Что такое защитное зануление. Как работает защитное зануление. Где применяется зануление. В чем отличие зануления от заземления. Какие требования предъявляются к занулению.
Что такое защитное зануление
Защитное зануление — это преднамеренное электрическое соединение открытых проводящих частей электроустановок с глухозаземленной нейтральной точкой источника питания в сетях трехфазного тока, с глухозаземленным выводом источника однофазного тока, с заземленной точкой источника в сетях постоянного тока.
Основное назначение защитного зануления — обеспечение электробезопасности путем быстрого отключения поврежденного участка цепи при замыкании на корпус электрооборудования.
Принцип действия защитного зануления
Как работает защитное зануление? Принцип действия основан на преобразовании замыкания на корпус в однофазное короткое замыкание. При этом происходит:
- Возникновение большого тока короткого замыкания
- Срабатывание максимальной токовой защиты (автомата, предохранителя)
- Быстрое отключение поврежденного участка от сети
Время срабатывания защиты при занулении не должно превышать 0,4 секунды. Это обеспечивает безопасность человека при случайном прикосновении к корпусу электроустановки, оказавшемуся под напряжением.
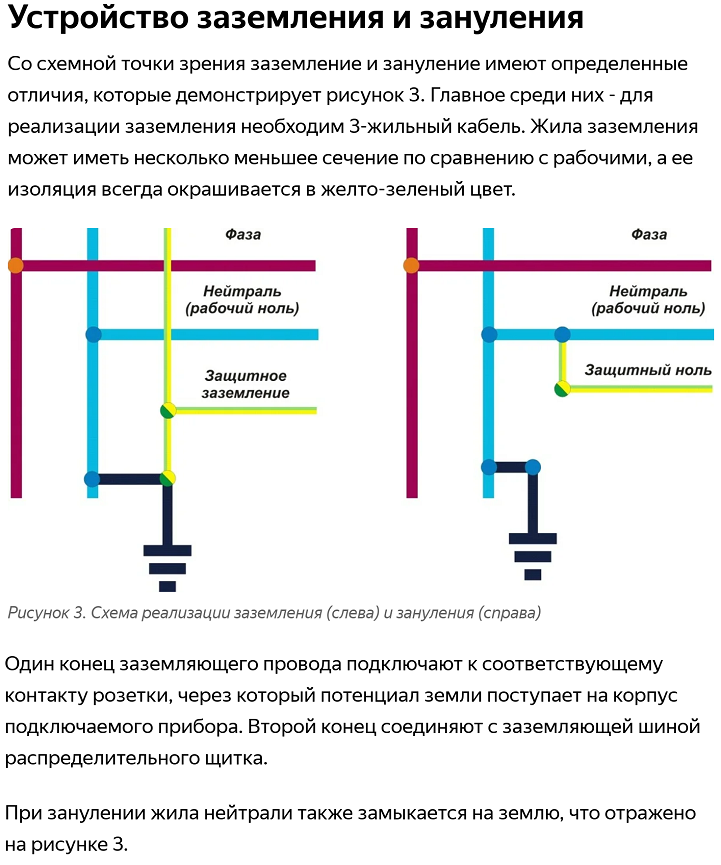
Области применения защитного зануления
Где применяется зануление для защиты от поражения электрическим током? Основные области применения:
- Электроустановки напряжением до 1000 В в трехфазных сетях с глухозаземленной нейтралью
- Однофазные сети переменного тока с глухозаземленным выводом
- Сети постоянного тока с глухозаземленной средней точкой
Зануление широко используется в промышленных и бытовых электроустановках для защиты персонала и населения от поражения электрическим током.
Отличия зануления от заземления
В чем основные отличия защитного зануления от защитного заземления? Сравним эти два вида защиты:
| Параметр | Зануление | Заземление |
|---|---|---|
| Принцип действия | Преобразование замыкания на корпус в КЗ | Снижение напряжения прикосновения |
| Подключение | К глухозаземленной нейтрали | К заземляющему устройству |
| Область применения | Сети до 1 кВ с глухозаземленной нейтралью | Сети свыше 1 кВ, сети до 1 кВ с изолированной нейтралью |
Таким образом, зануление и заземление имеют разные принципы действия и области применения, но выполняют общую функцию защиты от поражения электрическим током.

Требования к выполнению защитного зануления
Какие основные требования предъявляются к системе защитного зануления? Рассмотрим ключевые моменты:
- Сопротивление петли «фаза-ноль» должно обеспечивать ток КЗ, достаточный для срабатывания защиты за нормируемое время
- Нулевой защитный и нулевой рабочий проводники должны быть разделены на вводе в электроустановку
- Запрещается использовать в качестве нулевого защитного проводника металлические конструкции здания, трубопроводы и т.п.
- Площадь сечения нулевого защитного проводника должна быть не менее площади сечения фазного проводника
Соблюдение этих и других требований обеспечивает надежную работу системы зануления и защиту от поражения электрическим током.
Достоинства и недостатки защитного зануления
Как и любая система защиты, зануление имеет свои плюсы и минусы. Рассмотрим основные преимущества и недостатки зануления:
Достоинства:
- Простота устройства и надежность срабатывания
- Быстрое отключение поврежденного участка
- Возможность применения в разветвленных сетях
Недостатки:
- Возможность повышения потенциала на корпусах электрооборудования при обрыве нулевого провода
- Опасность поражения током до момента срабатывания защиты
- Необходимость в источнике питания с глухозаземленной нейтралью
Несмотря на некоторые недостатки, защитное зануление остается одной из основных и эффективных мер защиты от поражения электрическим током в сетях до 1000 В.
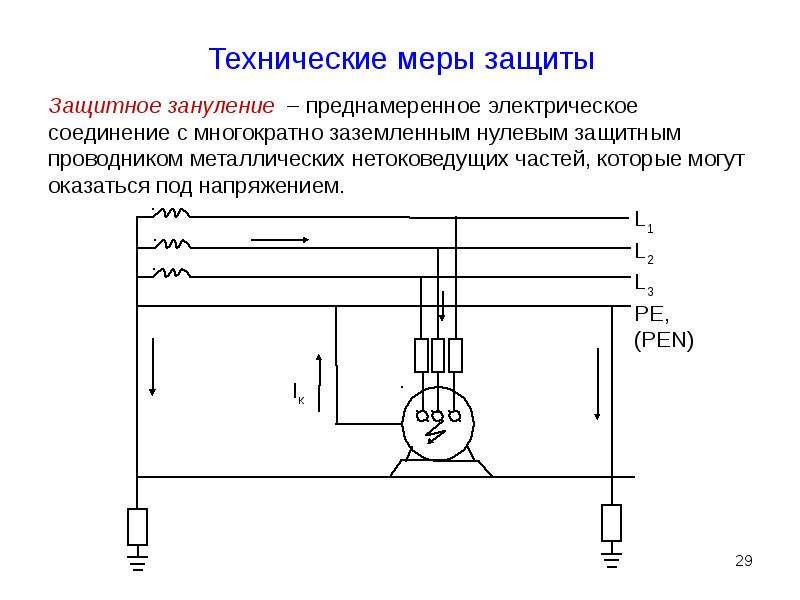
Проверка эффективности защитного зануления
Как проверить правильность и эффективность выполнения зануления? Основные методы проверки включают:
- Измерение сопротивления петли «фаза-ноль»
- Проверка целостности нулевого защитного проводника
- Проверка времени срабатывания защитных устройств
- Измерение напряжения между корпусом электроустановки и землей
Регулярная проверка эффективности зануления позволяет своевременно выявлять и устранять возможные нарушения в системе защиты.
Заключение
Защитное зануление является важной и эффективной мерой защиты от поражения электрическим током в электроустановках до 1000 В с глухозаземленной нейтралью. Правильное выполнение и регулярная проверка системы зануления позволяют обеспечить высокий уровень электробезопасности в промышленных и бытовых электроустановках.
Что такое защитное зануление — схема и принцип работы
Зануление представляет собой специальное подключение открытых металлических частей электрооборудования (электроустановок) к нейтрали. Это относится к металлическим не токоведущим частям оборудования, которые в нормальном (рабочем) режиме не находятся (и не должны находиться) под напряжением. Нейтраль, с которой происходит соединение, должна быть глухо заземлена.
В трёхфазных электрических сетях – это нейтраль генератора или силового трансформатора, в однофазной сети – это глухозаземлённый вывод источника питания.
Нулевым защитным проводником (не путать с нулевым рабочим проводником) является такой проводник, который соединяет металлические занулённые части электрооборудования с глухозаземлённой нейтралью, идущей от генератора или питающего силового трансформатора.
Цель защитного зануления – обеспечить электрическую безопасность в случае короткого замыкания на металлический корпус электрооборудования или электроустановки.
Принцип зануления
Защитное зануление работает следующим образом. Если при поданном электрическом питании происходит попадание фазы (случайное попадание или пробой изоляции фазного проводника) на металлический корпус с занулением, то возникает короткое замыкание, резко увеличивается значение электрического тока и срабатывает аппарат защиты (автоматический выключатель) или перегорает плавкая вставка защитного предохранителя, тем самым обесточивая электрооборудование или электроустановку.
Сопротивление защитного нулевого проводника должно быть очень низким. Это необходимо для того, чтобы обеспечить уровень тока короткого замыкания, достаточный для действия защиты. Т.е. значение тока к.з. должно быть достаточным для того, чтобы сработал защитный аппарат.
Если электрооборудование просто заземлить, то, например, в случае пробоя фазы на корпус ток короткого замыкания может быть недостаточным для того, чтобы сработал автоматический выключатель или перегорела плавкая вставка предохранителя.
Ввиду того, что нейтраль заземлена на генераторе или трансформаторе, благодаря защитному занулению обеспечивается достаточно малое напряжение прикосновения на корпусе. Т.е. защитное зануление можно считать своего рода разновидностью заземления.
Видео — Зануление и заземление — в чем разница?
Схемы защитного зануления
Существует несколько схем, по которым выполняется защитное зануление.
Система TN-C
Достаточно простая система, по которой выполняется защитное зануление. В ней нулевой проводник N и защитный проводник PE по всей длине объединены в один общий проводник PEN. Для реализации защитного зануления по системе TN-C необходимо соблюдать очень высокие требования к системе уравнивания потенциалов, а также к размеру поперечного сечения совмещённого PEN-проводника.
Зануление по системе TN-C применяется в трёхфазных электрических сетях, а в однофазных сетях такое зануление категорически запрещено.
Система TN-C-S
Данная система представляет собой соединённые N и PE проводники в части сети, начиная от электрического источника питания. По данной системе допускается зануление электрооборудования в однофазных сетях.
Область применения защитного зануления
Защитное зануление применяется в однофазных и трёхфазных сетях переменного тока до 1кВ. Сеть должна быть с глухозаземлённой нейтралью.
Проверка эффективности защитного зануления
Суть защитного зануления заключается в том, чтобы в случае короткого замыкания фазы на корпус электрооборудования произошло автоматическое отключение повреждённого участка цепи. Для того чтобы проверить на сколько эффективно выполнено защитное зануление, необходимо измерить сопротивление петли фаза-ноль в самой удалённой от источника питания точке. Это позволит определить, сработает ли аппарат защиты в случае однофазного к.з. на корпус.
Сопротивление петли фаза-ноль измеряется при помощи специальных измерительных приборов. Приборы для измерения петли фаза-ноль имеют два щупа. При измерении один щуп подключается к действующей фазе, а второй – к занулённой части электрооборудования.
В результате замера выясняется значение сопротивления петли фаза-ноль. Зная величину измеренного сопротивления и значение питающего напряжения, по формуле закона Ома для участка цепи можно рассчитать ток однофазного короткого замыкания, расчётное значение которого должно быть больше (или равно) тока срабатывания защитного устройства.
Допустим, для защиты цепи от токовых перегрузок и от коротких замыканий установлен автоматический выключатель, ток мгновенного срабатывания которого равен 100А. Измеренное значение сопротивления петли фаза-ноль равно 2 Ом, фазное напряжение в сети равно стандартному значению 220В.
Рассчитываем значение тока однофазного короткого замыкания. По закону Ома I = U/R = 220В/2Ом = 110А.
Т.к. расчётный ток к.з. больше чем ток мгновенного срабатывания (отсечки) автоматического выключателя, то защитное зануление будет эффективным. Если бы расчетный ток к.з. получился меньше тока мгновенного срабатывания автомата, то для эффективности защитного зануления пришлось бы или менять автоматический выключатель на устройство с меньшим током срабатывания, или искать решение по уменьшению сопротивления петли фаза-ноль.
Очень часто в расчётах ток срабатывания автоматического выключателя умножается на так называемый коэффициент надёжности Кн или коэффициент запаса. Дело в том, что отсечка автомата не всегда соответствует указанному значению, т.е. может быть некоторая погрешность, для этого и вводится в расчёты указанный коэффициент. Для старых автоматов Кн может равняться, например, 1,25 или 1,4. Для новых современных автоматов он может быть равен 1,1. Это связано с тем, что новые аппараты защиты работают более точно.
Защитное зануление электроустановок, назначение, принцип действия, схема заземления
Автор Фома Бахтин На чтение 3 мин. Просмотров 6.9k. Опубликовано Обновлено
Зануление – это специально предусмотренное электрическое подключение открытых токопроводящих частей потребителей электроэнергии:
- к нейтральной точке генератора (трансформатора) в сетях трехфазного тока,
- к глухозаземленныму выводу сети однофазного напряжения,
- к заземленной точке источника постоянного тока.
Такое подключение выполняется в целях обеспечения электробезопасности человека.
Для обеспечения подключения незащищенных от прикосновения токопроводящих частей электропотребителей к нейтральной точке источника электроэнергии предусмотрено применение нулевого защитного проводника.
Нулевой защитный проводник (обозначается PE – проводник для системы TN – S) – токопроводящая цепь, соединяющая открытые токопроводящие поверхности и глухозаземленную нейтральную точку на источнике питания в трехфазных сетях или заземленный вывод однофазной сети, или заземленную среднюю точку источника постоянного тока.
Следует различать понятия нулевого защитного проводника и нулевого рабочего или PEN – проводника. Рабочий нулевой проводник (обозначается, как N – проводник для системы TN – S) – это провод в электропотребителях напряжением до 1 кВ, применяемый для обеспечения электропитания, который соединен с глухозаземленным нейтральным выводом на генераторе или трансформаторе в сетях трехфазного тока, либо с глухозаземленной точкой на источнике однофазного тока, либо с глухозаземленным выводом на источнике в сети постоянного тока.
На практике допускается применение совмещенного (обозначается, как PEN – проводник для системы TN– C) нулевого защитного и нулевого рабочего проводника. Его роль выполняет цепь в электропотребителях напряжением до 1 кВ, совмещающая нулевой защитный и нулевой рабочий проводник. Использование зануления требуется для осуществления защиты человека от воздействия электрического тока при его прикосновении к токоведущим поверхностям за счет быстрого снижения напряжения на корпусе электропотребителя относительно земли, сопровождаемого быстрым отключением электроустановки от питающей сети
Зануление электроустановок
Обязательное защитное зануление необходимо выполнять на:
- электроустановках напряжением питания до 1 кВ (трехфазные сети переменного тока, имеющие заземленную нейтраль). Чаще всего это сети переменного тока напряжением 380/220, реже – 660/380 В;
- электроустановках напряжением питания до 1 кВ (однофазные сети переменного тока, имеющие заземленный вывод). Напряжение, как правило – 220 вольт;
- электроустановках постоянного тока с напряжением до 1 кВ в сетях, имеющих заземленную среднюю точку источника.
Физически зануление осуществляется специальным проводом, имеющим надежный электрический контакт с открытыми токоведущими поверхностями электропотребителей.
Принцип действия защитного зануления
В случае замыкания фазного провода на корпус электропотребителя, имеющий зануление, возникает электрическая цепь тока с коротким замыканием (происходит замыкание фазного и нулевого защитного проводников). Появление тока короткого замыкания приводит к срабатыванию токовой защиты. Как следствие, происходит отключение такой электроустановки от электропитающей сети. Попутно, до наступления срабатывания автоматической токовой защиты обеспечивается снижение напряжения на поврежденном корпусе относительно земли. Это связано с наличием защитного действия повторного заземления на нулевом защитном проводнике и перераспределения напряжений в сети вследствие протекания тока в короткозамкнутой цепи.
Принципиальная схема зануления
Рассмотрим схему заземления:
Мы искренне надеемся, что наша статья помогла вам понять определение заземления, его назначение и принцип действия.
Зануление и заземление. В чем разница между ними?
Рабочее и защитное заземление
Зануление и заземление
Защитное зануление. Работа и устройство. Применение и особенности
Во всем мире используется защита, основанная на соединении нетоковедущих проводящих частей оборудования с землей и заземленной нейтралью источника. В России эта система называется защитное зануление. Защитное действие этой системы основано на принципе достижения нулевого напряжения на корпусе прибора, за счет многократного заземления и соединения нетоковедущих частей с нейтралью источника.
Несмотря на ряд недостатков, зануление продолжает служить основным электрозащитным средством во всем мире. Открытые части установки соединяют отдельным нулевым защитным проводником.
Зануление – соединение металлических частей электрооборудования с нулевым защитным проводом. Зануление служит мерой защиты от случайного попадания под напряжение.
Защитное зануление рассчитано на случай короткого замыкания. Распределение нагрузки на предприятии осуществляется равномерно, нулевой провод исполняет функции защиты. Ноль соединяется с корпусом электродвигателя. Когда происходит короткое замыкание, то возникает напряжение на корпусе электродвигателя.
При этом происходит срабатывание автоматического выключателя. При применении заземляющей шины промышленные электроустановки соединяются.
Принцип действия
Замыкание случается при касании подключенного к напряжению фазного провода на корпус прибора, который соединен с нулем. Возникает большая сила тока, срабатывают аппараты защиты, отключающие питание неисправного прибора.
Время срабатывания защиты и отключения неисправной линии по правилам не должно быть более 0,4 секунды. Для зануления можно применить третью неиспользуемую жилу в кабеле для 1-фазной сети питания.
Фаза и ноль должны быть с небольшой величиной сопротивления. Только тогда аппарат защиты отключит напряжение в установленное время. Чтобы было хорошее зануление необходимо обеспечить качественные контакты соединений.
Защитное зануление дает возможность создать быстрое выключение от сети неисправного питания. Вероятность удара током человека практически исчезает. Зануление считается одним из видов заземления.
Порядок зануленияЗануление для защиты в доме начинается с нейтрали, соединенной с заземленной нейтралью трансформатора.
Нейтраль с 3-фазной линией приходит в здание дома в шкаф ввода. Далее, она разветвляется по щиткам на разных этажах. От нее используется рабочий ноль, образующий 1-фазное напряжение. Ноль имеет название рабочего, так как он применяется для работы.
Зануление для защиты создается отдельным нулем в щитке. Ноль соединен с заземленной нейтралью. Нужно знать, что в схеме соединения ноля с нейтралью не должно быть аппаратов коммутации (рубильников, автоматов).
Как известно в цепях трехфазного переменного напряжения обмотка трансформатора может соединяться в треугольник и в звезду. Рассмотрим звезду. Звезда имеет нулевую точку, или нейтраль. Это та точка, в которой сумма всех трех напряжений сети будет равна нулю.
При такой схеме трансформатора могут быть две возможные схемы. Схема с изолированной нейтралью показана на нашем рисунке. Такая схема обычно используется при работе трехфазных систем, а также однофазных систем, но используется именно изолированная нейтраль.
Также есть еще глухозаземленная нейтраль.
Нейтраль трансформатора соединяется с землей. Эта схема может быть использована не только для работы в трехфазной или однофазной системе, но также для защитного зануления.
Схема состоит из переменного источника напряжения 220 В, его датчика напряжения, нагрузки, сопротивления, которое в нормальном состоянии отключено. Но когда возникает пробой изоляции при выполнении неправильного монтажа, на корпусе появляется напряжение. Измерим напряжение на нагрузке относительно земли. Рассмотрим схему на базе однофазного источника напряжения.
Мы заземляем нулевую точку. Делаем имитацию пробоя изоляции на корпус. На корпусе установилось напряжение, которое будет равно напряжению источника. При таком состоянии если прикоснуться к корпусу, то человека ударит током. Как избежать этой ситуации? Все очень просто. Используют схему защитного зануления, а именно, корпус соединяют с глухозаземленной нейтралью трансформатора. Напряжение на корпусе становится равным нулю.
Почему опасно защитное зануление в квартиреЕго используют для защиты людей и животных от поражения электрическим током, а также для срабатывания защитной аппаратуры в случае возникновения утечки тока на землю. Возникает вопрос: если мы используем глухозаземленную нейтраль, то можно соединить точку защитного заземления с нейтралью?
Этого делать нельзя. По правилам это запрещено. Если при выполнении монтажных работ будут перепутаны местами фаза и ноль, а мы поставим перемычку для соединения заземления с нейтралью, получим следующую неприятную ситуацию. При подключении устройства к сети, корпус оказывается под напряжением относительно земли. Как гласит ПУЭ использование нулевого рабочего проводника в качестве защитного зануления категорически запрещено.
Для защитного зануления отводится специальная шина, которая будет соединена с заземляющим устройством или с глухозаземленной нейтралью. Все заземляющие провода подключаются к этой шине параллельно. Поэтому, не нужно ставить перемычки. А перед тем, как реализовывать защитное заземление или зануление нужно ознакомиться с правилами.
Некоторые специалисты делают заземление приборов перемычкой клеммы ноля в розетке на контакт защиты.
Такой способ запрещен.На входе в квартиру устанавливают аппарат, служащий для подключения питания сети. Это может быть пакетный выключатель или автомат. Опасность самодельного заземления с помощью перемычки в том, что корпус устройства, подключенного к этой розетке, в случае повреждения изоляции нуля станет доступным напряжению фазы. А если оборвется провод нуля, то работа прибора прекратится. Возникнет ложная видимость провода, как обесточенного. Это опасно для жизни.
Такая розетка сделает много неприятностей, если в нее запитать стиральную машину. Если отгорит ноль, то стиральная машина может убить человека в случае прикосновения к ней.
Если человек принимает душ из электрического водонагревателя, а в это время нулевой провод в розетке отсоединится, то человека ударит током. Такое зануление очень опасно выполнять в квартире.
Применение зануленияПрименяется в электроустановках до 1 кВ в:
- Сетях постоянного тока со средней точкой заземления.
- 1-фазных сетях с заземленным выводом.
- 3-фазных сетях с заземленным нулем.
Защитное зануление служит для защиты от удара током. Если внутри электроприбора повредилась изоляция и корпус прибора оказался под током, то отреагирует защита и отключит сеть питания.
Образование тока КЗ возникает, если произошло замыкание нулевого и фазного провода на зануленный корпус. Для скорейшего отключения устройства применяют автоматы, предохранители, магнитные пускатели с защитой от перегрева, контакторы с реле.
Похожие темы:
Зануление — принцип действия и область применения
Помимо заземления, обеспечивающего защиту от короткого замыкания, имеется также процедура зануления, которая кардинально отличается. Это специализированное соединение электрических частей открытого типа, имеющих отношение к силовым установкам, рабочим состоянием для которых является отсутствие напряжения, с глухозаземленной нейтралью генерирующих или преобразующих устройств, что особенно характерно для трехфазного тока. Также возможно сопряжение с глухим заземлением на выходе из источника с одной фазой или такой же точкой в сетях с постоянным током. Единственной целью данного действия является обеспечение высшей степени безопасности рабочего персонала на потенциально опасных объектах.
Зануление, сооружаемое с целью защиты, часто является единственной мерой защиты на электрических установках мощность до 1 киловольта, но только при условии наличия в них глухозаземленной нейтрали.
Основной принцип работы
Зануление заключается в том, что при наличие фазового перехода между корпусом (пробивается фаза), то происходит КЗ. В этом случае сила тока имеет огромные значения, что вызывает срабатывание защитных вспомогательных приборов. Неисправный прибор при этом быстро обесточивается. Если ПУЭ сработает без зануления, то тогда сети будет нанесён очень сильный ущерб. Просто автоматика не успеет выключить всё быстро. А так происходит выигрыш, иногда до 1 секунды. Этого достаточно, чтобы КЗ не сожгло всю проводку на объекте и в оборудовании.
Процедура может быть проведена только на базе специализированных проводников, где жила имеет точное расчётное сопротивление. В однофазной сети нужно использовать трехжильные провода, чтобы добиться успеха. Также можно с этой целью использовать третий провод.
Чтобы вовремя отключить петлю типа «фаза-ноль» и снизить её сопротивление, нужно предварительно проектировать данную систему, иначе зануление потеряет свою эффективность.
Быстрое обесточивание линий электроснабжения не является единственной мерой. Нейтраль имеет глухое заземление, поэтому при касании корпуса человек может почувствовать только неприятное пощипывание. Это строго необходимая мера для всего промышленного оборудования. Зануление сильно отличается от заземления, эти два процесса нельзя сравнивать. Данная мера очень эффективна, известны случаи, когда люди выживали при касании неисправного оборудования, работающего от 10000 вольт.
Главные ошибки в организации
Некоторые люди, далекие от сферы электрификации, думают, что заземление нужно выносить на отдельный провод, который не сопряжен с нулевым проводником электрической сети, но это неверно. Они просто пренебрегают сопротивлением бесконечного длинного проводника, которые идёт от раздающей электрической установки потребителя к ближайшей КТП. Это неверно, ведь если сопротивление заземления будет ниже, чем у нулевой жилы, то тогда весь ток при КЗ пойдёт на корпус, а это приведёт к известным печальным последствиям. Опасный потенциал будет искать наиболее кратчайший и простейший путь для распространения. Поток электронов пойдёт туда, где сопротивление будет значительно меньше. Чем большее время будет обеспечено для срабатывания ПУЭ, тем лучше будет работать вся система. Некоторые владельцы собственного бизнеса даже раскошеливаются на проводники, состоящие из смеси серебра и меди, чтобы снизить сопротивление на порядок, а также гарантировать 100% срабатывание.
Сейчас многие владельцы производственных линий разбалованы современными УЗО. Они полностью списывают на эти устройства всю ответственность, что делать запрещено. Серьезность отношения к процедуре зануления была очень велика в советское время. Тогда вообще запрещалось делать без этой меры подключение любого промышленного оборудования по системе ТТ. Именно этим и поясняется столь малое количество несчастных случаев на производстве, связанных с электричеством. Когда всё возложено на УЗО, то это также хорошо работает, но шанс получить удар током всегда есть. Лучше делать двойную защиту.
Где купить товары для зануления
Все необходимые приспособления вы можете приобрести прямо сейчас в нашем интернет-магазине «ПрофЭлектро». Мы являемся признанными экспертами в сфере создания систем электрификации бытового и промышленного назначения. Поэтому наш ассортимент состоит далеко не только из розеток, выключателей и проводов. У нас также имеется множество сопряженного оборудования, инструментов и вспомогательных аксессуаров, необходимых профессиональным электрикам для ежедневной работы. Всё тщательно проверяется нашими специалистами на предмет соответствия заявленным характеристикам и наличие фабричного брака перед отправкой. Доставка возможна в любую точку России.
Защитное зануление, назначение и принцип действия.
Защитное заземление и зануление электроприборов.
В России используется система с глухозаземленной нейтралью, хотя в мире существуют и другие системы электроснабжения потребителей напряжением до 1000 В. В наших домах используется именно такая система.
Несмотря на сложное название, все довольно просто. В этой системе на подстанции нейтральная точка трансформатора соединена непосредственно с землей. В данном случае, главной мерой защиты от случайного воздействия напряжения является защитное зануление, то есть соединение корпуса и любой другой металлической части электроприбора с нейтралью трансформатора.
- Выше было отмечено, что в таких системах, нейтраль глухо заземлена, то, в принципе, защитное зануление является одной из разновидностей заземления.
Розетка с заземляющим контактом
У нас дома в каждой розетке есть заземляющий контакт (при правильно выполненной электропроводке). Именно с помощью него мы при включении соединяем корпус бытового электроприбора с нейтральной точкой трансформатора.
Защитное заземление работает следующим образом. По нормативным документам, допустимое время, за которое при коротком замыкании происходит отключение поврежденной линии, не более 0,4 секунд. Считается, что именно за это время человек, попавший под напряжение, может остаться в живых.
При защитном занулении происходит заметное снижение сопротивления цепи «фаза-ноль», чем обеспечивается достаточный уровень тока короткого замыкания для того, чтобы сработал аппарат защиты (автоматический выключатель или предохранитель) за время не более 0,4 секунд.
Защитное зануление.
Если зануление, которое в быту называют «заземлением», отсутствует, за счет высокого сопротивления, ток короткого замыкания может быть недостаточным для срабатывания защиты и появится опасность поражения человека напряжением при прикосновении к поврежденному бытовому прибору.
Защитное зануление должно выполняться в соответствие с требованиями Правил Устройства Электроустановок (ПУЭ). Обычно для этого может использоваться медный проводник сечением не менее 4 кв. мм, проложенный отдельно, либо третья жила провода.
Кроме этого, категорически запрещается заземлять бытовые электроприборы на отдельный контур заземления в сетях с глухозаземленной нейтралью, если контур заземления не связан с нейтральной точкой трансформатора. То есть, просто заземлив соответствующий контакт розетки на вбитый под окном металлический стержень.
Защитное зануление.
С этим же связаны последствия попыток «заземления» на систему водоснабжения или отопления квартиры. В этом случае, земля и дополнительный контур заземления (обычно самопальный) могут иметь гораздо большее сопротивление, чем защитный нулевой проводник, в следствие чего ток короткого замыкания будет достаточно низким.
