Каковы основные параметры транзистора КТ817Б. Где применяется данный транзистор. Какие есть отечественные и зарубежные аналоги КТ817Б. Как правильно выбрать замену для КТ817Б.
Основные характеристики транзистора КТ817Б
КТ817Б — это кремниевый эпитаксиально-планарный биполярный транзистор структуры n-p-n. Он относится к мощным низкочастотным транзисторам и имеет следующие ключевые параметры:
- Максимально допустимое напряжение коллектор-эмиттер: 45 В
- Максимально допустимый ток коллектора: 3 А
- Максимальная рассеиваемая мощность: 25 Вт (с теплоотводом)
- Коэффициент усиления по току: 25-275
- Граничная частота коэффициента передачи тока: не менее 3 МГц
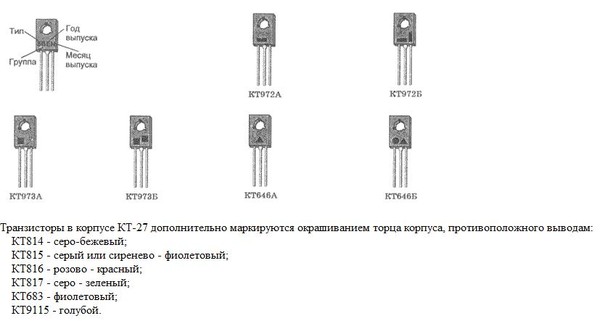
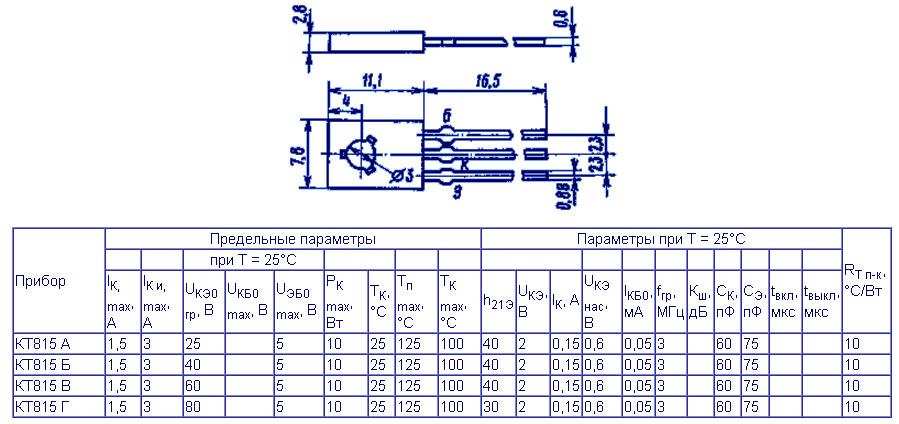
- Корпус: TO-126 (КТ-27)
Транзистор КТ817Б выпускается в пластмассовом корпусе TO-126 (отечественное обозначение КТ-27) с тремя выводами. Масса транзистора не превышает 0,7 г.
Области применения транзистора КТ817Б
Благодаря своим характеристикам, транзистор КТ817Б находит широкое применение в различных электронных устройствах:

- Усилители низкой частоты
- Выходные каскады усилителей мощности звуковой частоты
- Импульсные источники питания
- Схемы управления электродвигателями
- Стабилизаторы напряжения
- Преобразователи напряжения
- Коммутирующие и ключевые схемы
КТ817Б часто используется в паре с комплементарным транзистором КТ816Б для построения двухтактных усилителей мощности. Такая пара позволяет получить симметричный выходной каскад с хорошими характеристиками.
Отечественные аналоги транзистора КТ817Б
Среди отечественных транзисторов близкими аналогами КТ817Б являются:
- КТ817В — имеет более высокое напряжение коллектор-эмиттер (60 В)
- КТ817Г — рассчитан на напряжение коллектор-эмиттер до 80 В
- КТ815Б — близок по параметрам, но имеет меньший ток коллектора (2 А)
- КТ819Г — более мощный аналог на ток до 5 А
- КТ8116А — современный аналог с улучшенными характеристиками
При выборе отечественного аналога следует обращать внимание на соответствие основных параметров — напряжения, тока и мощности. Также важно учитывать тип корпуса для обеспечения совместимости при замене.

Зарубежные аналоги КТ817Б
Транзистор КТ817Б имеет множество зарубежных аналогов, среди которых можно выделить:
- BD233 — прямой аналог от Philips/NXP
- BD235 — аналог с немного большим напряжением
- TIP31A — популярный аналог от Texas Instruments
- 2SC2073 — японский аналог от Toshiba
- BDX33C — мощный аналог от STMicroelectronics
При выборе зарубежного аналога важно сравнивать не только основные электрические параметры, но и особенности применения транзистора в конкретной схеме. Некоторые зарубежные аналоги могут иметь отличия в усилительных свойствах или температурных характеристиках.
Как правильно выбрать замену для КТ817Б
При подборе замены для транзистора КТ817Б следует учитывать несколько ключевых факторов:
- Соответствие основных электрических параметров:
- Максимальное напряжение коллектор-эмиттер
- Максимальный ток коллектора
- Максимальная рассеиваемая мощность
- Коэффициент усиления по току
- Граничная частота усиления
- Совместимость корпуса и расположения выводов
- Соответствие теплового сопротивления переход-корпус
- Близость частотных и шумовых характеристик
- Совместимость по коэффициенту усиления в рабочей точке схемы
При замене КТ817Б на более современные транзисторы нужно учитывать, что они могут иметь лучшие характеристики. Это может потребовать корректировки режимов работы схемы, например, изменения сопротивления эмиттерного резистора для установки правильного тока покоя.
Особенности применения транзистора КТ817Б
При использовании транзистора КТ817Б в схемах следует учитывать некоторые особенности:
- Требуется обеспечить хороший теплоотвод для работы на максимальной мощности
- Рекомендуется использовать термокомпенсирующие цепи для стабилизации режима
- Необходимо учитывать разброс параметров и подбирать пары для симметричных схем
- При работе на высоких частотах может потребоваться коррекция АЧХ
Правильное применение транзистора КТ817Б с учетом его особенностей позволяет создавать надежные и эффективные электронные устройства различного назначения.
Сравнение КТ817Б с современными аналогами
Хотя транзистор КТ817Б остается популярным, современная электроника предлагает ряд улучшенных аналогов. Рассмотрим сравнение КТ817Б с некоторыми современными транзисторами:
| Параметр | КТ817Б | BD243C | TIP41C |
|---|---|---|---|
| Макс. напряжение К-Э | 45 В | 100 В | 100 В |
| Макс. ток коллектора | 3 А | 6 А | 6 А |
| Макс. рассеиваемая мощность | 25 Вт | 65 Вт | 65 Вт |
| Коэффициент усиления | 25-275 | 25-250 | 15-150 |
| Граничная частота | 3 МГц | 3 МГц | 3 МГц |
Как видно из сравнения, современные аналоги предлагают более высокие показатели по напряжению, току и мощности при сохранении основных характеристик усиления. Это позволяет создавать более эффективные и надежные устройства.
КТ817Б — Транзисторы — Радиодетали — Каталог
КТ817БКТ817Б
Транзисторы КТ817Б кремниевые эпитаксиально-планарные структуры n-p-n усилительные.
Предназначены для применения в усилителях низкой частоты, операционных и дифференциальных усилителях, преобразователях и импульсных устройствах.
Выпускаются в пластмассовом корпусе с жесткими выводами.
Тип прибора указывается на корпусе.
Масса транзистора не более 0,7 г.
Тип корпуса: КТ-27-2 (ТО-126).
Технические условия: аА0.336.187 ТУ/02.
Характеристики транзистора КТ817Б
Структура n-p-n
Максимально допустимое (импульсное) напряжение коллектор-база 45 В
Максимально допустимое (импульсное) напряжение коллектор-эмиттер 45 В
Максимально допустимый постоянный(импульсный) ток коллектора 3000(6000) мА
Максимально допустимая постоянная рассеиваемая мощность коллектора без теплоотвода (с теплоотводом) 1(25) Вт
Статический коэффициент передачи тока биполярного транзистора в схеме с общим эмиттером 25-275
Обратный ток коллектора Граничная частота коэффициента передачи тока в схеме с общим эмиттером =>3 МГц
Коэффициент шума биполярного транзистора Аналоги транзистора КТ817Б
2SC790, 2SD235, BD933, TIP31A, BD175, BD233, BD437, BD615
Uкбо — Максимально допустимое напряжение коллектор-база
Uкбои — Максимально допустимое импульсное напряжение коллектор-база
Uкэо — Максимально допустимое напряжение коллектор-эмиттер
Uкэои — Максимально допустимое импульсное напряжение коллектор-эмиттер
Iкmax — Максимально допустимый постоянный ток коллектора
Iкmax и — Максимально допустимый импульсный ток коллектора
Pкmax — Максимально допустимая постоянная рассеиваемая мощность коллектора без теплоотвода
Pкmax т — Максимально допустимая постоянная рассеиваемая мощность коллектора с теплоотводом
h31э — Статический коэффициент передачи тока биполярного транзистора в схеме с общим эмиттером
Iкбо — Обратный ток коллектора
fгр — граничная частота коэффициента передачи тока в схеме с общим эмиттером
Uкэн — напряжение насыщения коллектор-эмиттер
Транзистор кт 817 технические характеристики – 4apple – взгляд на Apple глазами Гика
Транзистор КТ817 – это кремниевый биполярный транзистор n-p-n типа, изготовленный по эпитаксиально-планарной технологии. Основное назначение КТ817 — применение в линейных и ключевых электрических схемах, модулях и блоках радио-электронных устройствах предназначенных для широкого использования.
Основное назначение КТ817 — применение в линейных и ключевых электрических схемах, модулях и блоках радио-электронных устройствах предназначенных для широкого использования.
Транзистор КТ817 – это кремниевый биполярный транзистор n-p-n типа, изготовленный по эпитаксиально-планарной технологии. Основное назначение КТ817 — применение в линейных и ключевых электрических схемах, модулях и блоках радио-электронных устройствах предназначенных для широкого использования.
Кремниевые эпитаксиально – планарные биполярные транзисторы. Предназначены для использования в ключевых и линейных схемах, блоках и узлах радиоэлектронной аппаратуры широкого применения.
Аналоги КТ817
- Прототип КТ817 Б – BD233
- Прототип КТ817 В – BD235
- Прототип КТ817 Г – BD237
Особенности
- Диапазон рабочих температур корпуса от – 60 до + 150 °C
- Комплиментарная пара – КТ816
Обозначение технических условий
Корпусное исполнение
- пластмассовый корпус КТ-27 (ТО-126) – КТ817 А, Б, В, Г
- пластмассовый корпус КТ-89 (DPAK) – КТ817 А9, Б9, В9, Г9
Назначение выводов
| Вывод (корпус КТ-27) | Назначение (корпус КТ-27) | Вывод (корпус КТ-89) | Назначение (корпус КТ-89) |
|---|---|---|---|
| №1 | Эмиттер | №1 | База |
| №2 | Коллектор | №2 | Коллектор |
| №3 | База | №3 | Эмиттер |
Технические характеристики транзистора КТ817
Таблица 1. Основные электрические параметры КТ817 при Токр.среды=25 °С
Основные электрические параметры КТ817 при Токр.среды=25 °С
| Паpаметpы | Обозн. | Ед.изм. | Режимы измерения | Min | Max |
|---|---|---|---|---|---|
| Граничное напряжение колл- эмит | |||||
| КТ817 А, А9 | Uкэо гp. | B | Iэ =0,1 A tи =0,3 – 1 мс | 25 | |
| КТ817 Б, Б9 | 45 | ||||
| КТ817 В, В9 | 60 | ||||
| КТ817 Г, Г9 | 80 | ||||
| Обратный ток коллектора | |||||
| КТ817 А, А9 | Iкбо | мкА | Uкэ =40 В | 100 | |
| КТ817 Б, Б9 | Uкэ =45 В | 100 | |||
| КТ817 В, В9 | Uкэ =60 В | 100 | |||
| КТ817 Г, Г9 | Uкэ =100 В | 100 | |||
| Обратный ток коллектор – эмиттер | |||||
| КТ817 А, А9 | I кэr | мкА | Uкэ =40 В, Rбэ≤1 кОм | 200 | |
| КТ817 Б, Б9 | Uкэ =45 В, Rбэ≤1 кОм | 200 | |||
| КТ817 В, В9 | Uкэ =60 В, Rбэ≤1 кОм | 200 | |||
| КТ817 Г, Г9 | Uкэ =100 В, Rбэ≤1 кОм | 200 | |||
| Статический коэффициент передачи тока | h21э | Uкб =2 B, Iэ =1 A | 25 | 275 | |
| Напряжение насыщения коллектор – эмиттер | Uкэ нас | В | Iк=1 A, Iб =0,1 A | 0,6 | |
Таблица 2. Предельно допустимые электрические режимы КТ817
Предельно допустимые электрические режимы КТ817
|
|
|||||||||||||||||||||||||||||||||||||||||||||||

Транзистор КТ814, КТ815, КТ816, КТ817
Транзистор КТ814, КТ815, КТ816, КТ817
Справочник содержания драгоценных металлов в радиодеталях основанный на справочных данных различных организаций занимающихся переработкой лома радиодеталей, паспортах устройств, формулярах и других открытых источников. Стоит отметить, что реальное содержание может отличатся на 20-30% в меньшую сторону.
Радиодетали могут содержать золото, серебро, платину и МПГ (Металлы платиновой группы, Платиновая группа, Платиновые металлы, Платиноиды, ЭПГ)
Содержание драгоценных металлов в транзисторе: КТ814, КТ815, КТ816, КТ817
Золото: 0.0043
Серебро: 0
Платина: 0
МПГ: 0
По данным: Справочник по драгоценным металлам ПРИКАЗ №70
Транзистор, полупроводниковый триод — радиоэлектронный компонент из полупроводникового материала, обычно с тремя выводами, способный от небольшого входного сигнала управлять значительным током в выходной цепи, что позволяет его использовать для усиления, генерирования, коммутации и преобразования электрических сигналов. В настоящее время транзистор является основой схемотехники подавляющего большинства электронных устройств и интегральных микросхем.
Типы транзисторов
Существует два основных типа транзисторов: биполярные и полевые.
1. Биполярные транзисторы. Они являются, вероятно, более распространенным типом (именно о них, например, шла речь в предыдущих разделах этой главы). В базу такого транзистора подается небольшой ток, а он, в свою очередь, управляет количеством тока, протекающего между коллектором и эмиттером.
2. Полевые транзисторы. Имеют три вывода, но они называются затвор (вместо базы у биполярного), сток (вместо коллектора) и исток (вместо эмиттера). Аналогично воздействие на затвор транзистора (но на этот раз не тока, а напряжения) управляет током между стоком и истоком. Полевые транзисторы также имеют разную полярность: они бывают N-канальные (аналог NPN-биполярного транзистора) и Р-канальные (аналог PNP).
Маркировка транзисторов СССР
Обозначение транзисторов до 1964 года
Первый элемент обозначения – буква П, означающая, что данная деталь и является, собственно, транзистором. Биполярные транзисторы в герметичном корпусе обозначались двумя буквами – МП, буква М означала модернизацию. Второй элемент обозначения – одно, двух или трехзначное число, которое определяет порядковый номер разработки и подкласс транзистора, по роду полупроводникового материала, значениям допустимой рассеиваемой мощности и граничной(или предельной) частоты.
От 1 до 99 – германиевые маломощные низкочастотные транзисторы.
От 101 до 199 – кремниевые маломощные низкочастотные транзисторы.
От 201 до 299 – германиевые мощные низкочастотные транзисторы.
От 301 до 399 – кремниевые мощные низкочастотные транзисторы.
От 401 до 499 – германиевые высокочастотные и СВЧ маломощные транзисторы.
От 501 до 599 – кремниевые высокочастотные и СВЧ маломощные транзисторы.
От 601 до 699 – германиевые высокочастотные и СВЧ мощные транзисторы.
От 701 до 799 – кремниевые высокочастотные и СВЧ мощные транзисторы.
Обозначение транзисторов после 1964 года
Первый символ необходим для обозначения типа используемого материала
Буква Г или цифра 1 – германий.
Буква К или цифра 2 – кремний.
Буква А или цифра 3 – арсенид галлия.
Второй символ обозначает тип транзистора
П – полевой транзистор
Т – биполярный транзистор
Третий символ необходим для обозначения мощности и граничной частоты
1 – транзисторы маломощные(до 0,3 ватт) низкочастотные(до 3 МГц).
2 – транзисторы маломощные(до 0,3 ватт) средней частоты(до 30 МГц).
3 – транзисторы маломощные(до 0,3 ватт) высокочастотные.
4 – транзисторы средней мощности(до 1,5 ватт), низкочастотные(до 3 МГц).
5 – транзисторы средней мощности(до 1,5 ватт),средней частоты(до 30 МГц).
6 – транзисторы средней мощности(до 1,5 ватт),высокочастотные и СВЧ.
7 – транзисторы мощные(свыше 1,5 ватт), низкочастотные(до 3 МГц).
8 – транзисторы мощные(свыше 1,5 ватт), средней частоты(до 30 МГц).
9 – транзисторы мощные(свыше 1,5 ватт), высокочастотные и СВЧ.
Четвертый и пятый элементы обозначения – определяют порядковый номер разработки.
Изменения в маркировке вступившие в силу в 1978 году. Изменения коснулись обозначения функциональных возможностей – третьего элемента.
Для биполярных транзисторов:
1 – транзистор с рассеиваемой мощностью до 1 ватта и граничной частотой до 30 МГц.
2 – транзистор с рассеиваемой мощностью до 1 ватта и граничной частотой до 300 МГц.
4 – транзистор с рассеиваемой мощностью до 1 ватта и граничной частотой более 300 МГц.
7 – транзистор с рассеиваемой мощностью более 1 ватта и граничной частотой до 30 МГц.
8 – транзистор с рассеиваемой мощностью более 1 ватта и граничной частотой до 300 МГц.
9 – транзистор с рассеиваемой мощностью более 1 ватта и граничной частотой свыше 300 МГц.
