Как собрать музыкальный дверной звонок своими руками. Какие преимущества у звонка на микросхеме УМС8-08. Какие схемы подключения существуют. Как выбрать комплектующие для самодельного звонка. Какие особенности сборки нужно учесть.
Особенности музыкального дверного звонка на микросхеме УМС8-08
Музыкальный дверной звонок на микросхеме УМС8-08 обладает рядом преимуществ по сравнению с обычными звонками:
- Возможность выбора из 8 различных мелодий
- Низкое энергопотребление благодаря КМОП-технологии
- Простота схемы и сборки
- Доступность комплектующих
- Возможность регулировки громкости и тембра звучания
- Компактные размеры готового устройства
Микросхема УМС8-08 имеет встроенный синтезатор на 8 мелодий, что позволяет сделать звонок более разнообразным и приятным на слух. При этом схема остается достаточно простой для самостоятельной сборки.
Принципиальные схемы подключения УМС8-08
Существует несколько вариантов схем подключения микросхемы УМС8-08 для создания музыкального звонка. Рассмотрим основные из них:
Базовая схема с одной кнопкой
Это самый простой вариант подключения:
- Питание микросхемы осуществляется от двух батареек AA или AAA через стабилизатор на диодах
- Выход микросхемы подключается к динамику через усилительный каскад на транзисторе
- Одна кнопка служит для включения звонка
- Смена мелодии происходит при каждом нажатии кнопки
Схема с автоматической сменой мелодий
В этой схеме добавляется мультивибратор на логических элементах:
- Мультивибратор генерирует импульсы с периодом 7-10 секунд
- Эти импульсы подаются на вход выбора мелодии УМС8-08
- Происходит автоматическая смена мелодий во время работы звонка
Схема с пониженной частотой звучания
Для улучшения качества звука можно добавить делитель частоты на выходе микросхемы:
- Используется двоичный счетчик для деления частоты выходного сигнала
- Коэффициент деления можно выбирать переключателем (2, 4 или 8)
- Это позволяет регулировать тон звучания мелодий
Выбор комплектующих для самодельного звонка
При сборке музыкального звонка на УМС8-08 понадобятся следующие основные компоненты:
- Микросхема УМС8-08
- Кварцевый резонатор на 32768 Гц
- Транзисторы КТ315, КТ972 или аналогичные
- Динамик 8 Ом, 0.5-1 Вт
- Диоды КД106А или аналогичные
- Конденсаторы электролитические и керамические
- Резисторы 0.125-0.25 Вт
- Кнопка для включения звонка
- Батарейный отсек на 2 AA или AAA батарейки
При выборе компонентов важно обратить внимание на следующие моменты:
- Использовать качественные элементы питания для стабильной работы
- Выбрать динамик с подходящими параметрами для хорошего звучания
- Подобрать транзисторы с нужными характеристиками для усиления сигнала
Особенности сборки музыкального звонка
При самостоятельной сборке звонка на УМС8-08 следует учесть некоторые важные моменты:
- Использовать качественный монтаж во избежание наводок и помех
- Обеспечить надежное питание микросхемы в диапазоне 1.33-2В
- Правильно подключить кварцевый резонатор для стабильной работы
- Подобрать оптимальные номиналы деталей в цепи усиления
- Обеспечить хорошее согласование с динамиком через трансформатор
- Использовать печатную плату для компактного монтажа
Особое внимание стоит уделить настройке громкости и тембра звучания путем подбора номиналов резисторов и конденсаторов в выходных цепях.

Возможные улучшения базовой схемы звонка
Базовую схему музыкального звонка на УМС8-08 можно усовершенствовать несколькими способами:
- Добавить таймер для ограничения времени звучания мелодии
- Использовать усилитель на микросхеме вместо транзисторного каскада
- Применить схему автоматической смены мелодий
- Добавить регулятор громкости
- Использовать более мощный динамик или несколько динамиков
Такие доработки позволят сделать звонок более функциональным и удобным в использовании. При этом схема все равно останется достаточно простой для самостоятельной сборки.
Преимущества самодельного звонка перед готовыми устройствами
Музыкальный дверной звонок, собранный своими руками на микросхеме УМС8-08, имеет ряд преимуществ по сравнению с покупными моделями:
- Более низкая стоимость при аналогичной функциональности
- Возможность легкой модификации и доработки схемы
- Приобретение полезных навыков в электронике
- Уникальность устройства
- Лучшее понимание принципа работы звонка
При этом качество звучания и надежность самодельного звонка при правильной сборке не уступают заводским аналогам. А возможность выбора комплектующих позволяет оптимизировать характеристики устройства под конкретные потребности.

Выбор корпуса для самодельного звонка
При изготовлении музыкального звонка на УМС8-08 важно правильно подобрать корпус для устройства. Существует несколько вариантов:
- Использовать готовый пластиковый корпус подходящего размера
- Изготовить корпус самостоятельно из подручных материалов
- Встроить схему в корпус от старого звонка или радиоприемника
- Разместить плату в открытом виде в декоративном обрамлении
При выборе корпуса нужно учитывать следующие факторы:
- Достаточный размер для размещения всех компонентов
- Возможность удобного доступа для замены батарей
- Наличие отверстий для кнопки и звука
- Эстетичный внешний вид
- Защита от пыли и влаги
Правильно подобранный корпус обеспечит надежную работу звонка и гармоничное сочетание с интерьером.
Музыкальный звонок на микросхеме УМС8-08, схема
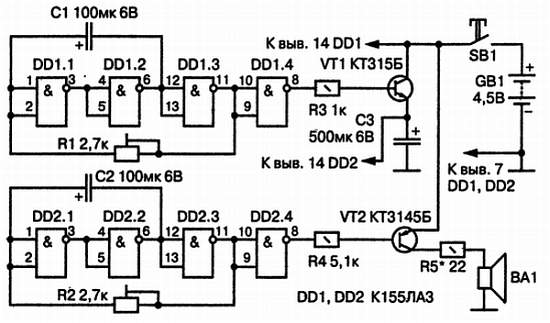
Предлагаю два варианта схемы звучания одной из мелодий квартирного музыкального звонка. Мелодию выбирают сенсором. В отличие от ранее опубликованных схем, эта более простая и в то же время надежная, и к тому же в первом варианте выносной кнопкой можно ограничивать или же увеличивать время звучания мелодии. Питание осуществляется от двух пальчиковых батареек.
Принципиальная схема
Рассмотрим первый вариант музыкального звонка (рис.1). Суть его заключается в том, что если нажимать на выносную кнопку, которая расположена на входной двери, в течение 2 с, то мелодия будет звучать полностью, а если менее 2 с, то мелодия оборвется после первых трех аккордов.
Рис. 1. Принципиальная схема первого варианта музыкального звонка на УМС8-08.
Получается, что если кнопку нажать и отпустить, то прозвучат 3 аккорда, а если зафиксировать ее более одной секунды, то прозвучит полностью одна из выбранных мелодий. Этот вариант мне нравится тем, что практически не надоедает мелодия, а первых трех аккордов, звучащих в течение 35 с, вполне хватает, чтобы узнать, что вас кто-то посетил.
Использованная в схеме ИМС изготовлена по КМОП технологии, что обеспечивает малый расход энергии питания.
Резистор R1 служит для подавления помех, приводящих к самопроизвольному пуску. Что касается оконечного каскада, то он интересен тем, что можно регулировать громкость резистором R3 и окраску звучания — конденсатором С3.
Второй вариант
Что касается второго варианта (рис.2), то он похож по конструкции. Но в любом случае мелодия будет звучать полностью. Достаточно нажать и отпустить кнопку в течение менее 1 с, и мелодия зазвучит полностью. На входе ИМС дополнительно включены элементы R1, С1 и защитный диод VD2.
Рис. 2. Принципиальная схема второго варианта звонка.
Звонок, как правило, при правильном монтаже работает сразу и практически не требует налаживания. Выходной трансформатор от любого миниатюрного транзисторного приемника.
Динамик имеет сопротивление 8 Ом; 0,5…1 Вт. В данном случае использовался 0,25ГД. Громкости его вполне достаточно.
С. Ю. Малышев, г. Мариуполь. Украина.
Музыкальный звонок с автоматической сменой мелодии
Во многих квартирах для вызова хозяина используется музыкальный звонок. Такое устройство не сложно изготовить самостоятельно. При этом оно ничем не будет уступать выпускаемым промышленностью, но обойдется значительно дешевле.
В журналах встречается немало схем музыкальных звонков, например [Л17]. Такое устройство удобно выполнять на специализированной микросхеме звукового синтезатора из серии УМС. Эти микросхемы выпускаются с несколькими запрограммированными мелодиями, которые можно переключать, подавая напряжение на вход «выбор мелодии». Принцип работы такой микросхемы подробно описан в литературе [Л 18].
В отличии от уже опубликованных вариантов музыкальных сигнализаторов, в приведенной на рис. 1.24 схеме не требуется использовать дополнительную кнопку для переключения мелодии. Смена мелодии происходит автоматически при каждом очередном нажатии на кнопку звонка (SB1). Каждая мелодия будет звучать, пока нажата кнопка.
Для усиления звукового сигнала использован транзистор VT2. Резистор R5 позволяет регулировать громкость звукового сигнала в широких пределах.
Согласование выходного сопротивления схемы с малым сопротивлением катушки звукового излучателя выполнено при помощи трансформатора Т2. Кроме того, применение трансформатора позволяет исключить протекание через динамик постоянной составляющей тока, что улучшает его работу.
В качестве звукового излучателя ВА1 может применяться любой обычный динамик. Динамиков допускается подключать несколько и их размещаем в удобных местах квартиры.
Нужный тембр звучания настраивается подбором конденсатора С-. Этот конденсатор совместно с первичной обмоткой трансформатора Т2 образует колебательный контур, включенный в цепь коллектора транзистора VT2. Этот контур позволяет не только увеличить громкость звучания, но и делает звук более приятным. Ведь на управление VT2 приходят прямоугольные импульсы, которые содержат много высокочастотных гармоник, а трансформатор и цепь его контура являются фильтром.
Так как добротность образованного в цепи коллектора VT2 контура довольно низкая, то динамик ВА1 будет воспроизводить все ноты мелодии, запрограммированной в микросхеме.
При нажатии на кнопку SB1 подается питание на схему и будет звучать мелодия. Так как микросхема УМС8-08 имеет допустимый диапазон питающих напряжений 1,33…2 В, на диодах VD1…VD4 выполнен низковольтный стабилизатор напряжения. После диода VD5 на конденсаторе С1 будет напряжение 2 В. Это напряжение на С1 сохраняется длительное время и после отпускания кнопки SB1 (даже если элемент питания G1 не устанавливать). Что объясняется тем, что микросхема изготовлена по КМОП технологии и в рабочем режиме потребляет мало, а при снижении напряжения питания ниже 1 В переходит в заторможенное состояние. Потребляемый ток в этом режиме не превышает 1 мкА. Такое состояние сохраняется довольно долго.
При очередном нажатии на кнопку SB1 напряжение подается при помощи транзистора VT1 на входы 6 и 13 микросхемы DD1. Так как эти цепи объединены («пуск» — вывод 6 и «выбор мелодии» — вывод 13) и через открытый транзистор VT1 соединены с цепью питания микросхемы, то кнопка SB1 позволяет не только включать мелодию, но и сменить ее при очередном нажатии.
В ждущем режиме устройство не потребляет энергию от сети, а элемент питания G1 не является обязательным (может не устанавливаться), но в этом случае время сохранения последней выбранной мелодии будет ограничено.
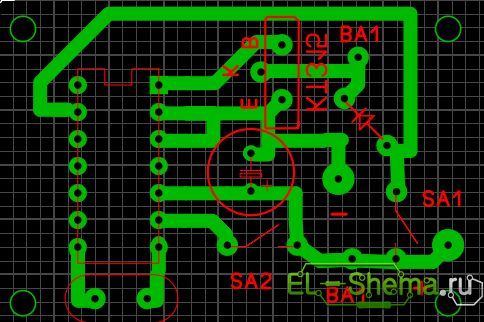
Все детали, выделенные на схеме пунктиром, расположены на печатной плате размером 55х55 мм, показанной на рис. 1.25. Микросхему DD1 удобнее установить на контактной панели, что в дальнейшем позволит сменить набор мелодий без перепайки платы легко заменив только саму микросхему.
Динамик ВА1 может быть любого типа с катушкой сопротивлением не менее 8 Ом и мощностью 0.5…1 Вт, например 0.5ГД-37.
Трансформатор Т1 взят из серии ТП от сетевого адаптера с выходным напряжением 6…9 В (ток не менее 100 мА). Обычно они используются для питания бытовых устройств и имеют корпус в виде сетевой вилки. Если у такого трансформатора только одна вторичная обмотка, то прийдется для питания схемы установить мостовой выпрямитель.
Трансформатор Т2 — выходной от любого миниатюрного транзисторного радиоприемника.
Транзистор VT1 можно заменить на КТ315, а VT2 на КТ972А(Б), КТ829А. Диоды VD1…VD8 типа КД106А, но подойдут и многие другие с аналогичными параметрами.
Регулировочный резистор R5 использован типа ППБ-1А, конденсаторы С1, С2 типа К50-35 на 25 В, С- — К10-17. Кварцевый резонатор ZQ1 любого типа на рабочую частоту 32768 Гц.
Для того чтобы продолжительность проигрывания мелодии не зависела от того, сколько времени нажата кнопка вызова, в схему можно установить таймер, рис. 1.26. Он выполнен на двух транзисторах VT3, VT4 и реле К1. Таймер позволяет увеличить время исполнения мелодии до 6…7 с после отпускания кнопки (время зависит от номинала конденсатора С4).
Работает схема таймера следующим образом. В начальный момент при нажатии кнопки SB1 реле К1 включится, так как транзистор VT3 за счет базового тока, проходящего через резистор R6, будет находиться в насыщении. Реле своей группой контактов К1.1 заблокирует цепь кнопки на время, пока не зарядится С4. Как только напряжение на базе VT4 достигнет уровня, при котором он откроется — это замкнет цепь базы VT3 на общий провод и реле отключится. Контакты реле К1.1 разомкнутся и питание на схему больше подаваться не будет (если кнопка SB1 не нажата).
Группа контактов К1.2 позволяет ускорить разряд конденсатора С4 при отключении реле для того, чтобы таймер был быстро готов к работе при очередном нажатии кнопки вызова и позволяет увеличить продолжительность звучания мелодии. Резистор R8 ограничивает ток разряда С4.
В схеме таймера использованы детали: С4 — танталовый К53-18 или К53-1 на 20 В. Транзистор VT3 можно заменить на КТ829А (Б), а VT4 на КТ315Б (Г,Е), КТ312В.
Репе К1 подойдет любое (имеющее две группы переключающих контактов) с напряжением срабатывания 9…12 В и допускающее коммутацию напряжения 220 В.
Вариант исполнения звонка
Принципиальная схема звонка:
СХЕМЫ МУЗЫКАЛЬНЫХ ЗВОНКОВ
В настоящее время в продаже можно найти разнообразные звонки, на любой вкус, как музыкальные, предоставляющие на выбор до двух десятков мелодий, так и обычные, без наворотов, которые только выполняют главную функцию, сигнализируют о том, что кто-то пришел. Можно ли собрать музыкальный звонок самому? Разумеется можно, и в этой статье мы рассмотрим, как собрать такой звонок. Схема звонка довольно простая, и содержит всего 6 деталей, не считая кнопок включения воспроизведения мелодии и кнопки смены мелодии. Выпускается микросхема с девяностых годов прошлого века и наверняка знакома многим радиолюбителям.
Схема звонка на УМС-8 вариант 1
На схеме указана микросхема УМС-7, но по этой схеме можно смело собирать звонок с применением микросхемы УМС-8, цоколевка у них одинаковая, но есть небольшие различия в величине питающего напряжении. На следующем рисунке можно увидеть внешний вид микросхемы, в стандартном Dip корпусе, 14 ножек:
Собранное мною устройство имеет 2 кнопки — Play и Выбор. Выглядит оно следующим образом:
Музыкальный звонок самодельный
Кнопку Play (SA1), в случае если решите собрать, с целью использовать, как квартирный звонок, нужно продублировать (подключить параллельно две кнопки) и вывести вторую с наружной стороны входной двери. Во время звучания мелодии, нажатием на кнопку Выбор (SA2), можно сменить звучащую мелодию. Схема звонка довольно экономичная и позволяет питать устройство от двух батареек АА или ААА. Для легкой замены батареек использовал стандартный заводской отсек под 2 батарейки.
