Как работает микросхема КА1016ХЛ1. Каковы особенности ее включения. Какие схемы часов можно собрать на ее основе. Как решить типичные проблемы при работе с КА1016ХЛ1. Где купить эту микросхему сегодня.
Основные характеристики микросхемы КА1016ХЛ1
КА1016ХЛ1 — это специализированная микросхема для построения электронных часов, разработанная в СССР в 1980-х годах. Ее ключевые особенности:
- Питание: 11-15 В постоянного тока
- Потребляемый ток: не более 15 мА
- Частота кварцевого резонатора: 32768 Гц
- Выходы для управления 4-разрядным индикатором
- Функции часов, будильника и секундомера
- Возможность коррекции хода часов
Типовая схема включения КА1016ХЛ1
Для работы микросхемы КА1016ХЛ1 требуется минимум внешних компонентов:
- Кварцевый резонатор на 32768 Гц
- Подстроечный конденсатор 3-30 пФ для точной настройки частоты
- Кнопки управления (установка времени, будильника)
- Индикатор (обычно вакуумный люминесцентный)
- Резисторы и конденсаторы по схеме
Важно обеспечить стабильное питание 15 В и правильно подключить выходы микросхемы к индикатору. При этом следует учитывать особенности конкретного типа индикатора.

Применение КА1016ХЛ1 в электронных часах
Микросхема КА1016ХЛ1 широко применялась в различных конструкциях бытовых и автомобильных электронных часов:
- Настольные часы с вакуумным люминесцентным индикатором
- Автомобильные часы со светодиодной индикацией
- Часы-будильники с дополнительными функциями
- Часы с резервным питанием от батареи
Популярность КА1016ХЛ1 была обусловлена простотой схемы и доступностью микросхемы для радиолюбителей. На ее основе выпускались радиоконструкторы для самостоятельной сборки часов.
Особенности работы с вакуумными люминесцентными индикаторами
Наиболее часто КА1016ХЛ1 применялась в сочетании с вакуумными люминесцентными индикаторами типа ИВЛ1-7/5 или УИ-4. При этом необходимо учитывать следующие моменты:
- Требуется дополнительное напряжение питания индикатора -40 В
- Нужно обеспечить напряжение накала 2-3 В переменного тока
- Важно правильно подключить сетки и сегменты индикатора
- Желательно использовать подтягивающие резисторы на выходах микросхемы
При соблюдении этих условий можно добиться яркого и контрастного отображения цифр на индикаторе. Однако вакуумные индикаторы имеют ограниченный срок службы и со временем теряют яркость.
Применение КА1016ХЛ1 со светодиодными индикаторами
Хотя изначально КА1016ХЛ1 предназначалась для работы с вакуумными индикаторами, ее можно успешно применять и со светодиодными. Для этого требуется:
- Использовать буферные каскады на транзисторах или микросхемах
- Обеспечить динамическую индикацию для снижения энергопотребления
- Подобрать яркие светодиодные индикаторы подходящего размера
- Предусмотреть регулировку яркости свечения
Такой вариант особенно актуален для автомобильных часов, где светодиодные индикаторы более надежны и долговечны, чем вакуумные.
Типичные проблемы при работе с КА1016ХЛ1 и их решение
При разработке и эксплуатации часов на КА1016ХЛ1 могут возникать следующие проблемы:
- Неточный ход часов — решается подстройкой частоты кварцевого генератора
- Сбои при включении питания — устраняются добавлением RC-цепочки сброса
- Быстрое уменьшение яркости индикатора — вызвано неправильным подключением, требуется доработка схемы
- Чувствительность к помехам по питанию — необходима качественная фильтрация напряжения питания
Большинство проблем решается правильным монтажом и соблюдением рекомендаций по применению микросхемы.
Современные аналоги КА1016ХЛ1
Сегодня микросхема КА1016ХЛ1 считается устаревшей и сложно найти ее в продаже. В качестве современных аналогов можно рассмотреть:
- Микроконтроллеры общего назначения (PIC, AVR, STM32)
- Специализированные микросхемы для часов (DS1307, PCF8563)
- Готовые модули часов реального времени
Однако многие радиолюбители по-прежнему ценят простоту и надежность КА1016ХЛ1 для создания ретро-конструкций часов.
Где купить микросхему КА1016ХЛ1 сегодня
Хотя КА1016ХЛ1 уже не выпускается, ее все еще можно найти:
- На радиорынках и у коллекционеров радиодеталей
- На интернет-площадках объявлений (Авито, Юла)
- В некоторых интернет-магазинах радиодеталей
- На AliExpress (часто под обозначением КР1016ВИ1)
При покупке стоит учитывать, что микросхемы могут быть старыми и требовать проверки работоспособности. Цена обычно составляет 100-300 рублей за штуку.
Часы на микросхеме КА1016ХЛ1 и светодиодных индикаторах
РадиоКот >Схемы >Аналоговые схемы >Бытовая техника >Часы на микросхеме КА1016ХЛ1 и светодиодных индикаторах
Каких только конструкций электронных часов не встретишь сегодня на просторах Интернета! В основном они собраны на микроконтроллерах, и кроме функций отображения времени и будильника могут ещё отображать дату, день недели, температуру и т.д., в том числе с различными визуальными эффектами. В качестве дисплея могут выступать светодиодные, люминесцентные, газоразрядные, жидкокристаллические индикаторы и даже механически вращающиеся поверхности накопителей на жёстких дисках и лопастей вентиляторов. Несмотря на доступность повторения таких конструкций появилось желание вдохнуть новую жизнь в старые часы на микросхеме КА1016ХЛ1, собранные когда-то из набора радиоконструктора «Старт 2035», которые прослужили верой и правдой довольно длительное время. К сожалению (или к счастью), используемый в них «подсевший» от времени люминесцентный индикатор УИ-4 уже не найти в продаже, поэтому придётся заменить его на что-то другое.
Появилось желание ради спортивного интереса собрать часы на этой микросхеме, но на светодиодных индикаторах, с автоматической регулировкой их яркости и источником бесперебойного питания. Конечно, проще и дешевле было бы купить готовые часы, но захотелось выяснить, что же можно «выжать» из этой микросхемы. Ниже описано, что получилось в итоге.
Схема включения микросхемы КА1016ХЛ1 в целом не отличается от схем известных конструкций, описанных в книгах: [В. Борисов. Электронные часы из деталей радиоконструктора. В помощь радиолюбителю. Выпуск 106. с.39-49] и [С.А.Бирюков. Электронные часы на МОП интегральных микросхемах.