В итоге, защитное зануление может сыграть большую роль в обеспечении электробезопасности вашей квартиры. Необходимо уделять большое внимание качеству и правильности его выполнения.
Что лучше заземление или зануление? Посмотреть наглядно в чём отличие и сравнить можно в этой статье. Так же вы узнаете, как можно сделать своими руками.
Назначение, принцип действия и расчет зануления
В соответствии с требованиями ГОСТ Р50571.3-94 «Электроустановки зданий» и «Правил устройства электроустановок» в электрических сетях типа TN-C-S для предотвращения электротравматизма при эксплуатации электрооборудования, конструктивные нетоковедущие металлические части которого оказались под напряжением вследствие замыкания тока на корпус, а также при других аварийных режимах сети, применяют зануление (рис.).
Рис. 10.6. Принципиальная схема зануления.
Физическая сущность зануления заключается в возникновении тока короткого замыкания между нулевым проводом и поврежденной фазой. Ток короткого замыкания может достигать сотен ампер -в результате плавкая вставка расплавляется или отключается тепловое реле и система отключается.
Нулевым защитным проводником называют проводник, соединяющий зануляемые части с глухозаземленной нейтральной точкой источника тока.
Основное требование безопасности к занулению заключается в уменьшении длительности отключения замыкания — оно должно быть не более долей секунды.
Так как время срабатывания плавких вставок предохранителей и тепловых расцепителей автоматов обратно пропорционально силе тока, то малое время срабатывания возможно при большой силе тока. Каждый отключающий аппарат имеет свою заводскую токов-ременную характеристику. Так, предохранитель срабатывает за 0,1 с, если ток короткого замыкания превысит его уставку (значение входной величины тока) в 10 раз и за 0,2 с — если в 3 раза. Время отключения предохранителя резко возрастает до 9 … 10 с при небольшой силе тока короткого замыкания (в 1,3 раза). По условиям безопасности такая система зануления недопустима.
Для надежного и быстрого отключения электроустановки, находящейся в аварийном состоянии, необходимо, чтобы ток короткого замыкания (А) превосходил ток уставки отключающего аппарата.
Коэффициент кратности короткого замыкания в помещении с нормальными условиями окружающей среды при защите предохранителями или автоматами с тепловым расцепителем должен быть следующий: к ≥ 3; для автоматов с электромагнитным расцепителем -к ≥1,4; для прочих автоматов — к ≥ 1,25.
Во взрывоопасных помещениях в расчете системы зануления принимают значение к ≥ 4 при защите предохранителями и к ≥ 6 — при защите автоматами.
Схема зануления требует наличия в сети нулевого защитного проводника РЕ, глухого заземления нейтрали источника тока и повторного заземления нулевого защитного проводника.
Нулевой защитный проводник в схеме обеспечивает необходимое для отключения электроустановки значение тока однофазного короткого замыкания путем создания для него цепи с малым сопротивлением.
Заземление нейтрали в сети до 1000 В снижает напряжение зануленных корпусов электрооборудования и нулевого защитного проводника относительно земли до малого значения при замыкании фазы на землю.
Повторное заземление нулевого защитного проводника практически не влияет на отключающую способность схемы зануления.
Однако при отсутствии повторного заземления нулевого защитного проводника возникает опасность для людей, прикасающихся к зануленному оборудованию в период замыкания фазы на корпус. Кроме того, в случае обрыва нулевого защитного проводника эта опасность повышается, поскольку напряжение относительно земли других подключенных в этот участок сети зануленных корпусов электродвигателей может достигать фазного напряжения. Повторное заземление нулевого защитного проводника значительно уменьшает опасность поражения током, но не может устранить ее полностью.
Что такое зануление и для чего оно нужно?
В настоящее время существует несколько различных систем электроснабжения потребителей напряжением до 1000 В, однако в России основной в данном случае является система с глухозаземленной нейтралью. Именно такая система используется в каждом нашем доме.
При кажущейся сложности названия все предельно просто. В такой системе нейтральная точка трансформатора на подстанции имеет непосредственное соединение с землей. Основной мерой защиты от случайного попадания под напряжения в данном случае служит защитное зануление, то есть специальное соединение любой металлической части бытового электроприбора с нейтралью трансформатора.
Поскольку, как и было отмечено выше, в таких системах нейтраль глухо соединена с землей то по сути своей защитное зануление не что иное, как одна из разновидностей заземления.
В каждой нашей домашней розетке при правильно выполненной в доме электропроводке имеется заземляющий контакт. Именно через него при включении электроприбора мы соединяем его корпус с нейтральной точкой трансформатора.
Суть работы защитного заземления заключается в следующем. Нормативные документы регламентируют допустимое время отключение поврежденной линии при коротком замыкании не более 0,4 секунд. Именно за это время, как считается, человек имеет все шансы остаться в живых при попадании под напряжение.
При выполнении защитного зануления значительно снижается сопротивление петли «фаза-ноль» и обеспечивается достаточное значение тока короткого замыкания для срабатывания аппарата защиты (предохранитель или автоматический выключатель) за время не более 0,4 секунд.
При отсутствии защитного зануления, или как его еще в быту называют «заземления» ток короткого замыкания за счет высокого сопротивления может оказаться недостаточным для срабатывания защиты и поврежденный бытовой прибор может надолго оказаться под опасным для человека напряжении.
Выполняется защитное зануление в соответствии с требованиями действующих Правил Устройства Электроустановок (ПУЭ). Как правило для этого используется третья жила провода, либо отдельно проложенный медный проводник сечением не менее 4 мм.кв.
Кроме того, в сетях с глухозаземленной нейтралью категорически запрещается выполнять заземление бытовых приборов на отдельный контур заземления, не связанный с нейтральной точкой трансформатора. Например, просто соединив заземляющий контакт розетки с самостоятельно вбитым под окном металлическим стержнем.
Защитное зануление
То же самое и касается попыток «заземления» на систему отопления или водоснабжения квартиры. В этом случае ток короткого замыкания может оказаться достаточно низким за счет того, что земля и дополнительный контур заземления (как правило самодельного производства) имеют значительно большее сопротивление нежели специальный нулевой защитный проводник.
В целом можно сказать, защитное зануление играет огромную роль в обеспечении электробезопасности вашего дома, а качеству и правильности его выполнения следует уделять максимум внимания.
Нейтрализующие антитела — обзор
Нейтрализующие антитела
Реакции нейтрализующих антител на ВИЧ-1 необычно запаздывают и возникают через несколько недель после заражения. 305 , 306 Кроме того, большинство антител, вырабатываемых во время ВИЧ-инфекции, не нейтрализуют или нейтрализуют только узкий круг циркулирующих вирусных штаммов, 73 и многие нацелены на сильно изменчивые петли в gp120, которые действуют как антигенные ловушки, такие как в качестве цикла V1, 307 цикла V2, 308 или цикла V3. 309–314 Широкие эпитопы нейтрализации действительно существуют на gp120 и gp41 (см. Следующий текст), но они остаются минимально доступными для иммунной системы из-за конформационно-зависимого воздействия на молекулу, препятствий со стороны «гликанового щита» и маскировки с помощью гипервариабельные петли в gp120. 315 Однако у некоторых инфицированных вырабатываются широко нейтрализующие антитела. 316–320 Они появляются через большее время после заражения, обычно становятся очевидными в среднем 2.Через 5 лет после заражения, что может означать необходимость созревания аффинности антител, 32 , 321 и у людей с более высокой вирусной плазменной нагрузкой, что указывает на необходимость хронического воздействия антигена. 322–325 От 20% до 30% субъектов в конечном итоге вырабатывают нейтрализующие антитела различной степени широты. 326 Однако только около 1% доноров, обозначенных как элитные нейтрализаторы, проявляют необычно сильную нейтрализующую сывороточную активность против большинства клад. 327 Широко нейтрализующие ВИЧ-1 антитела были также идентифицированы у макак-резусов, инфицированных R5 SHIV SF162P4 , и обнаружено, что они распознают четвертичные эпитопы на gp120. 328
Эксперименты на модели макака показали, что инфекцию SIV или SHIV можно предотвратить путем пассивной инфузии широко нейтрализующих антител 329–335 или с помощью опосредованной аденоассоциированным вирусом (AAV) непрерывной экспрессии иммуноадгезинов, происходящих из широких масс. нейтрализующие антитела. 336 Полная защита от инфекции SHIV 89.6P у взрослых макак была получена путем пассивной иммунизации ВИЧ-1 нейтрализующими моноклональными антителами 2 G12 и 2 F5 и иммуноглобулинами против ВИЧ-1 332 , 337 или моноклональное антитело 2 F5 или 4E10. 338 Аналогичным образом неонатальные макаки могут быть защищены от перорального заражения SHIV путем пассивной иммунизации широко нейтрализующими моноклональными антителами b12, 2 G12 и 2 F5. 330 , 339 Таким образом, широко нейтрализующие антитела не связаны с контролем виремии у хозяина, 323 , 340–342 , но они могут защищать от заражения живым вирусом.
Тем не менее, остается задача воплотить эти открытия в разработке иммуногенов, которые могли бы вырабатывать широко нейтрализующие антитела при включении в вакцину. 66 , 343–349
Профилактическое и терапевтическое применение нейтрализующих антител к вирусу иммунодефицита человека типа 1 (ВИЧ-1)
Ther Adv Vaccines. 2013 июл; 1 (2): 67–80.
иRajesh Ringe
Weill Medical College of Cornell University, New York, NY, USA
Jayanta Bhattacharya
International AIDS Vaccine Initiative (IAVI), THSTI-IAVI HVTR Laboratory, Translational Health Science and Technology Institute (THSTI ), Гургаон-122016, Харьяна, Индия.
Раджеш Ринге, Медицинский колледж Вейля Корнельского университета, Нью-Йорк, Нью-Йорк, США;
Автор, ответственный за переписку. Эта статья цитируется в других статьях в PMC.Abstract
Разработка профилактической вакцины для нейтрализации очень вариабельного и антигенно разнообразного вируса иммунодефицита человека типа 1 (ВИЧ-1) была непреодолимой целью. Недавнее открытие ряда перекрестно нейтрализующих и мощных моноклональных антител от элитных нейтрализаторов дало важные идеи в этой области.Нейтрализующие антитела (NAb) полезны для идентификации нейтрализующих эпитопов, пригодных для вакцины, и для понимания механизма сильной и широкой перекрестной нейтрализации, таким образом обеспечивая профилактическое и терапевтическое значение. В этой статье мы рассматриваем текущее понимание потенциального использования широко нейтрализующих антител (bNAbs) в их полноразмерной структуре IgG, сконструированных доменных антителах или биспецифических версиях для профилактических и терапевтических применений. Возможные последствия NAb обсуждаются в свете последних достижений в качестве ключевых компонентов вакцинации против ВИЧ-1.Разработка иммуногена для вакцины, который вырабатывает bNAb и обеспечивает защитный иммунитет, остается реальной проблемой.
Ключевые слова: ВИЧ-1, нейтрализующие антитела, вакцины
Введение
Синдром приобретенного иммунодефицита (СПИД), вызванный вирусом иммунодефицита человека 1 типа (ВИЧ-1), является серьезной проблемой общественного здравоохранения и требует срочной помощи разработка профилактической вакцины. Свидетельства, собранные в результате недавних открытий, подкрепленных фундаментальными научными исследованиями и клиническими испытаниями, действительно вселяют оптимизм в отношении возможности разработки вакцины против ВИЧ-1.Антитела, которые блокируют проникновение вируса, называются нейтрализующими антителами (NAb), и они считаются решающим коррелятом защиты в некоторых успешных вирусных вакцинах, а также могут быть эффективными против ВИЧ-1 [Haynes and Montefiori, 2006; Маскола, 2003; Montefiori et al. , 2007а; Плоткин, 2008]. Однако для полной защиты, по-видимому, необходимы обе стороны адаптивного иммунного ответа, ответ антител и ответ Т-клеток [Walker and Burton, 2008]. Оба этих ответа работают согласованно: антитела контролируют бесклеточное проникновение вируса в клетки-мишени, тогда как Т-клеточный ответ контролирует вирусные резервуары и необходим для проверки репликации вируса [McMichael and Hanke, 2002; Панталео и Куп, 2004].Передача вируса после нарушения слизистого барьера в подслизистую среду приводит к инфицированию окружающих CD4 + Т-клеток, а затем распространяется в системную инфекцию. Устойчивая репликация вируса приводит к разрушению CD4 + Т-клеток, что приводит к СПИДу [Letvin, 2006; МакМайкл, 2006]. Белок оболочки (gp160), кодируемый ВИЧ env , является самым внешним белком, экспрессируемым на ВИЧ. Он действует как молекулярная машина, которая связывает вирус с рецепторами клетки-мишени, таким образом опосредуя слияние клеточной мембраны и проникновение вируса [Wyatt et al. 1998]. Env является важным компонентом проникновения вируса и представляет собой привлекательную мишень для индуцированных вакциной антител, которые могут связываться с Env и блокировать проникновение вируса в клетку-мишень [Burton et al. 2004; Хейнс и Монтефиори, 2006; Montefiori et al. 2007b]. В последние годы был достигнут значительный прогресс в открытии антител, и у хронически инфицированных ВИЧ-положительных пациентов были выделены высокоактивные и широкие NAb с широко нейтрализующей сывороточной активностью, также называемые элитными нейтрализаторами.Эти антитела при пассивной иммунизации животных обеспечивали защиту нечеловеческих приматов и гуманизированных мышей [Burke and Barnett, 2007; Klein et al. 2012b; Mascola, 2003, Moldt et al. ., 2012]. Экспрессия in vivo широко нейтрализующих антител (bNAbs) посредством векторно-опосредованного переноса генов также показала высокую эффективность у гуманизированных мышей [Balazs et al. 2012]. Однако попытки вызвать такие антитела с помощью иммунизации не увенчались успехом [Burton et al. 2004; Хейнс и Монтефиори, 2006]. Исходная рекомбинантная белковая вакцина на основе белка gp120 индуцировала только иммуноген-специфические антитела, которые могли нейтрализовать лабораторно адаптированные вирусные штаммы, но не первичные изоляты, и, таким образом, не показала клинической значимости [Flynn et al . 2005; Gilbert et al. 2005; Грэм и Маскола, 2005]. Однако недавнее испытание вакцины против вируса оспы канареек (ALVAC-HIV) против вируса RV144 в сочетании с вакциной gp120 AIDSVAX B / E продемонстрировало умеренную эффективность и обещает, что антитела, индуцированные вакцинацией, могут обеспечить защитный иммунитет против ВИЧ-1 [Baden and Dolin , 2012; Rerks-Ngarm et al. 2009]. Интересно, что антитела в испытании RV144 в основном не нейтрализуют; однако именно связывание антител IgG с областью V1V2 Env gp120, вероятно, было коррелятом защиты в этом испытании [Haynes et al. 2012a]. Хотя этот режим не привел к появлению NAb, результаты этого испытания могут предоставить ценное руководство в отношении усилий по улучшению иммуногена и антител, необходимых для защиты от инфекции ВИЧ-1. Предпринимаются усилия по созданию улучшенных иммуногенов на основе новейших подробных структурных представлений о белке Env, которые демонстрируют лучший ответ антител [Kovacs et al. 2012; Phogat and Wyatt, 2007]. Улучшенное знание структуры Env и нейтрализующих эпитопов поможет улучшить рациональную структуру иммуногена, чтобы вызвать сильные bNAb [Dormitzer et al. 2008; Квонг и Уилсон, 2009 г .; Montefiori et al. 2007b; Phogat and Wyatt, 2007; Stamatatos et al. 2009]. В данной статье рассматривается текущее понимание прогресса в открытии широких и сильнодействующих NAb к ВИЧ-1, а также их потенциала в терапевтических и профилактических целях против ВИЧ-1.
Нейтрализующие эпитопы на оболочке ВИЧ-1
Хотя антитела вырабатываются против большинства вирусных белков, те, которые связываются с белком Env и предотвращают проникновение вируса, называются NAb [Mascola and Montefiori, 2010; Пантофлет и Бертон, 2006; Золла-Пазнер, 2004]. Особенно сложно достичь уникальной субъединичной архитектуры тримера Env ВИЧ-1, индуцирующего NAb [Mao et al. 2012]. Антитела на ранней стадии инфекции, как правило, специфичны к штамму, но у некоторых пациентов bNAb развиваются на хронической стадии инфекции.Примерно у 20% пациентов с ВИЧ с хронической инфекцией развиваются НАТ, способные нейтрализовать различные штаммы ВИЧ-1, а 2–4% таких субъектов обладают еще большей сывороточной нейтрализующей активностью, которая нейтрализует большинство штаммов ВИЧ-1 из разных кладов [Simek et al. . 2009]. С антигенной точки зрения белок Env очень вариабелен, и вирус может быстро ускользнуть от селективного давления со стороны существующих NAb. Тем не менее сыворотки некоторых хронически инфицированных пациентов проявляют более широкую нейтрализующую активность, которая объясняется единственной, небольшой или множественной специфичностью [Scheid et al. 2009; Уокер и др. 2009; Уокер и др. 2010; Wu et al. 2010]. Первое широко нейтрализующее человеческое моноклональное антитело (mAb) b12 было выделено от пациента, инфицированного кладой B, и оно связывается с gp120 на своем сайте связывания CD4 (CD4bs) [Burton et al. 1994]. Было обнаружено, что b12 нейтрализует более 50% изолятов вируса клады B и около 30% вирусов, не относящихся к кладе B [Binley et al . 2004; Kulkarni et al. 2009]. Недавно с использованием подходов обратной вакцинологии были выделены новые формы широких и сильных антител к CD4bs из элитного нейтрализатора [Falkowska et al. 2012; Wu et al. 2011a]. Эти человеческие mAb были выделены с использованием способности стабилизированного корового белка gp120 с обновленной поверхностью, имитирующего сайт связывания CD4, захватывать только широкие NAb на основе аффинности и глубокого распознавания (слабосвязанные не-NAb не допускались). Используя это модифицированное ядро gp120 в качестве флуоресцентно меченного зонда для захвата антиген-специфических В-клеток памяти, также были выделены CD4bs-специфические антитела против VRC. VRC01 и VRC02 являются самыми широкими, нейтрализующими 91% первичных псевдовирусов Env, в то время как VRC03 нейтрализует 57% циркулирующих вирусов панели () [Wu et al. 2010]. NIh55-46 является более мощным клональным вариантом VRC01 и имеет вставку из четырех остатков в области 3, определяющей комплементарность тяжелой цепи (CDR3), которая усиливает связывание gp120 [Scheid et al. 2011]. 3BNC117 [Scheid et al. 2011] и VRC-PG04 [Wu et al. 2011b] также являются антителами к CD4bs с широтой нейтрализации и эффективностью, аналогичными VRC01. Хотя все эти антитела связываются с CD4bs, существуют различия в том, как они взаимодействуют с этим сайтом.Структурный анализ взаимодействия CD4bs с mAb b12 (прототип CD4bs NAb), F105 (ненейтрализующий) и VRC01 (очень широко нейтрализующий) выявил необычное связывание с узким сайтом [Schief et al. 2009; Zhou et al. 2010]. VRC01 приближается к конформационно инвариантному сайту после начального присоединения CD4, избегая препятствий за счет конформационного маскирования, которое испытывает большинство антител против CD4bs, тем самым уменьшая их нейтрализующую способность. Таким образом, частичная мимикрия рецептора и обширное созревание аффинности способствуют эффективной нейтрализации ВИЧ-1 естественными человеческими антителами, такими как VRC01 [Zhou et al. 2010]. Два прототипа NAb gp41, проксимальная к мембране (MPER) — это 2F5 и 4E10 [Cardoso et al. 2005; Офек и др. 2004; Stiegler et al. 2001]. Субъединица gp41 гораздо более консервативна, чем gp120, но кинетические и стерические ограничения потенциально защищают ее уязвимые области от атаки NAb. MAb 2F5, 4E10 и Z13 были выделены от ВИЧ-положительного пациента и показали значительную эффективность, при этом 4E10 является наиболее нейтрализующим веществом [D’Souza et al. 1997]. Эти MAb связываются с промежуточной конформацией gp41 в процессе слияния [Frey et al. 2008]. Нативный gp41, по-видимому, недоступен для антител и выявляет эпитопы в процессе слияния. Следовательно, против этих эпитопов вырабатываются ограниченные антитела, поскольку их воздействие слишком короткое, чтобы их могли распознать В-клеточные рецепторы для выработки антител против них, либо у антитела есть небольшое окно возможностей для связывания с короткоживущими эпитопами, которые подвергаются воздействию только во время слияния [Alam et al. 2011; Ринге и Бхаттачарья, 2012; Шен и др. 2010]. 2F5 связывается с основной мишенью в ELDKWA в области гептадного повтора-2 gp41, тогда как 4E10 связывается с последовательностью NWFDIT в MPER [Cardoso et al. 2005; Офек и др. 2004]. Эти антитела связываются как с комплексами липид-пептид MPER, так и с вирионами ВИЧ [Alam et al. 2009] в двухэтапной модели изменения конформации, в которой они сначала связываются с липидами вириона, затем перемещаются по вирусной мембране, ожидая временного воздействия своих нейтрализующих эпитопов во время слияния с клеткой-мишенью.Эти антитела являются аутореактивными из-за их реакционной способности с липидами [Alam et al. 2009]. Недавно Хуанг и его коллеги сообщили об очень сильном и широко перекрестно нейтрализующем MPER-специфическом антителе gp41, названном 10E8 [Huang et al. 2012]. 10E8 нейтрализует 98% протестированных вирусов (). Более того, в отличие от 2F5 или 4E10, 10E8 не связывает фосфолипиды, не является аутореактивным и связывается с оболочкой клеточной поверхности, что позволяет предположить, что высококонсервативный эпитоп в эктодомене gp41 напрямую доступен для 10E8 и потенциально может увеличить окно возможностей для доступа эпитоп.Исследование также показало, что 8% сывороток здоровых ВИЧ-1-позитивных людей содержали 10E8-подобную специфичность, что свидетельствует о том, что соответствующий эпитоп является иммуногенным по природе, а 10E8-подобные антитела не удаляются из репертуара B-клеток из-за аутореактивности. Структура 10E8 в комплексе с полным MPER и исследования мутагенеза выявили сайт связывания, содержащий узкий участок высококонсервативных gp41-гидрофобных остатков и критический аргинин или лизин непосредственно перед трансмембранной областью [Huang et al. 2012]. Высококонсервативный MPER является мишенью для мощных, не самореактивных NAb. Частое образование 10E8-подобных антител у ВИЧ-1-инфицированных людей предполагает, что эта специфичность может быть вызвана вакцинацией у большей части ВИЧ-отрицательных или ВИЧ-положительных людей, чем у других специфичных к gp41 антител. Другой иммунодоминантной областью в Env является петля V3 на gp120, и часто вырабатываются антитела против эпитопов на конце этой петли. Специфические антитела к V3 обычно имеют узкую реактивность и могут нейтрализовать только штаммы уровня 1 или адаптированные к линии Т-клеток [Davis et al. 2009]. Многие V3-специфические антитела с некоторой широкой реактивностью от хронически инфицированных ВИЧ-1 субъектов были выделены, которые распознают штамм-специфический четвертичный эпитоп, включающий gp120 V2 и петли V3 в контексте тримера Env [Zolla-Pazner and Cardozo, 2010]. Новые технические достижения в области стимуляции В-клеток с одной памятью и высокопроизводительного анализа микронейтрализации облегчили скрининг антител до более высоких уровней. Выделение сильнодействующих и широко нейтрализующих mAb PG9 и PG16 (близкородственных соматических мутантов) от пациента было выполнено Уокером и его коллегами с использованием этой методологии [Walker et al. 2009]. PG9 и PG16 распознают новый эпитоп, состоящий из петель V1V2 и V3 на тримере Env, и проявляют замечательную широту нейтрализации и эффективность [McLellan et al. 2011; Уокер и др. 2011]. Совсем недавно, используя ту же технологию, многие дополнительные моноклональные антитела, называемые антителами PGT, были выделены из четырех элитных нейтрализаторов, которые нацелены на гликан-зависимый эпитоп и проявляют примерно в 10 раз большую эффективность, чем VRC01, PG9 и PG16, и в 100 раз больше, чем прототипные антитела старого поколения.Среди этих антител к PGT PGT141-145 нацелен на гликан-зависимые четвертичные эпитопы на gp120, тогда как PGT 125-128 и PGT130-131 специфически взаимодействуют с гликанами Man8 / 9 на gp120. Эти антитела очень эффективны, при этом некоторые нейтрализуют> 70% панели псевдотипов вируса env ВИЧ-1 из 162 оболочек () [Pancera et al. 2010; Уокер и др. 2009, 2011]. Кристаллическая структура PGT128, наиболее мощного и широкого PGT, в комплексе с полностью гликозилированным внешним доменом gp120, показывает, что антитело проникает через гликановый щит и распознает два консервативных гликана, а также короткий сегмент β-цепи петли V3 gp120, что объясняет его высокую аффинность связывания и широкую специфичность [Pejchal et al. 2011]. 2G12, прототипное специфичное к углеводам mAb, необычно по своей структуре и специфичности связывания. В отличие от PG9 или PGT mAb, это канонический гликан-связывающий bNAb с уникальной структурой обмена доменами. Его Fab-область состоит из тяжелой цепи и легкой цепи, которые являются общими для другого плеча антитела 2G12 [Calarese et al . 2003; Scanlan et al. 2002]. Гликаны на gp120 являются результатом посттрансляционной модификации гликозидаз клетки-хозяина и поэтому напоминают углеводы хозяина, возможно, снижая иммуногенность gp120 [Scanlan et al. 2002, 2003]. Гликаны, распознаваемые 2G12, содержат уникальный конформационный эпитоп олигоманнозных гликанов во внешнем домене gp120, который является слабо иммуногенным [Astronomo et al. 2008]. Однако новые гликановые антитела распознают эти домены намного сильнее и нейтрализуют многие первичные изоляты [Walker et al. 2011]. Это говорит о том, что иммунный ответ при ВИЧ-инфекции проходит сложный путь развития и распознавания таких сложных и трудных эпитопов [Mouquet et al. 2012]. Таким образом, оболочка ВИЧ-1 имеет по крайней мере пять консервативных областей, каждая с перекрывающимися эпитопами, которые могут быть мишенями для bNAb ().
Таблица 1.
Характеристики широких и мощных нейтрализующих антител к ВИЧ-1
| Название антитела | Место контакта | Исследование | Тип антитела | Гликановая зависимость | Четвертичная структура Нейтрализация | (%)Инфекционный подтип | Терапевтический потенциал | |
|---|---|---|---|---|---|---|---|---|
| IgG1b12 | CD4bs | Burton et al. (1991) Proc. Natl. Акад. Sci. США; 88: 10134-10137 | Целое антитело | № | № | 35 (n = 190) | B | Эффективно против вагинального заражения SHIV и защищает макак. |
| VRC01,02,03 | CD4bs | Wu et al. (2010) Science329: 856-861 | Целое антитело | № | № | 91, 91, 57 (n = 190) | B | Эффективно против заражения слизистой оболочкой SHIV и защищает макак. |
| NIh55-46 | CD4bs | Scheid et al. (2011) Science, 333: 1633-1637 | Целые антитела | Нет | Нет | 96 (n = 118) | B | Не тестировалось |
| VRC-PG04 | CD267s | и другие. (2011) Наука; 333: 1593-602Целое антитело | Нет | Нет | 76 (n = 178) | Рекомбинантный A-D | Не тестировалось | |
| 3BNC117 | CD4bs и др. | 902 (2011) Science, 333: 1633-1637Целые антитела | Нет | Нет | 96 (n = 118) | B | Не тестировалось | |
| VHH J3 | CoRbs | . и Вайс Р. (2013). J Exp Med 210: 209-223.Доменное антитело | NA | NA | 96 (n = 100) | — | Не тестировалось | |
| m36 | CoRbs | Chen (2008) Proc Natl Acad Sci | Домен Домен НетНет | 91 (n = 11) | — | Не тестировалось | ||
| 2008; 105: 17121-17126 | ||||||||
| 2G12 | Гликан | Тркола и др. (1996) J. Virol. 70: 1100-1108) | Целое антитело | Да | Нет | 32 (n = 162) | B | Снижает вирусную нагрузку и увеличивает количество Т-лимфоцитов CD4 в сочетании с 2F5, 4E10 |
| PGT121 123 | Гликан | Walker et al. (2011) Природа; 477: 466-470) | Целые антитела | Да | Нет | 65-70 (n = 162) | A | Не тестировались |
| PGT125-128,130-131 | etlycan | al. (2011) Природа; 477: 466-470)Целые антитела | Да | Нет | 40-72 (n = 162) | CRF02_AG | Не тестировалось | |
| PGT135 | Gly67 et al. | (2011) Природа; 477: 466-470)Целое антитело | Да | Нет | 33 (n = 162) | C | Не тестировалось | |
| PGT141-145 | Glycan | Walker (2011) Природа; 477: 466-470) | Целое антитело | Да | 38-78 (n = 162) | A или D | Не тестировалось | |
| PG9, PG16 | Четвертичная структура, включая V1V268 | Walker et al. (2009) Science 326: 285-289 | Целые антитела | Да | Да | 79 и 73 (n = 190) | A | Не тестировалось |
| 2F5 | MPER | Целое антитело | № | № | 57 (n = 177) | B | Испытано в комбинации с 2G12. Помогите снизить вирусную нагрузку и увеличить количество CD4 + Т-лимфоцитов у ВИЧ-инфицированных. | |
| 4E10 | MPER | Stiegler et al. (2001) AIDS Res. Гм. Ретровирусы 17: 1757-1765 | Целое антитело | Нет | Нет | 98 (n = 180) | B | Умеренно подавляет вирусную нагрузку в комбинации с 2F5 |
| 10E8 | и т.д. al (2012) Nature, 491: 406-412) | Целое антитело | Нет | Нет | 98 (n = 180) | B | Не тестировалось |
Потенциальные сайты ответа антител на Env .Тример оболочки состоит из трех субъединиц поверхностного белка gp120 (фиолетовый) и трех трансмембранных субъединиц gp41 (зеленый), как показано. gp120 имеет несколько потенциальных кластеров (обозначенных кружками), против которых у ВИЧ-инфицированных вырабатываются нейтрализующие антитела (многие из них были выделены из широко нейтрализующих сывороток). Белок gp41 до сих пор имел один сайт в области эктодомена (обозначен пунктирными кружками), против которого выделяются широко нейтрализующие антитела. Тример оболочки показан в открытой конформации (после взаимодействия с CD4), чтобы выявить кластер сайтов связывания корецепторов, который экспонируется после взаимодействия gp120 с CD4.MPER, мембранно-проксимальная наружная область. Иллюстрация любезно предоставлена Алессандро Балиани. Copyright © 2013.