Поделиться ссылкой:
Понравилось это:
Нравится Загрузка…
Похожее
Скачать статью в одном файле Описан полосовой усилитель мощности с корректором. В качестве корректирующего элемента корректора использована управляемая нелинейная емкость коллектор-эмиттер закрытого низкочастотного транзистора. Технические характеристики усилителя:
Высокие требования к линейности амплитудной характеристики полосовых усилителей мощности предъявляются при усилении телевизионных, однополосных, многочастотных, амплитудно-модулированных радиосигналов [1, 2]. Методы повышения линейности основаны на оптимизации базового смещения транзисторов усилителя, работающих в режиме с отсечкой коллекторного тока [3-5], использовании предыскажений входного сигнала [6-8] и адаптивной компенсации [8-10]. Несмотря на теоретические преимущества схем адаптивной компенсации [9], они достаточно сложны в реализации, поскольку содержат в своем составе такие элементы как синхронный детектор, направленные ответвители, линии задержки, управляемый аттенюатор, сумматор, усилитель искажений. Поэтому, например, в усилителях телевизионных передатчиков эти схемы не находят применения. Для реализации предыскажений входного сигнала используются корректоры, в качестве основного элемента которых может быть применен варикап [11]. Недостатком использования корректора на варикапе является невозможность осуществления плавной регулировки и подстройки его дифференциального коэффициента усиления [12]. На рис. 1 приведена принципиальная схема полосового усилителя мощности с корректором амплитудной характеристики на транзисторе. Рис. 1. Принципиальная схема полосового усилителя мощности с повышенной линейностью амплитудной характеристики. (Щелкните мышью для получения большого изображения) , Т1, Т10 — КТ315В, Т2 — КТ816А, Т3 — КТ630А, Т4 — КТ361В, Т5 — КТ913В, Т6 — КТ930А, Т7, Т9 — КТ817Б, Т8 — КТ971А; D1 — КС224Ж, D2,D4 — КД509А,D3 — КС215Ж,D5 — КД213А; НО – направленный ответвитель. Усилитель содержит корректор, три каскада усиления, фильтр нижних частот, схемы защиты от перегрузки по входу, от рассогласования по выходу, от превышения напряжением питания номинального значения, термозащиту. В качестве корректирующего элемента корректора использована управляемая нелинейная емкость коллектор-эмиттер Скэ закрытого низкочастотного транзистора Т3. Экспериментальные исследования зависимости емкости Скэ ряда транзисторов от напряжений на переходах коллектор-эмиттер Uкэ и база-эмиттер Uбэ показали следующее. В транзисторе с закрытыми переходами емкость Скэ практически не зависит от напряжения Uкэ, если это напряжение больше или равно двум вольтам. В случае дальнейшего уменьшения напряжения Uкэ значение емкости Скэ плавно нарастает. Скорость нарастания значения емкости Скэ, при этом, зависит от величины напряжения Uбэ и увеличивается с уменьшением указанного напряжения. Работа корректора основана на увеличении коэффициента его передачи при увеличении емкости Скэ. Потенциометром R3 устанавливается напряжение Uкэ, потенциометром R4 устанавливается напряжение Uбэ транзистора Т3. Резистор R5 обеспечивает линеаризацию скорости нарастания коэффициента передачи корректора при увеличении значения емкости Скэ. При условии, что амплитуда сигнала подаваемого на вход корректора меньше или равна значению Uкэ -2 В, сигнал на выходе корректора оказывается пропорциональным входному воздействию. Дальнейшее увеличение сигнала подаваемого на вход корректора приводит к увеличению среднего за период значения емкости Скэ и соответствующему увеличению коэффициента передачи корректора. Таким образом, изменением напряжения Uкэ можно регулировать величину линейного участка амплитудной характеристики корректора, а изменением напряжения Uбэ скорость нарастания его коэффициента передачи в нелинейной области работы. Входной усилительный каскад на транзисторе Т5 работает в режиме класса А, предоконечный и выходной каскады на транзисторах Т6 и Т8 в режиме с отсечкой коллекторного тока. В усилителе использован стабилизатор напряжения базового смещения на транзисторах Т7 и Т9. Требуемая величина базового смещения транзисторов выходного и предоконечного каскадов устанавливается выбором номинала резистора R6. Методика настройки подобных усилительных каскадов подробно описана в [5]. Использование корректора может быть эффективным, если начальный участок амплитудной характеристики разрабатываемого усилителя линеен. Это достигается оптимизацией базового смещения транзисторов T6 и T8 с помощью резистора R6. На входе усилителя включен резистивный делитель напряжения, обеспечивающий его согласование с сопротивлением генератора при срабатывании схем защиты. С целью сохранения работоспособности усилителя при перегрузке по входу, на выходе резистивного делителя включен биполярный транзистор T2, играющий роль самоуправляемого ограничителя входных сигналов. Порог срабатывания ограничителя устанавливается делителем на резисторах R1 и R2. Подробное описание физики работы ограничителя и методика его настройки даны в [13]. Для устранения влияния емкости Cкэ закрытого транзистора Т2 на амплитудно-частотную характеристику усилителя, она включена в фильтр нижних частот третьего порядка, образуемый указанной емкостью и элементами L1 и C1. Ограничитель на транзисторе T2 применяется также в качестве управляемого ограничителя при срабатывании защит от рассогласования по выходу, от превышения напряжением питания номинального значения, термозащиты. Выбором номинала резистора R7 устанавливается величина коэффициента стоячей волны по напряжению со стороны нагрузки, при котором мощность сигнала на выходе усилителя начинает падать пропорционально росту этого коэффициента. Защита от превышения напряжением питания номинального значения выполнена на стабилитроне D1. Установка схемы термозащиты на заданную температуру срабатывания осуществляется с помощью резистора R8. Описание работы используемых схем защиты и методика их настройки приведены в [13]. На выходе усилителя установлен чебышевский фильтр нижних частот пятого порядка [14], обеспечивающий подавление высших гармонических составляющих в спектре выходного сигнала. На рис. 2 представлены экспериментальные амплитудные характеристики разработанного усилителя, показывающие возможности их коррекции с помощью предлагаемого корректора и полученные при условии оптимального выбора величины базового смещения транзисторов T6 и T8. Рис. 2 Экспериментальные амплитудные характеристики усилителя. В процессе перестройки корректора происходит незначительное изменение коэффициента передачи усилителя. Кроме того, при отключенном корректоре коэффициент передачи усилителя возрастает на 21 дБ. Поэтому на рис. 2, для удобства сравнения линейности амплитудных характеристик усилителя при различных режимах его работы, по оси ординат отложено эффективное значение амплитуды первой гармоники сигнала в нагрузке Uвых.эф., а по оси абсцисс отложена эффективная амплитуда ЭДС источника сигнала Ег.эф., нормированная относительно своего значения , соответствующего величине напряжения Uвых.эф.= 30 В. Прямая 1 на рис. 2 соответствует идеальной амплитудной характеристике усилителя, кривая 2 – амплитудная характеристика усилителя при отключенном корректоре, кривая 3 – при напряжениях на транзисторе корректора Uбэ = -2 В, Uкэ = 6 В, кривая 4 – при напряжениях на транзисторе корректора Uбэ = -1 В, Uкэ = 7 В, кривая 5 – при напряжениях на транзисторе корректора Uбэ = 0 В, Uкэ = 8 В. Анализ характеристик приведенных на рис. 2 позволяет установить следующее. Предлагаемый корректор дает возможность плавно изменять наклон амплитудной характеристики усилителя в области больших напряжений, обеспечить постоянство дифференциального коэффициента усиления вплоть до выходного напряжения равного 85% от максимального значения, в два раза увеличить выходную мощность, ограниченную областью линейной работы усилителя. Технические характеристики усилителя:
Усилитель сохраняет работоспособность при внезапном длительном отключении или коротком замыкании нагрузки; при эксплуатации устанавливается на радиатор с использованием принудительной вентиляции. Литература
Советую попробовать, не пожалеете! Удачи и 73 !!! |
Простой усилитель ЗЧ на трех транзисторах, схема (КТ3102, КТ816, КТ817)
Усилитель построен по простой схеме на трех транзисторах. На выходе, на нагрузке сопротивлением 4 Ом выдает мощность 2W при питании от источника напряжением 12V. Входное сопротивление усилителя мало, и составляет 470 Ом.
Столь малое входное сопротивление позволяет ему хорошо согласовываться с выходами портативной аппаратуры, рассчитанными на работу на головные телефоны. Не секрет, что у многих «гаджетов» телефонные усилители сделаны так, что не могут работать без нагрузки.
Если с такого телефонного выхода просто снять аудиосигнал, как с линейного выхода, то он будет искажен, наполнен треском и другими помехами. Если же к нему подключить резистор сопротивлением до 1 кОм, и снимать аудиосигнал уже с этого резистора, то все работает нормально.
Принципиальная схема
Рис. 1. Схема очень простого усилителя звукового сигнала, собранного на трех транзисторах.
Схема показана на рисунке. Практически, это типовая схема двухкаскадного УНЧ «из учебника». На транзисторе VТ1 сделан предварительный каскад усиления.
С коллектора VТ1 сигналы поступают на двухтактный выходной каскад на разноструктурных мощных транзисторах VТ2 и VT3. Диоды VD1 и VD2 создают разность постоянных напряжений на базах транзисторов выходного каскада, таким образом, устраняя искажения типа «ступенька», и снижая коэффициент нелинейных искажений.
Детали и печатная плата
Усилитель собран на небольшой печатной плате. Транзисторам КТ817 и КТ816 при такой выходной мощности особо крупных радиаторов не требуется, достаточно собрать «экспромтом» по пирамидке из болта, гаек и шайб.
Для стерео варианта нужно два таких усилителя.
В выходном каскаде можно пару КТ817А-КТ816А заменить парой КТ817-КТ816 или КТ815-КТ814 с любыми буквенными индексами, но одинаковыми.
Рис. 2. Печатная плата для усилителя звука, выполненного на трех транзисторах.
Транзистор КТ3102 тоже может быть с любым буквенным индексом.
Налаживание
Налаживание сводится к подбору сопротивления R2 по напряжению на эмиттерах VТ2 и VT3, равному половине напряжения питания.
Такой усилитель можно использовать и как ремонтный модуль.
Попцов Г. РК-04-18.
MJ2254
Аннотация: 2N3740 motorola MJ3701 CI 8002 MJ2249 MJ310 MJ2250 Motorola германиевый транзистор pnp MJ2253 MJ3101
|
OCR сканирование |
MJ2253 MJ2254 MJ3701 MJ2249, MJ2250, MJ3101 MJ3701 2Н3740, г. 2N3740 моторола CI 8002 MJ2249 MJ310 MJ2250 Motorola германиевый транзистор pnp MJ3101 | |
2010 — КТ835А
Абстракция: b0330 MJ032 2N3171 b0362 2SA699A matsua MJ-03 2sa1009am M0810 BO150
|
Оригинал |
3000н 126вар 202вар КТ835А b0330 MJ032 2N3171 b0362 2SA699A мацуа MJ-03 2sa1009am M0810 BO150 | |
Нет в наличии
Аннотация: абстрактный текст недоступен
|
OCR сканирование |
PL500 2Н3740, г. 2Н3741, г. 2N4898 2N4699 2N4900 MJ2253 MJ2254 MJ3701 MJ3702 2Н3741 МОТОРОЛА 2N3740 моторола motorola транзистор PNP 2C4900 | |
2010 — 2sc630
Абстракция: KT817B 2SD130 IDB434 2SB1003 2SC790R MJE32 2SD130R 2sd1369 bd57
|
Оригинал |
КТ817А 2SD91 IDB1023 IDD1413 2SB1003 2SD1369 BDY34 2N3632 See00n 2sc630 КТ817Б 2SD130 IDB434 2SC790R MJE32 2SD130R bd57 | |
2010 — B0813
Абстракция: 8C440 PT9787 MJ3237 MJ2841 MOTOROLA MM1758 trw PT9787 MM4048 MJE42C MM1712
|
Оригинал |
2SC109S 2NS714 92PU01 2С0180С MPSU01 НСОУ01 92GU01 NA31KY B0813 8C440 PT9787 MJ3237 MJ2841 MOTOROLA MM1758 trw PT9787 MM4048 MJE42C MM1712 | |
mje 3001
Аннотация: MJ3583 2N3583 MOTOROLA транзистор SE 431 MJ3761 MJ3760 MJ2253 2N3583 MJ3701 MJ3585
|
OCR сканирование |
MJ3583 MJ3585 2N3583 MJ3701 MJ2253 MJ3760 MJ3761 MJ3760 MJ3761 16-КОЛЛЕКЦИОННАЯ БАЗА mje 3001 2Н3583 МОТОРОЛА транзистор SE 431 MJ3701 MJ3585 | |
MJE13005A
Аннотация: MJ3237 MJ4361 MJE10001 MJ2901 MJE240 MJ430 MJ9000 MJ2252 MJ3247
|
Оригинал |
MJ13330 О-204АА / ТО-3: MJ13101 MJ13331 MJ13333 MJE13005A MJ3237 MJ4361 MJE10001 MJ2901 MJE240 MJ430 MJ9000 MJ2252 MJ3247 | |
JE1100
Аннотация: MFC8030 MFC8040 MC1316 Triac 9707 схема транзистора mc1466 MJE 5332 AN294 National Semiconductor Linear Data Book MC4039P
|
OCR сканирование |
||
sx3704
Аннотация: AP239 Транзистор 80139 8C547 2N50B IN2222A 6C131C радио AC176 AC126 sft353 2N2064
|
OCR сканирование |
||
hep 154 кремниевый диод
Реферат: zy 406 транзистор hep R1751 motorola HEP 801 hep 154 диодный симистор zd 607 2sb337 F82Z hep 230 pnp RS5743.3
|
OCR сканирование |
MY110B Z0206 Z0208 Z0210 Z0211 Z0212 Z0214 Z0215 Z0217 Z0219 кремниевый диод hep 154 zy 406 транзистор гепатит R1751 Motorola HEP 801 hep 154 диод симистор zd 607 2sb337 F82Z hep 230 pnp RS5743.3 | |
Powec RM 1110
Реферат: инвертор rm 1100 powec сварочный аппарат 4 схема транзисторов JX 6822 A MPC1000 as11 dc hee nv германиевый транзистор PL 15Z DIODE stg 8810 транзистор SI 6822
|
OCR сканирование |
111ii MZ5558 Z5555, Z5556, MZ5557 Powec RM 1110 RM 1100 Powec схема инвертора сварочного аппарата 4 транзисторы JX 6822 A MPC1000 as11 dc hee nv германиевый транзистор PL 15Z ДИОД stg 8810 транзистор СИ 6822 | |
TNY 176 PN ЭКВИВАЛЕНТ
Аннотация: Справочник по свободным транзисторам SN76477 2n4401 Справочник по данным по полупроводникам mj802 2N3866 s2p tis43 bc149c Примечания по применению XR2206 Руководство по транзисторам Siemens LM131
|
OCR сканирование |
2114L 6116P3 6116LP3 AY-3-1270 AY-3-1350 AY-3-8910 AY-3-8912 AY-5-1230 CA3080E CA3130E TNY 176 PN ЭКВИВАЛЕНТ SN76477 2n4401 книга эквивалентов свободных транзисторов Справочник по полупроводникам mj802 2N3866 s2p tis43 bc149c Замечания по применению XR2206 LM131 siemens транзистор руководство | |
Германиевый дрейфовый транзистор
Реферат: 2N4895 германиевый транзистор эпитаксиальный меза-транзистор sec tip31A транзистор халблайтера транзистор BD222 BD699 EQUIVALENT kd 2060 транзистор
|
OCR сканирование |
орпорация / 464 CH-8105 Германиевый дрейфовый транзистор 2N4895 германиевый транзистор эпитаксиальная меза транзистор sec tip31a транзистор с индексом халблайтера транзистор BD222 BD699 ЭКВИВАЛЕНТ kd 2060 транзистор | |
NT101
Реферат: БФ503 КТ-934 КТ606 ПОЛУЧАЙНЫЕ ИНДЕКСЫ Mps56 транзисторы 2SA749 72284 2sk81 2SB618
|
OCR сканирование |
||
транзистор индекса халблайтера
Аннотация: германиевая электрическая схема аудиоусилителя IC 6283 Transistor Shortform Datasheet & Cross References Inverter Welder 4 Схема транзистора BF 175 1N5159 de ic LG 8838 Z620 BRIDGE германиевый транзистор
|
OCR сканирование |
4L3052 4L3056 транзистор с индексом халблайтера германий схема подключения аудиоусилителя ic 6283 Краткое описание транзистора и перекрестные ссылки схема инвертора сварочного аппарата 4 транзистор bf 175 1N5159 диаграммы де IC LG 8838 Z620 МОСТ германиевый транзистор | |
T35W
Аннотация: Транзистор 65е9 2SC965 транзистор кт 606а ср 6863 D CS9011 sr1k диод КТ850 транзистор 130001 8d транзистор st25a
|
OCR сканирование |
10DB2P 10DB4P 10DB6P 180B6A T35W 65e9 транзистор 2SC965 транзистор кт 606а SR 6863 D CS9011 sr1k диод КТ850 транзистор 130001 8d ТРАНЗИСТОР СТ25А | |
БТИЗ M16 100-44
Аннотация: Ericsson RBS 6102 ASEA HAFO AB GM378 Transistor B0243C Kt606 Ericsson SPO 1410 SEMICON INDEXES transistor 8BB smd tr / NEC Tokin 0d 108
|
OCR сканирование |
W211d W296o W211c БТИЗ M16 100-44 Ericsson RBS 6102 ASEA HAFO AB GM378 Транзистор B0243C Kt606 Ericsson SPO 1410 ПОЛУЧЕННЫЕ ИНДЕКСЫ транзистор 8BB smd tr / NEC Tokin 0d 108 | |
aeg диод Si 61 L
Реферат: aeg диоды D6 BC23B SILICONIX U315 SGS Transistors 2N3750 2N339 2N3299 транзистор bf 175 2N2197 GENERAL ELECTRIC
|
OCR сканирование |
11tA0A12 A025A A0290 U0U55 A0291 A0292 A0305 A0306 A0A56 A0A59 aeg диод Si 61 L aeg диоды D6 BC23B СИЛИКОНИКС U315 Транзисторы SGS 2N3750 2N339 2N3299 транзистор bf 175 2N2197 ОБЩАЯ ЭЛЕКТРИКА | |
Схема усилителя мощности 2n3773
Аннотация: Справочник по SCR, RCA HC2000H RCA транзистор npn a13 RCa T2850D 40659 npn транзистор RCA 467 B0241C triac t6440m DIAC D3202U
|
OCR сканирование |
Ан-6671 G4000) G4000 Схема усилителя мощности 2N3773 Справочник по SCR, RCA HC2000H rca транзистор npn a13 RCa T2850D 40659 npn транзистор RCA 467 B0241C симистор t6440m DIAC D3202U | |
SEM 5027A
Аннотация: транзистор BD 540 LYS MC710G 131-80 wj 89 LN4005 обратный ток и напряжение диода 2n328 C844P MC707G mc708g Транзистор, связанный с h2 3003A
|
OCR сканирование |
||
т110 94в 0
Аннотация: PTC SY 16P 2N2955T диод Philips PH 37m 35K0 trimble R8 модель 2 2sc497 2SA749 2n6259 ssi 2N4948 NJS
|
OCR сканирование |
Барселона-28, С-171 CH-5400 t110 94v 0 PTC SY 16P 2N2955T Philips диод PH 37m 35K0 trimble R8 модель 2 2sc497 2SA749 2n6259 SSI 2N4948 NJS | |
Схема усилителя la 4440 300 ватт
Реферат: схема усилителя la 4440 300 ватт DA 3807 pdf транзисторный диод LT 7229 2sd323 YM 7137 3D DIAC 1N5761 инверторный сварочный аппарат 4 схема LA 4301 THYRISTOR br 403
|
OCR сканирование |
Ан-784А Схема усилителя la 4440 300 ватт Схема усилителя la 4440 300 ватт DA 3807 pdf транзистор диод LT 7229 2sd323 YM 7137 3D DIAC 1N5761 схема инвертора сварочного аппарата 4 LA 4301 ТИРИСТОР BR 403 | |
MC2259
Аннотация: Спецификация германиевого диода MC9713P MC880P mc2257 MC9718P 1N4465 MC9802P 1N4003 3N214 2N1256 S P
|
OCR сканирование |
||