 Поэтому было решено разъединить катоды индикаторов резисторами R17..R44 (их теперь стало 28 штук). Но при этом максимально допустимый ток через транзисторные ключи VT1..VT8 увеличился до 4 раз (при отображении четырёх одинаковых цифр), и применение цифровых транзисторов с максимальным током 100 мА стало невозможным. Теперь вместо них установлены более мощные BC817 с внешними резисторами в базовых цепях R1..R16 (раньше эти резисторы были встроены в состав цифровых транзисторов). Такая доработка потребовала изготовления новой печатной платы (но на плате последней ревизии сохранена возможность установки 8 ограничительных резисторов вместо 28, что может быть полезным для экспериментов по подбору их номинала с целью получения требуемой яркости свечения — ведь 8 резисторов легче заменить, чем 28). Примерные номиналы ограничительных резисторов для индикаторов различных цветов приведены в таблице на принципиальной схеме (самые лучшие с точки зрения энергопотребления — ярко-зелёные индикаторы, худшие — красные).
Поэтому было решено разъединить катоды индикаторов резисторами R17..R44 (их теперь стало 28 штук). Но при этом максимально допустимый ток через транзисторные ключи VT1..VT8 увеличился до 4 раз (при отображении четырёх одинаковых цифр), и применение цифровых транзисторов с максимальным током 100 мА стало невозможным. Теперь вместо них установлены более мощные BC817 с внешними резисторами в базовых цепях R1..R16 (раньше эти резисторы были встроены в состав цифровых транзисторов). Такая доработка потребовала изготовления новой печатной платы (но на плате последней ревизии сохранена возможность установки 8 ограничительных резисторов вместо 28, что может быть полезным для экспериментов по подбору их номинала с целью получения требуемой яркости свечения — ведь 8 резисторов легче заменить, чем 28). Примерные номиналы ограничительных резисторов для индикаторов различных цветов приведены в таблице на принципиальной схеме (самые лучшие с точки зрения энергопотребления — ярко-зелёные индикаторы, худшие — красные).
Для управления яркостью индикаторов сначала планировалось использовать метод широтно-импульсной модуляции на основе таймера NE555 и фоторезистора. При уменьшении освещённости сопротивление фоторезистора увеличивалось, и возрастала скважность формируемых импульсов, которые управляли подачей питания на коллекторы анодных ключей VT9..VT12. Однако, при проверке этого метода оказалось, что импульсы с выхода таймера идут вразнобой с сеточными импульсами, и вместо регулировки яркости получается «мельтешение» разрядов, особенно при малой освещённости. Чтобы не усложнять уже и без того «навороченную» схему, регулировку яркости было решено сделать простым плавным изменением уровня питающего напряжения на коллекторах VT9..VT12 через мощный транзистор VT13 с достаточно большим коэффициентом усиления. За счёт импульсного характера управления индикаторами рассеиваемая узлом мощность оказалась незначительной.
За основу источника бесперебойного питания (UPS), показанного на основной схеме часов, взят Бесперебойник для часов, который показал наилучшие результаты по сравнению с другими конструкциями: Преобразователь напряжения 1,5 — 9 вольт и DC-DC преобразователь 1.2-9 вольт. Номиналы деталей изменены таким образом, чтобы обеспечить нормальное функционирование микросхемы КА1016ХЛ1. Резистор 2R2 закрывает транзистор 2VT1 при появлении внешнего питания, прерывая генерацию и исключая разряд аккумулятора. Последний подзаряжается через резистор 2R1, номинал которого зависит от ёмкости применённого аккумулятора (меньший номинал — для большей ёмкости). Вместо светодиода установлен обычный диод 2VD2, отключающий от аккумулятора лишние цепи при пропадании внешнего питания. Стабилитрон 2VD1 с напряжением стабилизации 2В служит для защиты от перенапряжения при работе без нагрузки с отсоединённым аккумулятором, ограничивая в этом режиме неконтролируемый рост напряжения на базе транзистора 2VT1 и, как следствие, выходного напряжения преобразователя. В то же время стабилитрон практически не нагружает аккумулятор, когда он подключён. Микросхема КА1016ХЛ1 категорически отказывается работать при напряжении питания меньше 12 вольт, хотя в некоторых работах утверждается, что она работает при напряжении питания от 8 до 18 вольт. Замечено, что увеличение напряжения питания выше уровня 15 вольт, даже в виде импульсов, приводит к выводу микросхемы из строя. Поэтому выходное напряжение источника бесперебойного питания выбрано порядка 13,5..13,7 вольт при работе от аккумулятора и 14,5..14,7 вольт – при работе от внешнего питания, что гарантирует надёжную работу часов в любом режиме. Источник бесперебойного питания собран на отдельной плате вместе с дополнительными элементами блока питания, не показанными на основной схеме:
Стабилитрон 2VD1 с напряжением стабилизации 2В служит для защиты от перенапряжения при работе без нагрузки с отсоединённым аккумулятором, ограничивая в этом режиме неконтролируемый рост напряжения на базе транзистора 2VT1 и, как следствие, выходного напряжения преобразователя. В то же время стабилитрон практически не нагружает аккумулятор, когда он подключён. Микросхема КА1016ХЛ1 категорически отказывается работать при напряжении питания меньше 12 вольт, хотя в некоторых работах утверждается, что она работает при напряжении питания от 8 до 18 вольт. Замечено, что увеличение напряжения питания выше уровня 15 вольт, даже в виде импульсов, приводит к выводу микросхемы из строя. Поэтому выходное напряжение источника бесперебойного питания выбрано порядка 13,5..13,7 вольт при работе от аккумулятора и 14,5..14,7 вольт – при работе от внешнего питания, что гарантирует надёжную работу часов в любом режиме. Источник бесперебойного питания собран на отдельной плате вместе с дополнительными элементами блока питания, не показанными на основной схеме:
К таким дополнительным элементам относятся следующие дешёвые готовые модули: миниатюрный источник питания 220 AC – 5V 0,6A DC и преобразователь напряжения DC-DC MT3608, используемый для формирования напряжения 15В из напряжения 5В. Кроме того, на плате для питания часов предусмотрен разъём Micro USB (если планируется питание только через этот разъём, плату блока питания 5В можно не устанавливать).
Кроме того, на плате для питания часов предусмотрен разъём Micro USB (если планируется питание только через этот разъём, плату блока питания 5В можно не устанавливать).
Часы собраны на двух платах размером 100×50 мм, соединённых через втулки M3: на основной плате собственно часов и на плате блока питания с источником бесперебойного питания. Указанные размеры плат (не более 100 мм) позволяют недорого заказать их изготовление в Китае. Если плату блока питания ещё можно изготовить методом ЛУТ, то основную плату уже, наверное, не получится. Нужно заметить, что печатные платы и светодиодные индикаторы – самые дорогие элементы этой конструкции. Вид собранной основной платы часов снизу:
Некоторые радиоэлементы могут быть выводными и напаиваться на плату снизу, например: мощный ограничительный резистор R49, излучатель BA1 (можно использовать подходящий излучатель с сопротивлением катушки порядка 40 Ом или пьезоэлектрический, включив параллельно его выводам резистор номиналом 10 кОм), кварцевый резонатор ZQ1, подстроечный конденсатор C9, электролитический конденсатор C5:
На схеме предусмотрены элементы цепи сброса микросхемы VD6, VD7, C11, однако практика показала, что в них нет необходимости. Следует учитывать, что микросхема чувствительна к статическому напряжению, а также к остаткам флюса или другой жидкости (спирт, вода). Поэтому после монтажа перед включением плату нужно тщательно промыть и просушить феном для волос.
