Что такое резистор и как он работает. Какие виды резисторов существуют. Где применяются резисторы в электронике и технике. Как выбрать подходящий резистор для конкретной задачи. На что обращать внимание при покупке резистора.
Что такое резистор и принцип его работы
Резистор — это пассивный радиоэлектронный компонент, основное назначение которого — создание определенного электрического сопротивления в цепи. Принцип работы резистора основан на преобразовании электрической энергии в тепловую за счет столкновения свободных электронов с ионами кристаллической решетки проводника.
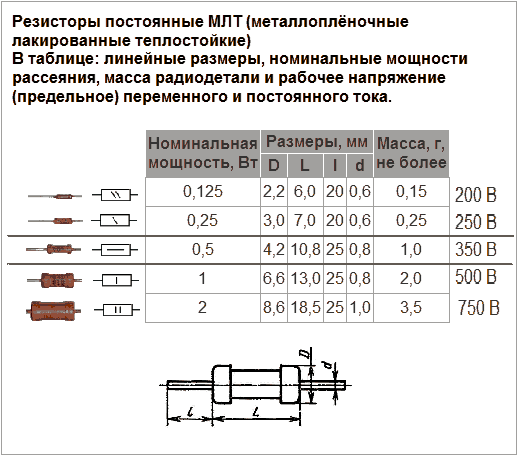
Основные характеристики резистора:
- Номинальное сопротивление (измеряется в Омах)
- Допустимая мощность рассеивания
- Температурный коэффициент сопротивления
- Предельное рабочее напряжение
- Точность (допуск)
Как выбрать подходящий резистор? Нужно учитывать следующие факторы:
- Требуемое номинальное сопротивление
- Мощность, которую должен выдерживать резистор
- Допустимый разброс сопротивления (точность)
- Условия эксплуатации (температура, влажность и т.д.)
- Габариты резистора
Основные виды и типы резисторов
Существует множество разновидностей резисторов, которые различаются по конструкции, материалам и назначению:

По конструкции:
- Проволочные — изготавливаются из проволоки с высоким удельным сопротивлением
- Пленочные — на диэлектрическую подложку наносится тонкая пленка резистивного материала
- Композиционные — изготавливаются из смеси проводящего порошка и изолирующего наполнителя
По назначению:
- Постоянные — имеют фиксированное сопротивление
- Переменные — позволяют регулировать сопротивление
- Подстроечные — для точной настройки схем
- Прецизионные — с высокой точностью сопротивления
Какой тип резистора выбрать? Это зависит от конкретной задачи и требований схемы. Для большинства бытовых приборов подходят недорогие постоянные резисторы. В измерительной технике часто используются прецизионные резисторы. Для регулировки громкости в аудиотехнике применяются переменные резисторы.
Применение резисторов в электронике и технике
Резисторы находят широкое применение практически во всех областях электроники и электротехники:
- Ограничение тока в цепях
- Деление напряжения
- Создание времязадающих цепей
- Согласование импедансов
- Регулировка усиления в усилителях
- Балластные резисторы в осветительных приборах
- Датчики температуры (терморезисторы)
- Датчики освещенности (фоторезисторы)
Где конкретно применяются резисторы? Вот несколько примеров:
- Бытовая техника — телевизоры, компьютеры, стиральные машины
- Автомобильная электроника — системы управления двигателем, климат-контроль
- Промышленное оборудование — станки с ЧПУ, роботизированные линии
- Измерительные приборы — мультиметры, осциллографы
- Медицинская техника — томографы, кардиографы
Особенности подстроечных резисторов
Подстроечные резисторы (также называемые триммерами) — это особый вид переменных резисторов, предназначенных для точной настройки электронных схем. Их основные особенности:
- Компактные размеры
- Возможность плавной регулировки сопротивления
- Высокая стабильность установленного значения
- Многооборотная конструкция для точной настройки
Где применяются подстроечные резисторы? Основные области применения:
- Калибровка измерительных приборов
- Настройка частоты генераторов
- Регулировка усиления в усилителях
- Балансировка мостовых схем
- Коррекция погрешностей в АЦП и ЦАП
Как выбрать подстроечный резистор? Нужно учитывать следующие параметры:
- Диапазон регулировки сопротивления
- Точность установки
- Температурный коэффициент сопротивления
- Количество оборотов регулировки
- Способ монтажа (для печатного монтажа или проводной)
Резисторы в автомобильной электронике
Автомобильная электроника — одна из важнейших областей применения резисторов. Они используются в различных системах управления и контроля:

- Системы управления двигателем
- Антиблокировочная система тормозов (ABS)
- Система стабилизации (ESP)
- Климат-контроль
- Системы безопасности (подушки безопасности, преднатяжители ремней)
Какие требования предъявляются к автомобильным резисторам? Основные из них:
- Высокая надежность и долговечность
- Устойчивость к вибрациям и ударам
- Работоспособность в широком диапазоне температур
- Стойкость к воздействию влаги и агрессивных сред
- Компактные размеры
Как выбрать резистор для автомобильной электроники? Нужно обращать внимание на следующие параметры:
- Соответствие автомобильным стандартам (например, AEC-Q200)
- Расширенный температурный диапазон (обычно от -40°C до +125°C)
- Высокая стабильность характеристик во времени
- Устойчивость к перегрузкам
- Низкий уровень шумов
Прецизионные резисторы в измерительной технике
Прецизионные резисторы — это резисторы с высокой точностью номинального значения сопротивления и малым температурным коэффициентом сопротивления. Они широко применяются в измерительной технике и высокоточной электронике.

Основные характеристики прецизионных резисторов:
- Точность (допуск) от ±0.01% до ±0.001%
- Температурный коэффициент сопротивления менее 5 ppm/°C
- Высокая долговременная стабильность
- Низкий уровень шумов
- Малая индуктивность
Где применяются прецизионные резисторы? Основные области применения:
- Измерительные мосты
- Эталонные источники тока и напряжения
- Высокоточные АЦП и ЦАП
- Калибраторы электрических величин
- Медицинская диагностическая аппаратура
Как выбрать прецизионный резистор? Нужно учитывать следующие факторы:
- Требуемая точность номинального значения
- Температурный коэффициент сопротивления
- Долговременная стабильность
- Рабочий диапазон температур
- Максимальное рабочее напряжение
Резисторы в силовой электронике
В силовой электронике используются специальные мощные резисторы, способные рассеивать большую мощность. Они применяются в преобразователях энергии, системах электропривода, устройствах защиты.
Основные особенности силовых резисторов:
- Высокая допустимая мощность рассеивания (от десятков ватт до киловатт)
- Способность выдерживать кратковременные перегрузки
- Низкая индуктивность
- Эффективный теплоотвод
- Высокая надежность и долговечность
Где применяются силовые резисторы? Основные области применения:
- Пусковые и тормозные резисторы в электроприводах
- Нагрузочные резисторы в системах электропитания
- Балластные резисторы в сварочных аппаратах
- Шунты для измерения больших токов
- Резисторы в системах защиты от перенапряжений
На что обращать внимание при выборе силового резистора?
- Номинальная и пиковая мощность рассеивания
- Максимальное рабочее напряжение
- Температурный коэффициент сопротивления
- Способ охлаждения (естественное или принудительное)
- Конструкция и способ монтажа
Сопротивление, вентилятор салона TOPRAN 104 117 для VW POLO (6KV) CLASSIC/ESTATE AL14220511
- Главная
- Каталог Все марки машин
- Volkswagen Все модели Volkswagen
-
POLO (6KV) SEDAN/ESTATE
11.

- Резистор кондиционера

Код товара: AL14220511
| Производитель | TOPRAN |
| Код | 104 117 |
| Товар | Сопротивление, вентилятор салона |
| Количество полюсов | 4 |
| Форма штепсельного корпуса | прямоугольный |
- ОЕМ коды:
-
SEAT 701 959 263A VW 701 959 263A - Показать ещё
Производители запчастей используют коды, чтобы правильно опознать запчасть.
Ниже увидите список кодов, с которыми эта запчасть совместима:
| A.I.C. Competition | 52474 |
| ALVADI | AL14220511 |
| ACR | 160201 |
| AUTOGAMMA | GA15283 |
| AUTOMEGA | 150114410 |
| BBR Automotive | 002-60-02049 |
| CASCO | CRS73005AS |
| CSV electronic parts | CRV9042 |
| EAN | 4063926060547 |
| EAN | 959 030 |
| EAN | 9590300000011 |
| ELTA AUTOMOTIVE | Eh2058 |
| ELTA AUTOMOTIVE | XHR0059 |
| ERA | 665013 |
| ESEN SKV | 94SKV023 |
| FEBI BILSTEIN | 19774 |
| FISPA |
10. 9042 9042
|
| FRIGAIR | 35.10080 |
| HERTH+BUSS ELPARTS | 75614920 |
| JP GROUP | 1196850800 |
| KRIOS |
10. 9042 9042
|
| MAXGEAR | 57-0030 |
| METZGER | 0917165 |
| MEYLE | 100 800 0035 |
| NRF | 342070 |
| OSSCA | 00325 |
| PATRON | P15-0013 |
| QUATTRO FRENI | QF00T01343 |
| QUATTRO FRENI | QF10Q00035 |
| SANDO |
SRS73005. 0 0
|
| SEAT | 701 959 263A |
| SIDAT | 10.9042 |
| SWAG | 30 91 9774 |
| THERMOTEC | DEW007TT |
| TOPRAN | 104 117 |
| VEMO | V10-79-0003 |
| VIKA | 89590787301 |
| VW | 701 959 263A |
| WE PARTS | K109042 |
| WILMINK GROUP | WG1778222 |
| WILMINK GROUP | WG1795248 |
- Главная
- Каталог Все марки машин
- Volkswagen Все модели Volkswagen
- POLO (6KV) SEDAN/ESTATE 11.1995-06.2001 Все запчасти для VW POLO (6KV) CLASSIC/ESTATE
 org/ListItem»>
Резистор кондиционера
org/ListItem»>
Резистор кондиционера
 org/BreadcrumbList»>
org/BreadcrumbList»>57691-14: 90 мод. 902030/10-402-1001-1-6-100-104/391 Термопреобразователи сопротивления платиновые
Назначение
Термопреобразователи сопротивления платиновые серии 90 модели 902030/10-402-1001
1-6-100-104/391 (далее по тексту — термопреобразователи или ТС) предназначены для измерений температуры в воздуховоде перед теплообменником на ТЭЦ ПГУ «ГСР ЭНЕРГО», Санкт-Петербург, г. Колпино.
Описание
Принцип действия термопреобразователей основан на преобразовании измеряемой температуры в изменение электрического сопротивления чувствительных элементов (ЧЭ) ТС.
Термопреобразователи конструктивно выполнены в виде измерительной вставки с одним ЧЭ, внутренних соединительных проводов, помещенных в защитный чехол из нержавеющей стали (1.4571), а также клеммной головки, изготовленной из алюминия. ЧЭ ТС имеют номинальную статическую характеристику преобразования (НСХ) типа «Pt100» по ГОСТ 6651-2009 (МЭК 60751).
ЧЭ ТС имеют номинальную статическую характеристику преобразования (НСХ) типа «Pt100» по ГОСТ 6651-2009 (МЭК 60751).
Схема соединения внутренних проводников термопреобразователей с чувствительными элементами: 3-х проводная.
Внешний вид термопреобразователя представлен на рисунке 1.
Рис 1. Внешний вид термопреобразователя сопротивления платинового серии 90 модели 902030/10-402-1001-1-6-100-104/391.
Технические характеристики
Таблица 1
|
Наименование характеристики | |
|
Диапазон измеряемых температур, °С |
от минус 50 до плюс 400 |
|
Температурный коэффициент ТС а, °С-1 |
0,00385 |
|
У словное обозначение номинальной статической характеристики преобразования (НСХ) по МЭК 60751/ГОСТ 6651-2009 |
Pt100 |
|
Номинальное значение сопротивления термопреобразователя при 0° С (Ro), Ом |
100 |
|
Класс допуска ТС по МЭК60751/ГОСТ 6651-2009 |
B |
|
Пределы допускаемого отклонения сопротивления ТС от НСХ в температурном эквиваленте (допуск) по ГОСТ 6651-2009, °С |
±(0,3+0,005|ф |
|
Электрическое сопротивление изоляции при температуре плюс (25±10) °С и относительной влажности воздуха от 30 до 80%, МОм (при 100 В), не менее |
1000 |
|
Диаметр монтажной части ТС, мм |
6 |
|
Длина монтажной части ТС, мм |
100 |
|
Рабочие условия эксплуатации: | |
|
Температура окружающей среды, °С |
от минус 40 до плюс 100 |
|
Относительная влажность воздуха, %, не более |
80 |
Знак утверждения типа
Знак утверждения типа наносится на титульный лист паспорта (в правом верхнем углу) методом штемпелевания и на корпус прибора при помощи наклейки.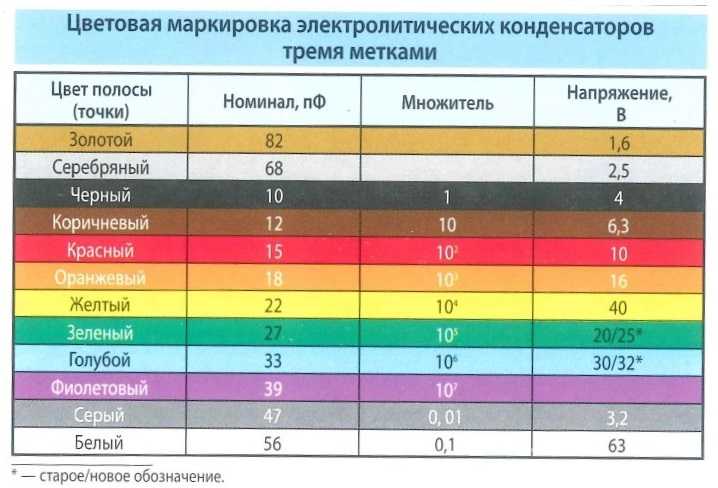
Комплектность
Термопреобразователь сопротивления платиновый серии 90
модели 902030/10-402-1001-1-6-100-104/391 6 шт;
Паспорт 6 экз.
Поверка
осуществляется в соответствии с документом ГОСТ 8.461-2009 «ГСИ. Термопреобразователи сопротивления из платины, меди и никеля. Методика поверки».
Основные средства поверки:
— термометр цифровой прецизионный DTI-1000, пределы допускаемой абсолютной погрешности: ±0,031 °С в диапазоне температур от минус 50 до плюс 400 °С, ±0,061 °С в диапазоне температур св. плюс 400 до плюс 650 °С;
— термостаты жидкостные прецизионные переливного типа моделей ТПП-1.0, ТПП-1.2 с диапазоном воспроизводимых температур от минус 60 до плюс 300 °С и нестабильностью поддержания заданной температуры ±(0,004.0,02) °С;
— многоканальный прецизионный измеритель температуры МИТ 8.10(М) с пределами допускаемой основной абсолютной погрешности измерения сопротивления ±(10-5R+5 10-4), где R — измеряемое сопротивление, Ом;
— мегомметр М4100/3, рабочее напряжение до 500В.
Сведения о методах измерений
приведены в соответствующем разделе паспорта на термопреобразователь сопротивления платиновый серии 90 модели 902030/10-402-1001-1-6-100-104/391.
Нормативные и технические документы, устанавливающие требования к термопреобразователю сопротивления платиновому серии 90 модели 902030/10-402-1001-1-6-100-104/391
ГОСТ 6651-2009 «ГСИ. Термопреобразователи сопротивления из платины, меди и никеля. Общие технические требования и методы испытаний»;
ГОСТ 8.558-2009 «ГСИ. Государственная поверочная схема для средств измерений температуры»;
ГОСТ 8.461-2009 «ГСИ. Термопреобразователи сопротивления из платины, меди и никеля. Методика поверки».
Рекомендации к применению
Осуществление производственного контроля за соблюдением установленных законодательством Российской Федерации требований промышленной безопасности к эксплуатации опасного производственного объекта на ТЭЦ ПГУ «ГСР ЭНЕРГО», Санкт-Петербург, г. Колпино.
Глава 104.
 Мощное сопротивление технике уровня Средний Хуан:Водяной Питон. / Расколотая битвой синева небес
Мощное сопротивление технике уровня Средний Хуан:Водяной Питон. / Расколотая битвой синева небес Увидев молниеносные действия на поле боя, все зрители уставились на Сяо Яна с удивлением. Они не ожидали, что этот парень в прямом столкновении с Великим Мастером возьмет инициативу в свои руки и проведет собственную атаку.
Конечно, атака не была очень эффективной, однако Сяо Ян не особо переживал из-за этого. Он понимал, что если бы не владение комбинацией «Рукой Пустоты» и «Огнедышащей ладони», то его бы быстро раздавили атакой кнута. Так как Сяо Ян не имел точки опоры в воздухе, его тело стало быстро падать. Тем не менее, когда он был в двух-трех метрах от земли, тот же самый кнут, подобно ядовитой змее, закрутился прямо в сторону Сяо Яна.
Повернув свою ладонь, он направил её к земле. Применив технику, его тело в мгновение ока притянулось к земле.
Применив технику, его тело в мгновение ока притянулось к земле.
Ему пришлось снова применить технику «Руки Пустоты», чтобы избежать удара. Коснувшись земли, он тут же оттолкнулся от нее и снова ринулся вперед. В мгновенье он оказался в том радиусе атаки, в котором мог проявиться себя лучше всего.
Сяо Ян не был хорош в использовании какого-либо оружия, предпочитая использовать собственное тело для сражений. В случае контактного боя: кулаки, голова, локти, ноги… Каждая часть тела становилась смертельным орудием. До тех пор, пока он мог поддерживать свою огромную скорость, то мог создать целый шторм резких и быстрых атак за крайне малое время.
Как только он достиг учителя Жо Лин, выражение его лица стало бесстрастным.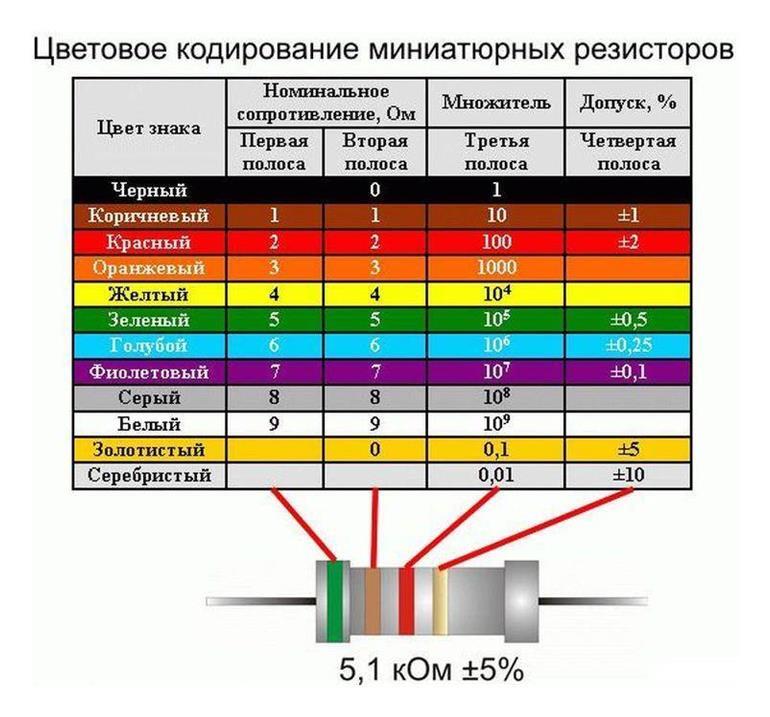 Его кулак, локоть и ноги быстро и яростно атаковали её, но каждый удар был с легкостью отбит.
Его кулак, локоть и ноги быстро и яростно атаковали её, но каждый удар был с легкостью отбит.
«Ладонь Останавливающее Сердце!»
«Пинок Раскалывающий Горы!»
«Удар Тяжелого Локтя!»
Получив возможность нанести быстрые атаки после стольких усилий, Сяо Ян применял все доступные ему техники Доу Ци, которые он изучил. Однако, ни одна из них не достигла цели.
Благодаря своему восприятию Сяо Ян понял, что тело учителя было будто бы покрыто слоем крема. Во время каждой атаки, часть тела, которой он атаковал, ускользала в сторону. Его усилия оказались пусты.
Во время каждой атаки, часть тела, которой он атаковал, ускользала в сторону. Его усилия оказались пусты.
Во время очередной атаки он встретился с глазами Жо Лин. Все его тело охватила дрожь… В тех глазах он увидел сплошную насмешку.
Заподозрив неладное, Сяо Ян хотел отпрыгнуть, однако его ноги оказались покрыты странной клейкой силой, не давая ему возможности переместиться.
Внезапное трудное положение заставило его сузить глаза и направить свой взгляд на учителя. Мельком увидев её ликующее лицо, он приготовился. Потеряв возможность двигаться, он напряг свой кулак. Используя всю оставшуюся у него Доу Ци, он направил в учителя Жо Лин тяжелый удар.
«Октановый Взрыв!»
Мысленно произнося название техники, на кулаке Сяо Яна вздулись вены. Бледная желтая энергия Доу Ци окутала кулак. И, наконец, рассекая ветер, разрушительный кулак ударил в учителя Жо Лин.
Бледная желтая энергия Доу Ци окутала кулак. И, наконец, рассекая ветер, разрушительный кулак ударил в учителя Жо Лин.
Внезапно усиленная атака Сяо Яна вызвала вспышку удивления в глазах учителя. Повернув руку, она создала компактный циклон водной энергии на своей ладони и приготовилась встретить удар Сяо Яна.
«Бэнг!»
Громоподобный звук взрыва окутал всю площадь, заставляя всю аудиторию озираться по сторонам.
Кулак и ладонь оставались в контакте всего несколько мгновений, после чего учитель Жо Лин сделала несколько шагов назад. Её лицо озарила улыбка, «Кажется, тебе не удастся получить отпуск.»
Её лицо озарила улыбка, «Кажется, тебе не удастся получить отпуск.»
Только после того, как его тело пару раз тряхнуло, Сяо Ян смог полностью отойти от шока, вызванного столкновением. Он опустил голову и бросил взгляд на ноги. Он осознал, что неосознанно сделал шаг в водную ловушку, созданную её водяным кнутом.
«Неудивительно, что она ничего не предпринимала во время моей атаки. Она пыталась заставить меня попасть в эту ловушку…» Обдумав произошедшее, он понял цель учителя. Она пыталась придумать метод остановить его молниеносную скорость, которой он так гордился.
