY296P КД266А
BY297P КД226Б
BY298P КД226В
BY299P КД226Д
DL4148 КД521А, 522Б-SMD
ESP5300 Д245Б
F0100 КД509А
F1E3 Д245Б
F1K3 Д248Б
F2B3 Л242
F2h4 КД206Б
F2M3 КД203Г
F2N3 КД210Б
FD600 КД521А
FDN600 КД521А
FPZ5V6 КС456А
FR101 КД247Е
FR102 КД247А
FR103 КД247Б
FR104 КД247В
FR105 КД247Г
FR106 КД247Д
FR153 КД258А
FR154 КД258Б
FR155 КД258В
FR156 КД258Г
FR157 КД258Д
FR202 КД226А
FR203 КД226Б
FR204 КД226В
FR205 КД226Г
FR206 КД226Д
FR303 КД257А
FR304 КД257Б
FR305 КД257В
FR306 КД257Г
FR307 КД257Д
G65HZ Д248Б
G1010 Д242
G3010 Д245
G4010 Д246
GP15d КД258А
GP15g КД258Б
GP15j КД258В
GP15k КД258Г
GP15m КД258Д
HDS901 КД521Г
HDS9003 КД509А
HMG626A Д220
HMG662 Д220Б
HMG662A Д220Б
HMG663 Д220Б
HMG844 Д220Б
HMG904 КД521Г
HMG904A КД521Г
HMG907 КД521Г
HMG907A КД521Г
HMG2873 КД509А
HMG3064 КД521А
HMG3596 КД521Г
HMG3598 КД521А
HMG3600 КД509А
HMG4150 КД509А
HMG4319 КД521А
HMG4322 КД509А
HR9 Д818А
HS033A КС133А
HS033B КС133А
HS2039 КС139А
HS7033 КС133А
HS9010 КД521Г
HS9501 КД521А
HS9504 КД521А
HS9507 КД521А
JE2 КД205Л
LAC2002 КС147А
LD57C АЛ336В
LDD5 КД521Б
LDD10 КД521Б
LDD15 КД521Б
LDD50 КД521Б
LR33H КС133А
M1B1 КД208А
M1B5 КД208А
M1B9 КД208А
M4HZ Д229Е
M14 Д229В
M68 Д229Ж
M69B КД205Л
M69C КД205Г
M500B КД205Е
M500C КД205А
R604 Д246
R606 КД206В
R612 Д243
R614 Д246
R616 КД206В
RGP10a КД247Е
RGP10b КД247А
RGP10d КД247Б
RGP10g КД247В
RGP10j КД247Г
RGP10k КД247Д
RGP15d КД258А
RGP15g КД258Б
RGP15j КД258В
RHP15k КД258Г
RGP15m КД258Д
RGP30d КД257А
RGP30g КД257Б
RGP20j КД257В
RGP30k КД257Г
RGP30m КД257Д
RL204 КД411ЕМ
RL205 КД411ВМ-ДМ
RL206 КД411АМ, БМ, НМ
RZ18 КС218Ж
RZ22 КС222Ж
RZZ11 КС211Ж
S1,5-0,1 КД208А
S2A-12 Д243
S2E20 КД205Г
S2E60 КД205Ж
S5A1 Д304
S5A2 Д243Б
S5A3 Д245Б
S5A6 Д248Б
S5AN12 КД206Б
S6AN12 КД206В
S7AN12 КД203Г
S8AN12 КД210Б
S15 КД205А
S17 КД205Г
S18 КД205А
S18A КД205А
S19 Д7Ж
S20-06 Д248Б
S23A КД205Ж
S26 Д229К
S28 КД105Г
S30 КД205Ж
S31 КД205В
S83 Д229К
S92A КД205Л
S101 КД205Г
S106 Д7Ж
S205 Д210
S206 Д211
S208 МД217
S210 МД218
S219 Д7Ж
S222 КД205Г
S223 КД205В
S234 КД105Г
S252 КД205Г
S253 КД205В
S256 КД105Ж
S425 КД206В
S427 КД210Б
S65250 КД509А
SD1A КД205Ж
SD11 Д101
SD17Z КД205Г
SD91A Д229Ж
SD92A КД205Л
SD93 Д229К
SE05B КД205Ж
SE05S КД205Г
SE1,5SS КД208А
SFD43 КД521Г
SFD83 КД521Г
SG203E, K Д243Б
SG5200 КД521А
SG5260 КД521А
SJ103E, K Д304
SJ104E, K Д242
SJ204E, K Д243
SL3 Д245Б
SM20 КД205Л
SM230 Д229К
SV131 Д818А
SV134 Д811
SVM91 Д818А
SVM905 Д818А
SVM9010 Д818А
SVM9011 Д818А
SVM9020 Д818А
SVM9021 Д818А
SW05B КД205Ж
SW05S КД205Г
SW1S Д229Ж
SW1SS КД205Л
SZ9 Д818А
SZ11 Д811
TIC106 КУ223И
TF24 Д226В
TK20 КД205Л
TK40 Д229Л
TMD45 Д207
TS1 Д229Ж
TS2 КД205Л
TS4 Д229Л
UR215 Д303
UP12069 КД205Л
UP12070 Д229Л
UP12070A Д229Л
URE100X Д304
URF100X Д304
URG100X Д304
UT112 Д229Ж
UT113 КД205Л
UT114 Д229К
UT115 Д229Л
UT212 Д229К
UT213 Д229Л
XS10 Д229Ж
XS17 КД205Л
Z1550 КС156А
Z1555 КС156А
Z1560 КС156А
Z1565 КС156А
Z1570 КС156А
Z1A5,6 КС156А
Z1A6,8 КС168А
Z1A11 Д811
Z1B5,6 КС156А
Z1B6,8 КС168А
Z1B11 Д811
Z1C5,6 КС156А
Z1C11 Д811
Z1D6,8 КС168А
Обозначение, применение и параметры диодов Шоттки
К многочисленному семейству полупроводниковых диодов названных по фамилиям учёных, которые открыли необычный эффект, можно добавить ещё один. Это диод Шоттки.
Немецкий физик Вальтер Шоттка открыл и изучил так называемый барьерный эффект возникающий при определённой технологии создания перехода металл-полупроводник.
Основной «фишкой» диода Шоттки является то, что в отличие от обычных диодов на основе p-n перехода, здесь используется переход металл-полупроводник, который ещё называют барьером Шоттки. Этот барьер, так же, как и полупроводниковый p-n переход, обладает свойством односторонней электропроводимости и рядом отличительных свойств.
В качестве материала для изготовления диодов с барьером Шоттки преимущественно используется кремний (Si) и арсенид галлия (GaAs), а также такие металлы как золото, серебро, платина, палладий и вольфрам.
На принципиальных схемах диод Шоттки изображается вот так.
Как видим, его изображение несколько отличается от обозначения обычного полупроводникового диода.
Кроме такого обозначения на схемах можно встретить и изображение сдвоенного диода Шоттки (сборки).
Сдвоенный диод – это два диода смонтированных в одном общем корпусе. Выводы катодов или анодов у них объединены. Поэтому такая сборка, как правило, имеет три вывода. В импульсных блоках питания обычно применяются сборки с общим катодом.
Так как два диода размещены в одном корпусе и выполнены в едином технологическом процессе, то их параметры очень близки. Поскольку они размещены в едином корпусе, то и температурный режим их одинаков. Это увеличивает надёжность и срок службы элемента.
У диодов Шоттки есть два положительных качества: весьма малое прямое падение напряжения (0,2-0,4 вольта) на переходе и очень высокое быстродействие.
К сожалению, такое малое падение напряжения проявляется при приложенном напряжении не более 50-60 вольт. При дальнейшем его повышении диод Шоттки ведёт себя как обычный кремниевый выпрямительный диод. Максимальное обратное напряжение для Шоттки обычно не превышает 250 вольт, хотя в продаже можно встретить образцы, рассчитанные и на 1,2 киловольта (VS-10ETS12-M3).
Так, сдвоенный диод Шоттки (Schottky rectifier) 60CPQ150 рассчитан на максимальное обратное напряжение 150V, а каждый из диодов сборки способен пропустить в прямом включении 30 ампер!
Также можно встретить образцы, выпрямленный за полупериод ток которых может достигать 400А максимум! Примером может служит модель VS-400CNQ045.
Очень часто в принципиальных схемах сложное графическое изображение катода попросту опускают и изображают диод Шоттки как обычный диод. А тип применяемого элемента указывают в спецификации.
К недостаткам диодов с барьером Шоттки можно отнести то, что даже при кратковременном превышении обратного напряжения они мгновенно выходят из строя и главное необратимо. В то время как кремниевые силовые вентили после прекращения действия превышенного напряжения прекрасно самовосстанавливаются и продолжают работать. Кроме того обратный ток диодов очень сильно зависит от температуры перехода. На большом обратном токе возникает тепловой пробой.
К положительным качествам диодов Шоттки кроме высокого быстродействия, а, следовательно, малого времени восстановления можно отнести малую ёмкость перехода (барьера), что позволяет повысить рабочую частоту. Это позволяет использовать их в импульсных выпрямителях на частотах в сотни килогерц. Очень много диодов Шоттки находят своё применение в интегральной микроэлектронике. Выполненные по нано технологии диоды Шоттки входят в состав интегральных схем, где они шунтируют переходы транзисторов для повышения быстродействия.
В радиолюбительской практике прижились диоды Шоттки серии 1N581x (1N5817, 1N5818, 1N5819). Все они рассчитаны на максимальный прямой ток (IF(AV)) – 1 ампер и обратное напряжение (VRRM) от 20 до 40 вольт. Падение напряжения (VF) на переходе составляет от 0,45 до 0,55 вольт. Как уже говорилось, прямое падение напряжения (Forward voltage drop) у диодов с барьером Шоттки очень мало.
Также достаточно известным элементом является 1N5822. Он рассчитан на прямой ток в 3 ампера и выполнен в корпусе DO-201AD.
Также на печатных платах можно встретить диоды серии SK12 – SK16 для поверхностного монтажа. Они имеют довольно небольшие размеры. Несмотря на это SK12-SK16 выдерживают прямой ток до 1 ампера при обратном напряжении 20 – 60 вольт. Прямое падение напряжения составляет 0,55 вольт (для SK12, SK13, SK14) и 0,7 вольт (для SK15, SK16). Также на практике можно встретить диоды серии SK32 – SK310, например, SK36, который рассчитан на прямой ток 3 ампера.
Применение диодов Шоттки в источниках питания.
Диоды Шоттки активно применяются в блоках питания компьютеров и импульсных стабилизаторах напряжения. Среди низковольтных питающих напряжений самыми сильноточными (десятки ампер) являются напряжения +3,3 вольта и +5,0 вольт. Именно в этих вторичных источниках питания и используются диоды с барьером Шоттки. Чаще всего используются трёхвыводные сборки с общим катодом. Именно применение сборок может считаться признаком высококачественного и технологичного блока питания.
Выход из строя диодов Шоттки одна из наиболее часто встречающихся неисправностей в импульсных блоках питания. У него может быть два «дохлых» состояния: чистый электрический пробой и утечка. При наличии одного из этих состояний блок питания компьютера блокируется, так как срабатывает защита. Но это может происходить по-разному.
В первом случае все вторичные напряжения отсутствуют. Защита заблокировала блок питания. Во втором случае вентилятор «подёргивается» и на выходе источников питания периодически то появляются пульсации напряжения, то пропадают.
То есть схема защиты периодически срабатывает, но полной блокировки источника питания при этом не происходит. Диоды Шоттки гарантированно вышли из строя, если радиатор, на котором они установлены, разогрет очень сильно до появления неприятного запаха. И последний вариант диагностики связанный с утечкой: при увеличении нагрузки на центральный процессор в мультипрограммном режиме блок питания самопроизвольно отключается.
Следует иметь в виду, что при профессиональном ремонте блока питания после замены вторичных диодов, особенно с подозрением на утечку, следует проверить все силовые транзисторы выполняющие функцию ключей и наоборот: после замены ключевых транзисторов проверка вторичных диодов является обязательной процедурой. Всегда необходимо руководствоваться принципом: беда одна не приходит.
Проверка диодов Шоттки мультиметром.
Проверить диод Шоттки можно с помощью рядового мультиметра. Методика такая же, как и при проверке обычного полупроводникового диода с p-n переходом. Но и тут есть подводные камни. Особенно трудно проверить диод с утечкой. Прежде всего, элемент необходимо выпаять из схемы для более точной проверки. Достаточно легко определить полностью пробитый диод. На всех пределах измерения сопротивления неисправный элемент будет иметь бесконечно малое сопротивление, как в прямом, так и в обратном включении. Это равносильно короткому замыканию.
Сложнее проверить диод с подозрением на «утечку». Если проводить проверку мультиметром DT-830 в режиме «диод», то мы увидим совершенно исправный элемент. Можно попробовать измерить в режиме омметра его обратное сопротивление. На пределе «20кОм» обратное сопротивление определяется как бесконечно большое. Если же прибор показывает хоть какое-то сопротивление, допустим 3 кОм, то этот диод следует рассматривать как подозрительный и менять на заведомо исправный. Стопроцентную гарантию может дать полная замена диодов Шоттки по шинам питания +3,3V и +5,0V.
Где ещё в электронике используются диоды Шоттки? Их можно обнаружить в довольно экзотических приборах, таких как приёмники альфа и бета излучения, детекторах нейтронного излучения, а в последнее время на барьерных переходах Шоттки собирают панели солнечных батарей. Так, что они питают электроэнергией и космические аппараты.
Диод IN 4007 — мощный полупроводниковый гаджет, который часто применяется в блоках питания, то есть непосредственно в их выпрямительной части (в диодном мосту).
Ключевая задача этих полупроводниковых элементов состоит в том, что они действительно принимают участие в преобразовании переменного напряжения в непрерывное, так как на этом напряжении функционируют практически все микроэлектронные составляющие.
После того как были высказаны слова о назначении и о самом предмете, который может быть интересен пользователю (юзеру), можно «перекинуться» напрямую к самим характеристикам (параметрам) вышеназванного диода.
Характеристика
Знание его данных сможет помочь любому мастеру грамотнее и практичнее применять диод по его прямому назначению. Таким образом, диод IN 4007 обладает оптимальными параметрами:
- Вес элемента — 0,35 грамма;
- Температура пайки устройства — 250 градусов по Цельсию;
- Допустимое обратное напряжение 1000 В;
- Вместимость диода — 15 пФ;
- Предельный долговременный прямой ток 1 А;
- Диапазон температур (рабочих) -65…+175 °С;
- Мощный элемент, который сможет выполнять работу с 220 В и с 380 В;
- Наибольшее (прямое) напряжение 1,1 В;
- Тип корпуса — DO -41.
Маркировка диода 1n4007:
- AL– изготовитель
- 1N
- 400х – 1N400х, где х – 1,2,3,4,5,6,7
- YYWW – YY – год выпуска, WW – неделя выпуска.
Вследствие всего, рассматривая эти данные, можно понять, что диоды выпускаются фактически для блоков питания. Зачастую эти детали можно повстречать в выпрямительной части схемы.
После разбора главных признаков этого диода, можно детально затронуть назначения данного элемента, чтобы юзер, ещё не знакомый с ним, сумел лучше понять, как использовать его в будущем.
Использование
Основная область распространения, в которой прилагаются указанные устройства (конструкции) — это, само собой разумеется, диодные мосты. Об этом было рассказано ещё в начале статьи. Кстати, в качестве другой сферы их использования, но уже менее востребованной, можно представить силовую электронику. В данной сфере деятельности они употребляются в качестве всевозможных аналоговых выпрямителей.