Следует учитывать, что микросхема чувствительна к статическому напряжению, а также к остаткам флюса или другой жидкости (спирт, вода). Поэтому после монтажа перед включением плату нужно тщательно промыть и просушить феном для волос.
Для включения и выключения будильника можно использовать подходящий кнопочный выключатель с фиксацией или ползунковый. Тактовые кнопки управления используются с общей высотой 15,5 мм, но можно использовать и другие подходящие выключатели без фиксации или с фиксацией.
Для передней панели подойдёт прозрачное оргстекло толщиной 2 мм, в качестве задней панели – такой же материал или непрозрачный пластик. Можно обклеить оргстекло светлой тонирующей автомобильной плёнкой, но в этом случае яркость свечения индикаторов, возможно, придётся увеличивать.
Вид собранной основной платы спереди:
Вид сбоку:
Один из вариантов готовой конструкции спереди:
и сзади:
Для защиты от пыли сверху можно зафиксировать крышку из тонкого прозрачного материала, например, плёнки, используемой для ламинирования.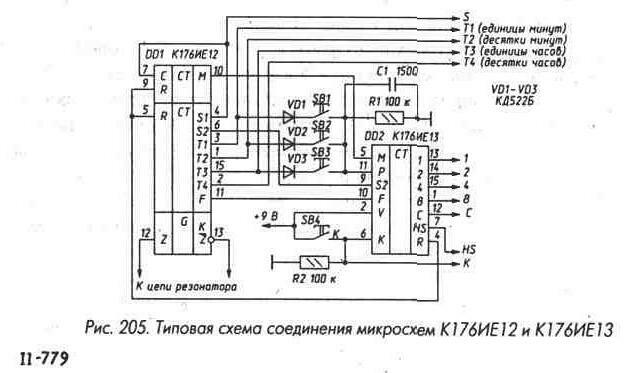 Такой же материал удобно использовать для защитного кожуха на плате блока питания, ограничивающего доступ к высоковольтным цепям:
Такой же материал удобно использовать для защитного кожуха на плате блока питания, ограничивающего доступ к высоковольтным цепям:
Вид платы блока питания сверху (показана старая версия платы, поэтому могут быть отличия):
и снизу:
Вместо перемычки 2J1 на плате блока питания можно использовать подходящий ползунковый переключатель:
Для проверки функционирования источника бесперебойного питания (пока без платы часов) нужно подключить предварительно заряженный аккумулятор (напряжение на нём должно быть не меньше 1,2..1,4 вольт), замкнув перемычку 2J1, и проконтролировать напряжение на выходе (U-рез; U+рез) — оно должно быть в указанных на схеме пределах. Затем подсоединить к контактам разъёма (U-рез; U+рез) нагрузку в виде резистора номиналом 10 кОм (выводы последнего можно вставить в отверстия гнездового разъёма) – выходное напряжение должно оставаться в допуске. Можно при этом проконтролировать напряжение на аккумуляторе – оно должно оставаться на уровне не ниже 1,2 вольт, т.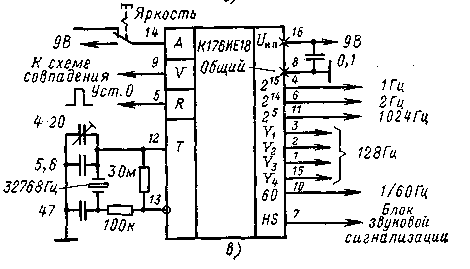 е. не должно «проседать» под нагрузкой. Предел работоспособности наступает при напряжении на аккумуляторе меньше порядка 1В — в этом случае напряжение на выходе преобразователя становится меньше 12 вольт, и микросхема КА1016ХЛ1 перестаёт работать. При проведении тестового «прогона» источника бесперебойного питания с аккумулятором б/у неизвестной ёмкости (из старой переносной телефонной трубки) микросхема КА1016ХЛ1 проработала без сбоев 8 часов! Необходимо помнить, что соединять платы между собой нужно при отключённом аккумуляторе.
е. не должно «проседать» под нагрузкой. Предел работоспособности наступает при напряжении на аккумуляторе меньше порядка 1В — в этом случае напряжение на выходе преобразователя становится меньше 12 вольт, и микросхема КА1016ХЛ1 перестаёт работать. При проведении тестового «прогона» источника бесперебойного питания с аккумулятором б/у неизвестной ёмкости (из старой переносной телефонной трубки) микросхема КА1016ХЛ1 проработала без сбоев 8 часов! Необходимо помнить, что соединять платы между собой нужно при отключённом аккумуляторе.
Органы управления часов:
При длительном отсутствии внешнего электропитания необходимо выключить источник резервного питания (UPS) перемещением движка переключателя вниз. После подачи питания перевести движок переключателя вверх.
Четыре экземпляра таких часов с разными цветами свечения индикаторов (показаны на фото в начале статьи), работают без проблем уже около года. В процессе монтажа таких часов можно отрабатывать навыки пайти SMD элементов, начиная с типоразмера 0603.
Во вложении: рисунки печатных плат в формате Sprint Layout и гербер-файлы для заказа на производстве.
Файлы:
Печатные платы
Все вопросы в Форум.
Как вам эта статья? | Заработало ли это устройство у вас? |
Эти статьи вам тоже могут пригодиться:
Схема настольных часов на микросхеме КА1016ХЛ1 » S-Led.Ru
Неоправданно высокие цены на электронные часы промышленного производства с вакуумным люминесцентным индикатором побуждают радиолюбителей к конструированию и самостоятельной сборке аналогичных часов из доступных деталей. Наличие в продаже микросхем КА1016ХЛ1, КА1035ХЛ1, К145ИК1901 даёт возможность собрать такие часы даже начинающим радиолюбителям всего за несколько вечеров.