y51 h 120c
Абстракция: ac128 bd192 bd124 MM1711 BD214 al103 KT368 AFY18 BFQ59
|
OCR сканирование |
500 мА 500 мА 240 МВт 240 МВт y51 h 120c ac128 bd192 bd124 MM1711 BD214 al103 КТ368 AFY18 BFQ59 | |
MJE2370
Аннотация: 2sc 1473
|
OCR сканирование |
MJE2370 MJE2S20 MJE2370 2сб 1473 | |
MJE2370
Аннотация: MJE2520 2sc 043 538 NPN транзистор
|
OCR сканирование |
MJE2520 MJE2370 MJE2370 MJE2520 2sc 043 538 NPN транзистор | |
MJE2370
Аннотация: транзистор MJE29 MJE30 2N5190 MOTOROLA MJE31 mje29 Транзисторы Motorola 2N5192 MJE32 MJE4918 2N6122 транзистор
|
OCR сканирование |
PL500 2C5192 2C5195 2N5190 2N5191 2N5192 2N5869 2NS870 2N6121 MJE2370 транзистор mje29 MJE30 2N5190 MOTOROLA MJE31 mje29 Транзисторы Motorola 2N5192 MJE32 MJE4918 2Н6122 транзистор | |
2010 — 2SA490-0
Абстракция: 2SA699A matsua B0934 MJE2491 MJE2370 2SA1359Y MJ2253 043C4 2sb856 1S2525
|
Оригинал |
О-220АБ О-220 ОТ-186 О-126вар О-127вар 2SA490-0 2SA699A мацуа B0934 MJE2491 MJE2370 2SA1359Y MJ2253 043C4 2sb856 1С2525 | |
2010 — 2sc630
Абстракция: KT817B 2SD130 IDB434 2SB1003 2SC790R MJE32 2SD130R 2sd1369 bd57
|
Оригинал |
КТ817А 2SD91 IDB1023 IDD1413 2SB1003 2SD1369 BDY34 2N3632 See00n 2sc630 КТ817Б 2SD130 IDB434 2SC790R MJE32 2SD130R bd57 | |
2010 — B0813
Абстракция: 8C440 PT9787 MJ3237 MJ2841 MOTOROLA MM1758 trw PT9787 MM4048 MJE42C MM1712
|
Оригинал |
2SC109S 2NS714 92PU01 2С0180С MPSU01 НСОУ01 92GU01 NA31KY B0813 8C440 PT9787 MJ3237 MJ2841 MOTOROLA MM1758 trw PT9787 MM4048 MJE42C MM1712 | |
1998 — DK53
Резюме: dk52 2SC4977 MJE102 BD699 2SA1046 BU808DFI эквивалент 2n3055 замена BUH513 замена MJ2955
|
Оригинал |
2N3016 2N3021 2N3022 2N3023 2N3024 2N3025 2N3026 2N3055 2N3076 2N3171 DK53 dk52 2SC4977 MJE102 BD699 2SA1046 Эквивалент BU808DFI 2n3055 замена BUH513 Замена MJ2955 | |
1997 — DK53
Абстракция: dk52 BU724AS mje2055 2n3055 замена BUX98PI BD263 BD699 BD292 2N5037
|
Оригинал |
2N3016 2N3021 2N3022 2N3023 2N3024 2N3025 2N3026 2N3055 2N3076 2N3171 DK53 dk52 BU724AS mje2055 2n3055 замена BUX98PI BD263 BD699 BD292 2N5037 | |
BUV48I
Аннотация: BU808DXI BD699 buv18a BD241CFI транзистор 2SA1046 BUW52I BU808DFI эквивалент BU724AS 2SA1046
|
Оригинал |
2N3016 2N3021 2N3022 2N3023 2N3024 2N3025 2N3026 2N3055 2N3076 2N3171 BUV48I BU808DXI BD699 buv18a BD241CFI транзистор 2SA1046 BUW52I Эквивалент BU808DFI BU724AS 2SA1046 | |
B0411
Резюме: B0733 2SC4977 THD200F1 dk52 2N5415 ЗАМЕНА НАКОНЕЧНИКА 2n3055 BD68D SGS-Thomson перекрестная ссылка BUX37 THOMSON
|
OCR сканирование |
2N3016 2N3021 2N3022 2N3023 2N3024 2N3025 2N3026 2N3055 2N3076 2N3171 B0411 B0733 2SC4977 THD200F1 dk52 2Н5415 ЗАМЕНА СОВЕТ 2n3055 BD68D Перекрестная ссылка SGS-Thomson BUX37 THOMSON | |
2001 — 2SA1046
Резюме: mje15033 замена 2SC1030 BD417 BD415 BU108 2SB528 BD262 2SC2080 BD295
|
Оригинал |
2N6057 2N6059 2N6050) 2Н6111, г. 2N6288 2N6109 2N6107, г. 2N6292 2SA1046 mje15033 замена 2SC1030 BD417 BD415 BU108 2SB528 BD262 2SC2080 BD295 | |
2001 — эквивалент tip162
Абстракция: 2N3055 BU108 2SA1046 2n6258 2N5981 pnp транзистор 2sd314 BD262 bd876 BU100
|
Оригинал |
2N6338 2N6339 2N6340 2N6341 2N6436 Продолжить32 TIP73B TIP74 TIP74A TIP74B эквивалент tip162 2N3055 BU108 2SA1046 2n6258 2N5981 транзистор pnp 2sd314 BD262 bd876 BU100 | |
2001 — 2SA1046
Абстракция: BU108 BU326 BU100 BD262 2sb688 y 2n5632 bd876
|
Оригинал |
2N6497 2N6498 2N6498 * TIP73B TIP74 TIP74A TIP74B 2SA1046 BU108 BU326 BU100 BD262 2сб688 г 2n5632 bd876 | |
2001 — замена mje15033
Резюме: Замена BD262 DARLINGTON 2SB554 Замена 2sc1079 2SD323 Замена 2N6254 Замена BD262A DARLINGTON 2n3055 Замена 2SC1013 Замена MJ2955
|
Оригинал |
2N3055 2N6576 2N6577 2N6578 TIP73B TIP74 TIP74A TIP74B TIP75 mje15033 замена BD262 ДАРЛИНГТОН 2SB554 замена 2sc1079 замена 2SD323 2Н6254 ЗАМЕНА BD262A ДАРЛИНГТОН 2n3055 замена 2SC1013 замена Замена MJ2955 | |
2001 — БУ108
Аннотация: транзистор Bc 574 MJ * 15033 BU100 BU326 2SC1419 MJ3055 to220 2SC1943 2n6107 MOTOROLA 2N6277
|
Оригинал |
BUV21 TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A TIP75B TIP75C 2N6488 BU108 транзистор Bc 574 МДж * 15033 BU100 BU326 2SC1419 MJ3055 — 220 2SC1943 2н6107 МОТОРОЛА 2N6277 | |
2001 — Д45х21 эквивалентная замена
Аннотация: Эквивалент 2N5036 BU108 Эквивалент BDW93C MJ10009 BU100 BU326 Эквивалент 2SB557 Эквивалент BD420 Эквивалент MJE6044
|
Оригинал |
MJ10009 Вольт32 TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A TIP75B TIP75C Эквивалентная замена Д45х21 2N5036 эквивалент BU108 Эквивалент BDW93C BU100 BU326 2SB557 эквивалент Эквивалент BD420 Эквивалент MJE6044 | |
2001 — транзистор 3569
Реферат: Т4 3570 дпак БУ 508 транзистор БУ108 МДЖ15015 ТРАНЗИСТОР ЗАМЕНЫ РУКОВОДСТВО MJ * 15033 БУ100 БУ326 транзистор Т4 3570 СТ Т4 3580
|
Оригинал |
Bandwi32 TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A TIP75B TIP75C 2N6488 транзистор 3569 t4 3570 дпак БУ 508 транзистор BU108 РУКОВОДСТВО ПО ЗАМЕНЕ ТРАНЗИСТОРА MJ15015 МДж * 15033 BU100 BU326 транзистор t4 3570 СТ Т4 3580 | |
2001 — БУ108
Аннотация: 2N3055 AN415A mje13005 BU100 BU326 2n3055 примечания по применению аудиоусилителя BDX54 MJE2955T ST BDV64
|
Оригинал |
MJE2955T MJE3055T TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A TIP75B TIP75C BU108 2N3055 AN415A mje13005 BU100 BU326 Примечание по применению усилителя звука 2n3055 BDX54 MJE2955T ST BDV64 | |
2001 — БД127
Аннотация: транзистор 2SA1046 2SD630 электронный балласт MJE13005 BD388 транзистор bd4202 BU108 motorola AN485 2SC122 MJE340 MOTOROLA
|
Оригинал |
MJE / MJF18206 MJE18206 MJF18206 POW32 TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A BD127 транзистор 2SA1046 2SD630 электронный балласт MJE13005 BD388 транзистор bd4202 BU108 моторола AN485 2SC122 MJE340 MOTOROLA | |
2001 — конфигурация выводов NPN транзистора tip41c
Аннотация: Эквивалентный транзистор TIP41C. Конфигурация выводов tip41c. Конфигурация выводов. NPN-транзистор.
|
Оригинал |
TIP41A, TIP42A TIP41B, TIP42B TIP41C, TIP42C TIP41A TIP41B * TIP41C * Конфигурация контактов NPN-транзистор tip41c ЭКВИВАЛЕНТ TIP41C КОНФИГУРАЦИЯ ПИН ТРАНЗИСТОРА tip41c Конфигурация контактов NPN-транзистор tip42c ЭКВИВАЛЕНТ TIP42C Все аналогичные транзисторы 2SA715 BU108 MJE520 МОТОРОЛА Полупроводники TIP41A 2СА634 М | |
2001 — конфигурация выводов транзистора bd140
Аннотация: 2SD669 эквивалентный транзистор BUV44 bd140 эквивалент MJE15020 bd140 эквивалентный BD250C эквивалентный RCA122 2N6045 NPN POWER Дарлингтон транзистор BD 136
|
Оригинал |
BD136 BD138 BD140 BD140-10 TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A конфигурация выводов транзистора bd140 2SD669 эквивалент BUV44 bd140 эквивалентный транзистор MJE15020 эквивалент bd140 BD250C ЭКВИВАЛЕНТ RCA122 2N6045 NPN Пауэр Дарлингтон ТРАНЗИСТОР BD 136 | |
2001 — IR642
Аннотация: 2N6410 2SC931 2SD375 IR3001 BU108 bu180 BD241 2SD675 2SC1986
|
Оригинал |
BD787, г. BD788 BD787 TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A IR642 2N6410 2SC931 2SD375 IR3001 BU108 bu180 BD241 2SD675 2SC1986 | |
2001 — техас 2n3055
Аннотация: BU108 BU100 BU326 2N5655 эквивалент 2n3055 замена MJ2955 замена BDX54 2SD424 2N5981 pnp транзистор
|
Оригинал |
2N3055 MJ2955 * TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A TIP75B TIP75C техас 2n3055 BU108 BU100 BU326 2N5655 эквивалент 2n3055 замена Замена MJ2955 BDX54 2SD424 2N5981 транзистор pnp | |
2001 — IC 3526
Аннотация: Транзистор BU108 MJ15024 RCA1C13 tip3055, эквивалентная схема на основе MJE13005 2N3791 аналога Motorola MJ3000 2N5981 pnp-транзистора MJ3247
|
Оригинал |
BD802 BD802 TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A TIP75B TIP75C IC 3526 BU108 транзистор MJ15024 RCA1C13 tip3055 эквивалент схема на базе MJE13005 2N3791 эквивалент моторола MJ3000 2N5981 транзистор pnp MJ3247 | |
2001 — эквивалент BDV65B
Аннотация: buv48 эквивалент BU108 tip127 pin детали 2SD424 BDX54 BU326 BU100 mje340
|
Оригинал |
BDV65B BDV64B TIP73B TIP74 TIP74A TIP74B TIP75 TIP75A TIP75B TIP75C Эквивалент BDV65B эквивалент buv48 BU108 tip127 pin детали 2SD424 BDX54 BU326 BU100 mje340 | |
Динамика развития морфогенеза эноцитов в Tribolium
Контекст 1
…содержаться в оеноцитах личинок Tribolium. Поскольку антитело Drosophila HNF4 не смогло распознать специфический паттерн в эмбрионах Tribolium, мы затем создали антисыворотку против белка Tribolium HNF4. Как показано на рис. 1B, мы обнаружили паттерн экспрессии HNF4, который маркирует многообещающий набор кластеров клеток, которые ограничены брюшными сегментами у 46h эмбрионов. Используя оптические срезы, эти клетки можно отличить от подлежащей мезодермы HNF4 + как по интенсивности окрашивания HNF4, так и по их латеральному положению.В соответствии с тем, что эти HNF4-положительные клетки являются личиночными оеноцитами, они также коэкспрессируют липид-модифицирующий фермент (увы, приложение, рис. 1), который экспрессируется в оеноцитах дрозофилы, а также несколько других маркеров эноцитов, описанных ниже. Более того, совместное окрашивание с антителом Spalt подтвердило, что в отличие от сильной активации, наблюдаемой в зрелых оеноцитах D. melanogaster, оеноциты в более старых эмбрионах T. castaneum не обладают экспрессией Spalt (рис. 1B ’’ ’). В процессе подтверждения того, что HNF4-экспрессирующие абдоминально-специфические клетки в эмбрионах Tribolium являются личиночными оеноцитами, мы по счастливой случайности идентифицировали новый биомаркер для эноцитов.При тестировании множества антител на перекрестную реактивность в Tribolium (полный список антител см. В дополнительной таблице 1) мы попытались усилить сигнал с помощью системы мечения биотин-стрептавидин. Вместо этого мы обнаружили, что всякий раз, когда использовался биотин-стрептавидин, устойчивый устойчивый сигнал специфически маркирует цитоплазму тех же кластеров клеток брюшной полости, которые экспрессируют HNF4, а также отросток плевропода в сегменте 1 брюшной полости (Рис. 1C). Последующее поэтапное удаление как первичных, так и вторичных антител показало, что добавление одной только молекулы стрептавидина, конъюгированной с флуорофором, приводит к этой специфической для брюшной полости паттерну.Сильное мечение личиночных оеноцитов стрептавидином, вероятно, связано с этим типом клеток, содержащим высокие уровни биотина, который служит важным коферментом для метаболизма жирных кислот (Fletcher and Myant, 1960). Таким образом, флуоресцентный конъюгированный стрептавидин является полезным красителем для распознавания личиночных эноцитов в эмбрионах Tribolium. Затем мы определили общую применимость стрептавидина в качестве биомаркера эноцитов, протестировав окраску на двух других насекомых. Хотя стрептавидин не распознавал личиночные эноциты во время эмбриогенеза D melanogaster, это окрашивание было очень эффективным для мечения эноцитов у личинок третьего возраста (рис.1D). Более того, способность стрептавидина распознавать этот тип клеток имеет дополнительное преимущество, заключающееся в том, что не требуется рассечение личинок (см. Методы). Чтобы дополнительно проверить применимость окраски стрептавидином к насекомым, мы собрали эмбрионы полуокрылых видов рода Anasa (часто называемые жуками-кабачками) из тыквенной тыквы в юго-западном Огайо. Яичная скорлупа вручную отделялась от каждого эмбриона на поздней стадии (фиг. 1E), и нанесение флуоресцентно-конъюгированного стрептавидина выявило сильное окрашивание областей живота, соответствующее скоплениям эноцитов (стрелки, фиг. 1F ’).Однако следует отметить, что окрашивание стрептавидином у Anasa, а также у Drosophila и Tribolium также метило подмножество других клеток, включая нейроны. Следовательно, по возможности следует использовать дополнительные маркеры для определения клеток, меченных стрептавидином, как эноцитов. Тем не менее, эти данные предполагают, что стрептавидин, вероятно, будет широко применимым маркером личиночных оеноцитов у многих видов насекомых. Мощным инструментом для маркировки определенных типов клеток в модельных организмах является использование линий ловушек энхансеров.Недавняя работа нескольких исследовательских групп сделала множество таких ловушек энхансеров GFP в Tribolium castaneum доступными через базу данных GEKU ((Trauner et al., 2009). Сканирование доступных линий ловушек энхансеров выявило несколько интересных паттернов экспрессии, специфичных для брюшной полости. из этих линий, KT817, был выбран для анализа, и было обнаружено, что он экспрессирует GFP по той же схеме, что и клетки, меченные HNF4 (рис. 2B) и стрептавидином (рис. 2C). (Обратите внимание на дополнительное слабое окрашивание GFP в каждом сегменте. к неинсерционной сайт-специфической активности конструкции GFP).Используя протокол iPCR (см. Методы), мы определили, что piggybac-GFP интегрирована на расстоянии ~ 60 килобайт от гомолога Tribolium в вентральных венах, лишенного гена (vvl), который, как известно, экспрессируется в ранних личиночных оеноцитах в D melanogaster (Inbal et al. др., 2003). Таким образом, мы полагаем, что специфический для эноцитов паттерн экспрессии, скорее всего, обусловлен энхансером Tribolium vvl, несмотря на присутствие нескольких других предсказанных генных моделей в локусе. Этот вывод становится более правдоподобным из-за относительно большого расстояния между генами в геноме Tribolium, а также из-за предыдущих находок, описывающих энхансеры более чем на 100 kb от их генов-мишеней (Levine, 2010).Следовательно, ловушка энхансера KT817 в дальнейшем упоминается как vvl-GFP. Затем мы использовали vvl-GFP и антитело HNF4 в качестве маркеров для изучения развития личиночных оеноцитов Tribolium во время эмбриогенеза (Рис. 3 и Дополнение Рис. 2). Антитело против фактора транскрипции Cut также использовали в качестве маркера близлежащих трахеальных ямок (рис. 3E). Для этого анализа мы выполнили синхронизированные коллекции эмбрионов и определили несколько временных точек и ориентиров развития, которые коррелируют с морфологическими переходами для эноцитов.Во-первых, начальные следы экспрессии vvl-GFP можно увидеть вскоре после завершения расширения зародышевой полосы (26 часов), и это окрашивание быстро усиливается, выявляя кластер примерно из 15 клеток в каждом сегменте брюшной полости (Рис. 3B на 28 часах). В это время GFP-положительные клетки лишены экспрессии HNF4, а сигнал GFP имеет небольшую, но постоянно более высокую интенсивность в передних (A1-A3) по сравнению с задними (A4-A8) сегментами брюшной полости (рис. 3B на 28 часах). Этот результат согласуется с тем, что Tribolium — насекомое с короткими зародышевыми полосами, которое в процессе развития добавляет новые (более молодые) задние сегменты брюшка (Schroder et al., 2008). К 34 часам каждый из сегментов брюшной полости от A1 до A8 имеет устойчивые клетки, экспрессирующие vvl-GFP, которые совместно экспрессируют HNF4 (рис. 3B-D), и эти клетки образуют кластер клеток в форме полумесяца вокруг Cut-положительной трахеальной ямки. По завершении ретракции зародышевой ленты (46 ч) количество GFP- и HNF4-положительных оеноцитов увеличивается до более чем 20 клеток на кластер в большинстве сегментов брюшной полости, и эти клетки присутствуют в одном кластере дорсальнее трахеальной ямки в каждом из них. сегмент. К 50 часам суставы ног образуют четкие границы, и удлинение кишечной трубки можно увидеть у эмбрионов, наклоненных в сторону дорсальной проекции (не показано).На этом этапе скопления эноцитов начинают распространяться как кзади, так и внутрь от покровов, что приводит к формированию характерной формы акульих зубов. Кроме того, многие клетки, не являющиеся эноцитами, начинают экспрессировать высокие уровни HNF4, но эти клетки не распознаются стрептавидином и не экспрессируют vvl-GFP (фиг. 3D через 50 часов и данные не показаны). К 58 часам кишечная трубка образует полную петлю, и дорсальное закрытие завершается. На этой стадии GFP-положительные клетки все еще сильно окрашиваются стрептавидином, но экспрессия HNF4 значительно снизилась.Более того, эноциты образуют непрерывную полосу, проходящую через сегментарные границы, и мигрировали дорсально, морфология соответствует классическому описанию личиночных оеноцитов жесткокрылых (Snodgrass, 1993; Wheeler, 1892). Оеноциты дрозофилы первоначально специфицируются у эмбрионов на стадии 11, когда клетки дорсальной эктодермы, которые экспрессируют Spalt, получают временный сигнал EGF (Elstob et al., 2001; Rusten et al., 2001). Клетка-отправитель EGF, специфический предшественник сенсорного органа (SOP), может быть помечена с использованием трансгенной репортерной линии, взятой из ромбовидного локуса: Rho654-LacZ, а принимающие EGF клетки могут быть визуализированы с использованием антитела фосфо-ERK, которое распознает активированный Путь передачи сигналов MAP-киназы (рис. 4A) (Gabay et al., 1997; Ли-Крегер и др., 2008; Witt et al., 2010). Клетки, принимающие EGF, впоследствии активируют и поддерживают экспрессию Spalt во время созревания эноцитов. Напротив, мы обнаружили, что зрелые оеноциты Tribolium (отмеченные HNF4 и стрептавидином) не поддерживают экспрессию Spalt (рис. 1B). Однако возможно, что оеноциты Tribolium первоначально рекрутируются из Spalt-экспрессирующих клеток дорсальной эктодермы и впоследствии подавляют этот фактор во время созревания. Чтобы проверить эту идею, мы тщательно проанализировали раннюю экспрессию Spalt, фосфо-ERK и vvl-GFP в эмбрионах жуков.Как показано на рис. 4В, небольшой вентральный выход Spalt-положительных клеток слабо, но постоянно окрашивается с использованием антитела фосфо-ERK. Хотя эти эмбрионы не экспрессируют трансген vvl-GFP на этой стадии развития, самая ранняя экспрессия vvl-GFP обнаруживается внутри Spalt-положительного выхода на поверхность, который ограничен брюшными сегментами (Рис. 4C). Однако, в отличие от Drosophila, экспрессия Tribolium Spalt быстро теряется в этих клетках, при этом белок не обнаруживается в GFP-положительных клетках к 34 часам развития.Другим полезным маркером, относящимся к спецификации оеноцитов Drosophila, является белок гомеодомена Engrailed (En), который специфически маркирует клетки заднего компартмента в каждом сегменте (Kornberg, 1981). Как показано на фиг. 5A, оеноциты, которые помечены линией энхансеров Seven-up-lacZ (Svp-lacZ) и антителом Spalt, возникают группами из трех-четырех клеток внутри или в очень непосредственной близости от него. En-позитивный задний отсек (Gebelein and Mann, 2007). Рекрутирование эноцитов в Tribolium происходит по аналогичной траектории.На 26 часах развития самые ранние рекрутированные эноциты, как это отмечено vvl-GFP и Spalt out-cropping, содержатся в En-позитивных клетках заднего компартмента брюшных сегментов (Рис. 5C). По мере созревания оеноцитов Tribolium и начала экспрессии HNF4 и совместной метки со стрептавидином, однако, они теряют экспрессию En, перемещаются к переднему отделу и располагаются за пределами заднего компартмента (Рис. 5D). Эноциты дрозофилы аналогичным образом подавляют …