В случае введения таких диодов в обусловленное устройство, можно значительно усовершенствовать наличествующие свойства. Вдобавок диоды IN4007 прекрасно себя зарекомендовали в случае их встраивания в автоматические источники питания. По свидетельству профессионалов, представленные диоды являются в наибольшей степени предпочтительным вариантом для конструкций такого типа.
Аналоги
Не следует забывать, что предложенный элемент IN4007 предстает лишь одним из поверенных довольно огромного семейства устройств такого класса. Помимо этой модели, имеются и иные, наименования которых модифицируются от модели IN4001 до IN4006. Какие ещё модификации присутствуют в представленном диапазоне можно и без труда догадаться, так как во всей этой серии меняется исключительно — завершающий индекс.
По нему, между прочим, можно узнать все о самом устройстве. Оказывается, чем меньше заключительный индекс в названии диода, тем мельче полупроводниковый элемент, применяемый в конструкции. В частности, представители этого семейства конструкций, в процессе их работы продемонстрировали любопытное свойство — это менять свою ёмкость.
Этот показатель непосредственно находится в зависимости от величины возвратного напряжения, которое было приложено к устройству. Отталкиваясь из этой занятной особенности, эксперты пришли к заключению, что представленные элементы можно приспособлять в качестве временных заменителей варикапов.
Между прочим, IN4007 может быть применен и в качестве эрзаца всех предшествующих устройств (девайсов) данной серии. Так как является самым мощным из них, что можно узнать по самому последнему индексу. Поэтому, за неимением диодов этой серии, но с иным индексом, можно без проблем выйти из такой сложной, сменив их диодом IN4007, который является в наибольшей степени универсальным.
1n4007 по даташиту — прежде всего низкое падение напряжения в прямом направлении и высокая пропускная способность.
Можно вспомнить и об аналогах, которые имеются на рынке и готовы заменить данный элемент в случае необходимости. Если юзеру далеки все заграничные конструкции и сердцем он с отечественным производителем, то у него есть основание для радости, поскольку имеется российский аналог диоду выпрямительному IN4007, который всецело отвечает ему, по всем данным — модель КД258Д. Кстати, зарубежному она ничем не уступает, поэтому в случае покупки, юзер не рискует потерять в производительности:
- Diotec Semiconductor — модели IN3549, IN2070 и 10D4;
- Thomson — BYW27-1000, BY156;
- Philips — BYW43;
- Motorola — HEPR0056RT.
Здесь необходимо выделить и тот факт, что здесь далеко не все распространенные аналоги разбираемого устройства, но они уж определенно являются самыми известными.
Заключение
Диод IN 4007 очень часто употребляется для разнообразных модификаций блоков питания. Данный полупроводниковый элемент впору назвать и впрямь незаменимым, если нужна основа или восстановление таких девайсов. К тому же, в силу собственной универсальности, IN 4007 может заменить собой каждую модель из своего окружения.
Диод выпрямиткльный IN 4007 показал себя гаджетом чрезвычайно надёжным, многоцелевым, да и стоит он релятивно недорого. Цена вполне приемлема, что делает его недорогим для любого пользователя. При учёте абсолютно всех вышеуказанных преимуществ, неудивительно почему представленный полупроводниковый элемент настолько востребован, даже несмотря на обилие дешевых и надежных диодных матриц.
Выпрямительные диоды серии КД2хх
Выпрямительные диоды серии КД2ххВЫПРЯМИТЕЛЬНЫЕ ДИОДЫ серии КД2хх
87 типономиналов на 8-май-2021
Предприятия, отмеченные таким цветом, прекратили свое существование.
| Подробная информация о производителях — в ПУТЕВОДИТЕЛе и о корпусах — здесь | |||||||||
| тип | аналог | Uобр, В | Iпр, А | fmax, кГц | Uпр, В | tвос, мкс | корпус | производитель | подробности |
|---|---|---|---|---|---|---|---|---|---|
| КД204А | 400 | 0.4 | 50 | ТОР | # | ||||
| КД206В | 600 | 10 | 1 | 1.5 | 10 | ОРБИТА | # | ||
| КД208А | 100 | 1.5 | 1 | 1 | КД7 | БАРИОН | # | ||
| КД209В | 800 | 0.5 | 1 | 1 | БАРИОН | # | |||
| КД210В1 | 1000 | 10 | 1 | 1 | КТ28 | СТАРТ | |||
| КД212А | 200 | 1 | 100 | 1 | 0.05 | КД16 | ФОТОН | # | |
| КД213А | 100 | 10 | 100 | 1.2 | 0.17 | КД23 | ФОТОН | # | |
| КД215Б | 600 | 1 | 10 | ТОР | # | ||||
| КД216Б | 200 | 10 | 100 | 1.4 | # | ||||
| КД217Б | 200 | 3 | 100 | 1.3 | # | ||||
| КД218 | 100 | 10 | 100 | 1.5 | ФОТОН | ||||
| КД219В | 30 | 10 | 200 | 0.55 | ТОР | # диод Шоттки | |||
| КД220Г | 1000 | 3 | 50 | 1.3 | 0.5 | ТОР | # | ||
| КД221Г | 600 | 0.3 | 50 | 1.4 | 1.5 | КД7 | ФОТОН | # | |
| 2Д222ВС | 40 | 3 | 200 | 0.6 | 4116.4-3 | ТОР, ЭЛТОМ | # пара ОК Шоттки | ||
| КД223А | 200 | 2 | 1.5 | 1.4 | БФТЗПП | # | |||
| КД225ВС | 35 | 3 | 200 | 0.6 | # пара | ||||
| КД226Д | 800 | 2 | 50 | 1.3 | 0.25 | ТОР | # | ||
| КД227Ж | 800 | 10 | 1 | 1.6 | КТ28-2 | СТАРТ | # | ||
| КД227ЖС | 800 | 5 | 1 | 1.6 | КТ28-2 | СТАРТ | пара | ||
| КД228А | 100 | 1 | 100 | 1.1 | ФОТОН | ||||
| КД229ВС | 35 | 3 | 200 | 0.6 | # пара | ||||
| КД230Г | 1000 | 3 | 20 | 1.5 | 0.5 | ТОР | # | ||
| КД231Г | 250 | 10 | 200 | 1 | 0.1 | ТОР | # | ||
| КД232В | 25 | 10 | 200 | 0.7 | ТОР | # | |||
| КД234В | 400 | 3 | 50 | 1.5 | 0.4 | ТОР | # | ||
| КД235А | 40 | 1 | 200 | 0.9 | ДНЕПР | # | |||
| КД236Б | 800 | 1 | 100 | 1.5 | 0.15 | КД7Б | ФОТОН | # | |
| КД237А2/ПМ | 200 | 1 | 300 | 1.35 | 0.05 | нестан. | ПУЛЬСАР | ||
| КД237Б | 200 | 1 | 300 | 1.3 | 0.05 | КД14А | БзПП, ФОТОН | # АЕЯР.432120.437ТУ | |
| КД238ВС | 45 | 7.5 | 200 | 0.65 | КТ28 | ЭЛЕКТРОНИКА | диод Шоттки | ||
| КД239В | 200 | 20 | 500 | 1.4 | 0.05 | КТ28 | ФОТОН | ||
| КД243Ж | 1000 | 1 | 1 | 1.1 | ТОР | ||||
| КД244Г | 200 | 10 | 200 | 1.3 | 0.035 | КТ28-1 | ФОТОН | ||
| КД245А | 400 | 10 | 200 | 1.4 | 0.07 | КД23 | ФОТОН | ||
| КД247Г | 600 | 1 | 150 | 1.3 | 0.15 | КД4Б | ТОР | ||
| КД248А | 1000 | 3 | 100 | 1.4 | 0.25 | КД16 | ФОТОН | ||
| КД249А | 40 | 3 | 0.48 | КД7Е | диод Шоттки | ||||
| КД251Е | 100 | 10 | 200 | 1 | ТОР | ||||
| КД253А | 800 | 3 | 100 | 1.5 | 0.22 | КД16 | ФОТОН | ||
| КД255А5 | ТОР | ||||||||
| КД257Д | 1000 | 1.5 | 150 | ТОР | |||||
| КД265А | 1000 | 1 | 1.5 | 0.2 | ФОТОН | ||||
| КД266А | 200 | 0.5 | 1.4 | 0.1 | ФОТОН | ||||
| КД267А | 1000 | 10 | 1.7 | 0.1 | ФОТОН | ||||
| КД268A | 30WQ03F | ЭЛЕКТРОНИКА | |||||||
| КД268Л1 | 400 | 3 | 200 | 1.1 | 0.05 | КТ28 | ЭЛЕКТРОНИКА | диод Шоттки | |
| КД268Е|ЕС | 200 | 6 | 2х3 | 1.0 | КТ28-1 | КТ28-2 | ВЗПП-С | АДБК.432120.380ТУ | |||
| КД269Е|ЕС|ЕС91 | 200 | 10 | 2х5 | 1.0 | КТ28-1 | КТ28-2 | КТ90 | ВЗПП-С | АДБК.432120.381ТУ | |||
| КД270Е | ЕС | 200 | 15 | 2х7,5 | 1,0 | КТ28-1 | КТ28-2 | ВЗПП-С | АДБК.432120.382ТУ | |||
| КД271Е|ЕС|ЕС91 | 200 | 20 | 2х10 | 1.0 | КТ28-1 | КТ28-2 | КТ90 | ВЗПП-С | АДБК.432120.383ТУ | |||
| КД272Е|ЕС|ЕС91 | 200 | 30 | 2х15 | 1.2 | КТ28-1 | КТ28-2 | КТ90 | ВЗПП-С | АДБК.432120.384ТУ | |||
| КД273Е|ЕС|ЕС91 | 200 | 40 | 2х20 | 1.15 | КТ28-1 | КТ28-2 | КТ90 | ВЗПП-С | АДБК.432120.385ТУ | |||
| КД275Е | 800 | 2.2 | 1.4 | 0.25 | капD=7 | ФОТОН | |||
| КД278АС | ТОР | ||||||||
| 2Д288ДС | 200 | 10 | 500 | 1.0 | КТ-28-2 | ЭЛТОМ | пара ОК Шоттки | ||
| КД289Е|ЕС | 200 | 1 | 2х1 | 1.0 | КТ28-1 | КТ28-2 | ВЗПП-С | АДБК.432120.714ТУ | |||
| КД290Е|ЕС | 200 | 4 | 2х2 | 1.0 | КТ28-1 | КТ28-2 | ВЗПП-С | АДБК.432120.715ТУ | |||
| КД296К | 5KQ100 | 100 | 5.0 | 0.79 | КТ28-1 | диод Шоттки | |||
| КД297ВС|91 | MBR(B)15100 | 100 | 2×7.5 | 0.85 | КТ28-2|КТ90 | ТРАНЗИСТОР | пара ОК Шоттки | ||
| КД298ВС | 100 | 2×5 | 0.8 | КТ28-2/КТ90 | ТРАНЗИСТОР | пара ОК Шоттки | |||
| КД2117В-5 | 1200 | 100 | 2.5 | 0.2 | б/к | СИТ | |||
| 2Д2136АС | 600 | 5 | 1.2 | б/к | КРЕМНИЙ | мост АЕЯР.432120.479ТУ | |||
| 2Д2137АС | 1000 | 5 | 1.5 | б/к | КРЕМНИЙ | 3ф.мост АЕЯР.432120.479ТУ | |||
| 2Д2138АС9 | 200 | 0,1 | 1.9 | КТ48 | ПЛАНЕТА | диод+стабилитрон | |||
| КД2149Б9 | 200 | 1 | 0.95 | 12 | 4601.3-1 | КРЕМНИЙ | fast диод ЮФ3.438.070-04ТУГК | ||
| 2Д2160А9 | 300 | 1 | 1.0 | 1 | КТ47 | ПЛАНЕТА | |||
| 2Д2187А|А19 | 1200 | 5 | 1.0 | КТ28А-2,02 | КТ28-1 | КРЕМНИЙ | ||||
| 2Д2946АС | 600 | 30 | 1.4 | КРЕМНИЙ | мост АЕЯР.432120.480ТУ | ||||
| 2Д2947АС | 1000 | 30 | 1.7 | КРЕМНИЙ | мост АЕЯР.432120.480ТУ | ||||
| КД2957В | 60EPS12 | 1200 | 60 | 1.1 | КТ43-1 | ||||
| КД2958В | 40EPS12 | 1200 | 40 | 1.1 | КТ43-1 | ||||
| КД2959В | 1200 | 30 | 1.15 | КТ43-1 | |||||
| КД2960В | 20ETS12 | 1200 | 20 | 1.1 | КТ28-1 | ||||
| КД2961Г | 1600 | 10 | 1.1 | КТ28-1 | |||||
| КД2970А | MBR10100 | 100 | 10 | 0.6 | КТ28-1 | ТРАНЗИСТОР | диод Шоттки | ||
| КД2972Б2 | 50 | 35 | КТ28-1 | ТРАНЗИСТОР | трансил | ||||
| КД2990А | 400 | 20 | 200 | 1.4 | 0.15 | КД23 | ФОТОН | # | |
| КД2991А | 45 | 60 | 200 | 0.8 | ТОР | диод Шоттки | |||
| КД2992А | SF304 | 200 | 30 | 100 | 1 | 0.1 | КД23 | ФОТОН, ПУЛЬСАР | |
| КД2993А | 200 | 20 | 100 | 1 | 0.2 | КД23 | ФОТОН | ||
| КД2994А | 100 | 20 | 200 | 1 | 0.2 | КТ28 | ФОТОН | # | |
| КД2995Д | 200 | 30 | 200 | 1.1 | 0.05 | ТОР | # | ||
| КД2997А | 200 | 30 | 100 | 1 | 0.2 | КД23 | ФОТОН | # | |
| КД2998Д | 30 | 30 | 200 | 0.6 | ТОР | # диод Шоттки | |||
| КД2999А | 200 | 20 | 100 | 1 | 0.2 | КД23 | ФОТОН | # | |
- Пояснения:
- Диоды отмеченные «#» описаны в справочнике: Диоды; Радио и Связь, М.,1990 /МРБ1158/
- Буквенный суффикс соответствует, как правило, максимальному обратному напряжению.
Фруктоза-1,6-бисфосфатаза препятствует прогрессированию рака почки
Abstract
Светлоклеточный почечно-клеточный рак (ccRCC), наиболее частая форма рака почки 1 , характеризуется повышенным содержанием гликогена и отложением жира 2 . Эти последовательные метаболические изменения связаны с нормоксической стабилизацией факторов, индуцируемых гипоксией (HIFs) 3 , вторичных по отношению к мутациям von hippel-lindau ( VHL ), которые встречаются более чем в 90% опухолей ccRCC 4 .Однако почечно-специфическая делеция VHL у мышей не вызывает метаболических фенотипов, специфичных для ccRCC, и образования опухолей 5 , указывая на то, что необходимы дополнительные механизмы. Недавний крупномасштабный анализ секвенирования выявил потерю нескольких ферментов ремоделирования хроматина в подмножестве ccRCC ( полибром 1 [ PBRM1 ] ~40%, домен SET, содержащий 2 [ SETD2 ] ~15%, связанный с BRCA1. белок-1 [ BAP1 ] ~15% и т. д.) 6–9 , что указывает на то, что эпигенетические нарушения, вероятно, вносят важный вклад в естественное течение этого заболевания. Здесь мы использовали интегративный подход, включающий панметаболическое профилирование и анализ метаболического набора генов, и определили, что глюконеогенный фермент фруктоза-1,6-бисфосфатаза 1 (FBP1) 10 равномерно истощен в более чем шестистах исследованных опухолях ccRCC. Важно отметить, что локус FBP1 человека находится на хромосоме 9q22, потеря которой связана с плохим прогнозом для пациентов с ccRCC 11 .Наши данные также показывают, что FBP1 ингибирует прогрессирование ccRCC посредством двух различных механизмов: 1) FBP1 противодействует гликолитическому потоку в эпителиальных клетках почечных канальцев, предполагаемых клетках происхождения ccRCC 12 , тем самым ингибируя потенциальный «эффект Варбурга» 13,14 , и 2) в pVHL-дефицитных клетках ccRCC FBP1 сдерживает клеточную пролиферацию, гликолиз и пентозофосфатный путь независимым от каталитической активности образом, ингибируя функцию ядерного HIF посредством прямого взаимодействия с HIF «ингибирующим доменом».Эта уникальная двойная функция белка FBP1 объясняет его повсеместную утрату при скПКР, отличая FBP1 от ранее идентифицированных опухолевых супрессоров ( PBRM1, SETD2, BAP1 и т. д.), которые не всегда мутируют во всех опухолях 6,7,15 .