НК ВИЧ-1 и ингибиторы на основе антител
НК являются очень мощными агентами против большинства вирусов животных и в иммунных ответах, индуцированных вакцинами [Casadevall, 2002; Klasse, Sattentau, 2002]. Однако лечение ВИЧ-1 с помощью NAb не было успешным из-за быстрой мутабельности вируса [Manrique et al. 2007; Wei et al. 2003].Увеличение мощности и широты может преодолеть эту пугающую проблему. В этом направлении были предприняты значительные усилия, и было создано несколько ингибиторов на основе антител с желательными характеристиками. В дополнение к информационным стратегиям для разработки вакцинных иммуногенов и получения аналогичных ответов антител, терапевтическое применение bNAb у пациентов, не прошедших антиретровирусную терапию (АРТ), или у тех, у кого развилась резистентность, представляет собой многообещающий подход к искоренению вируса. Таким образом, bNAb можно использовать индивидуально или в комбинации с АРТ для эффективного устранения вируса [Chen et al. 2013; Corti, Lanzavecchia, 2013]. В этом обзоре мы обсуждаем, как модифицируются антитела для достижения желаемых характеристик и как моноклональные антитела в их нативной форме или в виде сконструированных версий могут быть использованы для защиты от ВИЧ-инфекции.
Нейтрализация ВИЧ-1 нативными формами mAb
Как обсуждалось ранее, было охарактеризовано несколько важных нейтрализующих эпитопов, против которых были выделены bNAb. Одним из наиболее хорошо охарактеризованных широко перекрестно-реактивных и сильнодействующих mAb человека, нацеленных на CD4bs gp120, является b12.Он был выбран почти 20 лет назад путем фагового дисплея из библиотеки антител, сделанной из костного мозга ВИЧ-1-инфицированного донора [Burton et al. 1994; Roben et al. 1994]. Было обнаружено, что внутривенное переливание b12 частично защищает макак от зараженного вагинальным путем R5 вируса SHIV162P4, а позже было обнаружено, что вагинальное введение b12 может частично защитить макак от передачи вагинального вируса иммунодефицита обезьян / человека (SHIV) [Parren et al. 2001; Veazey et al. 2003]. Эти наблюдения предполагают, что mAb могут быть потенциальным средством предотвращения передачи ВИЧ-1 людям половым путем. Синергетический эффект нескольких mAb вместе может улучшить защитный эффект и может поставить вирус под угрозу до уровня, с которого вирусный возврат затруднен [Klein et al. 2012a]. Однако механизм, с помощью которого NAbs обеспечивают защиту in vivo , не очень ясен. Помимо прямой нейтрализации, эффекторные функции, опосредованные рецептором Fc антитела, также важны для защиты от ВИЧ-1.Удаление связывания b12 с Fc-рецептором привело к потере его защитной функции, что свидетельствует о том, что антитело обладает способностью нейтрализации как бесклеточного, так и связанного с клетками вируса [Hessell et al. 2007]. Молдт и его коллеги [Moldt et al . 2012] продемонстрировали, что усиление взаимодействия b12 с FcgammaRIIIa усиливает противовирусную активность in vitro , но не может оказывать тот же эффект in vivo . Таким образом, точный противовирусный механизм действия NAbs in vivo остается неясным.Углеводно-специфическое антитело 2G12 было испытано внутривенно на животных моделях на предмет его эффективности против зараженного вагинальным вирусом SHIV. Результаты показали значительную защиту макак или снижение вирусной нагрузки и задержку потери CD4 + Т-клеток [Chen and Dimitrov, 2012; Маскола и др. . 1999, 2000]. Hessell и его коллеги [Hessell et al. 2009] также показали, что низкое внутривенное введение 2G12 может защитить животных от вагинального заражения SHIV, в отличие от наблюдения, что высокая доза b12 требовалась для создания защитного иммунитета.В этих исследованиях использованный контрольный вирус был очень чувствителен к тестируемым NAb. Исследования 2F5 и 4E10 in vitro показали, что нейтрализующая активность этих mAb усиливается при тестировании в комбинации [Mascola et al. 1997], и поэтому они были оценены на предмет их противовирусной активности in vivo в основном в комбинации на животных моделях и на ВИЧ-инфицированных. По отдельности ни 2F5, ни 2G12 полностью не защищали макак от внутривенного заражения, но леченные животные показали менее глубокое падение CD4 + Т-клеток, что позволяет предположить, что эти антитела нарушают репликацию вируса in vivo [Mascola et al. 1999]. Эти первоначальные исследования на животных моделях документально подтвердили некоторый успех в защите от инфекции или замедлении вирусного отскока / потери CD4 + Т-клеток при вагинальном заражении SHIV. Позже комбинация 2F5 / 2G12 была также оценена в клиническом испытании фазы I на семи ВИЧ-1-инфицированных пациентах [Armbruster et al. 2002], который подтвердил безопасность этих mAb. Кроме того, временное снижение вирусной нагрузки наблюдалось у пяти из семи субъектов, в то время как улучшение количества CD4 + Т-клеток и соотношения CD4 + / CD8 + наблюдалось у всех людей.Однако вирус ВИЧ может развить устойчивость к 2G12, что делает этот подход неэффективным. По этой причине комбинация трех mAb 4E10, 2F5 и 2G12 [Armbruster et al. 2004] также оценивали у семи человек. Исследование подтвердило безопасность этой комбинации и некоторую защиту. В клиническом исследовании фазы II [Trkola et al. 2005] оценивалась способность этих антител подавлять или задерживать вирусный отскок, вызванный АРТ. Существенная задержка рецидива вируса наблюдалась у четырех из шести остро инфицированных лиц и только у двух из восьми хронически инфицированных пациентов.Это говорит о том, что mAb могут предотвращать прогрессирование заболевания, когда ВИЧ-инфекция находится на ранней стадии и вирусное разнообразие ограничено [Bar et al. 2012]. При хронической инфекции вирусное разнообразие и вирусные резервуары могут быть слишком высокими, чтобы их можно было контролировать с помощью моноклональных антител. Устойчивость к 2G12, но не к 2F5 или 4E10, наблюдалась у 12 из 14 человек, что позволяет предположить, что паттерн гликозилирования можно легко изменить без ущерба для функции gp120, чтобы противостоять связыванию с 2G12, тогда как MPER имеет решающее значение в процессе слияния и приобретении устойчивости. к соответствующим антителам может потребоваться более высокая стоимость приспособления [Manrique et al. 2007]. Последующие исследования показали, что противовирусный эффект этих трех mAb является результатом прямой нейтрализации in vivo в дополнение к антителозависимой клеточной цитотоксичности (ADCC) и механизмам фагоцитоза [Huber et al. 2008].
Модификации антитела для усиления противовирусной активности
Целое антитело представляет собой большую молекулу, которая обычно связывается с большей частью антигенного белка для повышения его авидности. Проблема, связанная с этими большими молекулами, состоит в том, что они не могут получить доступ к эпитопам, когда они стерически закрыты.Это особенно характерно для многих эпитопов белков оболочки ВИЧ-1, что приводит к устойчивости к антителам, индуцированной ВИЧ-инфекцией [Burton et al. 2005; Маскола и Монтефиори, 2010]. Для таких изменяющихся эпитопов небольшие фрагменты антител могут быть более эффективными и более способными контролировать репликацию вируса. Сайт связывания для таких объектов меньше, и поэтому можно избежать стерической окклюзии [Labrijn et al. 2003]. Labrijn и его коллеги [Labrijn et al. 2003] показали, что эффективность нейтрализации CD4-индуцированного (CD4i) эпитоп-связывающего антитела обратно коррелирует с размером фрагментов антитела scFv (~ 25 кДа). Похоже, что меньшие форматы антител могут получить доступ к сайту связывания легче, чем целое антитело, поскольку доступ для последнего стерически ограничен из-за небольшого пространства между вирусной оболочкой и клеточными мембранами после или до взаимодействия тримера Env с рецептором CD4. [Labrijn et al. 2003].Небольшое количество NAb CD4i было выделено от пациентов с ВИЧ и представлено 17b (IgG), X5 [Chen and Dimitrov, 2009; Moulard et al. 2002], m9 и m16 [Zhang et al . 2004]. Эти антитела к CD4i обладают сильной нейтрализующей активностью и терапевтической значимостью, поскольку их сайт-мишень в основном консервативен в структуре Env. Более новый класс антител называется доменными антителами (dAbs), которые представлены сконструированными фрагментами антител, которые являются достаточно эффективными и имеют меньший размер по сравнению с Fab или scFv [Chen et al. 2008a, 2008b; Holt et al. 2003; Vanlandschoot et al. 2011]. Эти небольшие фрагменты антител размером от 11 до 15 кДа лишены домена VL или VH и направлены против консервативных доменов эпитопа. Это особенно важная характеристика, поскольку большинство иммуногенных консервативных эпитопов Env защищены конформационной маскировкой вариабельными петлями или гликановым экраном и ускользают от иммунного распознавания [Kwong et al. 1998; Пантофлет и Бертон, 2006; Wyatt et al. 1998]. Эпитопы, которые недоступны для полноразмерных антител или Fab / scFv из-за стерических ограничений, легко доступны для dAbs [Chen and Dimitrov, 2009; Чен и др. 2008a; Gong et al. 2012]. Сайт связывания корецептора (CD4i) является наиболее стерически закрытой иммуногенной структурой в gp120, поскольку его образование индуцируется только после связывания gp120 с CD4. Поэтому была выдвинута гипотеза, что небольшие фрагменты антител (полученные из доменов целых антител), нацеленные на такие эпитопы, могут нейтрализовать вирус с широтой и эффективностью [Chen et al. 2010]. Высокоэффективный dAb m36, описанный Ченом и его коллегами, направлен на консервативную структуру сайта связывания корецептора (CD4i) [Chen et al. 2008a]. Это антитело широко нейтрализует перекрестную реакцию, обладает мощной нейтрализующей активностью и показало более высокую эффективность, чем антитело scFv m9 [Chen and Dimitrov, 2009]. Такие dAb могут быть особенно эффективными против кинетических сигнатур, таких как эпитопы CD4i и MPER. Некоторые dAb также были недавно выделены при иммунизации лам на основе рекомбинантного gp120 и проявили сильную нейтрализующую активность против ВИЧ-1 [Forsman et al. 2008]. Некоторые вариабельные домены, происходящие из антител тяжелой цепи (VHH), такие как A12, D7 и C8, были способны нейтрализовать изоляты клады B и C ВИЧ-1 [Forsman et al. 2008; Strokappe et al. 2012]. Используя более строгие критерии отбора, VHH смог нейтрализовать 42% протестированных штаммов in vitro с IC 50 в диапазоне <0,2–2533 нМ, хотя в целом они имеют более высокую специфичность в отношении иммунизирующего штамма или клады. Эти dAb блокируют связывание CD4 с gp120 и могут конкурировать со связыванием mAb CD4bs с gp120 [Forsman et al. 2008]. Кристаллическая структура фрагмента антитела тяжелой цепи ламы VHH D7 выявила наличие двух канонических CDR1 и CDR2, но более длинного и высокомобильного CDR3, который, вероятно, необходим для распознавания и предоставления большей энергии связывания для взаимодействия с gp120 CD4bs и нейтрализации вируса. [Hinz et al. 2010; Koh et al. 2010; Маккой, Вайс, 2013]. Используя семейно-специфический подход, Ко и его коллеги недавно выделили максимально возможное разнообразие родственных VHH-антител, которые конкурируют с растворимым CD4 за связывание с гликопротеином оболочки ВИЧ-1 [Koh et al. 2010; Strokappe et al. 2012]. Затем Matz и его коллеги расширили список dAb за счет иммунизации лам с использованием тримерного gp140, свободного или связанного с миметиком CD4, для выделения dAbs CD4bs и CoRbs [Matz et al. 2013]. Однодоменные антитела (sdAbs), выделенные в этом исследовании, эффективно нейтрализовали вирусы подтипа B, но также показали нейтрализующую активность против вирусов, несущих оболочки от A, C, G, CRF01_AE и CRF02_AG, подтипов, включая вирусы уровня 3.Новый модифицированный процесс скрининга, способный различать нейтрализующие и не-NAb, позволил выделить чрезвычайно широкий и мощный VHH J3 путем скрининга фагмидной библиотеки VHH, полученной из ламы, иммунизированной двумя рекомбинантными белками gp140 ВИЧ. Этот dAb VHH нейтрализовал 96% большой панели штаммов ВИЧ-1 и представляет собой потенциального терапевтического кандидата [McCoy and Weiss, 2013]. Эта работа продемонстрировала, что широкие и мощные антитела меньшего формата могут быть получены при иммунизации и рассмотрены для нескольких применений, таких как микробицид против ВИЧ-1 и для рационального дизайна иммуногена ВИЧ для ВИЧ для определения уязвимых эпитопов на белке Env.Однако с этими антителами связаны некоторые ограничения. У них меньший период полувыведения и низкое удерживание in vivo. Кроме того, противовирусное действие фрагмента антитела заключается в конкурентном связывании и нейтрализации. Однако другие функции IgG, такие как Fc-опосредованная ADCC или фагоцитарные эффекты, отсутствуют, что может снизить противовирусный эффект этих антител. Наконец, развитие устойчивости к одному способу противовирусного действия может быть менее трудным, чем у антител с несколькими противовирусными механизмами.
Биспецифические антитела
Решение вопросов эффективности на клиническом уровне и вирусной устойчивости к bNAbs вызвало необходимость повышения эффективности и расширения NAb. В нескольких исследованиях за последние годы описано получение продуктов конъюгации антител с двумя различными специфичностями в отношении белка Env. Растворимая форма CD4 представляет собой вирус-нейтрализующий белок и использовалась для конъюгирования с антителами, которые нацелены на эпитопы, которые индуцируются или экспонируются при связывании CD4 с Env [Chamow et al. 1992; Чен и др. 2013; Traunecker et al. 1992; West et al. 2010]. Внешние домены рецептора CD4 D1D2, конъюгированные с антителом, специфичным к эпитопу CD4i (антитело CD4-CD4i), показали повышенную эффективность и широту нейтрализации [Chen and Dimitrov, 2009; West et al. 2010]. Двухвалентный реагент, который сливает CD4 с тяжелой цепью антитела Е51 к CD4i, показал аналогичную или более высокую нейтрализующую способность, чем хорошо известные bNAb [West et al. 2010]. Сходным образом были исследованы гибриды иммуноадгезин-антитело, в которых scFv конъюгирован с Fc-доменом, и их эффективность нейтрализации по сравнению с таковыми у родительского IgG. Иммуноадгезины, полученные из PG9, PG16 и VRC01, показали пониженную активность, вероятно, из-за пониженного сродства родственного эпитопа. Однако присоединение scFv VRC01 к PG16 IgG дало биспецифический реагент, нейтрализующая активность которого объединяла активности обоих родительских антител, а также меньшее количество штаммов, избежавших нейтрализации [West et al. 2012]. Такой подход объединения антигенсвязывающих сайтов вместе в одно антитело увеличивает авидность антитела, что, вероятно, приводит к увеличению активности [Cavacini et al. 1994; Kausmally et al. 2004]. Помимо валентности, улучшенная гибкость и размер сайтов связывания антигена могут влиять на эффективность и широту нейтрализации. Было замечено, что эффективность b12 по нейтрализации панели оболочек клады B возрастает с увеличением валентности и гибкости между сайтами связывания антигена.Разработанный b12 продемонстрировал способность связывать бивалентно и сшивать шипы оболочки на поверхности вириона. Этого не наблюдали с аналогичным образом сконструированными антигенсвязывающими сайтами 4E10, вероятно, потому, что эпитоп 4E10 труднодоступен в нативном тримере, и такая конструкция может иметь наименьшее значение [Klein et al. 2009]. Недавние исследования установили тот факт, что большинство bNAb являются полиреактивными и гипермутированными [Mouquet et al. 2010]. BNAb образуются на хронической стадии ВИЧ-инфекции и созревают за счет соматических гипермутаций в основном, но не ограничиваясь ими, в сайтах, связывающих антиген [Klein et al. 2013; Муке и Нуссенцвейг, 2012; Mouquet et al. 2010; Scheid et al. 2011; Wu et al. 2011b]. Созревание аффинности и полиреактивность объясняются подтверждением связывания с высокоаффинным сайтом объединения против Env и вторым сайтом низкой аффинности на другой молекулярной структуре на ВИЧ-1 [Mouquet et al. 2010]. Методы инженерии антител, направленные на усиление кажущейся аффинности и эффективности нейтрализации, недавно были рассмотрены Mouquet и Nussenzweig [Mouquet and Nussenzweig, 2012].Биспецифические антитела против ВИЧ-1 (BiAbs), которые могут связываться бивалентно за счет одного плеча scFv, который связывается с gp120, и второго плеча с субъединицей gp41 gp160, показали усиленную нейтрализацию [Mouquet et al. 2012]. Таким образом, сконструированные антитела, содержащие разные сайты объединения, могут быть потенциальными нейтрализаторами и терапевтическими кандидатами против ВИЧ-1. Фактическая терапевтическая или профилактическая ценность in vivo также может быть измерена в ходе испытаний эффективности в будущем.
Резюме
Основной целью поиска эффективных мер по борьбе с ВИЧ / СПИДом является разработка действенной вакцины, которая может препятствовать новым приобретениям и лечению для уже инфицированных людей.Антиретровирусная терапия на основе низкомолекулярных препаратов против обратной транскриптазы принесла большие дивиденды и увеличила продолжительность жизни инфицированных людей; однако необходимо учитывать несколько ограничений. Циркулирующий вирус может развить устойчивость к терапии и восстановиться после прекращения терапии. Кроме того, побочные эффекты, связанные с противовирусной терапией, оборачиваются растущим экономическим бременем. Хотя в течение некоторого времени велись поиски вакцины против ВИЧ, разработке подходящего иммуногена препятствовали крайнее генетическое разнообразие, изменчивость белка оболочки и иммунологические ограничения, такие как созревание аффинности.В последние годы от ВИЧ-положительных пациентов было выделено несколько мощных bNAb, которые показали большой потенциал нейтрализации первичных вирусов, и их комбинация дала многообещающие результаты в качестве терапевтического вмешательства [Burton et al. , 2012b; Klein et al. 2012b]. Кроме того, было выделено много новых dAb, которые показали повышенную противовирусную активность. В то время как разработка соответствующих иммуногенов, вызывающих ответ на NAb, является высокоприоритетной задачей [Burton et al., 2012а; Haynes et al. 2012b], пассивный иммунитет, опосредованный комбинацией антител, может быть эффективным профилактическим средством против ВИЧ у людей с высоким риском и в то же время представляет собой альтернативу комбинированной терапии у инфицированных людей. Изменение свойств сконструированных антител, таких как улучшенная доступность утопленных эпитопов и усиленная доставка в лимфоидные и слизистые ткани, может повысить их эффективность. Включение таких антител в основной репертуар полноразмерных антител может усилить общий противовирусный эффект за счет сочетания эффекторной и нейтрализующей функций.Ни одна вакцина не обеспечила полной защиты от ВИЧ, но достигнут значительный прогресс и понимание того, как сильные антитела нейтрализуют вирус и каковы уязвимые эпитопы на оболочке ВИЧ-1. Однако менее ясно, как эти bNAb в конечном итоге развиваются у ВИЧ-положительных пациентов из-за чрезвычайно сложных путей активации и созревания репертуара антител. Более глубокое понимание важнейших особенностей и выделение более широких и эффективных антител могут объединиться, чтобы проложить путь к разработке эффективной терапии.
Сноски
Финансирование: Это исследование не получало специального гранта от какого-либо финансирующего агентства в государственном, коммерческом или некоммерческом секторах.
Заявление о конфликте интересов: Авторы заявляют об отсутствии конфликта интересов.
Информация для авторов
Раджеш Ринге, Медицинский колледж Вейля Корнельского университета, Нью-Йорк, Нью-Йорк, США.
Джаянта Бхаттачарья, Международная инициатива по вакцине против СПИДа (IAVI), Лаборатория THSTI-IAVI HVTR, Трансляционный институт науки и технологий здравоохранения (THSTI), Гургаон-122016, Харьяна, Индия.