«А это женщина не так проста…» Сяо Ян пытался освоюодить свою ногу, используя всю свою силу. Однако, с его текущим уровнем, мог ли он выбраться из ловушки, так тщательно подготовленной Великим Мастером.
«Хаха, Сяо Ян, все кончено. Это наш последний раунд!»
Смотря на непонятливое выражение лица Сяо Яна, Жо Лин рассмеялась. Она подняла свою свободную руку и заставила синий хлыст кольцом покрыть её.
Ладонью она держала закрытым рот змеи на кончике своего хлыста. Открыв рот, она стала вливать темно-синюю энергию Доу Ци в свое оружие.
Огромная синяя энергия заструилась в небо, словно водяной фонтан. Спустя секунду, собранная энергия преобразовалась в гигантскую водяную змею, длиной три или четыре метра. Змея уставилась в небо и испустила беззвучный рев. Большие капли воды отделялись от тела, проливаясь на всю площадь.
Большие капли воды отделялись от тела, проливаясь на всю площадь.
После рева, водная змея, управляемая учителем Жо Лин, излучая ужасающую силу, устремилась на неподвижного Сяо Яна.
Разглядывая гигантскую змею, появившуюся в воздухе, все кто её увидел, удивленно вскрикнули.
«Доу Сюань, средний уровень: Водяной Питон?»
«О боже, учитель использовала эту технику? Кажется Сяо Яну будет очень больно.» Удивленно произнесла Сяо Юй и бросила сочувственный взгляд на неподвижного Сяо Яна.
«Учитель пытается показать ему кто тут главный. С его неуправляемым характером, даже ей будет трудно в будущем дисциплинировать его, конечно, если она сможет его напугать.» Сяю Юй беспомощно вздохнула. Она вполне могла увидеть намерения Учителя.
С его неуправляемым характером, даже ей будет трудно в будущем дисциплинировать его, конечно, если она сможет его напугать.» Сяю Юй беспомощно вздохнула. Она вполне могла увидеть намерения Учителя.
Кроме того, что учитель использовала технику среднего уровня Сюань, Сяо Юй не особо волновалась. Она знала, что Жо Лин не хочет сильно навредить ему. Конечно, учитывая ее силу, её техника «Водного Питона» не была такой уж и слабой.
В академии, Сяо Юй улыбнулась удача и она уговорила учителя показать «Водного Питона» во всю её силу. Змея созданная Доу Ци оказалась семи или восьми метров в длину, что намного превышает эту малую версию.
Холодно смотря на проблемы Сяо Яна, уголки рта Ло Бу сформировали злорадную ледяную ухмылку. Он надеялся, что Сяо Ян встретит свои конец под атакой учителя Жо Лин.
Он надеялся, что Сяо Ян встретит свои конец под атакой учителя Жо Лин.
Гигантская змея падала на Сяо Яна. Чудовищный порыв ветра прижал одежды парня к телу.
Смотря на огромную силу, Сяо Ян беспомощно вздохнул. Сила Великого Мастера была безусловно чудовищной. Жо Лин не использовала и половины своей силы,в то время как он исчерпал свою.
Постепенно поднимая голову, он столкнулся лицом со змеей, появившейся в заходящих лучах солнца. Закрыв глаза и посмеявшись про себя он прошептал,» Ох, Яо Лао, помоги мне. Великий Мастер это не тот противник, с которым я могу справиться со своей текущей силой.»
Читайте ранобэ Расколотая битвой синева небес на Ranobelib. ru
ru
«Хаха, маленький мальчик. Ты, наконец-то понял свой текущий уровень. В глазах таких сильных людей ты словно ничто. Ты сделал только один шаг на пути становления сильным.» Произнес глубокий, мудрый голос в душе Сяо Яна.
«Действительно, очень сильна…»
Сяо Ян кивнул и сжал свои кулак. Его взгляд прошел через змею и уставился на мило улыбающуюся женщину, стоящую на расстоянии.» Но я верю, что в будущем я стану намного сильнее, чем она.»
«Бум!»
Гигантская змея, наконец, достигла Сяо Яна. Произошел взрыв, и бесчисленные капли воды брызнули в небо.
Наблюдая за областью столкновения питона с Сяо Яном, где все было скрыто от глаз водной завесой, учитель Жо Лин улыбнулась. Учитывая силу, с которой она атаковала, Жо Лин была уверена, что её должно было хватить, чтобы одолеть этого юношу.
«Юй Ер, пожалуйста, позаботься о нем. Нахождение в воде очень негативно скажется на…» Склонив голову, учитель попросила Сяо Юй, но перед тем как она успела закончить фразу, её лицо резко изменилось. Повернувшись обратно, она уставилась своими красивыми глазами на место столкновения воды и тела. Слабый туман распространился по всей площади. Внутри него были слышны легкие шаги и молодой человек неспешно вышел из тумана. Явив себя взору всем наблюдавшим, он остановился перед удивленной фигурой учителя. Рассмеявшись он произнес, «Учитель Жо Лин, мои извинения, но, кажется, вы не сможете не отпустить меня на годовой отпуск…»
Увидев улыбающееся лицо молодого человека, все были просто ошеломлены.
Наблюдая за одеждами молодого человека, которые не показывали и намека на то, что были атакованы водой, учитель Жо Лин медленно отошла от шока. Изучив его снова, она мягко произнесла, «Юноша, кажется, ты обладаешь еще какой-то способностью. Я недооценила тебя.»
«Ха-ха, мне просто повезло. Если бы Учитель использовала всю свою силу, я бы не продержался и трех раундов.» Покачал головой Сяо Ян.
«Если бы я использовала всю свою силу против нового студента с уровнем Ученика с четырьмя звездами, то как бы на меня смотрели в академии?» Ответила Жо Лин недовольным голосом, кинув на парня пренебрежительный взгляд.
«Так как вы выполнили мое условие, я гарантирую вам отпуск на год. Ох..» Беспомощно покачала Жо Лин. Было понятно, что хоть он и добился своего, она все еще не хотела его отпускать.
Ох..» Беспомощно покачала Жо Лин. Было понятно, что хоть он и добился своего, она все еще не хотела его отпускать.
«Ха-ха. Спасибо вам за помощь, учитель Жо Лин.» Сяо Ян поблагодарил Жо Лин за одобрение. Он был счастлив.
«Эх, другие люди хотят провести как можно больше времени в нашей академии, но ты, маленький поганец, захотел взять такой длинный отпуск? Ты действительно знаешь, как озадачить. После нашего возвращения, я займусь решением всех вопросов.» Возмущенно сообщила Жо Лин, смотря на возбужденную физиономию парня.
Сяо Ян показал восхитительную улыбку, однако промолчал. Касательно решения этого вопроса, он уже сделал то, что требовалось.
«Хорошо, давайте закончим на сегодня. В следующие семь дней мы продолжим набирать новых студентов.» Видя, что Сяо Ян не имел никакого желания объясняться, она вернула свое оружие обратно.
В следующие семь дней мы продолжим набирать новых студентов.» Видя, что Сяо Ян не имел никакого желания объясняться, она вернула свое оружие обратно.
Сяо Ян кивнул. Когда большинство дел в этом городе было закончено, ему потребовалось бы два или три дня для приготовлений к путешествию с Яо Лао.
Наблюдая, как учитель уходит, Сяо Юй схватила ее за руку и улыбаясь спросила, «Учитель, пока вы еще в городе Ву Тан, не хотите ли остановиться в Клане Сяо?»
«Приглашаешь пойти в Клан Сяо?»
Слегка удивившись, учитель Жо Лин слегка нахмурилась и спросила с беспокойством, «Для академии Цзя Нан уже выделено место для ночлега. И к тому же, если я приму это приглашение, не будет ли это слишком хлопотно?»
И к тому же, если я приму это приглашение, не будет ли это слишком хлопотно?»
«Ха-ха, никаких проблем. Это будет честь для нас, принять учителя из академии Цзя Нан. Я думаю все в клане будут рады видеть учителя в качестве гостя.» Усмехнулся Сяо Ян и шагнул вперед.
Став известной академией на континенте Доу Ци, влияние Академии Цзя Нан в империи Джи Ма было крайне велико. Если сравнивать её силу и возможности, то даже Главный Аукционный Дом уступал им.
Обладая огромной властью, академия Цзя Нан была снисходительна к малым силам в городе Ву Тан. Из-за этого отношения, академия слабо взаимодействовала с населением города ,еще меньше принимая приглашения посетить тот или иной клан.
При таких обстоятельствах, различные группы имели представления о своих возможностях. Они осознавали пропасть между ними. Мог ли слабый приглашать сильного и при этом надеяться на вежливость с его стороны?
Держа все это в уме, никакая сила в городе Ву Тан не была настолько сумасшедшей, чтобы пренебрежительно относиться к академии. Поэтому, когда группа от академии посещала их город, они держали дистанцию. Они не посмели бы их обидеть, но и не пытались к ним подлизаться, чтобы в итоге не стать целью насмешек.
Сяо Ян прожил в городе более десяти лет, поэтому естественно, что он понимал, что группа академии Цзя Нан вела себя отстраненно. Если учитель Жо Лин остановится в их доме, то это существенно поднимет влияние его клана на территории города и, возможно, оно будет не слабее, чем у Главного Аукционного Дома.
С особым статусом учителя Жо Лин, до тех пор, пока она благоприятно высказывается в сторону какой-либо организации, неважно насколько слабо будет её впечатление, на следующий день группе из академии следует ожидать повышенного количества посетителей.
Может это и преувеличенно, но если все пройдет гладко, учитель может повлиять на то, кто поедет в академию Цзя Нан, а кто нет. Академия не давала ни шанса тем, кто не подходит им по требованиям.
Поэтому, до тех пор, пока Учитель Жо Лин будет оставаться в Клане Сяо по приглашению Сяо Юй, каждая сила в городе будет стараться выражать доброжелательность клану. После их удачных продаж исцеляющей медицины некоторое время назад и имея благосклонность учителя, никто и не подумает о причинах, почему клан Сяо так возвысился.
Просто временное пребывание учителя на территории клана Сяо, предоставит им ряд преимуществ. И это была малая часть того, почему Сяо Ян рекомендовал ей посетить их клан.
Услышав приглашение от Сяо Юй, Жо Лин промолчала. По своему опыту, она знала, какие последствия произойдут в городе. Исходя из здравого смысла, учителя в прошлом всячески избегали подобных приглашений.
Однако, когда Сяо Ян лично её пригласил, ей было сложно отказать. Столкнувшись с этим студентом, который обладал самым чудовищным талантом за последние сто лет существования академии, они просто не могла проигнорировать его слова. Кроме того, если этот парень разозлится и откажется поступать, ей будет трудно снова отыскать другого студента, похожего на него.
Учитель Жо Лин нахмурилась, и что-то себе надумав, наконец, кивнула и сказала, улыбаясь, «Хорошо, следующие несколько дней я буду проводить в Клане Сяо.»
Увидев, как учитель соглашается, Сяо Юй бросилась к ней и крепко обняла её тонкую талию.
«Ло Бу, вы вернетесь с Гэ Ла и остальными в положенное место. Завтра мы продолжим здесь. Обращаю ваше внимание – не смейте создавать проблемы!» Отдала указания учитель Жо Лин, обнимая голову Сяо Юй.
«Хм.»
С угрюмым лицом, Ло Бу кивнул, смотря в спины группе веселящихся и смеющихся девушек, которые постепенно отдалялись от них. Он мог только беспомощно покачать головой. Без приглашения Сяо Юй, никто из них не мог последовать за ними. Поэтому, группа студентов мужчин проследовали к внешней части площади, без компании красавиц.
Он мог только беспомощно покачать головой. Без приглашения Сяо Юй, никто из них не мог последовать за ними. Поэтому, группа студентов мужчин проследовали к внешней части площади, без компании красавиц.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 — 4 с/дел (190-504, модель 500 МГц)
— 4 с/дел (190-504, модель 500 МГц)
VME-103/104, VME-Exi-105 (4-20мА+HART) | НПО «Вакууммаш» – Ижевск
ТУ 26. 51.43-003-24481731-2018
51.43-003-24481731-2018
Особенности
Конструктивно могут изготавливаться в корпусе для установки в головку первичного преобразователя либо в корпусе для щитового монтажа на рейку по ГОСТ Р МЭК 60715-2003. Преобразователи могут комплектоваться программным обеспечением и коммуникатором для настройки.
| Исполнение для установки в головку VME-103/104-3, VME-Exi-105-3 |
Исполнение для щитового монтажа VME-103/104-4, VME-Exi-105-4 |
Технические характеристики
| Напряжение питания для общепромышленного применения, В | 12…36 |
| Потребляемая мощность для общепромышленного применения, Вт, не более | 0,8 |
| Напряжение питания для искробезопасной цепи, В | 12…24 |
| Потребляемая мощность для общепромышленного применения, Вт, не более | 0,75 |
| Стандартный диапазон температур окружающей среды, °С | -40…+70 |
| Стандартный диапазон температур окружающей среды для общепромышленного применения, °С | -60…+85 |
| Версия протокола HART | 7. 2 2 |
Метрологические характеристики для исполнений с классом точности 0,05 и 0,1
| Тип ПП | Максимальный диапазон измерени, °С |
Пределы допускаемой основной приведенной погрешности (при температуре окружающей среды 20±5 °С, от максимального диапазона измерений) в ницаервале измерений меньшенентов |
|
|---|---|---|---|
| Класс точности 0,05 | Класс точности 0,1 | ||
| К | -200…+1300 | 0,05 % | 0,1 % |
| L | -200…+800 | ||
| N | -270…+1300 | ||
| J | -200…+900 | ||
| S | 0…+1600 | ||
| B | 300…+1800 | ||
| R | 0…+1600 | ||
| Напряжение | -15…+70 мВ | ||
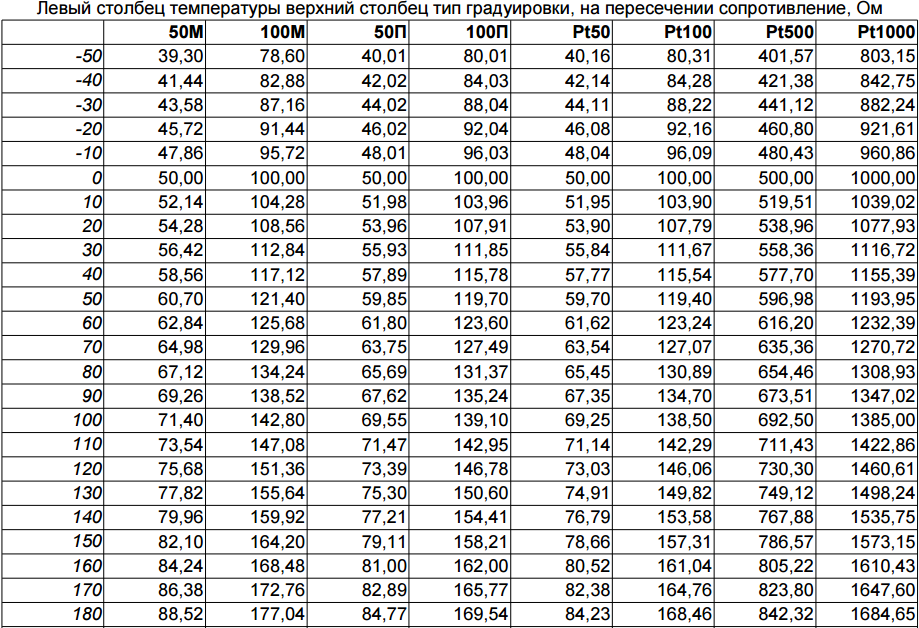
| 50М | -180…+200 | ||
| 100М | -180…+200 | ||
| 50П | -200…+850 | ||
| 100П | -200…+850 | ||
| Pt100 | -200…+850 | ||
| Pt500 | -200…+850 | ||
| Pt1000 | -200…+850 | ||
| Сопротивление | 1…400 Ом | ||
| Сопротивление | 1…4000 Ом | ||
| Потенциометр | 40…400 Ом | ||
| Потенциометр | 400…4000 Ом | ||
Примечание – допускается изготовление ПИ с диапазонами измерений, отличными от приведенных в таблице, но не превышающих нижней и верхней границы приведенных диапазонов. |
|||
| Подключение первичных преобразователей, внешнего датчика «холодного спая» и выходных цепей к преобразователям VMЕ в корпусе для установки в головку первичного преобразователя |
| Подключение первичных преобразователей и выходных цепей к преобразователям VMЕ в корпусе для щитового монтажа
Примечание – символ «x» в описании клемм представляет количество каналов. У одноканального исполнения символ «х» заменяется цифрой 1, у двухканального обозначает номер каналов: 1 или 2. |
|
Пример обозначения при заказе
| VM-Exi-105 | – | 3 | – | HART | – | (-60…+85) | – | K/2 | – | (-50/400) | – | 0,1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
-
- Обозначение модификации
- Конструктивное исполнение:
3 – в пластиковом корпусе для установки в головку ПП
4 – в пластиковом корпусе для щитового монтажа - Обозначение типа протокола передачи данных
- Обозначение температурного диапазона окружающей среды.
 При отсутствии обозначения принимается диапазон от минус 60 °С до плюс 85 °С.
При отсутствии обозначения принимается диапазон от минус 60 °С до плюс 85 °С. - НСХ первичного преобразователя (а также схема подключения для термопреобразователей сопротивления)
- Диапазон измерения
- Класс точности.
Подстроечные резисторы и потенциометры
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Цены в формате
. pdf,
.xls pdf,
.xls
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
- Функциональная характеристика подстроечного резистора…………………………….. А (линейная)
- Номинальная мощность подстроечного резистора при 70°С…………………………… 0,125 Вт
- Максимальное рабочее напряжение подстроечного резистора (постоянное)……..200 В
- Диапазон рабочих температур подстроечного резистора………………………………….-55° +125°С
- Температурный коэффициент сопротивления подстроечного резистора……………100 ppm/°С
- Допустимое отклонение номинала подстроечного резистора………………..
 …………. ± 20 %
…………. ± 20 % - Оборот подстроечного резистора…………………………………………………………………….. 250°
Подстроечные резисторы производства японской фирмы Nidec отличаются высокой надежностью и стабильным качеством. Резистор переменного сопротивления защищен от попадания влаги на резистивный слой при отмывки. Для этого в конструкции переменного резистора установлено резиновое кольцо препятствующее проникновению жидкости и парообразных остатков флюса между ротором и статором.
Технические характеристики и маркировка подстроечных потенциометров Nidec ST32 для поверхностного монтажа
Производитель — NIDEC.
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||
Цены в формате
. pdf,
.xls pdf,
.xls
|
||||||||||||||||||||||||||||||||||||||||||||||||||
| Маркировка Murata | Маркировка Bourns |
| PVZ3A201A01R00 | TC33X-1-201E |
| PVZ3A501A01R00 | TC33X-1-501E |
| PVZ3A102A01R00 | TC33X-1-102E |
| PVZ3A152A01R00 | TC33X-1-202E |
| PVZ3A502A01R00 | TC33X-1-502E |
| PVZ3A103A01R00 | TC33X-1-103E |
| PVZ3A153A01R00 | TC33X-1-203E |
| PVZ3A203A01R00 | TC33X-1-203E |
| PVZ3A503A01R00 | TC33X-1-503E |
| PVZ3A104A01R00 | TC33X-1-104E |
| PVZ3A105A01R00 | TC33X-1-104E |
- Функциональная характеристика подстроечного потенциометра.
 ……………………………… А (линейная)
……………………………… А (линейная) - Номинальная мощность подстроечного потенциометра при 70°С…………………………….. 0,1 Вт
- Максимальное рабочее напряжение подстроечного потенциометра (постоянное)………50 В
- Диапазон рабочих температур подстроечного потенциометра………………………………….-25°С +85°С
- Температурный коэффициент сопротивления подстроечного потенциометра……………±500 ppm/°С
- Допустимое отклонение номинала подстроечного потенциометра……………………………. ± 30 %
- Оборот подстроечного потенциометра…………………………………………………………………….. 250°
Серия PVZ3 является наиболее универсальной, одинаково подходящей для использования как в цветных ЖК-мониторах, проигрывателях DVD, системах проводной и безпроводной связи, так и в малошумящих усилителях и т. д. Универсальность этой серии обеспечивается наличием широкого диапазона сопротивлений, способом монтажа (SMD) и габаритными размерами корпуса.
д. Универсальность этой серии обеспечивается наличием широкого диапазона сопротивлений, способом монтажа (SMD) и габаритными размерами корпуса.