Контекст 2
… Затем проанализировали, какие абдоминальные сегменты эмбрионов Drosophila и Tribolium продуцируют эноциты. У эмбрионов D melanogaster эноциты формируются в первых семи абдоминальных сегментах, которые экспрессируют Abd-A Hox фактор (Brodu et al., 2002). Abd-A наиболее высоко экспрессируется в En-позитивных клетках заднего компартмента, которые продуцируют эноциты у эмбрионов дрозофилы на стадии 11 (Рис. 6A) (Gebelein and Mann, 2007). Однако после своего созревания оеноциты Drosophila экспрессируют относительно низкие уровни Abd-A по сравнению с соседними клетками брюшной полости (Рис. 6B).Предыдущий анализ экспрессии эмбрионов Tribolium показал, что сильная экспрессия abd-A обнаруживается в задних отделах первых восьми брюшных сегментов (Shippy et al., 1998). В этом исследовании мы продемонстрировали, что все восемь сегментов Tribolium продуцируют эноциты и что эти клетки возникают из заднего отдела. К сожалению, наши специфические антитела Drosophila Abd-A не смогли распознать Tribolium Abd-A. Однако мы смогли определить, экспрессируют ли оеноциты Tribolium белки Abd-A или Utx (гомолог Ultrabithorax в Tribolium), используя мышиное антитело UbdA (mAb FP6.87), который перекрестно реагирует с обоими в эмбрионах Tribolium. Как показано на фиг. 6, мы обнаружили, что антитело UbdA выявляет абдоминальные Hox-факторы во время ранней спецификации эноцитов (фиг. 6C) и что, как и оеноциты Drosophila, оеноциты Tribolium экспрессируют относительно низкие уровни абдоминальных Hox-факторов при созревании (фиг. 6D). Предыдущие публикации показали, что эмбрионы Drosophila melanogaster имеют скопления оеноцитов в брюшной полости, в среднем приблизительно шесть клеток в каждом полусегменте (Brodu et al., 2002, 2004; Gould et al., 2001). Используя HNF4 в качестве маркера, мы обнаружили схожее количество эноцитов у эмбрионов меланогастера стадии 16 D (5,79 ± 0,92 на кластер), а также определили, что количество эноцитов существенно не различается между брюшными сегментами вдоль передне-задней оси ( Рис 7A). Напротив, визуализация личиночных оеноцитов в Tribolium выявляет гораздо более крупные скопления клеток в сегментах брюшной полости. Чтобы оценить количество эноцитов, которые образуются в кластере в Tribolium, мы выбрали для анализа эмбрионов, меченных HNF4 и vvl-GFP, в момент времени (46 часов), в котором легко обнаруживается сильное ядерное окрашивание HNF (см. Рис. 4).Число эноцитов было сведено в таблицу для каждого сегмента брюшной полости девяти эмбрионов соответствующего возраста, что показало общее среднее значение 22,3 ± 0,6. Однако количество эноцитов, которые формируются между брюшными сегментами, варьировалось в зависимости от положения вдоль передне-задней оси в этот момент времени развития. Как показано на фиг. 7B, самый передний сегмент A1 в среднем содержал 28,3 ± 1,0 оеноцитов, тогда как самый задний сегмент A8 в среднем составлял 14,7 ± 1,3 клетки на кластер. К сожалению, нам не удалось определить количество эноцитов в более старых эмбрионах, потому что эноциты из разных сегментов находятся в тесном контакте друг с другом (см. Временные точки от 58 до 65 часов на Рис. 3 и Дополнение Рис. 2), а окрашивание ядер HNF4 использовалось для подсчета этих клеток. исчезает в процессе развития.Однако эта разница в количестве эноцитов вдоль передне-задней оси, вероятно, является временным событием, поскольку размер vvl-GFP-положительных кластеров у более старых эмбрионов Tribolium и ранних личинок сходен во всех сегментах брюшной полости (см. Suppl Fig 2). Таким образом, в отличие от эмбриона Drosophila с длинной зародышевой полосой, который одновременно продуцирует одинаковое количество оеноцитов во всех сегментах брюшной полости, эмбрион с короткой зародышевой полосой Tribolium продуцирует оеноциты с градиентом от высокого к низкому вдоль передне-задней оси…
характеристики, отзывы. Металлоискатель своими руками
В наши дни многие люди стремятся найти клады, а иногда и простой металлолом. Для кого-то это занятие стало интересным времяпрепровождением, а для кого-то — средством заработка.
Первый образец промышленного металлоискателя был создан в 1960-х годах и нашел широкое применение в горнодобывающей промышленности и других специальных работах.
Приборы используются для разминирования, для поиска оружия, в исследованиях геофизиков и археологов, в поисках сокровищ, а также для обнаружения инородных тел из металла в продуктах питания.В строительной отрасли они участвуют в обнаружении арматуры в бетонных блоках и трубопроводов в стенах. Металлоискатели также начали использовать шахтеры и старатели. А усовершенствование устройства позволило не прибегать к раскопкам при поиске золота.
В последние десятилетия этим устройством интересовалось множество людей. Поиск сокровищ и металлолома стал популярным хобби. Некоторые, например, отправляются с таким устройством на пляж в надежде найти ценную вещь.
Кто изобрел металлоискатель
На вопрос, какое устройство было первым, дать однозначный ответ сложно, потому что примерно в одно и то же время многие изобретатели в разных частях мира проводили собственные разработки названного устройства.
Но если говорить об одном конкретном человеке, которого можно считать родоначальником устройства, то это, несомненно, английский геолог и горный инженер Фокс. Он обнаружил свойство пропускать электричество через металлические руды и предметы.Примерно в 1830 году он разработал первый унифицированный локатор, который включал в себя батарею, несколько металлических стержней и проводов подходящей длины.
Первые методы поиска металла
Первый метод поиска заключался в следующем: один металлический стержень лежал в земле там, где должна была находиться руда. Он был подключен к одной клемме аккумулятора. Другой вывод был подключен к плавающему проводу. Металлические стержни втыкались в землю в разных точках и постоянно касались проводов.При обнаружении металлического предмета появились искры.
В 1870 году в устройстве использовались два отдельных стержня. Провод, подключенный через аккумулятор, упал на землю. При контакте с металлом прозвенел предупредительный звонок.
Аппарат «Пират»
А теперь посмотрим на современные инструменты. Некоторые из них — «Пират» — металлоискатель, работающий по проводимости электричества, индуктивным и магнитным свойствам металла. Кстати, свое интересное название устройство получило от изобретателей: ПИ — импульсный принцип работы, RAT — сокращенно от Radio Scot (сайт изобретателей).
Металлоискатель «Пират», фото которого представлены в этой статье, имеет унифицированный дизайн. Он включает в себя генератор, вырабатывающий переменный ток, который проходит через катушку с магнитным полем. Если металл, проводящий ток, находится слишком близко к катушке, то вихревые потоки будут направлены к металлу. Это способствует образованию переменного магнитного поля в металле. Обнаружение последнего позволяет использовать другую катушку для измерения магнитного поля.
Преимущества адаптации
«Пират» (металлоискатель) имеет простую конструкцию и унифицированную конфигурацию, не содержит программируемых элементов, которых так опасаются многие радиолюбители. Устройство отлично подойдет новичкам. И помните, что он не умеет различать металлы.
Металлоискатель «Пират», печатная плата которого представлена микросхемой NE555 (отечественный аналог КР1006ВИ1), не содержит дорогих или труднодоступных деталей. По техническим параметрам он не уступает зарубежным аналогам, цена которых достигает 300 у.е.е.
И главные преимущества этого устройства перед другими — стабильность работы и реакция на металл с большого расстояния.
Унифицированный «Пират» (металлоискатель для начинающих) имеет определенные технические характеристики. Его мощность 9-12 вольт, а уровень энергопотребления 3-40 мА. Устройство распознает предметы размером до 150 см.
Конструкция
Передающий и приемный узлы являются основными, металлоискатель «Пират». Печатная плата, представляющая собой модель NE555, и переключатель высокой мощности на транзисторе IRF740 входят в передающий узел.А приемный узел собран на базе микросхемы К157УД2 и транзистора BC547.
Намотка катушки осуществляется на оправке диаметром 190 мм и содержит 25 витков провода ПЭВ 0,5.
Биполярный транзистор NPN заменил модель Т2 и имеет напряжение не менее 200 вольт. Его можно взять от экономичной лампы или устройства для зарядки мобильного телефона. В крайнем случае Т2 можно заменить на КТ817.
В качестве Т3 можно использовать любую схему транзистора NPN.
Правильно собранный прибор не требует дополнительной настройки. Возможно, придется прибегнуть к применению резистора R12, чтобы щелчки при движении появлялись в среднем положении R13.
С помощью осциллографа можно контролировать длительность управляющего импульса на затворе Т2 и уровень частоты генератора. Оптимальная длительность импульса 130-150 мкс, частота 120-150 Гц.
Как работать с прибором
После включения прибора «Пират» (металлоискателя) следует подождать 15 или 20 секунд, после чего ручка чувствительности устанавливается в положение, при котором при движении слышны щелчки.Это послужит индикатором максимальной чувствительности.
Устройство имеет единую систему управления, поэтому приобрести навыки работы с ним не так уж и сложно.
Металлоискатель «Пират» своими руками
Многие задаются вопросом: как самому сделать металлоискатель «Пират»? Сборка такой сборки возможна людям с элементарными знаниями в области электроники.
Импульсный металлоискатель «Пират» имеет нестандартный и копируемый дизайн. Устройство содержит ряд компонентов и простую в использовании поисковую катушку.Если его диаметр составляет 280 мм, то он может обнаруживать объекты размером от 20 до 150 см.
Изготовление металлоискателя «Пиратские» руки — задача несложная, что является огромным преимуществом этого прибора. Компоненты сборки доступны и их легко найти. Они очень дешевые. Купить их можно в магазине радиодеталей или на рынке.
Перечень необходимых деталей для изготовления
Попробуем собрать металлоискатель «Пират» своими руками. Подробная инструкция поможет сделать это без ошибок даже неопытным радиолюбителям.
Устройство имеет две схематические модификации. В первом случае используется микросхема NE555 (отечественный аналог микросхемы КР1006ВИ1) — таймер. Но если у вас не получилось получить этот компонент, авторы также предусмотрели другой вариант схемы, основанный на транзисторах.
Тем не менее, рекомендуется собирать устройство по первой схеме, так как она имеет большую стабильность при работе.
При сборке на основе транзисторов выбирайте правильную частоту и длительность, так как они имеют довольно большой разброс технических характеристик.Для этого воспользуйтесь осциллографом.
Печатная плата прибора
Самодельный металлоискатель «Пират» имеет несколько вариантов разводки печатной платы, но чаще всего используют плату серии «Сприн Лайот».
После пайки к нему подключается питание. Для этого подайте любой источник питания с напряжением 9-12 Вольт. Можно прибегнуть к использованию батареек «Крона» (3 или 4 штуки) или батарейки. Использование одной «короны» не рекомендуется, так как это вызовет быстрое падение напряжения, что, в свою очередь, приведет к постоянному зависанию устройства.
Изготовление катушки для металлоискателя «Пират»
Как и другие модели импульсных устройств для поиска металла, прибор нетребователен к точности при изготовлении катушки. Вполне допустимо использовать ту, которая наматывается на оправку диаметром 190-200 мм — 25 витков. В этом случае используется обмоточный эмалированный провод сечением 0,5 мм.
Обмотки катушки обматываются изоляционной лентой или лентой. Кстати, чтобы увеличить глубину поиска устройства, можно прибегнуть к намотке названной детали, диаметром 260-270 мм, 21-22 витка тем же проводом.
Катушка прибора закреплена в твердом корпусе, который должен быть изготовлен, например, из пластика. Это необходимо для защиты устройства от ударов о землю или траву во время работы агрегата. Такой чехол можно приобрести в интернет-магазинах. Вообще, при изготовлении поисковых катушек не рекомендуется использование металлических деталей.
Выводы указанной детали припаяны к многожильному проводу сечением 0,5 — 0,75 мм. В идеале это две самоплетенные проволоки.Ваше устройство готово!
Обзоры
Имеющиеся на металлоискателе «Пират» отзывы свидетельствуют о том, что он стал очень популярным среди пользователей. По их словам, устройство обладает высокой степенью функциональности, а вещи из металла находят в короткие сроки и без ошибок. Легко наносится и не ощущается в руке.
Готовый прибор собран из нескольких частей, которые легко собираются. Нижняя часть — это его основание. Системная работа с устройством очень наглядная.Установить металлоискатель «Пират» не составит труда.
Развитие эноцитов у красного мучного жука Tribolium castaneum
Abstract
Эноциты — это особый тип клеток, необходимый для процессинга липидов, секреции феромонов и передачи сигналов в процессе развития. Их развитие хорошо охарактеризовано у Drosophila melanogaster , но остается неизвестным, сохраняется ли программа развития у других видов насекомых. В этом исследовании мы сравниваем спецификацию и развитие личиночных оеноцитов у Drosophila и красного мучного жука Tribolium castaneum .Во-первых, мы идентифицируем несколько полезных реагентов для мечения личиночных оеноцитов, включая линию улавливания энхансера Tribolium GFP и простой метод окрашивания конъюгированным с флуофором стрептавидином, который распознает эноциты у разных видов насекомых. Во-вторых, мы используем эти инструменты для описания развития оеноцитов в эмбрионах Tribolium , и наши результаты подтверждают консервативную роль передачи сигналов MAP-киназы, а также Spalt, Engrailed, HNF4 и вентральные вены, лишенные факторов в производстве специфичных для брюшной полости оеноцитов. клетки.Однако эмбрионы Tribolium продуцируют в четыре раза больше эноцитов на брюшной сегмент, чем Drosophila , и в отличие от Drosophila , эти клетки быстро подавляют экспрессию фактора транскрипции Spalt. Таким образом, эти результаты обеспечивают новое понимание молекулярных путей, регулирующих спецификацию эноцитов у разных видов насекомых.
Введение
Эноциты — это особый тип клеток, ответственный за множество важных метаболических и поведенческих ролей у насекомых (Chapman, 1998; Snodgrass, 1993).Функция эноцитов, пожалуй, лучше всего изучена у Drosophila melanogaster , который имеет отдельные популяции личинок и взрослых оеноцитов (Gould et al., 2001). Личиночные эноциты изначально формируются в эмбрионе и после созревания экспрессируют большое количество различных ферментов, необходимых для метаболизма липидов (Gutierrez et al., 2007). Фактически, избирательное удаление оеноцитов личинки приводит к неспособности личинки мобилизовать и перерабатывать липиды из жирового тела, что в конечном итоге приводит к серьезным дефектам роста и летальному исходу до окукливания (Gutierrez et al., 2007). Взрослые эноциты развиваются независимо от имагинальных тканей во время окукливания, и недавние работы показали, что они модулируют ухаживание и агрессивное поведение взрослых мух посредством выработки кутикулярных углеводородов, которые служат феромонами (Billeter et al., 2009; Fernandez et al., 2010 ). Следовательно, оеноциты личинок были уподоблены гепатоцитам позвоночных из-за их критической роли в гомеостазе липидов, тогда как взрослые оеноциты участвуют в коммуникации животных посредством продукции феромонов.В этой статье мы сосредоточены на сравнении и противопоставлении развития личиночных эноцитов у двух разных видов насекомых.