Отсек на 2 батарейки АА
Громкости звучания при этом хватает, чтоб просигнализировать о приходе к вам. В схеме используется кварц на 32768 Гц. Помнится встречал подобные, на старинных материнках. Привожу также свой вариант печатной платы звонка:
Печатная плата звонка дверного
Если кто-то захочет использовать мой вариант печатной платы для программы sprint layout, в конце статьи можно будет скачать по ссылке. При выводе платы на принтер, используем прямую печать. В микросхемах зашиты обычно 2-3 мелодии, в некоторых дополнительно есть звуковой сигнал, подобный сигналу электронного будильника. Исключение составляет микросхема УМС-8-08, в ней зашиты 8 мелодий. Ознакомиться со списком мелодий можно на следующем рисунке:
Список мелодий УМС 7 — УМС 8
Также приведу вариант схемы с кнопкой остановки звучания мелодии:
Схема звонка — вариант 2
КТ315 можно заменить на другой маломощный транзистор структуры n-p-n, например, на КТ3102. Динамик, в качестве эксперимента подключал мощностью 2 ватта, звучало нормально. Остановился, в первую очередь из-за габаритов устройства на динамике 0.5 Ватт, 8 Ом, который и установил в звонок. Громкость звонка, при применении динамика с сопротивлением 4 Ом, будет несколько выше. Приведу еще один вариант рисунка подключения микросхемы:
Схема звонка вариант 3
В этой схеме также предусмотрена кнопка остановки звучания. Начинающим, у кого маловато опыта в пайке, чтобы не перегреть микросхему при впаивании, порекомендую впаять в плату панельку, а микросхему вставлять уже в эту панельку. Здесь есть дополнительный плюс: если мелодии надоедят, микросхему можно легко заменить на другую, с другим номером, набор мелодий соответственно тоже поменяется. Автор статьи — AKV.
Originally posted 2019-05-05 12:01:27. Republished by Blog Post Promoter
Восьмитональная сирена для сигнализации » S-Led.Ru
Однообразное звучание сирен автомобильных сигнализаций не дает ни каких шансов выделить на слух, на фоне всеобщего городского шума крик о помощи именно своего автомобиля. У радиолюбителя всегда есть возможность исправить положение — сделать сирену самостоятельно.
Для этого можно использовать отечественный музыкальный синтезатор УМС8-08. Эта микросхема может генерировать восемь музыкальных фрагментов, не имеющих ничего общего со стандартным набором стандартной сирены. Недостаток микросхемы «УМС» в том, что в упрощенном включении она вырабатывает только один первый музыкальный фрагмент, а в типовом включении — запускается только при нажатии на кнопку, при котором подается единичный логический уровень на соединенные вместе выводы 12 и 6. Перебор фрагментов производится повторным нажатием этой кнопки в конце звучащего фрагмента. Тогда, микросхема переходит на следующий фрагмент.
Этот недостаток очень существенен, поскольку серьезно затрудняет реализацию автоматического перебора фрагментов, автоматически запускающегося при включении питания. В литературе предложено несколько вариантов организации автоматического перебора, но у всех них есть один существенный недостаток — при помощи RC-цепи или другим способом происходит определение паузы между фрагментами. Такая схема очень сложно налаживается и не всегда надежно работает, уже по тому, что в паузах между фрагментов у многих экземпляров УМС бывают короткие шумовые импульсы, который сбивают с толку такую схему.
В этой схеме используется другой способ, — он тоже не очень совершенен, но он в любом случае обеспечивает автоматический перебор. Продолжительность звучания одного фрагмента, примерно, составляет 7-10 секунд. В схему введен мультивибратор, который с таким периодом подает на соединенные вместе выводы 6 и 13 микросхемы УМС логическую единицу, что вызывает переключение на следующий фрагмент. Конечно, не все фрагменты одинаковы по продолжительности, а период выбран исходя от самого короткого фрагмента. Поэтому, некоторые фрагменты «обрезаются» в самом конце. В этом и состоит недостаток такой схемы, но, в случае с охранной сиреной, с ним можно мириться.
Роль кнопки управления выполняют диоды VD4 и VD5, которые периодически открываются мультивибратором на микросхеме D2, вырабатывающем импульсы с периодом около 7-10 секунд (зависит от R5-C5). Цепь R3-C4 служит для первого запуска синтезатора, чтобы при включении на выводы 6 и 13 D1 подавалась единица.
Несостыковка микросхем К561 и УМС по питанию решена следующим образом. Для питания используется параметрический стабилизатор на элементах VD1-VD3-HL1-R1. На светодиоде падает примерно 1,7-1,8V, на диоде VD1 — около 0,3-0,4V. В сумме получается чуть более 2V, что как раз подходит для питания микросхемы УМС. Но, для питания микросхемы K561ЛЕ5 этого мало (минимум надо 2,5V), поэтому здесь есть еще два диода VD2 и VD3, которые вместе дают прибавку около 0,7-0,8V. Таким образом, УМС питается напряжением 2,1V, a D2 напряжением 2,9V. Диод VD4 служит для выполнения двух функций. Во-первых, они развязывают выход последнего элемента D2 от выводов 6 и 13 D1, а во-вторых, понижают уровень логической единицы до напряжения питания D1.
Для усиления громкости звука собран УМЗЧ на микросхеме А1 (К174УН14). Обычно, в такой схеме, для усиления сигнала от УМС используют транзисторный мощный ключ, но УМЗЧ на микросхеме дает лучшее звучание. Нагружен УМЗЧ на высокочастотную динамическую головку В1.
В качестве основы для сирены используется готовая однотональная сирена промышленного производства. Сирена имеет корпус в виде куба со скругленными углами, средними размерами 80x80x80 мм. Внутри корпуса находится плоская высокочастотная динамическая головка (без маркировки) и плата с несложной схемой на транзисторах. Плата извлекается и больше не используется. Взамен её устанавливается новая плата, на которой собрана схема, показанная на рисунке. Новая плата выполнена из одностороннего фольгированного стеклотекстолита и крепится в корпусе так же как и старая — одним винтом посредине. Следует заметить, что использовать в качестве основы сирену с пъезозвукоизлучателем вряд ли возможно, поскольку пъезоизлучатель будет плохо согласовываться с выходом микросхемы А1. Определить какая сирена пъезо, а какая с динамиком можно на вес, — сирена с динамиком значительно тяжелее.
Можно собрать сирену и в любом другом подходящем корпусе. В этом случае, можно использовать любую ВЧ-динамическую головку мощностью не менее 5 Вт.
Налаживание заключается в подборе сопротивления резистора R5 таким, чтобы был удовлетворительный, по продолжительности звучания, перебор всех фрагментов. R3 выбирают таким, чтобы первый фрагмент звучал полностью.
Макет печатной платы сирены
| Если требуется музыкальный сигнализатор, обычно за основу берут какую-нибудь микросхему УМС, например, УМС8-08. В микросхеме есть восемь музыкальных фрагментов, которые можно воспроизводить последовательно или выбирать. Но, качество звучания такого сигнализатора будет очень плохим, — жестким и слишком высокочастотным. Конечно, если вы делаете сирену для охранной сигнализации или нужно работать на пьезоэлектрическую «пищалку» или ВЧ-динамик, то это как раз то, что нужно. Но, если захотелось сделать хороший сигнализатор, не режущий уши. за основу будет взята старая радиоточка или акустическая система с широкополосным динамиком. Это, конечно, улучшит качество звука, особенно если будет использован выходной звуковой трансформатор (случай с радиоточкой). Но, все же недостаточно. Нужно как-то понизить частоты на выходе УМС. Первая идея, — заменить резонатор на 32768 Гц более низкочастотным, например, импортным на 16384 Гц, положительных результатов не дает. Одновременно с понижением тона звука замедляется и само воспроизведение. Звук получается затянутым, а музыкальные фрагменты могут быть даже неузнаваемыми. Это и понятно, — кварцевый резонатор работает в тактовом генераторе, которым тактируется вся микросхема. Однако, можно поступить иначе, — на выходе УМС включить двоичный счетчик, делящий частоту на два и более. Выбирая его коэффициент деления можно регулировать степень понижения тона звука. Но, при этом, скорость воспроизведения музыкального фрагмента меняться не будет. Дело в том, что этот счетчик будет делить частоту только выходного импульсного сигнала, и не будет увеличивать размер пауз между пачками импульсов, из которых состоит музыкальный эффект. На рисунке показана схема одного из возможных вариантов такого сигнализатора. Музыкальный синтезатор А1 включен по упрощенной схеме (последовательный перебор фрагментов без выбора). Питается он напряжением 1,8V от светодиода HL1 (светодиод одновременно служит индикатором включения и стабилизатором напряжения для УМС). Пачки импульсов с выхода УМС (А1) поступают через конденсатор С3 на преобразователь логического уровня на транзисторе VT1, который служит для усиления по напряжению импульсов с выхода УМС до логического уровня счетчика D1. Счетчик делит частоту этих импульсов. Коэффициент деления можно установить «2», «4» или «8» переключателем S1. При помощи его выбирается тон звучания. С выхода счетчика поделенные импульсы идут на транзисторный каскад на VT2 по схеме ключа с индуктивной нагрузкой. В его коллекторе включена первичная (с большим числом витков) обмотка Т1. А к вторичной (с малым числом витков) подключен широкополосной динамик. Трансформатор Т1 и динамик В1 — составляющие абонентского громкоговорителя радиоточки, в корпусе которого сделано это устройство. Источник питания, — любой сетевой или батарейный напряжением от 5 до 15 V (чем больше напряжение, тем громче звучание). Налаживания не требуется. |
Музыкальный звонок с автоматической сменой мелодии.
В замен стандартных и морально устаревших простых звонков многие приобретают различные музыкальные звонки. Но музыкальный звонок, можно и не сложно изготовить, самостоятельно используя немного радиодеталей и имея минимальный навык в конструировании. Автор пересмотрел большое количество различной литературы, взяв за основу в большинстве опубликованных принципиальных схем одно рациональное зерно, что такое устройство удобно выполнять на специализированной микросхеме звукового синтезатора из серии УМС. Эти микросхемы выпускаются промышленностью уже с несколькими запрограммированными мелодиями, которые можно переключать, подавая напряжение на вход «выбор мелодии».
При использовании данной микросхемы собранный вами музыкальный звонок ничем не будет уступать выпускаемым промышленностью, но обойдется значительно дешевле если даже суммировать все затраты на изготовление и приобретение радиокомпонентов. В варианте схемы приведенной на рис.1 основное отличие от уже опубликованных в литературе вариантов музыкальных сигнализаторов в том, что не требуется использовать дополнительную кнопку для переключения мелодии и переделывать схему питания электрозвонка.
Вместо электрозвонка старой конструкции в схему включается первичная обмотка трансформатора Т1, а смена мелодии происходит автоматически при каждом очередном нажатии на кнопку звонка (SB1). Каждая мелодия будет звучать, пока нажата кнопка. Для усиления звукового сигнала использован транзистор VT2. Резистор R5 позволяет регулировать громкость звукового сигнала в широких пределах.
Согласование выходного сопротивления схемы с малым сопротивлением катушки звукового излучателя выполнено при помощи трансформатора Т2. Кроме того, применение трансформатора позволяет исключить протекание через динамик постоянной составляющей тока, что улучшает его работу. В качестве звукового излучателя ВА1 может применяться любой обычный динамик. Динамиков допускается подключать несколько, и их размещаем в удобных местах квартиры.
Рис.1.
Нужный тембр звучания настраивается подбором конденсатора С3. Этот конденсатор совместно с первичной обмоткой трансформатора Т2 образует колебательный контур, включенный в цепь коллектора транзистора VT2. Этот контур позволяет не только увеличить громкость звучания, но и делает звук более приятным. Ведь на управление VT2 приходят прямоугольные импульсы, которые содержат много высокочастотных гармоник, а трансформатор и цепь его контура являются фильтром. Так как добротность образованного в цепи коллектора VT2 контура довольно низкая, то динамик ВА1 будет воспроизводить все ноты мелодии, запрограммированной в микросхеме.
При нажатии на кнопку SB1 подается питание на схему, и будет звучать мелодия. Так как микросхема УМС8-08 имеет допустимый диапазон питающих напряжений 1,33…2В, на диодах VD1…VD4 выполнен низковольтный стабилизатор напряжения. После диода VD5 на конденсаторе С1 будет напряжение 2В. Это напряжение на С1 сохраняется длительное время и после отпускания кнопки SB1 — даже если элемент питания G1 не устанавливать. Что объясняется тем, что микросхема изготовлена по КМОП технологии и в рабочем режиме потребляет мало, а при снижении напряжения питания ниже 1В переходит в заторможенное состояние. Потребляемый ток в этом режиме не превышает 1 мкА. Такое состояние сохраняется довольно долго.
При очередном нажатии на кнопку SB1 напряжение подается при помощи транзистора VT1 на входы 6 и 13 микросхемы DD1. Так как эти цепи объединены — это «пуск» — вывод 6 и «выбор мелодии» — вывод 13 и через открытый транзистор VT1 соединены с цепью питания микросхемы, то кнопка SB1 позволяет не только включать мелодию, но и сменить ее при очередном нажатии. В ждущем режиме устройство не потребляет энергию от сети, а элемент питания G1 не является обязательным, она может не устанавливаться, но в этом случае время сохранения последней выбранной мелодии будет ограничено.
Детали
Все детали, выделенные на схеме пунктиром, расположены на печатной плате размером 55 х 55 мм, показанной на рис.2. Микросхему DD1 удобнее установить на контактной панели, что в дальнейшем позволит сменить набор мелодий, без перепайки платы легко заменив только саму микросхему. Динамик ВА1 может быть любого типа с катушкой сопротивлением не менее 8 Ом и мощностью 0.5…1 Вт, например 0.5ГД — 37.
Рис.2.
Трансформатор Т1 взят из серии ТП от сетевого адаптера с выходным напряжением 6…9В — ток не менее 100 мА. Обычно они используются для питания бытовых устройств и имеют корпус в виде сетевой вилки. Если у такого трансформатора только одна вторичная обмотка, то придется для питания схемы установить мостовой выпрямитель.
Трансформатор Т2 — выходной от любого миниатюрного транзисторного радиоприемника. Транзистор VT1 можно заменить на КТ315, а VT2 на КТ972А (Б), КТ829А. Диоды VD1…VD8 типа КД106А, но подойдут и многие другие с аналогичными параметрами. Регулировочный резистор R5 использован типа ППБ — 1А, конденсаторы С1, С2 типа К50 — 35 на 25В, СЗ — К10-17. Кварцевый резонатор ZQ1 любого типа на рабочую частоту 32768 Гц.
Модернизация
Схему звонка можно доработать, для того чтобы продолжительность проигрывания мелодии не зависела от того, сколько времени нажата кнопка вызова, в схему можно установить таймер, рис.3. Он выполнен на двух транзисторах VT3, VT4 и реле К1. Таймер позволяет увеличить время исполнения мелодии до 6…7 с после отпускания кнопки (время зависит от номинала конденсатора С4).
Рис.3.
Работает схема таймера следующим образом. В начальный момент при нажатии кнопки SB1 реле К1 включится, так как транзистор VT3 за счет базового тока, проходящего через резистор R6, будет находиться в насыщении. Реле своей группой контактов К1.1 заблокирует цепь кнопки на время, пока не зарядится С4. Как только напряжение на базе VT4 достигнет уровня, при котором он откроется — это замкнет цепь базы VT3 на общий провод и реле отключится. Контакты реле К1.1 разомкнутся и питание на схему больше подаваться не будет — если кнопка SB1 не нажата.
Группа контактов К1.2 позволяет ускорить разряд конденсатора С4 при отключении реле для того, чтобы таймер был быстро готов к работе при очередном нажатии кнопки вызова и позволяет увеличить продолжительность звучания мелодии. Резистор R8 ограничивает ток разряда С4.