Технические характеристики и маркировка подстроечных потенциометров Murata PVZ3A для поверхностного монтажа
Производитель — MURATA
переменный%20резистор%20104 спецификация и примечания по применению
 org/Product»>
org/Product»> переменная%20резистор%20104 Datasheets Context Search
| Каталог данных | MFG и тип | ПДФ | Теги документов |
|---|---|---|---|
Андрей FSJ4-50B
Аннотация: L4NM
|
OCR-сканирование |
ФСДЖ1-50А ФСДЖ2-50 Мужской/7-16 ФСДЖ4-50Б ЛДФ4П-50А-1, Эндрю FSJ4-50B L4NM | |
2001 — ЛМ117АК
Реферат: LM117.TO39 TO276AB LM117H-883B 60v регулятор напряжения LM117SMD 60v выходной регулятор
|
Оригинал |
ЛМ117″ ЛМ117 O39-8QR-B O66-8QR-B ЛМ117-220ФМ ЛМ117-220М ЛМ117-220М-8QR-Б ЛМ117АК ЛМ117.ТО39 ТО276АБ ЛМ117Х-883Б регулятор напряжения 60В LM117SMD Регулятор выхода 60В | |
7703401yx
Резюме: 7703407XX 7703402TX 7703405UX BS-94 F0436 IP117AG-DESC IP117MAHVH-883B
|
OCR-сканирование |
IP1060AD IP1060AJ IP1060AN IP1060BJ ИП1060БДЖ-883Б IP1060D IP1060J IP1060N IP117AG ИП117АГ-883Б 7703401yx 7703407ХХ 7703402TX 7703405UX БС-94 F0436 IP117AG-DESC ИП117МАХВХ-883Б | |
2010 — 5SGX
Аннотация: 16-битный умножитель 16-битный сумматор COMPRESSOR PLUG 16-битный сумматор с выбором переноса с использованием быстрых сумматоров
|
Оригинал |
СВ51004-1 5SGX 16-битный множитель 16-битный сумматор ЗАГЛУШКА КОМПРЕССОРА перенос выбора сумматора 16 бит с использованием быстрых сумматоров | |
2000 — 60В вход 15В выход регулятор
Реферат: для регулятора lm137 Высоковольтный отрицательный регулятор отрицательный РЕГУЛЯТОР НАПРЯЖЕНИЯ 3,7 В LM137AK 60-вольтовый регулятор напряжения LM137LCC4 LM137SMD05 Отрицательный РЕГУЛЯТОР 500MA регулятор напряжения
|
Оригинал |
ЛМ137-220М ЛМ137СМД9 O276AB) ЛМ137СМД05 LM137SMD05-8QR-B LM137SMD-8QR-B О276АА) 60В вход 15В выход регулятор для регулятора lm137 Отрицательный регулятор высокого напряжения отрицательный РЕГУЛЯТОР НАПРЯЖЕНИЯ 3,7 В ЛМ137АК регулятор напряжения 60В LM137LCC4 Отрицательный РЕГУЛЯТОР 500MA регулятор напряжения | |
2001 — ИП137АСМД05
Реферат: LM337HV IP337HVSMD05-8QR-B IP337K IP137AIG
|
Оригинал |
IP137A IP337A LM137A O276AB) О276АА) IP137ASMD05 LM337HV IP337HVSMD05-8QR-B IP337K IP137AIG | |
2000 — Регулируемый регулятор положительного напряжения vin 60V
Резюме: IP317 60v вход 15v регулятор выхода TO66 PLASTIC package IP117HVG-BSS2 ip117ig
|
Оригинал |
IP117, IP117A IP317, IP317A ЛМ117, LM117A СМД05 Регулируемый регулятор положительного напряжения vin 60V IP317 60В вход 15В выход регулятор TO66 ПЛАСТИКОВЫЙ пакет IP117HVG-BSS2 ip117ig | |
СТ72
Аннотация: 00A1 009F 00a6
|
Оригинал |
||
1998 — 009F
Реферат: 00A1 ОБУЧЕНИЕ МИКРОКОНТРОЛЛЕРОВ режимы адресации
|
Оригинал |
||
СТ-РЕАЛАЙЗЕР
Резюме: 009F 00A1 PC128 PC127 JRNC
|
Оригинал |
||
2010 — Недоступно
Резюме: нет абстрактного текста
|
Оригинал |
СВ51004-1 | |
ф0114
Реферат: отрицательный регулятор напряжения lm117ahvk BS-94 положительный отрицательный регулятор напряжения
|
OCR-сканирование |
IP79M12AH-BSS2 БС9430 F0646 -55к 5962-8874701ХХ IP79M12AH-DESC T0220 IP79М12АХ-СМ IP79M12AHLCC4 f0114 отрицательный РЕГУЛЯТОР НАПРЯЖЕНИЯ лм117ахвк БС-94 положительно-отрицательный регулятор напряжения | |
MC9S12DC128
Аннотация: 30F5013 MC9S12DG64 30f2010 30F6013 30f5011 30f4011 30f3011 30f4013 30f4011 захват
|
Оригинал |
16-битный ADSP2100A ADSP2101 ADSP2103 ADSP2104 ADSP2105 ADSP2109 ADSP2111 ADSP2115 ADSP2161 MC9S12DC128 30F5013 MC9S12DG64 30f2010 30F6013 30f5011 30f4011 30f3011 30f4013 30f4011 захват | |
Недоступно
Резюме: нет абстрактного текста
|
Оригинал |
СВ51004 18-сумматор | |
ПИ-МБУС-300
Реферат: modicon OPTO-22 daniel библиотека протокола modbus modbus modbus wiegand контроллер opto 22 протокол modbus код wiegand
|
Оригинал |
800-321-ОПТО 800-832-ОПТО опто22 800-ТЭК-ОПТО ПИ-МБУС-300 модикон ОПТО-22 Даниэль Modbus протокол библиотека Modbus модбус Виганд опто 22 контроллер протокол Modbus код Виганда | |
Ф0513
Резюме: отрицательный РЕГУЛЯТОР НАПРЯЖЕНИЯ 7703406TX F0814
|
OCR-сканирование |
ИП120Р-15 ИП120Р-15-883Б ИП120Р-15-БСС2 ИП123АГ-05 ИП123АГ-05-883Б ИП123АГ-05-БСС2 ИП123АГ-12 ИП123АГ-12-883Б ИП123АГ-15 ИП123АГ-15-883Б F0513 отрицательный РЕГУЛЯТОР НАПРЯЖЕНИЯ 7703406TX F0814 | |
2000 — 00FF
Реферат: режимы адресации 009F 00A1 микроконтроллера ОБУЧЕНИЕ МИКРОКОНТРОЛЛЕРОВ
|
Оригинал |
||
Ф0128
Реферат: РЕГУЛЯТОР ОТРИЦАТЕЛЬНОГО НАПРЯЖЕНИЯ BS-94 LM137aK LM140K-12/883B
|
OCR-сканирование |
ЛМ120Х-15-БСС2 ЛМ120К-05 ЛМ120К-05-БСС2 ЛМ120К-12 ЛМ120К-12-БСС2 ЛМ120К-15 ЛМ120К-15-БСС2 ЛМ120МАХ-05 ЛМ120МАХ-12 ЛМ120МАХ-15 F0128 отрицательный РЕГУЛЯТОР НАПРЯЖЕНИЯ БС-94 ЛМ137аК ЛМ140К-12/883Б | |
2001 — ТО276АА
Реферат: IP317 60v Регулятор выходного напряжения Регулятор напряжения SMD0
|
Оригинал |
IP317″ IP317AG ИП317АХВК IP317AHVSMD IP317AHVSMD05 IP317AHVSMD05-8QR-B IP317AHVSMD-8QR-B ИП317АК IP317AR IP317ASMD ТО276АА IP317 Регулятор выхода 60В SMD0 регулятор напряжения | |
2001 — опто s52
Реферат: nokia n8 2832 eeprom schneider HDD ручные турбинные расходомеры на основе gsm беспроводная доска объявлений расходомер Вентури ПИД-управление nokia c5 wiegand конвертер
|
Оригинал |
1301-060308—МАРТ, 800-321-ОПТО 800-832-ОПТО опто22 800-ТЭК-ОПТО CRC-16 опто s52 нокиа н8 2832 ЭСППЗУ Шнайдер жесткий диск ручные турбинные расходомеры Беспроводная доска объявлений на базе GSM расходомер Вентури ПИД-регулятор нокиа с5 преобразователь виганда | |
2010 — АН494
Аннотация: Прошивка IR 8500 AN366 Si1120 ИНФРАКРАСНЫЙ ДАТЧИК
|
Оригинал |
АН494 АН494 Прошивка ир 8500 АН366 Si1120 ИНФРАКРАСНЫЙ ДАТЧИК | |
2000 — АСМ 1042
Аннотация: MB89PV620
|
Оригинал |
СМ25-00320-1Е fcc896 fcc896 fcc896. АСМ 1042 МБ89ПВ620 | |
2010 — герконовый датчик
Реферат: Allen-Bradley Micrologix генератор последовательности pn с использованием jk-триггера Allen-Bradley slc 500 Allen-Bradley slc 150 F030 CRC-16 Square D от Schneider Electric ПИД-управление Allen-Bradley slc 500 modbus
|
Оригинал |
1701-101122 — ноябрь 800-321-ОПТО 800-832-ОПТО опто22 800-ТЭК-ОПТО CRC-16 язычковый датчик Аллен-Брэдли микрологикс генератор последовательности pn с использованием триггера jk Аллен-Брэдли slc 500 Аллен-Брэдли slc 150 F030 Square D от Schneider Electric ПИД-регулятор Allen-Bradley slc 500 Modbus | |
7703403ХХ
Реферат: F0548 7703403YX IP140AG
|
OCR-сканирование |
IP137K ИП137К-883Б ИП137К-БСС2 IP137K-DESC IP137MAH ИП137МАХ-883Б IP137MAH-BSS2 IP137MAH-DESC IP137MAH-LCC4 IP137MAHVH 7703403ХХ F0548 7703403YX ИП140АГ | |
Недоступно
Резюме: нет абстрактного текста
|
Оригинал |
РС485 РС485 | |
 org/Product»>
org/Product»>  org/Product»>
org/Product»>  org/Product»>
org/Product»>  org/Product»>
org/Product»>  org/Product»>
org/Product»>  org/Product»>
org/Product»>  org/Product»>
org/Product»>  org/Product»>
org/Product»>  org/Product»>
org/Product»> Предыдущий 1 2 3 … 23 24 25 Next
Платиновый регистратор данных сопротивления PT-104
Платиновый регистратор данных сопротивления PT-104
- Измеряет температуру, сопротивление и напряжение
- Высокое разрешение и точность
- Работает с датчиками PT100 и PT1000
- Поддерживает 2-, 3- и 4-проводные датчики
- Источник питания не требуется
- Интерфейсы USB и Ethernet
- Запуск нескольких устройств на одном ПК
Хотя точные датчики температуры широко доступны, их использование затруднено из-за ошибок, вызванных измерительным устройством. Однако ваш PT-104 по своей сути точен благодаря своей инновационной конструкции. Вместо того, чтобы полагаться на источники опорного напряжения, которые, как правило, чувствительны к температуре, он использует эталонные резисторы, которые чрезвычайно стабильны с низким температурным коэффициентом и дрейфом. Точное значение каждого резистора хранится в EEPROM , чтобы обеспечить максимальную точность (рекомендуется ежегодная повторная калибровка). Для достижения разрешения 0,001 °C используется высокопроизводительный 24-битный АЦП.
Однако ваш PT-104 по своей сути точен благодаря своей инновационной конструкции. Вместо того, чтобы полагаться на источники опорного напряжения, которые, как правило, чувствительны к температуре, он использует эталонные резисторы, которые чрезвычайно стабильны с низким температурным коэффициентом и дрейфом. Точное значение каждого резистора хранится в EEPROM , чтобы обеспечить максимальную точность (рекомендуется ежегодная повторная калибровка). Для достижения разрешения 0,001 °C используется высокопроизводительный 24-битный АЦП.
Температура
PT-104 измеряет температуру с помощью платиновых термометров сопротивления (ПТС). Поддерживаются оба общепринятых отраслевых стандарта (PT100 и PT1000). Устройство совместимо с двух-, трех- и четырехпроводными датчиками (для точных измерений рекомендуется использовать четырехпроводные датчики PT100). Широкий диапазон датчиков PT100 доступен для использования с вашим PT-104.
Напряжение
Для измерения напряжения каждый входной разъем можно рассматривать как дифференциальный вход с заземлением или как два несимметричных входа. На обоих входах должно быть 0 вольт или выше, хотя не имеет значения, на каком из входов напряжение выше.
На обоих входах должно быть 0 вольт или выше, хотя не имеет значения, на каком из входов напряжение выше.
Доступны два диапазона напряжения (от 0 до 115 мВ и от 0 до 2500 мВ). Для наиболее точных измерений используйте диапазон от 0 до 2500 мВ.
Сопротивление
При измерении сопротивления PT-104 использует четырехпроводную схему для обеспечения максимально возможной точности. Доступны два диапазона сопротивления (от 0 до 375 Ом и от 0 до 10 000 Ом). Устройство откалибровано в диапазоне от 0 до 375 Ом, поэтому этот диапазон следует использовать для точных измерений.
Одноканальный адаптер с винтовыми клеммами
Подключение датчиков сторонних производителей, не оснащенных 5-контактным разъемом mini-DIN, или измерение напряжения или сопротивления возможно с помощью адаптера с винтовыми клеммами PP660. Этот недорогой адаптер имеет 4 разъема и занимает один канал PT-104.
Программное обеспечение
Регистратор данных PT-104 поставляется с программным обеспечением для регистрации данных PicoLog 6. Это мощное, но гибкое программное обеспечение для сбора данных позволяет собирать, обрабатывать, анализировать, отображать и экспортировать данные.
Это мощное, но гибкое программное обеспечение для сбора данных позволяет собирать, обрабатывать, анализировать, отображать и экспортировать данные.
Разработанный с нуля, чтобы быть интуитивно понятным с самого начала, PicoLog 6 позволяет вам настроить регистратор и начать запись всего несколькими щелчками мыши, независимо от вашего уровня опыта регистрации данных.
Отсюда начать запись просто: подключите логгер, добавьте канал, нажмите «Запись» и вы записываетесь! PicoLog 6 также включает в себя ряд дополнительных функций для удовлетворения более сложных потребностей в регистрации данных.
Также можно загрузить PicoSDK, комплект для разработки программного обеспечения. SDK содержит ряд программных драйверов и примеры кода, которые можно использовать для написания собственного программного обеспечения или для использования регистратора данных PT-104 со сторонним программным обеспечением, таким как MATLAB, LabVIEW, C, C++, C# и Python.
Но мне нужно измерить больше каналов…
PT-104 может измерять до 4 каналов одновременно. Нужно больше каналов? Без проблем. Используя PicoLog, вы можете подключить до 20 PT-104 к одному ПК, что дает вам потенциальный 80-канальный регистратор данных PT100. В качестве альтернативы вы можете использовать свой регистратор PT-104 с 19 другими регистраторами данных Pico, такими как многоканальный регистратор данных PicoLog 1000, для создания собственной пользовательской системы сбора данных.
Нужно больше каналов? Без проблем. Используя PicoLog, вы можете подключить до 20 PT-104 к одному ПК, что дает вам потенциальный 80-канальный регистратор данных PT100. В качестве альтернативы вы можете использовать свой регистратор PT-104 с 19 другими регистраторами данных Pico, такими как многоканальный регистратор данных PicoLog 1000, для создания собственной пользовательской системы сбора данных.
Непревзойденная портативность
Имея интерфейсы USB и Ethernet, PT-104 можно использовать в различных ситуациях. Если вам нужно портативное решение, которое можно использовать в разных местах и быстро настроить и использовать, просто подключите свой ноутбук к PT-104 через USB. Нужно отслеживать ситуацию в течение нескольких часов или дней или из удаленного места? Подключите PT-104 к свободному порту в вашей сети, а затем получите к нему удаленный доступ либо из вашей локальной сети, либо через Интернет. Использование технологии Power over Ethernet (PoE) означает, что вам даже не нужен отдельный блок питания.
Когда точность имеет первостепенное значение, вам нужен PT-104
Платиновый регистратор данных сопротивления PT-104 обеспечивает высочайшее разрешение и точность. Вы можете использовать его для измерения температуры, напряжения и сопротивления. Наличие интерфейсов USB и Ethernet дает вам выбор способа подключения и питания PT-104, а также возможность удаленного доступа к PT-104 через локальную сеть или Интернет. Когда точность измерения температуры имеет первостепенное значение, PT-104 — это то, что вам нужно.
| Ввод/вывод | |||
|---|---|---|---|
| Тип | Температура | Сопротивление | Напряжение |
| Датчик | ПТ100, ПТ1000 | н/д | н/д |
| Диапазон | от –200 до +800 °С | от 0 до 375 Ом от 0 до 10 кОм |
от 0 до 115 мВ от 0 до 2,5 В |
| Погрешность (при 23 ±2 °C) | 0,015 °С + 0,01% от показания |
20 частей на миллион при 100 Ом | 0,4% |
| Температурный коэффициент | 5 частей на миллион/°C | 5 частей на миллион/°C | 100 частей на миллион/°C |
| RMS шум с фильтром | 0,01 °С | 10 частей на миллион | 10 частей на миллион |
| Разрешение | 0,001 °С | 1 мкОм | 0,156 мкВ |
| Защита от перегрузки | ±30 В | ||
| Количество входов | 4 | ||
| Разрешение преобразователя | 24 бита | ||
| Время преобразования | 720 мс на канал | ||
| Входные разъемы | 4-контактный мини-DIN | ||
| Входное сопротивление | >1 МОм | ||
| Выходные разъемы | USB и Ethernet | ||
| Требования к ПК | |||
|---|---|---|---|
| Минимум | Процессор : 1 ГГц Память : 512 МБ Свободное место на диске: 32-разрядная версия: 600 МБ, 64-разрядная версия: 1,5 ГБ Операционная система : 32- или 64-разрядная версия Microsoft Windows XP (SP3), Vista, Windows 7, Windows 8 (не Windows RT) или Windows 10 Порты : USB 1.  1 или Ethernet. 1 или Ethernet. |
||
| Рекомендуемый | Процессор : 1 ГГц Память : 512 МБ Свободное место на диске: 32-разрядная версия: 850 МБ, 64-разрядная версия: 2 ГБ Операционная система : 32- или 64-разрядная версия Microsoft Windows 7, Windows 8 (не Windows RT) или Windows 10 Порты : USB 1.1 (или выше) или Ethernet |
||
| Защита окружающей среды | |||
|---|---|---|---|
| Операционная среда Диапазон температур Диапазон влажности |
от 0 °C до 70 °C (от 20 °C до 30 °C для указанной точности) 20% до 90 % относительной влажности, без конденсации |
||
| Среда хранения Диапазон температур Влажность |
от -20 до +80°C относительная влажность от 5 до 95 %, без конденсации |
||
| Физические свойства | |||
|---|---|---|---|
| Размеры | 184 x 135 x 36 мм (примерно 5,31 x 7,24 x 1,42 дюйма) | ||
| Вес | 350 г (около 12,3 унции) | ||
| Программное обеспечение | |||
|---|---|---|---|
| ПикоЛог для Windows | Программное обеспечение для сбора данных PicoLog может собирать до 1 миллиона образцов. Несколько представлений — просмотр данных в виде графика, электронной таблицы или текста Масштабирование параметров — преобразование необработанных данных в стандартные инженерные единицы Математические функции — использование математических уравнений для расчета дополнительных параметров Пределы сигналов тревоги — программирование оповещения, если параметр выходит за пределы заданного диапазона IP-сеть — передача измерений по Поддерживаемые операционные системы — от Microsoft Windows XP (SP3) до Windows 10 (не Windows RT). |
||
| Языки PicoLog | Полная поддержка — English, Français, Deutsch Меню и диалоговые окна только для — Italiano, Español, Svenska |
||
| Комплект для разработки программного обеспечения | Включает пример кода. |
||
| Дополнительное бесплатное программное обеспечение | Драйверы для Linux и OS X. | ||
| Языки документации | |||
|---|---|---|---|
| Руководство пользователя Руководство программиста |
английский, французский, немецкий, итальянский, испанский Английский |
||
| Общий | |||
|---|---|---|---|
| Дополнительное оборудование (прилагается) | Кабель USB 2.0, кабель Ethernet, руководства пользователя, компакт-диск с программным обеспечением | ||
| Интерфейс ПК | USB 1. 1 (совместимость с USB 2.0 и USB 3.0) или Ethernet 1 (совместимость с USB 2.0 и USB 3.0) или Ethernet |
||
| Требования к питанию | Питание от порта USB или Ethernet | ||
| Соответствие | Европейские стандарты EMC и LVD Правила FCC, часть 15, класс A RoHS соответствует |
||
| Гарантия полного удовлетворения | В случае, если данный товар не полностью соответствует вашим требованиям, вы можете вернуть его для обмена или возврата денег. Чтобы подать заявку, товар должен быть возвращен в хорошем состоянии в течение 14 дней. | ||
| Гарантия | 5 лет | ||
Сальмонелла enterica Серотип Typhimurium DT 104 Резистентность к антибиотикам Геномный остров I в серотипе Paratyphi B
Emerg Infect Dis. 2002 апрель; 8(4): 430–433.
2002 апрель; 8(4): 430–433.
doi: 10.3201/eid0804.010375
, * , † , † , * , ‡ , § , * и *
Информация об авторе Информация об авторских правах и лицензии Отказ от ответственности
Мы идентифицировали Salmonella геномный остров I (SGI1) в изоляте Salmonella enterica серотипа Paratyphi B. Этот кластер генов устойчивости к антибиотикам, который обеспечивает множественную лекарственную устойчивость, был идентифицирован ранее. в S. enterica серотип Typhimurium фага типов DT 104 и DT 120 и в S. enterica серотип Agona.
Ключевые слова: Salmonella геномный остров I, Typhimurium DT 104, Paratyphi B
С множественной лекарственной устойчивостью Salmonella enterica серотип Typhimurium definitive phage type 104 (DT 104) за последнее десятилетие стал глобальной проблемой здравоохранения из-за своей связи с болезнями животных и человека ( 1 ). Штаммы этого типа фагов с множественной лекарственной устойчивостью были впервые идентифицированы у экзотических птиц в Соединенном Королевстве в начале 1980-х годов и у крупного рогатого скота и людей в конце 1980-х годов, но с тех пор стали обычным явлением у других видов животных, таких как домашняя птица, свиньи и овцы. Эпидемия DT 104 в настоящее время распространилась по всему миру, с несколькими вспышками с 1996 в США и Канаде ( 2 – 5 ).
Штаммы этого типа фагов с множественной лекарственной устойчивостью были впервые идентифицированы у экзотических птиц в Соединенном Королевстве в начале 1980-х годов и у крупного рогатого скота и людей в конце 1980-х годов, но с тех пор стали обычным явлением у других видов животных, таких как домашняя птица, свиньи и овцы. Эпидемия DT 104 в настоящее время распространилась по всему миру, с несколькими вспышками с 1996 в США и Канаде ( 2 – 5 ).
С множественной лекарственной устойчивостью S. Штаммы Typhimurium DT 104 обычно устойчивы к ампициллину, хлорамфениколу/флорфениколу, спектиномицину/стрептомицину, сульфонамидам и тетрациклинам. Гены устойчивости к антибиотикам сгруппированы в части геномного острова размером 43 т.п.н., называемого геномным островом I Salmonella (SGI1), между генами thdf и int2 хромосомы ( 6 – 10 ). Ген int2 является частью ретрона, обнаруженного только у серотипа Typhimurium ( 7 , 8 ). Ниже последовательности ретрона находится ген yidY , который также обнаружен в хромосоме других серотипов S. enterica ( 7 , 8 ). Кластер генов устойчивости к антибиотикам составляет примерно одну треть SGI1 и расположен на 3′-конце структуры ( 7 , 8 ).). Все гены устойчивости сгруппированы и заключены в скобки двумя интегронными структурами (). Первый интегрон несет ген aadA2 , придающий устойчивость к стрептомицину и спектиномицину, и укороченный ген устойчивости sulI . Второй содержит ген бета-лактамазы bla PSE-1 и полный ген sulI . Между этими двумя интегронными структурами находится ген floR ( 6 ), также называемый floSt .