Ссылки
- Алам С., Ляо Х., Деннисон С., Джегер Ф., Паркс Р., Анасти К. и др. (2011) Дифференциальная реактивность аллельных вариантов зародышевой линии широко нейтрализующего антитела ВИЧ-1 к промежуточной конформации слияния gp41. J Virol 85: 11725–11731 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Алам С., Морелли М., Деннисон С., Ляо Х., Чжан Р., Ся С. и др. (2009) Роль мембраны ВИЧ в нейтрализации двумя широко нейтрализующими антителами. Proc Natl Acad Sci U S A 106: 20234–20239 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Армбрустер К., Stiegler G., Vcelar B., Jager W., Koller U., Jilch R., et al. (2004) Пассивная иммунизация человеческим моноклональным антителом против ВИЧ-1 (hMAb) 4E10 и комбинацией hMAb 4E10 / 2F5 / 2G12. J Antimicrob Chemother 54: 915–920 [PubMed] [Google Scholar]
- Армбрустер К., Стиглер Г., Вцелар Б., Ягер В., Майкл Н., Веттер Н. и др. (2002) Фаза I испытания двух человеческих моноклональных антител (hMAb 2F5, 2G12) против ВИЧ-1. СПИД 16: 227–233 [PubMed] [Google Scholar]
- Астроном Р., Ли Х., Сканлан К., Пантофлет Р., Хуанг К., Уилсон И. и др. (2008) Гликоконъюгатный антиген, основанный на мотиве распознавания широко нейтрализующего антитела вируса иммунодефицита человека, 2G12, является иммуногенным, но вырабатывает антитела, неспособные связываться с собственными гликанами gp120. J Virol 82: 6359–6368 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Баден Л., Долин Р. (2012) Путь к эффективной вакцине против ВИЧ. N Engl J Med 366: 1343–1344 [PubMed] [Google Scholar]
- Балаш А., Чен Дж., Хонг К., Рао Д., Ян Л., Балтимор Д. (2012) Защита от ВИЧ-инфекции на основе антител с помощью векторной иммунопрофилактики. Природа 481: 81–84 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бар К., Цао К., Айер С., Деккер Дж., Янг Ю., Бонсиньори М. и др. (2012) Ранние нейтрализующие антитела с низким титром препятствуют репликации ВИЧ-1 и отбирают вирус. PLoS Pathog 8 (5): e1002721. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бинли Дж., Рин Т., Корбер Б., Цвик М., Ван М., Чаппи С. и др. (2004) Комплексный анализ перекрестной нейтрализации панели моноклональных антител против вируса иммунодефицита человека типа 1. J Virol 78: 13232–13252 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Берк Б., Барнетт С. (2007) Расширение нашего взгляда на защитные реакции антител против ВИЧ. Curr HIV Res 5: 625–641 [PubMed] [Google Scholar]
- Бертон Д., Ахмед Р., Баруш Д., Бутера С., Кротти С., Годзик А. и др. (2012a) План открытия вакцины против ВИЧ.Клеточный микроб-хозяин 12: 396–407 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бертон Д., Десрозье Р., Домс Р., Кофф В., Квонг П., Мур Дж. И др. (2004) Дизайн вакцины против ВИЧ и проблема нейтрализующих антител. Нат Иммунол 5: 233–236 [PubMed] [Google Scholar]
- Burton D., Poignard P., Stanfield R., Wilson I. (2012b) Широко нейтрализующие антитела открывают новые возможности для противодействия вирусам с высоким антигенным разнообразием. Наука 337: 183–186 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бертон Д., Пяти Дж., Кодури Р., Шарп С., Торнтон Г., Паррен П. и др. (1994) Эффективная нейтрализация первичных изолятов ВИЧ-1 рекомбинантным человеческим моноклональным антителом. Наука 266: 1024–1027 [PubMed] [Google Scholar]
- Бертон Д., Стэнфилд Р., Уилсон И. (2005) Антитело против ВИЧ в столкновении эволюционных титанов. Proc Natl Acad Sci U S A 102: 14943–14948 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Каларезе Д., Сканлан К., Цвик М., Дичонгкит С., Мимура Ю., Кунерт Р. и др.(2003) Обмен доменами антител — это иммунологическое решение распознавания углеводных кластеров. Наука 300: 2065–2071 [PubMed] [Google Scholar]
- Кардосо Р., Цвик М., Стэнфилд Р., Кунерт Р., Бинли Дж., Катингер Х. и др. (2005) Широко нейтрализующее антитело против ВИЧ 4E10 распознает спиральную конформацию высококонсервативного связанного с слиянием мотива в gp41. Иммунитет 22: 163–173 [PubMed] [Google Scholar]
- Casadevall A. (2002) Пассивное введение антител (немедленный иммунитет) как специфическая защита от биологического оружия.Emerg Infect Dis 8: 833–841 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cavacini L., Emes C., Power J., Duval M., Posner M. (1994) Влияние валентности антитела на взаимодействие с ВИЧ-1, экспрессируемым на клеточной поверхности, и нейтрализацию вируса. J Immunol 152: 2538–2545 [PubMed] [Google Scholar]
- Chamow S., Duliege A., Ammann A., Kahn J., Allen J., Eichberg J., et al. (1992) Иммуноадгезины CD4 в терапии против ВИЧ: новые разработки. Int J Cancer Suppl 7: 69–72 [PubMed] [Google Scholar]
- Чен В., Димитров Д. (2009) Человеческие моноклональные антитела и сконструированные домены антител как ингибиторы проникновения ВИЧ-1. Curr Opin ВИЧ СПИД 4: 112–117 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Чен В., Димитров Д. (2012) Терапевтические кандидаты на основе моноклональных антител против ВИЧ типа 1. Ретровирусы СПИДа Res Hum 28: 425–434 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Чен У., Ин Т., Димитров Д. (2013) Терапевтические кандидаты на основе антител против ВИЧ-1: значение для эрадикации вируса и дизайна вакцины.Мнение эксперта Biol Ther 13: 657–671 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Chen W., Zhu Z., Feng Y., Dimitrov D. (2008a) Антитела человеческого домена к консервативным стерически ограниченным областям на gp120 как исключительно мощные перекрестно-реактивные нейтрализаторы ВИЧ-1. Proc Natl Acad Sci U S A 105: 17121–17126 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Chen W., Zhu Z., Feng Y., Dimitrov D. (2010) Большая библиотека доменных антител человека, сочетающая разнообразие CDR3 тяжелой и легкой цепей.Мол Иммунол 47: 912–921 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Chen W., Zhu Z., Feng Y., Xiao X., Dimitrov D. (2008b) Конструирование большой библиотеки доменов человеческого антитела, отображаемой на фаге, с каркасом на основе недавно идентифицированного высокорастворимого стабильного вариабельного домена тяжелой цепи. Дж Мол Биол 382: 779–789 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Корти Д., Ланзавеккья А. (2013) Широко нейтрализующие противовирусные антитела. Анну Рев Иммунол 31: 705–742 [PubMed] [Google Scholar]
- Д’Суза М., Ливнат Д., Брадак Дж., Бриджес С. (1997) Оценка моноклональных антител к первичным изолятам вируса иммунодефицита человека типа 1 с помощью анализов нейтрализации: критерии эффективности для выбора антител-кандидатов для клинических испытаний. Рабочая группа по отбору антител группы клинических испытаний СПИДа. J заразить Дис 175: 1056–1062 [PubMed] [Google Scholar]
- Дэвис К., Грей Э., Мур П., Деккер Дж., Саломон А., Монтефиори Д. и др. (2009) Высокотитровые антитела к ВИЧ-1, специфичные к V3, с широкой реактивностью, но с низкой нейтрализующей способностью при острой инфекции и после вакцинации.Вирусология 387: 414–426 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Дормитцер П., Улмер Дж., Раппуоли Р. (2008) Дизайн антигена на основе структуры: стратегия для вакцин следующего поколения. Тенденции биотехнологии 26: 659–667 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Falkowska E., Ramos A., Feng Y., Zhou T., Moquin S., Walker L., et al. (2012) PGV04, антитело к участку связывания CD4 gp120 ВИЧ-1, является широким и мощным в нейтрализации, но не вызывает конформационных изменений, характерных для CD4.J Virol 86: 4394–4403 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Флинн Н., Фортал Д., Харро С., Джадсон Ф., Майер К., Пара М. (2005) Плацебо-контролируемое испытание фазы 3 рекомбинантной вакцины гликопротеина 120 для предотвращения инфекции ВИЧ-1. J заразить Дис 191: 654–665 [PubMed] [Google Scholar]
- Форсман А., Бейрнарт Э., Ааса-Чапман М., Хурелбек Б., Хиджази К., Ко В. и др. (2008) Фрагменты антител ламы с нейтрализующими свойствами вируса иммунодефицита человека типа 1 (ВИЧ-1) перекрестного подтипа и высокой аффинностью к gp120 ВИЧ-1.J Virol 82: 12069–12081 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Frey G., Peng H., Rits-Volloch S., Morelli M., Cheng Y., Chen B. (2008) Промежуточное состояние слияния gp41 ВИЧ-1, на которое нацелены широко нейтрализующие антитела. Proc Natl Acad Sci U S A 105: 3739–3744 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Гилберт П., Петерсон М., Фоллманн Д., Хадженс М., Фрэнсис Д., Гурвит М. и др. (2005) Корреляция между иммунологическими ответами на вакцину с рекомбинантным гликопротеином 120 и частотой инфицирования ВИЧ-1 в фазе 3 испытания превентивной вакцины против ВИЧ-1.J заразить Дис 191: 666–677 [PubMed] [Google Scholar]
- Гонг Р., Чен В., Димитров Д. (2012) Терапевтические препараты на основе антител против ВИЧ-1. BioDrugs 26: 143–162 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Грэм Б., Маскола Дж. (2005) Уроки неудач — подготовка к будущим испытаниям эффективности вакцины против ВИЧ-1. J заразить Дис 191: 647–649 [PubMed] [Google Scholar]
- Хейнс Б., Гилберт П., МакЭлрат М., Золла-Пазнер С., Томарас Г., Алам С. и др. (2012a) Иммунно-коррелятный анализ испытания эффективности вакцины против ВИЧ-1.N Engl J Med 366: 1275–1286 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Haynes B., Kelsoe G., Harrison S., Kepler T. (2012b) Дизайн иммуногена B-клеточного происхождения при разработке вакцины с ВИЧ-1 в качестве тематического исследования. Nat Biotechnol 30: 423–433 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Хейнс Б., Монтефиори Д. (2006) Стремление вызвать широко реактивные нейтрализующие ответы антител с кандидатами на вакцину против ВИЧ-1. Экспертные ревакцины 5: 579–595 [PubMed] [Google Scholar]
- Хессел А., Хангартнер Л., Хантер М., Хэвенит К., Бёрскенс Ф., Баккер Дж. И др. (2007) Fc-рецептор, но не связывание комплемента, важно для защиты антител от ВИЧ. Природа 449: 101–104 [PubMed] [Google Scholar]
- Hessell A., Rakasz E., Poignard P., Hangartner L., Landucci G., Forthal D., et al. (2009) Широко нейтрализующее человеческое антитело против ВИЧ 2G12 эффективно защищает от заражения SHIV слизистой оболочки даже при низких титрах нейтрализации сыворотки. PLoS Pathog 5 (5): e1000433. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Хинц А., Лютье Хулсик Д., Форсман А., Кох В., Белрхали Х., Горлани А. и др. (2010) Кристаллическая структура нейтрализующей ламы V (HH) D7 и ее способ взаимодействия с gp120 ВИЧ-1. PLoS One 5 (5): e10482. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Холт Л., Херринг К., Джесперс Л., Вулвен Б., Томлинсон И. (2003) Доменные антитела: белки для терапии. Тенденции биотехнологии 21: 484–490 [PubMed] [Google Scholar]
- Хуанг Дж., Офек Г., Лауб Л., Лаудер М., Дориа-Роуз Н., Лонго Н. и др.(2012) Широкая и мощная нейтрализация ВИЧ-1 человеческим антителом, специфичным к gp41. Природа 491: 406–412 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Huber M., von Wyl V., Ammann C., Kuster H., Stiegler G., Katinger H., et al. (2008) Сильная активность антител по нейтрализации вируса иммунодефицита человека и лизиса комплемента не обязательно связана. J Virol 82: 3834–3842 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Каусмалли Л., Ваален К., Лоберсли И., Хваттум Э., Berntsen G., Michaelsen T., et al. (2004) Нейтрализация человеческих антител к вирусу ветряной оспы (VZV), полученных из библиотеки рекомбинантных антител пациента VZV. Дж. Ген Вирол 85: 3493–3500 [PubMed] [Google Scholar]
- Klasse P., Sattentau Q. (2002) Занятие и механизм опосредованной антителами нейтрализации вирусов животных. Дж. Ген Вирол 83: 2091–2108 [PubMed] [Google Scholar]
- Кляйн Ф., Дискин Р., Шайд Дж., Гейблер К., Муке Х., Георгиев И. и др. (2013) Соматические мутации каркаса иммуноглобулина обычно необходимы для широкой и сильной нейтрализации ВИЧ-1.Клетка 153: 126–138 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Klein F., Gaebler C., Mouquet H., Sather D., Lehmann C., Scheid J., et al. (2012a) Широкая нейтрализация за счет комбинации антител, распознающих сайт связывания CD4, и нового конформационного эпитопа на белке оболочки ВИЧ-1. J Exp Med 209: 1469–1479 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Klein J., Gnanapragasam P., Galimidi R., Foglesong C., West A., Jr, Bjorkman P. (2009) Исследование вклада размера и авидности в механизмы нейтрализации антител b12 и 4E10 против ВИЧ.Proc Natl Acad Sci U S A 106: 7385–7390 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Klein F., Halper-Stromberg A., Horwitz J., Gruell H., Scheid J., Bournazos S. и др. (2012b) терапия ВИЧ комбинацией нейтрализующих антител широкого спектра действия у гуманизированных мышей. Природа 492: 118–122 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ко В., Стеффенсен С., Гонсалес-Пахуэло М., Хурелбеке Б., Горлани А., Шинол А. и др. (2010) Создание семейно-специфической фаговой библиотеки фрагментов одноцепочечных антител ламы, которые нейтрализуют ВИЧ-1.J Biol Chem 285: 19116–19124 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ковач Дж., Нколола Дж., Пенг Х., Чунг А., Перри Дж., Миллер К. и др. (2012) тример оболочки ВИЧ-1 вызывает более сильные нейтрализующие реакции антител, чем мономерный gp120. Proc Natl Acad Sci U S A 109: 12111–12116 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Кулькарни С., Лапедес А., Тан Х., Гнанакаран С., Дэниэлс М., Чжан М. и др. (2009) Сверхсложные детерминанты нейтрализации монофилетической линии недавно переданных клонов Env ВИЧ-1 подтипа C из Индии.Вирусология 385: 505–520 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Квонг П., Уилсон И. (2009) Антитела к ВИЧ-1 и гриппу: взгляд на антигены по-новому. Нат Иммунол 1: 573–578 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kwong P., Wyatt R., Robinson J., Sweet R., Sodroski J., Hendrickson W. (1998) Структура гликопротеина оболочки gp120 ВИЧ в комплексе с рецептором CD4 и нейтрализующим человеческим антителом. Природа 393: 648–659 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Лабрин А., Poignard P., Raja A., Zwick M., Delgado K., Franti M., et al. (2003) Доступ молекул антител к консервативному сайту связывания корецепторов на гликопротеине gp120 стерически ограничен для первичного вируса иммунодефицита человека типа 1. J Virol 77: 10557–10565 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Летвин Н. (2006) Прогресс и препятствия в разработке вакцины против СПИДа. Нат Рев Иммунол 6: 930–939 [PubMed] [Google Scholar]
- Манрике А., Русерт П., Джус Б., Фишер М., Кустер Х., Леманн К. и др. (2007) In vivo и in vitro избегают нейтрализующих антител 2G12, 2F5 и 4E10. J Virol 81: 8793–8808 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Мао Ю., Ван Л., Гу К., Хершхорн А., Сян С., Хаим Х. и др. (2012) Субъединичная организация мембраносвязанного тримерного гликопротеина оболочки ВИЧ-1. Нат Структ Мол Биол 19: 893–899 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Маскола Дж. (2003) Определение защитного ответа антител на ВИЧ-1.Курр Мол Мед 3: 209–216 [PubMed] [Google Scholar]
- Маскола Дж., Льюис М., Стиглер Г., Харрис Д., Ванкотт Т., Хейс Д. и др. (1999) Защита макак от патогенного вируса иммунодефицита обезьян / человека 89.6PD путем пассивного переноса нейтрализующих антител. J Virol 73: 4009–4018 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Маскола Дж., Лаудер М., Ванкотт Т., Сапан К., Ламберт Дж., Мюнц Л. и др. (1997) Мощная и синергетическая нейтрализация первичных изолятов вируса иммунодефицита человека (ВИЧ) типа 1 гипериммунным иммуноглобулином против ВИЧ в сочетании с моноклональными антителами 2F5 и 2G12.J Virol 71: 7198–7206 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Маскола Дж., Монтефиори Д. (2010) Роль антител в вакцинах против ВИЧ. Анну Рев Иммунол 28: 413–444 [PubMed] [Google Scholar]
- Маскола Дж., Стиглер Г., Ванкотт Т., Катингер Х., Карпентер К., Хэнсон К. и др. (2000) Защита макак от вагинальной передачи патогенного химерного вируса ВИЧ-1 / SIV путем пассивной инфузии нейтрализующих антител. Нат Мед 6: 207–210 [PubMed] [Google Scholar]
- Мац Дж., Кесслер П., Буше Дж., Комбес О., Рамос О., Барин Ф. и др. (2013) Прямой отбор широко нейтрализующих однодоменных антител, нацеленных на консервативные CD4 и сайты связывания корецепторов gp120 ВИЧ-1. J Virol 87: 1137–1149 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- McCoy L., Weiss R. (2013) Нейтрализующие антитела к ВИЧ-1, индуцированные иммунизацией. J Exp Med 210: 209–223 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Маклеллан Дж., Панчера М., Каррико К., Горман Дж., Жюльен Дж., Хаят Р. и др. (2011) Структура домена V1 / V2 gp120 ВИЧ-1 с широко нейтрализующим антителом PG9. Природа 480: 336–343 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- МакМайкл А. (2006) Вакцины против ВИЧ. Анну Рев Иммунол 24: 227–255 [PubMed] [Google Scholar]
- МакМайкл А., Ханке Т. (2002) В поисках вакцины от СПИДа: возможен ли подход CD8 + Т-клеток? Нат Рев Иммунол 2: 283–291 [PubMed] [Google Scholar]
- Молдт Б., Сибата-Кояма М., Ракаш Э., Шульц Н., Канда Ю., Данлоп Д. и др. (2012) Нефукозилированный вариант моноклонального антитела b12 против ВИЧ-1 обладает повышенной противовирусной активностью, опосредованной FcgammaRIIIa, in vitro, но не улучшает защиту от заражения SHIV слизистой оболочки у макак. J Virol 86: 6189–6196 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Молдт Б., Ракаш Э.Г., Шульц Н., Чан-Хуи П.Ю., Свидерек К., Вайсграу К.Л. и др. (2012) Высокоэффективная нейтрализация ВИЧ-специфических антител in vitro обеспечивает эффективную защиту от заражения SHIV слизистой оболочки in vivo.Proc Natl Acad Sci (США) 109: 18921–18925 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Монтефиори Д., Моррис Л., Феррари Г., Маскола Дж. (2007a) Нейтрализующие и другие противовирусные антитела при инфекции ВИЧ-1 и вакцинации. Curr Opin ВИЧ СПИД 2: 169–176 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Монтефиори Д., Саттентау К., Флорес Дж., Эспарза Дж., Маскола Дж. (2007b) Вакцины против ВИЧ-1 на основе антител: последние разработки и будущие направления. PLoS Med 4 (12): e348. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Мулар М., Phogat S., Shu Y., Labrijn A., Xiao X., Binley J., et al. (2002) Широко перекрестно-реактивный ВИЧ-1-нейтрализующий человеческий моноклональный Fab, выбранный для связывания с комплексами gp120-CD4-CCR5. Proc Natl Acad Sci U S A 99: 6913–6918 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Муке Х., Нуссенцвейг М. (2012) Полиреактивные антитела в адаптивных иммунных ответах на вирусы. Cell Mol Life Sci 69: 1435–1445 [PubMed] [Google Scholar]
- Муке Х., Шарф Л., Эйлер З., Лю Ю., Иден К., Шайд Дж., и другие. (2012) Распознавание N-гликанов сложного типа за счет мощных нейтрализующих антител к ВИЧ. Proc Natl Acad Sci U S A 109: E3268 – E3277 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Муке Х., Шейд Дж., Золлер М., Крогсгаард М., Отт Р., Шукаир С. и др. (2010) Полиреактивность увеличивает кажущуюся аффинность антител против ВИЧ за счет гетеролигирования. Природа 467: 591–595 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Офек Г., Танг М., Самбор А., Катингер Х., Маскола Дж., Wyatt R. и др. (2004) Структурный и механистический анализ антитела 2F5 против вируса иммунодефицита человека типа 1 в комплексе с его эпитопом gp41. J Virol 78: 10724–10737 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Панчера М., Маклеллан Дж., Ву Х., Чжу Дж., Чангела А., Шмидт С. и др. (2010) Кристаллическая структура PG16 и химерная диссекция с соматически родственным PG9: структурно-функциональный анализ двух четвертичных специфических антител, которые эффективно нейтрализуют ВИЧ-1. J Virol 84: 8098–8110 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Панталео Г., Куп Р. (2004) Корреляты иммунной защиты при ВИЧ-1-инфекции: что мы знаем, что мы не знаем, что мы должны знать. Нат Мед 10: 806–810 [PubMed] [Google Scholar]
- Пантофлет Р., Бертон Д. (2006) GP120: мишень для нейтрализации антител к ВИЧ-1. Анну Рев Иммунол 24: 739–769 [PubMed] [Google Scholar]
- Паррен П., Маркс П., Хесселл А., Лакей А., Харуз Дж., Ченг-Майер К. и др. (2001) Антитело защищает макак от вагинального заражения патогенным вирусом иммунодефицита обезьян / человека R5 на уровне сыворотки, обеспечивающем полную нейтрализацию in vitro.J Virol 75: 8340–8347 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Pejchal R., Doores K., Walker L., Khayat R., Huang P., Wang S. и др. (2011) Мощное нейтрализующее антитело широкого спектра действия распознает гликановый щит ВИЧ и проникает через него. Наука 334: 1097–1103 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Phogat S., Wyatt R. (2007) Рациональные модификации гликопротеинов оболочки ВИЧ-1 для дизайна иммуногена. Curr Pharm Des 13: 213–227 [PubMed] [Google Scholar]
- Плоткин С.(2008) Вакцины: корреляты иммунитета, вызванного вакциной. Clin Infect Dis 47: 401–409 [PubMed] [Google Scholar]
- Rerks-Ngarm S., Pitisuttithum P., Nitayaphan S., Kaewkungwal J., Chiu J., Paris R., et al. (2009) Вакцинация ALVAC и AIDSVAX для предотвращения заражения ВИЧ-1 в Таиланде. N Engl J Med 361: 2209–2220 [PubMed] [Google Scholar]
- Ringe R., Bhattacharya J. (2012) Ассоциация усиленной нейтрализации ВИЧ-1 с помощью одной замены Y681H в gp41 с повышенным взаимодействием gp120-CD4 и инфекционной активностью макрофагов.PLoS One 7 (5): e37157. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Roben P., Moore J., Thali M., Sodroski J., Barbas C., III, Burton D. (1994) Свойства распознавания панели рекомбинантных Fab-фрагментов человека на сайте связывания CD4 gp120, которые демонстрируют различные способности к нейтрализовать вирус иммунодефицита человека типа 1. J Virol 68: 4821–4828 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Scanlan C., Pantophlet R., Wormald M., Ollmann Saphire E., Stanfield R., Wilson I., и другие. (2002) Широко нейтрализующее антитело 2G12 против вируса иммунодефицита человека типа 1 распознает кластер альфа1-> 2 остатков маннозы на внешней поверхности gp120. J Virol 76: 7306–7321 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Сканлан К., Пантофлет Р., Вормолд М., Сапфир Э., Каларез Д., Стэнфилд Р. и др. (2003) Углеводный эпитоп нейтрализующего антитела 2G12 против ВИЧ-1. Adv Exp Med Biol 535: 205–218 [PubMed] [Google Scholar]
- Шайд Дж., Mouquet H., Feldhahn N., Seaman M., Velinzon K., Pietzsch J., et al. (2009) Широкое разнообразие нейтрализующих антител, выделенных из В-клеток памяти у ВИЧ-инфицированных людей. Природа 458: 636–640 [PubMed] [Google Scholar]
- Scheid J., Mouquet H., Ueberheide B., Diskin R., Klein F., Oliveira T. и др. (2011) Последовательность и структурная конвергенция широких и сильных антител к ВИЧ, имитирующих связывание CD4. Наука 333: 1633–1637 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Шиф В., Ban Y., Stamatatos L. (2009) Проблемы разработки структурной вакцины против ВИЧ. Curr Opin ВИЧ СПИД 4: 431–440 [PubMed] [Google Scholar]
- Шен X., Деннисон С., Лю П., Гао Ф., Джегер Ф., Монтефиори Д. и др. (2010) Длительная экспозиция проксимальной области мембраны gp41 ВИЧ-1 с заменой L669S. Proc Natl Acad Sci U S A 107: 5972–5977 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Симек М., Рида В., Придди Ф., Пунг П., Кэрроу Э., Лауфер Д. и др. (2009) Элитные нейтрализаторы вируса иммунодефицита человека типа 1: лица с широкой и сильной нейтрализующей активностью, идентифицированные с помощью высокопроизводительного анализа нейтрализации вместе с алгоритмом аналитического отбора.J Virol 83: 7337–7348 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Стамататос Л., Моррис Л., Бертон Д., Маскола Дж. (2009) Нейтрализующие антитела, образующиеся при естественной инфекции ВИЧ-1: хорошие новости для вакцины против ВИЧ-1? Нат Мед 15: 866–870 [PubMed] [Google Scholar]
- Стиглер Г., Кунерт Р., Пуртчер М., Вольбанк С., Фоглауэр Р., Штейндл Ф. и др. (2001) Мощное перекрестно-нейтрализующее человеческое моноклональное антитело против нового эпитопа на gp41 вируса иммунодефицита человека типа 1.Ретровирусы AIDS Res Hum 17: 1757–1765 [PubMed] [Google Scholar]
- Строкаппе Н., Шинол А., Ааса-Чапман М., Горлани А., Форсман Куигли А., Хулсик Д. и др. (2012) Фрагменты антител ламы, распознающие различные эпитопы CD4bs, нейтрализуют широкий спектр подтипов ВИЧ-1 A, B и C. PLoS One 7 (3): e33298. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Traunecker A., Lanzavecchia A., Karjalainen K. (1992) Янусин: новый молекулярный дизайн биспецифических реагентов. Int J Cancer Suppl 7: 51–52 [PubMed] [Google Scholar]
- Тркола А., Kuster H., Rusert P., Joos B., Fischer M., Leemann C., et al. (2005) Задержка рецидива ВИЧ-1 после прекращения антиретровирусной терапии за счет пассивной передачи нейтрализующих антител человека. Нат Мед 11: 615–622 [PubMed] [Google Scholar]
- Ванландшут П., Стортерлерс К., Бейрнарт Э., Ибанез Л., Шепенс Б., Депла Э. и др. (2011) Нанотела (R): новые боеприпасы для борьбы с вирусами. Противовирусный Res 92: 389–407 [PubMed] [Google Scholar]
- Визи Р., Шатток Р., Папа М., Кириджан Дж., Джонс Дж., Ху К. и др. (2003) Предотвращение передачи вируса макакам с помощью вагинального введения моноклональных антител к gp120 ВИЧ-1. Нат Мед 9: 343–346 [PubMed] [Google Scholar]
- Уокер Б., Бертон Д. (2008) К вакцине от СПИДа. Наука 320: 760–764 [PubMed] [Google Scholar]
- Walker L., Huber M., Doores K., Falkowska E., Pejchal R., Julien J., et al. (2011) Широкий охват нейтрализации ВИЧ множественными высокоактивными антителами. Природа 477: 466–470 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Уокер Л., Phogat S., Chan-Hui P., Wagner D., Phung P., Goss J., et al. (2009) Широкие и мощные нейтрализующие антитела от африканского донора раскрывают новую мишень для вакцины против ВИЧ-1. Наука 326: 285–289 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Уокер Л., Симек М., Придди Ф., Гач Дж., Вагнер Д., Цвик М. и др. (2010) Ограниченное количество специфичностей антител опосредует широкую и сильную нейтрализацию сыворотки у отдельных ВИЧ-1 инфицированных людей. PLoS Pathog 6 (8): e1001028. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Вэй X., Decker J., Wang S., Hui H., Kappes J., Wu X., et al. (2003) Нейтрализация антител и ускользание от ВИЧ-1. Природа 422: 307–312 [PubMed] [Google Scholar]
- West A., Jr, Galimidi R., Foglesong C., Gnanapragasam P., Klein J., Bjorkman P. (2010) Оценка архитектуры антител CD4-CD4i дает эффективные реагенты против вируса иммунодефицита человека с широкой перекрестной реакцией. J Virol 84: 261–269 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Вест А., младший, Галимиди Р., Гнанапрагасам П., Бьоркман П. (2012) Одноцепочечные анти-ВИЧ белки на основе Fv: потенциал и ограничения. J Virol 86: 195–202 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ву X., Чангела А., О’Делл С., Шмидт С., Панчера М., Янг Ю. и др. (2011a) Иммунотипы четвертичного сайта уязвимости к ВИЧ-1 и их распознавание антителами. J Virol 85: 4578–4585 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Wu X., Yang Z., Li Y., Hogerkorp C., Schief W., Seaman M., et al. (2010) Рациональный дизайн оболочки позволяет идентифицировать широко нейтрализующие человеческие моноклональные антитела к ВИЧ-1.Наука 329: 856–861 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- У X., Чжоу Т., Чжу Дж., Чжан Б., Георгиев И., Ван К. и др. (2011b) Сфокусированная эволюция нейтрализующих антител к ВИЧ-1, выявленная структурой и глубоким секвенированием. Наука 333: 1593–1602 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Wyatt R., Kwong P., Desjardins E., Sweet R., Robinson J., Hendrickson W., et al. (1998) Антигенная структура гликопротеина оболочки gp120 ВИЧ. Природа 393: 705–711 [PubMed] [Google Scholar]
- Чжан М., Шу Ю., Сидоров И., Димитров Д. (2004) Идентификация нового Fab моноклонального антитела человека к CD4i, которое нейтрализует первичные изоляты ВИЧ-1 из разных клад. Антивир Рес 61: 161–164 [PubMed] [Google Scholar]
- Чжоу Т., Георгиев И., Ву X., Ян З., Дай К., Финзи А. и др. (2010) Структурная основа широкой и мощной нейтрализации ВИЧ-1 антителом VRC01. Наука 329: 811–817 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Золла-Пазнер С. (2004) Идентификация эпитопов ВИЧ-1, индуцирующих защитные антитела.Нат Рев Иммунол 4: 199–210 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Золла-Пазнер С., Кардозо Т. (2010) Взаимосвязь между структурой и функцией вариабельных областей последовательности оболочки ВИЧ-1 позволяет переориентировать дизайн вакцины. Нат Рев Иммунол 10: 527–535 [Бесплатная статья PMC] [PubMed] [Google Scholar]
Старые проблемы в новых перспективах
Нейтрализующие антитела (NAbs) могут быть как достаточными, так и необходимыми для защиты от вирусных инфекций, хотя иногда они действуют согласованно с клеточным иммунитетом.Успешные вакцины против вирусов индуцируют NAb, но вакцины-кандидаты против некоторых основных вирусных патогенов, включая ВИЧ-1, не смогли вызвать сильные и эффективные такие реакции. Теории о том, как антитела нейтрализуют инфекционность вируса, были сформулированы и экспериментально проверены с 1930-х годов; и постоянно возникают споры о механистических и количественных основах нейтрализации. Растворимые версии нативных олигомерных вирусных белков, которые имитируют функциональные мишени нейтрализующих антител, теперь позволяют измерять соответствующие аффинности NAb.Таким образом, нейтрализующее заполнение вирионов можно оценить и связать с активностью NAb. Кроме того, кинетику и стехиометрию связывания NAb можно сравнить с нейтрализующей эффективностью. Недавнее фундаментальное открытие того, что внутриклеточный фактор TRIM21 определяет степень нейтрализации аденовируса, дало новые механистические и количественные идеи. Поскольку TRIM21 находится в цитоплазме, он не влияет на нейтрализацию вирусов в оболочке, но важно раскрыть диапазон его активности против «голых» вирусов.Эти разработки объединяют старые проблемы нейтрализации вирусов — механизм, стехиометрию, кинетику и эффективность — под неожиданными новыми углами.
1. Введение
Нейтрализующие антитела (NAbs) являются наилучшим коррелятом защиты от вирусной инфекции после вакцинации [1–8]. Точно так же они являются маркерами иммунитета против повторного заражения после того, как острая инфекция была устранена. Такой иммунитет может быть пожизненным [9–11]. Многие вакцины против вирусных инфекций являются хорошими индукторами защитных нейтрализующих реакций антител, но в области вирусной вакцинации остаются непреодолимые проблемы.Одна из проблем — это антигенная изменчивость. Антигенные мишени для нейтрализации антител к вирусу гриппа сильно различаются, поэтому новую вакцину необходимо готовить каждый новый сезон [7]. Вирус гепатита С (ВГС) и вирус иммунодефицита человека (ВИЧ) еще более изменчивы, и после многих лет исследований мы все еще не можем вызвать иммунные ответы, защищающие от них [7, 12–14].
Антитела являются продуктами адаптивного гуморального иммунного ответа; узнаваемые ими молекулы называются антигенами; молекулы, которые вызывают ответ антител, являются иммуногенами.Следовательно, некоторые белки, особенно чужеродные, являются как иммуногенами, индуцирующими выработку специфических антител против самих себя, так и антигенами, мишенями ответа. Другие молекулы, например, небольшие органические соединения, могут связываться с антителами с высокой специфичностью, не будучи способными вызывать эти антитела, за исключением случаев, когда они конъюгированы с более крупными носителями [15]. Такие небольшие молекулы называются гаптенами. Они иллюстрируют важное различие между иммуногенностью, способностью вызывать иммунный ответ, и антигенностью, способностью распознаваться иммунным ответом [15, 16].Более точным участком поверхности антигена, с которым непосредственно контактирует антитело, является эпитоп, а соответствующей поверхностью антитела, находящейся в прямом контакте с антигеном, является паратоп [17].
Некоторые успешные вакцины против вирусных инфекций основаны на ослабленных, но реплицирующихся вариантах патогенного вируса, например, вакцине против оспы и одной форме вакцины против полиомиелита. Ослабленные варианты вируса иммунодефицита обезьян (SIV), который тесно связан с ВИЧ, обеспечивают более сильную защиту, чем нереплицирующиеся экспериментальные вакцины против вируса дикого типа, но механизм защиты полностью не выяснен [18–20].В других случаях рекомбинантные белки, представляющие субъединицы вируса гепатита B (HBV) и вируса папилломы человека (HPV), вызывают сильную защиту [21, 22]. Вакцина против ВПЧ состоит из вирусоподобных частиц, которые могут обладать полезными свойствами как с антигенной, так и с иммуногенной точек зрения: они могут хорошо представлять нативные нейтрализующие эпитопы и восприниматься врожденной иммунной системой как молекулярные паттерны, ассоциированные с патогенами [23]. Но субъединичная иммунизация не смогла защитить от ВИЧ типа 1 (ВИЧ-1) [1, 7, 8, 14, 24].Только в клиническом испытании RV144, в котором сочетались вирусные белки, экспрессируемые из вектора канареек с рекомбинантными белками-бустами, наблюдалась некоторая умеренная защита. Но вакцина не вызвала NAb [8, 25, 26]. Поэтому ведется охота за другими антителами и иммунными реакциями, которые могли бы объяснить ограниченную защиту. Было описано множество различных противовирусных эффектов антител, которые не квалифицируются как нейтрализация [27, 28]. Это подводит нас к некоторым семантическим пояснениям.
2.Определение нейтрализации вирусов
Определения произвольны и не содержат более глубоких знаний, чем предлагаемое использование определенного термина [29]. Следовательно, единственная причина придерживаться строгого определения нейтрализации состоит в том, что оно может способствовать ясности и позволяет проводить полезные различия в области антивирусных исследований. Нейтрализация, как обсуждается здесь, определяется как снижение вирусной инфекционности за счет связывания антител с поверхностью вирусных частиц (вирионов), тем самым блокируя этап цикла репликации вируса, который предшествует кодируемой вирусами транскрипции или синтезу [30, 31] .Классически этот термин применялся только к антителам и фрагментам антител, Fab и F (ab ‘) 2, но позже его естественным образом распространили на однодоменные антигенсвязывающие рекомбинантные фрагменты и природные нанотела [32, 33]. Точно так же определение может быть расширено, чтобы охватить аналогичные активности растворимых форм вирусных рецепторов [34, 35], встречающихся в природе дефензинов или других молекул врожденной иммунной системы [36, 37]; его можно распространить на лектины, полученные из растений или растворимые рекомбинантные и гибридные формы рецепторов лектинов маннозы С-типа (MCLR) [38].Что касается небольших органических молекул, если о них говорят, что они нейтрализуют, следует уточнить, означает ли это, что они действуют путем связывания с поверхностью вириона [39–49]. В этом обзоре, однако, обсуждается только нейтрализация, опосредованная антителами.