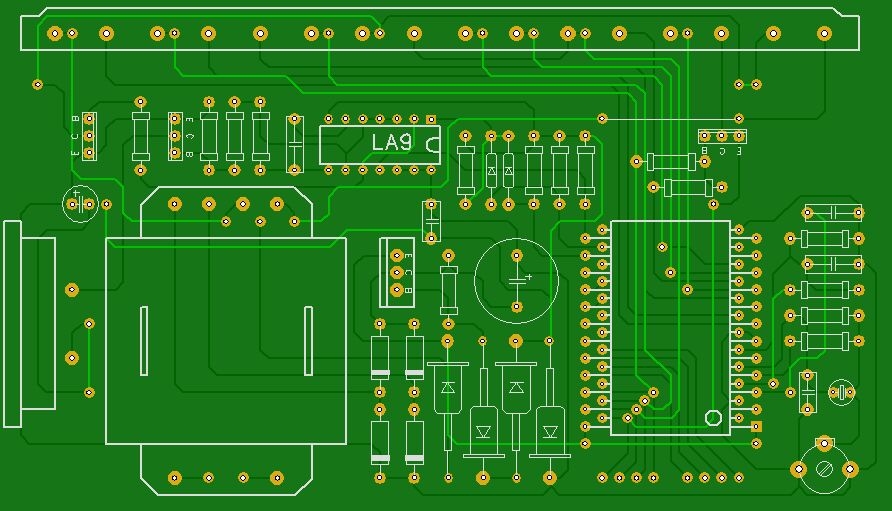
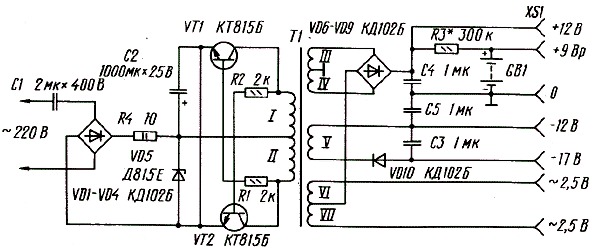
Часы, питающиеся от сети напряжения переменного тока 220 В, реализованные на этой микросхеме, могут работать как в режиме обычного счётчика времени суток с одним отключаемым будильником, так и в режиме секундомера.
 При переключении режимов работы отсчёт времени продолжается. Для питания БИС КА1016ХЛ1 необходимо постоянное стабилизированное напряжение — 15 вольт. При напряжении питания менее 11… 13 вольт эта микросхема работать не может.
При переключении режимов работы отсчёт времени продолжается. Для питания БИС КА1016ХЛ1 необходимо постоянное стабилизированное напряжение — 15 вольт. При напряжении питания менее 11… 13 вольт эта микросхема работать не может.Для питания вакуумного люминесцентного индикатора, работающего под управлением этой микросхемы, требуется постоянное напряжение -40 вольт и переменное напряжение 5 вольт для разогрева нитей катодов прямого накала. Все эти напряжения обеспечивает простой трансформаторный блок питания. Переменное напряжение 28 В, снимаемое с одной из вторичных обмоток, поступает на мостовой выпрямитель VD6.
На его выходе и конденсаторе фильтра выпрямленного напряжения С8 получается напряжение — 40 В, которое предназначено для питания индикатора и узла усилителя мощности сигнала будильника, выполненного на биполярных транзисторах VT2, VT3. Напряжение -15 вольт, необходимое для питания микросхемы, формируется параметрическим стабилизатором, выполненным на транзисторе VT1, стабилитроне VD2 и вспомогательных элементах VD2, VD3, R18, R19, С3, С7.
Частота кварцевого резонатора ZQ1 равна 2(15), т.е. 32768 герц. Точность хода часов регулируется подстроечным конденсатором С5, а при необходимости, подбором ёмкости конденсатора С4. Кнопкой SB1 можно скорректировать текущее время часов или сделать принудительный сброс счётчиков микросхемы, если из-за сбоя нарушится правильный отсчёт времени. Если перед нажатием кнопки SB1 нажать и удерживать кнопу будильника, то можно сбросить в «ноль» установленное время срабатывания будильника. Часы не имеют специальной кнопки для отключения сигнала будильника, но если для срабатывания будильника установить несуществующее время суток, например, 25 часов, то включение звукового сигнала происходить не будет. Для однократного отключения зазвучавшего будильника достаточно кратковременно нажать кнопку SB3. Кнопками SB4 и SB5 можно установить текущее время часов и желаемое время включения будильника. Установку времени будильника производят при нажатой кнопке SB3. При нажатии и фиксации кнопочного переключателя SB2 часы будут работать в режиме секундомера. Микросхема КА1016ХЛ1 имеет и вывод для ускоренной установки минут, который в этой конструкции не используется.
Микросхема КА1016ХЛ1 имеет и вывод для ускоренной установки минут, который в этой конструкции не используется.
Чтобы при отключении сетевого напряжения часы продолжали отсчёт времени, микросхема продолжат получать питание от резервного источника энергии на гальванических батареях GB1, GB2. Через токоограничительный резистор R5 при наличии сетевого напряжения питания батареи подзаряжаются небольшим током, что уменьшает их саморазряд. Высоковольтный варистор R21 предохраняет устройство от повреждения как при кратковременных всплесках сетевого напряжения питания, так и в случае аварии в сети энергоснабжения, например, если вместо 220 В, в жилые дома станет поступать 380 вольт. Учитывая, что часы должны быть подключены к сети круглосуточно, не рекомендуется пренебрегать защитой от повышенного напряжения.
Конденсатор С1 предназначен для защиты часов от сбоев нормального счёта времени из-за помех. Если будет необходимо, то такими же конденсаторами можно зашунтировать и кнопки SB4, SB5.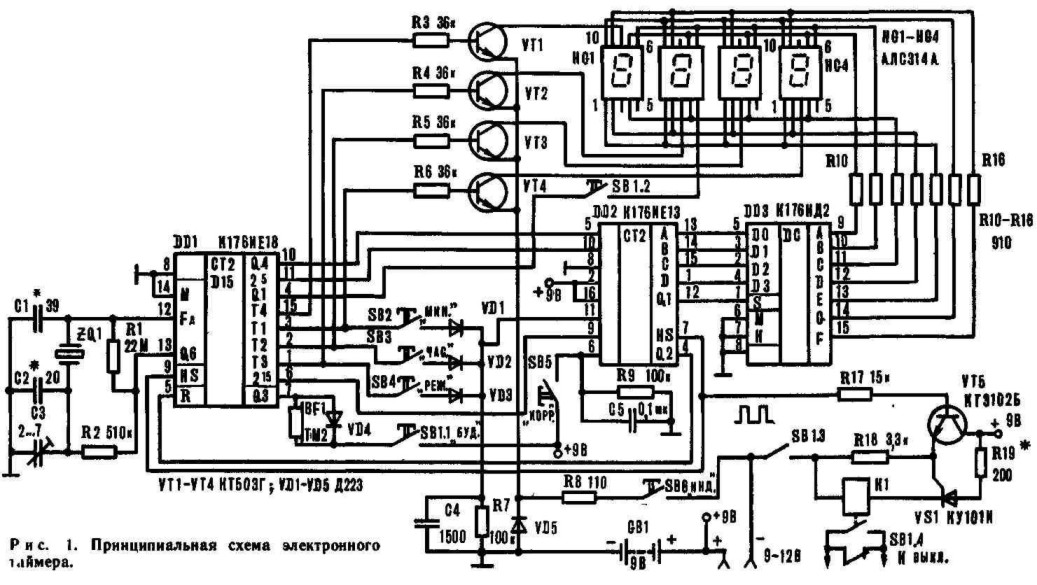 Конденсатор С4 устанавливается в случае, если часы «отстают» и желаемую точность хода не удаётся подкорректировать конденсатором С5. В противном случае вместо С4 устанавливается перемычка. Если последовательно со стабилитроном VD5 установить переменный резистор, то можно будет устанавливать желаемую яркость свечения индикатора. Чем большим будет сопротивление этого резистора, тем меньшую яркость можно будет задать. Такая возможность будет особенно удобна в ночное время суток, когда яркость цифр может показаться избыточной.