( 11) или cmlA -подобный (9), который придает перекрестную устойчивость к хлорамфениколу и флорфениколу, а также гены устойчивости к тетрациклину tetR и tet (G). Устойчивость к флорфениколу и обнаружение гена floR с помощью методов, основанных на полимеразной цепной реакции (ПЦР), были предложены в качестве средства для быстрой идентификации полирезистентных штаммов S.
Ниже последовательности ретрона находится ген yidY , который также обнаружен в хромосоме других серотипов S. enterica ( 7 , 8 ). Кластер генов устойчивости к антибиотикам составляет примерно одну треть SGI1 и расположен на 3′-конце структуры ( 7 , 8 ).). Все гены устойчивости сгруппированы и заключены в скобки двумя интегронными структурами (). Первый интегрон несет ген aadA2 , придающий устойчивость к стрептомицину и спектиномицину, и укороченный ген устойчивости sulI . Второй содержит ген бета-лактамазы bla PSE-1 и полный ген sulI . Между этими двумя интегронными структурами находится ген floR ( 6 ), также называемый floSt .
( 11) или cmlA -подобный (9), который придает перекрестную устойчивость к хлорамфениколу и флорфениколу, а также гены устойчивости к тетрациклину tetR и tet (G). Устойчивость к флорфениколу и обнаружение гена floR с помощью методов, основанных на полимеразной цепной реакции (ПЦР), были предложены в качестве средства для быстрой идентификации полирезистентных штаммов S. Typhimurium DT 104 ( 11 ), поскольку фаговое типирование является доступны только в специализированных лабораториях. Однако недавно сообщалось о SGI1 в другом фаговом типе серотипа Typhimurium и в серотипе Agona, что предполагает горизонтальный перенос SGI1 (9).1048 7 , 10 , 12 ). В серотипе Agona SGI1 имеет то же хромосомное расположение, что и в штаммах DT 104, за исключением того, что в нем отсутствует ретроновая последовательность, обнаруженная ниже SGI1; таким образом, он расположен между геном thdf и геном yidY хромосомы ( 7 ).
Typhimurium DT 104 ( 11 ), поскольку фаговое типирование является доступны только в специализированных лабораториях. Однако недавно сообщалось о SGI1 в другом фаговом типе серотипа Typhimurium и в серотипе Agona, что предполагает горизонтальный перенос SGI1 (9).1048 7 , 10 , 12 ). В серотипе Agona SGI1 имеет то же хромосомное расположение, что и в штаммах DT 104, за исключением того, что в нем отсутствует ретроновая последовательность, обнаруженная ниже SGI1; таким образом, он расположен между геном thdf и геном yidY хромосомы ( 7 ).
Открыть в отдельном окне
Генетическая организация кластера генов устойчивости к антибиотикам SGI1 Salmonella enterica серотипа Typhimurium DT 104. DR-L и DR-R представляют собой левый и правый прямые повторы соответственно, заключающие в скобки SGI1. Полимеразные цепные реакции (ПЦР), используемые для оценки генетической организации генов устойчивости к антибиотикам (ПЦР floR , A, B, C, D и E) и соединения SGI1 с хромосомой (ПЦР LJ и RJ для левого и правого соединений, соответственно). Сокращения для сайтов рестрикции: X, Xba I; H, Hin dIII; Xh, Xho I.
Сокращения для сайтов рестрикции: X, Xba I; H, Hin dIII; Xh, Xho I.
Недавно Nastasi и Mammina сообщили о наличии генов floR и tet (G), обнаруженных с помощью ПЦР в штамме S. enterica серотипа Paratyphi B биовара Java, выделенном из тропические рыбы в Сингапуре ( 13 ). Мы исследовали этот изолят, чтобы определить наличие SGI1.
Фенотип резистентности к антибиотикам серотипа Paratyphi B оценивали методом диско-диффузионного анализа. Все диски с антибиотиками, кроме флорфеникола, были приобретены у компании Bio-Rad (Marnes-La-Coquette, Франция). Диски с флорфениколом и само лекарство были получены от Schering-Plough Animal Health (Кенилворт, Нью-Джерси). Штамм серотипа Paratyphi B показал профиль множественной лекарственной устойчивости, типичный для штаммов серотипа Typhimurium DT 104 или серотипа Agona, несущих SGI1, то есть устойчивость к ампициллину, хлорамфениколу и флорфениколу, стрептомицину и спектиномицину, сульфонамидам и тетрациклинам. Более того, этот штамм показал такой же уровень устойчивости к флорфениколу, как и серотипы Typhimurium DT 104 и Agona, т. е. МИК флорфеникола 32 мкг/мл ( 12 ). Штамм был чувствителен к триметоприму и хинолонам налидиксовой кислоты, энрофлоксацину и ципрофлоксацину.
Более того, этот штамм показал такой же уровень устойчивости к флорфениколу, как и серотипы Typhimurium DT 104 и Agona, т. е. МИК флорфеникола 32 мкг/мл ( 12 ). Штамм был чувствителен к триметоприму и хинолонам налидиксовой кислоты, энрофлоксацину и ципрофлоксацину.
В штамме серотипа Paratyphi B не было обнаружено плазмид, что свидетельствует о хромосомной локализации всех генов устойчивости к антибиотикам. ПЦР-картирование типичных генов устойчивости к антибиотикам и интегронов, связанных с SGI1, выполняли, как описано () ( 12 ). ПЦР-амплификации дали продукты из геномной ДНК, выделенной из штамма серотипа Paratyphi B, размера, ожидаемого из ДНК серотипа Typhimurium DT 104 контрольного штамма BN9.181 (данные не показаны) ( 6 , 12 ). Частичное нуклеотидное секвенирование гена floR показало 100% идентичность с серотипом Typhimurium DT 104. Таким образом, эти результаты ПЦР-картирования показали, что штамм серотипа Paratyphi B содержит весь кластер генов устойчивости к антибиотикам серотипа Typhimurium DT 104.
Сохранность организации гена устойчивости к антибиотикам дополнительно оценивали с помощью Саузерн-блоттинга Hin dIII- или Xho 9.1049 I расщепленная геномная ДНК со вставкой 12 т.п.н. Xba I в качестве зонда (см. фрагмент Xba I) из рекомбинантной плазмиды pSTF3, содержащей кластер генов устойчивости к антибиотикам DT 104, как описано ( 6 , 12 ). Профили саузерн-блоттинга штамма Paratyphi B серотипа были идентичны профилям контрольного штамма DT 104 BN9181 и штамма Agona серотипа 959SA97 (), подтверждая, что организация генов устойчивости к антибиотикам в штамме Paratyphi B серотипа такая же, как и в серотип Typhimurium DT 104 или серотип Agona, содержащий SGI1 ( 7 , 12 ).
Открыть в отдельном окне. серотип Typhimurium DT 104, штамм BN9181 (дорожки 2), серотип Agona, штамм 959SA97 (дорожки 3), и серотип Paratyphi, штамм B (дорожки 4). Дорожка 1: лестница ДНК. B. Саузерн-блот-гибридизация с p1-9зонд Xba I расщепленных геномных ДНК S. enterica серотипа Typhimurium DT 104 штамма BN9181 (дорожки 2), серотипа Agona штамма 959SA97 (дорожки 3) и серотипа штамма Paratyphi B (дорожки 4). Дорожка 1: лестница ДНК.
enterica серотипа Typhimurium DT 104 штамма BN9181 (дорожки 2), серотипа Agona штамма 959SA97 (дорожки 3) и серотипа штамма Paratyphi B (дорожки 4). Дорожка 1: лестница ДНК.
Для оценки присутствия всего SGI1 и его локализации в хромосоме штамма Paratyphi серотипа B была проведена ПЦР с использованием праймеров, соответствующих левому и правому соединениям SGI1 в хромосоме Salmonella . ПЦР учитывал наличие или отсутствие int2 -ретронная последовательность, расположенная ниже SGI1 в серотипе Typhimurium DT 104, но не в серотипе Agona () ( 7 , 8 ). ПЦР была положительной для левого соединения SGI1, как и для серотипа Typhimurium DT 104 или серотипа Agona. Если в качестве обратного праймера использовали последовательность int2 гена ретрона, ПЦР была отрицательной для правого соединения SGI1, но положительной, если использовали последовательность yidY гена. Продукты ПЦР показали ожидаемые размеры примерно 500 п.н. как для ПЦР левого соединения, так и для ПЦР правого соединения без последовательности ретрона в .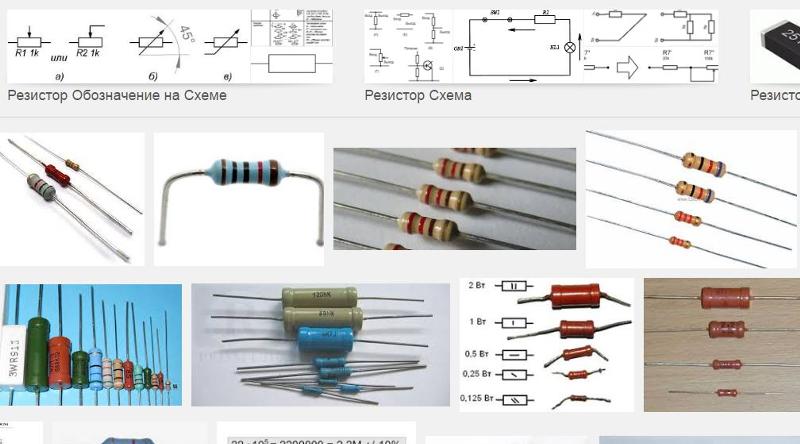 Эти данные показывают, что штамм Paratyphi B серотипа содержит SGI1 в том же хромосомном положении, что и у серотипа Typhimurium DT 104 или серотипа Agona, то есть между thdf и генов yidY , но отсутствует последовательность ретрона, обнаруженная в штаммах DT 104 и других штаммах серотипа Typhimurium ( 7 , 8 ). Присутствие всего SGI1 также было подтверждено Саузерн-блоттингом Xba I-расщепленной геномной ДНК с зондом p1-9, содержащим вставку 2 т.п.о. Eco RI. Это соответствует центральной области SGI1, включающей части открытых рамок считывания S023 и S024, которые кодируют предполагаемые белки геликазы и экзонуклеазы, выше кластера генов устойчивости к антибиотикам (9).1048 7 ). Этот зонд выявил фрагментов Xba I размером 4 и 9 т.п.н., ожидаемых в штамме серотипа Paratyphi B и контрольных штаммах серотипа Typhimurium DT 104 и серотипа Agona ().
Эти данные показывают, что штамм Paratyphi B серотипа содержит SGI1 в том же хромосомном положении, что и у серотипа Typhimurium DT 104 или серотипа Agona, то есть между thdf и генов yidY , но отсутствует последовательность ретрона, обнаруженная в штаммах DT 104 и других штаммах серотипа Typhimurium ( 7 , 8 ). Присутствие всего SGI1 также было подтверждено Саузерн-блоттингом Xba I-расщепленной геномной ДНК с зондом p1-9, содержащим вставку 2 т.п.о. Eco RI. Это соответствует центральной области SGI1, включающей части открытых рамок считывания S023 и S024, которые кодируют предполагаемые белки геликазы и экзонуклеазы, выше кластера генов устойчивости к антибиотикам (9).1048 7 ). Этот зонд выявил фрагментов Xba I размером 4 и 9 т.п.н., ожидаемых в штамме серотипа Paratyphi B и контрольных штаммах серотипа Typhimurium DT 104 и серотипа Agona ().
Макрорестрикционный анализ с помощью гель-электрофореза в пульсирующем поле ДНК штамма Paratyphi B серотипа, разрезанной Xba I, показал, что он генетически отличается от обоих серотипов Typhimurium DT 104 и Agona (), что дополнительно указывает на молекулярном уровне, что возникновение SGI1 в штамме Paratyphi B серотипа, вероятно, является результатом горизонтального переноса, а не сероконверсии известных серотипов S.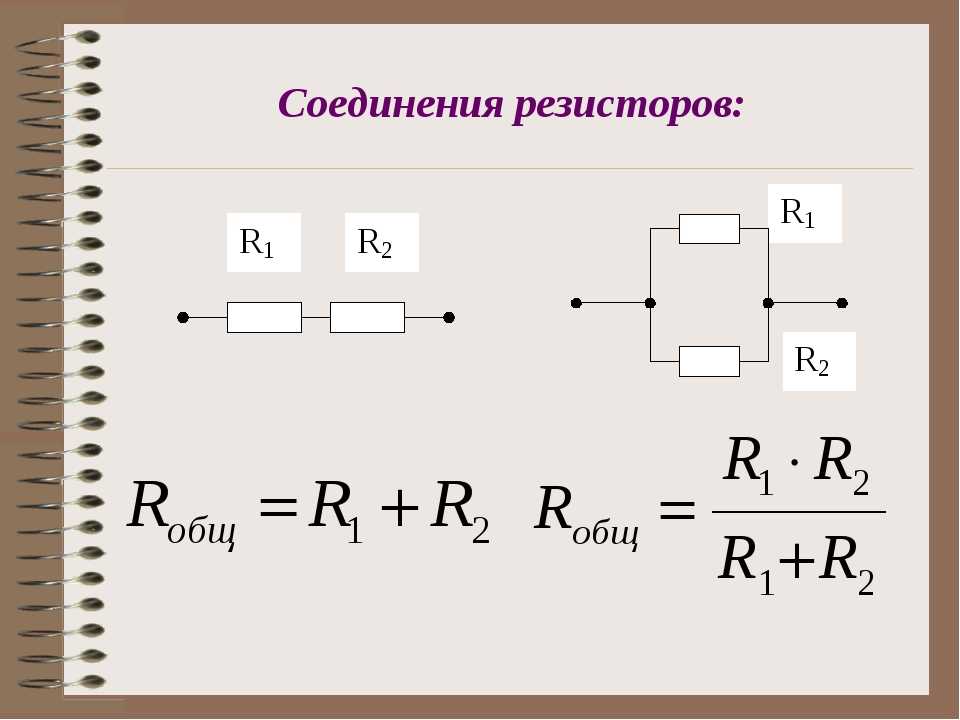 enterica , содержащих SGI1.
enterica , содержащих SGI1.
Открыть в отдельном окне. ) и штамм серотипа Paratyphi B (дорожка 4). Дорожка 1: лестница ДНК.
Мы идентифицировали SGI1 в штамме S. Paratyphi B. Эти данные в сочетании с идентификацией SGI1 в S. Agona и S. Typhimurium предполагают горизонтальный перенос этой области ( 7 , 10 , 12 ). Тот факт, что SGI1 имеет такое же хромосомное расположение у S. Typhimurium, S. Agona и S. Paratyphi B, предполагает, что его вставка произошла в результате гомологичной рекомбинации, возможно, посредством фаговой трансдукции ( 14 ). Эта гипотеза экспериментально подтверждается тем фактом, что гены резистентности серотипа Typhimurium DT 104 могут эффективно трансдуцироваться Р22-подобным фагом ES18 и фагом PDT17, который высвобождается всеми проанализированными изолятами DT 104 (9).1048 14 ). Однако остается вопрос, почему последовательность ретрона ниже SGI1 в штаммах Typhimurium DT 104 и DT 120 серотипа ( 7 ) не присутствует ниже SGI1 в других серотипах. Возможным объяснением может быть то, что в описанных здесь горизонтальных переносах штаммы-доноры SGI1 не являются штаммами серотипа Typhimurium. Как только SGI1 был приобретен, он может стать стабильным в хромосоме, поскольку эксперименты по вырезанию in vitro не смогли продемонстрировать его потерю в штамме DT 104 в отсутствие селективного давления антибиотиков (9).1048 8 ). Этот фактор может способствовать эволюции возбудителей S. enterica , подобно приобретению островов патогенности ( 15 , 16 ).
Возможным объяснением может быть то, что в описанных здесь горизонтальных переносах штаммы-доноры SGI1 не являются штаммами серотипа Typhimurium. Как только SGI1 был приобретен, он может стать стабильным в хромосоме, поскольку эксперименты по вырезанию in vitro не смогли продемонстрировать его потерю в штамме DT 104 в отсутствие селективного давления антибиотиков (9).1048 8 ). Этот фактор может способствовать эволюции возбудителей S. enterica , подобно приобретению островов патогенности ( 15 , 16 ).
Приобретение SGI1, возможно, было ключевым фактором, способствовавшим всемирной эпидемии DT 104, возможно, не только благодаря селекции противомикробных агентов в сельском хозяйстве ( 17 ), но и благодаря свойствам вирулентности SGI1 ( 10 ). Таким образом, требуется дальнейшее наблюдение за появлением горизонтального переноса SGI на S. enterica серотипов, важных для общественного здравоохранения.
Благодарим C. Mouline за квалифицированную техническую помощь.
Эта работа была поддержана грантом Французского национального института агрономических исследований (INRA, Action Transversalité 2001-2003).
•
Д-р Менье является научным сотрудником Национального института агрономических исследований во Франции. Ее основные интересы — механизмы антибиотикорезистентности и распространение генов антибиотикорезистентности среди энтеробактериальных патогенов.
Рекомендуемое цитирование: Meunier D, Boyd D, Mulvey MR, Baucheron S, Mammina C, Nastasi A, et al. Сальмонелла enterica Серотип Typhimurium DT 104 Устойчивость к антибиотикам Геномный остров I в серотипе Paratyphi B. Emerg Infect Dis. [сериал в Интернете]. 2002 г., апрель [дата указана]. Доступно на http://www.cdc.gov/ncidod/EID/vol8no4/01-0375.htm
1. Hancock D, Besser T, Gay J, Rice D, Davis M, Gay C. Глобальная эпидемиология мультирезистентных Сальмонелла энтерика серовар Typhimurium DT104. В: Браун С., Болин С., редакторы.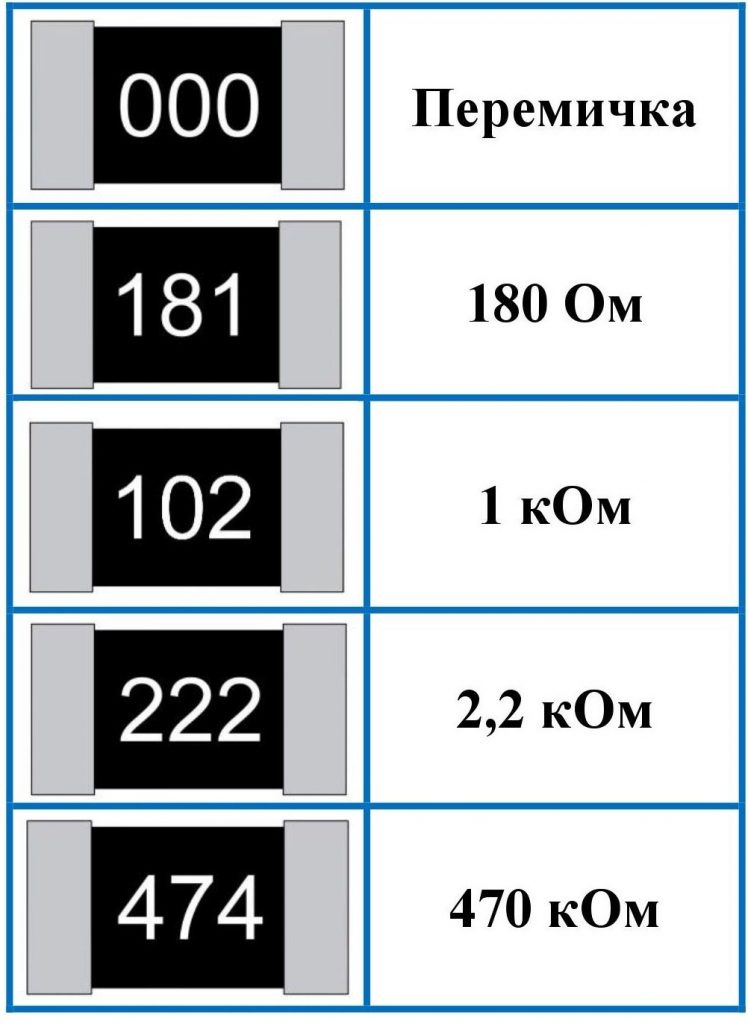 Новые болезни животных. Вашингтон: ASM Press; 2000. с. 217-43. [Google Scholar]
Новые болезни животных. Вашингтон: ASM Press; 2000. с. 217-43. [Google Scholar]
2. Besser TE, Goldoft M, Pritchett LC, Khakhria R, Hancock DD, Rice DH, et al. Мультирезистентные инфекции Salmonella Typhimurium DT104 человека и домашних животных на тихоокеанском северо-западе США. Эпидемиол инфекции. 2000; 124:193–200. 10.1017/S09502688983 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
3. Davis MA, Hancock DD, Besser TE, Rice DH, Gay JM, Gay C, et al. Изменения устойчивости к противомикробным препаратам среди изолятов Salmonella enterica серовара Typhimurium от людей и крупного рогатого скота на северо-западе США, 1982-1997 гг. Эмердж Инфекция Дис. 1999; 5: 802–6. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Glynn MK, Bopp C, Dewitt W, Dabney P, Mokhtar M, Angulo FJ. Появление инфекции с множественной лекарственной устойчивостью Salmonella enterica серотипа Typhimurium DT104 в Соединенных Штатах.
N Engl J Med. 1998;338:1333–8. 10.1056/NEJM199805073381901 [PubMed] [CrossRef] [Google Scholar]
1998;338:1333–8. 10.1056/NEJM199805073381901 [PubMed] [CrossRef] [Google Scholar]
5. Poppe C, Smart N, Khakhria R, Johnson W, Spika J, Prescott J. Salmonella typhimurium DT104: вирулентный и устойчивый к лекарствам патоген. Кан Вет Дж. 1998; 39: 559–65. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Арканджоли М.А., Леруа-Сетрин С., Мартель Дж.Л., Часлус-Данкла Э. Новый ген устойчивости к хлорамфениколу и флорфениколу, окруженный двумя интегронными структурами в Salmonella typhimurium ДТ104. FEMS Microbiol Lett. 1999; 174: 327–32. 10.1111/j.1574-6968.1999.tb13586.x [PubMed] [CrossRef] [Google Scholar]
7. Boyd D, Peters GA, Cloeckaert A, Sidi Boumedine K, Chaslus-Dancla E, Imberechts H, et al.
Полная нуклеотидная последовательность геномного острова размером 43 килобаз, связанного с областью множественной лекарственной устойчивости Salmonella enterica серовара Typhimurium DT104, и ее идентификация в фаговом типе DT120 и сероваре Agona. J Бактериол. 2001; 183: 5725–32. 10.1128/JB.183.19.5725-5732.2001 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
J Бактериол. 2001; 183: 5725–32. 10.1128/JB.183.19.5725-5732.2001 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Boyd DA, Peters GA, Ng LK, Mulvey MR. Частичная характеристика геномного острова, связанного с областью множественной лекарственной устойчивости Salmonella enterica Typhimurium DT104. FEMS Microbiol Lett. 2000; 189: 285–91. 10.1111/j.1574-6968.2000.tb09245.x [PubMed] [CrossRef] [Google Scholar]
9. Briggs CE, Fratamico PM. Молекулярная характеристика кластера генов устойчивости к антибиотикам из Salmonella typhimurium DT104. Противомикробные агенты Chemother. 1999;43:846–9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Cloeckaert A, Schwarz S. Молекулярная характеристика, распространение и эволюция множественной лекарственной устойчивости у Salmonella enterica Typhimurium DT104. Вет рез. 2001; 32: 301–10. 10.1051/vetres:2001126 [PubMed] [CrossRef] [Google Scholar]
11. Bolton LF, Kelley LC, Lee MD, Fedorka-Cray PJ, Maurer JJ. Обнаружение полирезистентного штамма Salmonella enterica серотип typhimurium DT104 на основе гена, придающего перекрестную устойчивость к флорфениколу и хлорамфениколу.