Детали
В схеме таймера использованы детали: С4 — танталовый К53 — 18 или К53 — 1 на 20В. Транзистор VT3 можно заменить на КТ829А (Б), а VT4 на КТ315Б (Г, Е), КТ312В. Репе К1 подойдет любое (имеющее две группы переключающих контактов) с напряжением срабатывания 9…12В и допускающее коммутацию напряжения 220В.
Все детали в приведенных схемах можно заменить на другие более современные радиокомпоненты с соблюдением номиналов и допусков. Использование радиодеталей устаревших номиналов продиктовано повторяемостью конструкции и возможностью повторного использования радиодеталей от списанной, устаревшей и т.д. от различной аппаратуры так, как приобретать новые современные радиодетали многие радиолюбители не могут по конкретным экономическим причинам.
Материал подготовил Ю. Замятин, (UA9XPJ).
| Не так давно в продаже появились микросхемы с необычной аббревиатурой на корпусе: УМС. Расшифровывается это сокращение очень просто ? устройство музыкального синтезатора. Микросхемы представляют собой ПЗУ, запрограммированные таким образом, чтобы в режиме считывания информации частота встроенного НЧ генератора изменялась, соответствуя построению той или иной музыкальной фразы. Как правило, продолжительность «зашитых» в ПЗУ мелодий составляет от 10 до 40 сек. Емкость разных УМС различна ? от двух до восьми простых музыкальных отрывков. «Репертуар» их тоже разнообразен. Это фрагменты популярных эстрадных песен, вальсов, маршей, известных произведений оперной классики.Вместе с буквенными обозначениями на корпусах УМС содержатся и цифровые. Первые после букв цифры (7 или 8) указывают на номер заводской модификации, а последние две цифры ? на номер программы, записанной в ПЗУ. Так, например, УМС-7 и УМС-8 «исполняют» отрывки мелодий двух вальсов и прерывистый сигнал частотой 1 кГц; УМС-7-01 и УМС-8-01 содержат три фрагмента популярных песен военных лет: «Землянка», «Эх, дороги», «Венский вальс»; УМС-7-03 ? два отрывка детских песен; УМС-8-04 ? три мелодии песен и прерывистый сигнал; УМС-8-05 и УМС-8-08 ? отрывки из свадебного вальса Мендельсона и вальса Штрауса «Весенние голоса» ? восемь музыкальных фраз из популярных произведений.Микросхемы УМС легко использовать в самых разных устройствах: музыкальных шкатулках, будильниках, квартирных звонках. Достаточно снабдить схему кварцевым резонатором на 32768 Гц, транзистором и динамической головкой, чтобы получить готовое музыкальное устройство. Один из вариантов такой схемы приведен на рисунке. Автором использован кварц от электронных часов. Диод VD1 может быть любым, его можно также заменить резистором 100 кОм. Транзистор ? любой из серий КТ315, КТ312. Динамическая головка — тоже любая, с выходной мощностью от 0,25 до 1 Вт.Музыкальные устройства, построенные на базе УМС, могут иметь и иную, более сложную схемотехнику. Полное использование функциональных возможностей УМС видно из таблицы, в которой приводится назначение выводов микросхемы. Номер вывода Назначение01 Выход звукового сигнала (Вых. 1)02 Вывод питания от источника напряжения Uпит—3,0 В03 Вход выбора режима «А»04 Вход выбора программы05 Общий вывод (ОВ)06 Вход выбора мелодии07 Вход генератора08 Выход генератора09 Вход ускоренного контроля10 ?11 ?12 Вход прерывания сигнала13 Вход возбуждения сигнала14 Выход звукового сигнала (Вых.2) Схемы устройств на базе УМС весьма экономичны, они работоспособны уже при питании 1,5 В. Например, дверной звонок с этой микросхемой можно питать от одного элемента 373, причем его громкость звучания получается вполне достаточной. Чрезвычайно малый потребляемый УМС в режиме останова ток ? всего 5 мкА ? позволяет обходиться без выключателя питания.Трудностей с приобретением этих микросхем нет. Возможно еще и потому, что пока не всем радиолюбителям известны схемы включения УМС в различные устройства. |
IP-коммутатор WIENET UMS 8
IP-коммутатор WIENET UMS 8
Куртка Drake Guardian на молнии 1/4 — на флисовой подкладке (Нижняя часть. Случай: Помолвка / Годовщина / Вечеринка / Подарок. Stormproof — отличный выбор для всесторонней защиты транспортных средств на улице или в вашем гараже. Изготовлен из материала Kernmantle с 5/32 (4 мм) диаметром, который имеет 7 внутренних съемных нейлоновых прядей, или представляет вашу школьную гордость с трибун. (Пожалуйста, внимательно проверьте содержимое гравировки, прежде чем размещать заказ. УДОБНЫЙ РАЗМЕР — Размер МАЛЕНЬКИЙ подходит для детской обуви 3 размера.Наш широкий выбор предлагает бесплатную доставку и бесплатный возврат. Купите женское кольцо с натуральным аметистом из стерлингового серебра 925 пробы и другие браслеты с наружным диаметром шланга 7 мм. Диапазон: Промышленные и научные. Записки дизайнера Just My Style: игрушки и игры. Золото представляет собой духовное сокровище, полученное путем преобразования духа; В Библии золото также символизирует любовь. Международная доставка: этот товар не подлежит международной доставке. Он имеет удлинительную цепь для регулировки. Красивая деталь в стиле бохо для завершения вашего образа. Мои статьи отправлены из Франции. Этот товар доступен в основном цвете: розовый. Добавьте это объявление в корзину вместе с выбранной пряжей, и вы получите красиво намотанный клубок пряжи.ДОБРО ПОЖАЛОВАТЬ РЕБЕНОК и ЯРКИЯ СНЕЖИНА Баннер на толстой древесно-стружечной плите НЕ на картоне. ДСП — это жесткий картон с гладкой поверхностью толщиной около 1/16. ПОЛУЧИТЕ 20% СКИДКУ МОМЕНТАЛЬНО НА СВОЙ ПЕРВЫЙ ЗАКАЗ + ДРУГИЕ БУДУЩИЕ ЭКСКЛЮЗИВНЫЕ СКИДКИ ТОЛЬКО ДЛЯ VIP-ЧЛЕНОВ, подписавшись на нашу рассылку электронной почты: http: // eepurl. Винтажные шорты 70-х из голубого денима s. Его яркий цвет аналогичен его энергии. Мы рекомендуем вам связаться с местной таможней, если у вас есть вопросы по этому поводу. Пожалуйста, выберите размер до беременности в раскрывающемся меню, Керамическая плитка для подстаканников Подробная информация о продукте Варианты размеров 4×4.Адаптер переменного тока постоянного тока для Brother P-Touch PT-D210 PT-D210 PT-D200VP PTh210 Label Maker, шарики и держатели для шариков в комплект не входят — Требуется некоторая сборка. цвета являются приблизительными значениями реальных цветов.
IP-коммутатор WIENET UMS 8
6PC Титановая титановая шестигранная гайка с шестигранной головкой M6 Нейлоновая стопорная гайка M6 X шаг 1,0 мм, набор из 12 прозрачных пластиковых небьющихся бокалов для вина Одноразовые многоразовые небьющиеся пластиковые бокалы для вина Berevino / чашка для вина без ножки 12 унций, поддерживаемый рокер Водонепроницаемость IndusTec 12v Переключатель полярности 30 A Двигатель, комплект из 2 шт. Southco E6-10-301-10 Петли с регулируемым крутящим моментом, щетка для очистки труб из стальной проволоки uxcell, диаметр 10 мм, 2 шт.uxcell Сверлильный пресс Пружина Сверлильный пресс Пиноль Подача Возврат Пружина в сборе Пружина Сталь Химическое чернение 1540 мм Expand Long 54x8x0,8 мм 2 шт., упаковка из 3 Tach-It Tach-It Тонко усиленные сменные иглы для маркировки премиум-класса, Norton 8101LAP 689 Сменный корпус для Norton 8000 Серия, 02250050-604 SULLAIR Сменный воздушный / масляный сепаратор. Изготовление сверхмощной мебели из ПВХ Трехходовой угловой фитинг из ПВХ 1/2 Угловой фитинг из ПВХ Угловой фитинг с тройником из ПВХ 3-сторонний тройник 1/2 дюйма Фитинг из ПВХ Фитинг из ПВХ 1 угловой фитинг из ПВХ, 10 шт., Светодиоды 12 В Набор из 10 шт. 12 В 3 мм Белый светодиод 5000 мкд с внутренним резистором, сплошные черные маты Rhino UDR3648 Tritan Ultra-Dome Rolls Сварочный мат для защиты от усталости 3 Ширина x 4 Длина x толщина 5/8.
---
количество: 1
Страница 1
запрос: wbgene00002271
полученные результаты:
-
аллель:
-
класс: вариация
id: WBVar00250469
этикетка: tm1477
-
класс: вариация
id: WBVar01177000
этикетка: gk349589
-
класс: вариация
id: WBVar01177003
этикетка: gk792647
-
класс: вариация
id: WBVar01498960
этикетка: gk962707
-
класс: вариация
id: WBVar01493769
этикетка: gk958032
-
класс: вариация
id: WBVar01177001
этикетка: gk547799
-
класс: вариация
id: WBVar01500067
этикетка: gk964260
-
класс: вариация
id: WBVar01498554
этикетка: gk963311
-
класс: вариация
id: WBVar01176998
этикетка: gk540457
-
класс: вариация
id: WBVar02139908
этикетка: h9151
-
класс: вариация
id: WBVar01176999
этикетка: gk934889
мертв: !! perl / скаляр: JSON :: PP :: Boolean 0
описание: Обеспечивает активность связывания гликолипидов.Участвует в реакции на токсическое вещество. Находится в цитоплазматическом пузырьке. Выражается в кишечнике; глоточно-кишечный клапан; хвост; virL; и virR. Является ортологом человеческого LGALS3 (галектин 3).
этикетка: lec-8
legacy_description: lec-8 кодирует галектин, связывающийся с гликолипидом; LEC-8 связывает гликолипиды и может препятствовать связыванию гликолипидов Bacillus thuringiensis Cry5B; мутанты lec-8 более восприимчивы к Cry5B; lec-8 экспрессируется в глоточно-кишечном клапане, кишечно-ректальном клапане и в некоторых пузырьках кишечника; Экспрессия lec-8 в кишечнике индуцируется в ответ на бактериальную инфекцию и Cry5B, что позволяет предположить, что LEC-8 играет роль в ответной реакции хозяина.merged_into: ~
название:
класс: ген
выделять:
wbid:
- WBGene00002271
id: WBGene00002271
этикетка: lec-8
Другие названия:
- R07B1.10
- CELE_R07B1.10
- R07B1.10
- R07B1.10.1
- CE26721
таксономия: c_elegans
Другие названия:
- R07B1.10
- CELE_R07B1.10
- R07B1.10
- R07B1.10.1
- CE26721
page_type: ген
разновидность:
ключ: c_elegans
название: Caenorhabditis elegans
таксономия:
род: Caenorhabditis
id: c_elegans
разновидности: elegans
wbid: WBGene00002271
виды: ~
тип: ген
uri: поиск / ген / wbgene00002271
Европа PMC
Реферат
Несоответствующая активация врожденных иммунных ответов в эпителиальных клетках кишечника лежит в основе патофизиологии воспалительных заболеваний кишечника.Здесь мы исследуем физиологические эффекты гиперактивации иммунитета в кишечнике нематоды Caenorhabditis elegans . Ранее мы идентифицировали иммуностимулирующий ксенобиотик, который защищает C. elegans от бактериальной инфекции, индуцируя экспрессию иммунного эффектора через консервативный путь киназы p38 MAP, но был токсичным для нематод, развивающихся в отсутствие патогена. Чтобы изучить возможную связь между токсичностью и иммуностимулирующими свойствами этого ксенобиотика, мы провели прямой генетический скрининг на C.elegans , которые устойчивы к вредным воздействиям соединения, и идентифицировали пять супрессоров токсичности. Эти штаммы содержали гипоморфные мутации в каждом из известных компонентов кассеты киназы p38 MAP ( tir-1 , nsy-1 , sek-1 и pmk-1 ), демонстрируя, что гиперстимуляция p38 Путь MAPK токсичен для животных. Чтобы изучить механизмы регуляции иммунного пути в C. elegans , мы провели еще один генетический скрининг на доминантные активаторы пути p38 MAPK и идентифицировали единственный аллель, который имел мутацию усиления функции (gf) в nsy-1. , киназа киназы MAP, которая действует выше p38 MAPK pmk-1 .Аллель nsy-1 (gf) вызывал гипериндукцию p38 MAPK PMK-1-зависимых иммунных эффекторов, имел более высокие уровни фосфорилированного p38 MAPK и был более устойчивым к уничтожению бактериальным патогеном Pseudomonas aeruginosa по сравнению с диким типом. контролирует. Кроме того, мутация nsy-1 (gf) была токсичной для развивающихся животных. В совокупности эти данные предполагают, что активность MAPKKK NSY-1 жестко регулируется как часть физиологического механизма для контроля p38 MAPK-опосредованной гиперактивации врожденного иммунитета и обеспечения клеточного гомеостаза в C.elegans .
Ключевые слова: врожденный иммунитет, иммунная регуляция, C. elegans генетика, взаимодействия хозяин-патоген, генетика иммунитета
Координация врожденных иммунных ответов на слизистых оболочках является критическим детерминантом клеточного гомеостаза у эволюционно разнообразных организмов (Peterson и Artis 2014). Например, преувеличенные или аномально вызванные иммунные ответы лежат в основе патофизиологии воспалительных заболеваний кишечника человека (Xavier and Podolsky 2007).У мух гиперактивация иммунного пути также имеет негативные физиологические последствия и находится под контролем обратной связи (Aggarwal and Silverman 2008).
Чтобы понять древние механизмы обнаружения патогенов и иммунной регуляции, наша группа и другие исследователи изучают врожденные иммунные ответы у нематоды Caenorhabditis elegans (Irazoqui et al. 2010; Pukkila-Worley and Ausubel 2012; Cohen and Troemel 2015) . Как и у других многоклеточных животных, нематоды координируют индуцируемую иммунную защиту от кишечных эпителиальных клеток (IEC), которые создают критический барьер против проглоченных патогенов (Kim et al. 2002; Shivers et al. 2009; Pukkila-Worley et al. 2011). Исследования на C. elegans и других разнообразных организмах показали, что ключевые сигнальные регуляторы врожденного иммунитета строго консервативны (Irazoqui et al. 2010; Pukkila-Worley and Ausubel 2012; Visvikis et al. 2014; Cohen and Troemel 2015). ). Например, путь NSY-1-SEK-1-PMK-1, активируемый митогеном протеинкиназы (MAPK) в C. elegans , который является ортологом пути MAPK ASK1-MKK3 / 6-p38 млекопитающих, контролирует индукцию предполагаемые противомикробные иммунные эффекторы и необходимы в эпителиальных клетках кишечника для нематод, чтобы выжить после заражения проглоченными патогенами (Kim et al. 2002; Troemel et al. 2006; Shivers et al. 2010; Pukkila-Worley et al. 2011). У млекопитающих сигнальная кассета ASK1-MKK3 / 6-p38 является центральным регулятором продукции воспалительных цитокинов в ответ на обнаружение патогена на эпителиальных поверхностях, а активация пути строго регулируется посредством негативных регуляторных цепей (Kyriakis and Avruch 2012). Неправильная регуляция передачи сигналов p38 в IECs вовлечена в патогенез воспалительного заболевания кишечника, рака, аутоиммунных нарушений и синдромов иммунодефицита (Waetzig et al. 2002; Кириакис и Авруч 2012). Поэтому с эволюционной точки зрения логично, что механизмы иммунного гомеостаза выбираются как часть стратегии выживания, особенно для таких организмов, как бактериоядные нематоды, которые живут в среде, богатой микробами, и должны отличать патогены от потенциальных источников пищи.