Мы провели панметаболомный анализ ( Methods ) 20 первичных опухолей человека ccRCC и соответствующих нормальных тканей почек. Уровни метаболитов, участвующих в гликолизе, глюконеогенезе и метаболизме сахара, связанном с глюкозой, были сильно повышены в опухолях, что свидетельствует о том, что перепрограммирование метаболизма глюкозы имеет решающее значение для прогрессирования скПКР (4).Кроме того, анализ набора метаболических генов 16 по данным TCGA ccRCC RNAseq показал, что группа «запасающих углеводы» была наиболее значительно недоэкспрессированной группой генов в опухолях ccRCC (), включая три гена, контролирующих почечный глюконеогенез 17 ( глюкоза-6- фосфатаза, каталитическая субъединица [ G6PC ], фосфоенолпируваткарбоксикиназа 1 [ PCK1 ] и фруктоза-1,6-бисфосфатаза 1 [ F121 90]). Повышенная активность HIF в опухолях ccRCC стимулирует аэробный гликолиз за счет увеличения экспрессии гликолитических генов, в том числе фосфоглицераткиназы 1 ( PGK1 ) и лактатдегидрогеназы A ( LDHA ), и шунтирования гликолитического потока от цикла TCA. активация киназы пируватдегидрогеназы 1 ( PDK1 ) 3 .Однако наш интегративный анализ выявил подавление глюконеогенеза как дополнительный компонент регуляции уровня глюкозы при скПКР. Затем мы определили, что ограничивающий скорость глюконеогенный фермент FBP1 был ингибирован на уровне накопления белка почти в 100% исследованных опухолей ccRCC (n> 200, и ) по сравнению с нормальной тканью почки. Сходные результаты наблюдались для гепатоцеллюлярной карциномы и нормальной ткани печени (4). Ингибирование FBP1 не опосредовано активацией HIF, связанной с ccRCC, потому что абляция HIF1α не смогла дерепрессировать экспрессию FBP1 в клетках RCC4 ccRCC ().Более того, клетки проксимальных канальцев HK-2 (из которых, по-видимому, произошли ccRCC 12 ) обнаруживают HIF1α-зависимую индукцию FBP1 при гипоксии (14). По сравнению с FBP1, два других глюконеогенных фермента G6PC и PCK1 были либо умеренно подавлены (G6PC), либо не проявляли устойчивых изменений (PCK1) в опухолях ccRCC (4). Интересно, что гликолитический фермент фосфофруктокиназа (печеночного типа, PFKL), который функционально противостоит FBP1 при гликолизе (1), экспрессировался на равных уровнях в тканях скПКР и нормальных тканях почек (4).Кроме того, более низкая экспрессия FBP1 значительно коррелирует с поздней стадией опухоли и худшим прогнозом пациента (1), тогда как экспрессия PFKL не имеет (2), что позволяет предположить, что FBP1 может выполнять новые неферментативные функции.
Интегративный анализ показывает, что FBP1 повсеместно ингибируется и проявляет опухолесупрессивные функции при ccRCCa , Анализ метаболического набора генов данных RNAseq, предоставленных проектом TCGA ccRCC (http://cancergenome.nih.gov). Было включено 480 опухолей ccRCC и 69 соседних нормальных тканей. 2752 гена, кодирующие все известные метаболические ферменты и транспортеры человека, были классифицированы в соответствии с KEGG (http://www.genome.jp/kegg/). Сгенерированные наборы метаболических генов были ранжированы на основе их средней кратности изменений экспрессии в опухоли ccRCC по сравнению с нормальной тканью и нанесены на график как медиана ± среднее абсолютное отклонение. b , Иммуногистохимическое окрашивание репрезентативного микрочипа ткани почки с антителом FBP1. T: опухоли скПКР; N: соседняя нормальная почка. c , нормализованные чтения RNASeq FBP1 в 69 нормальных почках и 480 опухолях ccRCC, сгруппированных по стадии I–IV по TCGA. d , кривая выживаемости Каплана-Мейера для 429 пациентов с ccRCC, зарегистрированных в базе данных TCGA. Пациенты были поровну разделены на две группы (верхняя и нижняя 50% экспрессии FBP1 ) на основе уровней экспрессии FBP1 в их опухолях. e , Рост клеток 786-O ccRCC в среде с низким содержанием сыворотки (1% FBS), с эктопической экспрессией FBP1 или без нее. f , Ксенотрансплантат опухолевого роста клеток 786-O с эктопической экспрессией FBP1 или без нее. Вес конечной точки опухоли измеряли и наносили на график. г , Рост клеток HK-2 проксимальных отделов почечных канальцев с ингибированием или без ингибирования FBP1 в 1% сывороточной среде. Значения представляют собой среднее ± стандартное отклонение. (четыре технические повторности, из двух независимых экспериментов). *р<0,01.
Чтобы исследовать функциональную роль FBP1 в прогрессировании ccRCC, мы эктопически экспрессировали FBP1 в опухолевых клетках 786-0 ccRCC до уровней, наблюдаемых в клетках проксимальных канальцев HK-2 ().Экспрессия FBP1 значительно ингибировала 2D-культуру (), независимый от прикрепления () и ксенотрансплантатный рост опухоли (и). Сходным образом усиленная экспрессия FBP1 ингибировала рост клеток RCC10 и 769-P ccRCC (14) и клеток рака легкого A549 преимущественно в условиях гипоксии (20). Эти результаты показали, что FBP1 может подавлять рост ccRCC и других опухолевых клеток, эффект значительно выражен в сочетании с активацией HIF. В клетках HK-2 истощения FBP1, но не абляции G6PC или эктопической экспрессии PFKL, было достаточно для стимулирования роста клеток HK-2 (и ).
Поскольку FBP1 является ферментом, ограничивающим скорость глюконеогенеза 10 , мы манипулировали экспрессией FBP1 в почечных клетках и измеряли метаболизм глюкозы. Ингибирование FBP1 увеличивало поглощение глюкозы и секрецию лактата в клетках HK-2, культивируемых в 10 мМ глюкозе (1), эффект усиливался при снижении уровня глюкозы до 1 мМ (3). Чтобы оценить гликолитический поток, мы провели анализ распределения изотопомеров, используя [1, 2- 13 C] глюкозу в качестве индикатора, который продуцирует промежуточные продукты гликолиза и TCA, содержащие два атома 13 C (виды M2), а также соответствующие виды M1. по пентозофосфатному пути (ПФП).Мы наблюдали повышенное обогащение M2 четырьмя промежуточными соединениями TCA (малатом, аспартатом, глутаматом и цитратом) в клетках HK-2, истощенных по FBP1. Напротив, ингибирование G6PC не способствовало обмену глюкозы и лактата (данные не показаны), что свидетельствует о том, что FBP1, но не G6PC, является критическим регулятором метаболизма глюкозы в почечных клетках. В соответствии с этим результатом, эктопическая экспрессия FBP1 в VHL -дефицитной клеточной линии ccRCC (RCC10) снижала поглощение глюкозы, секрецию лактата и производные глюкозы промежуточные продукты цикла TCA (11).Известно, что снижение глюкозозависимого потока ТСА увеличивает анаплеротический поток глутамина 18 , и мы также наблюдали повышенное поглощение глутамина и обогащение производными глутамина промежуточными продуктами цикла ТСА (виды M4) в [U- 13 C] меченных глутамином RCC10. клетки, экспрессирующие FBP1 (+).
FBP1 регулирует гликолиз и уровни NADPHa , поглощение глюкозы и секрецию лактата в клетках HK-2 с ингибированием FBP1 или без него. Распределение изотопомеров M2 указанных метаболитов ( b ) и цитрата ( c ) в клетках HK-2 с или без абляции FBP1, меченных [1, 2- 13 C] глюкозой.Обогащение M2 в процентах представляет собой молярный процент избытка видов M2 над естественной численностью. d , Поглощение глюкозы и секреция лактата в клетках RCC10 и RCC10VHL, эктопически экспрессирующих вектор или FBP1. Клетки RCC10VHL представляют собой клетки RCC10, в которые была повторно введена pVHL дикого типа. e , распределение изотопомеров M2 указанных метаболитов в клетках RCC10, экспрессирующих вектор или FBP1, меченных [1, 2- 13 C] глюкозой. f , Относительные уровни NADPH в клетках RCC10 и RCC10VHL, как указано в ( d ).Значения представляют собой среднее ± стандартное отклонение. (три экспериментальные повторности). *р<0,05.
Панметаболический анализ опухолей ccRCC выявил заметное повышение уровня восстановленного глутатиона (G-SH) 19 (). Для синтеза G-SH требуется восстановленная форма никотинамидадениндинуклеотидфосфата (NADPH), генерируемая главным образом посредством PPP в клетках человека 19 (). В соответствии с повышенным потоком PPP, в опухолях ccRCC наблюдается значительное накопление G-SH и связанных с PPP метаболитов (2), эффект частично повторяется в клетках HK-2, истощенных по FBP1 (2).Напротив, повторная экспрессия FBP1 в клетках RCC10 значительно снижала уровни NADPH и поток PPP (и ). Интересно, что FBP1-опосредованные изменения в потоке PPP (1) были сравнимы с изменениями в глюкозо-6-фосфате (G6P; ), входном метаболите пути PPP (2), что свидетельствует о том, что FBP1 влияет на поток PPP в первую очередь посредством регуляции гликолиза. Удивительно, но способность FBP1 снижать гликолиз и уровни NADPH была полностью устранена в клетках RCC10VHL (), где pVHL дикого типа была введена в RCC10, чтобы исключить нормоксическую экспрессию HIF (), что указывает на то, что белки HIF необходимы для опосредованных FBP1 эффектов на клетки RCC10. Метаболизм глюкозы в опухолевых клетках скПКР.
Чтобы исследовать механистическую связь между экспрессией FBP1 и активностью HIF, мы использовали две VHL -дефицитные линии ccRCC, RCC4 и RCC10, которые экспрессируют как HIF1α, так и HIF2α (). HIF1α и HIF2α индуцируются на разных стадиях ccRCC и играют взаимодействующие и противоположные роли в опухолевой прогрессии 3 . HIF1α и HIF2α функционируют путем связывания элементов реакции на гипоксию (HREs) в пределах генов-мишеней, включая те, которые модулируют клеточный метаболизм -20-. Интересно, что эктопическая экспрессия FBP1 подавляет активность HIF (1) и способствует потреблению кислорода в клетках RCC4 и RCC10 (4).Кроме того, экспрессия FBP1 в клетках RCC10 восстанавливала активность PDH (1), которая иначе ингибировалась HIF1α 3 . Наоборот, абляция FBP1 повышала активность HIF в клетках RCC10, которые экспрессируют обнаруживаемые уровни FBP1 (и ). Эта обратная корреляция между экспрессией FBP1 и активностью HIF была повторена в первичных опухолях ccRCC (). Напротив, экспрессия G6PC не коррелировала с активностью HIF в клетках ccRCC или в опухолевых тканях (4). Интересно, что FBP1 также ингибировал активность HIF в клетках рака легкого A549, культивируемых при 0.5% O 2 (), демонстрируя, что этот эффект не специфичен для почечных клеток. Более того, экспрессия FBP1 снижала уровни канонической мишени HIF ( PDK1, LDHA, транспортер глюкозы 1 [ GLUT1 , также известный как SLC2A1 ] и фактор роста эндотелия сосудов [ VEGF ]), уровни мРНК RCC1 при ПКР. и гипоксические клетки A549, но не в нормоксических клетках RCC10VHL (и). Анализ иммунопреципитации хроматина (ChIP) показал, что FBP1 был обогащен HRE промоторов PDK1, LDHA, GLUT1 и VEGF , но не в нечувствительном к гипоксии рибосомном белке L13A ( RPL13A 9001) ( RPL13A 9001) ).Анализы ChIP-reChIP выявили совместную локализацию HIF1α и FBP1 в этих HRE (), что позволяет предположить, что FBP1 непосредственно ингибирует HIF в ядре, вывод, подтвержденный клеточным фракционированием и иммунофлуоресцентным окрашиванием первичной ткани почки человека (и). Кроме того, форма FBP1 с исключенным ядром (FBP1 NES), содержащая мощную экспортную последовательность ядра 21 , слитую с С-концом FBP1 (1), не способна ингибировать экспрессию гена-мишени HIF так же эффективно, как FBP1 дикого типа (2). Экспрессия FBP1 NES не подавляла рост клеток RCC10 и не изменяла оборот глюкозы и лактата, в отличие от FBP1 дикого типа (и ).В совокупности эти данные показали, что ядерный FBP1 необходим для ингибирования HIF и метаболизма глюкозы в VHL -дефицитных клетках ccRCC ().
FBP1 ингибирует активность HIF в ядреa , репортерная активность HIF, измеренная в клетках RCC4 и RCC10, трансфицированных pHRE-люциферазой, в присутствии вектора, кДНК FBP1 или двух разных кшРНК FBP1. Эффективность трансфекции нормализовали относительно котрансфицированной pRenilla-люциферазы. b , 480 опухолей ccRCC из базы данных TCGA были поровну разделены на две группы (верхняя и нижняя 50% экспрессии FBP1 ) на основе уровней экспрессии FBP1 , и их относительная активность HIF была количественно определена и нанесена на график, как описано в Методах . .c , репортерная активность HIF в гипоксических клетках RCC4 и A549 (0,5% O 2 ) с эктопической экспрессией FBP1 или без нее. d , qRT-PCR анализ генов-мишеней HIF в клетках RCC10, экспрессирующих вектор или FBP1. e , анализы ChIP, оценивающие связывание хроматина FBP1 с HRE в промоторе GLUT1 или с нечувствительным к гипоксии участком локуса RPL13A . В качестве положительного контроля использовали антитело к РНК-полимеразе II. f , Иммунофлуоресцентное окрашивание первичной ткани почки человека (тубулярная область) антителом FBP1.Стрелки указывают на три репрезентативных участка с ядерным FBP1. IgG кролика использовали в качестве отрицательного контроля, а DAPI — флуоресцентный ядерный краситель. г , Рост клеток RCC10, экспрессирующих вектор, FBP1 или FBP1 NES (FBP1, связанный с С-концевой ядерной экспортной последовательностью), культивируемых в 1% сыворотке. Столбики погрешностей представляют sd. (три экспериментальных повтора), за исключением ( e ), который указывает с.э.м. (три технических повторения из репрезентативного эксперимента). *р<0,05.