Дж. Клин Микробиол. 1999; 37: 1348–51. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Обнаружение полирезистентного штамма Salmonella enterica серотип typhimurium DT104 на основе гена, придающего перекрестную устойчивость к флорфениколу и хлорамфениколу.
Дж. Клин Микробиол. 1999; 37: 1348–51. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Cloeckaert A, Sidi Boumedine K, Flaujac G, Imberechts H, D’Hooghe I, Chaslus-Dancla E. Возникновение Salmonella enterica серовара Typhimurium DT104- подобный кластеру генов устойчивости к антибиотикам, включая ген floR в S. enterica серовара Agona. Противомикробные агенты Chemother. 2000;44:1359–61. 10.1128/AAC.44.5.1359-1361.2000 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
серотипы, Италия. Эмердж Инфекция Дис. 2001; 7: 455–7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Schmieger H, Schicklmaier P. Трансдукция множественной лекарственной устойчивости Salmonella enterica серовара Typhimurium DT104.
FEMS Microbiol Lett. 1999; 170: 251–6. 10.1111/j.1574-6968.1999.tb13381.x [PubMed] [CrossRef] [Google Scholar]
1999; 170: 251–6. 10.1111/j.1574-6968.1999.tb13381.x [PubMed] [CrossRef] [Google Scholar]
15. Hansen-Wester I, Hensel M. Островки патогенности Salmonella , кодирующие системы секреции III типа. микробы заражают. 2001; 3: 549–59. 10.1016/S1286-4579(01)01411-3 [PubMed] [CrossRef] [Google Scholar]
16. Hentschel U, Hacker J. Острова патогенности: верхушка айсберга. микробы заражают. 2001; 3: 545–8. 10.1016/S1286-4579(01)01410-1 [PubMed] [CrossRef] [Google Scholar]
17. Angulo FJ, Griffin PM. Изменения устойчивости к противомикробным препаратам в Salmonella enterica серовар Typhimurium. Эмердж Инфекция Дис. 2000; 6: 436–7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Эталон стабильного сопротивления серии IETLAB SR102/103/104 (1 ppm/год)
(полные спецификации можно найти в техническом описании)
Высочайшая точность (1 ppm/год) ) и стабильность (обычно ±0,5 ppm/год) любого эталона сопротивления
Самый низкий температурный коэффициент (< 0,1 ppm/ºC)
Доступны 3 модели: 100 Ом, 1000 Ом, 10 000 Ом.
Остается передовым эталоном сопротивления, не имеющим аналогов, уже более 35 лет
Гарантия на 5 лет
Основной эталон национальных метрологических лабораторий во всем мире
Остается основным эталоном сопротивления, не имеющий эквивалента, более 35 лет
Маслонаполненный, герметичный; блоки могут поставляться без корпусов для использования в масляной ванне
Включена аккредитованная калибровка ISO/IEC-17025
Включены измерения альфа-бета
техническое описание
руководство
SR102, SR103, SR104 ОСНОВНЫЕ ХАРАКТЕРИСТИКИ 90136
Стабильность Первые 2 года: ±1 ppm/год, затем: ±0,5 ppm/год
Температурный коэффициент
Альфа: <0,1 ppm/°C при 23°C
Бета (1/2 скорости изменения TC ):<0,03 ppm/°C от 18°C до 28°Ct
Печи или внешнее питание не требуются
Коэффициент мощности <1 ppm/Вт
Корректировка до номинального значения ±1 ppm
Погрешность измерения <0,32 ppm
Максимальное напряжение 500 В от пика до корпуса
Номинальная мощность 1 Вт (Мгновенная перегрузка 100 Вт не приведет к отказу)
Термоэдс Термоэдс на клеммах не превышает ±0,1 мкВ при нормальных условиях.
Сопротивление изоляции Все клеммы имеют минимальное сопротивление 10 12 Ом относительно земли
Внутренний датчик температуры Резистор 100 Ом, 1 кОм или 10 кОм с температурным коэффициентом 1000 ppm/°C. Для калибровки предусмотрена встроенная гильза для термометра
Герметизация Для устранения воздействия влаги резистор герметизирован в масле металлическими стеклянными уплотнениями. Изменение сопротивления
<±0,1 ppm при нормальных изменениях атмосферного давления и влажности.
Самый стабильный в мире эталон сопротивления
IET продолжает производить SR102 и SR104 в соответствии с теми же строгими спецификациями, что и esi/Tegam
год) и точные (1 ppm), эталоны сопротивления 100 Ом, 1 кОм, 10 кОм. Эталоны SR102, SR103 и SR104 оборудованы для пятиконтактных измерений, используемых для показаний стандартных резисторов и показаний датчика температуры. Индивидуальные данные калибровки, которые определяют стандартное отклонение и температурные характеристики, поставляются с каждым устройством. Кроме того, каждый блок оснащен тепловым колодцем, чтобы была известна точная температура эталона сопротивления. Это позволяет пользователю проводить высокоточные измерения с температурной компенсацией.
Индивидуальные данные калибровки, которые определяют стандартное отклонение и температурные характеристики, поставляются с каждым устройством. Кроме того, каждый блок оснащен тепловым колодцем, чтобы была известна точная температура эталона сопротивления. Это позволяет пользователю проводить высокоточные измерения с температурной компенсацией.
esi SR102 Первичный эталон сопротивления 100 Ом
Эти эталоны сопротивления были разработаны, чтобы обеспечить важную связь между довольно странными значениями сопротивления квантового Холла и традиционными значениями сопротивления декады. 100 Ом обеспечивает уникальные эталонные возможности.
esi SR103 Первичный эталон сопротивления 1000 Ом
Новый эталон сопротивления, недавно разработанный для расширения ассортимента доступных первичных эталонов.
esi SR104 Первичный эталон сопротивления 10 кОм
Это давний отраслевой стандарт для 10 кОм. Этот неизменный стандарт является эталоном высокой точности, стабильности и низких температурных коэффициентов для калибровок, требующих прослеживаемости NIST.
Прецизионные переносные эталоны сопротивления SR102, SR103 и SR104 представляют собой маслонаполненные, герметичные эталоны сопротивления с пятью выводами, предназначенные для прецизионных настольных измерений или измерений в масляной ванне.
Долговременная стабильность этих эталонов сопротивления обычно составляет менее 0,2 промилле в год, а температурный коэффициент менее 0,1 промилле на градус Цельсия. Эта превосходная долговременная стабильность и низкий температурный коэффициент достигаются за счет использования согласованных групп резисторов, изготовленных из сплава Evanohm-R. Полученный в результате низкий температурный коэффициент позволяет применять эти стандарты с высокой эффективностью внутри или вне среды с точно контролируемой температурой.
Максимальная точность рассчитывается как значение сопротивления с поправкой на температуру. Это достигается за счет использования встроенного датчика температуры RTD для измерения внутренней температуры и обращения к диаграмме коррекции температуры, прилагаемой к каждому блоку. Точность измерения встроенного термометра сопротивления лучше 0,1 градуса Цельсия.
Точность измерения встроенного термометра сопротивления лучше 0,1 градуса Цельсия.
Очень низкий коэффициент мощности достигается за счет использования стандартных резисторов с большой площадью поверхности, таких как наша оригинальная модель SR104, стандартный резистор 10 кОм. Резисторы окружены маслом для отвода тепла, выделяемого измерительными токами, через корпус из нержавеющей стали.
Эти характеристики облегчают проведение точных лабораторных сравнений без критического контроля окружающей среды и используются везде, где требуется очень точный, стабильный резистор с низким температурным коэффициентом.
Модели
Переносной эталон сопротивления 100 Ом: SR-102
Переносной эталон сопротивления 1000 Ом: SR-103
Переносной эталон сопротивления 10 000 Ом: SR-104
Дополнительно:
Для исключенной версии добавить -DC в конце часть номер
Устойчивость к алюминию у Arabidopsis Mutantalr-104 вызвана индуцированным алюминием увеличением ризосферы ph2 | Физиология растений
Abstract
Был исследован механизм, обеспечивающий повышенную устойчивость к Al у мутанта Arabidopsis thaliana alr -104. Для измерения потоков H + , генерируемых вдоль поверхности мелких корней арабидопсиса, использовали модифицированную систему вибрирующих микроэлектродов. В отсутствие Al никаких различий в корне Н 9Выявлено 1026 + флюсов между диким типом и alr -104. Однако воздействие Al вызвало двукратное увеличение чистого притока H + в alr -104, локализованного на кончике корня. Увеличенный поток повысил pH поверхности корня alr -104 на 0,15 единицы. Анализ роста корней использовали для оценки устойчивости к Al alr -104 и дикого типа в питательном растворе с сильным pH-буфером. Повышение pH питательного раствора с 4,4 до 4,5 значительно повысило устойчивость к Al у дикого типа, что согласуется с идеей о том, что повышенный чистый H 9Приток 1026 + может объяснить большее сопротивление Al в alr -104. Различия в устойчивости к Al между диким типом и alr -104 исчезали, когда корни выращивали в pH-буферной среде, что позволяет предположить, что устойчивость к Al у alr -104 опосредована только изменениями pH в ризосфере.
Для измерения потоков H + , генерируемых вдоль поверхности мелких корней арабидопсиса, использовали модифицированную систему вибрирующих микроэлектродов. В отсутствие Al никаких различий в корне Н 9Выявлено 1026 + флюсов между диким типом и alr -104. Однако воздействие Al вызвало двукратное увеличение чистого притока H + в alr -104, локализованного на кончике корня. Увеличенный поток повысил pH поверхности корня alr -104 на 0,15 единицы. Анализ роста корней использовали для оценки устойчивости к Al alr -104 и дикого типа в питательном растворе с сильным pH-буфером. Повышение pH питательного раствора с 4,4 до 4,5 значительно повысило устойчивость к Al у дикого типа, что согласуется с идеей о том, что повышенный чистый H 9Приток 1026 + может объяснить большее сопротивление Al в alr -104. Различия в устойчивости к Al между диким типом и alr -104 исчезали, когда корни выращивали в pH-буферной среде, что позволяет предположить, что устойчивость к Al у alr -104 опосредована только изменениями pH в ризосфере. Насколько нам известно, этот мутант представляет собой первое свидетельство механизма устойчивости к алюминию, основанного на индуцируемом алюминием увеличении рН поверхности корней.
Насколько нам известно, этот мутант представляет собой первое свидетельство механизма устойчивости к алюминию, основанного на индуцируемом алюминием увеличении рН поверхности корней.
Al является наиболее распространенным металлом в земной коре и встречается в почве в различных формах. В нейтральных и основных почвах Al в основном находится в виде оксидных или силикатных осадков, нетоксичных для растений. Однако в очень кислых почвах (pH < 5,0) Al превращается в растворимую форму октаэдрического гексагидрата, обычно называемую Al 9.1026 3+ , который считается основным фитотоксичным видом Al (Kochian, 1995). Начальным и наиболее ярким симптомом отравления алюминием является торможение роста корней, что приводит к уменьшению и повреждению корневой системы и может привести к дефициту минералов и водному стрессу. Токсичность алюминия является основным фактором, ограничивающим сельскохозяйственное производство на кислых почвах, которые составляют более 30% пахотных земель в мире (Von Uexkull and Mutert, 1995). Верхушка корня является основной мишенью токсичности Al, и снижение роста корня можно обнаружить в течение нескольких минут после добавления Al (Ryan et al., 19).93; Джонс и Кочиан, 1995).
Верхушка корня является основной мишенью токсичности Al, и снижение роста корня можно обнаружить в течение нескольких минут после добавления Al (Ryan et al., 19).93; Джонс и Кочиан, 1995).
Сообщалось о различиях между сортами или сортами в устойчивости к Al ряда сельскохозяйственных культур (обзор см. в Carver and Ownby, 1995). Были предложены две категории механизмов устойчивости к Al: толерантность к более высоким концентрациям Al в симпласте корня и способность исключать Al из верхушки корня (Taylor, 1991; Delhaize and Ryan, 1995; Kochian, 1995). В то время как мало что известно о механизмах симпластической толерантности (Aniol, 1984), недавно был описан механизм Al-исключения. Делхайз и др. (1993a, 1993b) продемонстрировали на изогенных линиях пшеницы, что присутствие Al индуцирует высвобождение большего количества малата из верхушки корня у устойчивых к Al генотипов. Как и некоторые другие органические кислоты, малат хелатирует Al 3+ в ризосфере и предотвращает поглощение Al корнями.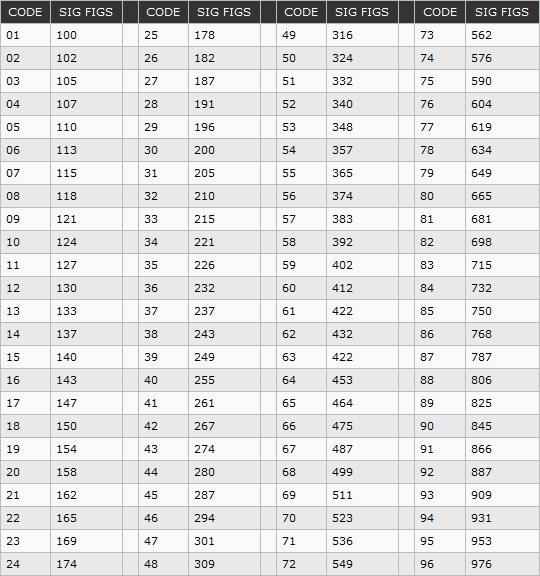 В этих исследованиях было показано, что резистентность сегрегируется как единый доминантный локус, названный alt1 . Впоследствии было показано, что высвобождение малата коррелирует с устойчивостью к Al у ряда других сортов пшеницы (Ryan et al., 19).95). Сходный механизм исключения наблюдался у кукурузы, у которой сообщалось об индуцированном Al высвобождении цитрата на верхушке корня (Pellet et al., 1994).
В этих исследованиях было показано, что резистентность сегрегируется как единый доминантный локус, названный alt1 . Впоследствии было показано, что высвобождение малата коррелирует с устойчивостью к Al у ряда других сортов пшеницы (Ryan et al., 19).95). Сходный механизм исключения наблюдался у кукурузы, у которой сообщалось об индуцированном Al высвобождении цитрата на верхушке корня (Pellet et al., 1994).
Еще в середине 1960-х годов Foy et al. (1965) предложили механизм исключения алюминия, который включает увеличение рН ризосферы. Подщелачивание ризосферы снизит концентрацию Al 3+ в пользу менее токсичных видов Al, таких как гидроксиды и фосфаты Al (Martell and Motekaitis, 1989). Было много сообщений об общей корреляции между устойчивостью к алюминию и кратковременным повышением pH ростового раствора для нескольких видов, включая пшеницу (Foy et al., 19).67, 1974; Mugwira et al., 1976, 1978; Mugwira and Elgawahry, 1979; Фой и Флеминг, 1982 г.; Fleming, 1983; Dodge, Hiatt, 1992), ячмень (Foy et al. , 1967), горох (Климашевский, Бернадская, 1973), рожь (Mugwira et al., 1976, 1978) и тритикале (Mugwira et al., 1976, 1978; Mugwira and Patel, 1977), но до сих пор не было прямых демонстраций этого механизма устойчивости к Al. В большинстве этих отчетов было неясно, были ли различия в pH причиной устойчивости к Al или они были результатом вызванного Al ингибирования корневой функции у чувствительных сортов.
, 1967), горох (Климашевский, Бернадская, 1973), рожь (Mugwira et al., 1976, 1978) и тритикале (Mugwira et al., 1976, 1978; Mugwira and Patel, 1977), но до сих пор не было прямых демонстраций этого механизма устойчивости к Al. В большинстве этих отчетов было неясно, были ли различия в pH причиной устойчивости к Al или они были результатом вызванного Al ингибирования корневой функции у чувствительных сортов.
Все эти исследования были основаны на измерениях pH нерасфасованного раствора, которые оказались проблематичными в двух отношениях. Во-первых, источник азота в питательной среде может оказывать значительное влияние на рН ризосферы, поскольку поглощение NO 3 − приводит к подщелачиванию среды, тогда как поглощение NH 4 + может вызывать подкисление ризосферы ( обзор см. Taylor, 1988). Следовательно, отношение NO 3 − к NH 3 в питательной среде каждого эксперимента может оказывать существенное влияние на рН ростового раствора (Taylor, 1991).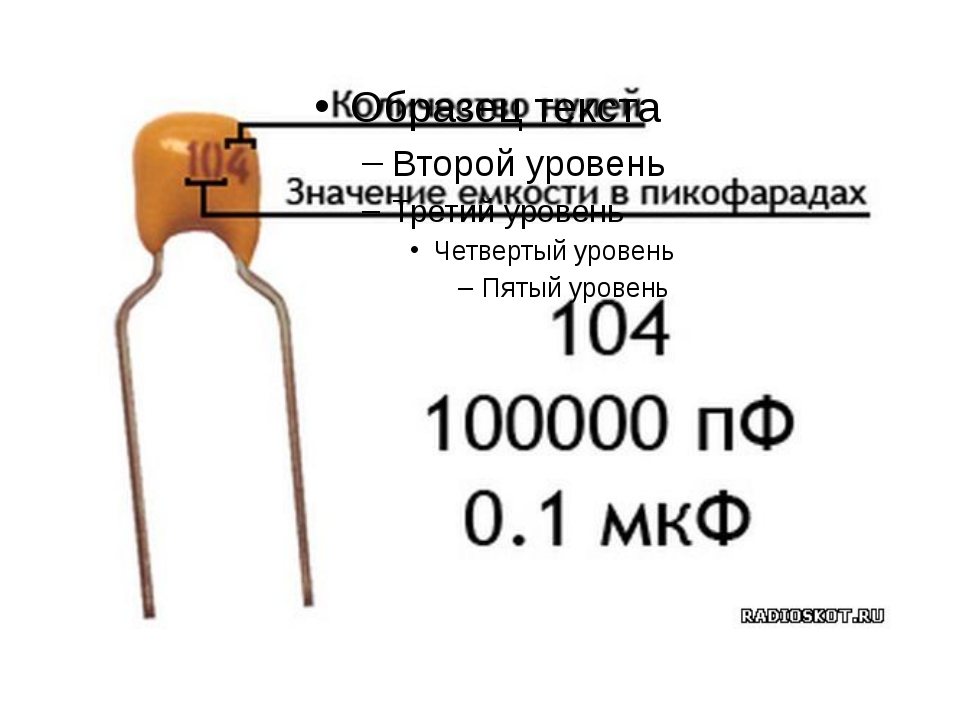 Во-вторых, измерения объемного раствора отражают изменения рН, связанные со всем корнем, а не конкретно с его кончиком, который является основным местом токсичности алюминия. По этой причине Миясака и соавт. (1989) использовали рН-микроэлектроды для картирования рН поверхности вдоль корней пшеницы и показали, что устойчивая к алюминию пшеница сорта Атлас поддерживает несколько более высокий рН (приблизительно 0,15 единицы рН) на кончике корня, но не в зрелых частях корня, по сравнению с с Al-чувствительным cv Scout. В этих экспериментах не было определено, приводили ли различия рН ризосферы на верхушке к устойчивости к Al или они просто отражали различия в функции корней после начала токсичности Al у сорта Scout.
Во-вторых, измерения объемного раствора отражают изменения рН, связанные со всем корнем, а не конкретно с его кончиком, который является основным местом токсичности алюминия. По этой причине Миясака и соавт. (1989) использовали рН-микроэлектроды для картирования рН поверхности вдоль корней пшеницы и показали, что устойчивая к алюминию пшеница сорта Атлас поддерживает несколько более высокий рН (приблизительно 0,15 единицы рН) на кончике корня, но не в зрелых частях корня, по сравнению с с Al-чувствительным cv Scout. В этих экспериментах не было определено, приводили ли различия рН ризосферы на верхушке к устойчивости к Al или они просто отражали различия в функции корней после начала токсичности Al у сорта Scout.
Недавно мы применили молекулярно-генетический подход, чтобы лучше понять токсичность и устойчивость к Al, изолировав Al-чувствительные и Al-толерантные мутанты в Arabidopsis thaliana . Ранее мы сообщали о Al-чувствительных мутантах Arabidopsis. Этот признак был описан восемью различными группами комплементации (Larsen et al.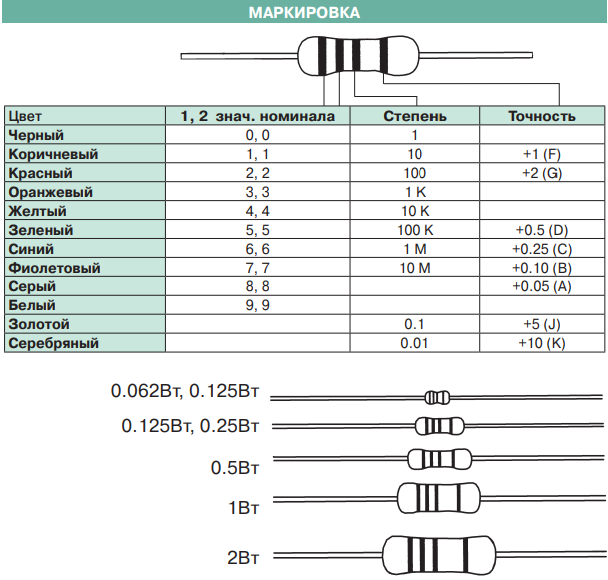 , 1996). В сопутствующей статье (Larsen et al., 1998) мы описываем семейство из пяти устойчивых к Al мутантов, которые картируются в двух разных локусах генома арабидопсиса. Все эти 9Мутанты 1048 alr исключают Al из верхушки корня в большей степени, чем дикий тип. Четыре мутанта картированы в одном и том же месте на хромосоме 1 и использовали механизм исключения Al, связанный с повышенным высвобождением малата и цитрата, тогда как другой мутант картирован в хромосоме 4.