Личиночные оеноциты в Drosophila melanogaster представляют собой специфический для брюшной полости тип клеток, который формируется в кластеры от трех до девяти клеток в каждом полусегменте первых семи сегментов брюшка (Gould et al., 2001). Элегантные исследования показали, что генетическое взаимодействие между Hox-фактором Abd-A (Abd-A), факторами транскрипции Spalt (Spalt-major (Salm) и Spalt-related (Salr)) и передачей сигналов рецептора эпидермального фактора роста (EGFR) пути необходимы для спецификации эноцитов (Brodu et al., 2002, 2004; Эльстоб и др., 2001; Рустен и др., 2001). В этой модели Abd-A индуцирует секрецию лиганда EGF Spitz из специфической клетки-предшественника сенсорного органа брюшной полости, которая находится внутри спинной эктодермы, экспрессирующей Spalt. Соседние клетки, которые получают сигнал EGF, дополнительно активируют экспрессию Spalt, расслаиваются в эмбрионе и передаются на судьбу личиночных клеток оеноцитов. Поскольку в этом контексте EGF ведет себя как морфоген ближнего действия, одновременно указываются только три-четыре клетки, контактирующие с сенсорным предшественником (Brodu et al., 2004; Шило, 2005). Однако, как только первые клетки расслаиваются на эмбрион, от трех до четырех новых клеток вступают в контакт с сенсорным предшественником, получают сигнал EGF и повторяют еще один цикл спецификации эноцитов. Поскольку сенсорный предшественник секретирует EGF в течение ограниченного периода времени во время эмбриогенеза, обычно происходит только два раунда рекрутирования оеноцитов, дающих в среднем шесть оеноцитов на кластер (Brodu et al., 2004; Witt et al., 2010). После спецификации оеноциты созревают и впоследствии активируют экспрессию фактора транскрипции ядерного фактора гепатоцитов-4 (HNF4), а также многочисленных липид-модифицирующих ферментов, необходимых для процессинга липидов во время роста личинок (Gutzwiller et al., 2010; Паланкер и др., 2009).
Эксперименты, сфокусированные на продукции личинок дрозофилы , предоставили множество генетических и молекулярных маркеров, специфичных для оеноцитов. Однако до сих пор использование этих реагентов ограничивалось исследованием D. melanogaster . Напротив, эноциты у других видов были описаны только на стадиях личинки / нимфы с использованием классических гистологических и анатомических подходов, где они идентифицируются по их большому размеру, политеническим ядрам и свойствам окрашивания (Jackson and Locke, 1989; Snodgrass, 1993).Даже без использования молекулярных маркеров эти исследования установили степень разнообразия в расположении и количестве эноцитов в разных кладах насекомых. В то время как большинство видов склонны группировать свои оеноциты в небольшие группы по 6–10, Истхэм описал паразитических ос с> 20 оеноцитов на кластер (Eastham, 1929; Gould et al., 2001). Более того, распределение эноцитов может сильно различаться. У большинства клад есть метамерные скопления непосредственно под покровом брюшных сегментов. Однако некоторые также рассредоточивают свои эноциты среди тканей жирового тела, в то время как другие, такие как жесткокрылые (жуки), организуют их непрерывной полосой по всей длине брюшка (Chapman, 1998; Wheeler, 1892).Есть даже описания кузнечиков и муравьев с эноцитами, распространившимися на грудные сегменты на поздних стадиях их соответствующих личинок / нимф (Wheeler, 1892; Zara and Caetano, 2004). Таким образом, различия в конечном местоположении и количестве оеноцитов личинок у разных видов насекомых представляют собой потенциально полезную модель для проведения сравнительных исследований биологии эволюционного развития.
Учитывая разнообразие насекомых и важную роль личиночных эноцитов в метаболических функциях, мы хотели изучить развитие эноцитов у отдаленно родственных видов насекомых.Для сравнительных исследований с Drosophila melanogaster мы выбрали красного мучного жука Tribolium castaneum. T castaneum — это новый модельный организм, имеющий секвенированный геном, и в отличие от Drosophila , Tribolium представляет собой насекомое с короткими зародышевыми полосками, которое последовательно добавляет сегменты во время эмбрионального развития (Richards et al., 2008; Roth and Hartenstein, 2008 ). Здесь мы идентифицируем несколько молекулярных маркеров, которые распознают личиночные оеноциты в Tribolium , в том числе линию ловушек-энхансеров, специфичных для oenoctye, около вентральных вен , лишенную гомолога ( vvl ), который также экспрессируется в оеноцитах Drosophila (Inbal и другие., 2003). Кроме того, мы определили, что конъюгированный с флуорофором стрептавидин действует как новый биомаркер, способный обнаруживать личиночные эноциты у нескольких видов насекомых. Наконец, мы используем эти биомаркеры для выявления как сходства, так и различий между эмбриональным развитием личиночных оеноцитов в пределах Tribolium по сравнению с таковыми в Drosophila . Наши находки показывают, что, подобно Drosophila , Tribolium оеноциты личинок являются абдоминально-специфическими и первоначально рекрутируются из Spalt-положительной эктодермы, которая впоследствии активирует экспрессию фактора HNF4.В отличие от Drosophila , однако, оеноциты Tribolium быстро подавляют экспрессию Spalt, рекрутируются в гораздо большем количестве (более 20 оеноцитов на полусегмент) и образуют непрерывную полосу оеноцитов между брюшными сегментами. Обсуждаются значение методов окрашивания эноцитов насекомых и результаты межвидового исследования.
Материалы и методы
Насекомые
Жуков выращивали в пластиковых бутылках при 30 ° C в инкубаторе Digitherm 47L (Tritech Research, DT2-MP-38), увлажняемом через небольшой лоток с водой.Жуки KT817 ( vvl-GFP ) были приобретены у Сью Браун в складе запасов Tribolium Университета штата Канзас (http://www.geku-base.uni-goettingen.de/) (Trauner et al., 2009 ). Сайт встраивания элемента piggybac-XP3-GFP определяли с помощью обратной ПЦР (iPCR) на кольцевых геномных фрагментах, полученных в результате расщепления Hha1. Продукт ПЦР секвенировали с использованием праймеров PRF1 5’-ACCGATAAAAACACATGCGTCAAT-3 ’и PRR1 5’-ATGCATTTGCCTTTCGCCTTAT-3’. Сайт интеграции и ориентация были подтверждены генотипированием ПЦР с использованием локус-специфичных праймеров (5′-AAGTCAGCCACCAATCATCG-3 ‘, 5′-GCTCAATTCGGGTGCTTGTT-3′) в паре со специфическими для piggybac праймерами (PRF1 и PLR1 5’-ACAGCGACGGATTCGCG соответственно) .Все стоки Drosophila выращивали при 25 ° C, и ранее были описаны трансгенные линии Rho654-lacZ (Li-Kroeger et al., 2008) и svp-lacZ (Elstob et al., 2001).
Продукция антител
Бактериальный экспрессионный вектор Tribolium castaneum HNF4 получали с использованием ПЦР для амплификации и клонирования кДНК HNF4, кодирующей аминокислоты 162-494 в рамке считывания с N-концевой меткой 6X-His (pET14b, Novagen). Плазмиду экспрессии трансформировали в бактерии BL21-CodonPlus (DE3) -RP (Stratagene), и экспрессию белка индуцировали с использованием 0.25 мМ IPTG в течение 2 часов при 37 ° C. Бактерии лизировали в 8М буфере для лизиса мочевины и очищали с помощью Ni-аффинной хроматографии, как описано ранее (Gutzwiller et al., 2010). Очищенный белок использовали для генерации антител к HNF4 у крыс с использованием стандартных протоколов (Cocalico Biologicals)
Фиксация эмбрионов и окрашивание антителами
Эмбрионы Tribolium отделяли от трехпрофильтрованной среды путем просеивания через сито # 50. Собранные эмбрионы подвергали дехорионированию в 50% растворе отбеливателя в течение 8 минут, промывали в H 2 O и фиксировали встряхиванием в 8% растворе формальдегид / гептан в течение 20 минут.Эмбрионы экстрагировали в метаноле и растирали через иглу 20 г с удалением голых эмбрионов через каждые ~ 10 проходов. Drosophila эмбрионов были собраны стандартными методами. Личинки Drosophila и яйца кабачка получали аналогично эмбрионам Tribolium , но вместо этого обрабатывали 100% отбеливателем и фиксировали встряхиванием в течение 30 мин.
Для окрашивания антител образцы инкубировали в течение ночи в коктейле первичных антител в PBX (PBS плюс 0.2% Triton-X) при 4 ° C, выдерживали 3 раза по 20 минут в PBX, инкубировали в течение 2 часов при комнатной температуре с соответствующими вторичными антителами, конъюгированными с флуорофором, промывали 4 раза по 20 минут в PBX и устанавливали для микроскопического анализа. Изображения получали либо на флуоресцентном микроскопе Zeiss, оборудованном апотомным фильтром, либо на конфокальном микроскопе Leica. Были использованы следующие первичные антитела: Cut (мышь, 1:20, DSHB, # 2B10), фосфогистон 3 (мышь, 1: 1000, Millipore, # 05-806), Dm HNF4 (крыса, 1: 2000, ( Gutzwiller et al., 2010)), Tc HNF4 (крыса, 1: 2000), Dm Spalt (кролик, 1: 1500, (Xie et al., 2007)), фосфо-ERK (дифосфорилированный MAPK; Mouse, 1:50, Sigma , M-8159) Engrailed (мышь, 1:20, DSHB, # 4D9), бета-галактозидаза (курица, 1: 1000, Abcam) Dm Abd-A (морская свинка, 1: 500, (Li-Kroeger et al. , 2008)) и UbdA (Mouse, 1: 5, DSHB FP6.87). Для обнаружения стрептавидина в оеноцитах образцы инкубировали в течение ночи с Cy5-конъюгированным стрептавидином (1: 1000) и промывали 4 раза по 20 мин, по крайней мере, с одной дополнительной промывкой в течение ночи в PBX.Подсчет эноцитов проводили на 10 совпадающих по возрасту эмбрионах Drosophila с использованием Dm анти-HNF4 и 9 соответствующих по возрасту эмбрионах Tribolium с использованием комбинации vvl-GFP и Tc анти-HNF4. Все результаты были проанализированы ANOVA с использованием Excel.
Результаты
Для проведения сравнительного исследования образования личиночных оеноцитов у разных видов насекомых мы сначала оценили способность двух антител, которые маркируют оеноциты Drosophila , перекрестно реагировать с консервативными белками в эмбрионах Tribolium .У эмбрионов Drosophila белки Spalt-major (Salm) и ядерного фактора гепатоцитов 4 (HNF4) коэкспрессируются в абдоминальных эноцитах на стадии 16 эмбрионов Drosophila (). Поскольку Tribolium содержит один гомолог генов HNF4 и Spalt и антигены Drosophila Salm и HNF4, используемые для создания каждого антитела, включающего высококонсервативные белковые домены (не показаны), мы протестировали эти реагенты на эмбрионах Tribolium (Gutzwiller et al. al., 2010; Xie et al., 2007). Как показано в, мы обнаружили, что антитело, индуцированное против белка Drosophila Spalt, но не против HNF4 (не показано), обнаруживает отчетливый паттерн экспрессии в эмбрионах Tribolium . В соответствии с паттерном, наблюдаемым у ранних эмбрионов Drosophila , антитело Spalt маркирует широкую полосу дорсальной эктодермы, которая включает область, из которой возникают эноциты в эмбрионах Tribolium (и см. Сравнение ранних эмбрионов D. melanogaster с эмбрионами начало т.castaneum ) (Li-Kroeger et al., 2008; Rusten et al., 2001). Кроме того, мы обнаружили, что это антитело Spalt демонстрирует паттерн экспрессии, идентичный опубликованному паттерну Spalt in situ в заднем крыле Tribolium (не показано и (Tomoyasu et al., 2005)). Однако, в отличие от Drosophila , не наблюдали отчетливых Spalt-положительных скоплений брюшных клеток у зрелых эмбрионов, что позволяет предположить, что этот фактор может не поддерживаться в личиночных оеноцитах Tribolium .
Молекулярные маркеры оеноцитов личинок в пределах видов насекомыхA-A » ‘) Вид сбоку эмбриона стадии 16 Drosophila melanogaster (Dm), иммуноокрашенного на Spalt (пурпурный) и HNF4 (зеленый), обнаруживает кластеры эноциты брюшной полости. Увеличенное изображение третьего грудного сегмента (T3) и первых двух брюшных сегментов (A1, A2 не обозначены) подчеркивает специфическое для брюшной полости образование этого типа клеток (A’-A ’’ ’). Обратите внимание, дополнительное близлежащее окрашивание Spalt выявляет клетки сенсорного органа, который является частью периферической нервной системы.
B-B » ‘) Вид сбоку 46h зародыша Tribolium castaneum (Tc), иммуномеченного на HNF4 (зеленый) и Spalt (пурпурный), показывает, что эти два фактора не локализуются совместно в эноцитах эмбриона жука. .
C-C » ‘) Вид сбоку 46h Tribolium castaneum (Tc) эмбриона, иммуномеченного на HNF4 (зеленый) и стрептавидина, конъюгированного с флуорофором (SA-Cy5), выявляет скопления оеноцитов в брюшной полости, но не на груди сегменты. Обратите внимание, что дополнительное сильное окрашивание SA-Cy5 наблюдается в плевроподе (P) в первом сегменте брюшной полости.
D) Крупным планом вид сбоку трех брюшных сегментов третьего возраста Личинка Drosophila melanogaster , окрашенная стрептавидином, конъюгированным с флуорофором (SA-Cy5), выявляет скопления примерно из шести эноцитов в каждом сегменте.
E) Изображение в светлом поле, показывающее вид сбоку зрелого эмбриона Hemipteran Anasa tristis .
F-F ’) Вид сбоку зрелого эмбриона Hemipteran Anasa tristis , меченного флуорофор-конъюгированным стрептавидином (SA-Cy5), выявляет скопления эноцитов в брюшных сегментах.Подмножество нейронов (n) также маркируется стрептавидином (F ‘).
Оеноциты происходят из зарезанной полосы как у Drosophila , так и у TriboliumA) Вид сбоку трех брюшных сегментов со стадии 11 svp-lacZ Эмбрион Drosophila , иммуноокрашенный на Engrailed (En, красный), β-галактозидазу (β-gal, зеленый) и Spalt (синий). svp-lacZ (β-gal) маркирует вновь определенные оеноциты Drosophila (стрелки), которые лежат в заднем компартменте, отмеченном En.
B) Вид сбоку трех брюшных сегментов на стадии 14 svp-lacZ Эмбрион Drosophila , иммуноокрашенный на Engrailed (En, красный), β-галактозидазу (зеленый) и Spalt (синий). На этой стадии оеноцитов Drosophila все еще положительны по svp-lacZ (β-gal) и Spalt, но они имеют подавляющую регуляцию En и лежат впереди заднего компартмента (стрелка).
C) Вид сбоку трех брюшных сегментов эмбриона 26h Tribolium , иммуноокрашенного на En (красный), vvl-GFP (зеленый) и Spalt (синий).Обратите внимание, что ранние vvl-GFP положительные клетки (стрелка) лежат в домене Spalt (C ’’, звездочка) и перекрываются с экспрессией En в брюшных сегментах.
D) Вид сбоку трех брюшных сегментов эмбриона 46h Tribolium , иммуноокрашенного на En (красный), vvl-GFP (зеленый) и Spalt (синий). Обратите внимание, что GFP-положительные клетки оеноцитов лежат впереди En-положительных клеток заднего компартмента и не экспрессируют Spalt.
Поскольку антитело Drosophila HNF4 не распознало специфический паттерн в эмбрионах Tribolium , мы затем создали антисыворотку против белка HNF4 Tribolium .Как показано в, мы обнаружили паттерн экспрессии HNF4, который маркирует многообещающий набор кластеров клеток, которые ограничены брюшными сегментами у 46-часовой эмбрионов. Используя оптические срезы, эти клетки можно отличить от подлежащей мезодермы HNF4 + как по интенсивности окрашивания HNF4, так и по их латеральному положению. В соответствии с тем, что эти HNF4-положительные клетки являются личиночными оеноцитами, они также коэкспрессируют липид-модифицирующий фермент (увы, приложение, рис. 1), который экспрессируется в оеноцитах Drosophila , а также несколько других маркеров оеноцитов, описанных ниже.Более того, совместное окрашивание с антителом Spalt подтвердило, что в отличие от сильной повышающей регуляции, наблюдаемой в зрелых оеноцитах D. melanogaster , оеноциты в более старых эмбрионах T. castaneum лишены экспрессии Spalt (’’ ’).
Новый биомаркер личиночных оеноцитов
В процессе подтверждения того, что HNF4-экспрессирующие абдоминально-специфические клетки в эмбрионах Tribolium являются личиночными оеноцитами, мы случайно определили новый биомаркер для оеноцитов.При тестировании множества антител на перекрестную реактивность в Tribolium (полный список антител см. В дополнительной таблице 1) мы попытались усилить сигнал с помощью системы мечения биотин-стрептавидин. Вместо этого мы обнаружили, что всякий раз, когда использовался биотин-стрептавидин, устойчивый устойчивый сигнал специфически маркирует цитоплазму тех же кластеров клеток брюшной полости, которые экспрессируют HNF4, а также отросток плевропода 1 сегмента брюшной полости (). Последующее поэтапное удаление как первичных, так и вторичных антител показало, что добавление одной только молекулы стрептавидина, конъюгированной с флуорофором, приводит к этой специфической для брюшной полости паттерну.Сильное мечение личиночных оеноцитов стрептавидином, вероятно, связано с этим типом клеток, содержащим высокие уровни биотина, который служит важным коферментом для метаболизма жирных кислот (Fletcher and Myant, 1960). Таким образом, флуоресцентный конъюгированный стрептавидин является полезным красителем для распознавания личиночных оеноцитов в эмбрионах Tribolium .
Затем мы определили общую применимость стрептавидина в качестве биомаркера эноцитов, протестировав окраску на двух других насекомых. В то время как стрептавидин не распознавал личиночные эноциты во время эмбриогенеза D melanogaster , это окрашивание было очень эффективным для мечения эноцитов в личинке третьего возраста ().Более того, способность стрептавидина распознавать этот тип клеток имеет дополнительное преимущество, заключающееся в том, что не требуется рассечение личинок (см. Методы). Чтобы дополнительно проверить применимость окраски стрептавидином к насекомым, мы собрали эмбрионы полуокрылых из рода Anasa (часто называемые жуками-кабачками) из тыквенной тыквы на юго-западе Огайо. Яичная скорлупа вручную отделялась от каждого эмбриона на поздней стадии (), и нанесение флуоресцентно-конъюгированного стрептавидина выявило сильное окрашивание областей живота, соответствующее скоплениям эноцитов (стрелки, ‘).Однако следует отметить, что окрашивание стрептавидином в Anasa , а также в Drosophila и Tribolium также пометило подмножество других клеток, включая нейроны. Следовательно, по возможности следует использовать дополнительные маркеры для определения клеток, меченных стрептавидином, как эноцитов. Тем не менее, эти данные предполагают, что стрептавидин, вероятно, будет широко применимым маркером личиночных оеноцитов у многих видов насекомых.
Линия улавливания энхансеров для отслеживания развития оеноцитов в
TriboliumМощным инструментом для мечения конкретных типов клеток в модельных организмах является использование линий ловушек энхансеров.Недавняя работа нескольких исследовательских групп сделала множество таких ловушек энхансеров GFP в Tribolium castaneum доступными через базу данных GEKU (http://www.geku-base.uni-goettingen.de/) (Trauner et al., 2009 ). Сканирование доступных линий ловушек энхансеров выявило несколько с интересными паттернами экспрессии, специфичными для брюшной полости. Одна из этих линий, KT817, была выбрана для анализа, и было обнаружено, что она экспрессирует GFP по той же схеме, что и клетки, меченные HNF4 () и стрептавидином (). (Обратите внимание, что дополнительное слабое окрашивание GFP в каждом сегменте связано с не-сайт-специфической активностью конструкции GFP).Используя протокол iPCR (см. Методы), мы определили, что piggybac-GFP находится на расстоянии ~ 60 килобайт от гомолога Tribolium вентральных вен без гена ( vvl ), который, как известно, экспрессируется в ранних личиночных оеноцитах в D melanogaster (Inbal et al., 2003). Таким образом, мы полагаем, что специфический для эноцитов паттерн экспрессии, скорее всего, обусловлен энхансером Tribolium vvl , несмотря на присутствие нескольких других предсказанных генных моделей в локусе.Этот вывод становится более правдоподобным из-за относительно большого расстояния между генами в геноме Tribolium , а также из предыдущих находок, описывающих энхансеры более чем на 100 килобайт от их генов-мишеней (Levine, 2010). Следовательно, ловушка энхансера KT817 в дальнейшем обозначается как vvl-GFP .