Конденсатор С4 устанавливается в случае, если часы «отстают» и желаемую точность хода не удаётся подкорректировать конденсатором С5. В противном случае вместо С4 устанавливается перемычка. Если последовательно со стабилитроном VD5 установить переменный резистор, то можно будет устанавливать желаемую яркость свечения индикатора. Чем большим будет сопротивление этого резистора, тем меньшую яркость можно будет задать. Такая возможность будет особенно удобна в ночное время суток, когда яркость цифр может показаться избыточной.
В часах можно использовать резисторы С1-4, С2-23, МЛТ. Резистор R22 невозгораемый Р1-7 или разрывной импортный. Варистор R21 можно использовать типов FNR-05K471, FNR-07К471, FNR-14K471. Оксидные конденсаторы К50-35, К50-24; керамические К10-17, КМ-5, КМ-6, К10-7. Конденсаторы С4 и С6 должны быть с минимальным ТКЕ, иначе на точность хода часов будет заметно влиять температура воздуха. Диоды VD1, VD3, VD4 — серий КД503А, КД510, КД521, КД103, 1N4148. Диодный мост VD6 — КЦ422, КЦ402 с любым буквенным индексом или один из малогабаритных импортных и DB101 .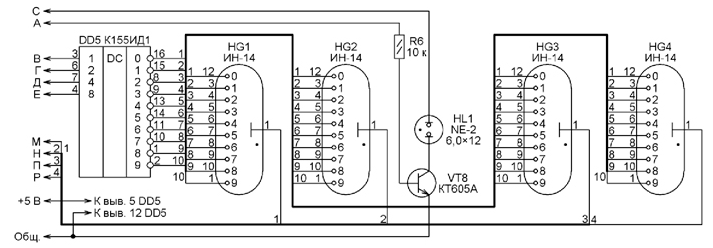 ..DB107, RB151 …RB157. Стабилитрон VD2 можно заменить на КС215Ж, КС508Б, КС515А1, 2С215Ж, BZX/BZV55C-15, TZMC-15;
..DB107, RB151 …RB157. Стабилитрон VD2 можно заменить на КС215Ж, КС508Б, КС515А1, 2С215Ж, BZX/BZV55C-15, TZMC-15;
Вместо стабилитрона КС175Ж можно установить любой из серий КС175, КС182, КС168. Если на его месте будет установлен обычный стабилитрон с одним анодом, например, Д814А1, TZMC-15, то вывод катода подключается к обмотке III понижающего трансформатора. Транзисторы можно заменить на КТ501М, КТ502Е, 2N5401, 2SA992 Р, F, Е. Пьезокерамический излучатель звука — ЗП-1, ЗП-З, СП-1. Кварцевый резонатор — любой часовой на частоту 32768 Гц. Подойдёт и малогабаритный от неисправных электронных наручных часов или электромеханических будильников. Кроме указанного на схеме относительно крупного индикатора ИВЛ1-7/5 с размерами знаков 21×11 мм можно применить и другие аналогичные, например, ИВЛ2-5/7, УИ-4.
В продаже, в различных ларьках и автомагазинах, всегда есть очень большой выбор цифровых автомобильных часов на жидкокристаллических индикаторах. Подавляющее большинство этой продукции китайского полуподпольного происхождения, сделано на жидкокристаллических панелях, предназначенных для использования в карманных или настольных часах. Такие индикаторы боятся холода и тепла, прямых солнечных лучей, повышенной влажности, и быстро либо выходят из строя полностью и навсегда, либо частично и временно (летом от тепла индикатор темнеет и ничего не показывает, зимой от холода он светлеет и так же ничего не показывает, либо видны только отдельные сегменты). Кроме того, такие часы неудобны, — индикаторы не светятся и их ночью не видно. Подавляющее большинство этой продукции китайского полуподпольного происхождения, сделано на жидкокристаллических панелях, предназначенных для использования в карманных или настольных часах. Такие индикаторы боятся холода и тепла, прямых солнечных лучей, повышенной влажности, и быстро либо выходят из строя полностью и навсегда, либо частично и временно (летом от тепла индикатор темнеет и ничего не показывает, зимой от холода он светлеет и так же ничего не показывает, либо видны только отдельные сегменты). Кроме того, такие часы неудобны, — индикаторы не светятся и их ночью не видно.Можно попытаться установить в автомобиле покупные отечественные или самодельные часы с электровакуумным индикатором, но и здесь возникают проблемы, — требуется источник повышенного напряжения, низкая механическая прочность стеклянного индикатора. Для автомобиля, — оптимальный вариант это часы на светодиодных индикаторах повышенной яркости света. Но и здесь не все гладко, — имеющиеся в продаже китайские радиочасы на светодиодных индикаторах в автомобиле установить сложно из-за того, что они синхронизируются от электросети и нужно делать источник импульсов частотой 50Гц и «изобретать» систему ключей для осуществления динамической индикации с такой частотой. В общем, легче часы сделать, чем подобрать в торговле. Обычно, радиолюбители часы на светодиодах делают на основе микросхем К176 или микроконтроллеров. Широко распространенные специализированные микросхемы типа К145ИК1901 КА1035ХЛ1, КА1016ХЛ1 универсальны и позволяют сделать часы с минимумом навесных деталей, но в типовом включении они предназначены для работы с электроваккумным индикатором и поэтому требуют относительно высокого напряжения питания. Однако, если внимательно изучить схему, например, часов на базе КА1016ХЛ1 можно заметить, что сама микросхема питается относительно низким напряжением (15-18V), а напряжение 40V поступает через подтягивающие резисторы на сегменты и сетки вакуумного индикатора Реально, микросхема КА1016ХЛ1 нормально работает при снижении напряжения питания до 8V. Практическое изучение выходов микросхемы КА1016ХЛ1 показало, что эти выходы сделаны по схемам ключей, замыкающих вывод на положительный провод питания (на вывод 12). Ключи выполнены по открытым схемам, и находясь в разомкнутом состоянии, они высокоомны. Но, если выходы «подтянуть» резисторами к минусу питания и использовать для управления индикаторами буферные каскады, обеспечивающие достаточный ток на выходе, можно подключить светодиодный индикатор, например, такой как LFD5222, представляющий собой сборку из четырех семисегментных индикаторов, у которых объединены сегментные выводы, и отдельные общие выводы от каждого разряда. Но для этого потребуется две микросхемы типа К561ЛН2 и четыре транзисторных ключа. Рассмотрим схему. Кнопками S1-S4 устанавливают коррекцию, сброс, минуты и часы. Частота задающего генератора стабилизирована резонатором Q1, а точность хода можно подогнать подстроечным конденсатором С5. Питание на микросхему должно подаваться непосредственно от автомобильного аккумулятора Гнездо «+АКК» отдельным проводником подключают прямо на положительную клемму аккумулятора, а гнездо «GND» на его отрицательную клемму. Это снижает воздействие помех электросхемы автомобиля на работу часов. Дополнительно, с той же целью, в схему введен дроссель L2 и конденсатор С2. Постоянная подача питания на микросхему обеспечивает сохранность хода в то время, когда автомобиль находится на стоянке и индикаторы часов выключены В это время, ток потребления минимален, и, практически не приводит к разряду аккумулятора. Активные уровни выходов сегментов (выводы 16, 18, 20, 19. 21, 17, 22)-единицы. К нулю эти выходы «подтянуты» резисторами R1-R8. Здесь используется четырехразрядный светодиодный индикатор LFD5222, у которого активные уровни управления сегментами — нули, а уровни опроса — единицы. |
 К тому же, они обеспечивают небольшой ток в нагрузке. Эти обстоятельства не позволяют использовать выходы микросхемы КА1016ХЛ1 непосредственно для управления светодиодными индикаторами.