Что не входит в определение? Антитело может связываться с зарождающимся вирионом, таким образом действуя на поздних этапах вирусного цикла, тем самым блокируя высвобождение вновь образованного вируса с поверхности инфицированной клетки. Примерами таких антител являются антитела, направленные к нейраминидазе на поверхности вируса гриппа [50].Фермент высвобождает потомство вируса путем переваривания фрагмента нейраминовой кислоты рецептора вируса. Позже мы увидим, почему такое антитело, хотя его антиген украшает поверхность вириона, не вмешивается в начало репликативного цикла и, следовательно, по определению, не нейтрализует.
Другой семантический момент заключается в том, что антитело к рецептору вируса на поверхности клетки может блокировать вирусную инфекцию, но не нейтрализует согласно определению: оно не связывается с вирионами, каждый из которых, диффундируя во внеклеточной среде. космос, потенциально был бы столь же заразным, как и в отсутствие антитела; вместо этого клетки-мишени станут невосприимчивыми.Следовательно, для ясности в этом особом случае следует использовать другой термин, например, антитело, блокирующее инфекцию.
Полезность строгого определения становится очевидной в исследованиях вакцин, когда мы задаемся вопросом, с помощью какого механизма индуцированные вакциной антитела защищают от инфекции. Антителозависимая клеточная цитотоксичность (ADCC) — это хорошо известный эффект, который требует наличия эффекторных клеток и противодействует вирусной инфекции, убивая вирус-продуцирующую клетку [27, 28]. Участвующие антитела могут быть нейтрализующими при тестировании в отсутствие эффекторных клеток (естественных клеток-киллеров), но часто это не так.И некоторые нейтрализующие антитела не относятся к тому изотипу, который опосредует этот эффект. Антитело-зависимое клеточно-опосредованное подавление вируса (ADCVI) имеет более сложную связь с нейтрализацией, которая может одновременно происходить в анализе, используемом для измерения эффекта. Но поскольку подавление зависит от эффекторных клеток, это не нейтрализация. Опять же, изотип антитела будет влиять на компонент ADCVI, который включает взаимодействия Fc-Fc-рецептора. Кроме того, расположение эпитопа на гемагглютинине вируса гриппа определяет, является ли антитело просто нейтрализующим или также способным опосредовать взаимодействия Fc-рецептора и, таким образом, ADCC и защиту in vivo на модели мыши [51].
ВИЧ-1 может передаваться напрямую от клетки к клетке через индуцированное вирусом соединение, известное как вирусологический синапс [52–55]. Однако удобно называть ингибирование передачи от клетки к клетке чем-то другим, кроме нейтрализации, даже если оно опосредуется NAb [56]. Причина в том, что не-NAb, включая антитела, которые мешают формированию синапса, связываясь с клеточными структурами, и антитела, которые противодействуют образованию или высвобождению вириона, могут блокировать этот способ переноса.Также следует отметить, что относительная эффективность нейтрализации и блокирования передачи от клетки к клетке различается между NAb [57]. Соблюдение этих различий вносит ясность.
Обычно нейтрализация измеряется в отсутствие комплемента, и определение, безусловно, можно сделать более строгим, сделав это критерием. Но может быть более полезным разрешить опосредованное комплементом усиление нейтрализации в качестве законной категории [58]; Тем не менее, полезно количественно оценить этот эффект [59, 60].Опять же, если факторы комплемента снижают вирусную инфекционность за счет связывания непосредственно с вирусной поверхностью, а не с антителами, уже находящимися в комплексе с вирионами [61], это также можно квалифицировать в рамках более широкого определения нейтрализации.
Одно из соображений биологической и медицинской важности состоит в том, что нейтрализация не должна приравнивать только то, что измеряется в анализах нейтрализации in vitro . Хотя нейтрализация должна быть измерена in vitro , мы можем плодотворно обсудить, как работает in vivo .Сделать вывод о том, что он отвечает за защиту in vivo , сложнее и требует использования антител без других эффекторных функций или экспериментальных моделей, в которых клеточно-зависимые механизмы нокаутируются другими способами [62–64]. И эффекты на поздних этапах вирусного цикла может быть труднее предотвратить в экспериментах in vivo : как всегда, гипотезы могут пройти только более строгие тесты, но никогда не будут подтверждены. Семантически, однако, не должно быть никаких препятствий для обсуждения нейтрализации, которая происходит как in vitro , так и in vivo .
Часть репликативного цикла, ограниченная определением, включает в себя множество различных шагов, некоторые из которых являются общими для всех вирусов, а другие специфичны для определенных групп. Чтобы заразить клетку, вирион должен прикрепиться к поверхности клетки. Это может происходить через вспомогательные факторы прикрепления или непосредственно через основной рецептор, используемый вирусом для проникновения; некоторые вирусы затем взаимодействуют со вторым рецептором или корецептором [65–79]. Голым вирусам необходимо проникнуть через клеточную мембрану, чтобы попасть в цитоплазму, или вместо этого они могут ввести свой геном через мембрану [80]; Оболочечные вирусы должны сливать свою оболочку с клеточной мембраной, чтобы переместить ядро и геном в цитоплазму [79, 81–85].Некоторые вирусы попадают в цитоплазму непосредственно с поверхности клетки; некоторые зависят от эндоцитоза для продуктивного входа, иногда потому, что пониженный pH в эндосоме запускает проникновение или слияние [86]. Если вирус проникает непосредственно с поверхности клетки, содержащая геном частица должна снова проникнуть через барьер, кортикальный цитоскелет, который представляет собой актиновую сетку с масками, иногда меньшими, чем голые вирионы или вирусные ядра [70, 87]. Проникая после эндоцитоза, вирус преодолевает это препятствие, доставляя капсид в более центральное место.После этих шагов ядро может потребоваться без покрытия или его миграция в определенные места в клетке до того, как произойдут события транскрипционного или трансляционного синтеза, за пределами которых любые ингибирующие эффекты по определению больше не представляют собой нейтрализацию.
3. Механизмы нейтрализации
Нейтрализация имеет много механистических аспектов: как связываются NAb, вызывают ли они конформационные изменения, необратимо ли инактивируют вирусные белки, которые опосредуют проникновение, и являются ли они наиболее эффективными против вирионов в суспензии или после вириона. прикрепление к клеткам.Однако здесь механизм нейтрализации относится конкретно к тому, какая ранняя стадия репликативного цикла вируса блокируется (рис. 1).
Если NAb препятствуют прикреплению вирионов к клеткам-мишеням, то это механизм нейтрализации. Если NAb блокируют необходимые рецепторные взаимодействия после прикрепления, это также является механизмом нейтрализации; так же мешает любой другой обязательный этап проникновения отдельных вирусов, такой как взаимодействие корецепторов, эндоцитоз, слияние или проникновение.Один из способов изучения нейтрализации после прикрепления (PAN) — позволить вирусу адсорбироваться на клетках при низкой температуре, не допускающей слияния или интернализации, а затем добавить NAb. Однако не следует делать вывод, что PAN не препятствует взаимодействию рецепторов: некоторые контакты рецепторов могут сначала устанавливаться при более низкой температуре, а затем NAb связывается и предотвращает дальнейшее необходимое рекрутирование рецепторов во время разминки [88, 89] . PAN может даже отменить привязку. Тем не менее, исследование емкости PAN может способствовать характеристике NAb.То, что NAb способно к PAN, тем не менее, не демонстрирует PAN как преобладающий механизм при более физиологических температурах, равно как и отсутствие PAN при низких температурах не исключает того, что это произойдет при более высоких температурах: индуцированное рецептором воздействие эпитопа на входе — опосредующим вирусным белкам может потребоваться температура, не блокирующая проникновение. Если, тем не менее, PAN обнаруживается, но менее эффективен, чем нейтрализация до прикрепления, этому может быть несколько объяснений. Во-первых, возможно, что, когда рецепторный контакт уже установлен, требуется более высокая занятость NAb на оставшихся рецептор-связывающих белках для остановки входа, чем на свободном вирионе.Во-вторых, в игру вступает кинетика: скорость связывания NAb должна быть выше, когда начался процесс присоединения-вхождения, чем когда NAb связывается в отсутствие клеток, и, следовательно, требуется более высокая концентрация NAb, если включено -скоростная константа такая же. В-третьих, PAN может частично действовать за счет конкуренции с рецепторами, тем самым обращая прикрепление, что может потребовать более высоких концентраций NAb, чем предотвращение, из-за валентности взаимодействий вирус-рецептор и усиления начального прикрепления за счет последующего рекрутирования рецептора.
Многие из этих механизмов подтверждены экспериментально для различных вирусов и для инфицирования клеток-мишеней в различных условиях. Например, было продемонстрировано, что NAb блокируют прикрепление риновируса к клеткам HeLa [90]. Сложнее обстоит дело с полиовирусом. Но более недавнее выяснение механизмов проникновения пикорнавирусов может пролить свет на то, какие именно необходимые шаги предотвращаются нейтрализацией и различаются ли механизмы нейтрализации внутри вирусного семейства [39–41, 65, 91, 92].В исследованиях нейтрализации поливирусов было обнаружено, что некоторые NAb не блокируют прикрепление, а частично блокируют эндоцитоз, индуцируют сдвиг pI в вирионе или предотвращают транскрипцию [58, 93–96]. Одно исследование показало, что среди NAb, инактивирующих непривязанный вирус, только бивалентно связывающееся меньшинство было способно к PAN; другие были сделаны способными к PAN посредством сшивания их частей Fc. Более того, нейтрализация совпадала с предотвращением структурных изменений в вирионе, содержащем РНК 135S, и геномной экструзии, которая обычно производит 80S пустые капсиды в процессе незафиксированной инфекции [97].Наконец, способность агрегировать частицы полиовируса была зарегистрирована как исключение среди NAb [97].
Агрегация вирионов антителом рассматривается как эффект, отличный от нейтрализации [58]. Но поскольку он снижает инфекционность, его можно квалифицировать как ограниченный нейтрализующий эффект, хотя и со сложной зависимостью от концентраций антител и вирионов. Обычно агрегация вирионов как функция концентрации антител описывает куполообразную кривую: при низких концентрациях антител перекрестное сшивание антигенов на отдельных вирионах благоприятно, при более высоких концентрациях вирионы соединяются мостиком, но когда эпитопы на вирионах приближаются к насыщению, сшивание больше не может происходить.Для измерения потери инфекционности может потребоваться количественный анализ: крупный агрегат будет диффундировать медленнее, чем вирионы, но может осаждаться на чувствительных клетках; меньше клеток может оказаться инфицированным, чем монодисперсная суспензия вирионов, но те, которые это делают, могут получить более высокую множественность инфекции. В количественном анализе инфекционности, то есть с подсчетом фокуса, это может дать отчетливое снижение инфекционности [98]; в анализах, основанных на продукции вирусных антигенов или активации репортерных генов, эффект может быть меньше или отсутствовать.Тем не менее, снижение инфекционности будет считаться нейтрализацией, потому что это связано с тем, как вирус не может достичь и проникнуть в свои клетки-мишени. In vivo агрегация может иметь различные эффекты в зависимости от доступности клеток-мишеней и требований к диффузии, но их фагоцитоз агрегатов, вспомогательное ингибирование, не связанное с самой нейтрализацией, может усиливать опосредованное антителами ингибирование.
Ситуация иная, когда антитела не агрегируют вирионы, а блокируют прикрепление и проникновение, конкурируя с рецепторными взаимодействиями вируса или иным образом противодействуя функции вирусных белков, которые опосредуют слияние или проникновение: не было бы основы для купола. форма кривой ингибирования.Тогда мы могли бы вместо этого предсказать, что чем выше уровень заселения такими антителами поверхности вирионов, тем меньше вероятность вирусной инфекции; в качестве альтернативы существует определенный порог занятости, выше которого инфекционность полностью устраняется. Эти соображения сформулированы в рамках теории оккупации для нейтрализации , которая имеет правдоподобные связи с блокировкой различных шагов прикрепления и входа в качестве нейтрализующих механизмов [31, 99, 100].
4.Теория нейтрализации занятости
Есть два типа научных гипотез: некоторые из них можно проверить; другие отвратительны. Теории нейтрализации присутствия существуют в разных версиях с различным потенциалом для прямого экспериментального тестирования. Сильные версии предполагают, что молекулы IgG из-за своей массы препятствуют функции белков, опосредующих прикрепление или проникновение вируса. Одной молекулы антитела на субъединицу вирусного белка или даже на олигомер может быть достаточно. Если бы для заражения требовалось определенное количество незанятых вирусных белков, то все вирионы с более высокой степенью занятости были бы нейтрализованы.Минимальная нейтрализующая занятость будет составлять порог нейтрализации. Пока все хорошо, но есть несколько сложностей.
Связывание с дефектными белками, обеспечивающими вход, на вирионах не имеет прямого отношения к нейтрализации (рис. 2). Антитела, которые могут связываться только с нефункциональными формами белков, опосредующих проникновение, могут никогда не нейтрализовать сами по себе [101–104], хотя они могут усиливать NAb, делая стерически блокирующую оболочку на вирионе более толстой.Но некоторые NAb могут связываться как с функциональными, так и с дефектными целями, и их дополнительная способность выполнять последние может не иметь отношения к нейтрализации: это может увеличивать их жадность и, таким образом, косвенно увеличивать занятость функциональных целей.
Как показано на рисунке 2, эффекты связывания с функциональными олигомерами, опосредующими вход, также являются сложными. Незанятые функциональные сайты могут нуждаться в кластеризации в одной области поверхности вириона для функционирования. Но они могут быть подвижными, так что достаточное количество может быть рекрутировано в комплекс входа вместе с рецепторами после первоначальной стыковки вириона с одной или слишком небольшим количеством рецепторных молекул [31, 84, 105–113].Однако, если эти вирусные белки неподвижны и неравномерно распределены по поверхности вириона, они могут иметь нейтрализующий эффект только тогда, когда NAb связываются в кластерах. Ожидается, что все эти осложнения смягчат очевидные пороги нейтрализации, даже если постулируется, что вирион либо полностью нейтрализован, либо нет [84, 105, 108–110].
Другая точка зрения заключается в том, что любое занятие антителами снижает склонность к инфицированию, и чем меньше незанятых функциональных молекул, опосредующих проникновение, тем ниже инфекционность.Такое пропорциональное соотношение, применяемое ко всему диапазону занятости, мы можем назвать инкрементальной моделью , в отличие от моделей, которые постулируют порог, который мы можем назвать предельным [105, 114]. Теперь обе модели могут быть сформулированы математически в их чистом виде, но сопоставление их с данными предполагает, что смешанные модели являются наиболее реалистичными [31, 84, 105–110]. Таким образом, может существовать абсолютный минимум тесного созвездия незанятых молекул, опосредующих проникновение. Но когда созвездие больше, чем необходимо, любое связывание антител по-прежнему снижает инфекционность.И если избыточность входных молекул огромна, это может быть незначительным в относительном выражении. Другими словами, нет ни строгой пропорциональности, ни резкого порога.
Одна из версий теории заселенности предполагает, что нейтрализация произойдет, когда NAb достигнет достаточно плотной оболочки на вирионах [99]. Эта теория покрытия имеет несколько различных значений. Это вызывает стерическое затруднение доступа не только к занятой молекуле, опосредующей вход, но также и к соседним.Он подразумевает линейную зависимость между площадью поверхности вириона и минимальным количеством молекул NAb, необходимых для нейтрализации. Эта линейность приблизительно сохраняется [99]. Это могло бы объяснить, почему антитела к нейраминидазе вируса гриппа не нейтрализуют [50, 115]: плотность этого антигена слишком мала; гемагглютинина на поверхности вируса в четыре раза больше, чем молекул нейраминидазы [99]. Однако некоторая степень нейтрализации является результатом перекрестного связывания антител, связанных с нейраминидазой, анти-Fc-антителами [116].Возможно, двухслойная оболочка из антител дает большее стерическое вмешательство.
Точно так же вирус бешенства не нейтрализуется антителом, которое распознает конформацию меньшинства его поверхностного гликопротеина. Но когда большинство из них превращается в антигенную конформацию за счет снижения pH или повышенной температуры, вирус нейтрализуется этим антителом [117]. Это серьезная поддержка версии теории занятости с покрытием. Аналогичным образом теория могла бы объяснить, почему антитела к некоторым клеточным антигенам-пассажирам, таким как ICAM-1 и MHC на поверхности вирионов ВИЧ или SIV, действительно нейтрализуют, хотя и в значительной степени зависимым от комплемента образом [118–120]: антигена так много. что его покрытие частично затрудняет доступ к нескольким тримерам гликопротеина оболочки.
Если стерическое вмешательство может происходить межмолекулярно, блокирование рецептор-связывающего сайта связыванием молекулы IgG в другом месте того же вирусного белка, опосредующего проникновение, является еще более вероятным. NAb, которые не связываются с основным сайтом связывания рецептора на Env ВИЧ-1, по-видимому, блокируют взаимодействия с рецепторами: если в клетках-мишенях отсутствуют вспомогательные факторы прикрепления, связывания антител где-либо на рецепторсвязывающей субъединице Env достаточно, чтобы блокировать прикрепление вируса к клеткам. то есть, когда прикрепление опосредуется первичным рецептором.Таким образом, в этих условиях стерические или прямые препятствия могут помешать первой стадии репликации [121]. Тем не менее, имеет значение, на какой субъединице расположен эпитоп. Некоторые NAb направлены на эпитопы, расположенные близко к вирусной мембране в трансмембранном белке. Они не мешают прикреплению к клеткам, у которых отсутствуют вспомогательные факторы прикрепления, и, следовательно, по-видимому, не мешают связыванию рецепторов [121]. В заключение, связывание близко к сайту связывания рецептора может потребоваться для непрямого блокирования взаимодействий рецептора.
Теория также подразумевает, что активность NAb тесно коррелирует с их сродством к нативной олигомерной форме Env на вирусной поверхности, и это также в значительной степени справедливо для ВИЧ-1 [100, 103]. Опять же, исключением могут быть NAb, направленные в проксимальную область мембраны в трансмембранном белке [122, 123]. Эти эпитопы могут стать экспонированными или полностью антигенными только после того, как рецепторные взаимодействия вызвали некоторые конформационные изменения в Env. Аналогично, эффективность связывания пептидных ингибиторов с трансмембранным белком коррелирует не со сродством к белку, а с константой скорости [124].Таким образом, сродство NAb к природным молекулам, опосредующим проникновение, в целом хорошо коррелирует с эффективностью нейтрализации, но в некоторых случаях сродство к индуцированной рецептором временной форме вирусного белка определяет нейтрализацию.
Теория покрытия, кроме того, подразумевает, что, хотя связывание NAb может вызывать конформационные изменения, такие эффекты не должны быть необходимы для их нейтрализующей способности. Это предсказание противоречит давней традиции исследований, особенно пикорнавирусов [58, 94–96].Новые данные действительно указывают на отсутствие корреляции между нейтрализацией и способностью вызывать конформационные изменения у риновирусов [125], что указывает на то, что конформационные изменения являются эпифеноменами, сопровождающими связывание одних NAb, но не других. Мощный NAb против ВИЧ-1, направленный на эпитоп, отдельный от сайта связывания CD4, по-видимому, блокирует взаимодействия CD4 аллостерически, что обнаружено с помощью растворимых форм тримера Env [126]; Необходимы ли такие конформационные изменения также на уровне вирионов, покрытых NAb, будет труднее исследовать.Возможно, аллостерическое вмешательство во взаимодействия рецепторов может добавить тормозящую силу к простому стерическому затруднению.
Еще одно следствие теории занятости в целом и ее версии покрытия в частности заключается в том, что не-NAb не должно быть возможности блокировать NAb. Хотя исследования вируса японского энцефалита предполагают такое вмешательство [127], было показано, что не-NAb, направленные на белок Env ВИЧ-1, не блокируют нейтрализацию NAb, даже если они направлены на перекрывающиеся эпитопы.Какие объяснения? Теория занятости подразумевает, что все NAb, которые связываются с функциональными вирусными белками, опосредующими проникновение, должны нейтрализоваться. В самом деле, не-NAb, которые связываются с эпитопами, перекрывающими известные нейтрализующие эпитопы на субъединице Env ВИЧ-1 gp120, как было показано, не связываются с gp120 в его тримерном нативном контексте [102, 103].
Для гемагглютинина гриппа и капсидных белков полиовируса также описаны перекрывающиеся эпитопы NAb и не-NAb, но ключевой вопрос заключается в том, действительно ли не-NAb связываются с нативным белком на поверхности инфекционных вирионов [93, 128] .Интересно, что также было обнаружено, что связывающий маннозу лектин может блокировать нейтрализацию NAb, направленную на эпитоп маннозы на Env ВИЧ-1 [129]. Если когда-либо произойдет блокирование нейтрализации не-NAb, объяснение, возможно, придется искать в конформационных изменениях, которые NAb дает, а не-NAb нет, и наоборот . Фокус снова сместится к тому, способствуют ли конкретные конформационные изменения антигена нейтрализации. Таким образом, не-NAb будет защищать антиген до тех пор, пока он не сможет конкурировать с рецепторами.Но почему бы ему не посоревноваться с НБ?
Совсем недавно проведенное исследование показывает, как могут действовать не-NAb, блокирующие нейтрализацию, хотя в нем сообщается об эффектах в более серой области, связанной с изменениями активности и эффективности. Определенные мутации в Env ВИЧ-1 снижают степень нейтрализации широко активным и мощным NAb (10E8 [122]), направленным на проксимальный к мембране эпитоп в трансмембранном белке. Хотя эти мутации, таким образом, не превращают NAb в не-NAb, можно изучить, как связывание с мутантными вирионами влияет на их чувствительность к другим NAb, то есть как остаточно инфекционный вирус в комплексе с этим все еще частично активным NAb. нейтрализуется [123].NAb усиливает нейтрализацию одних антител к другим эпитопам в gp41, но снижает нейтрализацию другими, в том числе направленными на соседние эпитопы, а также антителами, специфичными для сайта связывания CD4. Стехиометрия связывания NAb с мутантными тримерами ниже, чем у Env дикого типа: вместо трех связываются два паратопа [123]. Эта низкая степень связывания стабилизирует тример и может иметь отдаленные аллостерические эффекты, так что связывание других NAb облегчается или затрудняется. Следовательно, этот интригующий случай может однозначно показать, что слабо нейтрализующие антитела могут противодействовать, а не добавлять к действию более сильных.Если аллостерический механизм включает снижение сродства к удаленным затронутым эпитопам, это скорее подтверждает теорию занятости, чем подрывает ее.
Также имеет отношение к теории занятости то, что Abs может захватывать вирионы. Как NAb, так и не-NAb могут захватывать вирионы ВИЧ-1, но NAb преимущественно захватывают инфекционные вирионы [130]. Как это ни парадоксально, но не-NAb, которые не блокируют нейтрализацию, блокируют захват [131]. Это поднимает вопрос, почему связывание NAb с функциональными тримерами само по себе не опосредует захват вирионов.Возможно, некоторое связывание NAb с gp120 заставляет его диссоциировать от трансмембранного белка gp41, который закрепляет его на вирусной мембране [132, 133]. Опять же, если бы все связывание NAb имело такой эффект, было бы трудно объяснить предпочтительный захват инфекционных вирионов NAb.
Как упоминалось, в случае ВИЧ-1 некоторые эпитопы нейтрализации находятся в трансмембранном белке, но другие эпитопы там блокируются gp120; и хотя последние являются иммуногенными, направленные к ним антитела не нейтрализуют, поскольку они могут связываться с нефункциональными культями тримеров только после выделения gp120 [103, 134, 135].Все это согласуется с основными принципами размещения. Даже усиленное связывание NAb во время временного воздействия эпитопов вблизи мембраны согласуется с теорией. Хотя эти эпитопы присутствуют на функциональных тримерах, они обладают лишь слабой антигенностью в нативной форме шипа Env.
Несколько NAb для вирусов в оболочке блокируют поздние стадии процесса проникновения, то есть слияние вируса с клеточной мембраной. Причем этот блок не обязательно должен возникать на поверхности клетки.Если связывание антител позволяет до некоторой степени взаимодействовать с рецепторами, вирус может подвергнуться эндоцитозу, и слияние, которое в некоторых случаях запускается снижением pH в эндосоме, задерживается, а затем вирус направляется в сторону разрушения лизосомы до того, как он слился. и вытеснил его ядро и геном в цитоплазму. Этот сценарий был проиллюстрирован для вируса Западного Нила [136].
ВИЧ-1 может также зависеть от интернализации для полного слияния [83–85, 137] и, следовательно, может подвергаться нейтрализации антителами, которые разрешают эндоцитоз вириона, но препятствуют поздним стадиям слияния.Мы могли бы назвать это внутриклеточной нейтрализацией с тем важным отличием, что блокируется вход — слияние, предшествующее перемещению ядра в цитоплазму. Но определение нейтрализации, как обрисовано в общих чертах вначале, позволит блокировать несколько более поздних этапов, а именно, на ранней постентрийной стадии, в цитоплазме, до транскрипции вирусного генома или трансляции вирусных продуктов. Встречаются ли когда-нибудь такие механизмы?
5. Голая правда о постентарной нейтрализации
Постентарные механизмы нейтрализации неоднократно заявлялись о пикорнавирусах и вирусах гриппа [138, 139].Но окончательных доказательств не хватало, и многие вирусологи могли рассматривать нейтрализацию на практике как синоним опосредованного антителами ингибирования прикрепления и проникновения. Затем был открыт новый внутрицитоплазматический механизм нейтрализации [140–142]. Изучая аденовирус, Джеймс и его коллеги обнаружили, что его нейтрализация во многом зависит от присутствия в клетках-мишенях белка 21, содержащего трехкомпонентные мотивы (TRIM21). TRIM21 расположен в цитоплазме, имеет сильное сродство к IgG и убиквитинирует комплексы антитело-антиген, которые он захватывает, направляя их для разрушения протеасомой [141, 142].
Один удивительный аспект того, как работает внутрицитоплазматическая нейтрализация, заключается в том, что частица аденовируса в виде интактного комплекса с NAb должна перемещаться через везикулярную мембрану в цитоплазматический компартмент. По крайней мере, одним из важных путей проникновения аденовирусов является интернализация через макропиноцитоз с последующим проникновением через везикулярную мембрану. Следовательно, пока макропиноцитарная везикула остается интактной и капсид, лигированный NAb, не проникает оттуда, TRIM21 не получает доступа к комплексу.Только после проникновения можно установить контакт NAb-TRIM21. Вполне вероятно, что одной молекулы NAb может быть достаточно для нацеливания комплекса на TRIM21, но также кажется возможным, что нацеливание будет усилено за счет большего количества связанных NAb [143]. Эти соображения также вызывают вопросы о том, сколько молекул NAb может проходить через мембрану вместе с вирусной частицей; одна из гипотез заключается в том, что высокая степень занятости NAb предотвращает проникновение, блокируя прикрепление, взаимодействия рецепторов или более позднюю стадию проникновения, но что TRIM21 действует как страховочная сетка, позволяя нейтрализовать вирионы с низкой занятостью NAb.Это, однако, означало бы существенную TRIM21-независимую нейтрализацию при самых высоких концентрациях NAb, чего, по-видимому, не происходит [143]. Эти новые проблемы будут дополнительно исследованы при анализе стехиометрии и эффективности нейтрализации ниже.
Насколько общим может быть этот механизм? Казалось бы, голые вирусы строго ограничиваются. Поскольку при слиянии вирусов в оболочке, NAb, связанные с вирусными поверхностными белками, не получают доступа к цитоплазме. И хотя капсидные белки вирусов в оболочке вызывают сильные реакции антител во время инфекции и при экспрессии из вакцинных векторов (см., Например, e.g., [144–146]), эти антитела не могут связываться со своими антигенами, если вирион не поврежден. Им придется самостоятельно перемещаться в цитоплазму. В самом деле, открытие механизма TRIM21, кажется, создает новую дихотомию нейтрализации вируса, механистическое разделение между потенциальными эффектами NAb на вирусы без оболочки и вирусы в оболочке. Но тогда механизм TRIM21 может применяться не ко всем «голым» вирусам. Если пикорнавирус вводит свой геном в цитоплазму, а капсид, следовательно, никогда не проникает [80, 97], результатом будет отсутствие комплексов антитело-капсид в цитоплазме и, следовательно, отсутствие мишеней для TRIM21.
Некоторые голые вирусы и капсулы вирусов в оболочке должны быть без покрытия после входа для продолжения репликации. Поэтому нетрудно предположить, что вирус мог использовать убиквитинирование с помощью TRIM21 и других факторов, чтобы облегчить этот шаг. Если деградация голого вириона в комплексе с NAb происходит слишком медленно, чтобы предотвратить выход генома на следующий этап репликации, или если капсид оболочечного вируса убиквитинируется посредством альтернативного взаимодействия, вирус может получить пользу от клеточной помощи его непокрытие.Но ядро ВИЧ-1 разрушается протеасомой в значительной степени при незащищенной инфекции: деградация вызывает чистую потерю инфекционности [147]. И, по крайней мере, в случае аденовируса нейтрализующий эффект, кажется, преобладает над любым потенциальным преимуществом вируса.