Чтобы определить, необходима ли ферментативная активность FBP1 для ингибирования HIF, мы экспрессировали ранее описанный каталитически неактивный мутант FBP1 G260R 22,23 (и ) в клетках RCC10 и 786-O ().FBP1 G260R ингибировал рост клеток до сопоставимого уровня FBP1 дикого типа при 10 мМ глюкозы (и ). FBP1 также ингибирует метаболизм глюкозы, продукцию NADPH и экспрессию гена-мишени HIF в той же степени, что и FBP1 дикого типа в клетках RCC10 (10). Эти результаты свидетельствуют о том, что FBP1 вмешивается в функцию HIF через механизм, независимый от каталитической активности. В нормоксических клетках RCC10VHL (низкая активность HIF) способность мутанта FBP1 G260R ингибировать рост клеток, метаболизм глюкозы, продукцию NADPH и экспрессию гена-мишени HIF была устранена (), что еще раз подтверждает, что FBP1 влияет на метаболизм и рост клеток ccRCC, регулируя HIF, независимо от его ферментативной активности.Тем не менее, FBP1 дикого типа подавлял рост клеток RCC10 и 786-O () сильнее, чем мутант G260R в условиях низкого уровня глюкозы, по-видимому, потому, что ферментативное ингибирование гликолиза FBP1 дикого типа более выражено, когда поступление глюкозы становится ограниченным.
FBP1 ингибирует HIF независимо от его ферментативной активности посредством прямого взаимодействия с «ингибирующим доменом» HIFαa , Кристаллическая структура (идентификатор PDB ID: 1EYJ) свиного FBP1 в комплексе с AMP (синий) и фруктозо-6-фосфатом (F6P, красный).N-концевой регуляторный домен FBP1 окрашен в зеленый цвет, а С-концевой каталитический домен — в фиолетовый. Остаток G260 выделен желтым цветом. b , Рост клеток RCC10, эктопически экспрессирующих вектор, FBP1 или FBP1 G260R в 1% сывороточной среде. c , репортерная активность HIF в клетках RCC10, экспрессирующих вектор, FBP1, FBP1 G260R, регуляторный домен FBP1 (домен R) и каталитический домен FBP1 (домен C). Лизаты клеток RCC10 подвергали иммунопреципитации с помощью IgG, антитела HIF1α ( d ) или антитела HIF2α ( e ) и подвергали блоттингу на эндогенный FBP1. f , анализ GST pull-down между рекомбинантным FBP1 и рекомбинантным GST или HIF1α, меченным GST. ИБ: иммуноблот. g , анализ GST pull-down между рекомбинантным HIF1α и рекомбинантным GST-меченым укорочением экзона FBP1. h , анализ GST pull-down между рекомбинантным FBP1 и GST-мечеными мотивами HIF1α. Значения представляют собой среднее ± стандартное отклонение. (три экспериментальные повторности). *р<0,01.
Для изучения молекулярного механизма (механизмов), с помощью которого FBP1 ингибирует активность HIF, мы разделили белок FBP1 на N-концевой регуляторный («R») домен, содержащий аллостерические регуляторные сайты, и C-концевой («C») домен, содержащий каталитический центр ().Интересно, что эктопической экспрессии домена «R» FBP1 в клетках RCC10, RCC4 и 786-O было достаточно для ингибирования активности HIF, тогда как экспрессии домена «C» не было ( и ). Поскольку клетки 786-O экспрессируют HIF2α, но не функциональный HIF1α 24 , мы заключаем, что FBP1 ингибирует как HIF1α, так и HIF2α, предположительно посредством сходного механизма. Для дальнейшего картирования критических областей FBP1 для распознавания HIF мы систематически удаляли каждый экзон из полноразмерного FBP1 (10). Все семь укорочений FBP1 проявляли минимальную каталитическую активность (1), тогда как только N-концевые укорочения экзона 1 и экзона 2 значительно потеряли свою способность ингибировать HIF (1).
Далее мы продемонстрировали ассоциацию FBP1 и HIF1α с помощью ко-иммунопреципитации меченых эпитопом и/или эндогенных белков из лизатов клеток 293T или RCC10 ( и ). FBP1 также связан с HIF2α (и ), но не с PHD2 или FIh2, двумя хорошо задокументированными регуляторами HIFα 3 (). Интересно, что анализ GST pull-down показал, что белки HIF1α или HIF2α напрямую связываются с полноразмерным FBP1 (и ), а взаимодействие между HIF1α и FBP1 зависит от экзона 1 или экзона 2 FBP1 ().Кроме того, FBP1 ассоциируется с относительно не охарактеризованным «ингибиторным доменом» (ID) HIF 25 (и ). Чтобы проверить, ингибирует ли FBP1 активность HIF посредством распознавания ID, мы заменили ДНК-связывающий домен HIFα ДНК-связывающим доменом GAL4 (GBD) и провели анализы трансактивации GAL4 () (подробности см. в Methods ). В соответствии с репортерными анализами HIF (, и), FBP1 подавлял активность полноразмерного HIF1α-GBD примерно на 50% (красная колонка). Важно отметить, что удаление HIF1α ID в значительной степени ослабляло ингибирующий эффект FBP1 (1).В HIF2α критическая область, опосредующая ингибирование FBP1, распространяется на весь С-конец. Следовательно, FBP1 подавляет активность HIF1α и HIF2α, взаимодействуя с их С-концевыми областями, особенно с мотивом ID.
Помимо потери VHL , ccRCC демонстрируют значительную генетическую гетерогенность 26 . Недавний крупномасштабный анализ выявил частые мутации в трех эпигенетических генах PBRM1, SETD2 и BAP1 , каждый из которых находится в области 43 Mb на хромосоме 3p, которая охватывает VHL 6–9 .Гистологически ccRCC характеризуется фенотипом «светлых клеток», возникающим в результате накопления гликогена и липидов 2 , что позволяет предположить, что метаболические нарушения являются определяющей чертой этих опухолей. Здесь мы демонстрируем, что глюконеогенный фермент FBP1 повсеместно истощен в ccRCC, что согласуется с нашими предыдущими анализами количества копий 27 . Более того, FBP1 проявляет двойную опухолесупрессирующую функцию, опосредованную двумя отдельными доменами, что объясняет универсальную потерю экспрессии FBP1 в опухолях ccRCC.В совокупности наши данные показывают интригующую регулирующую связь между FBP1 и гипоксическими реакциями при карциноме почки, которая имеет значение для метаболической регуляции всех глюконеогенных тканей (дополнительное обсуждение).
Методы
Антитела и реагенты
Антитела, определяющие HDAC1 (5356), HSP90 (4877), GST (2625), HA (3724) и нормальные изотопные контроли IgG (кролик-2729, мышь-5415) были приобретены у компании Cell Сигнальные технологии. Антитела, определяющие HIF1α (NB100-134) и HIF2α (NB100-122), были получены от Novus, а антитела против PCK1 (ab28455), G6PC (ab83690) и PFKL (ab37583) были получены от Abcam.D-фруктоза-1, тринатриевая соль 6-бисфосфата (47810), 2-гидроксиэтилагароза (A4018) и антитела, детектирующие FBP1 (SAB1405798 и HPA005857), были приобретены у Sigma.
Культура клеток
Клетки 786-O, RCC4, RCC10, 769-P, A549 и 293T были проверены на отсутствие микоплазмы и культивированы в среде DMEM, содержащей 10% FBS и антибиотики. Клетки эпителия проксимальных канальцев HK-2 культивировали в среде Keratinocyte-SFM с добавлением человеческого рекомбинантного эпидермального фактора роста 1–53 и экстракта бычьего гипофиза (Life Technologies).Гипоксические условия (0,5% O 2 ) были достигнуты на рабочей станции Ruskinn in viv O2 400 путем добавления в окружающий воздух сбалансированных N 2 и CO 2 . Для анализа метаболического мечения клетки поддерживали в среде DMEM без глюкозы (Life Technologies) с добавлением 10% диализированного FBS (Gemini Bio Products) и 10 мМ [1, 2- 13 C] глюкозы (Кембриджский изотоп) или глутамин- свободный DMEM (Life Technologies) с добавлением 10% диализированного FBS и 2 мМ [U- 13 C] глутамина (Кембриджский изотоп).
Конструкции
Репортерную конструкцию HRE (элемент реакции на гипоксию) получали путем введения трех повторов мотивов HRE, полученных из промотора PGK1 человека, перед кассетой экспрессии люциферазы в векторе pGL2-TK. Плазмиды ShRNA, нацеленные на человеческие мРНК FBP1, G6PC и HIF1A , были приобретены у Open Biosystems. Антисмысловые последовательности короткой шпильки против человеческого FBP1 представляют собой 5’-ATGTTGGAAGATCCATCAAGG-3’ (SH-1) и 5’-AACATGTTCATAACCAGGTCG-3’ (SH-2).Антисмысловая последовательность против человеческого G6PC представляет собой 5′-TTCAAGGAGTCAAAGACGTGC-3′, а последовательности против человеческого HIF1A представляют собой 5′-TAACTTCACAATCATAACTGG-3′ (SH-1) и 5′-ATTCGGTAATTCTTTCATCAC-3′ (SH-2). . Экспрессионные плазмиды FBP1, G6PC и PFKL конструировали путем клонирования открытой рамки считывания каждой кДНК в сайт множественного клонирования вектора PCDNA3.1-V5. Мутант FBP1 G260R был получен с использованием набора для мутагенеза QuikChange II компании Stratagene (Agilent). FBP1 NES был получен путем связывания эффективной ядерной экспортной последовательности (LALKLAGLDIGS) с С-концом FBP1 его экспрессионной кассеты.Укорочения экзонов FBP1 были получены путем субклонирования следующих кодирующих областей из полноразмерного FBP1 (1–338): ΔE1 (58–338), ΔE2 (1–57, 112–338), ΔE3 (1–111, 143–338) , ΔЕ4 (1–142, 189–338), ΔЕ5 (1–188, 236–338), ΔЕ6 (1–235, 276–338), ΔЕ7 (1–275). Полноразмерные FBP1, HIF1α и HIF2α клонировали в вектор pGEX-6P-1 для создания белковых конструкций, меченных GST. Различные GST-меченые мотивы HIF1α были созданы путем субклонирования следующих областей из полноразмерного HIF1α (1–826) в вектор pGEX-6P-1: bHLH (1–80), PASA (81–235), PASB (236–236). 329), ЛИНК (330–392), НОДД (393–531), НТАД (532–603), ИД (604–786), СТАД (787–826).Для анализов трансактивации GAL4 последовательности HIFα, в которых отсутствует мотив связывания ДНК bHLH, клонировали в вектор pBIND (Promega), который находится в рамке считывания с ДНК-связывающим доменом GAL4 (GBD) на N-конце. Серия усечений GBD-HIF1α была создана путем удаления указанных мотивов HIF1α. Точно так же укорочения GBD-HIF2α были созданы путем индивидуальной делеции следующих мотивов из последовательностей HIF2α (1–870): PASA (80–236), PASB (237–331), LINK (332–395), NODD (396–495). , NTAD (496–581), ID (582–830), CTAD (831–870).Конструкции GFP-FIh2 и GFP-PHD2 были получены от доктора Эрика Метцена через Addgene.
Трансфекция плазмид и вирусная инфекция
Экспрессионные конструкции трансфицировали в линии раковых клеток с использованием реагента Lipofectamine LTX (Life Technologies) в соответствии с протоколом производителя. Лентивирус был получен путем совместной трансфекции клеток 293T векторами экспрессии pRSV-Rev, pMD2.G, pMDLg/pRRE и pLKO.1-puro shRNA или pCDH-puro. Вирус собирали через 48 часов фильтрованием вируссодержащей среды через 0.Фильтр Steriflip 45 мкМ (Millipore). Заражение вирусом осуществляли путем инкубации клеток со средой, содержащей указанный вирус и 8 мкг/мл полибрена (Sigma) в течение 24 часов. Клетки восстанавливались в полной среде в течение 24 часов, а затем отбирались с помощью пуромицина в течение 24 часов (анализ мРНК) или 48 часов (анализ белков и фенотип). Выжившие пулы подвергали указанным экспериментам. Обратите внимание, что клеткам HK-2, подвергшимся вирусной инфекции и отбору пуромицином, обычно требуется 10–14 дней для восстановления перед тем, как их подвергнуть экспериментам.
Анализ TCGA RNAseq
Необработанные данные RNAseq для 480 опухолей ccRCC и 69 нормальных тканей почек были загружены из проекта TCGA ccRCC (http://cancergenome.nih.gov) 2 апреля 2013 г. Данные были проанализированы на дифференциальную экспрессию генов с помощью DeSeq (Bioconductor, версия 2.12). Были экспортированы нормализованные подсчеты, значения p, значения p с поправкой на частоту ложных открытий (p-adj) и кратность изменения экспрессии для каждого гена. Для проведения анализа набора генов полный список из 2752 генов, кодирующих все известные метаболические ферменты и переносчики человека, был сгруппирован в функциональные наборы в соответствии с метаболическими путями, аннотированными KEGG (http://www.genome.jp/kegg/). Пороговое значение p-adj было установлено равным 0,1, чтобы исключить гены, которые не были постоянно обнаружены с помощью RNAseq. Для изучения активности HIF, ассоциированной с опухолью, необработанные данные каждого секвенированного гена были перемасштабированы, чтобы установить медиану равной 1, и активность HIF была количественно определена путем усреднения нормализованной экспрессии 44 генов-мишеней HIF, включая те, которые кодируют IGFBP3, EDN2, PFKFB4, FLT1, TFR2, BNIP3L, TGFA, BNIP3, PGK1, EGLN1, LDHA, EGLN3, CP, TGFB3, PFKFB3, HK1, TFRC, EDN1, CDKN1A, CA9, ADM1, HMOX1, SERPINE1, LOX, NDRG1, CA12, PDK1, VEGFA, ERO1L, RORA, P4HA1, MXI1, SLC2A1(GLUT1), STC2, MIF, DDIT4, ENO1, CXCR4, PLOD1, P4HA2, GAPDH, PGAM1, TMEM45A и PIM1 28 .
Количественное определение метаболитов
Скорость поглощения глюкозы и секреции лактата определяли с помощью многопараметрической биоаналитической системы YSI 7100 (YSI Life Sciences). Уровни NAPDH измеряли с использованием набора для количественной оценки NADP + /NADPH (BioVision) в соответствии с протоколом производителя. Уровни глюкозо-6-фосфата (G6P) определяли с использованием набора для флуорометрического анализа PicoProb G6P (BioVision) в соответствии с протоколом производителя. Абсолютные уровни NADPH или G6P нормализовали по отношению к клеткам контроля вектора, установив уровни контроля вектора равными 1.Панметаболический анализ был выполнен на 20 первичных опухолях ccRCC и 20 соседних нормальных тканях почек (полученных из Cooperative Human Tissue Network) с помощью Metabolon. В частности, пробоподготовку проводили с использованием автоматизированной системы MicroLab STAR® (Hamilton). Стандарты добавляли перед первой стадией экстракции в целях контроля качества. Экстракты метаболитов разделяли на две фракции; одну фракцию анализировали с помощью газовой хроматографии-масс-спектрометрии (ГХ-МС), а другую с помощью жидкостной хроматографии-тандемной масс-спектрометрии (ЖХ-МС/МС).Образцы для анализа ГХ-МС высушивали в вакуумной сушке в течение 24 часов перед дериватизацией в атмосфере азота с обработкой бистриметилсилилтрифторацетамидом (БСТФА). Колонка ГХ содержала 5% фенила, а линейное изменение температуры составляло от 40 до 300°C за 16-минутный период. Образцы анализировали на быстросканирующем одноквадрупольном масс-спектрометре Thermo-Finnigan Trace DSQ с использованием ионизации электронным ударом. Часть этого анализа, связанная с ЖХ-МС/МС, была основана на УЭЖХ Waters ACQUITY UPLC и масс-спектрометре Thermo-Finnigan LTQ, который состоит из источника ионизации электрораспылением (ESI) и масс-анализатора с линейной ионной ловушкой (LIT).Экстракты образцов разделяли на две аликвоты, сушили и восстанавливали по отдельности в кислых или основных растворителях. Одну аликвоту анализировали в условиях, оптимизированных для кислых положительных ионов, а другую — в условиях, оптимизированных для основных отрицательных ионов, с использованием специальных колонок. Экстракты, восстановленные в кислых условиях, элюировали градиентом воды и метанола, содержащего 0,1% муравьиной кислоты, а основные экстракты элюировали водой и метанолом, содержащим 6,5 мМ бикарбоната аммония. Анализ МС чередовался между сканированием МС и МС/МС с использованием динамического исключения.Соединения идентифицировали путем сравнения с собственной метаболомической библиотекой с очищенными стандартами. Зарегистрировано более 3000 коммерчески доступных стандартных соединений для определения их аналитических характеристик. Комбинация хроматографических свойств и масс-спектров указывала на совпадение с конкретным соединением или изобарической единицей. Для исследований, охватывающих несколько дней, был включен этап нормализации данных, чтобы скорректировать отклонения, возникающие в результате различий в настройке инструментов.Для создания конечного отчета необработанные данные по каждому метаболиту были перемасштабированы, чтобы установить медиану, равную 1. Для попарного сравнения метаболитов, связанных с пентозофосфатным путем, были выполнены t-критерии Уэлча. Также сообщается значение q для описания частоты ложных обнаружений при множественном тестировании.