, 1996). В сопутствующей статье (Larsen et al., 1998) мы описываем семейство из пяти устойчивых к Al мутантов, которые картируются в двух разных локусах генома арабидопсиса. Все эти 9Мутанты 1048 alr исключают Al из верхушки корня в большей степени, чем дикий тип. Четыре мутанта картированы в одном и том же месте на хромосоме 1 и использовали механизм исключения Al, связанный с повышенным высвобождением малата и цитрата, тогда как другой мутант картирован в хромосоме 4.
Мы описываем устойчивый к Al мутант хромосомы 4 . alr -104, который не проявляет усиленного высвобождения корневой органической кислоты. В этом исследовании мы исследовали, повышена ли устойчивость к алюминию в alr -104 был вызван повышенным рН ризосферы вокруг верхушки корня. Для этих исследований мы использовали вибрирующую систему микроэлектродов H + , которая позволяла нам измерять потоки корневых ионов с очень высокой степенью пространственного и временного разрешения (Kochian et al. , 1992; Smith et al., 1994). Мы модифицировали этот метод для измерения очень маленьких корней, таких как корни проростков арабидопсиса, и разработали метод для количественного определения потоков корней H + в гелевой пленке, содержащей комплексный питательный раствор. alr -104 показал индуцируемый Al повышенный приток H + на кончике корня, что привело к более высокому pH ризосферы в этой области (по сравнению с диким типом). Повышение рН апикальной ризосферы у alr -104 объясняет большую толерантность к Al у этого мутанта. Этот отчет предоставляет первое прямое доказательство, насколько нам известно, механизма устойчивости к Al, основанного на модификации рН апикальной ризосферы.
, 1992; Smith et al., 1994). Мы модифицировали этот метод для измерения очень маленьких корней, таких как корни проростков арабидопсиса, и разработали метод для количественного определения потоков корней H + в гелевой пленке, содержащей комплексный питательный раствор. alr -104 показал индуцируемый Al повышенный приток H + на кончике корня, что привело к более высокому pH ризосферы в этой области (по сравнению с диким типом). Повышение рН апикальной ризосферы у alr -104 объясняет большую толерантность к Al у этого мутанта. Этот отчет предоставляет первое прямое доказательство, насколько нам известно, механизма устойчивости к Al, основанного на модификации рН апикальной ризосферы.
МАТЕРИАЛЫ И МЕТОДЫ
Генетические и общефизиологические характеристики устойчивости к Al у дикого типа и alr- 104 и alr- 128 мутантов Arabidopsis thaliana Heyn. Экотип Columbia (Col-0), используемый в этом исследовании, описан в сопутствующей статье (Larsen et al., 1998). Проростки арабидопсиса выращивали на 2-миллиметровом слое геля (0,15% геллановой камеди; Gell-Gro, ICN), покрывающего предметное стекло микроскопа; гель содержал питательный раствор, как описано Larsen et al. (1996)(2 мм KNO 3 , 0,1 мм KH 2 PO 4 , 2 мм MgSO 4 , 0,25 мм [NH 4 ] 2 SO 4 , 1 мМ CA [№ 3 ] 2 , 1 мМ CASO 4 , 1 мм K 2 SO 4 , 1 мкм Mnso 4 , SO 4 , 1 μM MNSO 4 , SO 4 , 1 μM MNSO 4 , SO 4 , 1 μM MNSO 4 , SO 4 , 1 µM MNSO 4 , SO 4 , 1 мм. 5 мкм ч 3 BO 3 , 0,05 мкм CUSO 4 , 0,2 мкм ZnSO 4 , 0,02 мкм NAMOO 4 , 0,1 мкм CACL 2 , 0,00111109, 0,1 мкм 2 , 0,00111109, 0,1 мкм 2 , 0,00111109, 0,1 мкм 2 , 0,00111109, 0,1 мкм 2 .
Экотип Columbia (Col-0), используемый в этом исследовании, описан в сопутствующей статье (Larsen et al., 1998). Проростки арабидопсиса выращивали на 2-миллиметровом слое геля (0,15% геллановой камеди; Gell-Gro, ICN), покрывающего предметное стекло микроскопа; гель содержал питательный раствор, как описано Larsen et al. (1996)(2 мм KNO 3 , 0,1 мм KH 2 PO 4 , 2 мм MgSO 4 , 0,25 мм [NH 4 ] 2 SO 4 , 1 мМ CA [№ 3 ] 2 , 1 мМ CASO 4 , 1 мм K 2 SO 4 , 1 мкм Mnso 4 , SO 4 , 1 μM MNSO 4 , SO 4 , 1 μM MNSO 4 , SO 4 , 1 μM MNSO 4 , SO 4 , 1 µM MNSO 4 , SO 4 , 1 мм. 5 мкм ч 3 BO 3 , 0,05 мкм CUSO 4 , 0,2 мкм ZnSO 4 , 0,02 мкм NAMOO 4 , 0,1 мкм CACL 2 , 0,00111109, 0,1 мкм 2 , 0,00111109, 0,1 мкм 2 , 0,00111109, 0,1 мкм 2 , 0,00111109, 0,1 мкм 2 . , рН доведен до 4,2). Гель был ограничен пластиковой подложкой, приклеенной к предметному стеклу. Четыре-шесть семян дикого типа и alr мутантов ( alr -104 и alr -128) высевали на слой геля, предметное стекло помещали под углом 30 o в ростовую камеру с 16-часовым днем/8-часовым ночным циклом. на 4-5 д.
, рН доведен до 4,2). Гель был ограничен пластиковой подложкой, приклеенной к предметному стеклу. Четыре-шесть семян дикого типа и alr мутантов ( alr -104 и alr -128) высевали на слой геля, предметное стекло помещали под углом 30 o в ростовую камеру с 16-часовым днем/8-часовым ночным циклом. на 4-5 д.
Во время инкубации нижний конец предметных стекол погружали в питательный раствор того же состава. Жидкую среду меняли ежедневно, чтобы предотвратить истощение питательных веществ в геле. За 12 часов до эксперимента предметные стекла ориентировали горизонтально, а среду в слое геля уравновешивали 200 мл питательного раствора. Для обработки алюминием питательный раствор также содержал 300 мкм AlCl9.1108 3 . pH растворов, содержащих Al, регулировали перед добавлением AlCl 3 , как подробно описано Larsen et al. (1996). При использовании забуференного питательного раствора добавляли 10 мм трубы Homo-Pipes (Research Organics, Cleveland, OH) перед регулированием pH и добавлением AlCl 3 .
Измерение роста корней
Рост корней измеряли на инвертированном микроскопе (модель IM35, Zeiss) с использованием объектива с большим рабочим расстоянием 40× (общее увеличение, ×400). Кончики корней выравнивали по шкале окулярного микрометра и регистрировали удлинение корней через 1, 2 и 4 мин после выравнивания. Скорость роста корней выражали в микрометрах в минуту как среднее значение для 12 и более проростков в группе. Чтобы предотвратить механическое повреждение корней во время манипуляций с предметным стеклом, для измерения выбирали только корни, полностью погруженные в гель.
Измерение корня H
+ Потоки с вибрирующим H + Микроэлектрод Предметное стекло с проростками помещали в прозрачную поликарбонатную камеру размером 30 мм × 80 мм × 4 мм. Камеру заполняли 5 мл соответствующего питательного раствора, который непрерывно меняли со скоростью 1 мл/мин. H + -селективные микроэлектроды (диаметр кончика 1 мкм), содержащие селективную смесь H + (каталожный номер 95297, Fluka), были сконструированы, как описано ранее (Lucas and Kochian, 19).86). Система вибрирующих микроэлектродов была подробно описана в другом месте (Kochian et al., 1992; Smith et al., 1994) и использовалась с модификациями. Если не указано иное, микроэлектрод был ориентирован перпендикулярно поверхности корня и вибрировал вдоль оси, перпендикулярной корню. Электрод вибрировали внутри геля между двумя положениями, 5 и 35 мкм от поверхности корня (рис. 1). Опять же, для измерения использовались только корни, полностью погруженные в гель.
H + -селективные микроэлектроды (диаметр кончика 1 мкм), содержащие селективную смесь H + (каталожный номер 95297, Fluka), были сконструированы, как описано ранее (Lucas and Kochian, 19).86). Система вибрирующих микроэлектродов была подробно описана в другом месте (Kochian et al., 1992; Smith et al., 1994) и использовалась с модификациями. Если не указано иное, микроэлектрод был ориентирован перпендикулярно поверхности корня и вибрировал вдоль оси, перпендикулярной корню. Электрод вибрировали внутри геля между двумя положениями, 5 и 35 мкм от поверхности корня (рис. 1). Опять же, для измерения использовались только корни, полностью погруженные в гель.
Рис. 1.
Открыть в новой вкладкеСкачать слайд
Экспериментальная установка для измерения потоков H + вокруг корней арабидопсиса. Корни росли в тонком слое геля, уравновешенного питательным раствором. Микроэлектрод устанавливали вертикально и перпендикулярно корню и вибрировали вдоль его длинной оси на расстоянии от 5 до 35 мкм от поверхности корня. Гель удерживали на предметном стекле микроскопа в камере, содержащей соответствующий питательный раствор. Раствор постоянно меняли с помощью перистальтического насоса.
Гель удерживали на предметном стекле микроскопа в камере, содержащей соответствующий питательный раствор. Раствор постоянно меняли с помощью перистальтического насоса.
Эффективность вибрирующего микроэлектрода H + для обнаружения градиента H + в геле определяли, следуя процедуре, модифицированной Smith et al. (1994). Искусственный градиент H + был установлен и измерен с помощью вибрирующего микроэлектрода H + , и измеренный градиент сравнивался с теоретическими значениями градиента, определенными из уравнений диффузии. Ниже описаны модификации, которые были сделаны для учета буферного эффекта геля и питательного раствора. Гель, состоящий из 0,15% геллановой камеди в питательном растворе, доводили до pH 4,0 и вводили в микропипетку (диаметр кончика примерно 5 мкм). Пипетку устанавливали на предметное стекло и заливали 2-мм слоем того же геля при рН 6,0. Ионную силу геля регулировали добавлением 99 мкм KOH для обеспечения катиона для встречной диффузии и минимизации осмотического потока воды между двумя фазами. Через 4 ч вокруг кончика микропипетки, служившего источником ионов, возник устойчивый диффузионный градиент H + — (данные не представлены). Градиент H + измеряли микроэлектродом pH и сравнивали с ожидаемым градиентом диффузии, полученным из соответствующего уравнения диффузии (Jaffe and Levy, 1987; Kühtreiber and Jaffe, 1990; Smith et al., 1994).
Через 4 ч вокруг кончика микропипетки, служившего источником ионов, возник устойчивый диффузионный градиент H + — (данные не представлены). Градиент H + измеряли микроэлектродом pH и сравнивали с ожидаемым градиентом диффузии, полученным из соответствующего уравнения диффузии (Jaffe and Levy, 1987; Kühtreiber and Jaffe, 1990; Smith et al., 1994).
Измерение pH поверхности корня с помощью микроэлектрода H
+ pH ризосферы вдоль корня измеряли с помощью стационарного микроэлектрода H + с использованием той же системы, которая описана выше для измерения потока. Концентрацию H + определяли в неперемешиваемом слое, прилегающем к корню, в радиальном направлении на расстоянии 20 мкм от поверхности корня. Вместе с измерением потока H + , направленного от этой точки к поверхности корня, по закону Фика рассчитывали рН на поверхности корня (Crank, 19). 75). Этот расчет предполагает радиальную диффузию H + в цилиндрический корень: + концентрация на расстоянии 20 мкм от поверхности корня, J H+ – поток H + на расстоянии 20 мкм от поверхности корня, r – диаметр корня, Δ r точка измерения и поверхность корня (20 мкм) и D H+ — коэффициент диффузии для H + (9,308 × 10 −5 см 2 с −1 ). Измерения концентрации H + на различных радиальных расстояниях от поверхности корня следовали экспоненциальной функции (данные не показаны) и, таким образом, подтверждают профиль радиальной диффузии, ранее задокументированный для корней кукурузы Kochian et al. (1992).
75). Этот расчет предполагает радиальную диффузию H + в цилиндрический корень: + концентрация на расстоянии 20 мкм от поверхности корня, J H+ – поток H + на расстоянии 20 мкм от поверхности корня, r – диаметр корня, Δ r точка измерения и поверхность корня (20 мкм) и D H+ — коэффициент диффузии для H + (9,308 × 10 −5 см 2 с −1 ). Измерения концентрации H + на различных радиальных расстояниях от поверхности корня следовали экспоненциальной функции (данные не показаны) и, таким образом, подтверждают профиль радиальной диффузии, ранее задокументированный для корней кукурузы Kochian et al. (1992).
РЕЗУЛЬТАТЫ
Эффективность Н
+ -селективного вибрационного микроэлектрода Вибрационный электрод был откалиброван с учетом буферной способности комплексной среды и задержки измерения градиентов H + , вызванной временем отклика ионофора H + (Smith et al.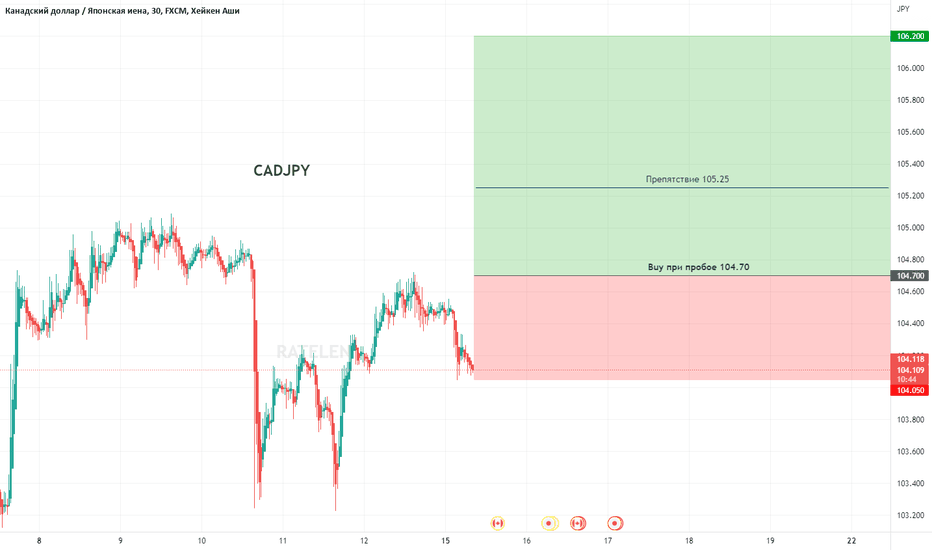 , 1994). Эффективность микроэлектрода представляет собой процент от общего потока ионов, который регистрируется электродом как разность потенциалов во время измерения. Определенный источник H + использовали для определения эффективности (см. Методы). рН-микроэлектрод вибрировали на разных расстояниях от этого источника и определяли разность потенциалов в этих положениях. Эти данные сравнивались с разностью потенциалов, рассчитанной по закону Фика и предполагающей 100% КПД электрода (рис.2). Эффективность описанной здесь системы составила 32%. Это можно увидеть как разницу в наклонах графиков, построенных на рисунке 2. Повторы этой калибровки показали, что между отдельными микроэлектродами существуют лишь небольшие различия. Эффективность микроэлектрода использовалась для корректировки измеренных значений потока с учетом того факта, что вибрация H 9Электрод 1026 + обнаружил только 32% общего градиента.
, 1994). Эффективность микроэлектрода представляет собой процент от общего потока ионов, который регистрируется электродом как разность потенциалов во время измерения. Определенный источник H + использовали для определения эффективности (см. Методы). рН-микроэлектрод вибрировали на разных расстояниях от этого источника и определяли разность потенциалов в этих положениях. Эти данные сравнивались с разностью потенциалов, рассчитанной по закону Фика и предполагающей 100% КПД электрода (рис.2). Эффективность описанной здесь системы составила 32%. Это можно увидеть как разницу в наклонах графиков, построенных на рисунке 2. Повторы этой калибровки показали, что между отдельными микроэлектродами существуют лишь небольшие различия. Эффективность микроэлектрода использовалась для корректировки измеренных значений потока с учетом того факта, что вибрация H 9Электрод 1026 + обнаружил только 32% общего градиента.
Рис. 2.
Открыть в новой вкладкеСкачать слайд
Эффективность вибрационного H + -селективный микроэлектрод. Электрод вибрировали в разных положениях от источника H + (микропипетка, содержащая гелеобразный питательный раствор с pH 4,0), помещенного в питательный раствор с pH 6,0. В каждой точке определялась разность потенциалов (представляющая градиент H + между конечными точками вибрации) (▴). Сравнение этих данных с теоретическими значениями для H 9Градиент 1026 + , рассчитанный по закону Фика (□), позволил определить эффективность системы.
Электрод вибрировали в разных положениях от источника H + (микропипетка, содержащая гелеобразный питательный раствор с pH 4,0), помещенного в питательный раствор с pH 6,0. В каждой точке определялась разность потенциалов (представляющая градиент H + между конечными точками вибрации) (▴). Сравнение этих данных с теоретическими значениями для H 9Градиент 1026 + , рассчитанный по закону Фика (□), позволил определить эффективность системы.
Al Воздействие на потоки H
+ на кончике корня арабидопсиса Для определения пространственной картины потоков H + вдоль кончика корня арабидопсиса дикого типа, двумерные измерения потоков с помощью H + -селективного микроэлектрод изготавливали на 5-дневных проростках. Измерения проводились в нескольких точках вдоль кончика корня на радиальном расстоянии 20 и 50 мкм от поверхности корня. В этих точках электрод вибрировали либо параллельно, либо перпендикулярно поверхности корня для измерения H + потоков в каждом направлении. Ортогональные векторы потоков суммировались в каждой позиции вдоль корня, чтобы создать двумерную карту суммарных потоков H + вдоль кончика корня. Как показано на рисунке 3, наблюдался сильный приток H + в апикальную область корня. Большая часть притока была локализована в области от 200 до 400 мкм назад от кончика корня (в зоне растяжения). Точка максимального потока находилась примерно в 250 мкм от кончика корня, где ток H + ориентирован перпендикулярно поверхности корня. Все остальные измеренные токи вдоль корня ориентированы на эту конкретную корневую зону, что позволяет предположить наличие либо сильного H + приток или локализованный отток H + -связывающего растворенного вещества в этой области. Корневой чехлик и более зрелые части корня поддерживают меньший чистый приток H + . Повторы этого эксперимента показали, что положение максимального притока H + несколько варьирует среди отдельных корней и находится в пределах 250–400 мкм позади кончика корня.
Ортогональные векторы потоков суммировались в каждой позиции вдоль корня, чтобы создать двумерную карту суммарных потоков H + вдоль кончика корня. Как показано на рисунке 3, наблюдался сильный приток H + в апикальную область корня. Большая часть притока была локализована в области от 200 до 400 мкм назад от кончика корня (в зоне растяжения). Точка максимального потока находилась примерно в 250 мкм от кончика корня, где ток H + ориентирован перпендикулярно поверхности корня. Все остальные измеренные токи вдоль корня ориентированы на эту конкретную корневую зону, что позволяет предположить наличие либо сильного H + приток или локализованный отток H + -связывающего растворенного вещества в этой области. Корневой чехлик и более зрелые части корня поддерживают меньший чистый приток H + . Повторы этого эксперимента показали, что положение максимального притока H + несколько варьирует среди отдельных корней и находится в пределах 250–400 мкм позади кончика корня.
Рис. 3.
Открыть в новой вкладкеСкачать слайд
Векторная диаграмма потоков сетки H + вдоль кончика корня арабидопсиса. Измерения ортогонального потока (показаны тонкими векторами для одной точки на расстоянии 600 мкм от кончика корня) проводились в нескольких точках на радиальном расстоянии 20 и 50 мкм от поверхности корня. Длина вектора представляет величину потока. Добавление компонентов ортогонального вектора определяет величину и направление сети H + тока (жирные векторы). Толщина корня не отображается в масштабе с измерениями потока.
Чтобы сравнить потоки корня H + между диким типом и alr- 104, мы сосредоточили наши измерения потока на области максимального притока (между 0 и 500 мкм от кончика корня). В этой области ток H + преимущественно перпендикулярен поверхности корня, что позволило измерить потоки микроэлектродом, вибрирующим под углом 90 o по отношению к корню. Данные от 8 до 12 корней усреднялись для учета различий между отдельными корнями. В питательном растворе без Al разницы в максимальном H 9 не было.Приток 1026 + между alr- 104 и диким типом (приблизительно 120 пмоль см -2 с -1 ) (фиг.4А). Кроме того, в регионах, прилегающих к точке максимального притока H + , не было обнаружено существенных различий в потоках между мутантным и диким типом.
Данные от 8 до 12 корней усреднялись для учета различий между отдельными корнями. В питательном растворе без Al разницы в максимальном H 9 не было.Приток 1026 + между alr- 104 и диким типом (приблизительно 120 пмоль см -2 с -1 ) (фиг.4А). Кроме того, в регионах, прилегающих к точке максимального притока H + , не было обнаружено существенных различий в потоках между мутантным и диким типом.
Рис. 4.
Открыть в новой вкладкеСкачать слайд
Влияние воздействия алюминия на чистый приток H + вдоль кончиков корней арабидопсиса. Приток H + измерялся вдоль линий дикого типа (□) и alr -104 (•) корней в отсутствие (A) и в присутствии (B) 300 мкм AlCl 3 . C, приток Root H + вдоль корней дикого типа (□) и alr -128 (▴) в присутствии 300 мкм AlCl 3 . Показан средний чистый приток и SE для 8-12 корней.