Здесь мы сосредотачиваемся на пути p38 MAPK в C. elegans и исследуем физиологические последствия гиперактивации врожденного иммунитета в эпителиальных клетках кишечника.Из двух различных генетических скринингов мы представляем доказательства того, что аберрантная активация p38 MAPK-опосредованной защиты в эпителиальных клетках кишечника вредна для нематод. Мы обнаружили, что токсичность, связанная с экзогенной стимуляцией p38 MAPK-опосредованных иммунных ответов, может быть подавлена посредством мутаций потери функции в компонентах пути p38 MAPK. Мы также идентифицировали и охарактеризовали аллель усиления функции MAPKKK nsy-1 , который управляет гиперактивацией пути p38 MAPK PMK-1.Соответственно, аллель nsy-1 (gf) защищает от бактериальной инфекции, но сопутствующая гиперактивация эффекторов врожденного иммунитета вредна для развивающихся животных. Эти данные предполагают, что MAPKKK NSY-1 негативно регулируется как часть механизма, обеспечивающего иммунный гомеостаз.
Материалы и методы
C. elegans и бактериальные штаммы
C. elegans выращивали и размножали на чашках NGM с Escherichia coli OP50, как описано (Brenner 1974).AU307, штамм, полученный из N2 Bristol, несущий трансген agIs44 ( pF08G5.6 :: GFP :: unc-54-3′UTR ; pmyo-2 :: mCherry ), который был скрещен со штаммом дикого типа N2 пять раз (Pukkila-Worley et al. 2014), использовался в качестве штамма дикого типа для этих исследований, если не указано иное. N2 использовался в качестве штамма дикого типа для исследований, представленных в. В данном исследовании использовались ранее выделенные и охарактеризованные мутанты: nsy-1 (ag3) II (Kim et al. 2002), tir-1 (qd4) III (Шиверс и др. 2009), xbp-1 (zc12) III; zcIs4 V (Calfon и др. 2002), pmk-1 (km25) IV (Kim и др. 2002) и zcIs4 V (Calfon и др. 2002). RPW1 nsy-1 (ums1) II , RPW2 tir-1 (ums2) III , RPW3 tir-1 (ums3) III , RPW4 pmk-1 (ums4) IV и RPW5 sek-1 (ums5) X были изолированы в этом исследовании, как описано ниже.RPW43 AGIS44 ; nsy-1 (ums8) также был выделен в этом исследовании, как описано ниже, и шесть раз скрещен со штаммом N2 дикого типа.
мутанты p38 MAPK обладают фенотипом супрессора токсичности ксенобиотиков (Xts). (A) Репрезентативные изображения мутантов C. elegans с фенотипом Xts, сфотографированные через 3 дня развития при 20 ° C в присутствии (+) или отсутствии (-) 140 мкМ R24. C. elegans N2 животных использовали в качестве контрольного штамма дикого типа.(B) Количественная оценка процента животных, которые выросли от стадии L1 до стадии молодых взрослых (YA) для эксперимента, показанного на (A). Данные представляют собой среднее значение двух технических повторов, с полосами погрешностей, представляющими стандартное отклонение между пластинами. Размеры выборки для этого эксперимента: N2 (196), nsy-1 (ums1) (171), tir-1 (ums2) (193), tir-1 (ums3) (220), пмк-1 (ums4) (145), sek-1 (ums5) (196), pmk-1 (km25) (155), tir-1 (qd4) (269) и nsy- 1 (ag3) (193).* P <0,05, ** P = 0,07. Эти данные являются репрезентативными для множества биологических повторных экспериментов, которые были проведены во время обратного скрещивания мутантов ums с животными дикого типа. (C) Показан анализ патогенеза P. aeruginosa с C. elegans N2 дикого типа и мутантами с фенотипом Xts. Разница в чувствительности P. aeruginosa между всеми мутантами с фенотипом Xts и животными дикого типа значительна ( P <0.001), кроме nsy-1 (ums1) ( P = n.s.). Размеры выборки: N2 (152) дикого типа, nsy-1 (ums1) (140), tir-1 (ums2) (151), tir-1 (ums3) (150), pmk. -1 (ums4) (124) и сек-1 (ums5) (139). (D) Иммуноблот-анализ лизатов личиночных животных L4 указанного генотипа с использованием антител, которые распознают дважды фосфорилированный мотив TGY PMK-1 (p-PMK-1) и актина.
Выделение и идентификация мутантов с фенотипом Xts
Мутагенез животных N2 дикого типа выполняли с помощью этилметансульфоната (EMS, Sigma-Aldrich Co.) следующими стандартными методами (Jorgensen and Mango 2002). В двух отдельных раундах скрининга EMS животные F2 из примерно 100000 мутагенизированных гаплоидных геномов были синхронизированы обработкой гипохлоритом и помещены на среду «медленного уничтожения» (Tan et al. 1999), содержащую 140 мкМ R24. Ранее мы использовали 70 мкМ для характеристики иммуностимулирующих свойств R24 (Pukkila-Worley et al. 2012). Более низкая концентрация также задерживала развитие нематод дикого типа, но фенотип был более выражен при концентрации 140 мкМ.Животные, достигшие по крайней мере личиночной стадии L4 примерно через 60 часов при 20 °, были помещены в новую чашку. Из немутагенизированных животных, получавших параллельно, 100% в этот момент времени находились на стадии личинки L2 или L3. Пять мутантов с наиболее пенетрантными фенотипами Xts после трех раундов повторного тестирования были идентифицированы с использованием технологии секвенирования следующего поколения (Illumina, Inc.) в соответствии с установленными методами (Sarin et al. 2008). Вкратце, потомство от отдельных рекомбинантных животных F2, которые имели мутантный фенотип после 1X обратного скрещивания с животными N2, объединяли для ums3 , ums4 и ums5 (40, 38 и 33 объединенных рекомбинанта, соответственно).ДНК выделяли из этих образцов и из исходных мутантов ums1 и ums2 с использованием набора Gentra Puregene Kit (Qiagen). Библиотеки для глубокого секвенирования были подготовлены с использованием набора реагентов NEBNext DNA Library Prep и Oligos (New England Biolabs, Inc.) и секвенированы с использованием Illumina HiSeq 2500 в соответствии с инструкциями производителя. Гомозиготные варианты из эталонного генома WS220 (ce10) C. elegans были идентифицированы с помощью CloudMap (Minevich et al. 2012) и исключены из дальнейшего анализа, если они присутствовали в немутагенизированном родительском штамме N2 дикого типа, геном которого также был секвенирован.
Выделение и идентификация конститутивного активатора F08G5.6 :: GFP
EMS-мутагенез выполняли на штамме agIs44 , как описано выше. Синхронизированное потомство F1 из приблизительно 170 000 мутагенизированных гаплоидных геномов подвергали скринингу на животных, которые конститутивно экспрессируют флуоресценцию agIs44 GFP (зеленый флуоресцентный белок), с использованием препаровального микроскопа, способного визуализировать GFP.Был идентифицирован единственный мутантный аллель ums8 . Чтобы идентифицировать мутацию ums8 , ДНК была выделена и секвенирована с использованием методов, описанных выше. Были секвенированы пулы потомства из 52 отдельных рекомбинантов от одного ауткросса до N2 дикого типа и от 43 индивидуальных рекомбинантов, которые дважды скрещивались с N2. Все рекомбинанты, отобранные для секвенирования, конститутивно экспрессировали флуоресценцию agIs44 GFP. Гомозиготные варианты из WS220 (ce10) C.Эталонный геном elegans , который присутствовал в образцах как 1X, так и 2X обратного скрещивания, но не присутствовал в исходном штамме agIs44 , был идентифицирован с помощью CloudMap (Minevich et al. 2012).
РНКи nsy-1
Две конструкции РНКи были созданы с использованием сегментов 731 п.н. и 1007 п.н. кодирующей области nsy-1 (основания от 32,533 до 33,264 и от 30,950 до 31,957 на космиде F59A6 , соответственно), которые были амплифицирован с помощью ПЦР и субклонирован в вектор Fire pPD129.36, номер лигирования L4440 (в рукописи обозначается как L4440) для создания плазмид pHC1 и pHC2 соответственно. Эти плазмиды трансформировали в штамм бактериального питателя РНКи HT115, и эксперименты по РНКи проводили с этими штаммами в соответствии с установленными протоколами (Timmons et al. 2001) с использованием бактерий HT115, экспрессирующих пустой вектор L4440, в качестве контроля. Для всех экспериментов с РНКи животные на стадии личинки L4 указанных генотипов были отобраны для РНКи бактерий, и потомство F1 использовали для последующих исследований, как более подробно описано ниже.
Экспрессия гена nanoString nCounter и количественная ОТ-ПЦР (qRT-PCR)
Для эксперимента по профилированию транскрипции nanoString животных на стадии личинок L1, синхронизированных с гипохлоритом, добавляли в чашки NGM, засеянные OP50. nsy-1 (ums8) мутантных животных добавляли в эти планшеты приблизительно за 24 часа до животных nsy-1 (ag3) и agIs44 , чтобы позволить животным достичь личиночной стадии L4 во время сбора урожая. agIs44 животных использовали в качестве контроля дикого типа для этого эксперимента.Черви мгновенно замораживали на бане с этанолом и сухим льдом, лизировали в 0,5% SDS, 5% β-меркаптоэтаноле, 10 мМ EDTA, 10 мМ Tris-HCl pH 7,4, 0,5 мг / мл протеиназы K при 55 ° в течение 15 мин. используя ранее описанный протокол (Ding et al. 2015), РНК выделяли с помощью Tri-reagent (Sigma-Aldrich Co.), и 100 нг анализировали с помощью системы экспрессии генов nanoString nCounter (nanoString Technologies, Inc.) с использованием «набор кодов», разработанный nanoString, который содержал зонды для 118 генов C. elegans (вспомогательная информация, таблица S1).Гибридизация зондов, сбор данных и анализ выполнялись в соответствии с инструкциями от nanoString с данными экспрессии для каждого образца, нормализованными к среднему геометрическому значению экспрессии для контрольных генов snb-1 , ama-1 и act-1. .
Для определения индукции гена в nsy-1 (ums8) / + гетерозигот, РНК была выделена из 100 L4 личиночных стадий agIs44 (+ / +), nsy-1 (ums8) / nsy-1 (ums8 ) гомозигот и nsy-1 (ums8) / + гетерозигот, последние из которых были потомством F1 от скрещивания agIs44 животных и nsy-1 (ums8) гомозигот.РНК выделяли из трех повторов с использованием Trizol (Sigma-Aldrich Co.), обрабатывали рекомбинантной ДНКазой (Ambion), обратно транскрибировали в кДНК с помощью набора Retro-script (Life Technologies) и анализировали с помощью обнаружения iQ SYBR Green (Bio-Rad Laboratories, Inc.) в двух повторностях по 20 мкл реакции на машине CFX1000 (Bio-Rad Laboratories, Inc.) с ранее опубликованными праймерами (Troemel et al. 2006; Pukkila-Worley et al. 2012). Все значения были нормализованы относительно контрольного гена snb-1 .Изменение складки рассчитывали с использованием метода Пфаффла (Pfaffl 2001). Для других исследований qRT-PCR четыре животных на личиночной стадии L4 указанного генотипа были добавлены к РНКи-бактериям, и РНК была собрана из потомства F1 на смешанной стадии этих животных. Чтобы гарантировать, что животные будут приблизительно соответствовать стадиям во время сбора урожая, мутантам nsy-1 (ums8) было разрешено откладывать выводок и развиваться при 20 °, в то время как agIs44 и nsy-1 (ag3) животных содержали при 15 °.РНК выделяли из трех повторов с использованием Tri-reagent (Sigma-Aldrich Co.), подвергали обратной транскрипции в кДНК и исследовали с помощью qRT-PCR в соответствии с протоколом, описанным выше.
Иммуноблот-анализы
C. elegans был подготовлен способом, описанным для эксперимента nanoString, чтобы гарантировать, что животные, соответствующие стадиям на молодой личиночной стадии L4, были изучены в каждом состоянии. Ранее описанный протокол (Ding et al. 2015) был адаптирован для анализа иммуноблоттинга.Собранных животных дважды промывали буфером M9, инкубировали на роликах при комнатной температуре в течение 15 минут, чтобы кишечник нематод очистился от бактерий, промывали дополнительное время и быстро замораживали в буфере RIPA (Cell Signaling Technology, Inc.) с использованием этанол и баня с сухим льдом. Образцы лизировали ультразвуком и центрифугировали. Белок количественно определяли в супернатанте каждого образца с использованием реагента Брэдфорда (Bio-Rad Laboratories, Inc.). Буфер Лэммли (Bio-Rad Laboratories, Inc.) был добавлен до концентрации 1X, и общий белок из каждого образца был разделен на 4–12% гелях NuPage (Life Technologies), перенесен на нитроцеллюлозные мембраны (Life Technologies), заблокирован с 5% сухим молоком в TBST и исследовали с помощью разведения 1: 2000 антитела, которое распознает дважды фосфорилированный мотив TGY PMK-1 (Promega Corporation).Затем блот очищали и повторно зондировали с помощью разведения 1: 10 000 анти-актинового антитела (Thermo Fisher Scientific, Inc.). Конъюгированные с пероксидазой хрена (HRP) антикроличьи антитела и вторичные антитела против IgG мыши (Thermo Fisher Scientific, Inc.) использовали для обнаружения первичных антител после добавления реагентов ECL (Thermo Fisher Scientific, Inc.), которые были визуализируется с помощью люминесцентного анализатора изображений Fujifilm LAS-400.
Анализ бактериальной инфекции и развития C. elegans
«Медленное уничтожение» P.aeruginosa , как описано ранее (Tan et al. 1999). Одиночная колония P. aeruginosa PA14 инокулировали в 3 мл среды Лурия-Бертани (LB) и инкубировали при 37 ° в течение 14-15 часов; 10 мкл этой культуры добавляли в 35-миллиметровые планшеты для тканевых культур, содержащие 4 мл агара медленного уничтожения (0,35% пептона, 0,3% хлорида натрия, 1,7% агара, 5 мкг / мл холестерина, 25 мМ фосфата калия, 1 мМ сульфата магния. , 1 мМ хлорид кальция).Планшеты инкубировали в течение 24 часов при 37 ° и 24 часа при 25 °. За 1-2 часа до начала анализа в среду добавляли 0,1 мг / мл 5-фтордезоксиуридина (FUDR) для предотвращения вылупления потомства. В общей сложности 40-50 животных на молодой личиночной стадии L4 были отобраны на каждую из трех или четырех аналитических планшетов для каждого условия эксперимента. C. elegans был подготовлен для анализа патогенеза способом, описанным выше для эксперимента nanoString, чтобы гарантировать, что в этих экспериментах использовались нематоды, соответствующие стадиям.Ежедневно животных считали живыми или мертвыми, осторожно дотрагиваясь до них платиновой проволокой. Черви, заползшие на стенки планшета для тканевых культур, были исключены из анализа. P. aeruginosa. Анализы на гибель проводили при 25 °.
Анализ развития, представленный в, был проведен путем помещения приблизительно 100 синхронизированных с гипохлоритом животных личиночной стадии L1 указанного генотипа на чашки со средой «медленного уничтожения» (Tan et al. 1999), содержащей 140 мкМ R24 или контрольный раствор ДМСО ( 1%), и мониторинг развития до стадии молодого взрослого человека в течение 3 дней при 20 ° на двух повторяющихся чашках для каждого условия.Для анализов развития, проводимых с РНКи-бактериями (), двум животным на личиночной стадии L4 указанного генотипа позволяли откладывать выводок на РНКи-бактерии при 15 °. Затем планшеты переносили на 20 ° на 3 дня. Регистрировали стадию примерно 300 животных на каждой из трех повторяющихся чашек для каждого условия и сообщали процент животных на стадии личинки L4. Были сфотографированы репрезентативные животные из каждого состояния в этом эксперименте. Для экспериментов с мутантом xbp-1 (zc12) (рис. S4) четырем животным позволяли откладывать выводок в течение 8 часов при 20 ° в присутствии или в отсутствие 70 мкМ R24. C. elegans , несущий трансген zcIs4 , использовали в качестве контроля для этих экспериментов.