Идентификация ловушки-энхансера, специфичной для эноцитов vvl-GFPA) Снимок экрана обзора GB, показывающий сайт интеграции (красная стрелка) вектора piggybac-GFP ~ 60 т.п.н. ниже по потоку от Tribolium без вентральной вены ( vvl ) гомолог.
до н.э.) Вид сбоку 46h эмбриона vvl-GFP Tribolium , иммуномеченного на GFP (зеленый) и либо HNF4 (B, пурпурный), либо совместно окрашенного стрептавидином, конъюгированным с Cy5 (SA-Cy5, пурпурный), показывает брюшную полость -специфические кластеры GFP-положительных клеток, которые метятся совместно с HNF4 и SA-Cy5. Сегменты T3 и A1 помечены.
Затем мы использовали vvl-GFP и антитело HNF4 в качестве маркеров для изучения развития личиночных оеноцитов Tribolium во время эмбриогенеза (и Дополнение на рис. 2).Антитело против фактора транскрипции Cut также использовалось в качестве маркера близлежащих ямок трахеи (). Для этого анализа мы выполнили синхронизированные коллекции эмбрионов и определили несколько временных точек и ориентиров развития, которые коррелируют с морфологическими переходами для эноцитов. Во-первых, начальные следы экспрессии vvl-GFP можно увидеть вскоре после завершения расширения зародышевой полосы (26 часов), и это окрашивание быстро усиливается, выявляя кластер примерно из 15 клеток в каждом сегменте брюшной полости (через 28 часов).В это время GFP-положительные клетки лишены экспрессии HNF4, а сигнал GFP имеет небольшую, но стабильно более высокую интенсивность в передних (A1-A3) по сравнению с задними (A4-A8) сегментами брюшной полости (через 28 часов). Это открытие согласуется с тем, что Tribolium представляет собой насекомое с короткими зародышевыми полосами, которое добавляет новые (более молодые) задние сегменты брюшка во время развития (Schroder et al., 2008). К 34 часам каждый из сегментов брюшной полости с A1 по A8 содержит клеток, экспрессирующих vvl-GFP , которые совместно экспрессируют HNF4 (), и эти клетки образуют кластер клеток в форме полумесяца вокруг Cut-положительной трахеальной ямки.По завершении ретракции зародышевой ленты (46 ч) количество GFP- и HNF4-положительных оеноцитов увеличивается до более чем 20 клеток на кластер в большинстве сегментов брюшной полости, и эти клетки присутствуют в одном кластере дорсальнее трахеальной ямки в каждом из них. сегмент. К 50 часам суставы ног образуют четкие границы, и удлинение кишечной трубки можно увидеть у эмбрионов, наклоненных в сторону дорсальной проекции (не показано). На этом этапе скопления эноцитов начинают распространяться как кзади, так и внутрь от покровов, что приводит к формированию характерной формы акульих зубов.Кроме того, многие неоеноцитные клетки начинают экспрессировать высокие уровни HNF4, но эти клетки не распознаются стрептавидином и не экспрессируют vvl-GFP (через 50 часов, данные не показаны). К 58 часам кишечная трубка образует полную петлю, и дорсальное закрытие завершается. На этой стадии GFP-положительные клетки все еще сильно окрашиваются стрептавидином, но экспрессия HNF4 значительно снизилась. Более того, эноциты образуют непрерывную полосу, проходящую через сегментарные границы, и мигрировали дорсально, морфология соответствует классическому описанию личиночных оеноцитов жесткокрылых (Snodgrass, 1993; Wheeler, 1892).
Динамика развития морфогенеза оеноцитов в TriboliumЭмбрионы были сопоставлены по стадиям и окрашены по общей морфологии (DAPI в A), маркерам эноцитов ( vvl-GFP в B и D, HNF в C и D), и Cut-антитело, которое маркирует развивающуюся трахеальную ямку (E). Эноциты, маркированные vvl-GFP , сначала появляются в виде слабых скоплений в задней части брюшных сегментов на 28 часах развития. Экспрессия GFP становится более устойчивой к 34 часам, клетки начинают экспрессировать HNF4, и кластер оеноцитов тянется к переднему и дорсальному в каждом сегменте брюшной полости.На 46 час. Большинство эноцитов располагаются в кластере дорсальнее трахеальной ямки, при этом небольшое количество клеток остается только кзади, что приводит к образованию характерной формы запятой. К 50 часам метка GFP показывает, что оеноциты образуют кластер клеток в форме «акульего зуба» в каждом сегменте брюшной полости, и хотя оеноциты поддерживают экспрессию HNF4, многие дополнительные клетки также начинают экспрессировать высокие уровни этого белка. После дорсального закрытия (58 ч) эноциты образуют непрерывную полосу клеток, и экспрессия HNF4 начала угасать.Более того, эноциты мигрировали внутрь и дорсально относительно наружных отверстий трахеальных ямок.
Сравнение спецификации оеноцитов между
D. melanogaster и T. castaneumОеноциты Drosophila первоначально определяются в эмбрионах стадии 11, когда клетки дорсальной эктодермы, экспрессирующие Spalt, получают временный сигнал EGF. (Elstob et al., 2001). ; Rusten et al., 2001). Клетка-отправитель EGF, специфический предшественник сенсорного органа (SOP), может быть помечена с использованием трансгенной репортерной линии, взятой из ромбовидного локуса : Rho654-LacZ , а принимающие EGF клетки можно визуализировать с помощью фосфо-ERK-антитела. который распознает активированный сигнальный путь MAP-киназы () (Gabay et al., 1997; Ли-Крегер и др., 2008; Witt et al., 2010). Клетки, принимающие EGF, впоследствии активируют и поддерживают экспрессию Spalt во время созревания эноцитов. Напротив, мы обнаружили, что зрелые оеноциты Tribolium (отмеченные HNF4 и стрептавидином) не поддерживают экспрессию Spalt (). Однако возможно, что оеноцитов Tribolium первоначально рекрутируются из Spalt-экспрессирующих клеток дорсальной эктодермы и впоследствии подавляют этот фактор во время созревания. Чтобы проверить эту идею, мы тщательно проанализировали раннюю экспрессию Spalt, фоспо-ERK и vvl-GFP в эмбрионах жуков.Как показано на, небольшой вентральный выход Spalt-положительных клеток слабо, но постоянно окрашивается с использованием антитела фосфо-ERK. Хотя эти эмбрионы не экспрессируют трансген vvl-GFP на этой стадии развития, самая ранняя экспрессия vvl-GFP обнаруживается внутри Spalt-позитивного выхода на поверхность, который ограничен брюшными сегментами (). Однако в отличие от Drosophila , Tribolium Spalt экспрессия быстро теряется в этих клетках, при этом белок не обнаруживается в GFP-положительных клетках к 34 часам развития.
Передача сигналов Spalt и pERK присутствует в предшественниках оеноцитов как у Drosophila , так и у TriboliumA) Вид сбоку стадии 11 Rho654-LacZ Drosophila эмбриона Spa ), β-галактозидаза (зеленый в A) и фосфо-ERK (pERK, зеленый в A ‘и A’ ‘). Rho654-lacZ специфически маркирует сенсорную клетку-предшественник C1, которая секретирует лиганд EGF для активации пути MAP-киназы (p-ERK) в соседних предшественниках эноцитов, которые впоследствии активируют Spalt.Обратите внимание, активность pERK также отмечает развивающиеся трахеальные ямки (TP).
B) Вид сбоку 24-часового эмбриона Tribolium , иммуноокрашенного на Spalt (пурпурный в B и B ’’) и фосфо-ERK (pERK, зеленый в B ’и B’ ’). Обратите внимание на присутствие небольшого выхода экспрессии Spalt, которая перекрывается с активностью pERK в задней части каждого абдоминального сегмента (стрелка).
C) Вид сбоку эмбриона 26h Tribolium , иммуноокрашенного на Spalt (пурпурный в C и C ’’) и vvl-GFP (зеленый в C ’и C’ ’).Обратите внимание на небольшое выступление экспрессии Spalt теперь перекрывается с ранней экспрессией vvl-GFP в сегментах брюшной полости.
Другим полезным маркером, относящимся к спецификации оеноцитов Drosophila , является белок гомеодомена Engrailed (En), который специфически маркирует клетки заднего компартмента в каждом сегменте (Kornberg, 1981). Как показано на фиг.1, оеноциты, которые совместно помечены энхансерной линией-ловушкой Seven-up-lacZ ( Svp-lacZ ) и антителом Spalt, возникают группами из трех-четырех клеток внутри или в очень непосредственной близости. to, En-позитивный задний отсек (Gebelein and Mann, 2007).Рекрутирование эноцитов в Tribolium происходит по аналогичной траектории. На 26 часах развития самые ранние рекрутированные оеноциты, как отмечено vvl-GFP и Spalt out-cropping, содержатся в En-позитивных клетках заднего компартмента брюшных сегментов (). Однако по мере созревания оеноцитов Tribolium и начала экспрессии HNF4 и совместной метки со стрептавидином они теряют экспрессию En, перемещаются к переднему отделу и располагаются за пределами заднего компартмента (). Оеноциты Drosophila сходным образом подавляют экспрессию En у старых эмбрионов и находятся за пределами заднего компартмента (). В целом эти исследования предполагают, что оеноцитов Tribolium аналогичным образом возникают из клеток заднего компартмента, которые активируют сигнальный путь MAP-киназы и экспрессируют Spalt, но, в отличие от Drosophila , экспрессия Spalt быстро теряется в оеноцитах Tribolium , что указывает на то, что этот фактор не требуется. для их созревания.
Затем мы проанализировали, какие абдоминальные сегменты эмбрионов Drosophila и Tribolium продуцируют эноциты.У эмбрионов D melanogaster эноциты формируются в первых семи абдоминальных сегментах, которые экспрессируют фактор Abd-A Hox (Brodu et al., 2002). Abd-A наиболее высоко экспрессируется в En-позитивных клетках заднего компартмента, которые продуцируют эноциты на стадии 11 эмбрионов Drosophila () (Gebelein and Mann, 2007). Однако после своего созревания оеноцитов Drosophila экспрессируют относительно низкие уровни Abd-A по сравнению с соседними клетками брюшной полости (). Предыдущий анализ экспрессии эмбрионов Tribolium показал, что сильная экспрессия abd-A обнаруживается в задних отделах первых восьми брюшных сегментов (Shippy et al., 1998). В этом исследовании мы продемонстрировали, что все восемь из этих сегментов Tribolium продуцируют эноциты и что эти клетки возникают из заднего компартмента. К сожалению, наши специфические антитела Drosophila Abd-A не смогли распознать Tribolium Abd-A. Однако мы смогли определить, экспрессируют ли оеноциты Tribolium белки Abd-A или Utx (гомолог Ultrabithorax в Tribolium ), используя мышиное антитело UbdA (mAb FP6.87), который перекрестно реагирует с обоими в эмбрионах Tribolium . Как показано в, мы обнаружили, что антитело UbdA выявляет абдоминальные Hox-факторы во время ранней спецификации оеноцитов () и что, подобно оеноцитам Drosophila , оеноциты Tribolium экспрессируют относительно низкие уровни абдоминальных Hox-факторов при созревании ().
Абдоминальная экспрессия Hox в оеноцитах Drosophila и TriboliumA) Вид сбоку трех брюшных сегментов на стадии 11 Drosophila эмбрион, иммуноокрашенный на Abd-A (пурпурный) и Spal.Высокие уровни spalt маркируют недавно определенные эноциты Drosophila , коэкспрессирующие Abd-A.
B) Вид сбоку трех брюшных сегментов на стадии 16 Эмбрион Drosophila , иммуноокрашенный на Abd-A (пурпурный) и Spalt (зеленый). На этом этапе оеноцитов Drosophila все еще положительны на Spalt, но экспрессируют меньше Abd-A.
C) Вид сбоку трех брюшных сегментов 28-часового эмбриона Tribolium , иммуноокрашенного на UbdA (пурпурный) и vvl-GFP (зеленый).Обратите внимание, что первые vvl-GFP положительные клетки экспрессируют абдоминальный Hox-фактор, обнаруживаемый антителом UbdA.
D) Вид сбоку трех брюшных сегментов эмбриона 46h Tribolium , иммуноокрашенного на UbdA (пурпурный) и vvl-GFP (зеленый). Обратите внимание, что GFP-положительные клетки оеноцитов экспрессируют низкие уровни абдоминальных Hox-факторов.
Подсчет эноцитов в
TriboliumПредыдущие публикации показали, что эмбрионов Drosophila melanogaster имеют скопления эноцитов в брюшной полости, в среднем приблизительно шесть клеток в каждом полусегменте (Brodu et al., 2002, 2004; Gould et al., 2001). Используя HNF4 в качестве маркера, мы обнаружили схожее количество эноцитов в эмбрионах D melanogaster стадии 16 (5,79 ± 0,92 на кластер), а также определили, что количество эноцитов существенно не различается между сегментами брюшной полости вдоль передне-задней части. ось (). Напротив, визуализация личиночных оеноцитов в Tribolium выявляет гораздо более крупные скопления клеток в сегментах брюшной полости. Чтобы оценить количество эноцитов, которые образуются на кластер в Tribolium , мы выбрали для анализа эмбрионы, меченные HNF4 и vvl-GFP , в момент времени (46 часов), в котором легко обнаруживается сильное ядерное окрашивание HNF (см.).Число эноцитов было сведено в таблицу для каждого сегмента брюшной полости девяти эмбрионов соответствующего возраста, что показало общее среднее значение 22,3 ± 0,6. Однако количество эноцитов, которые формируются между брюшными сегментами, варьировалось в зависимости от положения вдоль передне-задней оси в этот момент времени развития. Как показано на фиг.4, самый передний сегмент А1 в среднем составлял 28,3 ± 1,0 оеноцитов, тогда как самый задний сегмент А8 в среднем составлял 14,7 ± 1,3 клетки на кластер. К сожалению, нам не удалось определить количество эноцитов в более старых эмбрионах, потому что эноциты из разных сегментов находятся в тесном контакте друг с другом (см. Временные точки от 58 до 65 часов на рис. 2 и дополн.), А окрашивание ядер HNF4, используемое для подсчета этих клеток, исчезает во время разработка.Однако это различие в количестве эноцитов вдоль передне-задней оси, вероятно, является временным событием, поскольку размер положительных кластеров vvl-GFP- у более старых эмбрионов Tribolium и ранних личинок одинаков во всех сегментах брюшной полости (см. Рис. ). Таким образом, в отличие от эмбриона с длинным зародышем Drosophila , который одновременно продуцирует одинаковое количество оеноцитов во всех абдоминальных сегментах, эмбрион с коротким зародышевым поясом Tribolium продуцирует оеноциты с градиентом от высокого к низкому вдоль передне-задней оси живот.
Подсчет эноцитов в TriboliumA-B) Сравнение количества эноцитов на кластер в каждом брюшном сегменте эмбрионов Drosophila (A) и Tribolium (B) График показывает среднее значение и стандартную ошибку для количества эноцитов, образующихся в каждом сегменте брюшной полости. Drosophila эмбрионов в среднем составляет около 6 оеноцитов на кластер, при этом статистических различий между сегментами брюшной полости от А1 до А7 не наблюдается.Напротив, у эмбрионов Tribolium в среднем более 20 эноцитов на кластер, а сегменты A1 (*) и A8 (**) статистически отличаются от всех других сегментов брюшной полости (p <0,05).
CD) Вид сбоку трех брюшных сегментов 26h (C) или 40h (D) эмбриона Tribolium , иммуноокрашенного на vvl-GFP (зеленый) и фосфогистон3 (ph4, пурпурный), показывают, что клетки экспрессирующие GFP не могут колокализоваться с маркером ph4 G2-M.
Более высокое количество эноцитов, продуцируемых в Tribolium , может быть связано либо с привлечением большего количества эноцитов из пула компетентных клеток, либо с последующим размножением эноцитов посредством деления клеток. Чтобы различить эти возможности, мы окрасили эмбрионов vvl-GFP митотическим маркером фосфо-гистон 3 (ph4) (Mahadevan et al., 1991). Как показано на фиг.4, ph4 мечен множеством клеток внутри эмбриона Tribolium . Однако мы не обнаружили совместного мечения GFP-положительных клеток с ph4 на любой стадии эмбриогенеза ().Эти находки согласуются с предыдущими исследованиями на Drosophila , которые показали, что эноциты являются постмитотическими после их первоначальной спецификации (Gould et al., 2001). Таким образом, увеличенное количество vvl-GFP -положительных оеноцитов в Tribolium не происходит за счет пролиферации после спецификации и, вероятно, связано с повышенным рекрутированием клеток этого типа.
Обсуждение
Личиночные оеноциты представляют собой тип клеток, обнаруживаемый у насекомых, и функциональные исследования на Drosophila melanogaster показали, что они необходимы для правильного метаболизма липидов и роста личинок (Chapman, 1998; Gutierrez et al., 2007). В этом исследовании мы сравниваем спецификацию и развитие личиночных эноцитов между D melanogaster и красным мучным жуком Tribolium castaneum . Наши результаты имеют два важных значения: 1) Мы определили общий инструмент (конъюгированный с флуорофором стрептавидин), который можно использовать для распознавания эноцитов у других видов насекомых, а также несколько полезных маркеров для изучения эноцитов в триболии Tribolium . 2) Мы применили эти инструменты для описания спецификации оеноцитов у эмбрионов Tribolium , и наши результаты показывают как сходство, так и различия в их развитии по сравнению с Drosophila .
Стрептавидин является полезным маркером судьбы эноцитов у насекомых
Из-за своего относительно небольшого размера и высокого сродства к стрептавидину биотин долгое время был излюбленной молекулой для мечения и очистки биологических белков. Однако биотин также является важным коферментом, необходимым для многочисленных клеточных реакций, включая правильный синтез жирных кислот и метаболизм (Fletcher and Myant, 1960). Следовательно, неудивительно, что оеноциты личинок, которые экспрессируют многие липид-модифицирующие ферменты, необходимые для метаболизма жирных кислот, имеют относительно высокие концентрации биотина, который может быть помечен с использованием стрептавидина, конъюгированного с флуорофором.Мы показали полезность окрашивания стрептавидином у видов, встречающихся в пределах Diptera, Coleoptera и Hemiptera, предполагая, что этот метод мечения оеноцитов, вероятно, будет широко применим ко многим видам насекомых. Однако следует помнить о двух важных предостережениях при использовании стрептавидина для мечения клеток. Во-первых, способность окрашивания стрептавидином строго маркировать личиночные эноциты зависит от стадии. Например, окрашивание стрептавидином распознало эноциты в личинке третьей стадии Drosophila melanogaster , но не метило эти клетки в эмбрионе.Напротив, стрептавидин сильно пометил оеноциты Tribolium вскоре после их спецификации в эмбрионе жука. Это дифференциальное мечение личиночных эноцитов стрептавидином предполагает, что каждый организм может иметь особые потребности в концентрации биотина для ферментативных реакций внутри эноцитов в разные моменты времени развития. Во-вторых, поскольку биотин играет важную роль в цикле лимонной кислоты, стрептавидин, вероятно, метит большинство клеток и, следовательно, не является специфическим для эноцитов красителем.Однако клетки, такие как эноциты, которым требуется высокая концентрация биотина по сравнению с их соседями, будут легко обнаружены стрептавидином. Дополнительные типы клеток, включая подмножество нейрональных клеток, помечены стрептавидином в Drosophila , Tribolium и Anasa эмбрионах и / или личинках (не показаны) (Ziegler et al., 1995). Таким образом, для окончательной идентификации клеток как личиночных оеноцитов могут потребоваться другие маркеры и / или морфологические характеристики, такие как размер и положение клеток в организме.Тем не менее, простота получения и использования стрептавидина, конъюгированного с флуорофором, для мечения личиночных оеноцитов делает его мощным инструментом для изучения этого типа клеток.