H + — измерения потока были повторены в присутствии 300 мкм AlCl 3 . Поскольку гелевая матрица связывает Al 3+ , активность Al 3+ при 300 мкм AlCl 3 в системе гель-слой сравнима с активностью Al 3+ в жидком питательном растворе, содержащем 30 мкМ AlCl 3 , на основании биоанализа активности Al 3+ (ингибирование роста корней у проростков дикого типа; данные не показаны). После 12 ч инкубации в Al-содержащем питательном растворе чистый приток H + в alr- 104 корней увеличился до 180 пмоль см -2 с -1 в точке максимума H + приток, тогда как приток в корнях дикого типа оставался примерно на уровне 100 пмоль см -2 с -1 (рис. 4Б). Это увеличение притока H + примерно на 80% можно постоянно наблюдать вдоль первых 500 мкм корня. Проростки Al-устойчивого мутанта alr -128, который является одним из четырех мутантов alr , картированных в один и тот же локус на хромосоме 1 ( alr -104 картируется на хромосоме 4), сравнивали с проростками дикого типа.
Поскольку гелевая матрица связывает Al 3+ , активность Al 3+ при 300 мкм AlCl 3 в системе гель-слой сравнима с активностью Al 3+ в жидком питательном растворе, содержащем 30 мкМ AlCl 3 , на основании биоанализа активности Al 3+ (ингибирование роста корней у проростков дикого типа; данные не показаны). После 12 ч инкубации в Al-содержащем питательном растворе чистый приток H + в alr- 104 корней увеличился до 180 пмоль см -2 с -1 в точке максимума H + приток, тогда как приток в корнях дикого типа оставался примерно на уровне 100 пмоль см -2 с -1 (рис. 4Б). Это увеличение притока H + примерно на 80% можно постоянно наблюдать вдоль первых 500 мкм корня. Проростки Al-устойчивого мутанта alr -128, который является одним из четырех мутантов alr , картированных в один и тот же локус на хромосоме 1 ( alr -104 картируется на хромосоме 4), сравнивали с проростками дикого типа. в отдельном эксперименте. Эти проростки, которые выделяли повышенное количество Al-связывающих органических кислот, не имели заметного увеличения H 9Приток 1026 + в присутствии Al (рис. 4С).
в отдельном эксперименте. Эти проростки, которые выделяли повышенное количество Al-связывающих органических кислот, не имели заметного увеличения H 9Приток 1026 + в присутствии Al (рис. 4С).
Al Влияние на рН ризосферы на кончике корня арабидопсиса
Чтобы проверить, оказывает ли измененный приток H + вдоль кончика корня alr- 104 значительное влияние на рН ризосферы (и, таким образом, на видообразование Al внутри этой области) рН поверхности вдоль верхушки корня определяли с помощью статического и вибрационного микроэлектродов Н + . Как показано на рисунке 5A, pH поверхности вдоль alr- 104 и кончиков корней дикого типа был между pH 4,3 и 4,4 в отсутствие Al (pH основного раствора был 4,2). Не было обнаружено существенной разницы рН между мутантом и диким типом в отсутствие Al. Когда корни подвергались воздействию 300 мкм Al, pH поверхности корней составлял alr- 104 увеличилось до 4,53, в то время как pH поверхности корней у проростков дикого типа оставался на уровне около 4,39 в области наибольшего притока (рис. 5B). Так, в присутствии Al alr -104 подщелачивает ризосферу на поверхности корня.
5B). Так, в присутствии Al alr -104 подщелачивает ризосферу на поверхности корня.
Рис. 5.
Открыть в новой вкладкеСкачать слайд
Влияние воздействия алюминия на рН ризосферы вдоль поверхности кончиков корней арабидопсиса. pH поверхности корня измеряли вдоль корней дикого типа (□) и alr .-104 (•) корней в отсутствие (A) и в присутствии (B) 300 мкм AlCl 3 . Показаны средние pH поверхности корней и se для 8-12 корней.
Устойчивость к алюминию в
alr -104 зависит от изменения рН ризосферы Были проведены эксперименты, чтобы определить, было ли небольшое вызванное алюминием увеличение рН ризосферы в alr -104 достаточным для повышения устойчивости к алюминию. Устойчивость к алюминию определяли путем измерения скорости роста корней в присутствии алюминия в alr -104 и дикий тип. Пятисуточные проростки арабидопсиса, выращенные в тонком слое геля, инкубировали в течение 12 ч в питательном растворе, содержащем 300 мкМ AlCl 3 , в присутствии и в отсутствие 10 мм Homo-Pipes (рН 4,4 и 4,5 соответственно). Ранее мы показали, что Homo-Pipes буферизует растворы в диапазоне pH от 4,0 до 4,5, не образуя комплексов с Al и не нарушая нормальный рост корней (Pellet et al., 1997). Используя вибрирующий микроэлектрод H + , мы обнаружили, что включение 10 мм Homo-Pipes в раствор для купания корней устраняло любой обнаруживаемый градиент pH вдоль кончика корня дикого типа и алр- 104 сеянцев (данные не приведены).
Ранее мы показали, что Homo-Pipes буферизует растворы в диапазоне pH от 4,0 до 4,5, не образуя комплексов с Al и не нарушая нормальный рост корней (Pellet et al., 1997). Используя вибрирующий микроэлектрод H + , мы обнаружили, что включение 10 мм Homo-Pipes в раствор для купания корней устраняло любой обнаруживаемый градиент pH вдоль кончика корня дикого типа и алр- 104 сеянцев (данные не приведены).
Скорость роста корней проростков определяли в алюминийсодержащей среде, которая была либо небуферной, либо буферной при рН 4,4 или 4,5 соответственно (рис. 6). Как было показано ранее, скорость роста alr -104 в небуферной среде превышала скорость роста дикого типа в присутствии Al, что согласуется с повышенной устойчивостью к Al. При повышении рН среды на 0,1 ед. рН (с 4,4 до 4,5) скорость роста как дикого типа, так и alr- 104 корней в присутствии Al увеличилось почти вдвое. Таким образом, увеличение рН ризосферы на 0,1 единицы приводило к значительному увеличению устойчивости к Al.
Рис. 6.
Открыть в новой вкладкеСкачать слайд
Влияние Al и pH на скорость роста корней проростков дикого типа (WT), alr -104 и alr -128, выращенных в небуферизованных и буферная среда. Проростки выращивали в тонком слое геля, уравновешенного питательным раствором, содержащим 300 мкМ AlCl 9 .1108 3 с трубками Homo-Pipe диаметром 10 мм или без них, отрегулированными до pH 4,4 или 4,5. Показаны средняя скорость роста корней и SE 12 проростков после 12 ч инкубации в соответствующей среде.
В этот эксперимент был включен еще один мутант, устойчивый к Al, alr- 128. Этот мутант демонстрирует степень устойчивости к Al, аналогичную alr- 104, и было показано, что он выделяет повышенное количество органических кислот в ризосферу (Larsen et al. др., 1998). В буферных условиях, которые устраняли устойчивость к алюминию в alr -104, стойкость Al alr -128 сохранялась (рис. 6). Это говорит о том, что устойчивость к Al у alr- 128 не зависит от изменения рН ризосферы, тогда как устойчивость к Al у alr- 104, по-видимому, связана с pH-опосредованным механизмом, при котором корни мутанта подщелачивают ризосферу. Повышенный рН ризосферы вокруг верхушки корня alr -104 должен привести к формированию Al в сторону менее токсичных видов Al, что снизит токсичность Al. Аналогичный темп роста alr- 104 и дикого типа в буферных условиях также демонстрирует, что повышенная устойчивость к Al alr- 104 объясняется исключительно механизмом, основанным на изменении рН ризосферы.
Повышенный рН ризосферы вокруг верхушки корня alr -104 должен привести к формированию Al в сторону менее токсичных видов Al, что снизит токсичность Al. Аналогичный темп роста alr- 104 и дикого типа в буферных условиях также демонстрирует, что повышенная устойчивость к Al alr- 104 объясняется исключительно механизмом, основанным на изменении рН ризосферы.
Исследования роста корней были повторены без добавления Al, чтобы определить, влияет ли разница в кислотном стрессе между pH 4,4 и 4,5 на скорость роста корней. Результаты, показанные на рисунке 7, показывают, что эта небольшая разница рН в питательном растворе сама по себе не оказывает существенного влияния на скорость роста корней.
Рис. 7.
Открыть в новой вкладкеСкачать слайд
Влияние рН ризосферы на рост корней арабидопсиса в рН-буферной среде. Проростки дикого типа (WT) и alr -104 выращивали в тонком слое геля, уравновешенном средой, содержащей 10 мм Homo-Pipes, при pH 4,4 или 4,5. Показаны средняя скорость роста корней и SE 12 проростков после 12 ч инкубации в соответствующей среде.
Показаны средняя скорость роста корней и SE 12 проростков после 12 ч инкубации в соответствующей среде.
ОБСУЖДЕНИЕ
H
+ — Измерения потока вдоль кончика корня арабидопсисаВ этом исследовании мы исследовали механизм, который придает повышенную устойчивость к Al у мутанта арабидопсиса alr -104. Поскольку устойчивость к Al у alr -104 не связана с повышенным высвобождением органических кислот (Larsen et al., 1998), мы исследовали возможность того, что этот мутант изменяет pH ризосферы, чтобы исключить попадание токсичных ионов Al 3+ в корень. . Этот рН-опосредованный механизм устойчивости к алюминию часто выдвигался в литературе в качестве гипотезы, но его существование у наземных растений не было убедительно доказано.
Необходимо было модифицировать метод внеклеточного вибрирующего микроэлектрода для изучения потоков ионов в очень маленьких корнях арабидопсиса, погруженных в гелевую матрицу низкой плотности. Чтобы свести к минимуму нарушение геля, электрод вибрировали перпендикулярно корню и вдоль длинной оси микроэлектрода. Приток H + измеряли вдоль кончика корня, который достиг примерно 120 пмоль см -2 с -1 в точке максимального притока. Двумерное отображение H + притока показали, что большая часть притока локализована в пределах довольно небольшой области в зоне удлинения корня, длиной примерно от четырех до восьми эпидермальных клеток корня. Пространственная картина суммарного тока H + на кончике корня арабидопсиса была сходной с таковой общего ионного тока (выведенного из измерений внеклеточных электрических полей), измеренного у других видов, включая ячмень (Weisenseel et al., 1979), Lepidum. sativum (Behrens et al., 1982), Trifolium repens (Miller et al., 1986) и табак (Miller et al., 1988).
Чтобы свести к минимуму нарушение геля, электрод вибрировали перпендикулярно корню и вдоль длинной оси микроэлектрода. Приток H + измеряли вдоль кончика корня, который достиг примерно 120 пмоль см -2 с -1 в точке максимального притока. Двумерное отображение H + притока показали, что большая часть притока локализована в пределах довольно небольшой области в зоне удлинения корня, длиной примерно от четырех до восьми эпидермальных клеток корня. Пространственная картина суммарного тока H + на кончике корня арабидопсиса была сходной с таковой общего ионного тока (выведенного из измерений внеклеточных электрических полей), измеренного у других видов, включая ячмень (Weisenseel et al., 1979), Lepidum. sativum (Behrens et al., 1982), Trifolium repens (Miller et al., 1986) и табак (Miller et al., 1988).
У устойчивого к Al мутанта Arabidopsis alr -104 повышенный приток H + индуцировался Al. Несмотря на изменение интенсивности потока, изменения пространственной картины притока H + не наблюдалось. Поэтому вполне вероятно, что процессы ионного транспорта в alr -104 такие же, как и в кончике корня дикого типа, но сильнее в присутствии Al. Однако природа повышенного чистого H 9Приток 1026 + в alr -104 еще предстоит определить. Поскольку мы измеряем чистое поглощение H + верхушкой корня, стимуляция этого потока может быть вызвана стимуляцией однонаправленного притока H + или уменьшением оттока H + . Уменьшение оттока H + предположительно должно быть вызвано взаимодействием Al с плазматической мембраной H + -ATPase. Мутации в механизме регуляции H + -ATPase в alr -104 могут обеспечивать прямой или косвенный эффект Al.
Несмотря на изменение интенсивности потока, изменения пространственной картины притока H + не наблюдалось. Поэтому вполне вероятно, что процессы ионного транспорта в alr -104 такие же, как и в кончике корня дикого типа, но сильнее в присутствии Al. Однако природа повышенного чистого H 9Приток 1026 + в alr -104 еще предстоит определить. Поскольку мы измеряем чистое поглощение H + верхушкой корня, стимуляция этого потока может быть вызвана стимуляцией однонаправленного притока H + или уменьшением оттока H + . Уменьшение оттока H + предположительно должно быть вызвано взаимодействием Al с плазматической мембраной H + -ATPase. Мутации в механизме регуляции H + -ATPase в alr -104 могут обеспечивать прямой или косвенный эффект Al.
Очевидный повышенный приток H + в alr -104 также может быть связан с оттоком растворенных веществ, которые протонируются при их высвобождении в кислую ризосферу. Что касается органических и неорганических кислот, было обнаружено, что только пируват высвобождается с большей скоростью у alr -104, чем у дикого типа (см. Ларсен и др., 1998). Однако повышенное высвобождение пирувата было конститутивным и не индуцировалось Al в alr -104. Кроме того, пируват не подвергается значительному протонированию при транспортировке из нейтральной цитоплазмы в ризосферу с рН около 4,0, и поэтому маловероятно, что он окажет влияние на рН ризосферы.
Что касается органических и неорганических кислот, было обнаружено, что только пируват высвобождается с большей скоростью у alr -104, чем у дикого типа (см. Ларсен и др., 1998). Однако повышенное высвобождение пирувата было конститутивным и не индуцировалось Al в alr -104. Кроме того, пируват не подвергается значительному протонированию при транспортировке из нейтральной цитоплазмы в ризосферу с рН около 4,0, и поэтому маловероятно, что он окажет влияние на рН ризосферы.
Увеличение чистого притока H + в alr -104 также может быть вызвано изменением транспортной системы, связанной с H + (H + symport или antiport). Одним из возможных изменений является поглощение N (как NO 3 — или NH 4 + ), которое тесно связано с транспортом H + . Измерения pH нерасфасованного раствора показали, что поглощение NO 3 − и NH 4 + часто ассоциируется с изменениями pH (обзор см. в Miller and Smith, 1996). Во многих из этих исследований интерпретация часто затруднена, потому что NO 3 — и/или NH 4 + в конечном итоге истощались в гидропонной среде, вызывая резкие изменения pH основного раствора. В описанных здесь экспериментах корни растений арабидопсиса уравновешивали большим объемом питательного раствора, обеспечивая постоянный NH 4 + /NO 3 − соотношение. Возможно, однако, что вызванное Al различие в поглощении NH 4 + или NO 3 − в alr -104 является причиной изменения рН ризосферы. То есть возможно, что в alr -104 воздействие Al стимулирует приток NO 3 − или ингибирует поглощение NH 4 + , что, в свою очередь, может увеличить рН ризосферы.
в Miller and Smith, 1996). Во многих из этих исследований интерпретация часто затруднена, потому что NO 3 — и/или NH 4 + в конечном итоге истощались в гидропонной среде, вызывая резкие изменения pH основного раствора. В описанных здесь экспериментах корни растений арабидопсиса уравновешивали большим объемом питательного раствора, обеспечивая постоянный NH 4 + /NO 3 − соотношение. Возможно, однако, что вызванное Al различие в поглощении NH 4 + или NO 3 − в alr -104 является причиной изменения рН ризосферы. То есть возможно, что в alr -104 воздействие Al стимулирует приток NO 3 − или ингибирует поглощение NH 4 + , что, в свою очередь, может увеличить рН ризосферы.
Из-за сложной потребности в питательных веществах для поддержания роста корней арабидопсиса будет трудно идентифицировать процесс переноса ионов, связанный с устойчивостью к алюминию в алр -104. Мутация в alr -104 представляет собой единственную мутацию на хромосоме 4, которая была выделена из популяции семян, подвергнутых мутагенезу этилметилсульфонатом. Следовательно, вероятно, это точечная мутация, которая вызывает потерю функции, хотя индуцируемость притока H + с помощью Al предполагает усиление функции. Другой пример индуцируемости Al-резистентности можно найти в работе Delhaize et al. (1993a, 1993b) с пшеницей, в которой локус Al-резистентности 9Было показано, что 1048 Alt1 придает индуцированную алюминием устойчивость к алюминию на основе высвобождения органической кислоты. Клонирование гена alr -104 у арабидопсиса в настоящее время проводится в наших лабораториях и, в случае успеха, может пролить некоторый свет на механизм повышения рН ризосферы, вызванного алюминием.
Мутация в alr -104 представляет собой единственную мутацию на хромосоме 4, которая была выделена из популяции семян, подвергнутых мутагенезу этилметилсульфонатом. Следовательно, вероятно, это точечная мутация, которая вызывает потерю функции, хотя индуцируемость притока H + с помощью Al предполагает усиление функции. Другой пример индуцируемости Al-резистентности можно найти в работе Delhaize et al. (1993a, 1993b) с пшеницей, в которой локус Al-резистентности 9Было показано, что 1048 Alt1 придает индуцированную алюминием устойчивость к алюминию на основе высвобождения органической кислоты. Клонирование гена alr -104 у арабидопсиса в настоящее время проводится в наших лабораториях и, в случае успеха, может пролить некоторый свет на механизм повышения рН ризосферы, вызванного алюминием.
Различия pH на поверхности корня ответственны за устойчивость к алюминию в
alr -104 По мере увеличения pH раствора с 4,0 до 5,0 состав алюминия быстро меняется с токсичного Al 3+ видов в менее токсичные гидроксиды алюминия и осадки алюминия, так что небольшие изменения рН могут привести к значительным изменениям токсичности алюминия (Martell and Motekaitis, 1989). Измерения рН на поверхности корней дикого типа и alr -104 выявили разницу от 0,1 до 0,15 единиц рН вдоль верхушки корня. Было показано, что этот участок корня является основным местом токсичности Al в корнях (Ryan et al., 1993). В предыдущих исследованиях различия рН ризосферы аналогичной величины были обнаружены вдоль кончиков корней устойчивого к Al сорта пшеницы Atlas 66 и чувствительного к Al сорта Scout, выращенных в питательном растворе с Al (Miyasaka et al., 19).89; Пелле и др., 1997).
Измерения рН на поверхности корней дикого типа и alr -104 выявили разницу от 0,1 до 0,15 единиц рН вдоль верхушки корня. Было показано, что этот участок корня является основным местом токсичности Al в корнях (Ryan et al., 1993). В предыдущих исследованиях различия рН ризосферы аналогичной величины были обнаружены вдоль кончиков корней устойчивого к Al сорта пшеницы Atlas 66 и чувствительного к Al сорта Scout, выращенных в питательном растворе с Al (Miyasaka et al., 19).89; Пелле и др., 1997).
Различия от 0,1 до 0,2 единицы рН также были отмечены при измерениях в нерасфасованном растворе для нескольких других сортов пшеницы (Taylor and Foy, 1985a, 1985b). Во всех этих исследованиях не было показано, приводят ли небольшие различия pH к повышенной устойчивости к Al. Важно отметить, что величина различий рН, измеренных на поверхности корня, может быть даже больше на поверхности плазматической мембраны внутри клеточной стенки. Поскольку градиент pH создается на плазматической мембране клеток корня, электрически заряженная клеточная стенка может действовать как барьер для высвобождения ионов. Таким образом, возможно, что разница рН между поверхностью плазматической мембраны и внешним раствором больше, чем в неперемешиваемом слое, прилегающем к корню.
Таким образом, возможно, что разница рН между поверхностью плазматической мембраны и внешним раствором больше, чем в неперемешиваемом слое, прилегающем к корню.
Чтобы определить, действительно ли устойчивость к Al у alr -104 обусловлена небольшим (0,1–0,2 ед. с высокой концентрацией Homo-Pipes. Степень буферизации была достаточной, чтобы избежать образования градиентов pH на поверхности корня, но мы не знаем, насколько далеко буферность распространяется на корневой апопласт. Homo-Pipes — это биологический буфер с pK 4,29.и мы обнаружили, что он не связывает Al 3+ (данные не показаны).
Из этих исследований роста корней были получены две важные части информации. Во-первых, когда мы устранили градиенты рН на поверхности корней с помощью Homo-Pipes, устойчивость к алюминию в alr -104 исчезла, что позволяет предположить, что повышенная устойчивость к алюминию в alr -104 является исключительно результатом механизма, основанного на изменении рН в ризосфере. Во-вторых, когда pH ризосферы был забуферен до pH 4,5, наблюдалось значительное увеличение устойчивости к Al по сравнению с результатами исследований токсичности Al, проведенных в растворе, забуференном до pH 4,4. Эти результаты показывают, что в alr -104, небольшое увеличение рН ризосферы, вызванное алюминием (0,1–0,2 единицы), достаточно для объяснения наблюдаемой устойчивости к алюминию. В исследовании alr -128, в котором мы продемонстрировали корреляцию между устойчивостью к алюминию, выведением алюминия из кончика корня и повышенным высвобождением малата и цитрата (Larsen et al., 1998), мы смогли показать, что устойчивость к алюминию не не связаны с изменением рН ризосферы.
Во-вторых, когда pH ризосферы был забуферен до pH 4,5, наблюдалось значительное увеличение устойчивости к Al по сравнению с результатами исследований токсичности Al, проведенных в растворе, забуференном до pH 4,4. Эти результаты показывают, что в alr -104, небольшое увеличение рН ризосферы, вызванное алюминием (0,1–0,2 единицы), достаточно для объяснения наблюдаемой устойчивости к алюминию. В исследовании alr -128, в котором мы продемонстрировали корреляцию между устойчивостью к алюминию, выведением алюминия из кончика корня и повышенным высвобождением малата и цитрата (Larsen et al., 1998), мы смогли показать, что устойчивость к алюминию не не связаны с изменением рН ризосферы.
Повышенная устойчивость к Al мутанта Arabidopsis alr -104, по-видимому, вызвано подщелачиванием ризосферы, вызванным алюминием. Это повышенное подщелачивание локализовано в кончике корня, который является местом токсичности алюминия. Хотя этот механизм часто предлагался в литературе, эти результаты являются первым убедительным доказательством наших знаний о механизме устойчивости к Al, включающем рН-барьер ризосферы у высших растений. Этот механизм устойчивости к алюминию в alr -104 отличается от ранее описанных механизмов устойчивости к алюминию, которые были основаны на экссудации органических кислот, хелатирующих алюминий. В будущих исследованиях мы изучим роль ионно-транспортных процессов в Al-индуцированном подщелачивании ризосферы. Мы также уделяем особое внимание изоляции alr -104 путем клонирования на основе карты, чтобы лучше понять этот механизм толерантности к Al на молекулярном уровне.