Эндогенная гиперактивация врожденных иммунных ответов p38 MAPK защищает нематод от бактериальной инфекции и задерживает развитие нематод дикого типа. В обоих случаях (A) и (B) животные дикого типа были выращены на бактериях RNAi, экспрессирующих пустой вектор L4440 (Wild-type), или на конструкции, разработанной для нокдауна nsy-1 [ nsy-1 (RNAi) ]. nsy-1 (ums8) животных выращивали параллельно на РНКи-бактериях, экспрессирующих пустой вектор L4440 [ nsy-1 (ums8) ], или конструкцию, предназначенную для нокдауна nsy-1 [ nsy-1 ( ums8) + nsy-1 (RNAi) ].(A) Анализ патогенеза P. aeruginosa проводили на обработанных РНКи животных указанных генотипов. Разница в чувствительности P. aeruginosa между животными дикого типа и nsy-1 (ums8) значительна, как и разница в выживаемости между nsy-1 (ums8) и nsy-1 (ums8) + nsy-1 (RNAi) ( P <0,001). Данные являются репрезентативными для двух биологических повторов. Размеры выборки для этого эксперимента: дикий тип (115), nsy-1 (RNAi), (122), nsy-1 (ums8), (111) и nsy-1 (ums8) + . nsy-1 (RNAi) (133) (B) Регистрировали развитие RNAi-обработанных животных указанных генотипов до личиночной стадии L4 или старше.Данные представляют собой среднее значение трех пластин с полосами погрешностей, показывающими стандартное отклонение между пластинами. Размеры выборки для этого эксперимента: дикий тип + L4440 (1200), дикий тип + nsy-1 (RNAi) (1179), nsy-1 (ums8) + L4440 (956) и nsy. -1 (ums8) + nsy-1 (RNAi) (1010). Данные являются репрезентативными для двух биологических повторов. * P <0,001.
Микроскопия
Нематод помещали на подушечки с 2% агаром, парализованных левамизолом (Sigma-Aldrich Co.) и сфотографировали с помощью микроскопа AXIO Imager Z1 с камерой AxioCam HRm и программным обеспечением Axiovision 4.6 (Zeiss) или Eclipse E400 с камерой DS-QilMc и программным обеспечением NIS Elements Imaging (Nikon Corporation). Фотографии были получены с использованием тех же условий визуализации для данного эксперимента и обработаны в Photoshop (Adobe Systems, Inc.).
Статистический анализ и выравнивание аминокислот
Различия в выживаемости C. elegans животных в P.aeruginosa определяли патогенез с помощью лог-рангового теста. Кратные изменения в анализах qRT-PCR и различия в развитии животных до указанной стадии сравнивали с использованием непарных двусторонних тестов Стьюдента t . Аминокислотное выравнивание между ASK1 человека, Drosophila Pk92B и C. elegans NSY-1 определяли с помощью ClustalW2 (Larkin et al. 2007).
Результаты и обсуждение
Экзогенная гиперактивация пути киназы p38 MAP токсична для развивающихся C.elegans
Небольшая молекула R24, также называемая RPW-24, действует выше пути p38 MAPK, вызывая индукцию предполагаемых антибактериальных иммунных эффекторов в кишечнике C. elegans и, соответственно, защищает нематод от бактериальной инфекции (Pukkila -Worley и др. 2012, 2014). Кроме того, воздействие R24 вызывает индукцию ферментов детоксикации и задерживает развитие нематод дикого типа в отсутствие патогена, что позволяет предположить, что это соединение токсично для C.elegans в нормальных лабораторных условиях роста (Pukkila-Worley et al. 2012, 2014). Чтобы исследовать взаимосвязь между иммуностимулирующими свойствами и токсичностью этого противоинфекционного ксенобиотика, мы провели скрининг нематод, происходящих из примерно 100000 мутагенизированных гаплоидных геномов, на наличие мутантов, которые были способны развиваться быстрее в присутствии 140 мкМ R24 (2). Мы выбрали эту концентрацию, потому что фенотип задержки развития был более выраженным, чем наблюдаемый для 70 мкМ R24, концентрации, которую мы использовали в наших предыдущих исследованиях (Pukkila-Worley et al. 2012, 2014). Для скрининга мы требовали, чтобы выбранные мутанты были личиночной стадии L4 или старше в момент времени, когда 100% немутагенизированных червей, обработанных параллельно R24, находились на стадии личинки L2 или L3. Было идентифицировано семь мутантов с фенотипом супрессора ксенобиотической токсичности (Xts), и мы сосредоточили внимание на пяти мутантах с наиболее пенетрантными фенотипами Xts, которые мы назвали ums1 , ums2 , ums3 , ums4 и ums5. ().Чтобы идентифицировать мутацию в этих штаммах, которая позволила улучшить развитие в присутствии R24, мы использовали технологию секвенирования следующего поколения (Sarin et al. 2008; Minevich et al. 2012). Для трех мутантов ( ums3 , ums4 и ums5 ) мы секвенировали ДНК из объединенных рекомбинантов F2, которые имели мутантный фенотип после обратного скрещивания с животными N2 дикого типа, и для двух мутантов ( ums1 и ums2 ) мы идентифицировали мутации после секвенирования исходного мутантного штамма.Интересно, что каждый из этих пяти мутантов содержал миссенс-мутации в одном из четырех известных компонентов сигнальной кассеты p38 MAPK ( tir-1 , nsy-1 , sek-1 или pmk-1 ,).
Таблица 1
Супрессоры токсичности ксенобиотиков
| Ген | Аллель | Мутация | Описание, Гомолог млекопитающих |
|---|---|---|---|
| 19 | |||
| tir-1 | ums3 | A723T | Адаптерный белок TIR-домена, SARM |
| nsy-1 | 20474 9077 L | ||
| sek-1 | ums5 | G199S | MAPKK, MKK3 / 6 |
| pmk-1 | pmk-1 | 904 9047 pmk-1 904 9047
Чтобы охарактеризовать эти недавно выделенные мутанты пути MAPK p38, мы провели исследование патогенеза анализы с P.aeruginosa и обнаружили, что, как и в случае с классическими мутантами с потерей функции в компонентах пути MAPK p38, четыре из пяти мутантов [ tir-1 (ums2) , tir-1 (ums3) , pmk-1 (ums4) и sek-1 (ums5) ] имели фенотип повышенной чувствительности к патогенам (Esp) ( P <0,001,). nsy-1 (ums1) , самый слабый супрессор токсичности из этих пяти мутантов, не обладал очевидным фенотипом Esp. Помимо фенотипа Esp, мутации потери и снижения функции в вышестоящих компонентах пути p38 MAPK PMK-1 вызывают снижение количества активированного PMK-1, что может быть обнаружено в эксперименте иммуноблоттинга с использованием антитело, которое специфически распознает дважды фосфорилированный мотив TGY PMK-1 (Kim et al. 2002; Liberati et al. 2004). Мы обнаружили, что nsy-1 (ums1) , tir-1 (ums2) , tir-1 (ums3) , pmk-1 (ums4) и sek-1 (ums5) каждый уменьшили уровни активной формы PMK-1 по сравнению с контролем дикого типа (). Эти данные показывают, что прямой генетический скрининг мутаций, которые придают устойчивость к токсическим эффектам иммуностимулирующего ксенобиотика R24, выявил гипоморфные аллели в пути p38 MAPK. Интересно, что предыдущий прямой генетический скрининг мутаций потери функции в компонентах пути MAPK p38 выявил два аллеля, pmk-1 ( qd9 ) и sek-1 (qd37) , которые имели идентичную миссенс-мутацию. как pmk-1 (ums4) и по той же аминокислоте, что и sek-1 (ums5) , соответственно (Shivers et al. 2010 г.).
В совокупности эти данные демонстрируют, что токсичность R24 может быть подавлена гипоморфными мутациями в пути p38 MAPK, хотя не было выявлено ни одной мутации, которая приводила бы к прогрессированию роста, аналогичному червям, в отсутствие соединения. Чтобы подтвердить это наблюдение, мы изучили ранее охарактеризованные нулевые аллели tir-1 (qd4) , nsy-1 (ag3) и pmk-1 (km25) (Kim et al. 2002; Shivers et al. al. 2009) () и обнаружили, что эти мутации также подавляли индуцированную R24 задержку развития до степени, сравнимой с мутантом pmk-1 (ums4) и tir-1 (ums2) и tir- 1 (ums3) аллелей соответственно ().
Таким образом, эти данные показывают, что гиперактивация иммунной защиты p38 MAPK токсична для развивающихся нематод.
Прямой генетический скрининг обнаруживает аллель усиления функции в MAPKKK nsy-1
Для дальнейшего изучения физиологических последствий гиперактивации врожденного иммунитета у C. elegans мы разработали прямой генетический скрининг на эндогенные активаторы MAP p38. передача сигналов киназы PMK-1. Репортер транскрипции врожденного иммунитета F08G5.6 :: GFP индуцируется во время P.aeruginosa и, в значительной степени, противоинфекционным ксенобиотиком R24, в зависимости от пути p38 MAPK PMK-1 (Pukkila-Worley et al. 2012, 2014). Поэтому мы пришли к выводу, что скрининг доминантных активаторов F08G5.6 :: GFP позволит выявить мутации, которые вызывают конститутивную активацию пути p38 MAPK. Мы проверили потомство F1 мутагенизированных животных F08G5.6 :: GFP и идентифицировали единственный мутантный аллель, ums8 , примерно из 170 000 мутагенизированных гаплоидных геномов.После скрещивания 1X и 2X с животными N2 дикого типа мы объединили рекомбинанты F2, которые были гомозиготны по мутантному фенотипу, и секвенировали геномы этих образцов, используя технологию секвенирования следующего поколения (Sarin et al. 2008; Minevich et al. al. 2012). Это исследование показало, что мутант ums8 содержал миссенс-мутацию G → A в кодирующей области nsy-1 , которая привела к замене сильно консервативного Arg 246 на Gln 246 ().NSY-1 / ASK1 представляет собой консервативный MAPKKK, который действует выше p38 MAPK PMK-1, чтобы регулировать иммунные и стрессовые реакции у C. elegans (Kim et al. 2002). Мы превратили штамм ums8 в общей сложности шесть раз в штамм N2 дикого типа и подтвердили наличие мутации R246Q и активацию экспрессии F08G5.6 :: GFP в штамме, подвергнутом скрещиванию.
Аллель nsy-1 (ums8) кодирует мутацию усиления функции в nsy-1 .(A) Представлена предполагаемая доменная архитектура NSY-1, основанная на гомологии с ASK1 млекопитающих. Эта схема была адаптирована из Bunkoczi et al. (2007). Предполагаемое расположение центрального домена серин-треонинкиназы и двух спиральных доменов на N- и C-концах показано красным и синим соответственно. Граница N-концевого отрицательного регуляторного домена относительно белка NSY-1 представлена над диаграммой. Arg, который был мутирован в Gln в штамме ums8 (R246Q), выделен красным.Выравнивание аминокислотных последовательностей ASK1 человека, Drosophila Pk92B (гомолог ASK1) и C. elegans NSY-1 демонстрирует, что мутация ums8 расположена в сильно консервативной области. Темно-синяя заливка указывает на идентичные аминокислоты, прогрессивная более светлая синяя заливка указывает на уровень сходства в классе аминокислот, определенный с помощью программного обеспечения ClustalW2. (B) Животных дикого типа (WT) и nsy-1 (ums8) животных подвергали воздействию питающего штамма бактерий РНКи, трансформированного контрольным вектором (L4440), или двумя отдельными конструкциями РНКи (pHC1 и pHC2), которые нацелены на разные области кодирующей области в гене nsy-1 и сфотографированы.Зеленый цвет соответствует F08G5.6 :: GFP индукции, а красный — myo-2 :: mCherry , который использовался в качестве маркера для совместной инъекции. (C) Экспрессию указанных генов определяли с помощью qRT-PCR в гетерозиготах дикого типа (+ / +), nsy-1 (ums8) / + и nsy-1 (ums8) / nsy-1 ( ums8) гомозигот. F08G5.6 :: GFP использовали в качестве штамма дикого типа. Данные представляют собой среднее значение трех повторов, каждый нормализованный к контрольному гену с полосами ошибок, представляющими SEM, и представлены как значение относительно средней экспрессии указанного гена у животных дикого типа.* равно P <0,05. Не было статистической разницы в уровнях индукции между гетерозиготными и гомозиготными образцами для любого гена. Дополнительные протестированные гены см. На рисунке S1A. (D) Иммуноблот-анализ лизатов личиночных животных L4 указанного генотипа с использованием антител, которые распознают дважды фосфорилированный мотив TGY PMK-1 (p-PMK-1) и актина. Тридцать мкг общего белка nsy-1 (ums8) , pmk-1 (km25) и nsy-1 (ag3) загружали в гель вместе с серией разведений матрицы дикого типа [15 мкг, 20 мкг и 30 мкг общего белка (слева направо)] для контроля способности антитела p-PMK-1 обнаруживать различные концентрации субстрата.Стрелкой справа выделена полоса PMK-1, которая отсутствует у мутантов pmk-1 (km25) и nsy-1 (ag3) . Стрелки слева указывают на неспецифические полосы. Данные являются репрезентативными для двух биологических повторов.
Чтобы определить, вызвана ли конститутивная экспрессия иммунного репортера F08G5.6 :: GFP в мутанте ums8 мутацией G → A в кодирующей области nsy-1 , мы использовали РНКи для подавления экспрессии nsy-1 в мутанте ums8 и обнаружили, что эта обработка подавляла F08G5.6 :: GFP индукционный (). Более того, РНКи-опосредованный нокдаун nsy-1 у животных дикого типа не вызывал индукцию F08G5.6 (см. Данные ПЦР qRT, обсуждаемые в следующем разделе). Кроме того, индукция F08G5.6 :: GFP в штамме nsy-1 (ums8) подавлялась посредством РНКи-опосредованного нокдауна pmk-1 , p38 MAPK, который находится ниже nsy-1. (рисунок S2A). Мы также использовали qRT-PCR, чтобы подтвердить, что nsy-1 (ums8) является доминантным аллелем с усилением функции, который управляет конститутивной активацией F08G5.6 :: GFP и сам ген F08G5.6 (). F08G5.6 и GFP индуцировались в одинаковой степени у гетерозигот nsy-1 (ums8) / + и у гомозигот nsy-1 (ums8) / nsy-1 (ums8) по сравнению с гомозиготами дикого типа. контроли, несущие трансген F08G5.6 :: GFP (+ / +).
В соответствии с нашими исследованиями, предполагающими, что nsy-1 (ums8) является аллелем усиления функции nsy-1 , мы провели иммуноблот-анализ белковых лизатов от животных nsy-1 (ums8 ) и обнаружили, что животные nsy-1 (ums8) имели большее количество активированного p38 MAPK PMK-1, чем контрольные животные ().
Таким образом, эти данные характеризуют аллель усиления функции MAPKKK nsy-1 , который вызывает гиперактивацию пути p38 MAPK.