Сравнительное развитие личиночных оеноцитов между
Tribolium и DrosophilaРазвитие эноцитов можно разделить на два ключевых этапа; спецификация клеток и созревание клеток. У Drosophila спецификация личиночных оеноцитов требует активации передачи сигналов EGF в дорсальных клетках эктодермы, которые экспрессируют фактор транскрипции Spalt.Сигнал EGF исходит от конкретной клетки-предшественника сенсорного органа в заднем отделе брюшных сегментов. В среднем шесть соседних клеток на полусегмент брюшной полости получают достаточную передачу сигналов EGF, чтобы дополнительно регулировать экспрессию Spalt и передавать судьбу клеток оеноцитов (Brodu et al., 2002, 2004). Во время клеточной спецификации оеноциты также экспрессируют несколько других факторов транскрипции, включая белки Ventral-veinless (Vvl) и Seven-up (Svp) (Brodu et al., 2002; Inbal et al., 2003). После спецификации эноцитов эти клетки становятся постмитотическими, остаются сгруппированными и в конечном итоге активируют высокие уровни фактора транскрипции HNF4, а также многочисленные липид-модифицирующие ферменты посредством в значительной степени неизвестных механизмов во время созревания (Gutierrez et al., 2007; Palanker et al. , 2009).
В этом исследовании мы создали и использовали несколько маркеров для изучения развития личиночных оеноцитов у Tribolium castaneum , и наши результаты выявили сходства и различия в развитии оеноцитов между этими двумя видами.Как и в случае Drosophila , личиночные оеноциты Tribolium специфичны для брюшной полости (хотя оеноциты образуются в первых 8 сегментах брюшной полости у Tribolium по сравнению с первыми 7 сегментами брюшной полости у Drosophila ), и эти клетки являются постмитотическими после клеток. Спецификация. Более того, эноциты возникают из Spalt-экспрессирующих клеток дорсальной эктодермы в заднем компартменте, и эти клетки также метят антитело фосфо-ERK. Однако, в отличие от Drosophila , оеноциты Tribolium не активируют Spalt, и его экспрессия не поддерживается в зрелых оеноцитах.Эти находки подтверждают, что Spalt может играть существенную роль в процессе спецификации, но не в созревании эноцитов. Вместо этого созревающие личиночные эноциты как в Tribolium , так и в Drosophila экспрессируют фактор транскрипции HNF4. Кроме того, наш анализ динамики развития vvl-GFP согласуется с оеноцитами Tribolium , участвующими в обширном паттерне миграции, который не наблюдается у Drosophila . После спецификации в заднем компартменте оеноцитов Tribolium становятся локализованными вокруг формирующейся трахеи, которая лежит в более передне-дорсальной области каждого абдоминального сегмента.Кластеры Tribolium впоследствии образуют непрерывную полосу эноцитов между брюшными сегментами, что согласуется с паттерном, описанным в классических исследованиях (Snodgrass, 1993; Wheeler, 1892).
Дополнительным отличием между Drosophila и Tribolium является подсчет эноцитов. Оба вида образуют кластеры оеноцитов, но кластеры Tribolium примерно в 4 раза больше — 22 клетки / кластер по сравнению с 6 в кластерах Drosophila .Это различие может быть результатом более длительного периода, в течение которого оеноциты набираются в Tribolium . Как и Drosophila , большинство оеноцитов Tribolium рекрутируются в начальный импульс, который происходит через 24-26 часов развития жука. Однако дополнительные vvl-GFP -положительные клетки ( оеноцитов Tribolium ) продолжают возникать из полосы Engrailed в небольших количествах до 46 часов развития. Продолжающаяся продукция в vvl-GFP -положительных клетках не является следствием клеточного деления, поскольку мы не обнаружили совместного мечения с маркером пролиферации клеток фосфо-гистоном-3.Следовательно, личиночные оеноциты являются постмитотическими после спецификации как у Drosophila , так и у Tribolium . Кроме того, мы обнаружили, что передние брюшные сегменты продуцируют больше эноцитов на сегмент, чем задние брюшные сегменты у Tribolium , но не у Drosophila . Это открытие предполагает, что насекомые с короткими зародышевыми полосками ( Tribolium ) производят меньше эноцитов в задних сегментах, которые добавляются позже в процессе развития, тогда как насекомые с длинными зародышевыми полосками ( Drosophila ), которые образуют все брюшные сегменты, одновременно производят одинаковое количество эноцитов в каждый сегмент.
Оставшийся без ответа вопрос, который поднимают наши открытия, заключается в том, что является источником сигнала фосфо-ERK в Tribolium и зависит ли он от передачи сигналов EGF? В отличие от Drosophila , по-видимому, не существует центральной одиночной сигнальной клетки, поскольку у Tribolium наблюдаются широкие полосы активности фосфо-ERK, которые покрывают более 10 клеток одновременно. Более того, активность фосфо-ERK может представлять активацию нескольких основных тирозинкиназозависимых сигнальных путей, и, таким образом, окрашивание фосфо-ERK не окончательно демонстрирует, что передача сигналов EGF участвует в спецификации оеноцитов в Tribolium .У Drosophila секретирующие EGF клетки можно идентифицировать путем анализа экспрессии ромбовидных сериновых протеаз , которые обрабатывают EGF (Elstob et al., 2001; Lage et al., 1997). Мы проанализировали эмбриональную экспрессию ромбовидных генов в эмбрионах Tribolium и обнаружили, что единственным ортологом ромбовидной формы с различимым паттерном экспрессии является ромбовидный-A (корневой ортолог ромбовидных образований 909–399 D.melanogaster ). Однако, хотя мы обнаружили экспрессию rhomboid-A в области образования трахеи (не показано), мы не обнаружили экспрессию, относящуюся к области рекрутирования оеноцитов. Это открытие может указывать на то, что либо ромбовидный не является источником передачи сигналов фосфо-ERK, чтобы индуцировать оеноциты, либо что уровни ромбовидного , экспрессируемого в этой области, ниже нашего порога обнаружения. В соответствии с более поздней возможностью, экспрессия Drosophila rhomboid в сенсорных клетках брюшной полости, которые индуцируют эноциты, намного ниже, чем экспрессия, обнаруженная в трахеальных ямках Drosophila .Следовательно, будущие исследования потери функции ромбовидных генов и пути EGF необходимы для окончательного установления их роли в спецификации эноцитов Tribolium .
Ультразвуковой туман. Ультразвуковой генератор тумана: характеристика, фото и отзывы
Не спорю, можно было бы быстрее купить готовый ультразвуковой увлажнитель, но у меня его так много, что вышел сам. В статье я покажу, как и из чего я сделал, а в конце расскажу, как бы я сделал это сейчас, исходя из опыта текущего запуска.
В марте приехал ультразвуковой опрыскиватель в пластиковый корпус, специально готовился к лету, собирали конструкцию, но в один прекрасный день в ультразвуковой головке не сработал датчик уровня, и в результате работы на сухой — корпус сенсора оплавился, и кое-где правда, заметил не сразу — стоило.
Неделю назад мне попала ультразвуковая головка в металлическом корпусе и поэтому пришлось разбирать всю конструкцию и переделывать заново.
Нужно
В наличии было:
- — Ведро офисное емкостью 10 литров;
- — блок питания на 12 вольт;
- — опрыскиватель ультразвуковой в металлическом корпусе;
- — монтажная коробка черного цвета размером 100 x 60 x 25 мм;
- — любой буст-модуль, у меня оказался модуль XL6009;
- — Регулятор опрокидывания 12 вольт;
- — Турбина;
- — выключатель питания, несколько гнезд и вилок к ним;
- — в процессе сборки появилась дурацкая мелочь;
- — А еще — случай с неисправным контроллером бунта — вы увидите дальше.
Визуальная составная схема
Пришлось постараться, чтобы все это не выглядело кашей и было удобно для восприятия.
- — на входе 12 вольт поднимается на модуль до 22 вольт и подается на ультразвуковой распылитель;
- — Также вход 12 вольт подается на вентилятор, регулирующий оборот;
- — оба подключены параллельно и через общий выключатель питания, подключенный к входному разъему.
Готовый регулятор revolt сразу пришел ко мне неисправным, и как это бывает, поставил его в кучу «до лучших времен», его корпус с четырьмя удобными отверстиями для крепления вошел в корпус.Вы видите, что случилось с начинкой. Некоторое время ток был настолько велик, что проводка, ведущая к гнезду на ультразвуковой головке, сгорела и оплавилась. Однако оба модуля оказались хорошими и потребовалось только заменить проводку.
В торце офисной корзины просверлил четыре отверстия для установки шурупов, к которым крепился блок управления, еще ниже вы видите отверстие для пропуска провода, идущего к ультразвуковой головке.
Первый вариант я собрал в секрете, и такое решение выявило его недостатки.Если заглянуть внутрь корзины, то видно, что винты шурупов были коррозией.
Для избавления от ржавчины потребовалось обработать ее тампонами с увлажненной жесткой пищевой лимонной кислотой
После чистки зубной щетки и сушки головки залили прозрачным акриловым клеем из строительного магазина.
Узел вентилятора
Он должен быть полностью защищен от брызг, и после недолгих размышлений я решил применить центробежную турбину, вклеенную в черный пластиковый ящик на стержнях двустороннего монтажного скотча из расчета 1 мм.
Через меньшее отверстие в нижней крышке коробки воздух подается внутрь корзины. Обратите внимание, что входное отверстие на верхней стороне коробки и градуировка внизу будут расположены напротив друг друга. Таким образом, никакие брызги не смогут попасть на газотурбинный двигатель. По периметру полученного узла наклеил уплотнительную ленту, применяемую для герметизации проема открывающихся пластиковых окон, а к самой турбине добавился отрезок скрученного шнура от заглушек на конце.Паяные контакты заделаны горячим клеем.
Узел накладки
Оборотная сторона.
Крышка офисного ведра, как известно, оснащена поворотным клапаном, и он доставил мне больше всего неудобств.
- — Сначала наметили и проделали дырку для наружного тумана;
- — Потом просушил отрезал прямоугольное окно для веерного узла;
- — Чтобы заглохнуть клапан, по всей внутренней части крышки наклеил водостойкий клей из поролона;
- — заготовка, чтобы пары не травили, пришлось хорошо смочить в одном клее в несколько приемов;
- — После высыхания на почти водоотталкивающую поролоновую заготовку наклеил заготовку из обрезки линолеума.
Крышка с лицевой стороной. В отверстие для выпуска мурены вставляется половинка от шоколадного яйца. С некоторым усилием он может вращаться. Советую прожигать в нем отверстия только с одной стороны, чтобы поток холодного пара мог быть направлен в сторону от вентилятора и отверстия для забора воздуха.
В итоге общий вид Моющая офисная корзина без установленного здесь вентиляторного узла.
Чтобы дать меньше колхоза, по контуру поворотной заслонки наклеил оставшийся росток.
Узел качающийся
Поплавок круглый вырезал из вспененного полиэтилена, в рамке из такого материала «одеваю» дисплеи и телевизоры.Чашка йогурта вставляется в поплавок, в который будет вставлен ультразвуковой распылитель.
Первые тесты сразу показали, что ультразвуковая головка должна быть утоплена под поверхностью воды, глубиной фаланги пальца, но при этом из фантазии тумана летели отдельные брызги.Поэтому пришлось подумать о капельке. Он сделан из крышки цилиндра с монтажной пеной, а на счастье он имел проушину с отверстием для полиэфирной трубки.
Следы ржавчины объясняются тем, что вместо нейлоновых стяжек я использовал металлический штифт, и после пропитки лимонной кислотой окончательной сборки наложу их на них.
Собственно, все — чистка завершена, потом серию фото с пояснениями собираюсь увидеть процесс окончательной комплектации запчастей.
Для нее я поделюсь своими мыслями, что я бы поступил иначе и видео увлажнителя воздуха в коллекции.
Узел электроники
Провода отремонтированы. При этом слева вы видите розетку для подключения блока клапанов.
И крышка закрыта. Две нижние розетки. Правый, выход на ультразвуковую головку, левый разъем предназначен для подключения внешнего блока питания +12 вольт.
Ультразвуковая головка и поплавковая система
Мне пришлось обрезать обычный провод из-за его плохой гибкости и бороться с ним гибкими проводниками в силиконовой оболочке.Игровые места щедро заклеены горячим клеем. И, обратите внимание — проволока продета через силиконовый колпачок слабой банки с антибиотиками.Вы пропилили отверстие в офисной корзине, в нее будет вставлена крышка с проводом поперек ее центра, что не только будет препятствием для выхода мелкого тумана, но и позволит всему узлу удалить этот узел без перекусов кондукторов.
А вот плавучую платформу пришлось полностью поменять. Металлический опрыскиватель был тяжелым для нее, а плавучесть была отрицательной.
Взял как вы видите пенопласт, мне повезло, это плотный пенопласт из пенопласта размером в ширину 24 мм и партии 100 на 115 мм.
Корзина для ультразвуковой головки также должна была заменить целую чашку из-под йогурта. Распылитель был плотно врезан в чашку до дна, а в паяльнике отверстия были прорезаны для доступа воды внутри этой небольшой емкости.
Придется экспериментально выяснить плавучесть платформы, но сразу скажу — альтернативы пене нет.
Пробный запуск
Корзина — нанита воды, ультразвуковой узел опущен на поверхность, заглушка ультразвукового узла через силиконовый колпачок прошла через стенку офисной корзины. Также вы видите, что по внутреннему периметру корзины наклеен все тот же уплотнительный шнур.
Система на средней оборачиваемости.
Потребление системы было при максимальном обороте вентилятора и внешнем источнике питания 12В — 1,92А. Без вентилятора 1.72a.
Итак, я изменился сейчас.
Во-первых, крышка, как мне кажется, не совсем удачная. Поднимитесь вверх, пока не появится картинка, на которой я показал перевернутую крышку. Будет лучше, если из цельного листа пластика вы вырежете заготовку по размеру с внутренней стороны (ступеньки) крышки. После подбора размеров и проверки герметичности вентиляторный блок можно поместить в получившееся место под крышкой вращающегося клапана офисной корзины. Думаю, места для другой электроники тоже хватило.Какой?
Например — датчик влажности. Есть модули с датчиками влажности, совмещенные с реле, и после калибровки и установки модуля на влажность 40% об играх с переключателем можно будет забыть. Влажность всегда будет автоматически поддерживаться на оптимальном уровне.
Во-вторых, система безопасности. Я догадываюсь, что в пластиковом корпусе звенел бывший генератор тумана. На нем (как и на этом) был установлен емкостной датчик в виде кронштейна и, вероятно, генератор тумана в силу его простоты — датчик бака находился в воде, а пьезомембрана оказалась частью воздух, что привело к перегреву всей головы.На микросхеме TTP222 есть компактные датчики-контейнеры, так же можно наклеить на минимальном уровне воды в корзину снаружи, что гарантировано этой ультразвуковой головкой хоть и жесткой, но все же, в воде она была бы. Сам датчик может управлять модернизированным модулем, модуль повышения имеет управляемый вход.
В-третьих, может быть дешевле увеличение модуля, не обязательно того, что я использовал — больше под рукой не было.
Примерная стоимость всего комплекта:
- — Корзина офисная — 2 шт.5 долларов.
- — Опрыскиватель ультразвуковой — 5,6 дол.
- — увеличение модуля XL6009, который может быть другим — 0,80 доллара.
- — Турбина — 1,43 доллара.
- — Черный ящик 100х60х25 мм — 1,08 доллара.
- — Готовый регулятор революции стоит 1,32 доллара.
Все остальное у меня было в наличии. Считаю, что эта самоделка, не относящаяся к центру праздничного стола как самовар, однако обладает всеми необходимыми потребительскими качествами, которых за эти деньги в готовом варианте, скорее всего, не найти.
Спасибо за внимание.
Руслан.
Ультразвуковая пушка собрана своими руками на всех двух логических инверторах и имеет минимальное количество компонентов. Несмотря на простоту сборки, конструкция довольно мощная и может применяться против пьяных пьяниц, собак или подростков, которые сидят и поют в чужих подъездах.
Схема ультразвукового пистолета
Для генератора микросхемы CD4049 (HeF4049), CD4069 или отечественные микросхемы K561LN2, K176PU1, K176PU3, K561PU4 или любые другие стандартные логические микросхемы с 6 или 4-мя логическими инверторами, но придется менять подвал.
Схема нашей ультразвуковой пушки выполнена на микросхеме HeF4049. Как уже было сказано, нам нужно использовать всего два логических инвертора, а какой из шести инверторов использовать — решать вам.
Сигнал с выхода последней логики усилен транзисторами. Для накатки последнего (силового) транзистора в моем случае применены два маломощных транзистора КТ315, но выбор огромный, можно поставить любые NPN транзисторы малой и средней мощности. .
Выбор ключа питания тоже не критичен, можно поставить транзисторы из серии КТ815, КТ817, КТ819, КТ805, КТ829 — последний составной и будет работать без дополнительного усилителя на маломощных транзисторах.Для увеличения выходной мощности можно использовать мощные композитные транзисторы типа CT827 — но для его раскачки все равно понадобится дополнительный усилитель.
В качестве излучателя можно использовать любые SC и RF головки мощностью 3-20 Вт, также можно использовать пьезоизлучатели от сирен (как в моем случае).
Выбор конденсатора и сопротивления сильного резистора — Частота настраивается.
Такой ультразвуковой пистолет, собранный своими руками, вполне подойдет для защиты загородного участка или частного дома. Но не нужно забывать — ультразвуковой диапазон опасен! Не слышно, но тело чувствует. Дело в том, что уши воспринимают сигнал, но мозг не в состоянии его отклонить, отсюда такая реакция нашего тела.
Собирайте, тестируйте, радуйтесь — но будьте предельно внимательны, и я прощаюсь с вами, но ненадолго — он же Касьян.
Климатическая техника постоянно развивается, предлагая потребителю новые возможности регулирования параметров микроклимата.Решение проблемы увлажнения воздуха давно вышло из разряда опциональной заправки массивных конденсаторов и реализовано в виде полноценных автономных устройств. Сегодня желающим освежить помещение в доме частицами влаги предлагается огромный ассортимент компактных увлажнителей воздуха. И отдельное место в этом классе техники занимает туман, который тоже подходит для бытового использования, но при этом имеет свои особенности эксплуатации.