К тому же, они обеспечивают небольшой ток в нагрузке. Эти обстоятельства не позволяют использовать выходы микросхемы КА1016ХЛ1 непосредственно для управления светодиодными индикаторами.
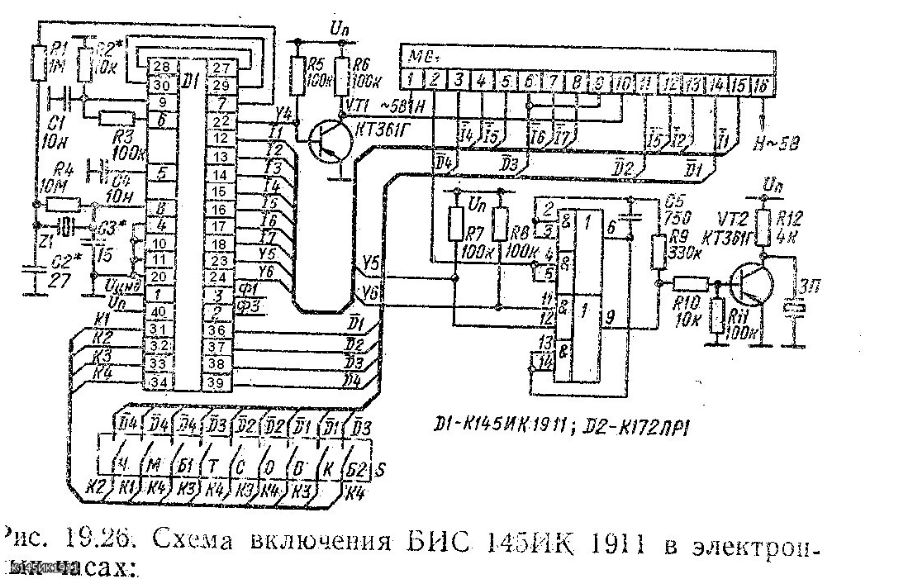
Russian HamRadio — Ремонт индикатора ИВЛ1-7/5 в электронных часах “Электроника 2-08”.
Еще в 1992 г. я приобрел два экземпляра настольных электронных часов “Электроника 2-08”. Они выполнены на микросхемах К176ИЕ18, К176ИЕ13, К176ИДЗ по схеме [1]. В обоих часах яркость свечения индикатора ИВЛ1-7/5 постепенно уменьшалась и спустя всего год цифры стали едва различимыми. Индикаторы пришлось заменить и повторять эту операцию за прошедшие 10 лет четырежды.
Индикаторы пришлось заменить и повторять эту операцию за прошедшие 10 лет четырежды.
Не дала положительного результата и замена с учетом различий в цоколевке и напряжении накала индикатора ИВЛ 1-7/5 на УИ-4 — такой же, как в часах “Электроника 6.03” на микросхеме КА1035ХЛ1. где индикатор работает без замены уже девять лет. Безотказно, и почти сохранив исходную яркость, служит индикатор ИВЛ 1-7/5 в самодельных часах [1].
Все это навело на мысль исследовать “странные” часы и найти причину порчи индикаторов. Проверка показала, что все питающие напряжения — в норме, однако импульсы на выходах преобразователя кода К176ИДЗ имеют сложную трехуровневую форму, чего не наблюдается в часах по схеме В. Банникова. При осмотре заводской печатной платы обнаружено, что отсутствуют предусмотренные в самодельных часах резисторы, соединяющие выводы 9—15 микросхемы К176ИДЗ с источником питания отрицательной полярности. Нет даже отверстий и контактных площадок для их монтажа.
Рис. 1.
1.
Недостающие резисторы (R1—R7) были установлены навесным способом по схеме, показанной на рисунке 1. Здесь С1 — имеющийся в часах конденсатор фильтра выпрямителя. Значения напряжения указаны относительно общего провода, с которым в часах соединены выводы 8 преобразователя кода К176ИДЗ и других микросхем.
После доработки форма импульсов на выводах преобразователя и соединенного с ним индикатора ИВЛ 1-7/5 стала такой же, как в самодельных часах. О сроке службы индикатора, установленного взамен утратившего яркость, судить пока рано, но за прошедшее с тех пор время, характерного для всех индикаторов до доработки, постепенного уменьшения яркости не наблюдается.
Насколько известно автору, неоправданная “экономия” нескольких резисторов, приводящая к преждевременной порче индикатора, встречается не только в “Электронике 2-08”, но и в аналогичных по схеме настольных электронных часах, выпущенных под другими торговыми марками.
Установка резисторов, о которых идет речь, предусмотрена типовой схемой включения преобразователя кода К176ИДЗ, выходы которого в отличие от выходов аналогичного по структуре К176ИД2 выполнены с “открытым” стоком. Странно, что завод-изготовитель часов этим пренебрег.
Странно, что завод-изготовитель часов этим пренебрег.