13 Анализ С-метаболического потока
Для измерения обогащения 13 С в промежуточных продуктах цикла ТХУ, [1, 2- 13 С], меченных глюкозой (48 часов при 37 °С) или [U- 13 C] клетки, меченные глутамином (3 часа при 37 °C), экстрагировали 4% хлорной кислотой (PCA), как описано ранее 29 .Вкратце, клетки дважды промывали PBS, а затем инкубировали с 4% PCA в течение 30 минут при 4 ° C на качающейся платформе. Экстракты клеток собирали и нейтрализовали с помощью 5М КОН. После центрифугирования надосадочные жидкости переносили на колонку AG-1 (100–200 меш, 0,5 × 2,5 см, Bio-rad) для обогащения органическими кислотами, глутаматом и аспартатом, которые затем превращали в трет-бутилдиметилсилильные производные. Изотопное обогащение изотопомеров глутамата 13 C контролировали с использованием ионов с m/z 432, 433, 434, 435, 436 и 437 для M0, M1, M2, M3, M4 или M5 (содержащих от 1 до 5 13 атомов C выше M0, естественной численности) соответственно.Изотопное обогащение изотопомеров аспартата 13 C контролировали с использованием ионов с m/z 418, 419, 420, 421 и 422 для M0, M1, M2, M3 и M4 (содержащих от 1 до 4 13 атомов C выше M0, естественное изобилие) соответственно. Изотопное обогащение лактата 13 C определяли с использованием ионов с m/z 261, 262, 263 и 264 для M0, M1, M2 и M3 (содержащих от 1 до 3 13 атомов C выше естественного содержания), соответственно. Изотопное обогащение 13 C изотопомеров пирувата контролировали с использованием ионов с m/z 259, 260, 261 и 262 для M0, M1, M2 и M3 (содержащих от 1 до 3 13 атомов C выше естественного содержания), соответственно.Изотопное обогащение 13 изотопомеров малата C оценивали с использованием ионов с m/z 419, 420, 421, 422 и 423 для M0, M1, M2, M3 и M4 (содержащих от 1 до 4 13 атомов C выше естественного содержания) , соответственно, и обогащение 13 C изотопомеров цитрата 13 C было проанализировано с использованием ионов с m/z 459, 460, 461, 462, 463, 464 и 465 для M0, M1, M2, M3, M4, M5 и M6 (содержащие от 1 до 6 13 атомов C сверх естественного содержания) соответственно. Изотопное обогащение изотопомеров 13 C рибозо-5-фосфата определяли с использованием системы ЖХ-МС, как описано ранее 29 .Для рибозы в режиме MRM мы измерили ионные пары 561-175, 562-175, 563-175, 564-175, 565-175 и 566-175 для определения М0, М1, М2, М3, М4 и М5 соответственно. (содержащие от 1 до 5 13 атомов C сверх естественного содержания).
Количественная оценка пентозофосфатного пути
Метаболический поток, проходящий через пентозофосфатный путь (PPP), был количественно определен на основе ранее описанного метода 30 с использованием [1, 2- 13 C] глюкозы в качестве метки .Вкратце, прямой гликолиз [1, 2- 13 C] глюкозы (без прохождения через PPP) дает лактат, меченый M2, в то время как поток, проходящий через окислительную часть PPP, удаляет первый углерод из глюкозы и высвобождает его в виде СО 2 . Полученные промежуточные продукты, меченые M1, возвращаются обратно в гликолиз для производства лактата, меченого M1, посредством неокислительного действия PPP. Соотношение лактата, меченого M1 и M2, указывает отношение потока, проходящего через PPP, к потоку, проходящему непосредственно через гликолиз.Таким образом, поток PPP рассчитывали на основе следующей формулы: поток PPP = скорость потребления глюкозы (измеренная с помощью анализатора YSI, как описано выше) × обогащение лактатом M1 / (обогащение лактатом M1 + обогащение лактатом M2). Рассчитанный поток PPP был затем нормализован для контрольных клеток-переносчиков путем установки потока контроля вектора равным 1. Важным предположением для этой модели является то, что большинство промежуточных продуктов M1, генерируемых PPP, возвращаются обратно в гликолиз, а не экспортируются для синтеза нуклеотидов 30 .Мы измерили изотопное распределение рибозо-5-фосфата с помощью ЖХ-МС и обнаружили, что обогащение рибозо-5-фосфата, меченого M1, было необнаружимым, подтверждая, что большинство меченых M1 промежуточных продуктов, образующихся в PPP, были рециркулированы обратно в гликолиз.
Анализ роста клеток
Множественные культуры клеток ccRCC или A549 высевали в 6-луночные планшеты с плотностью 5×10 4 клеток/лунку с добавлением DMEM, содержащей 1% FBS. Для анализов роста, проводимых в условиях низкого содержания глюкозы, клетки поддерживали в среде DMEM без глюкозы с добавлением 10% диализированного FBS и 1 мМ глюкозы.Каждый день собирали и подсчитывали один набор культур.
Анализ роста, не зависящего от прикрепления
Клетки 786-O, стабильно экспрессирующие векторный контроль или FBP1, высевали при плотности 6×10 3 клеток/мл в полной среде, содержащей 0,3% агарозы, на подложки, состоящие из среды, содержащей 0,6% агароза. Культуры подкармливали каждую неделю, и после трех недель роста колонии определяли количественно.
Иммунопреципитация и вестерн-блоттинг
Клетки лизировали буфером для лизиса клеток млекопитающих, состоящим из 25 мМ Трис, 100 мМ NaCl, 1% Тритон Х-100, 1 мМ ЭДТА, 1 мМ ДТТ, 1 мМ NaVO 4 , 1 мМ β-глицеринфосфат и смесь ингибиторов протеазы (Roche Diagnostics).Для экспериментов по иммунопреципитации клетки собирали в буфере для лизиса, содержащем 0,3% CHAPS вместо 1% Triton X-100 в качестве детергента. 2 мг цельноклеточных лизатов предварительно очищали с помощью 30 мкл гранул с белком G (Life Technologies), а затем инкубировали с 2 мкг соответствующего изотипа контрольного IgG или указанных антител в течение 2 часов на качающейся платформе. Добавляли 50 мкл гранул с белком G и перемешивали еще 2 часа. Иммунопреципитаты собирали центрифугированием и затем разделяли с помощью SDS-PAGE. Разделенные белки переносили на нитроцеллюлозные мембраны и фильтры инкубировали с указанными первичными антителами, разведенными в TBST (20 мМ Трис, 135 мМ NaCl и 0.02% Tween 20) с добавлением 5% бычьего сывороточного альбумина (Sigma). Первичные антитела выявляли с помощью вторичных антител, конъюгированных с пероксидазой хрена (Cell Signaling Technology), с последующим воздействием реагентов ECL (Pierce).
Количественная ОТ-ПЦР
Тотальную РНК выделяли из клеток с использованием набора для очистки RNAeasy (Qiagen). кДНК синтезировали с использованием мастер-микса High Capacity RNA-to-cDNA от Applied Biosystems. Анализ qRT-PCR проводили с использованием реагентов TaqMan Universal PCR, смешанных с указанными кДНК и праймерами TaqMan, в системе ViiA7 Real-Time PCR (Applied Biosystems).Предварительно спроектированные праймеры Taqman были получены из прикладных биосистем, обнаружение следующих генов: FBP1 (ID: HS00983323_M1), HIF1A (ID: HS00153153_M1), PDK1 (ID: HS01561850_M1), LDHA (ID: HS00855332_G1) , GLUT1 ( SLC2A1 , ID: HS00892681_M1), VEGF ( VEGFA , ID: HS005_M1), 18S ( RN18S1 , HS03928985_G1).
Иммунопреципитация хроматина (ChIP) и анализ ChIP-reChIP
Анализы ChIP проводились с использованием набора Imgenex QuikChIP (Novus).Вкратце, три миллиона клеток RCC10, стабильно экспрессирующих V5-FBP1, сшивали 1% формальдегидом в течение 15 минут при 37 °C. Реакцию перекрестного связывания гасили глицином, и клетки лизировали в SDS-буфере, содержащем смесь ингибиторов протеазы. Клеточные лизаты обрабатывали ультразвуком для разделения хроматиновой ДНК на фрагменты размером 200–1000 пар оснований, а затем подвергали иммунопреципитации с использованием 5 мкг антител IgG (Cell Signaling Technology), V5 (Life Technologies) или полимеразы II (Imgenex). После промывки серией промывочных буферов с низким содержанием соли и высоким содержанием соли иммунопреципитированные фрагменты ДНК депоперечно сшивали при 65 °C в условиях с высоким содержанием соли, очищали с использованием набора для очистки QIAquick PCR (Qiagen), а затем анализировали с помощью qRT-PCR.Для анализов ChIP-reChIP первый ChIP выполняли с использованием антитела HIF1α (5 мкг, Novus) до этапов промывки. Иммунопреципитированные комплексы белок-ДНК инкубировали в элюирующем буфере ChIP-reChIP (10 мМ Трис, 1 мМ ЭДТА, 2% ДСН и 20 мМ ДТТ, рН 7,5) в течение 30 мин при 37 °С. После центрифугирования выделенный супернатант разбавляли не менее чем в 20 раз и подвергали второй ЧИП с использованием 5 мкг антител IgG или V5. В обоих анализах ChIP и ChIP-reChIP qRT-PCR выполняли с использованием следующих праймеров: PDK1 -HRE: 5′-CGCGTTTGGATTCCGTG-3′ и 5′-CCAGTTATAATCTGCCTTCCCTATTATC-3′; LDHA -HRE: 5′-TTGGAGGGCAGCACCTTACTTAGA-3′ и 5′-GCCTTAAGTGGAACAGCTATGCTGAC-3′; GLUT1 -HRE: 5′-CAAATGTGTGGATGTGAGTTGC-3′ и 5′-CCATCACGGTCCTTCTTCATG-3′; VEGF -HRE: 5′-CAGGAACAAGGGCCTCTGTCT-3′ и 5′-GCACTGTGGAGTCTGGCAAA-3′; RPL13A -nonHRE: 5′-GAGGCGAGGGTGATAGAG-3′ и 5′-ACACACAAGGGTCCAATTC-3′.
Репортер люциферазы и анализ трансактивации GAL4
0,5 мг репортерной конструкции HRE трансфицировали в клетки, выращенные в 6-луночных планшетах, вместе с 0,05 мг люциферазы Renilla и 1 мг индицируемой экспрессии или плазмиды кшРНК. Активность люциферазы измеряли через 48 часов с использованием набора для анализа Dual Luciferase (Promega). Активность репортерной люциферазы нормализовали по отношению к люциферазе Renilla, а затем масштабировали, чтобы установить контрольные сигналы вектора, равные 1. В анализе трансактивации GAL4 0.3 мг конструкции GBD-HIFα (с использованием вектора pBIND в качестве основы) трансфицировали в клетки 769-P (GBD-HIF1α) или A549 (GBD-HIF2α) вместе с 0,3 мг репортера UAS (восходящей последовательности активации)-люциферазы и 0,4 мг указанной экспрессии. плазмиды. Активность люциферазы измеряли, как описано выше. Эффективность трансфекции нормализовали по люциферазе Renilla, экспрессируемой из основы pBIND.
Субклеточное фракционирование
Цитозольное и ядерное фракционирование указанных клеток и тканей проводили с использованием реагентов для выделения ядер и цитоплазмы NE-PER (Pierce) в соответствии с протоколом производителя.
Анализ ферментативной активности
20 мкг лизатов клеток 293T, экспрессирующих вектор FBP1 или FBP1 G260R, добавляли к 100 мкл реакционного буфера, содержащего 50 мМ Трис, 10 мМ MgCl 2 и 100 мкМ D-фруктозы 1,6- бисфосфат тринатриевая соль. Реакцию начинали путем инкубации растворов при 37 °C в течение 15 минут и останавливали по протоколу депротеинизации с использованием набора для подготовки образцов для депротеинизации (BioVision). Каталитическую активность FBP1 количественно определяли по выходу фруктозо-6-фосфата (ферментативный продукт), измеренному с использованием набора для флуорометрического анализа фруктозо-6-фосфата PicoProb (BioVision).
Анализ потребления кислорода
Клетки RCC4 или RCC10, эктопически экспрессирующие вектор или FBP1, высевали в черный непрозрачный 96-луночный планшет с плотностью 70 000 клеток на лунку. После прикрепления культуральную среду в каждой лунке заменяли 150 мкл свежей насыщенной кислородом среды, дополненной 10 мкл раствора зонда MitoXpess (Cayman Chemical). В качестве отрицательного контроля использовали антимицин А. В каждую лунку наносили 100 мкл минерального масла, и весь планшет контролировали с помощью считывателя микропланшетов SpectraMax m2e (Molecular Devices) при 37 °C.Длины волн установлены на 380 нм для возбуждения и 650 нм для излучения.
Очистка белков
Конструкции, кодирующие указанные GST-меченые белки, трансформировали в химически компетентную BL21-DE3 E. coli (Life Technologies), выращенную в среде LB при качании при 37 °C до значения OD600, равного 0,8. Экспрессию белка индуцировали добавлением 1 мМ IPTG (Denville Scientific) к культурам LB, которые встряхивали при 37°C в течение еще 3 часов или при комнатной температуре в течение ночи. После центрифугирования собранные бактериальные осадки ресуспендировали в буфере STE (50 мМ Трис, 150 мМ NaCl и 1 мМ ЭДТА), дополненном коктейлем ингибиторов протеазы (Sigma).Белки, меченные GST, были дополнительно очищены с использованием набора для солюбилизации и ренатурации телец включения GST (Cell Biolabs) в соответствии с протоколом производителя.
GST pull-down assay
Анализ проводили с использованием GST Protein Interaction Pull-Down Kit (Pierce) в соответствии с протоколом производителя. Кратко, 100 мкг указанных белков-приманок, меченных GST, инкубировали с 50 мкл предварительно промытой глутатион-агарозы (Pierce) в 500 мкл вытягивающего буфера GST (20 мМ Трис, 150 мМ NaCl, 2 мМ MgCl 2 , 0.1% NP-40 и 20 мкг/мл BSA) в течение 1 часа при 4 °C. Между тем, 50 мкг указанных белков-«жертв» предварительно очищали с помощью 50 мкл предварительно промытой глутатион-агарозы в 500 мкл вытягивающего буфера GST в течение 1 часа при 4 °C. Предварительно очищенные растворы «добычи» соединяли с растворами «приманка-агароза» и перемешивали еще 2 часа при 4 °С. Агарозу, связанную с белком, промывали 3 раза и кипятили в буфере для образцов SDS. Супернатанты подвергали SDS-PAGE и вестерн-блоттингу.