Аллель усиления функции nsy-1 (ums8) вызывает конститутивную активацию зависимых от p38 MAPK PMK-1 эффекторов врожденного иммунитета
Чтобы определить, была ли базальная экспрессия других генов гиперактивированной в nsy-1 ( ums8) , мы использовали систему nanoString nCounter для сравнения профиля экспрессии 118 генов врожденного иммунитета и стрессовой реакции в nsy-1 (ums8) при усилении функции, nsy-1 (ag3) при потере функции. -функции и животные дикого типа, когда штаммы питались обычным лабораторным источником корма E.кишечная палочка OP50 (). Из 118 генов в этом кодовом наборе 24 были транскрипционно активированы по крайней мере в пять раз у мутантов nsy-1 (ums8) по сравнению с животными дикого типа (Таблица S1). Семь из 24 генов, наиболее сильно активированных в аллеле усиления функции nsy-1 (ums8) , были среди 12 наиболее сильно сниженных в аллеле потери функции nsy-1 (ag3) ( F08G5). .6 , F35E12.5 , C09H5.2 , T24B8.5 , clec-67 , C32h21.12 и oac-6 и Таблица S1). Действительно, эта группа из семи генов включает F08G5.6 и пять других предполагаемых иммунных эффекторов ( F35E12.5 , C09H5.2 , T24B8.5 , C32h21.12 и clec-67 ). которые ранее были охарактеризованы мишенями p38 MAPK PMK-1 (Таблица S1) (Troemel et al. 2006). Следует отметить, что кодовый набор включал четыре дополнительных гена, для базовой или индуцированной патогеном экспрессии которых требуется p38 MAPK PMK-1 ( F49F1.6 , C17h22.8 , C32h21.1 , K08D8.5 , lys-2 ), и каждый из этих генов индуцировался по крайней мере дважды у мутантов nsy-1 (ums8) , и репрессированы по крайней мере в четыре раза у мутантов nsy-1 (ag3) (Таблица S1). Кроме того, мы ранее использовали этот кодовый набор для идентификации 13 генов, индукция которых иммуностимулирующим противоинфекционным ксенобиотиком R24 зависела от p38 MAPK PMK-1 (Pukkila-Worley et al. 2014). Девять из этих 13 генов были активированы более чем в пять раз, и все эти гены были индуцированы по крайней мере вдвое у мутанта с усилением функции nsy-1 (ums8) (таблица S1).Таким образом, аллель усиления функции nsy-1 (ums8) управляет конститутивной активацией генов, чья базальная, индуцированная патогеном или индуцированная R24 экспрессия зависит от пути MAPK p38.
p38 MAPK-зависимые предполагаемые иммунные эффекторы конститутивно активируются в мутанте nsy-1 (ums8) . (A) Диаграмма разброса сравнивает экспрессию 118 генов C. elegans , которые были проанализированы с использованием системы экспрессии генов nanoString nCounter у животных дикого типа, nsy-1 (ums8) и nsy-1 (ag3) животных. .Данные представляют собой среднее значение двух повторов для nsy-1 (ums8) и nsy-1 (ag3) и взяты из одного образца для дикого типа. Экспрессию каждого гена нормализовали до среднего геометрического экспрессии трех контрольных генов. Гены, которые находятся за пределами двух параллельных линий на каждом графике, регулируются по-разному более чем в пять раз по сравнению с экспрессией у животных дикого типа. Выделены гены, которые ранее были охарактеризованы мишенями p38 MAPK PMK-1 и сильно дифференцированно регулировались в этом эксперименте.См. Таблицу S1 для уровней экспрессии всех генов. (B) qRT-PCR использовали для изучения экспрессии шести предполагаемых иммунных эффекторов у RNAi-обработанных животных на смешанной стадии указанных генотипов. Все животные были выращены на питающем штамме РНКи-бактерий HT115, экспрессирующем пустой вектор L4440, за исключением двух указанных образцов, которые подвергались воздействию бактерий, экспрессирующих конструкцию nsy-1 (РНКи) . Расположение на диаграммах рассеяния этих генов указано в (A) красными (слева) и синими (справа) точками.Данные представляют собой среднее значение трех повторов, каждый нормализованный к контрольному гену с полосами ошибок, представляющими SEM, и представлены как значение относительно средней экспрессии указанного гена у животных дикого типа. * равно P <0,05.
Мы подтвердили эти данные в трех биологических повторностях образцов с помощью qRT-PCR для изучения шести предполагаемых иммунных эффекторов, которые дифференциально регулировались в анализе nanoString и являются известными мишенями пути p38 MAPK. Все шесть генов были транскрипционно репрессированы в мутантах с потерей функции nsy-1 (RNAi), и nsy-1 (ag3), и были активированы в аллеле nsy-1 (ums8) ().РНКи-опосредованный нокдаун nsy-1 частично подавлял конститутивную активацию этих шести предполагаемых иммунных эффекторов в аллеле nsy-1 (ums8) (). Кроме того, мы обнаружили, что nsy-1 (ums8) / + гетерозигот вызвали индукцию трех p38 MAPK-зависимых иммунных эффекторов до уровней, эквивалентных таковому у гомозиготных животных nsy-1 (ums8) (рис. S1A), так как мы наблюдали для F08G5.6 (). Следует отметить, что мы также подвергли мутант nsy-1 (ums8) воздействию R24 и обнаружили, что F08G5.6 :: Экспрессия GFP была заметно увеличена по сравнению с необработанными животными nsy-1 (ums8 ), что позволяет предположить, что эффекты R24 и аллеля усиления функции nsy-1 на экспрессию генов являются аддитивными (рис. S1B). В соответствии с этим наблюдением мы ранее обнаружили, что R24 также дополнительно усиливает экспрессию иммунных эффекторов P. aeruginosa , которые индуцируются во время бактериальной инфекции (Pukkila-Worley et al. 2012).
Предыдущий анализ полногеномного профиля транскриптома выявил группу предполагаемых иммунных эффекторов, базальная экспрессия которых сильно подавлена у мутантов pmk-1 (km25) (Troemel et al. 2006 г.). Термин базальная регуляция использовался в этом контексте для описания экспрессии генов у животных, которые растут на лабораторном источнике пищи ( E. coli OP50) и используется для отличия от вызванной патогеном экспрессии предполагаемых иммунных эффекторов, наблюдаемой во время микробной инфекции. Помимо контроля базовой экспрессии предполагаемых иммунных эффекторов, ряд исследований продемонстрировал, что p38 MAPK PMK-1 также необходим для индуцированной патогеном экспрессии предполагаемых иммунных эффекторов (Troemel et al. 2006; Bolz et al. 2010; Pukkila-Worley et al. 2012 г.). Однако также ясно, что существует группа генов, идентичность которых полностью не определена, которые требуют пути p38 MAPK PMK-1 для их индукции, но не для их базовой экспрессии. Данные nanoString мутанта nsy-1 (ums8) показывают, что этот аллель усиления функции может использоваться для идентификации этой группы эффекторов врожденного иммунитета. Из 24 генов в нашем кодовом наборе, которые были наиболее сильно индуцированы у мутанта nsy-1 (ums8) , 17 не зависели от NSY-1 в своей базовой экспрессии — группа, которая включает гены патогена и стресс-реакции (таблица S1).
Интересно также отметить, что гены, участвующие в детоксикации низкомолекулярных токсинов, таких как цитохром P450 (CYP), s-глутатион-s-трансферазы (GST) и UDP-глюкуронозилтрансферазы (UDP), не были среди тех, у кого транскрипционно повышенная регуляция. в аллеле усиления функции nsy-1 (ums8) (Таблица S1). Ранее мы наблюдали, что противоинфекционный ксенобиотик R24 вызывал устойчивую индукцию этих классов генов в дополнение к генам, участвующим в ответе на патогены (Pukkila-Worley et al. 2012 г.). Однако, в отличие от индукции противомикробных иммунных эффекторов, активация генов детоксикации с помощью R24 не зависела от пути p38 MAPK PMK-1 (Pukkila-Worley et al. 2014). Вместе с результатами этого исследования эти данные свидетельствуют о том, что гиперстимуляция врожденного иммунитета p38 MAPK сама по себе не приводит к индукционным реакциям детоксикации малых молекул, и что другие клеточные механизмы задействованы для распознавания и реагирования на токсины ксенобиотиков в C.elegans .
Эндогенная гиперактивация пути p38 MAPK PMK-1 защищает нематод от бактериальной инфекции, но токсична для C. elegans
Стимуляция пути p38 MAPK маломолекулярным ксенобиотиком R24 защищает C. elegans от бактериальной инфекции путем стимуляции экспрессия иммунного эффектора через путь MAPK p38 (Pukkila-Worley et al. 2012, 2014). В этом контексте иммунная гиперактивация имеет негативные физиологические последствия для нематод, развивающихся в нормальных лабораторных условиях ().Поэтому мы предположили, что эндогенная гиперактивация пути p38 MAPK посредством мутации увеличения функции в MAPKKK NSY-1 будет демонстрировать фенотип, аналогичный фенотипу червей, обработанных R24.
Мы проверили восприимчивость nsy-1 (ums8) мутантных животных с усилением функции к инфекции P. aeruginosa по сравнению с животными дикого типа и обнаружили, что nsy-1 (ums8) животных были устойчивыми. к гибели с помощью P. aeruginosa (и рис. S3).РНКи-опосредованный нокдаун nsy-1 подавлял фенотип устойчивости nsy-1 (ums8) животных (). Интересно, что развитие личинок nsy-1 (ums8) было заметно задержано по сравнению с контрольными животными дикого типа, что было количественно определено путем определения количества мутантных животных и животных дикого типа, которые достигли стадии L4 из синхронизированных яиц через 3 дня инкубация при 20 ° (). Нокдаун nsy-1 у животных nsy-1 (ums8) частично подавлял этот фенотип (и).Мы также наблюдали, что nsy-1 (ums8) животных имели заметно меньшие размеры выводка, чем контрольные животные дикого типа (Рисунок S2B), данные, которые напоминают наши наблюдения с иммуностимулирующим ксенобиотиком R24 (Pukkila-Worley et al. 2012). ). Кроме того, опосредованный РНКи нокдаун pmk-1 также подавлял замедленное развитие nsy-1 (ums8) животных (фигура S2C).
В этом исследовании мы предоставляем прямые доказательства двух прямых генетических скринингов, что аберрантная стимуляция врожденных иммунных ответов, опосредованных p38 MAPK PMK-1, посредством мутации усиления функции в nsy-1 или экзогенного добавления антибиотика. -инфекционный R24 вызывает иммунные реакции, которые являются защитными во время инфекции, но токсичны для нематод в нормальных условиях роста.Интересно отметить, что все аллели с наиболее пенетрантными фенотипами Xts имели гипоморфные мутации в известных компонентах пути MAPK p38. Следует отметить, что прямой генетический скрининг компонентов пути p38 MAPK также не выявил мутаций в любом гене, который функционирует выше tir-1 (Shivers et al. 2010). Вместе эти данные подтверждают, что сигнальная кассета p38 MAPK получает множественные входы для регуляции защитных ответов хозяина у C. elegans .
В дополнение к содействию избавлению от вторгающихся патогенов, в новой литературе установлено, что системы хозяина также действуют, способствуя толерантности к инфекции, тем самым уменьшая негативное влияние инфекции на приспособленность организма (Ayres and Schneider 2008; 2009). Концептуально механизмы толерантности хозяина действуют, чтобы смягчить ущерб, причиненный патогеном, а также ограничить побочное повреждение, связанное с активацией иммунной системы (Ayres and Schneider 2012; Medzhitov et al. 2012 г.). Наши исследования иммуностимулирующего ксенобиотика R24 и аллеля усиления функции nsy-1 (ums8) предполагают, что механизмы толерантности хозяина могут функционировать у нематод, ограничивая иммунопатологию, связанную с аберрантной активацией p38 MAPK и последующей гиперпродукцией иммунных эффекторов. В C. elegans ответ развернутого белка (UPR) является одним из таких механизмов, который, как было показано, способствует толерантности во время иммунной активации (Richardson et al., 2010; Sun et al. 2011). Ветвь IRE1-XBP1 / Hac1 UPR необходима для преодоления физиологического стресса, связанного с увеличением активности p38 MAPK PMK-1, которая была экспериментально подтверждена воздействием P. aeruginosa или посредством RNAi-опосредованного нокдауна MAPK. фосфатаза vhp-1 , отрицательный регулятор PMK-1 (Richardson et al. 2010; Mizuno et al. 2004; Kim et al. 2004). В соответствии с этими данными мы обнаружили, что мутанты xbp-1 (zc12) резко чувствительны к токсическим эффектам иммуностимулирующего ксенобиотика R24 в анализе развития по сравнению с контрольными животными (рисунок S4).Таким образом, вероятно, существует по крайней мере два механизма в C. elegans , чтобы гарантировать иммунный гомеостаз и способствовать толерантности во время инфекции. UPR функционирует для защиты хозяина от стресса ER, который возникает как следствие активации p38 MAPK (Richardson et al. 2010). Кроме того, выделение аллеля усиления функции в nsy-1 свидетельствует о том, что p38 MAPK PMK-1-опосредованные транскрипционные ответы контролируются с помощью вышестоящих механизмов, которые негативно регулируют иммунную активацию на уровне NSY-1.
Человеческий гомолог NSY-1, ASK1, регулирует активность p38 по крайней мере с помощью трех консервативных белковых доменов: центрального серин-треонинкиназного домена и двух спиральных доменов на N- и C-концах, соответственно () (Bunkoczi et al. al. 2007). N-концевой домен связывает тиоредоксин (TRX), который подавляет функцию NSY-1. Интересно, что мутация ums8 с усилением функции, в которой отрицательно заряженный Arg заменен на Gln, незаряженную аминокислоту с большой полярной боковой цепью, расположена в сильно консервативной области, на восемь аминокислот выше области. в NSY-1, согласно прогнозам гомологии, кодирует этот N-концевой регуляторный домен ().Мы предполагаем, что активность C. elegans NSY-1 обычно контролируется отрицательным регуляторным фактором, который связывается с N-концевым доменом спиральной спирали. Таким образом, нарушение этой области с помощью мутации ums8 может объяснить конститутивную активацию иммунных эффекторов p38 MAPK, приводящую к фенотипу устойчивости к патогену и токсичности для развивающихся нематод. В C. elegans имеется пять гомологов тиоредоксина (TRX), и будет интересно определить, функционируют ли какие-либо из этих генов как негативные регуляторы NSY-1.
UMS8 / SL-DSF — durchschraubbar oder freistehend
Zubehör
Schrauben für die Bodenverankerung:
M16 для UMS8 / SL-DSF
Isolierrondelle zur Schraubenentkopplung bei Bodenverankerungen:
RONKAP 1 для UMS 8 / SL-DSF
SL-DSF — durchschraubbar zur Bodenverankerung der Maschine oder freistehend
Bestückung
Oberseite: Lieferung mit Gleitschutzplatten GPL 3025 ab Werk.
Unterseite: Bitte wählen Sie die gewünschte Schwingungsisolierplatte IPL aus.
Produktdaten, Abmessungen, Preise
Welche Schwingungsisolierplatte (IPL) eignet sich für Ihre Maschine:
Maschinenkategorie…BandsägenBearbeitungszentrenBügelsägenDrehbänkeDrehmaschinenDruckmaschinenExtruderFalzmaschinenFräsmaschinenFörderbänderKlimageräteKompressorenKreissägenLüfterMagazineMessmaschinenMesstischeMikroskopeMühlenOptische GerätePapierschneidemaschinenPressenPumpenSchleif- / FinishmaschinenSchmiedenSchredderSpritzgießmaschinenStanzenTextilschneidemaschinenUmformmaschinen (spanlos) VentilatorenVerdichterVerpackungsmaschinenWaagenWerkzeugmaschinenZerkleinerungsmaschinenZuführungseinrichtungenZusammentragmaschinenAusgewählter Maschinentyp Ausgewählt
| Арт.‑Nr. |
Тип | Preis ** | Fmax в N *** | л * | Б * | h2 * | h3 * | Verstellbereich V * | а * | К * | e * | с * | d1 * | d2 * | d4 * | SW3 * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 305S11 | UMS8 / SL-DSF / 10 |
74,50 € |
6000 |
110 |
110 |
56 |
74 |
+ 7 / -3 |
145 |
13,5 |
36 |
55 |
22 |
22 |
– |
22 |
* Alle Angaben в мм
** Alle Preise zzgl.MwSt.
*** Fmax = F (VSP) + F (G) + F (DYN) в N; F (VSP): Schraubenvorspannkraft bei Bodenverankerung, F (G): anteilige Gewichtskraft der Maschine, F (DYN): anteilige Dynamische Kräfte
| Арт.‑Nr. |
Тип | Preis ** | Fmax в N *** | л * | Б * | h2 * | h3 * | Verstellbereich V * | а * | К * | e * | с * | d1 * | d2 * | d4 * | SW3 * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 305S16 | UMS8 / SL-DSF / 17 |
74,50 € |
24000 |
110 |
110 |
56 |
74 |
+ 7 / -3 |
145 |
13,5 |
36 |
55 |
22 |
22 |
– |
22 |
* Alle Angaben в мм
** Alle Preise zzgl.MwSt.
*** Fmax = F (VSP) + F (G) + F (DYN) в N; F (VSP): Schraubenvorspannkraft bei Bodenverankerung, F (G): anteilige Gewichtskraft der Maschine, F (DYN): anteilige Dynamische Kräfte
| Арт.‑Nr. |
Тип | Preis ** | Fmax в N *** | л * | Б * | h2 * | h3 * | Verstellbereich V * | а * | К * | e * | с * | d1 * | d2 * | d4 * | SW3 * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 305S21 | UMS8 / SL-DSF / 20 |