Этот механизм устойчивости к алюминию в alr -104 отличается от ранее описанных механизмов устойчивости к алюминию, которые были основаны на экссудации органических кислот, хелатирующих алюминий. В будущих исследованиях мы изучим роль ионно-транспортных процессов в Al-индуцированном подщелачивании ризосферы. Мы также уделяем особое внимание изоляции alr -104 путем клонирования на основе карты, чтобы лучше понять этот механизм толерантности к Al на молекулярном уровне.
БЛАГОДАРНОСТЬ
Авторы благодарят Джона Э. Шаффа за щедрую помощь с техническими аспектами вибрационного зонда. Доктор Питер Дж.С. Мы выражаем благодарность Смиту за советы по измерению эффективности.
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
1
Аниол
A
Индукция толерантности к алюминию у проростков пшеницы с помощью низких доз алюминия в питательном растворе.
Plant Physiol
123
1984
223
227
2
Behrens
HM
Weisenseel
MH
Sievers
A
Rapid changes in the pattern of electrical current around the root наконечник Lepidum sativum L. после гравистимуляции.
после гравистимуляции.
Завод Физиол
70
1982
1079
1083
3
Резчик
BF
Ownby
JD
Устойчивость пшеницы к кислой почве.
Adv Agron
54
1995
117
173
4
Crank
J
The Maiffusion
1975
Clarendon Press
Oxford, UK
5
Delhaize
E
Craig
S
Beaton
CD
Bennet
RJ
Джагадиш
VC
Randall
PJ
Устойчивость пшеницы к алюминию ( Triticum aestivum L.). I. Поглощение и распределение алюминия в верхушках корней.
Plant Physiol
103
1993a
685
693
6
Delhaize
E
Ryan
PR
Aluminum toxicity and tolerance in plants.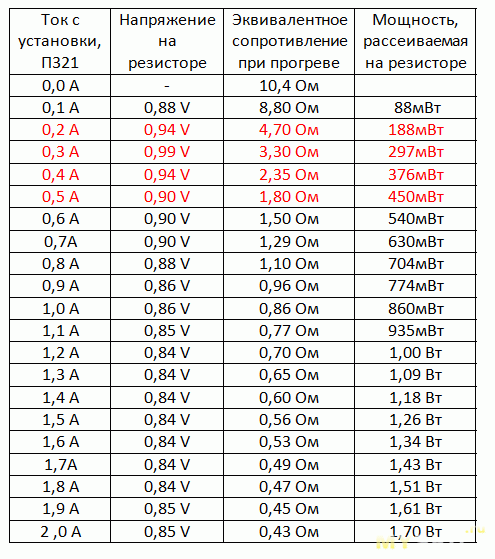
Завод Физиол
107
1995
315
321
7
Delhaize
E
Ryan
PR
. II. Стимулируемое алюминием выделение яблочной кислоты из верхушек корней.
Plant Physiol
103
1993b
695
702
8
Dodge
CS
Hiatt
AJ
Relationship of pH to ion uptake imbalance by varieties of wheat ( Triticum vulgare ).
Agron J
64
1992
476
481
9
Fleming
ALсорта пшеницы, отличающиеся устойчивостью к аммонию
AL
Agron J
75
1983
726
730
10
Burns
GR
Brown
JC
Fleming
AL
Differential aluminum tolerance of two wheat varieties associated with plant- индуцированные изменения рН вокруг их корней.
СВЯЗА SCI SOC AM PROC
29
1965
64
67
11
Fleming
AL
ALUMEN -ALERASE.
J Plant Nutr
5
1982
1313
1333
12
Fleming
AL
Burns
GR
Arninger
WH
Characterization of differential aluminum tolerance among varieties of wheat and ячмень.
Soil Sci Soc Am Proc
31
1967
513
521
13
Lafever
HN
Schwartz
JW
Fleming
AL
Aluminum tolerance of wheat cultivars related to region происхождения.
Agron J
66
1974
751
758
14
SACELIVE
LF
SACEDIMEL.
Proc 9th Annu Conf IEEE
9
1987
779
781
15
Jones
DL
Kochian
LV
Aluminum inhibition of the 1,4,5-trisphosphate signal transduction pathway в корнях пшеницы: роль в токсичности алюминия?
Растительная клетка
7
1995
1913
1922
16
Климашевская
ЭЛ
0137
МЛ
Активность АТФазы и кислой фосфатазы в зонах роста корней двух сортов гороха с разной устойчивостью к токсичным ионам Al.
SOV Plant Physiol
20
1973
257
263
17
Kochian
LV
.
Annu Rev Plant Physiol Plant Mol Biol
46
1995
237
260
18
Kochian
LV
Shaff
JE
Kühtreiber
WM
Jaffe
LF
Lucas
WJ
Use of an extracellular, ion-selective vibrating microelectrode system for the количественная оценка потоков K + , H + и Ca 2+ в корнях кукурузы и суспензионных клетках кукурузы.
Планта
188
1992
601
610
19
Kühtreiber
WM
Jaffe
LF
Обнаружение градиентов внеклеточного кальция с помощью кальций-специфического вибрирующего электрода.
J Cell Biol
110
1990
1565
1573
20
Larsen
PB
Degenhardt
J
Tai
C-Y
Stenzler
LM
Howell
SH
Kochian
LV
Мутанты арабидопсиса с повышенной устойчивостью к алюминию демонстрируют измененные модели накопления алюминия и выделения органических кислот из корней.
Plant Physiol
117
1998
9
17
21
Larsen
PB
Tai
C-Y
Kochian
LV
Howell
SH
Arabidopsis mutants with increased sensitivity к алюминию.
Plant Physiol
110
1996
743
751
22
Lucas
WJ
Kochian
LV
Ion transport processes in corn roots: an approach utilizing microelectrode techniques.
Gensler
WG
Передовые сельскохозяйственные инструменты: проектирование и использование.
1986
402
425
Мартинус Нейхофф
Дордрехт, Нидерланды
23
Martell AE, Motekaitis RJ (1989) Координационная химия и спецификация Al(III) в водном растворе. В Т. Е. Льюис, изд. Химия окружающей среды и токсикология алюминия. Lewis Publishers, Chelsea, MI, pp 3–17
Химия окружающей среды и токсикология алюминия. Lewis Publishers, Chelsea, MI, pp 3–17
24
Miller
AL
Raven
JA
Sprent
JI
Weisenseel
MH
Endogenous ion currents traverse growing roots and root hairs of Trifolium repens .
Plant Cell Environ
9
1986
79
83
25
Miller
AL
Shand
E
Gow
NAR
Ion currents associated with root tips, emerging laterals and индуцированные раневые участки в Nicotiana tabacum : предполагаемая пространственная связь между результирующими электрическими полями и заражением фитофторановыми зооспорами.
Среда растительных клеток
11
1988
21
25
26
Miller
AJ
Smith Tranpers
.
J Exp Bot
47
1996
843
854
27
Miyasaka
SC
Kochian
LV
Shaff
JE
Mechanisms of aluminum tolerance in wheat. Исследование генотипических различий рН ризосферы, К + и H + транспорт и мембранные потенциалы корневых клеток.
Plant Physiol
91
1989
1188
1196
28
Mugwira
LM
Elgawahry
SM
Aluminum accumulation and tolerance of triticale and wheat in relation to root cation exchange capacity.
Почвоведение Soc Am J
43
1979
736
740
29
Mugwira
LM
Elgawahry
SM
Patel
SU
Различная устойчивость тритикале, пшеницы, ржи и ячменя к алюминию в питательном растворе.
Agron J
68
1976
782
786
30
Mugwira
LM
Elgawahry
SM
Patel
SU
Aluminum tolerance in triticale, wheat and rye as measured by характеристики роста корней и концентрация алюминия.
Plant Soil
50
1978
681
690
31
Mugwira
LM
Patel
SU
Root zone pH changes and ion uptake imbalances by triticale, wheat, and rye.
Agron J
69
1977
719
722
32
Pellet
DM
Grunes
DL
Kochian
LV
Organic acid exudation as an aluminum tolerance mechanism in maize ( Zea Mays L.).
Planta
196
1994
788
795
33
Pellet
DM
Papernik
LA
Jones
DL
Darrah
PR
Grunes
DL
Kochian
LV
Участие множественных механизмов исключения алюминия в устойчивости пшеницы к алюминию.
Почва для растений
192
1997
63
68
34
Ryan
PR
Delhaize
E
Randall
PJ
Malate efflux from root apices: evidence for a general mechanism of Al-tolerance in wheat .
Aust J Plant Physiol
22
1995
531
536
35
Ryan
PR
DiTomaso
JM
Kochian
LV
Токсичность алюминия в корнях: исследование пространственной чувствительности и роли корневого чехлика.
J Exp Bot
44
1993
437
446
36
Smith
PJS
Sanger
RH
Jaffe
LF
The vibrating Ca 2+ electrode: a новый метод обнаружения областей плазматической мембраны Ca 2+ притока и оттока.
Методы Cell Biol 9(0137
40
1994
115
134
37
Taylor
GJ
Can J Bot
66
1988
694
699
38
Тэйлор
алюминиевая основа реакции на стресс.
Curr Top Plant Biochem Physiol
10
1991
57
93
39
Taylor
GJ
Mechanisms of aluminum tolerance in Triticum aestivum L. (wheat). I. Дифференциальный рН, вызываемый зимними сортами в питательных растворах.
Am J Bot
72
1985a
695
701
40
Механизм Taylor
GJ 9013 допуск 7 из алюминия
 org/Product»>048 Triticum aestivum L. (пшеница). II. Дифференциальный рН, вызываемый весенними сортами в питательных растворах.
org/Product»>048 Triticum aestivum L. (пшеница). II. Дифференциальный рН, вызываемый весенними сортами в питательных растворах. Am J Bot
72
1985b
702
706
41
Von Uexkull
HR
Mutert
E
Global extent, development and economic impact of acid soils.
Почва для растений
171
1995
1
15
42
Weisenseel
MH
Jaffe
LF
Natural H + токи пересекают растущие корни и корневые волоски ячменя ( Hordeum vulgare L.).
Plant Physiol
64
1979
512
518
Автор Примечания
1
. Эта работа была подтверждена. -010 к S.H.H. и L.V.K.).
*
Автор корреспонденции; электронная почта lvk1@cornell. edu; факс 1–607–255–2459.
edu; факс 1–607–255–2459.
Copyright © 1998 Американское общество физиологов растений
Эта статья публикуется и распространяется в соответствии с условиями публикации Oxford University Press, Standard Journals Publication Model (https://academic.oup.com/journals/pages/open_access/funder_policies/ chorus/standard_publication_model)
Copyright © 1998 Американское общество физиологов растений
RM3216B-104/204-PBVW10 техническое описание — Технические характеристики: Сопротивление (Ом): 100K, 200K; Допуск:
Детали, спецификация, цитата по номеру детали: RM3216B-104/204-PBVW10
| Деталь | RM3216B-104/204-PBVW10 |
| Категория | Резисторы |
| Название | Резистор 100K, 200K Ом 62,5 мВт 2 резистора [количество контактов] Штыревые сети, массивы |
| Описание | |
| Компания | Сусуму (Вирджиния) |
| Технический паспорт | Загрузить RM3216B-104/204-PBVW10 Лист данных |
| Specifications | |
| Resistance (Ohms) | 100K, 200K |
| Tolerance | 0. 1% 1% |
| Power Per Element | 62.5mW |
| Circuit Type | Изолированный |
| Количество контактов | 4 |
| Упаковка | Cut Tape (CT) |
| Number of Resistors | 2 |
| Package / Case | 1206 (3216 Metric) |
| Mounting Type | Surface Mount |
| Temperature Coefficient | 25ppm/C |
| Статус без свинца | Без свинца |
| Статус RoHS | Соответствует RoHS |
 Тип Диапазон сопротивления Абсолютное значение Доп. %(код)
Тип Диапазон сопротивления Абсолютное значение Доп. %(код) Отслеживание TCR ppm/ C(код) Макс. рабочее напряжение Пакет
0,02(P),0,05(W) (1 соотношение 0,02(P),0,05(W) (1 соотношение 0,05(W) (100 соотношение 0,05(W) (100 соотношение 2(W),5(V) (1 соотношение 2(W),5(V) (1 соотношение 5(V) (100 соотношение 5(V) (100 соотношение 1000 шт./катушка(10), 5000 шт./катушка(50)
Пожалуйста, свяжитесь с нами для TCR 5ppm/ C для 300 или более в RM2012, RM3216 Пожалуйста, свяжитесь с нами для другого разнообразия, кроме этих размеров и индивидуальных спецификаций.
Срок службы 0,15 0,10 Изменение сопротивления (%) Время (ч) Абсолютное отслеживание Условия испытаний: 90 мин ВКЛ/ 30 мин ВЫКЛ Образец: 0,15 0,10 Изменение сопротивления (%) Время (ч) Абсолютное отслеживание Условия испытаний: 90 мин ВКЛ/ 30 мин ВЫКЛ Образец: RM3216B-203/203 Срок службы под нагрузкой (85C)
Температура Влажность Смещение (85C Изменение сопротивления (%) Время (ч) Абсолютное отслеживание Условия испытаний: 90 мин ВКЛ/ 30 мин ВЫКЛ Образец: 0,15 0,10 Изменение сопротивления (%) Время (ч) Абсолютное отслеживание Условия испытаний: 90 мин ВКЛ/ 30 мин ВЫКЛ Образец: RM3216B-203/ 203 Смещение температуры и влажности (85C 85%)
Хранение при высокой температуре 0,15 0,10 Изменение сопротивления (%) Время (ч) Абсолютное изменение сопротивления отслеживания (%) Условия испытаний: Ta155C Без нагрузки Образец: RM2012B-103/103 Время (ч) Абсолютное отслеживание Условия испытаний: Ta155C Без нагрузки Образец: RM3216B -203/203 Высокотемпературное хранение (155°C)
Отслеживание упаковки Допуск сопротивления Отслеживание температурного коэффициента сопротивления Допуск температурного коэффициента сопротивления Размеры цепи Код детали
| Связанные продукты с тем же паспортом |
| РМ3216Б-104/204-ПБВВ10 |
| РМ3216Б-104/304-ПБВВ10 |
| РМ3216Б-104/304-ПБВВ10 |
| Номер детали того же производителя Susumu (VA) |
| RM3216B-104/304-PBVW10 Технические характеристики: Сопротивление (Ом): 100K, 200K; Допуск: 0,1%; Мощность на элемент: 62,5 мВт; Тип цепи: изолированный; Количество контактов: 4; Упаковка: Cut Tape (CT); Количество резисторов: 2; Упаковка/кейс: |
| RM3216B-202/103-PBVW10 Технические характеристики: Сопротивление (Ом): 2K, 10K; Допуск: 0,1%; Мощность на элемент: 62,5 мВт; Тип цепи: изолированный; Количество контактов: 4; Упаковка: Cut Tape (CT); Количество резисторов: 2; Упаковка/кейс: |
RM3216B-212/492-N48 Технические характеристики: Сопротивление (Ом): 2,1 кОм. 4,9К; Толерантность: — ; Мощность на элемент: 62,5 мВт; Тип цепи: изолированный; Количество контактов: 4; Упаковка: лента и катушка (TR); Количество резисторов: 2; Пакет 4,9К; Толерантность: — ; Мощность на элемент: 62,5 мВт; Тип цепи: изолированный; Количество контактов: 4; Упаковка: лента и катушка (TR); Количество резисторов: 2; Пакет |
| RM3216B-502/104-PBVW10 Технические характеристики: Сопротивление (Ом): 5K, 100K; Допуск: 0,1%; Мощность на элемент: 62,5 мВт; Тип цепи: изолированный; Количество контактов: 4; Упаковка: Cut Tape (CT); Количество резисторов: 2; Упаковка/кейс: |
| RM3216F-N50 Технические характеристики: Сопротивление (Ом): 500, 3,02 К; Толерантность: — ; Мощность на элемент: 62,5 мВт; Тип цепи: изолированный; Количество контактов: 4; Упаковка: лента и катушка (TR); Количество резисторов: 2; Пакет |
| RGh3012-2E-P-362-B Технические характеристики: Сопротивление (Ом): 3K ; Мощность (Ватт): 0,25 Вт, 1/4 Вт; Допуск: 0,1%; Упаковка: Cut Tape (CT); Состав: Тонкая пленка; Температурный коэффициент: 25ppm/C; Статус без свинца: без свинца |
| RG1608N-362-B-T1 Технические характеристики: Сопротивление (Ом): 3,6 кОм; Мощность (Ватт): 0,1 Вт, 1/10 Вт; Допуск: 0,1%; Упаковка: лента и катушка (TR); Состав: Тонкая пленка; Температурный коэффициент: 10ppm/C; Безсвинцовый статус: |
| RG1608N-362-P-T1 |
| RG1608N-362-W-T1 |
| RG1608P-362-B-T1 |
| RG1608P-362-P-T1 |
| RG1608P-362-W-T1 |
| RG1608V-362-B-T1 |
| RG1608V-362-P-T1 |
| RG1608V-362-W-T1 |
| RG1005N-362-B-T1 Технические характеристики: Сопротивление (Ом): 3,6 кОм; Мощность (Ватт): 0,063 Вт, 1/16 Вт; Допуск: 0,1%; Упаковка: лента и катушка (TR); Состав: Тонкая пленка; Температурный коэффициент: 10ppm/C; Безсвинцовый статус: |
| RG1005N-362-C-T10 |
| RG1005N-362-W-T1 |
| RG1005P-362-B-T1 |
| RG1005P-362-C-T10 |
| RG1005P-362-W-T1 |
|
RL3264SW4-006M-F-T5 : Чип-резистор 0,001 Ом, 2 Вт — поверхностный монтаж; RES 0,001 Ом 2 Вт 1% 2512 SMD Технические характеристики: Сопротивление (Ом): 0,001 Ом; Мощность (Ватт): 2 Вт; Допуск: 1%; Упаковка: Cut Tape (CT); Состав: Тонкая пленка; Температурный коэффициент: 300ppm/C; Статус без свинца: без свинца; Статус RoHS: Соответствует RoHS RL1220T-6R8-G : 6,8 Ом, 0,333 Вт, чип-резистор 1/3 Вт — поверхностный монтаж; RES 6,8 OHM 1/3 Вт 2% 0805 Технические характеристики: Сопротивление (Ом): 6,8 Ом; Мощность (Ватт): 0,333 Вт, 1/3 Вт; Допуск: 2%; Упаковка: лента и катушка (TR); Состав: Тонкая пленка; Температурный коэффициент: 0/+350ppm/C; Статус без свинца: без свинца; Статус RoHS: соответствует RoHS RG1005N-2431-W-T5 : 2,43 кОм, 0,063 Вт, чип-резистор 1/16 Вт — поверхностный монтаж; RES 2. RG2012V-9760-P-T1: 976 Ом, 0,125 Вт, чип-резистор 1/8 Вт — поверхностный монтаж; RES 976 OHM 1/8W .1% 0805 Характеристики: Сопротивление (Ом): 976; Мощность (Ватт): 0,125 Вт, 1/8 Вт; Допуск: 0,1%; Упаковка: лента и катушка (TR); Состав: Тонкая пленка; Температурный коэффициент: 5ppm/C; Статус без свинца: без свинца; Статус RoHS: Соответствует RoHS RM3216B-502/104-PBVW10: Резистор 5 кОм, 100 кОм, 62,5 мВт 2 резистора [количество контактов] Контакты Сети, массивы; RES MULT OHM 2 RES 1206 Технические характеристики: Сопротивление (Ом): 5K, 100K; Допуск: 0,1%; Мощность на элемент: 62,5 мВт; Тип цепи: изолированный; Количество контактов: 4; Упаковка: Cut Tape (CT); Количество резисторов: 2; Упаковка/кейс: 1206 (метрическая 3216); Тип монтажа: поверхностный монтаж; Температурный коэффициент: 25ppm/C; Ведущий RR0510P-3320-D : 332 Ом, 0,063 Вт, чип-резистор 1/16 Вт — поверхностный монтаж; RES 332 OHM 1/16W 0. PAT0816-C-3DB-T : 0 МГц — 10000 МГц ФИКСИРОВАННЫЙ ВЧ/СВЧ-АТТЕНЮАТОР Технические характеристики: Тип аттенюатора: Фиксированный; Частота: от 0,0 до 10 ГГц; КСВ: 1,3 1 ; Затухание: 0,0 дБ; Тип упаковки: для поверхностного монтажа, SURFACE MOUNT PACKAGE-3 PAT0816C-10DB-B-H-LF : 0 МГц — 10000 МГц ФИКСИРОВАННЫЙ ВЧ/СВЧ-АТТЕНЮАТОР Технические характеристики: Тип аттенюатора: Фиксированный; Частота: от 0,0 до 10 ГГц; КСВ: 1,3 1 ; Затухание: 0,0 дБ; Тип упаковки: для поверхностного монтажа, SURFACE MOUNT PACKAGE-3 HRG3216P-2490-B-T1 : Тонкопленочные резисторы — SMD 1,0 Вт 249 Ом 0,1% 1206 25ppm Тонкопленочные резисторы серии Susumu HRG представляют собой резисторы высокой мощности с короткой боковой обмоткой вокруг выводов. Похожие записи
|
 43K OHM 1/16W .1% 0402 Характеристики: Сопротивление (Ом): 2.43K; Мощность (Ватт): 0,063 Вт, 1/16 Вт; Допуск: 0,1%; Упаковка: лента и катушка (TR); Состав: Тонкая пленка; Температурный коэффициент: 10ppm/C; Статус без свинца: без свинца; Статус RoHS: Соответствует RoHS
43K OHM 1/16W .1% 0402 Характеристики: Сопротивление (Ом): 2.43K; Мощность (Ватт): 0,063 Вт, 1/16 Вт; Допуск: 0,1%; Упаковка: лента и катушка (TR); Состав: Тонкая пленка; Температурный коэффициент: 10ppm/C; Статус без свинца: без свинца; Статус RoHS: Соответствует RoHS  5% 0402 SMD Характеристики: Сопротивление (Ом): 332 Ом; Мощность (Ватт): 0,063 Вт, 1/16 Вт; Допуск: 0,5%; Упаковка: Digi-Reel; Состав: Тонкая пленка; Температурный коэффициент: 25ppm/C; Статус без свинца: без свинца; Статус RoHS: Соответствует RoHS
5% 0402 SMD Характеристики: Сопротивление (Ом): 332 Ом; Мощность (Ватт): 0,063 Вт, 1/16 Вт; Допуск: 0,5%; Упаковка: Digi-Reel; Состав: Тонкая пленка; Температурный коэффициент: 25ppm/C; Статус без свинца: без свинца; Статус RoHS: Соответствует RoHS 