Иммуногистохимия тканей и иммунофлуоресценция
Срезы нормальной ткани почки (из Cooperative Human Tissue Network) и предметные стекла с тканевым микрочипом (KD801, KD208, BC07114, BC07115, LV804, LV208 из US Biomax) депарафинизировали путем запекания предметных стекол при 60 °C в течение 30 мин.Предметные стекла регидратировали в серии растворов этанола, и активность эндогенной пероксидазы гасили 3% H 2 O 2 в дистиллированной воде в течение 30 мин. После трех промывок в ТТ-буфере (500 мМ NaCl, 10 мМ Trizma и 0,05% Tween-20) проводили выделение антигена путем кипячения предметных стекол в течение 20 минут в демаскирующем антигенном растворе на основе цитрата (Vector Labs). После охлаждения до комнатной температуры предметные стекла инкубировали с 4% нормальной козьей сывороткой в ТТ-буфере в течение 1 часа, а затем блокировали растворами авидина и биотина (набор для блокирования авидина/биотина, Vector Labs).Затем предметные стекла инкубировали с различными первичными антителами при 4°C в течение ночи. После трех промывок в ТТ-буфере на эти предметные стекла добавляли биотинилированные вторичные антитела на 1 час с последующей обработкой в течение 1 часа реагентами ABC (Vectatain ABC Kit, Vector Labs). После трех промывок ТТ слайды обрабатывали либо хромогеном (субстрат ImmPACT DAB, Vector Labs), либо растворами гематоксилина для иммуногистохимического окрашивания, либо тирамидом Alexa Fluor 488 (набор TSA № 22, Life Technologies) для иммунофлуоресцентных экспериментов.Наконец, предметные стекла были либо обезвожены и помещены в раствор Paramount (для иммуногистохимии), либо непосредственно помещены в реагенты против выцветания ProLong Gold (Life Technologies) с добавлением DAPI (для иммунофлуоресценции).
Исследования на животных
Эксперименты с ксенотрансплантатом опухоли были одобрены Комитетом по уходу и использованию животных Пенсильванского университета. Вкратце, пяти самкам голых мышей NIH-III (Charles River, 4–6 недель) вводили подкожно в оба бока пять миллионов клеток 786-O, стабильно экспрессирующих вектор или FBP1.Перед каждой инъекцией клетки ресуспендировали в 150 мкл DMEM, смешанного с равным объемом матригеля (BD Bioscience). После установления пальпируемых опухолей размер опухоли измеряли один раз в неделю с помощью цифрового штангенциркуля. Через четыре недели мышей умерщвляли путем ингаляции CO 2 , ксенотрансплантатные опухоли вырезали и взвешивали. Рандомизация и ослепление не применимы в этом эксперименте.
Субъекты-люди
Первичные образцы почек человека были получены из Кооперативной сети изучения тканей человека и обработаны в Пенсильванском университете с одобрения комитетов его институционального наблюдательного совета.Все ткани собираются с информированием и согласием донора. Сопутствующие процедуры были выполнены в соответствии с этическими и правовыми нормами.
Анализ выживаемости
Доступные данные о выживаемости пациентов были получены в рамках проекта TCGA ccRCC. Пациенты были ранжированы на основе их опухолевой экспрессии указанных генов, показанных в данных TCGA RNAseq. Верхняя половина ранжированных пациентов была определена как группа «высокие 50% экспрессии», а нижняя половина была определена как группа «нижние 50% экспрессии».Для этих двух групп пациентов были соответственно проведены анализы выживаемости Каплана-Мейера и тесты значимости логарифмического ранга.
Статистические данные
Все результаты, указанные в основных и расширенных данных, представлены как среднее значение ± стандартное отклонение. если не указано иное. Данные были представлены как биологические повторы, за исключением анализов qRT-PCR, где использовались технические повторы репрезентативного эксперимента. Данные за пределами двух стандартных отклонений от среднего значения рассматриваются как выбросы. Значения р рассчитывали на основе двусторонних непарных t-критериев Стьюдента.р<0,05 считали статистически значимым.
Политика конфиденциальности | Carolina Shores Vacation Rentals
Добро пожаловать на наш веб-сайт и благодарим за посещение. Ваша конфиденциальность важна для нас, и в этом обзоре подробно объясняется, как мы собираем, храним и передаем ваши данные на основе вашего взаимодействия с веб-сайтом. Обратите внимание, что эта политика конфиденциальности применяется исключительно к этому веб-сайту и информации, собранной на этом веб-сайте.
Сбор, использование и обмен информацией
Мы являемся исключительными владельцами информации, собранной на этом веб-сайте.У нас есть доступ только для сбора информации, которую вы добровольно предоставляете нам по электронной почте, при взаимодействии с контактной формой или при отправке, или через другие прямые контакты от вас. Мы не будем распространять эту информацию другим лицам.
Мы будем использовать вашу информацию, чтобы ответить на ваш запрос и указать причину обращения к нам. Мы не будем передавать вашу информацию какой-либо третьей стороне за пределами нашей организации, за исключением случаев, когда это необходимо для выполнения вашего запроса.
Если вы не откажетесь, мы можем связаться с вами в будущем, чтобы сообщить вам о рекламных предложениях, новых продуктах или услугах или изменениях в этой политике конфиденциальности.
Какую информацию мы собираем?
Мы собираем информацию от вас, когда вы вводите свою контактную информацию на нашем веб-сайте, включая заполнение контактной формы, запрос на бронирование или согласие на получение наших рекламных электронных писем.
Сюда входят: имя, фамилия, адрес электронной почты, почтовый адрес, номер телефона и/или другая информация.
Для чего используется эта информация?
Любая информация, которую мы получаем от вас, может быть использована одним из следующих способов:
Для персонализации вашего опыта
(ваша информация помогает нам лучше реагировать на ваши индивидуальные потребности)
Для улучшения обслуживания клиентов
(ваша информация помогает нам более эффективно реагировать на ваши проблемы)
Для обработки транзакций
Ваша информация, будь то общедоступная или частная, не будет продана, обменена, передана или передана какой-либо другой компании по какой-либо причине без вашего согласия, другое чем для явной цели обработки транзакции
Для отправки периодических электронных писем
Предоставленный вами адрес электронной почты может использоваться для отправки вам информации и обновлений, относящихся к вашему запросу, в дополнение к получению периодических рекламных предложений
Ваш доступ и управление Дополнительная информация
Вы можете отказаться от любых будущих контактов с нами в любое время.Вы можете сделать следующее в любое время, связавшись с нами по адресу электронной почты или номеру телефона, указанному на нашем веб-сайте.
Файлы cookie
Для дальнейшего улучшения вашего опыта на нашем веб-сайте мы используем файлы cookie для хранения ваших данных. Файл cookie — это часть данных, обрабатываемых нашим веб-сайтом и сохраняемых на вашем устройстве вашим веб-браузером во время вашего взаимодействия с нашим веб-сайтом. Сбор файлов cookie можно использовать для стратегических маркетинговых возможностей, как описано ниже.
Безопасность
Наш веб-сайт работает на уровне HTTPS, что означает, что любая посещаемая страница защищена.Независимо от того, заполняете ли вы контактную форму или отправляете запрос на бронирование, ваша информация полностью конфиденциальна и зашифрована.
Компьютеры/серверы, на которых мы храним личную информацию, хранятся в безопасной среде.
После запроса бронирования ваша личная информация не будет храниться на наших серверах.
Ссылки третьих лиц
Мы размещаем ссылки на другие стороны и услуги на нашем веб-сайте, и мы не несем ответственности за размещенное ими содержание.Эти веб-сайты имеют независимые политики конфиденциальности. Если вы считаете, что веб-сайт, на который мы ссылаемся, не заслуживает доверия, сообщите нам об этом, и мы проведем тщательное расследование.
Ваше согласие
Просматривая и взаимодействуя с нашим веб-сайтом, вы соглашаетесь с нашей политикой конфиденциальности.
Изменения в нашей политике конфиденциальности
Дополнительная информация или изменения в нашей политике конфиденциальности будут обновляться на этой странице.
Связаться с нами
По любым вопросам или опасениям по поводу этой политики конфиденциальности, пожалуйста, напишите или позвоните нам.
Ссылки для отказа
Мы используем ремаркетинг Google, Bing Ads, Facebook Ads и другие. Обратите внимание на следующую информацию о нашей рекламе:
- Сторонние поставщики, включая Google, Bing и Facebook, показывают нашу рекламу на веб-сайтах в Интернете.
- Сторонние поставщики, включая Google, Bing и Facebook, используют файлы cookie для показа рекламы на основе предыдущих посещений пользователем веб-сайта.
- Вы можете отказаться от ремаркетинга Google и использования ваших файлов cookie, посетив Настройки рекламы Google.
- Вы можете отказаться от персонализированной рекламы Bing, посетив Настройки рекламы Bing.
- Вы можете обновить настройки рекламы Facebook в настройках рекламы Facebook.
- Кроме того, у вас есть возможность отказаться от использования файлов cookie сторонними поставщиками, посетив Network Advertising Initiative (NAI) и Digital Advertising Allegiance (DAA).
- Google Analytics: мы используем Google Analytics, чтобы лучше понять, как посетители используют этот веб-сайт, и для рекламы на основе интересов, связанной с вашей учетной записью Google и другими используемыми вами устройствами.Информация, сгенерированная файлом cookie Google Analytics об использовании вами этого веб-сайта, передается и хранится в Google. Если вы не хотите, чтобы Google Analytics отслеживал ваши действия на этом веб-сайте, вы можете отказаться, перейдя по этой ссылке: http://tools.google.com/dlpage/gaoptout?hl=ru.
Если вы считаете, что мы не соблюдаем эту политику конфиденциальности, немедленно свяжитесь с нами по телефону 866-418-5263 или по электронной почте [email protected].
Металлургический дивизион История мировой войны
Номер: NP601 металлургический дивизион мировая война история Номер: NP602 Мэриленд, сентябрь 2002 г., низкий уровень воды Номер: NP603 казначейство архивов ussgl Номер: NP604 стеллваген банк Номер: NP605 Коллоквиум Солнечной системы в Пилчере Номер: NP606 автомобильная навигационная система и другие технологии Номер: NP607 каталог местных центров tasc Номер: NP608 грант штата Вашингтон Номер: NP609 Управление гражданских прав по политике в отношении сексуальных домогательств Номер: NP610 Соединенные Штаты против Дэвида Дж.kaiser стенограмма судебной апелляции Номер: NP611 Поправки к регламенту по туберкулезу крупного рогатого скота 2003 г. Номер: NP612 детский браслет с удостоверением личности напомнить об опасности удушья Номер: NP613 бюджетный комитет дома бюджет военного времени 2002 г. Номер: NP614 техподдержка ФМС Номер: NP615 Медицинская интенсивная программа ухода за инвалидами Номер: NP616 Шумер пресс-релиз кредитный спор о выбросах Номер: NP617 экономия с солнечным видео Femp 1999 Номер: NP618 штатное расписание центра тепловых систем зданий 2005 г. Номер: NP619 мы возражаем против заявлений аннана ирака Номер: NP620 страницы правительственного учреждения сша Номер: NP621 USGC Lands к 30-летию Номер: NP624 как добраться до музея альбукерке Номер: NP625 закон о труде на железнодорожном транспорте чрезвычайный совет по усмотрению президента Номер: NP626 Инициативы CDC и HRSA по профилактике ВИЧ в исправительных учреждениях Номер: NP627 Церемония перерезания ленточки Центр технических записей НАСА Номер: NP628 предварительные шаги запроса foia Номер: NP629 новости сома Номер: NP630 пищевой кодекс FDA 1997 г. Номер: NP631 должностная инструкция геодезиста Номер: NP632 правила для нас федеральное правительство жилье и налоговые льготы Номер: NP633 Годовой отчет городского транспорта за 1998 г. Номер: NP634 деятельность по реинтродукции хорьков Номер: NP635 конкурс стихов о рассеянном склерозе мать Номер: NP636 виза в сша москва Номер: NP637 как подать заявку на лицензию НАСА технологии Номер: NP638 Свидетельские показания Донны Тануэ Номер: NP640 Содержание статистического отчета о фертильности США Номер: NP641 информационный бюллетень Номер: NP642 Трибьют начальной школы Глисон-Лейк 11 сентября Номер: NP644 генетическое расстройство дефицита гемоглобина Номер: NP645 Краткий ответ США против Белл и Вестерн Электрик Номер: NP646 эффективность и экономическая эффективность вакцин в аквакультуре (25-й ежегодный семинар по здоровью рыб) Номер: NP647 программа конкурсных грантов Номер: NP648 национальная токсикологическая программа tr-403 резорцин Номер: NP649 Билл Белл и его программа 1996 года в Белом доме о Пасхе Номер: NP651 агентство по охране окружающей среды исследовательский центр Номер: NP652 Брифинг Мюллера, ФБР, Филиппины, терроризм, милфа Джи Номер: NP653 справочная страница ipcs Номер: NP654 изображение кофе НАСА Номер: NP655 план управления морским парком Сан-Франциско Номер: NP656 Джонсон контролирует биографию президента Номер: NP657 отчет о смертности среди диких животных за первый квартал 1995 г. в середине континента Номер: NP658 справочный глоссарий по генетике Номер: NP659 статистика веб-сервера национального центра данных космической науки, 10-16 февраля 1997 года Номер: NP660 Письмо назначающего органа cs-6884 от 31 июля 1998 г. от Джона Ф.лопес Номер: NP661 Меган Тиррелл, Нью-Гемпшир, реферат исследования 1998 2001 Номер: NP662 Документация по объекту распределителя С++ Номер: NP663 Ненаблюдаемость подходов для ресурсов и услуг Номер: NP664 Грег Киммел в emsl Номер: NP665 должностная инструкция страховщика промышленного страхования в Вашингтоне Номер: NP666 Законопроект о чрезвычайном положении в Ираке и Афганистане в Кантуэлле Номер: NP667 борьба с вредителями в перспективе Номер: NP668 Карибская экологическая программа Номер: NP669 награда за выдающиеся достижения в области образования для Летисии Меркадо Номер: NP670 Зеллик, речь, реформа вто, гана, корея Номер: NP671 органическая добавка для улучшения почвы Номер: NP672 часто задаваемые вопросы о кружащейся болезни Номер: NP673 Отчет о ценах на виноград в Калифорнии за 2002 г. Номер: NP674 домашняя страница вольфганга вагнера Номер: NP675 планы клиентов менеджера по выставлению счетов Номер: NP676 криоконсервация дескрипторов цитрусовых и зародышевой плазмы фиников Номер: NP677 кентукки kydjj 2003 пресс-релизы Номер: NP678 аукцион федеральная земля округ дуглас Номер: NP679 стенограмма Буш Путин Договор о сокращении ядерных боезарядов 2002 г. Номер: NP681 охота за мусорщиком Номер: NP682 время сбора урожая национальный парк Маунт-Рейнир Номер: NP683 исследовательский профиль Раджана Гупты Номер: NP684 ресурсная комната для ваучерной программы wtw Номер: NP685 Национальный парк Бунакен Номер: NP686 программа землепользования Номер: NP687 проекты, финансируемые сосудистыми растениями Номер: NP688 Обязанности регулятора энергетики Калифорнии Номер: NP689 mem_fun1_ref_t ссылка на stl Номер: NP690 браун сетцер аннотация землепользование бразилия Номер: NP691 Грузия критерии для получения Medicaid Номер: NP692 пособие по смерти на работе Номер: NP693 Статистика населения округа Кэрролл Номер: NP694 ответ сената на ошибку 2000 года Номер: NP695 плюшевый мишка леди Номер: NP696 бостон кофе день матери Номер: NP697 опрос льгот по медицинскому обслуживанию Номер: NP698 Документ о работе совета по условно-досрочному освобождению штата Невада Номер: NP699 Еженедельный архив Рудольфа о роли Бернарда Керика в снижении преступности в Нью-Йорке Номер: NP701 контактные данные проекта Richard E.