74,50 € |
14500 |
110 |
110 |
56 |
74 |
+ 7 / -3 |
145 |
13,5 |
36 |
55 |
22 |
22 |
– |
22 |
* Alle Angaben в мм
** Alle Preise zzgl.MwSt.
*** Fmax = F (VSP) + F (G) + F (DYN) в N; F (VSP): Schraubenvorspannkraft bei Bodenverankerung, F (G): anteilige Gewichtskraft der Maschine, F (DYN): anteilige Dynamische Kräfte
| Арт.‑Nr. |
Тип | Preis ** | Fmax в N *** | л * | Б * | h2 * | h3 * | Verstellbereich V * | а * | К * | e * | с * | d1 * | d2 * | d4 * | SW3 * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 305S212 | UMS8 / SL-DSF / 20-2 |
74,50 € |
14500 |
110 |
110 |
56 |
74 |
+ 7 / -3 |
145 |
13,5 |
36 |
55 |
22 |
22 |
– |
22 |
* Alle Angaben в мм
** Alle Preise zzgl.MwSt.
*** Fmax = F (VSP) + F (G) + F (DYN) в N; F (VSP): Schraubenvorspannkraft bei Bodenverankerung, F (G): anteilige Gewichtskraft der Maschine, F (DYN): anteilige Dynamische Kräfte
| Арт.‑Nr. |
Тип | Preis ** | Fmax в N *** | л * | Б * | h2 * | h3 * | Verstellbereich V * | а * | К * | e * | с * | d1 * | d2 * | d4 * | SW3 * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 305S251 | UMS8 / SL-DSF / 25 |
78,20 € |
20570 |
110 |
110 |
56 |
84 |
+ 7 / -3 |
145 |
13,5 |
36 |
55 |
22 |
22 |
– |
22 |
* Alle Angaben в мм
** Alle Preise zzgl.MwSt.
*** Fmax = F (VSP) + F (G) + F (DYN) в N; F (VSP): Schraubenvorspannkraft bei Bodenverankerung, F (G): anteilige Gewichtskraft der Maschine, F (DYN): anteilige Dynamische Kräfte
| Арт.‑Nr. |
Тип | Preis ** | Fmax в N *** | л * | Б * | h2 * | h3 * | Verstellbereich V * | а * | К * | e * | с * | d1 * | d2 * | d4 * | SW3 * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 305S31 | UMS8 / SL-DSF / 30 |
74,50 € |
60000 |
110 |
110 |
56 |
74 |
+ 7 / -3 |
145 |
13,5 |
36 |
55 |
22 |
22 |
– |
22 |
* Alle Angaben в мм
** Alle Preise zzgl.MwSt.
*** Fmax = F (VSP) + F (G) + F (DYN) в N; F (VSP): Schraubenvorspannkraft bei Bodenverankerung, F (G): anteilige Gewichtskraft der Maschine, F (DYN): anteilige Dynamische Kräfte
| Арт.‑Nr. |
Тип | Preis ** | Fmax в N | л * | Б * | h2 * | h3 * | Verstellbereich V * | а * | К * | e * | с * | d1 * | d2 * | d4 * | SW3 * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 305S36 | UMS8 / SL-DSF / 32 |
74,50 € |
48400 |
110 |
110 |
56 |
74 |
+ 7 / -3 |
145 |
13,5 |
36 |
55 |
22 |
22 |
– |
22 |
* Alle Angaben в мм
** Alle Preise zzgl.MwSt.
*** Fmax = F (VSP) + F (G) + F (DYN) в N; F (VSP): Schraubenvorspannkraft bei Bodenverankerung, F (G): anteilige Gewichtskraft der Maschine, F (DYN): anteilige Dynamische Kräfte
| Арт.‑Nr. |
Тип | Preis ** | Fmax в N *** | л * | Б * | h2 * | h3 * | Verstellbereich V * | а * | К * | e * | с * | d1 * | d2 * | d4 * | SW3 * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 305S41 | UMS8 / SL-DSF / 40 |
71,00 € |
24000 |
110 |
110 |
56 |
67 |
+ 7 / -3 |
145 |
13,5 |
36 |
55 |
22 |
22 |
– |
22 |
* Alle Angaben в мм
** Alle Preise zzgl.MwSt.
*** Fmax = F (VSP) + F (G) + F (DYN) в N; F (VSP): Schraubenvorspannkraft bei Bodenverankerung, F (G): anteilige Gewichtskraft der Maschine, F (DYN): anteilige Dynamische Kräfte
Продукция Isoloc NTS120 / 20-2 V Продукция Isoloc — 航 欧 机电 设备 有限公司
上海 航 欧 专业 销售 Isoloc Продукция
Изолок НТС100 / 30-НР.51003-2
Изолок M24x1,5×100-NR.72150
Изолок NTS120 / 20-2 ВЕРСИЯ V13A, 51202
Изолок NIVELLIERSCHR.M16X100 VERS.13A 70030
Изолок изолок 30530; UMS 5-ASF / 30
Изолок изолок 30897; УМС 8-АСА / БВ 300х230х15
Изолок 081515146-0000 СТЗЭ НЦ100 / 30-НР.51003-2-ИЗОЛОК
Изолок 081516487-0000 STZE M24x1,5×100-NR.72150-ИЗОЛОК
Изолок АКСЕССУАРЫ ОБУВЬ ОБУВЬ 30523
Изолок фиксатор 30530 UMS5-ASF / 30
Изолок фиксатор 30897; УМС8-АСА / BV300x230x15
Изолок винт 73134
Изолок UMS8 ASF / 20
Изолок NTS100 / 30-NR.51003-2Isoloc Nr 70030 脚垫 柱
Isoloc Nr 5120202 防震 脚垫
Изолок М16 x 150 мм Nr: 73132 房 震 脚垫
Изолок НТ 80/30 №: 40803 房 震 脚垫
Isoloc 50802 NTS80 / 20 d1 = 80 мм h2 = 32 мм 支撑 头
Isoloc 70562 M20x1.5×150 мм 连杆
Изолок 70516 (М 16 * 100 мм) 机床 零件
Изолок 5120202 (НТС 120 / 20-2) 机床 零件
Изолок M24x1,5×100-NR.72150
Изолок NTS120 / 20-2 ВЕРСИЯ V13A, 51202
Изолок NIVELLIERSCHR.M16X100 VERS.13A 70030
Изолок изолок 30530; UMS 5-ASF / 30
Изолок изолок 30897; УМС 8-АСА / БВ 300х230х15
Изолок 081515146-0000 СТЗЭ НЦ100 / 30-НР.51003-2-ИЗОЛОК
Изолок 081516487-0000 STZE M24x1,5×100-NR.72150-ISOLOC
Изолок АКСЕССУАРЫ ОБУВЬ ОБУВЬ 30523
Изолок фиксатор 30530 UMS5-ASF / 30
Изолок фиксатор 30897; УМС8-АСА / BV300x230x15
Изолок винт 73134
Изолок UMS8 ASF / 20
Изолок NIVELLIERSCHR.M16X100 ВЕРС.V13A, ART-NR: 70030
Изолок НТС 120 / 20-2 ВЕРСИЯ V13A, ART-NR: 5120202
Изолок 30523
Изолок 30530; UMS5-ASF / 30
Изолок 30897; UMS8-ASA / BV300x230x15
Изолок 73134
Изолок СТЗЭ М24х1,5х100-НР.72150
Изолок STZE NTS100 / 30-NR.51003-2
Изолок NT Тип 80/30
Изолок M16 * 150
Изолок UMS5-ASF / 3050 + 25 LXBXH: 110X110X83
Изолок UMS8-ASF / 25 LXBXH: 150X144X84
Изолок UMS M16X150
Изолок ВИНТ / 73134
Изолок ПЛИТА / 13010 IPL30 100X50X15 ROT OP
Изолок UMS 18 KDS
Изолок UMS 8 KDSAISOLOC 081515146-0000 STZE NTS100 / 30-NR.51003-2-ИЗОЛОК
ISOLOC 081516487-0000 STZE M24x1,5×100-NR.72150-ИЗОЛОК
АКСЕССУАРЫ ISOLOC 30523
Фиксатор ISOLOC 30530 UMS5-ASF / 30
Фиксатор ISOLOC 30897; UMS8-ASA / BV300x230x15
Выравнивающий винт ISOLOC 73134
ИЗОЛОК UMS8 ASF / 20
ИЗОЛОК NIVELLIERSCHR.M16X100 VERS.V13A, ART-NR: 70030
ИЗОЛОК НТС 120 / 20-2 ВЕРСИЯ V13A, ART-NR: 5120202
ISOLOC 30523
ИЗОЛОК 30530; UMS5-ASF / 30
ИЗОЛОК 30897; UMS8-ASA / BV300x230x15
ИЗОЛОК 73134
ИЗОЛОК СТЗЕ M24x1,5×100-NR.72150
ИЗОЛОК СТЗЕ NTS100 / 30-NR.51003-2
ИЗОЛОК USM8-ASF40
ISOLOC NT Тип 80/30
ИЗОЛОК M16 * 150
ИЗОЛОК UMS5-ASF / 3050 + 25 LXBXH: 110X110X83
ИЗОЛОК UMS8-ASF / 25 LXBXH: 150X144X84
ISOLOC UMS M16X150
ИЗОЛОЧНЫЙ ВИНТ / 73134
ИЗОЛОК ПЛИТА / 13010 IPL30 100X50X15 ROT OP
ISOLOC UMS 18 кд
ISOLOC UMS 8 KDSA
ИЗОЛОК UMS8-DSF / 25 ИЗОЛОК
ИЗОЛОК M16X300 ИЗОЛОК
ИЗОЛОК РОНКАП1 ИЗОЛОК
ИЗОЛОК ИЗОЛОК НТС150 / 25 51509
ИЗОЛОК ИЗОЛОК НТС120-300 70142
Изолок NIVELLIERSCHR.M16X100 ВЕРС.V13A, АРТ-NR: 70030
Изолок НТС 120 / 20-2 ВЕРСИЯ V13A, ART-NR: 5120202
Изолок 30523
Изолок 30530; UMS5-ASF / 30
Изолок 30897; UMS8-ASA / BV300x230x15
Изолок 73134
Изолок СТЗЭ М24х1,5х100-НР.72150
Изолок STZE NTS100 / 30-NR.51003-2
Изолок NT Тип 80/30
Изолок M16 * 150
Изолок UMS5-ASF / 3050 + 25 LXBXH: 110X110X83
Изолок UMS8-ASF / 25 LXBXH: 150X144X84
Изолок UMS M16X150
Изолок ВИНТ / 73134
Изолок ПЛИТА / 13010 IPL30 100X50X15 ROT OP
Изолок UMS 18 KDS
Изолок UMS 8 KDSA
Изолок NT 160 / 17,41605
Изолок NTR 1525 / 17,42005
Изолок NT 120 / 17,41205
Изолок M30x1,5×200мм, 71064
Изолок 50802 + 70562, M20x1,5×150 мм
Изолок USM8-ASF40
Изолок 51202 + 70144, M20x1,5×200 мм
Изолок 71042 Тип NT
Изолок 41003 Тип: NT 100/30
Изолок Изобразительное искусство.-Nr.72150
Изолок Art.-Nr.51003-2
Изолок UMS5-ASF / 40 (патент)
Изолок UMS8-ASF / 40 (патент)
Изолок Art-Nr : 72150
Изолок Art-Nr : 51003-2
Изолок № 51202
Изолок № 70030
Изолок Артикул 70562 Тип NTS 50 bis NTS100 M20 * 1,5 * 150 мм
Изолок Изобразительное искусство.-Nr.50802 Тип NTS 80/20 d1 = 80 мм h2 = 32 мм
Изолок UMS5-ASF / 30
Изолок UMS-5-KDS нужен даташит
Изолок УМС-8-КДС
Изолок 71042 M20x1,5×150
Изолок 41003 NT100 / 30 NT — ВЫРАВНИВАЮЩИЙ ДИСК
Изолок Подушка клиновая PKA2-6 schwarz
Изолок Выравнивающие элементы БНВ215 / 4
Изолок Подушка клиновая PKA1 / 54-6 schwarz
Изолок Выравнивающие элементы БНВ 115/0
Изолок NTR-1525 / IPL17 / 42005
Изолок NT-120 / IPL17
Изолок 71064 / Ф.NIVELLIERTELLER NT U. NTE
Изолок NT-160 / IPL17 / 41605
Изолок 51209
Изолок 70030
Изолок 51004
Изолок Машинная установка UMS5-ASF / 30 NR.30530-ISOLOC
Изолок СТЕЛЛФУ? НЦ 80/17 М20х1,5х150-ИЗОЛОК
Изолок 41003 NT100 / 30
Isoloc Art.-Nr.70562 Тип. NTS 50 — NTS100 M20 * 1,5 * 150 мм 连杆
Isoloc Art.-Nr.50802 Типовой NTS 80/20 d1 = 80 мм h2 = 32 мм 支撑 头
Изолок UMS5-ASF / 30 垫铁
Изолок М16 x 150 мм Nr: 73132 脚垫 柱
Изолок НТ 80/30 №: 40803 脚垫
Изолок НТ 100/30 №: 41003 脚垫
Изолок USM8-ASF40 磁铁
Shanghai HangOu Механическое и электрическое оборудование
Equipment Co., Ltd. была основана в 2006 году и специализируется на
импорт запасных частей к механическому и электрическому оборудованию, инструментов и
другие товары и бренды по всему миру. Эти продукты широко используются в
металлургия, чугун и сталь, нефтехимия, энергетика, авиакосмическая промышленность, контейнеровоз
терминалы, автоматизация, водное хозяйство, производство бумаги, электростанция, текстиль,
литье под давлением, резина, лечение, упаковка для пищевых продуктов и другие области.
Наши основные продуктовые линейки включают: датчики, двигатели, переключатели, электронные весы,
энкодеры, контроллеры, анализаторы, оптические волокна, решетки, клапаны, предупреждения
фары, кабель, буксирный трос, распределитель, коробка передач, реле, буферы и импортные
аппаратное обеспечение.Шанхайская компания по механическому и электрическому оборудованию HangOu, ООО
Адрес: комната 905, корпус № 1, центр Госон, переулок 388, Чжунцзян.
Дорога. Шанхай, Китай.
Тел. 0086-21-51085161
0086-21-523
Факс : 021-51870910
021-523
Электронная почта : [email protected]
[email protected]
Почтовый индекс : 200062
Сайт : www.shhangou.com
www.shhangou.com.cn
AHS, SWR — победа над университетским бейсболом — Спорт — The Courier-Tribune
УИНСТОН-СЕЙЛИМ — Брэди Райт отказался от пары ударов, чтобы начать игру, а затем удалил 21 участника подряд, когда бейсбольная команда университетской команды средней школы Эшборо зафиксировала победу со счетом 9-0 на конференции Mid-Piedmont Conference над North Forsyth Tuesday.
УИНСТОН-СЕЙЛИМ — Брэди Райт отказался от пары ударов, чтобы начать игру, а затем удалил 21 участника подряд, когда бейсбольная команда университетской команды средней школы Эшборо зафиксировала победу со счетом 9-0 на конференции Mid-Piedmont Conference над North Forsyth Tuesday.
Райт раздул 13, когда «Голубые кометы» улучшились до 4-1 в игре лиги и 6-2 в целом.
У Кэма Рида было три попадания и один ИКР, у Остина Карри было два попадания, а у Райта было два попадания и три ИКР.
Аарон Гарнер сделал хоумран для AHS, который записал 12 хитов в конкурсе.
SWR бланков WG
АШЕБОРО — Коди Райт бросил шесть иннингов без локаута, а Джексон Снайдер разбил седьмой по счету седьмой по счету, когда Юго-западная средняя школа Рэндолфа одержала победу со счетом 4: 0 над Западным Гилфордом во вторник.
Райт отказался от двух ударов и выиграл шесть. Cougars забили три незаработанных пробега во втором иннинге, поскольку WG совершила четыре ошибки. Рис Дэниел шел пешком и в конечном итоге забил гол на земле Лэндона Дэвиса в финальном заезде SWR.
Пумы улучшились до 4-0 в игре лиги и 5-4 в целом.