TRIM21-зависимая нейтрализация также влияет на то, могут ли некоторые антитела блокировать нейтрализацию. Если IgA и IgM, специфичные для нейтрализующих эпитопов, перекрывающих эпитопы IgG NAb, были неспособны лигировать TRIM21, они, вероятно, были бы способны блокировать TRIM21-зависимую нейтрализацию IgG, при условии, что они обладают достаточно высокой аффинностью и присутствуют в достаточных концентрациях.Но данные свидетельствуют о том, что эти классы Ab, как и IgG, могут взаимодействовать с TRIM21 [140, 142]. Эта способность объясняет их неспособность блокировать этот механизм нейтрализации, что сильно отличается от объяснения того, почему антитела редко блокируют нейтрализацию вирусов в оболочке.
Не может ли аналогичный механизм работать против вируса в оболочке? Один из эффектов, который может быть наиболее близким, — это связывание комплексов NAb-вирион с рецепторами Fc с последующим эндоцитозом и, в конечном итоге, лизосомной деградацией вируса.Этот зависимый от антител перенос вируса можно квалифицировать как нейтрализацию согласно определению. В случае ВИЧ-1, например, наблюдалась интернализация в зависимости от низкой занятости NAb и последующая потеря инфекционности, хотя и не классифицируемая как нейтрализация, возможно, потому, что эффект был слишком слабым по сравнению с обычной нейтрализацией [148]. Но следует отметить, что этот механизм будет препятствовать проникновению: жизнеспособные ядра, содержащие геном, не попадут в цитоплазму. Путь к лизосомной деградации может быть неэффективным, потому что он вытесняется продуктивным входом, который может происходить через эндосомальную мембрану.В самом деле, это может быть обычным местом продуктивного проникновения ВИЧ-1 [84, 85, 137]. Вероятно, антитело в комплексе с рецептором Fc будет блокировать взаимодействия фузогенного Env-рецептора за счет стерических затруднений и, таким образом, задерживать слияние до тех пор, пока не начнется деградация в лизосомном компартменте. Тем не менее, для этого может потребоваться более низкая заселенность вирионов молекулами антител, чем при обычном блокировке входа, так же как и гораздо более мощный механизм TRIM21. Оболочечные вирусы между собой также представляют собой противоположные примеры: низкая степень заселенности флавивирусов может опосредовать повышение инфекционности, тогда как высокая степень заселения теми же антителами вызывает нейтрализацию [81, 149–155].Мы вернемся к этим соображениям при более подробном обсуждении стехиометрии ниже.
TRIM21-зависимый постентрийный механизм нейтрализации и ограниченные по занятости механизмы блокировки входа имеют совершенно разные значения для классических и недавно изученных аспектов нейтрализации, и их необходимо постоянно противопоставлять. В качестве иллюстрации теория занятости может объяснить отсутствие нейтрализации антителами, даже если они связываются с инфекционными вирионами: они могут достигать слишком низких уровней занятости или связываться только с сайтами, которые не функционируют при входе.Напротив, TRIM21-зависимый механизм не может объяснить такое отсутствие нейтрализации, пока TRIM21 распознает Fc-часть антитела. Для убиквитинирования, по-видимому, не требуются пороговые уровни занятости антителами, хотя вполне вероятно, что чем больше связывается антитело, тем сильнее и быстрее будет деградация капсида. TRIM21 также не будет различать антитела, связанные с функциональными молекулами, опосредующими проникновение, и другими антигенами. Возможно, некоторые антитела, связанные с капсидами, не могут попасть в цитоплазму; они диссоциировали бы и позволили несвязанному капсиду проникнуть, иначе антитело нейтрализовалось бы на более ранней стадии.Ясно, что прорыв TRIM21 не только многое объясняет, но и поднимает новые интригующие вопросы.
В контексте нового механизма это провокационное наблюдение, что генетическая недостаточность антител, по-видимому, предрасполагает к большей уязвимости к заражению голыми вирусами, чем вирусами в оболочке [142]. Распространено ли постентрийное ингибирование среди «голых» вирусов? Обеспечивает ли он страховочную сетку или представляет собой главную линию защиты? Или вирусы в оболочке более уязвимы для клеточного иммунитета в дополнение к ответам антител?
Эти вопросы здесь придется оставить без ответа, но механизм, зависящий от TRIM21, имеет интригующие связи с количественным аспектом нейтрализации, к которому мы не обращаемся.
6. Кинетика нейтрализации
Причина этого раздела двоякая: во-первых, чтобы попытаться устранить ущерб, нанесенный ошибочными выводами из кинетики самой реакции нейтрализации; и во-вторых, чтобы прояснить, чем этот аспект отличается от других, менее тривиальных, кинетических аспектов нейтрализации или связан с ними.
Чтобы рассматривать нейтрализацию вирионов в суспензии антителами в растворе как химическую реакцию, требуются некоторые сомнительные предположения. Таким образом, следует предположить, что в какой-то момент события связывания между реагентами превращают вирионы из инфекционных в неинфекционные, эффект такой же черный и белый, как образование или разрыв ковалентной связи.Как уже упоминалось, вирионы (хотя здесь виды вирусов, вероятно, существенно различаются) могут иметь широкий спектр склонностей к заражению, и нейтрализация может быть сдвигом в таких склонностях, то есть не эффектом «все или ничего». Но если мы примем предпосылку, что нейтрализация — это полная потеря инфекционности отдельного вириона, мы могли бы попытаться узнать молекулярность реакции нейтрализации, то есть, сколько молекул антител должно связываться для достижения нейтрализации.Молекулярность связана с порядком реакции, который является кинетическим понятием, но не может быть выведен из него. Порядок может быть определен эмпирически, если концентрацию свободных реагентов контролировать вместе с расходами. Таким образом, скорость реакции нейтрализации будет равна где — константа скорости, [вирус] и [ Ab ] — концентрации свободных реагентов (которые, следовательно, оба уменьшаются), и порядок реакции по концентрации антител, которая не обязательно должна быть целым числом.Поскольку уже предполагается, что вирионы действуют в одиночку, реакция первого порядка по концентрации вирионов (нейтрализация посредством агрегации будет сложной в этой схеме). Что еще предстоит определить. В попытках сделать вывод о молекулярности реакции отслеживали скорость нейтрализации с течением времени как функцию концентрации антител. Наблюдались значения, близкие к 1, и был сделан вывод, что одна молекула антитела инактивирует один вирион [156–158]. Ошибка состоит в том, что концентрация свободных антител не регистрируется и ее изменения невозможно измерить из-за огромного молярного избытка антител над вирусом.Это классическая ситуация кинетики псевдопервого порядка. Поскольку [Ab] существенно не меняется, верно следующее приближение: где [ Ab ] — константа скорости псевдопервого порядка для концентрации вириона. Данные ничего не говорят о порядке концентрации антител. Второй недостаток — это предположение, что эпитопы можно разделить на критические и некритические, но каким-то образом только связывание с критическими эпитопами будет отражаться на кинетике нейтрализации.Это не имеет смысла: если связывание является случайным и равным сродством к двум типам эпитопов, определенное количество некритических эпитопов будет связано с той же занятостью, что и критические эпитопы. Следовательно, порядок был ошибочно получен из кинетики, и так называемая молекулярность одиночного попадания была ошибочно выведена из порядка; чтобы скрыть эти неоправданные скачки, иногда используется термин кинетика одиночного удара . Следует отметить, что в исходной статье изучались один «голый» и один вирус в оболочке и наблюдалась аналогичная кинетика для них.Оболочечный вирус представлял собой вирус западного конского энцефалита [159]. Голым вирусом был полиовирус. Могут ли эти старые предположения иметь какое-либо отношение к новому открытию эффекта TRIM21, который потенциально может применяться к нескольким голым вирусам?
Нет, это было бы ложным совпадением. TRIM21 может и не способствовать нейтрализации полиовируса, хотя вирус является «голым», если его геном вытесняется трансмембранной инъекцией, а не транслокацией капсида вместе с любым связанным NAb в цитоплазму.Следовательно, если выяснится, что TRIM21 позволяет единичным антителам опосредовать нейтрализацию некоторых вирусов, этот результат станет важным достижением в знаниях. Но это не пролило бы света на какие-либо неожиданные предполагаемые явления однократного попадания, основанные на кинетике нейтрализации, потому что таких явлений не было, только ошибочные интерпретации.
В более ранней статье правильно описаны последствия огромного молярного избытка антител над вирионами (и над эпитопами): поскольку концентрация вирионов варьируется в широком диапазоне, пропорция, которая нейтрализуется фиксированной концентрацией антител, остается постоянной.Это соотношение было названо законом процента [160]. Эти наблюдения иллюстрируют основу кинетики псевдопервого порядка: доля антител, потерянных в результате связывания, незначительна.
Другой подход был использован для изучения того, сколько молекул антител должно связываться, прежде чем произойдет нейтрализация. Таким образом, на кривой нейтрализации с течением времени первый сегмент кривой был тщательно исследован на предмет признаков любого плеча. Если наблюдалось плечо, это означало, что должно было связываться более одного антитела, прежде чем инфекционность была устранена для любого вириона.Иногда такое плечо наблюдалось, иногда нет [161]. Сложность заключается в получении необходимой точности данных для достаточно ранней части кривой. И даже при самых точных таких данных толкования не бесспорны. Сам метод остановки реакции нейтрализации, а именно быстрое разбавление, является неудовлетворительным, поскольку делает результаты зависимыми от степени необратимости. И если NAb используется при концентрации ниже его, может быть плечо даже при очень низкой занятости, потому что скорость связывания будет очень низкой.Кроме того, одна NAb, связанная с самыми маленькими вирусами, будет представлять собой более высокую занятость, чем несколько NAb, связанных с более крупными вирусами. Таким образом, пороги одиночного и множественного попадания могут выглядеть одинаково.
Другой возможной причиной плеча является то, что антитело должно вызывать изменения вирусного антигена, которые происходят медленнее, чем связывание; отсутствие плеча может означать, что вирионы теряют некоторую склонность к инфицированию с помощью первых событий связывания, но не нейтрализуются полностью, как предполагает гипотеза однократного попадания; данные не позволят отличить частичную вмятину в инфекционности многих от полной потери некоторых.Рассуждения здесь аналогичны различию между инкрементальной и лиминальной моделями. Только высокий и однородный порог может отображаться как широкое плечо на кинетической кривой.
Ошибочные интерпретации одиночного попадания привели к поиску таких механизмов, как инактивация сигналов от связанных NAb внутрь вириона для вирусов в оболочке и конформационные сдвиги во всем капсиде для «голых» вирусов. В общем, охота велась за постентрийными механизмами. Ибо блок рецепторных взаимодействий и проникновения казался труднее объяснимым молекулярностью однократного попадания, по крайней мере, в случае вирусов в оболочке.
Как практическое следствие гипотезы об однократном попадании, нейтрализующая занятость будет низкой, за исключением случаев, когда предполагается, что вирус имеет только единственную релевантную молекулу антигена [84, 105, 106, 111]. И чем ниже требуемая занятость, тем легче будет задача индуцирования титров защитного связывания NAb путем вакцинации: титры и заполненность являются продуктами аффинностей и концентраций антител. Неправильное толкование может ввести в заблуждение исследования вакцины.
7.Кинетика связывания: пример ВИЧ-1 Env
Кинетика связывания NAb и других антител с поверхностными белками вирусов более изучена, чем кинетика нейтрализации. Одним из методов, который позволяет проводить кинетические измерения, является поверхностный плазмонный резонанс (SPR) [162–167]. С помощью этого метода антиген или антитело можно иммобилизовать на сенсорном чипе. Если антиген иммобилизован, раствор антитела протекает через него. Связывание вызывает изменение угла отражения поляризованного света, который отслеживается и преобразуется в резонансный сигнал, пропорциональный массе связавшегося белка.Этот метод использовался для измерения связывания антител с мишенями нейтрализации различных вирусов, включая гликопротеины Env ВИЧ-1. Моделирование связывания при различных концентрациях дает константу скорости включения, константу скорости отклонения, и их соотношение, константу диссоциации, обратную меру аффинности; кроме того, поскольку достигается максимальное равновесное связывание и его можно экстраполировать, можно также оценить стехиометрию связывания. Такие исследования связывания с конформационно гибкими олигомерами гликопротеинов вирусной оболочки, которые опосредуют проникновение в чувствительные клетки, в последнее время стали более актуальными для нейтрализации за счет улучшения мимикрии нативных антигенов.
Антигенность рецепторсвязывающей субъединицы, gp120, тримера Env ВИЧ-1 была тщательно изучена, но, как уже упоминалось, многие из эпитопов, которые экспонирует gp120, экранированы нативным тримером [168]. Точно так же нерасщепленный предшественник Env, хотя и тримеризируется, антигенно отличается от нативных функциональных тримеров [102]. Тем не менее, для получения растворимых тримеров, усеченных на N-конце трансмембранного сегмента, которые не разбираются, общим подходом является удаление сайта расщепления между субъединицами и добавление дополнительных мотивов тримеризации на С-конце усечения [169–16]. 174].Но эти нерасщепленные растворимые тримеры не принимают нативные структуры и поэтому плохо имитируют антигенные функциональные шипы.
В качестве альтернативного подхода протеолитический процессинг вместо этого был усилен за счет модификации сайта расщепления и за счет коэкспрессии с протеазой фурин; но для сохранения целостности тримера гетеродимеров была добавлена дисульфидная связь для ковалентного связывания gp120 с усеченным трансмембранным белком gp41 [103, 126, 175–180]. Эти растворимые тримеры структурно имитируют нативные тримеры на поверхности вириона, как было определено с помощью электронной микроскопии [102]; кроме того, их трехмерная структура была определена с разрешением, близким к атомному, в комплексе с Fab различных NAbs [181, 182].
Что касается исследований SPR, то в различных подходах кроется несколько опасностей, связанных с ошибочными результатами. Даже с тримерами, которые структурно имитируют нативные шипы, если они иммобилизованы ковалентно непосредственно на чипах SPR, например амидом, связывание их антигенности будет нарушено. Следовательно, выгодно добавлять His- или эпитопные метки на С-конце при усечении, чтобы тримеры могли быть захвачены антителом или иммобилизованным на чипе антителом. Когда расщепленные и стабилизированные тримеры Env иммобилизуются посредством такого захвата, они связывают NAb, активные против соответствующего штамма вируса, превосходно, а не-NAb — незначительно.В качестве примеров возьмем различные антитела, направленные на сайт связывания CD4 и вариабельную область V3. Обе группы содержат нейтрализующие и ненейтрализующие антитела против определенных штаммов ВИЧ-1. Они одинаково хорошо связываются с мономерной субъединицей Env gp120 и с нерасщепляемыми мутантными тримерными формами Env, полученными из одних и тех же штаммов. Но только нейтрализующие вещества хорошо связываются с тримерами, полученными из штамма, который они нейтрализуют. Некоторые NAb с конкретными эпитопами не связываются с мономерными или неродными формами Env: они специфичны для тримеров, тогда как не-NAb, независимо от эпитопа, неспособны связывать нативные тримеры (рис. 3) [102, 103, 175, 183] .Следовательно, общим для NAb является то, что они распознают нативные вирусные белки, опосредующие проникновение; Что общего у не-НАБов, так это то, что их нет. В этом случае не имеет значения, насколько они отличаются или похожи друг на друга в распознавании других форм Env. Это подтверждает теорию нейтрализации оккупации [31, 99, 106, 184]. Возможно, некоторые антитела, которые не нейтрализуют другие вирусы, несмотря на связывание с вирионами, могут также распознавать только нефункциональные формы поверхностных белков вирусов.
Из-за богатства информации, полученной с помощью SPR, можно идентифицировать NAb с аналогичной аффинностью, но сильно различающейся кинетикой связывания. Такая характеристика связывания выходит за рамки простого занятия и открывает возможность проверки более динамичных теорий нейтрализации: как кинетика связывания NAb и рецептора вместе влияет на эффективность нейтрализации?
Когда связывание IgG и Fabs сравнивается с помощью SPR, можно исследовать конкретные модели двухвалентного или моновалентного связывания.Нам мало известно о плотности тримеров Env на инфекционных вирионах ВИЧ-1, не говоря уже об отношении функциональных тримеров к дефектным или разрушенным. Это соотношение также может меняться с возрастом вирионов. Но иммобилизацию Env на корабле SPR можно точно контролировать и преобразовывать в плотности тримеров, которые можно сравнивать и корректировать с наблюдаемыми плотностями Env на вирионах ВИЧ-1. С улучшенными знаниями, моделирование плотности тримеров на вирионах и, таким образом, реалистичных средних расстояний тримеров, может сделать измеренную степень двухвалентности связывания с антигеном на чипе SPR релевантной для нейтрализации.Некоторые NAb, однако, могут связываться как с дефектными, так и с нативными тримерами, и это может усиливать связывание за счет бивалентности; другие НАБ распознают только тримеры, подобные нативным, и это ограничивает их жадность.
Другие вирусы, чем ВИЧ-1, как правило, имеют более высокие отношения полумаксимальных ингибирующих концентраций Fab по сравнению с IgG [185]. Вклад основной части части Fc, возможно, за счет стерических эффектов, может быть установлен путем сравнения Fab и (Fab ‘) 2 в нейтрализации. Но связывание IgG также может быть усилено взаимодействиями Fc-Fc, и это можно оценить с помощью SPR [186, 187].Аналогичным образом можно сравнивать Fab и одноцепочечные конструкции меньшего размера. Посредством этих комбинированных сравнений можно было бы различить жадность и объемные эффекты. Такие исходные знания можно сравнить и подтвердить с помощью моделирования плотности тримеров на поверхности вириона.
Следует отметить, что степени двухвалентного связывания, которая увеличивает активность NAb за счет снижения скорости их связывания, благоприятствует высокая плотность антигена на поверхности вириона [185]. Такой эффект мог бы противодействовать относительной устойчивости к нейтрализации, возникающей из-за высокой избыточности функциональных молекул, опосредующих проникновение [106, 188–191].Но противодействующие селективные силы не являются симметричными: некоторые NAb одинаково хорошо связываются с функциональными и нефункциональными молекулами, опосредующими проникновение, увеличивая бивалентность, когда присутствуют дополнительные нефункциональные олигомеры, но эффект избыточности требует функциональных олигомеров. Эти факторы могут играть роль в эволюции естественных вирусных вариантов с разной степенью чувствительности к нейтрализации.
Одно основанное на SPR исследование вируса иммунодефицита обезьян (SIV) и его белка Env дало несколько неожиданных результатов.NAb и не-NAb связываются с аналогичной кинетикой и, следовательно, схожим сродством с рекомбинантным растворимым белком Env [192]. Но это был нерасщепленный белок Env, который, как позже было обнаружено, обнажал не нейтрализующие эпитопы; и SPR был основан на иммобилизованном антителе с трехвалентным Env в растворе, который не моделирует потенциально двухвалентное, но часто моновалентное связывание IgG с вирионами. Также удивительным на момент публикации было то, что Env из штамма, устойчивого к нейтрализации, связывался с более высокими скоростями включения и выключения с NAb (и не-NAb), чем Env из чувствительного варианта.Различия по ставкам и вне их компенсировали друг друга, не давая чистой разницы в сходстве. Это указывало бы на большую важность низких процентных ставок в обеспечении чувствительности нейтрализации, чем высоких ставок включения, хотя другие предостережения относительно условий SPR могут сделать недействительными сравнения с нейтрализацией. Конечно, относительное влияние постоянных скорости включения и выключения может быть разным. Но то, как они изменяются, будет определяться продолжительностью предварительной инкубации с NAb в анализе нейтрализации.Это вызывает вопросы о том, какие условия наиболее важны для защиты in vivo .
8. Кинетика проникновения и распада вириона: режим нейтрализации
Классический анализ нейтрализации состоит из четырех этапов. Сначала вирус и антитело инкубируются вместе. Во-вторых, вирусу позволяют адсорбироваться на клетках-мишенях. В-третьих, вирусная репликация продолжает продуцировать вирусный продукт или индуцировать экспрессию репортерной молекулы. В-четвертых, продукт измеряется в анализе и преобразуется в сигнал и сравнивается с контролями без антител и вирусов (фон, шум) [31].Первые две фазы можно варьировать; первый даже может быть устранен. Второй может быть выполнен при пониженной температуре, чтобы предотвратить интернализацию и проникновение.
Результат может сильно отличаться в зависимости от таких вариаций. Некоторые NAb значительно повышают эффективность, если их предварительно инкубировать с вирусом в течение длительных периодов времени. Яркими примерами являются NAb, направленные на внутренние эпитопы в поливирусных белках VP1 и VP4, которые временно экспонируются, когда белки «дышат» при физиологической температуре [193].Другие NAb, по-видимому, действуют после прикрепления вируса к клеткам-мишеням, даже после рецепторных взаимодействий [31, 99, 194]. Если эпитопы временно экспонируются только после взаимодействий с рецепторами, целые молекулы IgG могут быть частично закрыты от доступа к таким точкам в мультимолекулярных комплексах входа; Было показано, что Fab, направленные на такие эпитопы, более эффективны, чем IgG [195, 196].
Следовательно, мы можем различать разные способы нейтрализации: индуцированный распад в суспензии и замедление входа на поверхность клетки или из эндосомы.Строго говоря, тогда режим отличается от механизма, поскольку два режима могут блокировать один и тот же этап репликации, например, взаимодействия рецепторов. Примечательно, однако, что TRIM21-зависимый механизм нейтрализации аденовируса и, возможно, других «голых» вирусов [142] будет иметь другое отношение к предложенным двум режимам нейтрализации: в той степени, в которой инфекционность снижается уже в суспензии или при проникновении. задерживается, цитоплазматическое действие NAbs становится избыточным. Но он может составлять важный третий уровень защиты, по-видимому, доминирующий при определенных обстоятельствах [143].Третий способ — интрацитоплазматическая ингибирующая активность NAb.
Эффекты NAb, действующие на вирионы в суспензии, различаются в зависимости от специфичности эпитопа и вируса, и некоторые из этих эффектов обратимы, другие необратимы [95, 160]. Сами по себе вирионы обычно имеют определенный инфекционный период полужизни в суспензии, и режим нейтрализации в суспензии можно измерить как сокращение этого периода полужизни [194, 197]. Например, для вирусов в оболочке с нековалентно связанными субъединицами поверхностных белков, опосредующих проникновение, эффект NAb может заключаться в индукции диссоциации внешней субъединицы, так что вирус теряет способность связываться с рецепторами; в случае ВИЧ-1 растворимые формы основного рецептора CD4 вызывают диссоциацию субъединицы gp120 от гликопротеинового спайка оболочки.Такой индуцированный шеддинг, также опосредованный некоторыми NAb, может представлять собой преждевременный запуск, аналогичный или более радикальный, чем тот, который индуцируется взаимодействием CD4 и корецепторов на поверхности клетки или в эндосоме в отсутствие антител во время не ингибированного входа [84, 132, 133, 198].
Таким образом, индуцированный распад можно рассматривать как аспект кинетики нейтрализации: он будет представлять собой вторую стадию инактивации после связывания NAb. Его значимость для защиты in vivo будет зависеть от продолжительных периодов воздействия вируса на антитела до контакта с чувствительными клетками; эта значимость также будет зависеть от того, оказывает ли связывание антитела ингибирующие эффекты, не нейтрализующие его, такие как опсонизация вирусных частиц для разрушения клеток посредством фагоцитоза.Последние могут доминировать. Следует также отметить, что спонтанный, а также индуцированный распад могут влиять на количество молекул, опосредующих проникновение в вирион, которые должны быть инактивированы NAb, чтобы блокировать инфекцию: значение нейтрализации IC 50 не является фиксированным, но может варьироваться в зависимости от времени предварительной инкубации. Судя по всему, NAbs могут откусить и отколоть на пороге нейтрализации.
Хотя связывание антитела обратимо, замедление проникновения может иметь необратимый результат, как и индуцированный распад в суспензии.Замедляя продуктивный вход, NAbs могут по умолчанию шунтировать вирус на абортивный путь [199]. Вирусы, которые используют проникновение или слияние на клеточной поверхности в качестве обязательного шага, могут быть направлены на прерванный эндоцитарный путь посредством взаимодействий с Fc-Fc-рецептором или просто путем отсроченного проникновения. Это решило бы судьбу вириона, покрытого NAb. В конце этого пути кроется угроза разрушения лизосом.
С вирусами, которые используют эндоцитотический путь проникновения, ситуация иная.Например, сообщалось, что некоторые NAb, связанные с вирусом Западного Нила, замедляют интернализацию вируса через эндоцитарный путь и, таким образом, делают вирус более доступным для PAN [200]. Напротив, другие NAb или, по крайней мере, меньшая степень заполнения NAb, могут усиливать инфекцию вируса Западного Нила, способствуя эндоцитозу и обеспечивая слияние с эндосомальной мембраной [151, 201].
При прочих равных условиях любой необратимый аспект нейтрализации вируса, будь то режим индуцированного распада в суспензии или направление вирионов на разрушение клетками в других режимах, должен способствовать защите хозяина.Остаются вопросы, является ли необратимость инактивации избыточной или критической.
9. Молекулярность: сколько совпадений?
Как мы видели, ошибочные интерпретации кинетики нейтрализации посеяли непокорную путаницу в отношении того, сколько молекул NAb требуется для нейтрализации, то есть его молекулярность , минимальное количество молекул, участвующих в определяющей скорость стадии реакции. Кинетика нейтрализации не покажет его молекулярность, но кинетика связывания может дать сродство и, следовательно, позволить обоснованные оценки занятости.Заполнение является эквивалентом стехиометрии, и если известно абсолютное количество функциональных, релевантных для нейтрализации сайтов вирусных белков, опосредующих проникновение, молекулярность определяется следующим образом.
При изучении кинетики связывания с помощью SPR также могут быть получены стехиометрические значения, но они относятся к максимальному связыванию, например, паратопов на олигомерный антиген; эти данные будут полезны, но недостаточны для правильного определения молекулярности на уровне вириона. И есть предостережения в отношении определений SPR; например, некоторые NAb диссоциируют так медленно, что невозможно получить значительные значения, или они настолько высоки, что ограничение массопереноса приводит к неопределенности.Как сродство, так и стехиометрия, однако, также могут быть определены калориметрическим методом, тогда как стехиометрия, кроме того, может быть установлена с помощью электронной микроскопии (рис. 4) [103, 126, 182], и эти данные могут обеспечить необходимые дополнения и корректировки данных SPR. .
Фенотипически смешанный вирус был использован для более точного определения молекулярности проникновения и нейтрализации. Такие вирусные препараты содержат разные пропорции антигенных и неантигенных, функциональных и дефектных протомеров олигомерных белков, опосредующих проникновение [111–114, 202].Следовательно, комбинаторные математические уравнения, полученные из биномиальной теоремы, могут применяться для интерпретации данных [105, 107, 108, 110]. Но также этот подход влечет за собой подводные камни. Если входной белок олигомерный и каждый вирион, как обычно, имеет более одного олигомера, моделирование должно одновременно совмещать два уровня анализа, каждый со своим собственным потенциальным порогом. Одним из следствий является то, что совершенно разные модели могут предполагать неразличимые эмпирические данные, потому что изменения в модельных предпосылках на уровнях олигомера и вириона компенсируют друг друга [84, 105].Примечательно, что предпосылка о том, что каждый вирион обладает только одним олигомером, опосредующим проникновение, и, следовательно, нуждается только в одном, переводится в точно такое же уравнение, что и самая экстремальная инкрементная модель, упомянутая ранее, а именно, что каждый олигомер вносит равный вклад в инфекционность, независимо от того, сколько нетронутых олигомеров остается на один вирион или как они группируются.
Предположение о том, что инфекционность является бинарной, все вирионы полностью или совсем не заразны, и что нет степени нейтрализации отдельного вириона, вероятно, следует изменить оба.Эндрюс и Элфорд уже были на удивление близки к предсказанию возрастающих нейтрализующих эффектов [203]: если бы к их анализу прислушались, некоторые заблуждения, вытекающие из предпосылок «все или ничего», не получили бы распространения. Другая крайность, то есть все незанятые функциональные белковые олигомеры вносят инкрементальный вклад в точную пропорцию их количеству, может быть столь же нереалистичным. Вирион, вероятно, может быть наполовину нейтрализован, но это не означает, что не может быть избыточности в количестве входных белков на вирионах или что все вирионы пропорционально заразны вплоть до своего последнего олигомерного шипа или молекулы, опосредующей вход [84, 105] .Имея в виду эти сложности, можно, тем не менее, сделать информативные выводы из данных о смешанных фенотипах. И это важно, потому что это помогает оценить, какие концентрации и сродство NAb должны индуцировать вакцины для защиты.
Другими подходами к определению присутствия нейтрализующих антител являются электронная микроскопия и различные биохимические измерения связывания NAb с вирионами. Последние методы дают только среднюю степень связывания, но аналогичные анализы нейтрализации просто предоставляют данные о средней инфекционности для популяции вирионов.Таким образом, электронная микроскопия имеет преимущество, поскольку она может описать полное распределение заселенностей NAb по популяции вирионов.
Источником артефактов этих анализов связывания в совокупности является то, что они не различают инфекционные и неинфекционные вирионы. Информация о соотношении инфекционных и неинфекционных вирионов будет информативной: чем выше это соотношение, тем выше актуальность измерений. Но, как и в случае с нейтрализацией, инфекционность вряд ли будет свойством «все или ничего».