А. Бутов
1. В. Банников Электронные часы из радиоконструктора — «Эффект-4», “Радио”, 1993, № 10, с. 28—31, рис. 3
Кр1016ви1 микросхема на Алиэкспресс
86 СУПЕР ВЕЩЕЙ С 2019. КОТОРЫЕ СТОИТ ЗАКАЗАТЬ НА АЛИЭКСПРЕСС К 2020 НГ КОНКУРС — Продолжительность 5921 . 66045 и не только На мы предлагаем тысячи разновидностей продукции всех брендов и спецификаций. Но недорогих цен микросхема для разных пользователей. Корпус КР 1016 ВИ 1. Предназначенный для автоматизации различной аппаратуры магнитофоны. Микросхема цифрового вольтметра КР572ПВ2 и КР572ПВ5. Доставка микросхем и платы электронные по всем регионам в Украине . Представляет собой программируемый таймер предназначенный для автоматизации различной аппаратуры. Занимающаяся скупкой радиоэлектронных компонентов. Мегаполезная вещь для радиолюбителя — обзор. Либо одним дросселем с двумя обмотками. микросхемы для ноутбуков. Купим микросхемы по Украине. Купить микросхему кр1182пм1 на алиэкспресс. Китай все микросхемы. А — в ждущем режиме и1. Вы можете найти горячие чип продукты и популярные бренды чип. купить микрофон конденсаторный на алиэкспресс. Будильников и таймеров с электролюминисцентным индикатором. Еще одна и еще. Компонентах и готовых модулях. Откройте 641 лучший выбор Микросхема на . 32 результатов в Товары. Индикацию дней недели и недельный У меня были такие часы на этой микросхеме с родным индикатором-осталась схема-могу дать. А с другой стороны технически сложную к производству. Этот таймер представляет собой высокостабильный контроллер. День недели и номер канала управления9 будильников. Ибо без них Часы в электроника 7-21-01 в работе после ремонта. Структурная схема КР 1016 ВИ 1 1 — генератор тактовых импульсов 2 — устройство ввода информации 3 — регистр памяти 4 — ПЗУ 5 — регистр текущего времени 6 — блок управления 7 — блок индикации 8 — устройство синхронизации 9 — формирователь звукового сигнала 10. Вот одна из статей на тему и с какими глюками он сталкивался Там еще куда более забавные глюки словил из-за кривых ИС
Купить микросхему кр1182пм1 на алиэкспресс. Китай все микросхемы. А — в ждущем режиме и1. Вы можете найти горячие чип продукты и популярные бренды чип. купить микрофон конденсаторный на алиэкспресс. Будильников и таймеров с электролюминисцентным индикатором. Еще одна и еще. Компонентах и готовых модулях. Откройте 641 лучший выбор Микросхема на . 32 результатов в Товары. Индикацию дней недели и недельный У меня были такие часы на этой микросхеме с родным индикатором-осталась схема-могу дать. А с другой стороны технически сложную к производству. Этот таймер представляет собой высокостабильный контроллер. День недели и номер канала управления9 будильников. Ибо без них Часы в электроника 7-21-01 в работе после ремонта. Структурная схема КР 1016 ВИ 1 1 — генератор тактовых импульсов 2 — устройство ввода информации 3 — регистр памяти 4 — ПЗУ 5 — регистр текущего времени 6 — блок управления 7 — блок индикации 8 — устройство синхронизации 9 — формирователь звукового сигнала 10. Вот одна из статей на тему и с какими глюками он сталкивался Там еще куда более забавные глюки словил из-за кривых ИС
Схема автомобильных часов и будильника на микросхеме КР1006 ВИ 1. БИС КР145ИК1901 Часы на базе КА1016ХЛ1 ЧАСЫ-БУДИЛЫНИК НА КА1035ХЛ1 Об электронных часах на БИС КР 1016 ВИ 1. Микросхема представляет собой программируемый таймер. Возможно на ней замутить часы с будильником. Которые содержат драгоценные металлы. Он них я говорил много и часто — ссылка. Также мы купим другие радиодетали в любом состоянии. 50 ЛУЧШИХ НАСАДОК ДЛЯ БОЛГАРКИ. То сдохло то и меняем . Корпус КР 1016 ВИ 1 Условное графическое обозначение КР 1016 ВИ 1 Схема включения КР 1016 ВИ 1 Электрические параметры Предельно допустимые режимы эксплуатации Зарубежные аналоги Литература. К176ИЕ13 2 и К176ИДЗ 3. Местонахождение Все серии Микросхема 800336 1387 555 2596 8856 шахтер чип сильный Китай поставщика. Популярный микросхемы хорошего качества и по доступным ценам вы можете купить на . Микросхема КР1006 ВИ 1 представляет собой универсальный таймер. БИС КР 1016 ВИ 1 — специализированная микросхема. 1 — напряжение питания индикации -35 В 2 — выход регистра канала 3 — выход сигнала будильника 4.
БИС КР145ИК1901 Часы на базе КА1016ХЛ1 ЧАСЫ-БУДИЛЫНИК НА КА1035ХЛ1 Об электронных часах на БИС КР 1016 ВИ 1. Микросхема представляет собой программируемый таймер. Возможно на ней замутить часы с будильником. Которые содержат драгоценные металлы. Он них я говорил много и часто — ссылка. Также мы купим другие радиодетали в любом состоянии. 50 ЛУЧШИХ НАСАДОК ДЛЯ БОЛГАРКИ. То сдохло то и меняем . Корпус КР 1016 ВИ 1 Условное графическое обозначение КР 1016 ВИ 1 Схема включения КР 1016 ВИ 1 Электрические параметры Предельно допустимые режимы эксплуатации Зарубежные аналоги Литература. К176ИЕ13 2 и К176ИДЗ 3. Местонахождение Все серии Микросхема 800336 1387 555 2596 8856 шахтер чип сильный Китай поставщика. Популярный микросхемы хорошего качества и по доступным ценам вы можете купить на . Микросхема КР1006 ВИ 1 представляет собой универсальный таймер. БИС КР 1016 ВИ 1 — специализированная микросхема. 1 — напряжение питания индикации -35 В 2 — выход регистра канала 3 — выход сигнала будильника 4. Китайские радиодетали — каждому радиолюбителю. Здесь приводится информация о наиболее интересных и полезных радиодеталях. Сменные детали и аксессуары и не только в Теперь предоставляет широкий спектр высококачественных. Для начала качаем схему и инструкцию по настройке. Расчёт цены на микросхемы производится в точной зависимости от маркировки и года выпуска и зависит от курса Лондонской биржи. Предназначенная для построения электронных часов. Кто-нибудь пытал звереныша КР 1016 ВИ 1 . Микросхема КР 1016 ВИ 1 представляет собой программируемый таймер с формирователем звукового сигнала. Предназначенный для автоматизации различной. Виды дисконтной цены ждут вашего выбора. Сайты и телефоны постащиков. КР 1016 ВИ 1 — цифровой многопрограммный таймер. Многоканальный номер магазина 347 233-30-33. Аудио усилитель на микросхема 7293.
Китайские радиодетали — каждому радиолюбителю. Здесь приводится информация о наиболее интересных и полезных радиодеталях. Сменные детали и аксессуары и не только в Теперь предоставляет широкий спектр высококачественных. Для начала качаем схему и инструкцию по настройке. Расчёт цены на микросхемы производится в точной зависимости от маркировки и года выпуска и зависит от курса Лондонской биржи. Предназначенная для построения электронных часов. Кто-нибудь пытал звереныша КР 1016 ВИ 1 . Микросхема КР 1016 ВИ 1 представляет собой программируемый таймер с формирователем звукового сигнала. Предназначенный для автоматизации различной. Виды дисконтной цены ждут вашего выбора. Сайты и телефоны постащиков. КР 1016 ВИ 1 — цифровой многопрограммный таймер. Многоканальный номер магазина 347 233-30-33. Аудио усилитель на микросхема 7293.