Национальный центр экологических исследований Стивенсона США эпа Номер: NP702 программа работы с молодежью темпе Номер: NP703 лаборатория реактивного движения Номер: NP704 Письмо Маккарти JSE Securities Южная Африка 2002 г. Номер: NP705 Биография Уинфилда Хэнкока Номер: NP706 cbar класса java Номер: NP708 hctc часто задаваемые вопросы Номер: NP709 содержание сайта по сохранению природных ресурсов Номер: NP710 кобайси теллез даян герцог 2001 callicarpa japonica Номер: NP711 cdc it помощь описание Номер: NP712 Политика Белого дома Клинтона Гора поднять отчет об успеваемости учащихся Номер: NP713 Преимущества посещения кадровой службы для бывших военнослужащих Номер: NP715 симулятор строительства зеленого жилья Номер: NP716 Кэти Спаркс, апелляция, январь 2000 г. Номер: NP718 стенограмма интервью armitage nbc крайний срок инспекции оружия Номер: NP719 имя генерального директора BusinessLinc Номер: NP720 Доклад 1999 г., Румыния, права человека Номер: NP721 уведомление об отзыве тренажера для кроватки Номер: NP722 Микробиологическая монография Ассоциации мыльных детергентов Номер: NP723 программа усыновления темпе дерева Номер: NP724 dusd информационные системы дома в арлингтоне Номер: NP725 Политические подразделения штата Миссури статуты 2003 г. Номер: NP726 11 сентября объявлено чрезвычайное положение Номер: NP729 описание опроса о заработной плате fltcip 2002 г. Номер: NP730 политика исключения претензий dofa Номер: NP731 факты и правила морфологии lvg Номер: NP732 р-4 р-5 зонирование Номер: NP733 радио адрес лаура буш афганистан белый дом Номер: NP734 война мир толстой отрывок Номер: NP735 патент на поворотный зажимной блок Номер: NP736 описание разрыва бронха из медицинской энциклопедии Номер: NP737 проект пуаиохи демография населения охрана дикой природы Номер: NP738 Статистика округа Митчелл для лиц, не являющихся работодателями (бюро переписи населения США) Номер: NP739 малый бизнес помощь правительства Индианы Номер: NP740 кодекс поведения в швейной промышленности Номер: NP741 Заявление Уайдена Маккейна Лихи о налоговом законодательстве в отношении электронной коммерции Номер: NP742 речь байдена саудовская ближний восток мир Номер: NP743 заявление сенатора веллстоуна о смерти аберкромби Номер: NP744 абстрактные функции Dias Prata Widner Номер: NP745 Скачать форму государственной службы штата Мичиган Номер: NP746 Должностная инструкция инспектора по сбору взысканий Майами Дейд Каунти Номер: NP747 тигровые жуки округ боллинджер Номер: NP748 серия отчетов по статистике здоровья матери и ребенка, российская федерация, сша отдельные годы 1985-95 гг. Номер: NP749 Роланд Знаменитый Викторианский По Письма 4543 Альянс Атлантида Номер: NP750 Конгрессмен Хергер открытое письмо Лауден компенсация пожара Номер: NP751 предвыборная рассылка ограничения массовая рассылка Номер: NP752 заявление форель неограниченное западное водосбережение тедди рузвельт Номер: NP753 отчет о государственных расходах средней страны Номер: NP754 Понятовский лишение лицензии стоматолога Номер: NP755 история драгунских казарм Номер: NP756 Финансовый отчет «Зеленый город Миссури» за 1998–2003 гг. Номер: NP757 сообщает m’cleland 1825 kd208 1825.м2 Номер: NP758 атмосферные ядерные осадки Национальный институт рака оценил облучение Номер: NP759 Среднее количество осадков в Форт-Смит в апреле 2003 г. Номер: NP760 факты о кондиционере автомобиля Номер: NP761 предполагаемое воздействие атмосферных осадков ядерной бомбы на американцев Номер: NP762 eeoicpa рассмотрение иска о возмещении ущерба заводу в скалистых квартирах Номер: NP763 Закон об отмене налога на смерть 2001 года Джим Нассл Номер: NP764 страница поиска межведомственных контрактов Номер: NP765 ссылка на команду lsrpnode Номер: NP766 домашняя страница администрации социального обеспечения Номер: NP767 Эдгар закрытый новогодний праздник Номер: NP768 Кентукки Motorcoach программа пресс-релиз Номер: NP769 Иск Министерства труда США против попечителей пенсионного фонда «Аврора» 22 января 2001 г. Номер: NP770 ежемесячная передача школьных финансов Номер: NP771 Отчет о посещаемости учебного года долины Патнэм для иногородних за 1998-1999 гг. Номер: NP772 инфраструктура национальный институт рака центр биоинформатики Номер: NP773 Статистика лиц, не являющихся работодателями, в мегаполисах Вирджинии в 1999 г. Номер: NP774 табак беременность аризона cdc Номер: NP775 обзор национальной инфраструктуры пространственных данных Номер: NP776 Домашняя страница обслуживания Casil штата Калифорния Номер: NP777 посещаемость в Монетт Хай Миссури с 1999 по 2003 год Номер: NP778 стандарты данных о раке cadsr ncicb Номер: NP779 надзор федеральных ведомств за растратой государственных средств Номер: NP780 стандарты данных рака oracle java Номер: NP781 Дизайн и методология обследования демографической переписи Номер: NP782 Национальная служба погоды для Мельбурна Номер: NP783 общедоступная инфраструктура пространственных данных Номер: NP785 эпоха Номер: NP786 Льюис и Кларк рабочий лист Номер: NP787 Окружной суд Висконсина: часто задаваемые вопросы Номер: NP788 приливный глоссарий океан Номер: NP789 бюро переписи металлический бак бизнес занятость Номер: NP790 orbz проверяет эффективность перехвата спама Номер: NP791 марс следопыт имп изображение аэрозоль непрозрачность солнце Номер: NP792 руководство по установке mips debian Номер: NP793 документ управления водоразделом агентства по охране окружающей среды Номер: NP794 научные обзоры индекса агентства по охране окружающей среды Номер: NP795 глобальный геодезист Марса Номер: NP796 физиологическая эффективность экстракта растения индиго polygonum tinctorium Номер: NP798 стратегия нсди Номер: NP799 специалист по закупкам жилья и градостроительства Номер: NP800 передовые методы совместной работы для научного моделирования и управления Номер: NP801 офис по исследованию СПИДа Номер: NP802 последовательный обмен географическими данными с меньшим дублированием и затратами Номер: NP803 us-31 resurfacing берриен Номер: NP804 национальная медицинская библиотека гепатоэритропоэтическая порфирия Номер: NP805 обработка свалки дисковых труб Номер: NP806 национальная медицинская библиотека поиск Номер: NP809 пищевая цепь кратерного озера Номер: NP810 кратерное озеро национальный парк растения живая природа Номер: NP811 национальная инфраструктура пространственных данных nsdi Номер: NP813 Программа управления твердыми отходами Департамента природных ресурсов штата Миссури Номер: NP814 заявление о конфиденциальности для грузии.домен gov Номер: NP815 cdc it помощь описание Номер: NP816 ресурсы разработчика lims Номер: NP817 Страница вакансий штата Орегон Номер: NP818 Совет акушерок Теннесси Номер: NP819 Джордж Бохарски против компании Aetna, занимающейся несчастными случаями и порукой, установление фактов, выводы закона и судебного решения Номер: NP820 новости губернатора штата иллинойс Номер: NP821 Часто задаваемые вопросы об агентстве по контролю за загрязнением окружающей среды Миннесоты Номер: NP822 Делавэр Номер: NP823 часто задаваемые вопросы о компенсации работникам Номер: NP824 члены объединенного судебного комитета Номер: NP825 Парк штата Пенсильвания, бассейн, доступ для инвалидов Номер: NP826 Дорожный патруль Флориды округ Гейнсвилл Номер: NP827 критерии оценки учебной программы в Нью-Гемпшире Номер: NP828 Годовой отчет Орегонского центра статистики здравоохранения за 1998 г. Номер: NP829 озеро монтана чем заняться сток Номер: NP830 Контактные данные Канзасского отдела водных ресурсов Номер: NP831 брюшной тиф птицы Номер: NP832 pmp поселение износа и разрыва мэриленд Номер: NP833 протокол тестирования стека Номер: NP834 Условия рыбалки в Уолнат-Крик Номер: NP835 советы по безопасному вождению в офисе Номер: NP836 течение реки Бойсе Номер: NP837 Награды дорожного патруля Флориды Номер: NP838 показания Дениз Кроуфорд Номер: NP839 Служба социального обеспечения Техаса, февраль 2003 г., протоколы Номер: NP840 Руководство для практикующих врачей по программе скрининга новорожденных в штате Небраска, второе издание, 1998 г. Номер: NP841 Законодатели штата Миннесота с двумя офисами Номер: NP842 Стивен Манхарт экзамен ptoe Миннесота Номер: NP843 ключевой сотрудник племени рекомендация соискателя казино Номер: NP844 правила для нас федеральное правительство жилье и налоговые льготы Номер: NP845 сша заявляет последние законодательные инициативы Номер: NP846 кораблекрушение Чарльза Хейта Номер: NP847 Иллинойс, отчеты об образовании для карьеры, 1997 г. Номер: NP848 телефон джеймса сауэрса Номер: NP849 История пенсионной системы Аризоны Номер: NP850 dhmh пройти тест 2000 года Номер: NP852 Статистические данные о занятости и заработной плате в Нью-Мексико в 2002 г., исследование OES Номер: NP853 официальные результаты выборов в грузии 5 ноября 1996 г. Номер: NP854 программы по производству одежды Номер: NP855 Рекомендации губернатора Миннесоты по капитальному бюджету на 2004 г. Номер: NP856 Джим Хилл Республиканская предвыборная кампания Номер: NP859 парк штата маккарти-бич история живая природа Номер: NP860 обзор школьного округа Киллин управление объектами аутсорсинга Номер: NP861 страницы правительственного учреждения сша Номер: NP862 список артистов программы резидентуры Номер: NP863 уровень безработицы в грузии саванна 2000 Номер: NP864 фонды городского озеленения патаки Номер: NP865 Закон о банках и трастовых компаниях штата Миссури Номер: NP866 мемориал палмера студенческая жизнь Номер: NP867 Годовой отчет DHS за 2000 год, штат Орегон Номер: NP868 Департамент труда Коннектикута, высокие ставки заработной платы, Нью-Хартфорд Номер: NP869 индекс сайта lepc Номер: NP870 заполнение формы 6251 альтернативный минимальный налог Номер: NP871 Закон штата Колорадо о безопасности детей-пассажиров Номер: NP872 меморандум медицинского консультативного комитета правило об исключении идетKO88 — Внутренний заказ (полная структура распределения A1)
Привет гуру,
Я искал решение своей проблемы, но ничего из этого не могло мне помочь.
Когда я запускаю KO88 для полного расчета, я получаю следующую ошибку:
================================================ ================================================== ========
Полная структура распределения A1
Номер сообщения. КД503
Диагностика
Во время расчета система распределяет дебеты отправителя по группам (присвоениям), которые рассчитываются с использованием одного и того же вида затрат для расчета.Назначение расчета выполняется в структуре распределения, которая хранится в правиле расчета для отправителя (в параметрах расчета).
Вид затрат 981550 не может быть присвоен присвоению расчетных затрат в структуре распределения A1 и, следовательно, не может быть присвоен расчетному виду затрат.
Процедура
Возможные решения:
- Если проводка по виду затрат 981550 была выполнена отправителю из-за неправильного ввода, можно отменить проводку и не нужно обновлять структуру перерасчета.Вам нужно только отменить неправильно назначенную проводку, прежде чем повторять расчет. Исключение: для инвестиционного показателя с расчетом по отдельным позициям необходимо расширить структуру распределения даже после сторнирования.
- Если проводка была сделана правильно, то можно сделать следующее:
-
a) Отправителю можно присвоить другую структуру перерасчета: в ведении основных данных для отправителя выберите «Правило расчета -> Перейти к -> Параметры расчета», а затем введите другую структуру перерасчета.
- При необходимости также проверьте, правильно ли введена структура распределения в профиле расчета. Структура перерасчета устанавливается по умолчанию в профиле расчета при создании основных данных отправителя или при ведении правила расчета.
- Вы можете обновить структуру распределения A1
============================================== ================================================== =========
Что я сделал до сих пор:
1.Создать структуру распределения
2. Группа видов затрат (код T: KAh2) следующим образом
Благодарю за помощь и заранее большое спасибо
С уважением,
Кензо
US Biomax — Избранные ссылки
ССЫЛКИ
Тканевые микроматрицы и продукты US Biomax упоминались в более чем 1200 научных статьях с 2006 г., из них 600 с 2013 г. .Наши продукты были использованы в нескольких статьях ведущих журналов, таких как Nature и Cell.
Чтобы просмотреть последние ссылки, проверьте наши списки в Google Scholar или просмотрите полный список.
Избранные ссылки
Топ-200 по импакт-фактору:
Фруктозо-1,6-бисфосфатаза препятствует прогрессированию рака почкиB Li, B Qiu, DS Lee, ZE Walton, … — Nature, 2014
… предметные стекла с микрочипами (KD801, KD208, BC07114, BC07115, LV804 и LV208 от US Biomax) депарафинизировали путем запекания предметных стекол при 60 °C в течение 30 минут.Слайды… Потеря передачи сигналов TGFbeta дестабилизирует гомеостаз и способствует плоскоклеточному раку в многослойном эпителии
G Guasch, M Schober, HA Pasolli, EB Conn, … — Cancer Cell, 2007
… и 41 женский генитальный SQCC (UV241, UV801, PE241, PE801) были получены от US Biomax, Rockeville, MD. Парафиновые препараты депарафинизировали и окрашивали… Одиночные соматические удары инактивируют сцепленный с Х-хромосомой супрессор опухоли FOXP3 в простате
L Wang, R Liu, W Li, C Chen, … — Cancer Cell, 2009
… Информация.Образцы микрочипов тканей поступили из Мичиганского университета, компании Biomax (US Biomax, Inc., Роквилл, Мэриленд) и Target Biotech (Target Biotech, … Раскрытие роли KIAA1199, нового белка эндоплазматического ретикулума, в миграции раковых клеток
NA Evensen, C Kuscu, HL Nguyen, K Zarrabi, … — J. Natl. Институт рака, 2013
… образцы микрочипов (BR804, BC08013a и BR10010a) были приобретены у US Biomax Inc (Rockville, MD). Анализ SNAP Pull-Down и протеомный анализ. … Рецептор активированной С-киназы 1 способствует росту гепатоцеллюлярной карциномы за счет усиления активности митоген-активируемой протеинкиназы 7
Y Guo, W Wang, J Wang, J Feng, … — Hepatology, 2013
… Окрашивание RACK1 (BD Biosciences, Сан-Хосе, Калифорния) на предметных стеклах тканевых микрочипов (US Biomax, Inc., Роквилл, Мэриленд; видеть … Болезнеспецифическое репозиционирование генов при раке молочной железы
К. Дж. Миберн, П. Р. Гудла, С. Хан, С. Дж. Локетт, … — J. Cell Biol., 2009
… были получены из Ресурса образцов СПИДа и рака или приобретены у US Biomax, Inc., Imgenex Деконденсация хроматина и ядерная реорганизация … CXCR7 (RDC1) способствует росту опухолей молочной железы и легких in vivo и экспрессируется в ассоциированной с опухолью сосудистой сети
Z Miao, KE Luker, BC Summers, R Berahovich, … — Proc.Натл. акад. науч. США, 2007 г.