Общие сведения о генераторах тумана
Внешне такие устройства напоминают большие увлажнители воздуха или малогабаритные мобильные кондиционеры.По конструкции это небольшой блок, который подключается к электросети и работает в заданном режиме. Намного интереснее принципы работы устройства. Есть два типа генераторов — для образования матовости (декоративные) и непосредственно для увлажнения холодного тумана. В первом случае генерация осуществляется механически под давлением — через насос и компрессор буквально выдавливается и мелкие частицы жидкости выбрасываются в помещение. Особенность таких систем в том, что они работают не с обычной водой, а с аэрозолями и модифицированными смесями.Конечно, они тоже содержат воду, но в качестве разбавителя. В качестве активных ингредиентов обычно выступают глицерин или гликоль. В свою очередь, ультразвуковой генератор тумана направлен на распыление частиц холодной воды с целью изменения микрокламатических характеристик. Принцип работы этого устройства заслуживает отдельного внимания.
Как работают ультразвуковые модели?
Чтобы понять особенности работы таких устройств, важно выделить две составляющие рабочего процесса — это непосредственно генерирующий ультразвуковой аппарат и рабочая среда.Первый компонент воздействует на рабочую жидкую среду колебательными волнами (ультразвуком) с частотами, достаточными для разрушения поверхностного слоя на мелкие элементы. В процессе работы водная среда называется озвученной — в данный момент ультразвуковой генератор тумана выполняет над ней следующие операции:
- Воздействие на поверхность.
- Увеличение поверхности взаимодействия звуковых волн с жидкостью.
- Дисперсия. Дисперсия жидких частиц на фоне тонкого помола.
- Эмульгирование. Генерация эмульсий.
Не всегда обязательно выполнять полный цикл описанных операций. Более того, не каждый генератор в принципе может осуществлять, например, диспергирование и эмульгирование. Конкретные режимы работы со списком операций устанавливает сам пользователь.
Технические характеристики
В характеристиках устройства можно проследить отличия от обычных увлажнителей. В первую очередь это сила.Для квартиры или небольшого дома вполне хватит инструмента на 700-1000 Вт. Но если мы говорим о больших студиях, площадках или беседках, то мощность должна быть порядка 1200-1500 Вт. Из этого показателя производительность, составляющая 250-300 м 2 / мин и далее. То есть это количество выработки и выпуска пара. Иногда производители подсчитывают эту сумму в «кубиках». В этом случае средний показатель производительности составит 50-70 м 3 / мин. Что касается характеристик блока питания, то для обслуживания бытового ультразвукового увлажнителя воздуха достаточно 220 В.А вот генератор тумана из профессионального сегмента может питать трехфазную сеть напряжением 380 В. Это промышленные установки, которые также отличаются большими размерами.
Положительные отзывы
С точки зрения обычного потребителя, генераторы ультразвука отличаются способностью быстро заполнять помещения до тумана. Если тому же увлажнителю или даже парогенератору для обеспечения желаемого результата требуется от 30 минут до 1,5-2 часов, то в случае с генератором тумана — 8-10 минут.К достоинствам также можно отнести качество распыления, которым отличается даже бюджетный туманообразователь. Ультразвук хорошо проявляет себя с точки зрения защитных качеств. Он устойчив к перегрузкам в сети и внешним воздействиям, в том числе температурным.
Отрицательная обратная связь
Несмотря на привлекательные характеристики, эргономика большинства генераторов этого типа оставляет желать лучшего. И в плане дизайнерских качеств, и в управлении пользователи больше ценят аккуратные и компактные устройства для увлажнения и промывки воздуха.Кроме того, генераторы требуют особого обслуживания. Например, во время работы следует периодически очищать или заменять мембрану ультразвукового увлажнителя тумана генератора, но компоненты для такого оборудования не всегда легко найти на рынке. С другой стороны, многие владельцы отмечают высокую степень надежности конструкции, поэтому поломки случаются редко.
Самостоятельное производство генератора
Для сборки генератора вам понадобится ультразвуковой модуль для функции колебания, источник питания, пластиковый контейнер вентилятора и водопроводная арматура для подачи жидкости.В процессе работы ультразвуковой модуль будет создавать колебательный эффект с выбросом мелких частиц воды. В свою очередь, перед этим модулем устанавливается вентилятор, выводящий поднятые частицы за пределы помещения. Соответственно, вода все это время будет находиться в емкости. Какие функции еще должен иметь ультразвуковой генератор тумана? Своими руками можно собрать поплавок, который будет подниматься и опускаться в зависимости от уровня воды. Эта функция важна, если вы планируете оставить устройство в рабочем состоянии несколько часов или даже дней.По мере того, как жидкость рассеивается, система подойдет к точке отключения, которая просто вызывает поплавок с механикой пресса на соответствующую кнопку источника питания.
Как выбрать ультразвуковой генератор тумана?
Многое зависит от назначения генератора. Чаще всего такие устройства приобретаются для увлажнения помещения, если до этого оно было сухим. В этом случае важны основные параметры, среди которых интенсивность образования тумана, производительность и качество распыления как таковые, чтобы образовавшееся облако не привело к чрезмерному приближению обоев или мебели.Опять же, есть модели, предназначенные исключительно для декоративной функции. Они создают только дымку определенной плотности, которую можно использовать на студийных съемках. В этом случае необходимо отдать предпочтение ультразвуковому генератору тумана с подсветкой, который сделает то же облако визуально ярче и эффектнее. Важно учитывать возможность использования генератора вне дома, то есть без возможности подключения к электросети. Такие устройства есть — на них нужно будет рассчитать потенциал аккумулятора.В среднем аккумулятор обеспечивает автономную работу в течение 30-60 минут.
Заключение
При всех достоинствах генераторы с функциями формирования тумана еще не полностью вписались в сферу бытового использования. Об этом говорит и отсталая эргономика, и устаревшая система управления. И это понятно, поскольку инструменты пришли из промышленного сегмента, в котором такие качества практически не учитываются. И напротив, это решение более чем подходит для жилищного строительства.Что касается стоимости, то она тоже немаленькая — в среднем 5-6 тысяч рублей. Однако на ресурсе Avito ультразвуковой генератор тумана можно найти за 2 тысячи здесь, кстати, можно будет договориться с продавцами комплектующих с расходниками к устройству. Стоят они тоже недорого, но важно учитывать соотношение технических параметров Компоненты с конструкцией генератора. Для тех же мембран размеры стандартизированы, но найдены современные модификации, к которым предъявлены новые нормативные требования.Поэтому перед покупкой стоит обязательно проверить соответствие отдельных деталей и комплектующих.
Всем привет!
Зимой особенно актуальным становится вопрос увлажнения воздуха в квартире. На курорт поступает огромный ассортимент всевозможных устройств, некоторые с задачей справляются, а некоторые нет.
Итак, в погоне за экономией, 3 года назад не совсем правильно выбрала, и приобрела прибор нулевой эффективности по увлажнению воздуха. После нескольких дней попыток использования это «ведро с вентилятором» отключили и сняли с глаза.
Позже на просторах Алиэкспресс был замечен дешевый аппарат — герой сегодняшнего обзора. Было решено приобрести его и модернизировать вышеупомянутый «ковш».
Подробнее Из этого получилось под кат.
Внешний вид, размеры и вес
В доставке и упаковке нет ничего особенного. Доставка длительная (2 месяца), упаковка — пуховая обычная упаковка. Купил сразу 3 штуки на случай, если одна не справится или зарезервирует.Устройство выглядит как небольшой цилиндр с проволокой.Сверху есть отверстие, в которое вкручивается пластиковая гайка, откручивая, мы увидим мембрану, которая естественно не прилипает. Больше ничего интересного. С нижней стороны есть прорези для забора воды.
Возможно снизу это тоже что-то можно открутить, но не смог, но извините.
Есть модификации устройств со встроенными светодиодами.
Габаритные размеры устройства:
Диаметр — 35мм.
Высота — 29мм
На блоке питания есть подвижная резиновая заглушка диаметром 16мм.
Вес устройства 58 грамм
Длина кабеля — 93 сантиметра
Разъем для подключения блока питания круглый, центральный плюс, внешний минус.
Блок питания 24 В и устройство 1a приобретались отдельно.
Штепсельная вилка имеет диаметр около 6 мм.
Мощность кабеля питания 91 см.
Демонстрация работы
После включения устройства в сеть и погружения в воду , он автоматически начинает работать. Как только вода закончится, устройство автоматически выключится. Для нормальной работы его необходимо погрузить в воду на глубину 3-3,5 сантиметра. Если опустить на 5 см ниже, то небольшой фонтан выйдет из воды без тумана (пара).Демонстрация работы устройства на небольшом видео.
Колхозная плавающая корзина
Идея доработки заключалась в том, чтобы поставить рабочий опрыскиватель в его бак с водой, а штатная система обдува влажным воздухом сделает все остальное.Для того, чтобы прибор постоянно находился на определенной глубине, было решено взять обычный пластиковый стакан, сделать на дне стакана отверстие и прикрепить к нему любой нехостинговый материал. Под рукой не было обычного поролона нужного размера, поэтому было решено использовать имеющийся герметик для окон и дверей как необычный материал.
Чтобы не нервничать! Дикий колхоз !!!
Допирование имеющегося «увлажнителя»
Далее в пластиковом резервуаре для воды просверливаем отверстие под провод с заглушкой.Сверла больше 12мм у меня не получилось поэтому отверстие получилось не очень ровным.
Все остальное собираем и проверяем как работает.
Отлично, можно верх с вентилятором поставить.
Получившийся результат
Собираем «ведро», включаем сначала разбрызгиватель воды, потом вентилятор на «ведро».Малая демонстрация на видео
Наружная решетка сделана таким образом, что он направил увлажненный воздух вниз, поэтому я ее снял, перевернул и приклеил обратно на двусторонний скотч.
выводы
В результате доработок внешний вид Первоначальный увлажнитель практически не изменился. Оцифровать КПД устройства не могу, к сожалению, гигрометра нет, но в результате работы допированного увлажнителя наблюдается следующее:1. После нескольких часов работы начали запотевать окна и это при — 20 градусов по ул.
2. Утром после пробуждения сухости в носу нет и т.д …
3.8 литров воды, вмещающей емкость, испаряются за 2-е сутки. В этом случае устройство работает только половину времени.
Все вышеперечисленное говорит об эффективности полученного увлажнителя.
В увлажнителе я использую воду из-под фильтра с обратным осмосом, чтобы на поверхностях в помещении не появлялись известковые образования.
Работой получившегося увлажнителя доволен.
Из минусов:
1. Требуется еще розетка.
2. При работе аппарата слышен небольшой «буффель».
Вот и все.
Всем здоровья и спасибо за внимание!
Экология потребления. Подробности: Многие считают приобретение увлажнителя воздуха для квартиры делом неудачным и ненужным. Но это совсем не так. Это устройство вовсе не роскошь. Тем более что сделать своими руками ультразвуковой увлажнитель воздуха можно.
Многие считают, что приобретение увлажнителя воздуха для квартиры — дело неудачное и ненужное.Но это совсем не так. Это устройство вовсе не роскошь. Тем более что сделать своими руками ультразвуковой увлажнитель воздуха можно.
Очень часто увлажнители приобретают в связи с необходимостью регулировки качества воздуха в тех помещениях, в которых постоянно проживают люди. Такая необходимость может быть вызвана появлением заболеваний, причиной которых может стать некачественный сухой воздух, затрудняющий дыхание и негативно влияющий на общее самочувствие. Кроме того, сухость на воздухе может навредить не только людям, но и предметам мебели из дерева, натурального паркета, а также растениям и цветам.
Для детской комнаты увлажнитель воздуха просто незаменимый прибор, так как недостаточная влажность воздуха вредит детскому организму. Сухой воздух повреждает слизистые оболочки и увеличивает вероятность респираторных заболеваний.
Любые увлажнители воздуха для дома и офиса не требуют установки и имеют возможность круглые сутки в закрытом помещении, которым может быть офис, квартира или офис. Такие устройства работают достаточно тихо, поэтому их можно установить в спальне.
Типы увлажнителей
По устройству и принципу работы можно выделить пять основных типов увлажнителей воздуха:
Это наиболее традиционный вид увлажнителей. Функционирование этого устройства заключается в холодном испарении жидкости. Вода заливается в емкость, затем направляется на поддон, а оттуда в испарительные картриджи. Самые простые модели имеют сменные бумажные фильтры. От испарительных элементов воздух, увлажняясь, уходит в комнату.
Такие устройства не только увлажняют воздух, но и очищают его. Вся грязь скапливается в картриджах и фильтрах. Для увлажнителей холодного воздуха используется дистиллированная вода. Достоинством таких устройств является еще и то, что они очень экономят электроэнергию. Важно, чтобы они работали практически без шума. Увлажнители этого вида принято устанавливать в детской и спальне, а также в офисах. Недостаток такого устройства заключается только в том, что они имеют небольшую емкость.
Функционирование этих устройств основано на испарении жидкости за счет ее нагрева. Такие устройства работают по принципу электрочайника. Если вода в испарителе полностью перевернулась, сам прибор отключается. Поэтому такие устройства полностью пожаробезопасны. Этот тип увлажнителей имеет гидростат, который позволяет прибору контролировать влажность воздуха и выключаться самостоятельно, когда этот показатель достигает желаемого значения.
Паровой или ультразвуковой увлажнитель воздуха лучше всего использовать для создания комфортного климата в Апельсинах.Положительные качества паровых приборов — удобство эксплуатации и использования их для ингаляций. Вам просто нужно добавить в прибор настой из лекарственных компонентов или ароматических масел. Ароматические масла позволяют применять прибор также в качестве освежителя воздуха.
Принцип действия ультразвукового увлажнителя воздуха заключается в переводе частиц воды в состояние «водяного облака». Это происходит не за счет закипания жидкости, а за счет высокочастотных колебаний. Сухой воздух попадает в увлажнитель.Затем с помощью вентилятора выводятся в виде холодного тумана. Такие увлажнители не нагревают воду, поэтому их можно использовать в помещениях, где находятся маленькие дети.
Ультразвуковые увлажнители воздуха подходят для помещений, в которых находятся объекты, требующие поддержания определенного уровня влажности. К таким предметам относятся антиквариат, антикварная мебель, музыкальные инструменты. Также в жилом помещении можно установить ультразвуковой увлажнитель воздуха.
Принцип работы ультразвукового увлажнителя воздуха позволяет очень точно контролировать влажность.Конечно, такие устройства довольно дороги, но их достаточно просто сделать самостоятельно.
Такие увлажнители помогают не только увлажнять, но и очищать воздух от любых загрязнений. Процесс увлажнения проходит по принципу работы паровых приборов. Фильтр, вставленный в устройство, чаще всего является антиаллергенным и антибактериальным. Если устройство оборудовано угольным фильтром, это поможет уберечь помещение от неприятных запахов и табачного дыма. Эти комплексы можно устанавливать в офисах и детских комнатах.В некоторые модели можно вставлять капсулы для ароматерапии.
Хотя все эти типы увлажнителей имеют свои преимущества, но все же используются ультразвуковые увлажнители, которые можно изготовить самостоятельно. Помочь в этом может пошаговая инструкция, описанная ниже.
Такие увлажнители воздуха распыляют водную суспензию, в которой находятся мельчайшие частицы воды. Извлеченная из увлажнителя суспензия переходит в состояние пара. Такие устройства очень мощные, но стоят очень дорого. Поэтому такие увлажнители, которые еще называют форсунками, устанавливают только в производственных помещениях, где они могут себя оправдать.
Преимущества и недостатки ультразвуковых увлажнителей воздуха
Положительные стороны таких устройств видны невооруженным глазом. К ним можно отнести высокую степень безопасности, простоту использования, малошумный показатель и небольшие размеры, что очень важно для российских квартир. К тому же такие увлажнители, несмотря на хорошую производительность, экономят электроэнергию. Точная регулировка влажности также является важным преимуществом.
Единственным недостатком таких устройств является жесткая потребность в воде.Лучше всего, если он будет дистиллированным.
Создание ультразвукового увлажнителя воздуха
Для того, чтобы сделать ультразвуковой увлажнитель воздуха своими руками, вам потребуется:
- Ультразвуковой отпариватель.
- Вентилятор для компьютера.
- Пластиковая тара на 5 или 10 литров.
- Пластиковый стаканчик.
- Деталь детской пирамиды в виде бублика.
- Электропитание на 24В 7.
- Гибкая труба, гофрированная.
- Стабилизатор.
- Алюминиевый уголок
Каждый необходимый для создания увлажнитель воздуха можно найти дома или купить в магазине. Общая стоимость такого самодельного приема составит не более одной тысячи рублей, что значительно ниже стоимости заводского увлажнителя воздуха. Итак, как сделать ультразвуковой увлажнитель воздуха? Опишем весь процесс.
Подпишитесь на наш YouTube канал Ekonet.ru, который позволяет смотреть онлайн, скачивать с YouTube бесплатно видео о выздоровлении, омоложении человека..
Нравится, поделись с друзьями!
https://www.youtube.com/channel/ucxd71u0w04qcwk32c8ky2ba/videos.
Подписаться —
Лабораторный блок питания TL431 — Buzz On Live
Всем привет, сегодня мы сделаем лабораторный блок питания на корпусе старого компьютерного блока питания ATX. Он не идеален, но имеет право на жизнь!
Технические характеристики устройства следующие: Стабилизированное регулируемое выходное напряжение 2,7-17В Регулируемый выходной ток 0-3А (5А) Мощность 50 (80) Вт Линейная схема
Инструменты и материалы:
— Неисправен блок питания ATX
— Конденсаторы 2шт. 100nF
-Конденсаторы 1шт 10мкФ
-Конденсаторы 2шт 2200мкФ
-Мощный транзистор npn 3шт MJE13007
-Транзистор KT817 (kt815) BD139
-Резисторы разного номинала и мощности
-12V Zeneriometer
-12V Zeneriometer
-12V
— Микросхема TL431
— Транзистор 1шт 2N5551
— Переменный резистор 1шт 1К
— Переменный резистор 1шт 10К
— Предохранитель 2А
— Трансформатор TN-7 (любой TN) или другой
— Диодный мост не менее 6А
— Паяльник
— Паяльник
Припой
-Flux
Схема:
Диоды D1 и D2 — 1N4007, C6 и C7 на 100 нФ.Т1 и Т5, Т3 любой мощный n-p-n транзистор (13007).
R6 — шунт 0,22 Ом, T2 — KT817 (KT815, BD139). А Т4 — 2N5551, Br1 — диодный мост на 6А и больше!
P1 для 10 кОм, P2 для 1 кОм, R2 для 240 Ом 2 Вт, R4 для 1 кОм 1 Вт и R8 10 кОм 2 Вт (5 Вт).
D4 — стабилитрон 12В, охладитель 12В. R7 на 220 Ом (не 1КОМ), также трансформатор Тр1 ТН-7 или любой другой!
Creation
Разберем блок питания ATX и возьмем корпус и радиатор, кулер, возможно, еще и диодный мост.
Ставим диодный мост и транзистор на термопасту!
Паяем конденсатор и радиатор с деталями на макетную плату.Просверливаем отверстия под клей, трансформатор и потенциометр.
Ввинтить трансформатор в корпус, припаивать первичную обмотку через предохранитель к кабелю.
Перед этим подключить обмотки на 220В и 12,6В.
Паяем схему и кулер.
Закрепляем клеммы и потенциометр, закрываем крышкой! Подключаемся к сети.
Формулы и расчеты
Для начала необходимо определить сопротивление резистора. При максимальном входном напряжении 19 В согласно закону Ома сопротивление рассчитывается следующим образом:
R = U / I = 19 В / 0.