Исследования, основанные на радиоактивно меченных NAb, показали, что частицы полиовируса с одной бивалентно связанной молекулой IgG были нейтрализованы [98]. Было предложено полностью нейтрализовать аденовирус: в среднем на один вирион приходится 1,4 молекулы NAb [204]. Это вызывает вопросы относительно точности или актуальности измерений. Фактически, нейтрализация и связывание не проводились в тех же условиях в последнем исследовании [204]: вирус и NAb инкубировали в течение 3 часов при 37 ° C, а затем еще 21 час при 4 ° C перед анализом инфекционности.Результаты инфекционности наносили на график как логарифмическую выживаемость как функцию линейной концентрации в сыворотке. В анализе связывания NAb и вирус инкубировали только 30 мин при 37 ° C, а затем комплексы вирион-NAb отделяли от свободных NAb центрифугированием в градиенте сахарозы. Восстановленный вирус не был заразным. Вполне вероятно, что процедура разделения может нарушить как равновесное связывание, так и инфекционность вириона. Анализ также ставит теоретические проблемы.
Была предпринята попытка анализа Пуассона для объяснения взаимосвязи связывания-нейтрализации в двух различных условиях, длительной и короткой инкубации.Есть две проблемы. Во-первых, нейтрализация за один удар с распределением по Пуассону связыванием и в среднем 1,4 молекулы NAb на вирион подразумевает остаточную инфекционность около 25%. Поскольку инфекционность не была обнаружена, либо вирионы были неспецифически повреждены процедурой, либо точность анализа была недостаточной для такого рода анализа. Во-вторых, пуассоновский анализ нейтрализации требует знания количества связанных NAbs и вытекающей из этого остаточной инфекционности. Переход от простой концентрации в сыворотке к множеству связанных NAb потребует гораздо большего количества информации, а именно средней аффинности и концентрации [106, 205].И тогда теоретическая кривая логарифмической остаточной инфекционности как функции концентрации NAb не является линейной даже для нейтрализации за один удар [106, 205]. Только кривая, постулирующая нейтрализацию однократным попаданием со средней множественностью связывания NAb на оси β, является линейной [205]. Однако на практике приблизительно линейная кривая может возникнуть, если самые низкие разведения сыворотки дают концентрации NAb, намного превышающие среднее значение. Следовательно, линейность, наблюдаемая в таких условиях, вопреки аргументу, приведенному в исследовании [204], не является доказательством молекулярности однократного попадания.
Предлагаемое связывание одиночного NAb с полиовирусом, упомянутое выше, не было основано на анализе Пуассона [98]. Исследование, в котором применялся анализ Пуассона, опровергло нейтрализацию полиовируса за один удар и предложило минимум четыре NAb на вирион [206]. Примечательно, что также наблюдалась кинетика первого порядка, и к настоящему времени должно быть ясно, что в этих наблюдениях нет ничего противоречащего. В этом исследовании было получено минимальное количество молекул NAb, считая значение на оси при относительной инфекционности 1/ e .Эта процедура была обычной практикой, но не оправдана в теории. Только для кривой с одним совпадением 1/ e точно соответствует минимальному числу NAb. Чем выше фактическое число, тем больше отклонение. Тем не менее, с помощью этого метода была определена молекулярность 70 молекул IgG на вирион гриппа [158].
Анализ Пуассона обеспечил бы наилучшую проверку данных путем сравнения с разрозненными теоретическими кривыми для различных молекулярностей: одно, два, три, четыре, пять и так далее попаданий [106, 205].Таким образом можно обоснованно оценить соответствие данных конкретной модели. Или, по крайней мере, можно рассчитать прогнозируемую относительную инфекционность для каждой модели при пороговом значении, а показания можно сравнить с теоретическим значением. Для молекул с молекулярностью, отличной от 1 (для полной нейтрализации требуется минимум один NAb на вирион), показания на оси -оси при / e теоретически столь же произвольны, как и при, скажем, 50% нейтрализации, что не означает это малоинформативно; по крайней мере, это могло бы опровергнуть молекулярность однократного попадания [31, 106].
Для записи, анализ Пуассона включает в себя следующее. На оси -оси отложен натуральный логарифм все еще заразной фракции вируса, как функция среднего числа NAb на вирион,,, на оси -оси. Минимальное количество NAb на вирион, необходимое для нейтрализации, должно иметь целочисленное значение. Инфекционная фракция вирионов будет равна кумулятивным фракциям с меньшим количеством связанных с ними NAb:. Таким образом, с оспариваемой молекулярностью единичного попадания и в среднем 1 NAb на вирион, то есть, так что: если бы гипотеза единственного попадания была верна, примерно 37% инфекционности сохранялось бы, когда вирионы в среднем имели один NAb привязан к ним.Но очевидно больше будет отличаться от значения при / e , чем выше.
Преобладающие ошибки в анализе Пуассона могут, однако, иметь меньшее значение по сравнению с ошибочной предпосылкой подхода: что все вирионы имеют одинаковую инфекционность в отсутствие NAb и что порог нейтрализации абсолютен; помещения ниже не имеют никакого эффекта; как только он будет достигнут, нейтрализация будет полной, и более высокая занятость не имеет никакого значения.
Причина, по которой мы сосредоточили внимание на подводных камнях выводов и ошибках при выполнении пуассоновского анализа нейтрализации, заключается в том, что эти недостатки оказали большое влияние на поле нейтрализации.Молекулярность единичного попадания была возведена в категорию виртуальных догм на надуманных основаниях; он распространен по учебникам [207]. Открытие TRIM21-зависимого механизма предполагает, что нейтрализация за один удар возможна на молекулярном уровне; тогда это могло бы считаться ранее по неправильным причинам и в основном о неправильных вирусах. В действительности нейтрализация аденовирусной инфекции эмбриональных фибробластов мыши и линии эпителиальных клеток человека HeLa происходила при среднем количестве NAb на вирион, равном 1.6 и 4.8, соответственно, когда NAb был мышиным. Обе клеточные линии стимулировали IFN, чтобы максимизировать экспрессию TRIM21. Объяснение различий в результатах между клеточными линиями могло бы состоять в том, что мышиный TRIM21 имеет более высокое сродство к мышиным NAb, чем человеческий TRIM21 [143]. На более низких уровнях TRIM21 требовалось гораздо большее количество людей. Или, в крайнем случае, даже насыщающие концентрации NAb не давали измеримой нейтрализации; аналогично Fc-мутанты NAb, неспособные взаимодействовать с TRIM21, не смогли нейтрализовать.Эти новые открытия проливают свет на контекстно-зависимые и неабсолютные молекулярные массы, которых достаточно для нейтрализации. Они также предполагают заметную неэффективность нейтрализации на этапах до входа в цитоплазму. Поэтому следует вернуться к более старым наблюдениям, с которыми они иногда кажутся противоречащими.
В более раннем исследовании частицы аденовируса, связанные с помощью NAb, не могли проникнуть из эндосомального пузырька и, следовательно, не могли быть подвергнуты TRIM21-опосредованной маркировке для деградации; наблюдаемый блок находился на более раннем этапе.Предполагаемый механизм заключался в том, что связывание NAb с основанием пентона предотвращало взаимодействия с эндосомальной мембраной и, следовательно, блокировало вход. Антифибровые антитела агрегируют вирионы и тем самым обеспечивают некоторую степень нейтрализации. НК антигексона блокировали вызванные pH конформационные изменения в капсиде, которые, как считается, способствовали проникновению. Антигексоновые NAb также опосредуют PAN [204].
Прямые доказательства подтверждают молекулярную молекулярную нейтрализацию многих вирусов, не только вирусов в оболочке.Таким образом, для нейтрализации полиовируса, одного из самых маленьких вирусов, требуется 4–5 молекул NAb, 36–38 для вируса папилломы, 70 для вируса гриппа и 225 для вируса бешенства [99]. Эти возрастающие числа предполагают примерно линейную зависимость между площадью поверхности вириона и минимальным количеством молекул NAb, необходимых для нейтрализации [99, 201]. Эта корреляция приблизительно согласуется с теориями нейтрализации оккупации, но не демонстрирует их общей обоснованности. В сравнении есть два «голых» вируса, но, как уже говорилось, разные принципы применяются к механизмам и молекулярной массе нейтрализации «голых» вирусов и вирусов в оболочке.Гипотетически приблизительная линейность может лучше всего применяться к вирусам в оболочке, но среди них будут играть такие факторы, как плотность, функциональность и хрупкость белка оболочки; и плотность клеток-хозяев также может определять эффективность нейтрализации, то есть зависимость от концентрации Nab [208]. В этом отношении наблюдения не основаны на сопоставимых условиях для различных вирусов. Поэтому грубая корреляция между размером вириона и количеством связанных NAbs, необходимых для нейтрализации, становится еще более поразительной.
Оболочечный вирус Западного Нила, который принадлежит к роду флавивирусов семейства Flaviridae , вероятно, представляет собой количественно наиболее понятный пример нейтрализации антител [149]. Одни и те же антитела могут как нейтрализовать, так и усилить вирусную инфекцию: их эффект зависит от достигнутой занятости. На вирион доступно около 120 эпитопов для некоторых конкретных NAb. Когда эти NAb занимают 25 эпитопов, инфекционность вириона снижается вдвое. При более низкой заселенности эти же антитела вместо этого способны усиливать инфекцию, направляя вирус по пути эндоцитов через рецепторы Fc, таким образом, без стерического блокирования окончательного слияния оболочки с эндосомальной мембраной [151, 201].
Другие NAb как вируса Западного Нила, так и вируса денге требуют более высокой степени заполнения их эпитопов для нейтрализации. Объяснение, которое элегантно иллюстрирует теорию занятости, состоит в том, что меньшее количество этих эпитопов доступно для связывания NAb, поэтому для достижения аналогичного порогового количества NAb или Fab на вирион необходимо лигировать большую часть экспонированных эпитопов [149]. .
Как уже упоминалось, было высказано предположение, что низкая заселенность пиков Env на вирионах ВИЧ-1 шунтирует вирус на непродуктивный эндоцитарный путь, также посредством взаимодействий с Fc-рецептором [148].Этот механизм инактивации имел смысл, когда считалось, что ВИЧ-1 продуктивно проникает через прямое слияние с мембраной на поверхности клетки. Но теперь, когда убедительные доказательства указывают на то, что эндоцитотический путь продуктивен и даже обязателен [83–85, 137, 209–211], этот эффект низкой занятости антител стал интригующим. Также наблюдалось усиление эффектов низкой занятости NAb на ВИЧ-1 [106]. С точки зрения вакцины было бы обнадеживающим знать, как различное количество NAb на вирионах влияет на инфекционность.
10. Степень, эффективность и стойкая фракция
Постоянная фракция (PF) вирусной инфекционности — это плато инфекционности, к которому асимптотически приближается по мере того, как инкубация с NAb продлевается или концентрация NAb увеличивается (рис. 5). Здесь основное внимание будет уделено последнему, но необходимо сделать несколько общих замечаний по поводу различных гипотез, которые были сформулированы на протяжении многих лет для объяснения этих явлений [58, 99, 156, 184, 212–214].
Правдоподобное объяснение может заключаться в том, что вирус неоднороден, а PF просто представляет собой устойчивый вариант.Хотя обобщения были бы необдуманными, по крайней мере в некоторых случаях вирус, который вырос из устойчивых фракций, тем не менее, показал чувствительность к нейтрализации, аналогичную чувствительности исходного вируса [58, 99, 184, 212, 214]. Если исключить генетически обусловленные причины, эпигенетические причины, такие как процессинг гликанов, могут быть ответственны за сохранение. Если нейтрализация требует конформационных изменений в вирусных белках, они могут быть индуцированы в разной степени, и тогда некоторые вирионы могут быть более устойчивыми к эффекту связывания NAb [58].Спектр плотностей функционально сохраненных молекул нейтрализующего антигена в популяции вирионов также может дать, если не абсолютную устойчивость, по крайней мере, хвост менее чувствительных вирионов.
До того, как PF были продемонстрированы с моноклональными NAb, была выявлена гетерогенность среди антител в сыворотке. Также упоминалось о существовании антител, блокирующих нейтрализацию [58, 127]. Даже моноклональные NAb могут быть гетерогенными из-за вариации посттрансляционных модификаций, например сульфатирования тирозина, влияющего на аффинность [215].Тем не менее, PF являются более общим явлением, и их возникновение не может быть привязано к конкретным NAb, хотя их размеры варьируются в зависимости от NAbs.
Большое внимание в ранних исследованиях PF уделялось агрегатам и тому, как они сохраняют некоторую инфекционность и прикрепляются с большей жадностью, чем отдельные вирионы [58]. Но PF наблюдались и с монодисперсным вирусом [213]. Следует отметить, что закон действия масс не подразумевает какой-либо PF (рис. 5): только когда подмножество целевых молекул имеет аберрантное, существенно сниженное сродство может закончиться нейтрализацией.Бернет плодотворно предположил, что диссоциация NAb ответственна за PF [212]. Это согласуется с выводами о том, что как добавление вторичных антител, перекрестно связывающих антитела в нейтрализующей сыворотке, так и комбинация NAb с отдельными эпитопами может уменьшить размер PF [213]. Гипотеза диссоциации становится еще более правдоподобной, когда применяется к динамической конкуренции между рецепторами и NAb в месте проникновения. Также важно, что степень моновалентного связывания IgG повышается, когда связывание приближается к насыщению, как и ожидалось из хорошо установленного эффекта прозоны [216].Таким образом, авидность снижается, и конкурентное связывание с рецепторами предпочтительнее, чем с паратопами.
Недавно было показано, что PF ВИЧ-1 коррелирует с константами отклонения NAb, измеренными с помощью нативных тримеров Env ВИЧ-1 с помощью SPR [183]. Кроме того, стехиометрия была связана с PF (Рисунки 4 и 5). Большинство NAb с тримером Env ВИЧ-1 связываются со стехиометрией, приближающейся к трем паратопам на тример; но некоторые специфичные для тримеров NAb имеют только один эпитоп на вершине тримера (рис. 4) [175].Таким образом, стехиометрия по оценке калориметрии и SPR коррелировала с размером PF: чем выше стехиометрия, тем ниже PF [183]. Это открытие также может быть связано с диссоциацией, потому что при стехиометрии один паратоп на вирусный олигомер уязвимость к диссоциации будет выше. В самом деле, другие данные указывают на то, что одной молекулы IgG на тример некоторых NAb достаточно, чтобы блокировать функцию этого тримера [84, 105, 108, 110, 111, 114, 202]. Таким образом, когда три связаны, два являются избыточными, но будут действовать как сети безопасности, особенно в динамическом соревновании с рецепторами, когда требования к занятости самые высокие.
Исследования причин PF возродились после разоблачения TRIM21-зависимой нейтрализации. Таким образом, нейтрализация аденовируса одним и тем же NAb оставляет более высокие или более низкие PF обратно пропорционально уровням TRIM21 в клетке-мишени [143]. Но в конце концов устанавливается двойное плато: увеличение занятости NAb и уровни экспрессии TRIM21 достигают убывающей отдачи, а остаточная инфекционность выходит на плато. Действительно, чем выше вирусная доза, тем легче преодолеть комбинированную защиту NAbs и TRIM21.Вполне вероятно, что др. Клеточные факторы, ответственные за деградацию комплексов NAb-вирус после TRIM21-опосредованного убиквитинирования, становятся ограничивающими на этом этапе. Ясно, что старая проблема ПФ в значительной степени прояснилась открытием роли вспомогательных клеточных факторов в нейтрализации. Вопрос в том, насколько далеко эти идеи могут выйти за рамки нейтрализации аденовируса [142].
11. Синергия и взаимодействие
На молекулярном уровне два NAb могут потенциально усиливать связывание друг друга [217].Это будет означать синергизм в нейтрализации, способствующий индукции антител к множественным эпитопам путем вакцинации. Но синергизм в нейтрализации также наблюдается, когда кажется, что на уровне связывания антитела с молекулой антигена нет никакого синергизма, и действительно, когда не может быть никакого синергизма в связывании, потому что синергизирующие NAb направлены на перекрывающиеся эпитопы [218]. Тогда объяснение должно быть другим. Вероятно, это неоднородность популяции молекул-мишеней, то есть антигенов, которая может быть обширной для некоторых вирусов [219].Эта гетерогенность не обязательно должна быть генетически детерминирована одна, но также может происходить из-за изменчивости посттрансляционных модификаций. Таким образом, синергизм будет возникать из-за того, как два NAb дополняют друг друга, охватывая разные эпитопы или иногда разные варианты одного и того же эпитопа, к которым NAb имеют различное сродство. Такое объяснение в равной степени применимо к синергии в эффективности, то есть эффективности, как и в эффективности, причем последнее отражается в снижении PF.
При определении синергизма были допущены как концептуальные, так и практические ошибки и ошибки моделирования.Но классический индекс синергии, который, вопреки многим утверждениям, действительно имеет общую ценность, может пролить свет как на синергетические, так и на антагонистические эффекты, при условии, что он рассчитывается с помощью подходов, которые избегают некоторых распространенных источников артефактов и ошибок [219, 220]. Тем не менее, это просто мера синергии или антагонизма в потенции. Эффективность может быть не менее важной, чем in vivo , и потребует столь же строгой основы для анализа.
Кооперативность отдельных ингибиторов, действующих на несколько сайтов на олигомерные молекулы-мишени, классически измерялась как наклон, коэффициенты Хилла, кривых ингибирования [221].Однако ясно, что неоднородность может уменьшить наклон этих кривых [219, 222, 223]. И то, что комбинация двух NAb с уменьшенными наклонами дает более высокий наклон, может объяснить, как антигенная гетерогенность приводит к синергетическим явлениям [219]. Наклон кривых нейтрализации изучен гораздо меньше, чем средняя точка, то есть эффективность, но он отражает важное свойство NAb и частично является продуктом гетерогенности эпитопа; он также может быть предметом паратопной гетерогенности даже для моноклональных антител, когда внутренняя аффинность модулируется, например, с помощью сульфатирования тирозина, что не может быть достигнуто равномерно в популяциях НК, полученных естественным путем [215].
Обычно в исследованиях ингибирования, когда одна молекула инертна, а другая активна, но комбинация все еще более активна, подходящим термином для комбинаторного эффекта является потенцирование [224]. Что касается постентарной нейтрализации, можно отметить, что интерфероны могут усиливать нейтрализацию, повышая TRIM21 [141–143]. Еще неизвестно, в какой степени TRIM21-зависимое ингибирование просто обеспечивает безопасность, когда NAb не в состоянии предотвратить проникновение, или же оно, скорее, составляет основной механизм нейтрализации, и против каких голых вирусов оно активно.В этой области исследований может возникнуть множество новых сложных проблем, касающихся того, как клеточные факторы ограничивают или усиливают нейтрализацию, и дают ли комбинации аддитивные, синергические или антагонистические сетевые эффекты.
12. Выводы: редукционизм Redux
Было бы большим преимуществом при разработке вакцины знать, что для нейтрализации антител не требуется никаких других специальных свойств, кроме его способности связывать функциональные вирусные белки, опосредующие проникновение, на поверхность. вириона: вакцинация упрощается, если достаточно создать имитаторы таких вирусных белков и сделать их иммуногенными.Для некоторых вирусов, таких как ВИЧ-1, уже очень трудно получить достаточно хорошую имитацию нативного белка Env и сфокусировать иммунный ответ на консервативных областях, чтобы индуцированные антитела также распознавали тримеры Env встречающихся в природе дивергентных вариантов белка. вирус. Поэтому любые дальнейшие осложнения, такие как потребность в антителах для индукции определенных конформационных изменений в антигенах или действительно сложных посттрансляционных модификаций, необходимых для наиболее сильного связывания, будут крайне нежелательными.Точно так же приятно, если нейтрализация работает таким образом, что антитела, способные ее блокировать, не возникают быстро: все еще может быть невозможно избежать индукции некоторых нерелевантных, инертных антител или, в случае некоторых вирусов, антител, усиливающих инфекцию.
Комплексный взгляд на нейтрализацию как на вмешательство в различные этапы проникновения вируса и как результат критической степени покрытия вирусной частицы NAb, подтверждается несколькими линиями доказательств для многих вирусов.Но открытие роли цитоплазматического фактора TRIM21 в нейтрализации аденовирусов предоставило интригующий пример подлинной постентарной нейтрализации и рациональное объяснение того, почему для нейтрализации иногда достаточно очень небольшого количества NAb на вирион. Еще неизвестно, представляет ли этот механизм исключение или порождает более общую дихотомию между незащищенными вирусами и вирусами в оболочке. Всякий раз, когда голые вирусные ядра могут проникать через клеточные мембраны, сохраняя при этом связанные NAb, существует потенциал для внутрицитоплазматической нейтрализации, ускоренной клеточными факторами.
Улучшенная биофизическая характеристика связывания NAb с нативными вирусными антигенами дает много информации, потенциально имеющей отношение к эффективности нейтрализации in vitro и защиты NAb in vivo. Грубая корреляция между аффинностью и эффективностью нейтрализации может, однако, дать упрощенную картину взаимосвязи между связыванием NAb и нейтрализацией. Стехиометрия и кинетика связывания могут также влиять на эффективность нейтрализации, которая может быть более важной, чем эффективность для предотвращения распространения инфекции в организме.Хотя улучшение знаний о том, как защищают NAb, пойдет на пользу при разработке вакцины, любые дополнительные требования, помимо достаточного связывания с функциональными антигенами, снова усложнят задачу.
Ранние попытки свести нейтрализацию к химическим принципам были ошибочными, отчасти потому, что инфекционность и ее ингибирование чрезмерно упрощенно рассматривались как явления «все или ничего». Но теперь появляющееся количественное понимание того, как вирусные белки вносят вклад в инфекционность, вместе с биофизической характеристикой связывания NAb с вирусными антигенами, может объяснить многие аспекты нейтрализации вирусов.Хотя и блокировка входа, и механизмы постентрии должны быть поняты в их различных молекулярных деталях, общие принципы могут преобладать, такие как необходимая занятость и повсеместная конкуренция с динамическими событиями вирусной репликации.
Конфликт интересов
Автор заявляет об отсутствии конфликта интересов в отношении публикации данной статьи.
Благодарность
Работа автора в этой области поддержана грантом NIH R37 AI36082.
Измерение суррогатов защиты от смертельного заболевания
Цитирование: Мур С.М., Хэнлон Калифорния (2010) Антирабические антитела: Измерение суррогатов защиты от смертельного заболевания. PLoS Negl Trop Dis 4 (3): e595. https://doi.org/10.1371/journal.pntd.0000595
Редактор: Джеффри М. Бетони, Университет Джорджа Вашингтона, Соединенные Штаты Америки
Опубликован: 9 марта 2010 г.
Авторские права: © 2010 Мур, Хэнлон.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Специального финансирования для написания этой статьи получено не было.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Желает ли работник службы контроля животных определить, необходима ли бустерная вакцина против бешенства для установления приемлемого предконтактного статуса, или врач рассматривает причины энцефалита у ребенка, или владелец собаки с ослабленным иммунитетом обеспокоен тем, что реакция собаки на вакцинацию против бешенства будет недостаточной для прохождения серологического теста, позволяющего ей отправиться в зону, свободную от бешенства, или исследователь пытается определить, адекватна ли реакция на вакцину против бешенства в популяции енотов, или нужно присвоить значение эффективности продукту иммуноглобулина против бешенства, все это требует точной оценки, основанной на измерении циркулирующих антител.В каждой из этих ситуаций измерение или просто обнаружение специфичных для вируса бешенства нейтрализующих или других антител поможет решить вопрос. Однако точно так же, как обстоятельства в каждом из этих сценариев различны, особенности метода, выбранного для измерения антител, нормативные требования к тестированию и цели тестирования в каждой из этих ситуаций различны. Антитела возникают в результате гуморального иммунного ответа на антигены бешенства, процесс которого контролируется многими факторами, включая количество антигена, путь доставки, экспрессию и участие генов главного комплекса гистосовместимости (MHC) и состояние здоровья индивидуальный, среди прочего.Понимание иммунного ответа хозяина, включая подкласс (тип) иммуноглобулинов (Ig), а также кинетику и продолжительность ответа, необходимо для определения меры или степени иммунитета против бешенства.
Первоначально измерение нейтрализующих антител к вирусу бешенства (РВНК) проводили in vivo с использованием теста нейтрализации на мышах (MNT). Впоследствии быстрый тест подавления флюоресцентного фокуса (RFFIT) был признан «золотым стандартом» in vitro [1]. Методы измерения иммунитета к бешенству различаются в зависимости от измеряемого гуморального компонента (т.е. подкласс Ig или функциональная активность) и рабочие характеристики (то есть специфичность или чувствительность), а также стоимость и сложность метода. Понимание каждого из этих уникальных факторов необходимо при выборе и правильном использовании метода, а также важно при интерпретации результатов, полученных с помощью методов. Более того, правила (например, Управления по санитарному надзору за качеством пищевых продуктов и медикаментов США (USFDA), Европейского союза, Всемирной организации здравоохранения животных), требующие проверенных и утвержденных методов тестирования для измерения образования антител к вирусу бешенства, актуальны при транспортировке домашних животных и при бешенстве. производство и оценка биопрепаратов для использования как людьми, так и животными.В этом обзоре мы обсуждаем следующее (1) роль специфических антител к бешенству в профилактике заболеваний, (2) методы, которые можно использовать для обнаружения и измерения, и (3) соображения и текущие требования к стандартизации и валидации методов. .
Методы
Обзор литературы был проведен с использованием онлайн-базы данных PubMed с 1975 по 2008 год с заголовками медицинских дисциплин Национальной медицинской библиотеки США (MeSH). Справочные списки избранных статей и обзоров также были исследованы индивидуально.Кроме того, были проанализированы неопубликованные серологические данные по бешенству из лаборатории бешенства Канзасского государственного университета.
Роль специфических антител против бешенства в профилактике заболеваний
Животные модели защиты от бешенства продемонстрировали важную роль РВНК [2], [3]. В самом деле, одна только РВНК может привести к выведению вируса из центральной нервной системы (ЦНС) экспериментально инфицированных мышей [4]. Большинство антирабических антител направлено на эпитопы гликопротеина вируса бешенства, хотя некоторые могут специфически распознавать нуклеопротеин [5], [6].Как специфические антитела против бешенства нейтрализуют вирус, не совсем понятно с точки зрения того, какие конкретные эпитопы или подклассы Ig задействованы [7, 8]. Один вирион бешенства может связывать до 1000 молекул некоторых антигликопротеиновых моноклональных антител без нейтрализации, предполагая, что нейтрализация вируса, вероятно, включает больше, чем просто связывание антител с эпитопами вириона [7], [8]. Можно предположить, что антитела, нейтрализующие вирус in vitro, более эффективны в процессе, который приводит к защите от вирусной инфекции in vivo, чем антитела, которые не нейтрализуют вирус in vitro.Только Fc-часть антител выполняет специфические биологические функции, включая активацию антигенпрезентирующих клеток и каскад комплемента. Таким образом, ожидается, что цельный IgG будет более активным, чем эпитоп-специфичные части нейтрализующих антител F (ab) 2 [9]. С развитием молекулярных методов конструирования специфичности Fab-фрагментов антител и интактных моноклональных антител для диагностических и терапевтических целей важность понимания того, как антитела механически нейтрализуют вирус бешенства, становится все более важной.Поскольку с практической точки зрения трудно предсказать способность препарата поликлональных или моноклональных антител нейтрализовать вирус путем измерения только авидности или аффинности антител, нейтрализационная активность должна быть сначала определена некоторыми способами in vitro, даже если только для того, чтобы обеспечить оценку in vitro. vivo активность. Тем не менее, препарат антирабических антител, демонстрирующий сильную нейтрализацию in vitro, может все же не защищать in vivo [10].
Вирус бешенства очень нейротропен.Внутри ЦНС инфекция вируса бешенства развивается по большей части незаметно адаптивными иммунными реакциями хозяина, пока инфекция не перейдет в завершающую стадию. В дополнение к инфекции, которая развивается в этом иммунном привилегированном месте, вирус бешенства способен подорвать любой иммунный ответ хозяина, и, таким образом, исход инфекции почти в 100% случаев заканчивается смертельным исходом. Напротив, вакцина против бешенства способна стимулировать высокие уровни циркулирующей РВНК. Вот почему иммунитет после контакта с вирусом бешенства у невакцинированного человека, инициированный немедленным очищением раны для снижения вирусной нагрузки в месте воздействия и введением антирабического Ig с последующей серией вакцинации против бешенства, практически на 100% эффективен в предотвращении инфекционное заболевание.Пассивная защита с помощью иммуноглобулина бешенства (RIG) имеет решающее значение для немедленной нейтрализации большинства инфекционных вирионов, предотвращая распространение вируса в ожидании индукции адаптивного иммунного ответа хозяина. В целом, антирабические вакцины исторически представляют собой убитые цельновирусные вакцины, которые, как ожидается, будут способствовать развитию специфических антител против вируса бешенства наряду с ответом Т-лимфоцитов CD4 + , который включает продукцию цитокинов. Ответ поликлональных антител каждого человека на вакцинацию против бешенства представляет собой уникальное сочетание специфичности к антигенам вируса бешенства.Хотя у большинства людей вырабатываются различные измеримые антитела в ответ на вакцинацию до или после контакта, могут быть существенные различия в нейтрализующей активности и количестве продуцируемой РВНК.