… Микрочипы опухолей были приобретены у Imgenex, Zymed/Invitrogen, Cybrdi, US Biomax, Biochain и … NCYM, цис-антисмысловой ген MYCN, кодирует возникший de novo белок, который ингибирует GSK3β, что приводит к стабилизации MYCN в нейробластомах человека
Y Suenaga, SM Islamic, J Alagu, Y Kaneko, … — PLoS Genet., 2014
… панель) при истощении MYCN в массивах тканей (FDA808a-1 и FDA808a-2, US Biomax, Rockville, MD, USA) использовались для анализа NCYM или MYCN… Сверхэкспрессия ANCCA/ATAD2 идентифицирует пациентов с раком молочной железы с плохим прогнозом, действуя, чтобы стимулировать пролиферацию и выживание тройных негативных клеток посредством контроля B-Myb и EZh3
Е.В. Калашникова, А.С. Ревенко, А.Т. Гемо, Н.П. Эндрюс, … — Cancer Res., 2010
… микрочипы опухолевой ткани (ТМА; BR1002, BR961 и BR208) были получены от US BioMax, Inc., выдержаны при 60°C в течение 2 часов. Клетки показали, что большинство этих генов…
Технология тканевых микрочипов:
- Хольгер Мок, Юха Кононен, Олли-П. Каллиониеми и Гвидо Заутер, Тканевые микрочипы: что они привнесут в молекулярную и анатомическую патологию. Достижения в области анатомической патологии, том 8, № 1, 14–20, (2001 г.).
- Рональд Саймон, Ян Рихтер, Урс Вагнер и др., Анализ количества копий 3p25 (RAF1) и 8p12 (FGFR1) на тканевых микрочипах изменения при раке мочевого пузыря. Рак рез. 61:4514-4519, (2001).
- Марк А. Рубин. Использование микродиссекции с лазерным захватом, микрочипов кДНК, и тканевые микроматрицы в продвижении нашего понимания предстательной железы Рак. Дж. Патол. 195:80-86, (2001).
- Олли-П. Каллиониеми, Урс Вагнер, Юха Кононен и Гвидо Заутер, Технология тканевых микрочипов для высокопроизводительного молекулярного профилирования рака.Молекулярная генетика человека, том 10, № 7, 657–662, (2001).
- Дэвид Л. Римм, Роберт Л. Кэмп, Лори А. Шаретт и др., Тканевой микрочип: новая технология увеличения тканевых ресурсов. Раковый журнал, том 7, № 1 (2001 г.).
- Лукас Бубендорф, Антонио Ноцито, Хольгер Мок и Гвидо Заутер, Технология микрочипов тканей (ТМА): миниатюрные архивы патологий для высокопроизводительных исследований in situ. Дж. Патол. 195:72-79, (2001).
- Чад Р. Энглерт, Галина В. Байбакова и Майкл Р.Эммерт-Бак, Cancer Res. 60:1526-1530, (2000).
- Ф. фон Эггелинг, Х. Дэвис, Л. Ломас, В. Фидлер, К. Юнкер, У. Клауссен и Г. Эрнст, Тканеспецифическая микродиссекция в сочетании с технологиями массивов белковых чипов: применение в исследованиях рака. Биотехнологии Том 29, № 5, 1066-1070, (2000).
- Olivier Braissant и Walter Wahli, Упрощенный протокол гибридизации in situ с использованием нерадиоактивно меченных зондов для обнаружения обильных и редких мРНК на срезах тканей. Биохимия №1, 10-16, (1998).
Тестирование — Все производители — eTesters.com
Показаны последние результаты 1 — 15 из 7974 найденных продуктов.
-
Тестирование
Underwriters Laboratories Inc.
Наш независимый и строгий процесс тестирования помогает убедиться, что ваша продукция соответствует применимым требованиям и ожиданиям. Мы предлагаем инновационные индивидуальные решения, направленные на оптимизацию процессов тестирования, снижение затрат и ускорение выхода продукции на рынок по всему миру.А знания нашей глобальной команды помогут вам оставаться на шаг впереди меняющихся требований, материалов и технологий.
-
Тестирование
ТЮФ ЗЮД АГ
TÜV SÜD — мировой лидер в области тестирования и сертификации продукции. Поэтому, когда вы сотрудничаете с нами, ваш бизнес в надежных руках.
-
Тестирование
Лаборатории Апогей
Apogee Labs предлагает широкий выбор типов модульных шасси, используемых для тестирования приложений с различными модулями тестирования канала передачи данных.
-
Испытание на летальность, Испытание на арене, Испытание взрывом
Национальные технические системы
Чтобы оценить поражающую способность усовершенствованных боеголовок, необходимо точно охарактеризовать распределение массы и скорости осколка боеголовки, а также пространственное распределение избыточного давления взрыва. Оценка поражающей способности усовершенствованных боеголовок требует точной характеристики распределения массы и скорости осколка боеголовки. , а также пространственное распределение избыточного давления взрыва.
-
Тестирование SOA, облачное тестирование, тестирование веб-приложений
SOAtest — Parasoft Corp.
Автоматизируйте сквозное тестирование критически важных для бизнеса и безопасности транзакций. Parasoft SOAtest широко известен как ведущее решение корпоративного уровня для тестирования API и проверки целостности API. Тщательно тестируйте составные приложения с надежной поддержкой REST и веб-сервисов, а также 120+ лучших в отрасли протоколов/типов сообщений.
-
Испытание на молниезащиту и испытание на устойчивость к импульсным перенапряжениям
Соответствие трапецеидальным искажениям
Keystone Compliance предлагает инженерные услуги для выполнения стандартных и пользовательских требований к испытаниям на устойчивость к грозовым импульсам и помехоустойчивости. Молниеносные испытания могут быть выполнены в виде одиночных, многократных и многократных испытаний. Прямые молниезащитные испытания обычно требуются для внешних систем самолета.Косвенное тестирование молниезащиты требуется для большей части бортовой электроники, как внутренней, так и внешней. Непрямая молния имитирует вторичные токи и напряжения, которые проходят через схемы и кабели. Испытание также называют испытанием на устойчивость к импульсным перенапряжениям.
-
Услуги по тестированию производительности и нагрузочному тестированию
Enhops Solutions Pvt Ltd.
Мы в Enhops предлагаем комплексные услуги по тестированию производительности, которые гарантируют доступность, надежность, масштабируемость и скорость систем и приложений наших клиентов.Наш Центр передового опыта по тестированию производительности анализирует требования наших клиентов к производительности, определяет стратегии тестирования производительности, дорожную карту и показатели, оценивает отчеты и предоставляет рекомендации. Наша команда по тестированию обладает глубоким опытом работы с различными открытыми и коммерческими инструментами для тестирования производительности.
-
Услуги по тестированию на электростатический разряд и тестированию с защелкой
Аналитическая группа Эванс®
EAG является лидером в области испытаний на электростатический разряд (электростатический разряд) и испытаний на фиксацию.Наша высококвалифицированная команда инженеров использует свои ведущие в отрасли знания и многолетний или реальный опыт работы с новейшими полупроводниковыми технологиями, проектированием схем и физикой устройств для оптимизации результатов наших клиентов в отношении электростатического разряда и защелкивания. Модель человеческого тела (HBM) и модель машины (MM), модель заряженного устройства (CDM), блокировка, импульс линии передачи (TLP).
-
Испытание на электростатический разряд / испытание на электростатический разряд
Соответствие трапецеидальным искажениям
Испытание на электростатический разряд, также известное как испытание на электростатический разряд, является важным испытанием для многих продуктов.Электростатический разряд может вызвать ряд проблем, включая, помимо прочего, механические неисправности, взрывы угольной пыли, взрывы паров топлива и телесные повреждения. Электростатический разряд — это внезапная передача электричества между электрически заряженными объектами. Этот электростатический перенос может быть вызван контактом, пробоем диэлектрика или коротким замыканием. Испытания на электростатический разряд имитируют различные электростатические воздействия, которые оборудование может испытать во время транспортировки или эксплуатации. Воздействие электростатического разряда может быть значительным, независимо от того, является ли электростатический разряд настолько незначительным, что его невозможно обнаружить на слух или зрение, или столь же впечатляющим, как электрические искры или молния.Испытание на электростатический разряд определяет, соответствует ли продукт требованиям зоны защиты от электростатического разряда и процедурам. Производители электроники должны определить меры по снижению электростатического разряда, если их продукция либо подвержена электростатическому разряду, либо может создавать электростатический разряд. Эти меры могут включать создание электростатических защитных зон, свободных от статического электричества, контроль влажности, использование мер по устранению заряда, таких как отказ от сильно заряжающихся материалов и принятие мер по удалению статического электричества, таких как заземление рабочих или требование антистатических устройств.Испытание на электростатический разряд подтверждает, что оборудование работает правильно в соответствии с требованиями испытаний и рабочими процедурами производителя.
-
Наборы для тестирования
Наборы для тестирования — Детектортестеры
ТестерыSolo доступны либо в виде отдельных компонентов, либо в виде полных комплектов, сконфигурированных как по высоте, так и по области применения. Цена полных комплектов обычно меньше суммы составных частей.
-
Нагрузочное тестирование и стресс-тестирование
Дотком-Монитор
Нагрузочный тест — это запланированный тест для выполнения определенного количества запросов к системе с целью проверки функциональности системы при определенных уровнях одновременных запросов. Нагрузочный тест гарантирует, что веб-система способна обрабатывать ожидаемый объем трафика, поэтому его иногда называют объемным тестированием. Цель нагрузочного теста — доказать, что система может обрабатывать ожидаемый объем с минимальным или приемлемым снижением производительности.Порог приемлемого снижения производительности должен быть определен тестировщиками как некоторое значение, которое считается приемлемым для конечного пользователя, чтобы пользователи не уходили с сайта.
-
Юзабилити-тестирование | Проверка внешнего вида и ощущений
Национальные технические системы
NTS может помочь вам определить удобство использования вашего продукта, предоставляя анализ конечного пользователя с различным пользовательским опытом (например,г., новичок, средний уровень, эксперт и т. д.). NTS придумала фразу «Взгляд и ощущение» из термина «Дизайн, ориентированный на пользователя» или (UCD) Уэсли Э. Вудсона. Вудсон определил UCD как «практику проектирования продуктов таким образом, чтобы пользователь мог выполнять необходимые задачи по использованию, эксплуатации, обслуживанию и поддержке с минимальным стрессом и максимальной эффективностью». UCD — это углубленный процесс анализа юзабилити как в качественном, так и в количественном отношении, который требует большого количества времени и денег. А на сегодняшнем конкурентном рынке большинство разработчиков не могут себе позволить такой подход.
-
Испытание материалов
Gaynes Labs, Inc.
*Испытание на истирание*Испытание на адгезию*Темп. Испытание на твердость*Теплопрогиб*Гидростатическое испытание*Испытание на удар*Испытание на жизненный цикл*Испытание на индекс плавления*Испытание на пластичность*Разделение слоев*Содержание смолы*Испытание на резину*Испытание на прочность на сдвиг*Испытание на трещины под напряжением*Испытание на разрыв*Температурное испытание*Растяжение/удлинение* Воздействие УФ-излучения*Влагопоглощение*Испытание древесины
(PDF) Фруктоза-1,6-бисфосфатаза противостоит прогрессированию рака почки
Последовательности HIFα, лишенные ДНК-связывающего мотива bHLH, были клонированы в вектор pBIND
(Promega), который находится в рамке считывания с ДНК-связывающим доменом GAL4 (GBD) на N-конце.
Серия усечений GBD-HIF1α была создана путем удаления указанных мотивов HIF1α.
Аналогичным образом, укорочения GBD-HIF2α были получены путем удаления по отдельности следующих
мотивов из последовательностей HIF2α (1–870): PASA (80–236), PASB (237–331), LINK (332–
395), NODD (396–495), NTAD (496–581), ID (582–830), CTAD (831–870). Конструкции GFP-FIh2
и GFP-PHD2 были получены от доктора Эрика Метцена через Addgene.
Плазмидная трансфекция и вирусная инфекция
Экспрессионные конструкции трансфицировали в линии раковых клеток с использованием реагента Lipofectamine LTX
(Life Technologies) в соответствии с протоколом производителя.Лентивирус
был получен путем совместной трансфекции клеток 293T векторами экспрессии pRSV-Rev, pMD2.G, pMDLg/pRRE и
pLKO.1-puro shRNA или pCDH-puro. Вирус собирали через 48 часов
путем фильтрации вируссодержащей среды через 0,45 мкМ фильтр Steriflip (Millipore). Заражение вирусом
проводили путем инкубации клеток со средой, содержащей указанный вирус и полибрен 8
мкг/мл (Sigma) в течение 24 часов. Клетки восстанавливали в полной среде
в течение 24 часов, а затем отбирали пуромицином в течение 24 часов (анализ мРНК) или 48 часов
(анализ белков и фенотип).Выжившие пулы подвергали указанным экспериментам.
Обратите внимание, что клеткам HK-2, подвергшимся заражению вирусом и отбору пуромицином, обычно требуется 10–
14 дней для восстановления, прежде чем их можно будет подвергнуть экспериментам.
Анализ TCGA RNAseq
Исходные данные RNAseq для 480 опухолей ccRCC и 69 нормальных тканей почек были загружены
из проекта TCGA ccRCC (http://cancergenome.nih.gov) 2 апреля 2013 г. Данные были проанализированы
для дифференциальной экспрессии генов с помощью DeSeq (Bioconductor, версия 2.12).
Экспортированы нормализованные подсчеты, значения p, значения p с поправкой на частоту ложных открытий (p-adj) и
изменения экспрессии для каждого гена. Для проведения анализа наборов генов полный список
из 2752 генов, кодирующих все известные метаболические ферменты человека и
переносчиков, был сгруппирован в функциональные наборы в соответствии с метаболическими путями
, аннотированным KEGG (http://www.genome.jp). /кегг/). Пороговое значение p-adj было установлено равным 0,1
, чтобы исключить гены, не обнаруживаемые последовательно с помощью RNAseq.Для изучения активности HIF
, ассоциированной с опухолью, необработанные данные каждого секвенированного гена были перемасштабированы, чтобы установить медиану равной 1, и активность
HIF была количественно определена путем усреднения нормализованной экспрессии 44 генов-мишеней HIF
, включая те, которые кодируют IGFBP3, EDN2, PFKFB4, FLT1, TFR2, BNIP3L, TGFA,
BNIP3, PGK1, EGLN1, LDHA, EGLN3, CP, TGFB3, PFKFB3, HK1, TFRC, EDN1,
CDKN1A, CA9, ADM1, HMOX1, LOX, SERPINE1 NDRG1, CA12, PDK1, VEGFA,
ERO1L, RORA, P4HA1, MXI1, SLC2A1(GLUT1), STC2, MIF, DDIT4, ENO1, CXCR4,
PLOD1, P4HA2, GAPDH, PGAM1, TMEM45A и PIM94
9004
.
Количественное определение метаболитов
Скорость поглощения глюкозы и секреции лактата определяли с помощью многопараметрической биоаналитической системы YSI 7100
(YSI Life Sciences).