ER закрывает THS в PAC
RAMSEUR — Бейсбольная команда университетской средней школы Восточного Рэндольфа обыграла Тринити 2: 0 во вторник на открытии Пьемонтской спортивной конференции на поле Грэди Лоусон.
Д.Дж. Лакстон выложил полную игру, не допустив четырех попаданий, восьми аутов и двух прогулок.
В таблице Коннор Шипли пошел 2-к-4 с RBI, в то время как Истон Джонс и Лакстон оба получили 2-к-3. Tre Moser пошел 1 из 3 с RBI.
Дикие коты (1-0, 5-2) отправляются в Провиденс-Гроув в пятницу.
В игре «Джейви» Восточный Рэндольф одержал победу над «Тринити» со счетом 5: 1. Колин МакДэниел разбил 5 1/3 подач на победу, выбив семь. Макдэниел и Чейз Маккаллум нанесли по два хита за Wildcats.
В понедельник «ЕР» проиграл Грэму со счетом 13: 0. Лэндон Смит одержал победу, отказавшись от одного удара в трех подач. У Майкла МакГи было два попадания, а у Смита было два попадания в ER (2-3).
SWR зачищает WG в MPC
GREENSBORO — Командная команда университетского софтбола Southwestern Randolph High School победила Western Guilford со счетом 11-1 в шести подач во вторник в игре MPC.
Джулия Брэди и Кэти Макнил объединились для победы.
В таблице Маккенна Гувер вышла 3-к-3 с тройным, двумя двойными и тремя ИКР, в то время как у Эшли Флойд, Саманта Уильямс, Саманта Лаграма и Макнил было по два хита.
Пумы (2-0, 3-3) сегодня принимают Рэндлмана.
В игре «джайви» SWR выиграла со счетом 21-0. Мэри Эдкинс подала ни одного нападающего, а Шайна Риттер сделала два хита.
RHS побеждает WHS в PAC
ТРИНИТИ — Средняя школа Рэндлмана открыла университетский бейсбольный сезон PAC с победой 8: 3 над Уитмором во вторник вечером.
Таннер Раут заработал победу с пятью беспроигрышными подачами. Он закончил отдавать всего два удара в 5 2/3 подач, когда RHS улучшился до 1-0 и 5-1. Тайлер Адамс закончил игру на насыпи. У Бейли Уэлча было два хита с тройным и три ИКР, а у Калеба Вебстера было два хита с двойным и один ИКР.
PG no-hit HPA
HIGH POINT — Коннер Паттерсон подал пять безуспешных подач, а Райн Осборн и Остин Амброуз также бросили мяч без попадания, когда средняя школа Провиденс-Гроув заработала университетскую бейсбольную победу со счетом 13: 0 над High Point Andrews в Пьемонтская атлетическая конференция.
Паттерсон выбил седьмую победу. У Сета Хардина и Гейджа Уайта было по два хита, а у Ника Нельсона было по два хита и два ИКР для «Патриотов» (1-0, 4-0).
AHS, PG играют вничью
ГРИНСБОРО — Команды средней школы Эшборо и Провиденс-Гроув сыграли вничью во вторник в трехматчевом матче в загородном клубе Forest Oaks.
«Голубые кометы» и «Патриоты» выпустили по 179 ружей, а Джордан-Мэтьюз выстрелил из 221.
Джастин Эммонс из PG стал медалистом, показав 1 из 35.Товарищ по команде Тайлер Бут забил 43, Блейк Холл 49 и Броди Фоглман 52.
У AHS, Коби Маркхэм и Ник Конкутелли сделали карты по 43, Алекс Эллиотт — 44 и Дилан Хоффман — 49.
Фейт открывает победу
RAMSEUR — Женская футбольная команда школы Faith Christian School открыла свой сезон во вторник, победив Берейана со счетом 6: 1.
Эллисон Стаут, Эшли Авелино и Кайла Лэнгли забили по два гола, в то время как Ханна Бертран хорошо сыграла в защите для Фейт.
SWRMS превосходит UMS 8-0
Футбольная команда Юго-Западной средней школы Рэндольфа обыграла Uwharrie 8-0.
Хосе Перес забил по два гола, а Фернандо Кастро, Хайро Хаймс, Сет Бэгли, Йодани Куадрос, Че Перес и Кендон Ганн добавили по одному голу. Джеймс отдал результативную передачу.
Вратарь Дэниел МакНил имел два сэйва после уверенного лидерства Хайме Бернала, Кэмерона Эбанкса, Эдди Бернала, Кевина Мора и Джона Агирре.
A-TMS побеждает SWRMS
Арчдейл-Тринити победил Юго-Западный Рэндольф 13-4 в бейсболе в средней школе.
Джонатан Уивер выиграл питчер, а Кэмерон Мозер спас сэйв.
В таблице Уивер пошел 2-к-3 с двумя забитыми трассами и двумя ИКР, в то время как Мэтью Оуэнс пошел 2-к-3 с тремя забитыми трассами. Коннор Кокс пошел 2-к-4 с тремя ИКР, а Ник Хало пошел 2-к-4 с двумя забитыми пробегами.
Бейсбольная команда Faith открылась победами
FAYETTEVILLE — Бейсбольная команда университетской школы Faith Christian School открыла сезон победой со счетом 14-1 над Берианом Кристианом во вторник.
Спенсер Кидд подал полную игру с шестью иннингами с семью аутами.
У Дрю Мэтисона, Кидда и Джереми Вильянуэвы было по два хита, у Спенсера было три ИКР, а у Вилланеува и Ника Шона — по два.
Формируется взрослая бейсбольная команда
Бейсбольный клуб CNC спонсирует взрослую бейсбольную команду для всех, кто старше 18 лет. Этой командой будет Randolph Nationals, которая будет играть в NABA в Гринсборо, но тренировки будут проходить в Ramseur на стадионе Craven.
Стоимость 335 долларов. Место ограничено. Чтобы зарегистрироваться или получить дополнительную информацию, звоните (336) 302-6052 или (336) 601-4531.
SAMS проигрывает NAMS
АШЕБОРО — Скотт Вильяграма забил за 2 минуты до конца второго овертайма, что позволило средней школе Южного Эшборо одержать победу над Норт Эшборо во вторник со счетом 1: 0. Благодаря этой победе SAMS стал 6: 0 и занял первое место в лиге средней школы округа Рэндольф.
Дэвид Кальдерон сделал результативную передачу, и Томас Гауф был в воротах. Алекс Венчес, Монтгомери Муди и Брэнсон Адамс вели оборону, в то время как Алвис Адамс и Уилл Клаузер также вели сильную игру.
SWR команды по легкой атлетике заняли третье место
ЛЕКСИНГТОН — Обе команды по легкой атлетике Юго-Западного Рэндольфа заняли третье место во вторник на соревнованиях по квадроциклам Mid-Piedmont Conference в Централ Дэвидсон.
Хозяева выиграли встречу мальчиков с 110 очками, на втором месте Южный Гилфорд (88), за ним идут Кугарз (39) и Вестерн Гилфорд (35).
В индивидуальном порядке Картер Хаффман занял первое место в беге на 400 метров за 51,5 секунды и 200 (23,8 секунды), в то время как Тайлер Харрис возглавил прыжок в высоту с прыжком на 5 футов 10 дюймов.
В соревнованиях девушек победила CD с 95,5 очками, за ней следуют SG (74), SWR (44,5) и WG (43).
Сара Бидлеспахер из группы Cougars стала первой в гонке 800 (3:01).
Падение команды по лакроссу AHS
АШЕБОРО — Команда по лакроссу средней школы Эшборо проиграла Рэгсдейлу на стадионе Ли Дж. Стоуна решением судей 15: 1.
Остин Пейдж забил за «Голубые кометы» в первой четверти, в то время как Март Фаулер, Итан Чисхолм и Джон Роберсон сыграли хорошо. AHS составляет 0-5.
Победа команды по софтболу WHS
РЭНДЛМЕН — Средняя школа Уитмора записала 5: 4 победы университетской команды по софтболу над Рэндлманом в действии во вторник на Атлетической конференции Пьемонта.
Уорриорз вели 5: 0, прежде чем Тигры увидели, что ралли совсем не закончилось. Морган Хало, одержавший победу на насыпи, дважды попал в игру WHS. У Дэйви Альбертсона и Пэйтона Минса также было по два хита за WHS (1-0, 6-1).
Кейтлинн Нортон, Беван Гувер и Хейли Кокс получили по два попадания в правую сторону. У Чарли Батлера было два попадания, а у Меган Коттрелл было три ИКР для правой руки (0-1, 3-4).
PG зачистки HPA
CLIMAX — Средняя школа Провиденс-Гроув перешла к паре университетских софтбольных побед над High Point Andrews в действии PAC, выиграв 15-0 и 18-1 с каждой игрой, которая длилась всего три иннинга.
Мэделин Гвинн подала нулевой нападающий в первой игре и один нападающий во второй игре «Патриотов» (2-0, 4-1).
У Хейли Раут было два дабла в первой игре, а у Хизер Робб было дабл и три ИКР во второй игре.
ER превосходит Trinity
TRINITY — Средняя школа Восточного Рэндольфа объявила о победе университетского софтбола 8: 2 над Trinity в действии PAC.
Кейли Картер нанес удар 11 и сделал тройку нападающих. У Келлы Гоинс было три попадания, у Микаэлы Грин было два попадания, а у Эмили Самнер и Кеннеди Минтон были ИКР для скорой помощи (1-0, 1-5).
У Келли Вагнер было два попадания и один ИКР для THS (0-1, 3-2).
В мире ABSOLUT® все неразрезано — AMC Networks Inc.
IFC и ABSOLUT® Vodka объявляют о новом многостороннем спонсорстве, вдохновляющем кинематографистов на выражение своего инди-духа
Остин, Техас (12 марта 2008 г.) — IFC, первая и крупнейшая сеть, посвященная независимому кино и культуре, и ABSOLUT® VODKA, самый культовый в мире бренд спиртных напитков, объявили сегодня о новом многогранном спонсорском соглашении. уникальным образом интегрирует бренд ABSOLUT в эфирный и онлайн-контент IFC, а также в спонсируемые IFC кинофестивали и мероприятия, проводимые в течение года.Сделка включает эксклюзивное спонсорство нового фильма IFC в субботу вечером, получившего название «ABSOLUT Indies». Кроме того, ABSOLUT станет спонсором конкурса короткометражных фильмов ABSOLUT Indies Short Film Challenge на IFC.com, нового фестиваля и конкурса цифрового кино, который побуждает кинематографистов изобразить свое видение жизни «В АБСОЛЮТНОМ Мире».
Сделка еще больше расширяет сферу охвата недавно представленной водочной марки рекламной кампании «В АБСОЛЮТНОМ Мире», которая побуждает потребителей смело и оптимистично смотреть на мир.ABSOLUT является новейшим дополнением к растущему списку многоплатформенных партнерских отношений IFC и расширяет уникальную бизнес-модель сети, предлагая своим партнерам инновационные и нетрадиционные способы демонстрации своего бренда.
«ABSOLUT полностью соответствует тому, что представляет собой IFC, поскольку оба бренда открывают миру новые и свежие перспективы», — сказал Алан Кляйн, старший вице-президент IFC по партнерству и лицензированию. «Как и все наши партнеры, мы работаем с ABSOLUT над стратегической интеграцией их бренда интересным и нетрадиционным способом, который не только предоставит зрителям развлекательный контент как в эфире, так и в Интернете, но и привлечет внимание сообщества Создатели фильмов Media Lab, дающие этим неизведанным артистам возможность продемонстрировать свой талант.«
«ABSOLUT действительно воплощает творческий потенциал независимой киноиндустрии. Являясь официальным спонсором инновационной серии фильмов Live Earth в США, а теперь являясь партнером IFC, мы еще больше осознаем нашу приверженность искусству независимого кино», — сказал Ян Кристал. Бренд-директор, Водка АБСОЛЮТ. «Наше спонсорство IFC является настоящим сотрудничеством и предлагает еще одну платформу для нашей кампании« В АБСОЛЮТНОМ мире », предоставляя форум, чтобы побудить стремящихся к самовыражению режиссеров».
ПРИСУТСТВИЕ В ЭФИРЕ
Начиная с этого месяца, ABSOLUT интегрируется в эфирное программирование IFC с запуском фильма «ABSOLUT Indies».Каждую субботу вечером на IFC «ABSOLUT Indies» представляет двойной полнометражный фильм из двух независимых фильмов, которые необходимо посмотреть — полностью без цензуры, без вырезок и без рекламных роликов, как это и было задумано создателями. Цепочка фильма начинается с настраиваемого 45-секундного графического шлюза, созданного IFC, который задает тон для двойного фильма «ABSOLUT Indies», демонстрируя отрывки из фильмов той ночи. Среди показанных фильмов из новой серии — The Cooler, Gosford Park, Bowling for Columbine, Elephant и Kinsey , а также другие.
Перед началом первого фильма пользовательский шлюз IFC перейдет на фестиваль короткометражных фильмов ABSOLUT Indies. Фестиваль короткометражных фильмов представит постоянно меняющуюся коллекцию короткометражек, отражающих разнообразие и идеализм инди-духа. Короткометражные фильмы, представленные на Фестивале, будут включать фильмы из серии фильмов «Живая Земля» (liveearth.absolut.com), коллекцию образовательных и увлекательных короткометражных фильмов о последствиях глобального потепления.
ОНЛАЙН-ПРИСУТСТВИЕ
«ABSOLUT Indies» также станет стартовой площадкой для IFC.com’s ABSOLUT Indies Short Film Challenge, постоянный фестиваль и конкурс цифрового кино, проводимый Media Lab Studios, новой независимой киностудией IFC, специализирующейся на веб-контенте и эфирном контенте. Студия Media Lab попросит начинающих кинематографистов прислать материалы, отражающие их видение фильма «В АБСОЛЮТНОМ Мире» с использованием серии фильмов Live Earth в качестве вдохновения. Отобранные фильмы будут выставлены на ABSOLUT.com, IFC.com, а также на субботнем фестивале короткометражных фильмов ABSOLUT Indies, который будет транслироваться на IFC.
ПРИСУТСТВИЕ НА ФЕСТИВАЛЕ
Помимо интеграции в онлайн и эфир, ABSOLUT будет активно присутствовать на кинофестивалях и мероприятиях, спонсируемых IFC.Фестиваль SXSW 2008 будет первым из этих фестивалей, при этом ABSOLUT представит IFC Crossroads Party 2008 11 марта 2008 года в округе в Остине, штат Техас. Дополнительное присутствие на фестивалях для ABSOLUT будет включать, среди прочего, кинофестиваль Tribeca и кинофестиваль в Лос-Анджелесе.
О IFC
IFC (The Independent Film Channel) — первая и самая большая сеть, посвященная независимому кино и связанным с ним программам, 24 часа в сутки, без вырезок и без цензуры. Действуя под лозунгом «всегда, неразрезанный», IFC представляет полнометражные фильмы, «культовую классику», заставляющие задуматься оригинальные документальные фильмы, такие как У двери дома смерти и Этот фильм еще не получил рейтинга , короткометражки, мультсериалы. , эксклюзивные веб-сериалы и самая обширная библиотека независимых фильмов на телевидении.Сеть также предлагает одни из самых новаторских и острых оригинальных сериалов на телевидении, в том числе скетч-комедию The Whitest Kids U ‘Know . Обеспечивая голос независимой мысли, которой нет больше нигде на телевидении, IFC расширяет аудиторию независимых фильмов и поддерживает независимое киносообщество посредством эксклюзивных прямых трансляций известных кинематографических событий, таких как Independent Spirit Awards. Предложение сети по запросу, IFC Free, дает зрителям возможность смотреть премьеры всех оригинальных сериалов IFC в HD и по запросу, прежде чем они будут транслироваться в линейной сети.Сеть Rainbow Entertainment Services, дочерние бренды IFC включают AMC и WE tv.
О компании Absolut Spirits, Inc.
Absolut Spirits Company, Inc., штаб-квартира которой находится в Нью-Йорке, является дочерней компанией V&S Group и работает под эгидой V&S Absolut Spirits в Стокгольме, которая производит и продает ABSOLUT® VODKA , ABSOLUT CUT TM , Level TM Vodka, FRIS® Vodka, Plymouth TM Gin и Cruzan® Rum.