Антитела развиваются из внутренних ответов хозяина и в ответ на внешние факторы, такие как количество введенного антигена, тип антигена и путь воздействия или вакцинации. Например, было продемонстрировано, что чем сильнее вакцина, тем выше уровни РВНК, индуцируемые у людей [11].Другие исследования показали, что тип вакцины и способ вакцинации влияют на уровень антител как через 14 дней, так и через год после вакцинации [12]. Кроме того, иммунный ответ Т-хелперов типа 2 (Th3) на убитую вакцину по своей сути отличается от активации иммунитета Т-хелперов типа 1 (Th2) вакциной на основе живого вируса. В дополнение к факторам хозяина, таким как возраст и общее состояние здоровья, адаптивный иммунный ответ индивидуума требует связывания чужеродных пептидов с молекулой MHC на антигенпрезентирующих клетках и стимуляции соответствующего клона Т-клеток этой конкретной комбинацией MHC-пептид.Разнообразие генов в комплексе MHC, состоящем из сотен аллелей, влияет на вариацию иммунного ответа на вакцинацию [13]. Иммунные ответы гуморального (Th3) и клеточного (Th2) типа в значительной степени направляются выработкой цитокинов, включая созревание ответа антител и образование подкласса Ig. Также наблюдалась дихотомия ответов при вакцинации против бешенства среди людей на людей с высоким или хорошим ответом и с плохим или низким ответом [14]. Прогнозы относительно продолжительности ответа РВНК могут быть сделаны на основе уровня и времени пикового ответа на вакцинацию [12], [14].
Методы обнаружения и измерения антител к вирусу бешенства
Доступные методы обнаружения и измерения антител, специфичных к вирусу бешенства, — это либо анализы связывания антигена, либо анализы нейтрализации вируса. В анализах связывания антигена антитела в сыворотке или спинномозговой жидкости (ЦСЖ) выявляются, количественно определяются и характеризуются их способностью связываться с различными антигенами вируса бешенства из цельного вируса, очищенных субъединиц или специфических пептидов, имитирующих эпитопы.Такие анализы определяют аффинность, авидность и специфичность связывающихся антител. Обычно эти методы включают фиксацию антигена на твердой поверхности, то есть на пробирке, пластине или шарике. Затем взаимодействие между антителом и антигеном визуализируется и количественно оценивается с помощью различных систем обнаружения, которые включают развитие окраски посредством фермент-субстратной реакции или связывания вторичного антитела, конъюгированного с флуоресцентным маркером или стафилококковым белком A / G, с первичным антителом. связаны с антигеном.Эти анализы можно использовать для измерения антител, которые реагируют на внутренние вирусные белки, структурные или ферментативные, а также для оценки уровней антител, которые связываются с внешними белками, против которых направлены нейтрализующие антитела. Напротив, современные анализы нейтрализации вирусов — это клеточные анализы, которые определяют функциональную активность антител в сыворотке или спинномозговой жидкости против живого вируса. Механизм нейтрализации вируса в культуре клеток (анализ in vitro) зависит от взаимодействия между эпитопом вириона и паратопом (специфическим сайтом противодействия) антитела, а также может зависеть от рецепторных характеристик клеток, на которых проводится анализ. выполняется [15].Имеющиеся данные указывают на то, что вирус инфицирует клетки нейронального происхождения, используя рецепторы никотинового ацетилхолинового рецептора, рецептор низкоаффинного фактора роста нервов или молекулу адгезии нервных клеток, одновременно инфицируя клетки, не относящиеся к ЦНС, с помощью других неидентифицированных рецепторов [16], [16] [ 17]. Результат анализа нейтрализации вируса основан на измерении роста вируса в культуре клеток, то есть на определении того, ускользает ли вирус от нейтрализации или нет. С другой стороны, анализы связывания измеряют другой набор характеристик антирабического ответа Ig по сравнению с анализами нейтрализации.Следовательно, результаты анализов связывания априори не могут быть напрямую сопоставимы с результатами нейтрализации [7]. При соответствующей валидации и пригодности для намеченной цели анализы связывания можно использовать для определения наличия или отсутствия антиген-специфических антител, а в некоторых случаях подклассов Ig, которые можно использовать в качестве приближения или подтверждения ответа нейтрализующих антител. Цель тестирования определит наиболее подходящие методы. Например, обнаружение специфических антител к вирусу бешенства в спинномозговой жидкости является диагностикой инфекции бешенства, независимо от того, является ли проведенный тест одним из анализов связывания (например, для фиксированного целого вируса на слайде, который обнаруживается с помощью антивидовых IgG или IgM) или анализ нейтрализации (например, RFFIT).Активность антител, обнаруженная в сыворотке крови, может указывать на предшествующую вакцинацию или, при наличии клинических признаков бешенства, на активную инфекцию или даже, в очень редких случаях, на предшествующий контакт с вирусом бешенства, который не привел к клинической инфекции [18] или к выживанию. заражения индивидуумом [19]. Чтобы полностью определить иммунитет к бешенству, требуется высокоспецифический анализ и способность обнаруживать чрезвычайно низкие уровни антител, а часто и специфических подклассов, таких как IgM и IgG. Для достижения этих требований проще разработать анализы связывания за счет использования специфических, чистых антигенов и надежных систем обнаружения (считывания).Тем не менее, мера защиты от заражения бешенством лучше всего определяется тестом на нейтрализацию вируса. Поскольку экспериментальные методы заражения вирусом никогда не будут применяться на людях, используются суррогатные экспериментальные модели защиты на животных, основанные на полевых наблюдениях и количестве и продолжительности действия нейтрализующих антител в сыворотке, измеренных методами in vitro.
Изменяя различные компоненты анализа, такие как штамм вируса, систему обнаружения и т. Д., Можно разработать индивидуальный дизайн анализа, «соответствующий назначению».Например, обнаружение РВНК в конкретной колонии видов летучих мышей, ассоциированных с вариантом вируса бешенства, может быть «специфически» измерено с использованием в анализе in vitro предполагаемого варианта вируса бешенства, циркулирующего среди летучих мышей, в качестве заражающего вируса. Чувствительность метода тестирования можно регулировать, варьируя количество антигена или дозы контрольного вируса для более точного определения и определения низких уровней антител или наличия неспецифических или перекрестно реагирующих антител или веществ.Для анализов связывания, таких как твердофазный иммуноферментный анализ (ELISA), меченый анти-IgM в качестве вторичного антитела обнаруживает только антитела к вирусу бешенства IgM в качестве первичного антитела. Уровень анти-IgM можно сравнить с уровнем анти-IgG в том же образце, выполнив тот же анализ, но с использованием меченого анти-IgG в качестве вторичного антитела. В конкурентном методе ELISA используется конкурирующее моноклональное антитело к эпитопу белка бешенства и измеряется способность антител в тестовом образце (сыворотки или спинномозговой жидкости) конкурировать с моноклональным антителом за прикрепление к этому конкретному эпитопу [20].Это делает анализ очень специфичным для измерения антител, которые связываются со специфическим эпитопом на данном белке бешенства [21]. Кроме того, можно внести изменения в методы анализа, чтобы упростить их выполнение и стандартизацию для использования в самых разных условиях и лабораториях. Например, молекулярные методы с использованием псевдотипированных вирусов в качестве векторов, таких как лентивирусная векторная система, для экспрессии гликопротеина бешенства, имеют несколько преимуществ. Они обеспечивают возможность (а) стандартизировать целевой эпитоп; (б) тест на нейтрализацию против множества штаммов или генотипов вирусов; (c) использовать более низкий уровень практики биологической опасности (т.д., делая их потенциально более безопасными, чем использование живого вируса бешенства), и (г) повысить экономичность методов тестирования [22]. Вот некоторые из факторов, которые следует учитывать при выборе метода, который «соответствует назначению».
Определение порогового значения (т. Е. Точки, указывающей на сероконверсию или определение адекватной вакцинации) специфично для каждого метода тестирования и имеет решающее значение для интерпретации результатов. В общем, титр нейтрализации сыворотки 1–100 (титр конечной точки 90%) приемлем как эффективный, даже если уровни антител в ткани могут быть ниже [23].Признанный во всем мире титр (уровень) антител 0,5 МЕ (международных единиц) / мл был впервые упомянут в Восьмом отчете Комитета экспертов Всемирной организации здравоохранения (ВОЗ) по бешенству, 1992 г. Что касается предконтактной вакцинации. в отчете говорится: «Всем лицам, работающим с живым вирусом бешенства в диагностической, исследовательской лаборатории или лаборатории по производству вакцин, следует сдать образец сыворотки, проверенный на антитела, нейтрализующие вирус бешенства, и ввести бустер, когда титр опускается ниже 0.5 МЕ / мл ». В отчете также рекомендуется, чтобы уровень РВНК определялся MNT или RFFIT с использованием обычного штамма контрольного вируса [24]. Понятно, что уровень 0,5 МЕ / мл был установлен как показатель адекватной вакцинации (не защиты!) У людей, подверженных риску заражения бешенством, и что он относится к использованию теста нейтрализации сыворотки с использованием стандартного контрольного штамма вируса. В США рекомендации Консультативного комитета по практике иммунизации (ACIP) гласят: «Если титр опускается ниже минимально допустимого уровня полной нейтрализации антител при разведении сыворотки 1-5, однократная бустерная доза вакцины перед воздействием. рекомендуется для людей с постоянным или частым риском заражения бешенством »(минимально допустимый уровень антител не определяется в МЕ / мл) и рекомендовать метод RFFIT для тестирования [25].Следовательно, когда используются другие методы для измерения специфических антител к бешенству у людей или когда RFFIT применяется для других видов (не людей), принятый уровень 0,5 МЕ / мл может не применяться. В обзоре исследований заражения бешенством были предложены допустимые пороговые значения для RFFIT у кошек и собак, равные 0,1 и 0,2 МЕ / мл соответственно [2]. Уровень 0,5 МЕ / мл методами RFFIT или FAVN признан регулирующими органами из большинства свободных от бешенства территорий как доказательство адекватного ответа на вакцинацию при ввозе кошек и собак [26].Когда сравниваются результаты тестирования сыворотки енота с помощью RFFIT и коммерческого ветеринарного набора для ELISA, аналогичные показатели чувствительности и специфичности получаются только при использовании разных пороговых значений (см. Таблицу 1). Для сывороток енотов, протестированных RFFIT, установка порогового уровня 0,5 МЕ / мл допускает возможность присутствия неспецифических ингибиторов в сыворотках диких животных, что может дать ложноположительные результаты, если также установлен пороговый уровень. низкий. Часто метод ELISA менее чувствителен к мешающим веществам, потому что разведение сыворотки, используемое в анализе, обычно выше.Измерения выше 0,1 ЕС (эквивалентных единиц) / мл кажутся специфичными для этого набора образцов с использованием этого конкретного набора для тестирования. При мониторинге поглощения наживки енотами в программах оральной наживки важнее использовать метод оценки уровней потенциального иммунитета популяции, чем определение степени защиты у отдельного енота. Использование разных пороговых значений в исследованиях при сравнении результатов, полученных с помощью одного и того же метода, может привести к неверным выводам. Таким образом, анализ и пороговое значение, позволяющее отличить вакцинированных енотов от невакцинированных, становятся наилучшим «подходящим» методом.Точно так же уровень отсечения является значимым в исследовании реакции человека на вакцину против бешенства, в котором сравниваются три метода ELISA с методом RFFIT с использованием порогового значения сероконверсии 0,5 МЕ / мл [27]. Расчеты и, следовательно, выводы, полученные на основе результатов RFFIT сероконверсии, будут варьироваться в зависимости от используемого порогового уровня, либо 0,5 МЕ / мл, либо принятого ACIP уровня полной нейтрализации при разведении 1-5. Например, определение показателей чувствительности и специфичности методов ELISA по сравнению с результатами RFFIT, интерпретируемыми как отрицательные или положительные для сероконверсии по уровню ACIP, а не 0.Уровень 5 МЕ / мл, используемый в статье, даст разные значения [27]. В этих двух примерах соответствующее пороговое значение должно определяться и индивидуально для каждого метода, а также в соответствии с целью тестирования и нормативными требованиями. Помимо установления соответствующих пороговых уровней для конкретных методов, регулирующие органы, такие как Европейская фармакопея, USFDA или национальные органы по импорту в районах, свободных от бешенства, могут также потребовать лабораторную валидацию и одобрение или сертификацию.
Стандартизация и проверка методов
Стандартизация методов серологических тестов на бешенство имеет важное значение для обеспечения значимого сравнения характеристик и активности антител у вакцинированных или подвергшихся воздействию людей. Это становится особенно актуальным, когда результаты серологического исследования бешенства сравниваются между различными лабораториями и разными исследованиями с течением времени. Тест нейтрализации вируса флуоресцентными антителами (FAVN) был разработан, в частности, для стандартизации различных модификаций используемого RFFIT [28].Переменные в тесте, которые включают контрольный вирус, антитела, клетки-мишени и пороговые значения, должны быть стандартизированы, чтобы результаты теста были сопоставимы. Например, с цельным вирусом бешенства по сравнению с гликопротеином вируса бешенства в качестве антигена в непрямом ИФА-тесте, где все другие переменные стандартизированы, нельзя ожидать, что результаты будут сопоставимыми. В большинстве тестовых образцов большая часть анализируемых антител к вирусу бешенства будет направлена против гликопротеина, но антитела к нуклеопротеиновым и фосфопротеиновым антигенам вируса не редкость и будут обнаружены с помощью непрямого ИФА на основе всего вируса [29]. .При измерении эффективности моноклональных антител важно учитывать, что специфичности антитела к одному эпитопу может быть недостаточно для нейтрализации всех эпитопных (антигенных) вариаций штамма контрольного вируса или даже ряда квазивидов в пределах одного запас вируса [30], [31]. Точно так же различные штаммы контрольного вируса могут влиять на результаты, полученные при тестировании нейтрализации сыворотки. В исследовании с участием людей, которое сравнивало титры сыворотки, измеренные in vitro, против штаммов контрольного вируса, которые были либо гомологичны, либо гетерологичны вакцинному штамму, показало, что у большинства субъектов были обнаружены более высокие титры против гомологичного штамма [32].Кроме того, как высокие, так и низкие дозы заражающего вируса могут влиять на определение активности иммуноглобулина бешенства [33]. Использование признанного стандартного контроля сыворотки против бешенства в качестве эталона стандартизирует результаты метода и позволит преобразовать измерения (например, титры, показания оптической плотности и т. Д.) В признанные единицы, такие как МЕ / мл или ЕС. / мл.
Международные эталоны для продуктов RIG, которые используются сегодня, включают первый международный стандарт ВОЗ на иммуноглобулин против бешенства, второй международный стандарт ВОЗ на иммуноглобулин против бешенства и эталонную сыворотку RIG для собак, разработанную Международным бюро эпидемиологии (МЭБ).Эффективность каждого из этих продуктов была оценена с помощью методов нейтрализации сыворотки [34] — [36]. Первоначальная международная стандартная эталонная сыворотка RIG лошадиного происхождения была создана в 1955 году. Исходя из веса лиофилизированного продукта 86,6 мг на ампулу, было присвоено значение активности 86,6 МЕ. Первая эталонная сыворотка ВОЗ для RIG человеческого происхождения была приготовлена из объединенных сывороток вакцинированных людей. Эффективность RIG для человека была установлена межлабораторным тестированием по сравнению со стандартным RIG для лошадей в RFFIT, проведенном в шести лабораториях.Тест включал два разведения продукта (низкое и высокое) в четырех повторных анализах. После статистического анализа, в 1984 г. эффективность 59 МЕ была отнесена к первому международному стандарту ВОЗ для RIG. Аналогичным образом второй пул сывороток вакцинированных людей был протестирован в 1993 году, ему была присвоена эффективность 30 МЕ, и он стал вторым международным стандартом ВОЗ для эталонной сыворотки RIG. Эталонная сыворотка RIG, используемая в США, Lot R-3, является частью партии, которая стала первым международным стандартом ВОЗ для эталонной сыворотки RIG.Эталонная сыворотка RIG для собак МЭБ имеет эффективность 6,7 МЕ, и партии этого продукта откалиброваны по второму международному стандарту ВОЗ для эталонной сыворотки RIG. Относительная эффективность этих двух референсных сывороток ВОЗ человека RIG сравнивалась USFDA в 1997 г. и снова Лабораторией борьбы с бешенством Университета штата Канзас в 2006 г. Это сравнение показало снижение относительной активности первого международного стандарта ВОЗ по отношению к эффективности ВОЗ второй международный стандарт 2.5% в 1997 г. и 14% в 2006 г. Эти различия не считаются статистически значимыми, но они иллюстрируют эффект использования различных эталонных стандартов при сравнении результатов серологического исследования бешенства и важность мониторинга и стандартов контроля качества.
Использование значений эффективности для RIG, которые назначаются методами нейтрализации сыворотки, в отличие от значений, установленных с помощью анализов связывания, неуместно и проблематично. На рисунке 1 показаны расхождения, которые могут возникнуть, когда различные контрольные сыворотки RIG с одинаковой эффективностью, определенной путем нейтрализации сыворотки, используются для расчета значений активности, полученных методом ELISA.Когда первая и вторая эталонные сыворотки ВОЗ RIG разводятся до 2,0 МЕ / мл для получения аналогичных значений активности с помощью RFFIT, результаты ELISA расходятся. Таким образом, если первый RIG ВОЗ используется в качестве эталонного контроля для стандартной кривой в анализе ELISA, будут получены более низкие результаты, чем при использовании второго RIG ВОЗ.
Рис. 1. Международные стандартные эталонные сыворотки для RIG.
Первая SRIG ВОЗ и вторая SRIG ВОЗ были разбавлены до 2,0 МЕ / мл в соответствии с указанной эффективностью. Препараты SRIG оценивали в трех независимых тестовых партиях методами RFFIT и ELISA (Bio-Rad Platelia Rabies Kit II).Значение МЕ / мл рассчитывалось по первому SRIG ВОЗ, а ЕС / мл — по стандарту набора. Отображаются средние значения МЕ / мл или ЕС / мл с одним стандартным отклонением.
https://doi.org/10.1371/journal.pntd.0000595.g001
Расчет «метода результата» является дополнительным фактором, который следует учитывать при стандартизации анализа. Например, использование титров конечной точки 50% или 100%, оба из которых могут быть рассчитаны для анализов нейтрализации сыворотки, даст разные значения титра.Для любого сравнения результатов важно знать и всегда указывать, какой расчет использовался для получения значений титра.
Выводы
Важно, чтобы характеристики и вариабельность, связанная с различными методами анализа, которые определяют титры антител, продолжали определяться по мере того, как мы продвигаемся в нашем понимании иммунитета и профилактики заболеваний. Измерение антител, специфичных к бешенству, in vitro имеет важное значение и должно быть первым шагом для установления успешности иммунитета против бешенства после вакцинации.Даже в этом случае измерения in vitro могут никогда полностью не коррелировать с тем, что можно считать защитным in vivo. Даже использование «золотого стандарта» измерения нейтрализующей активности вируса бешенства in vitro обеспечивает только оценку защиты. Несмотря на долгую историю использования тестов на нейтрализацию вируса, до сих пор не существует международно признанных стандартных протоколов для измерения антирабических антител, для конкретного определения активности RIG для составов биопрепаратов против бешенства человека или для оценки поглощения оральной вакцины против бешенства дикой природой.Стандартизация методов требует тщательного изучения и оценки результатов лабораторных испытаний, которые могут включать аудиты лабораторных операций, публикацию и совместное использование стандартных операционных процедур, а также меж- и внутрилабораторную оценку квалификации при проведении тестов. Проверка квалификации, обучение и сертификация, а также тенденции в выполнении анализов являются компонентами программ обеспечения качества, чтобы гарантировать постоянное соблюдение признанных и принятых стандартов. Эти усилия потребуют сотрудничества всех организаций, проводящих тесты, включая национальные лаборатории, регулирующие органы, коммерческие компании, а также диагностические и исследовательские лаборатории.
Для смертельного заболевания, такого как бешенство, где вакцинация и пассивный иммунитет (использование HRIG) абсолютно необходимы для защиты от воздействия, жизненно важна проверка точности и эффективности каждого анализа, связанного с прогнозированием соответствующей защиты от вируса in vivo. Более того, существует основное требование для подтверждения пригодности и применимости любой используемой тест-системы нейтрализации или анализа связывания антигена в качестве предиктора защиты in vivo с каждым новым типом профилактического биологического средства, будь то вакцина, Ig или моноклональные антитела. формулировка.В случае вакцин крайне важно понимать, как антитела нейтрализуют вирусную инфекционность, чтобы внести свой вклад в разработку и представление иммуногенов вакцины, которые выявляют специфичность и изотип продуцируемых антител, чтобы обеспечить максимальную нейтрализацию и, в конечном итоге, защитную способность. Мероприятия.
Вставка 1. Очки обучения
1. Было продемонстрировано, что РВНК имеет решающее значение для защиты от бешенства. Даже в этом случае измерения in vitro — это лишь частичное определение степени защиты, обеспечиваемой in vivo.Не все методы измерения антител, специфичных к бешенству, определяют нейтрализующую функцию антител.
2. При выборе наиболее подходящего анализа для обнаружения антител к бешенству рассмотрение цели и использования результатов так же важно, как и установленные рабочие характеристики анализа.
3. Стандартизация анализов включает как компоненты анализа, так и условия испытаний. Изменения приведут к изменению результатов; поэтому использование определенного анализа (например,g., ELISA или нейтрализация сыворотки) не гарантирует сопоставимых результатов, если анализы не были стандартизированы.
4. Поскольку бешенство является смертельным заболеванием, для которого развитие достаточного ответа РВНК имеет первостепенное значение для защиты, специфичность, чувствительность и точность анализа должны быть проверены для принятия значимых клинических решений на основе результатов.
5. Шаги к лучшему пониманию и использованию серологических тестов на бешенство будут включать сотрудничество национальных лабораторий, регулирующих органов, а также коммерческих и исследовательских лабораторий.Более тесное сотрудничество и стандартизация серологических анализов на бешенство приведет к лучшему пониманию взаимосвязи между измерением in vitro и защитой in vivo.
Вставка 2. Ключевые ссылки в поле
1. Smith JS, Yager PA, Baer GM (1973) Быстрый воспроизводимый тест для определения нейтрализующих бешенство антител. Bull World Health Organ 48: 535–541.
2. Cliquet F, Aubert M, Sagne L (1998) Разработка теста нейтрализации вируса флуоресцентными антителами (тест FAVN) для количественного определения антител, нейтрализующих бешенство.J Immunol Methods 212: 79–87.
3. Обер М.Ф. (1992) Практическое значение антител против бешенства у кошек и собак. Rev Sci Tech 11: 735–760.
4. Grassi M, Wandeler AI, Peterhans E (1989) Иммуноферментный анализ для определения антител к гликопротеину оболочки вируса бешенства. J Clin Microbiol 27: 899–902.
5. Ири Т., Каваи А. (2002) Исследования различных условий нейтрализации вируса бешенства моноклональными антителами № 1-46-12 и № 7-1-9.J Gen Virol 83: 3045–3053.
Нейтрализация вируса антителами
Антительный ответ имеет решающее значение для предотвращения многих вирусных инфекций и может также способствовать разрешению инфекции. Когда позвоночное животное инфицировано вирусом, вырабатываются антитела против многих эпитопов на нескольких вирусных белках. Подмножество этих антител может блокировать вирусную инфекцию с помощью процесса, который называется нейтрализацией .
Антитела могут нейтрализовать вирусную инфекционность несколькими способами, как показано на иллюстрации.Они могут препятствовать связыванию вириона с рецепторами, блокировать захват клетками, предотвращать снятие оболочки геномов в эндосомах или вызывать агрегацию вирусных частиц. Многие вирусы в оболочке лизируются, когда противовирусные антитела и сывороточный комплемент разрушают мембраны.
Нейтрализующие антитела также образуются после вирусной инфекции. Такие антитела специфически связываются с вирусными частицами, но не нейтрализуют инфекционность. Они могут повышать инфекционность, поскольку антитела могут взаимодействовать с рецепторами макрофагов.Весь комплекс вирус-антитело попадает в клетку посредством эндоцитоза. Затем может продолжаться репликация вируса, поскольку антитело не блокирует инфекционность. Этот путь может позволить проникнуть в клетки, которые обычно не несут специфические вирусные рецепторы.
Где в организме происходит нейтрализация вирусов антителами? Вирионы, инфицирующие поверхности слизистой оболочки, сталкиваются с секреторными антителами IgA, присутствующими на апикальных поверхностях эпителиальных клеток. Вирусы, которые распространяются в крови, будут подвергаться воздействию антител IgG и IgM.Фактически, тип продуцируемого антитела может повлиять на исход вирусной инфекции. Инфекция полиовирусом вызывает ответы IgM и IgG в крови, но IgA слизистой оболочки жизненно важен для блокирования инфекции. Это антитело может нейтрализовать полиовирус в кишечнике, в очаге первичной инфекции. Живая аттенуированная вакцина против полиовируса Сабина эффективна, потому что она вызывает сильный иммунный ответ слизистой оболочки и обеспечивает иммунитет кишечника. Напротив, введенная вакцина против полиомиелита (Салк) не вызывает кишечного иммунитета и, следовательно, менее эффективна для предотвращения распространения полиовируса среди населения.
Подавляющее большинство вакцин против гриппа вводятся путем инъекции и стимулируют выработку антител IgG; они являются плохими индукторами антител IgA слизистой оболочки. Эффективность противогриппозных вакцин, вероятно, значительно повысилась бы, если бы они были разработаны для стимуляции иммунитета слизистых оболочек. Флумист — это лицензированная вакцина против инфекционных вирусов для интраназального введения, которая, как было доказано, стимулирует как слизистые, так и системный иммунитет. Было проведено девять клинических испытаний на детях, сравнивающих эффективность Флумиста с инактивированной вакциной или плацебо.Анализ результатов показывает, что вакцина, вводимая интраназально, более эффективна в профилактике гриппа. Эти результаты подчеркивают роль местного иммунитета в устойчивости к респираторным патогенам.
Рорер, Дж., Амброуз, К., Дикинсон, С., Гамильтон, Х., Олека, Н., Малиноски, Ф., и Виттес, Дж. (2009). Эффективность живой аттенуированной вакцины против гриппа у детей: метаанализ девяти рандомизированных клинических испытаний вакцины, 27 (7), 1101-1110 DOI: 10.1016 / j.vaccine.2008.11.093
Нейтрализующие антитела коррелируют с защитой от SARS-CoV-2 у людей во время вспышки болезни на рыболовных судах с высокой частотой атаки
Abstract
Разработка вакцин против SARS-CoV-2 будет значительно облегчена путем определения иммунологических коррелятов защиты в людях. Однако до настоящего времени исследования защитного иммунитета проводились только на животных моделях, а корреляты защиты у людей не установлены. Здесь мы описываем вспышку SARS-CoV-2 на рыболовном судне, связанную с высоким уровнем атак.Серологические и вирусные ОТ-ПЦР перед отъездом и повторное тестирование после возвращения на берег были доступны для 120 из 122 человек на борту в течение среднего периода наблюдения 32,5 дня (диапазон от 18,8 до 50,5 дней). В общей сложности 104 человека имели положительный вирусный тест с помощью ОТ-ПЦР с Ct <35 или сероконверсией в течение периода последующего наблюдения, что дало показатель атаки на борту 85,2% (104/122 человека). Метагеномное секвенирование 39 вирусных геномов показало, что вспышка возникла в основном из-за одной вирусной клады.Только три члена экипажа дали серопозитивный результат до отплытия лодки при первоначальном серологическом скрининге, а также имели нейтрализующие и спайк-реактивные антитела в последующих анализах. Ни у одного из этих членов экипажа с титрами нейтрализующих антител не было доказательств истинной вирусной инфекции и не было никаких симптомов во время вирусной вспышки. Таким образом, присутствие нейтрализующих антител от предшествующей инфекции было в значительной степени связано с защитой от повторного заражения (точный тест Фишера, p = 0.002).
Заявление о конкурирующем интересе
Авторы заявили об отсутствии конкурирующего интереса.
Отчет о финансировании
Работа, выполненная в клинической лаборатории, была поддержана Департаментом лабораторной медицины и патологии. Это исследование было поддержано следующими грантами NIAID NIH: R01AI141707 (для J.D.B.) и F30AI149928 (для K.H.D.C.). J.D.B. является исследователем Медицинского института Говарда Хьюза.
Заявления авторов
Я подтверждаю, что были соблюдены все соответствующие этические принципы и получены все необходимые разрешения IRB и / или комитета по этике.
Да
Подробная информация об IRB / надзорном органе, предоставившем разрешение или исключение для описанного исследования, приводится ниже:
Это исследование было одобрено IRB Вашингтонского университета.
Получено все необходимое согласие пациента / участника, а соответствующие институциональные формы заархивированы.
Да
Я понимаю, что все клинические испытания и любые другие проспективные интервенционные исследования должны быть зарегистрированы в одобренном ICMJE реестре, таком как ClinicalTrials.губ. Я подтверждаю, что любое такое исследование, указанное в рукописи, было зарегистрировано и предоставлен идентификатор регистрации исследования (примечание: при публикации проспективного исследования, зарегистрированного ретроспективно, просьба предоставить заявление в поле идентификатора исследования, объясняющее, почему исследование не было зарегистрировано заранее) .
Да
Я соблюдал все соответствующие руководящие принципы отчетности по исследованиям и загрузил соответствующие контрольные списки отчетов по исследованиям сети EQUATOR и другие соответствующие материалы в качестве дополнительных файлов, если применимо.
Да
Доступность данных
Данные в дополнительных таблицах
3-ct789
% PDF-1.6 % 228 0 объект > эндобдж 138 0 объект > эндобдж 572 0 объект > поток 2007-07-28T14: 13: 08ZAdobe PageMaker 6.522007-09-10T12: 42: 39 + 02: 002007-09-10T12: 42: 39 + 02: 00application / pdf