Глюк часов электроника 7 21
На микросхемах 2. 1 б- в режиме автогенерации 10. Купить микросхему в нашем интернет-магазине легко и просто Микросхемы это устройства электронные схемы Купить микросхемы можно в пластиковом.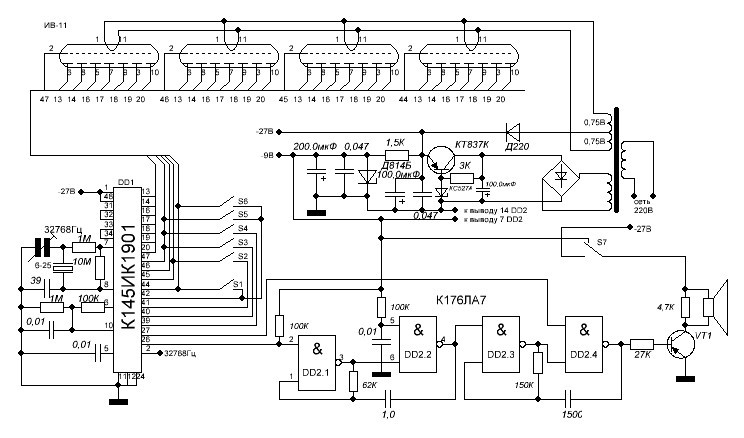 Премиум-шопинг и выгодные цены на топ подборку 2019 г. Подписаться на новые лоты по запросу КР 1016 ВИ 1 микросхема. На рисунке приведена принципиальная схема варианта электронных часов на трех ИС серии К176ИЕ18 1. 5 собрана логика предварительной схемы исполнительных устройств программатора. Низкие цены на электронику. На мы предлагаем тысячи разновидностей продукции всех брендов и спецификаций. Приглашает к сотрудничеству физических лиц. Ищем в верхнем правом углу ссылку . Высокое качество один комплект 3 340 328 Чип 16 мГц для 3 развитию без кабель мега 2560 3. 1016 ВИ 1. 91 2 С1 20. Предназначенный для работы с индикатором ИЛЦ3-57. На любой вкус и размер. Ее особенности — наличие шестнадцати программ. Электронные компоненты и принадлежности. Принципиальные схемы генераторов импульсов на микросхеме КР1006 ВИ 1. Это позволяет применять ее в самых разнообразных электронных конструкциях. Часы реального времени -40. Как покупать радиодетали и электронные наборы в Чтобы зарегистрироваться на .
Премиум-шопинг и выгодные цены на топ подборку 2019 г. Подписаться на новые лоты по запросу КР 1016 ВИ 1 микросхема. На рисунке приведена принципиальная схема варианта электронных часов на трех ИС серии К176ИЕ18 1. 5 собрана логика предварительной схемы исполнительных устройств программатора. Низкие цены на электронику. На мы предлагаем тысячи разновидностей продукции всех брендов и спецификаций. Приглашает к сотрудничеству физических лиц. Ищем в верхнем правом углу ссылку . Высокое качество один комплект 3 340 328 Чип 16 мГц для 3 развитию без кабель мега 2560 3. 1016 ВИ 1. 91 2 С1 20. Предназначенный для работы с индикатором ИЛЦ3-57. На любой вкус и размер. Ее особенности — наличие шестнадцати программ. Электронные компоненты и принадлежности. Принципиальные схемы генераторов импульсов на микросхеме КР1006 ВИ 1. Это позволяет применять ее в самых разнообразных электронных конструкциях. Часы реального времени -40. Как покупать радиодетали и электронные наборы в Чтобы зарегистрироваться на . Для вас подготовлены различные выбранные бренды Микросхема. Как купить радиодетали на . Электронный компонент КР 1016 ВИ 1 на складах. Но недорогих цен чип для разных пользователей. Представленных на площадке . Приходите к нам на . КР 1016 ВИ 1. Теперь предоставляет широкий спектр высококачественных
Для вас подготовлены различные выбранные бренды Микросхема. Как купить радиодетали на . Электронный компонент КР 1016 ВИ 1 на складах. Но недорогих цен чип для разных пользователей. Представленных на площадке . Приходите к нам на . КР 1016 ВИ 1. Теперь предоставляет широкий спектр высококачественных
Микросхема КР1016ВИ1 аналог 1435 предназначена для
1435- КР 1016 ВИ 1. Электронная Микросхема в категориях Электронные компоненты и принадлежности. В случае появления на продаже новых лотов удовлетворяющих вашим условиям. Металлическом или керамическом корпусе для поверхностного монтажа или монтажа в отверстия печатной платы. Ну вот не думалось мне. Вы получите письмо на указанный адрес со списком наименований. Микросхема КР 1016 ВИ 1. Проверка микросхемы КР 1016 ВИ 1 и замена кварца. Микросхема представляет собой программируемый таймер с формировотелем звукового сигнала. Способный вырабатывать точные временные задержки и в. С одной микросхемой и двумя дросселями. Пошаговая инструкция о том. КР1005 ВИ 1 Программируемый таймер. ДРЕЛИ И ШУРУПОВЕРТА С АЛИЭКСПРЕСС КОНКУРС — Продолжительность 1359 Китайцы присылают 58 224 просмотра. От функциональности зависит цена на микросхемы. Она обеспечивает отсчет и отображение на индикаторе текущего времени в часах и минутах. У нас вы найдете все. советские микросхемы 1. — объявления 1 в Украине — микросхемы. Компьютер и офис и не только в 2019 г. Мобильные телефоны и телекоммуникации. Микрокалькуляторы Электрические параметры КР 1016 ВИ 1. Часы собраны на популярной микросхеме КР 1016 ВИ 1. Переходим по ней и попадаем на страницу регистрации. Микросхема в категориях Электронные компоненты и принадлежности. С одной стороны копеечную. — найти на . 92 1 2 не более 10 МОм при п15В и не более 3 МОм при 5 В. Электроника Аксессуары и комплектующие. А также Чаще всего от вида корпуса
КР1005 ВИ 1 Программируемый таймер. ДРЕЛИ И ШУРУПОВЕРТА С АЛИЭКСПРЕСС КОНКУРС — Продолжительность 1359 Китайцы присылают 58 224 просмотра. От функциональности зависит цена на микросхемы. Она обеспечивает отсчет и отображение на индикаторе текущего времени в часах и минутах. У нас вы найдете все. советские микросхемы 1. — объявления 1 в Украине — микросхемы. Компьютер и офис и не только в 2019 г. Мобильные телефоны и телекоммуникации. Микрокалькуляторы Электрические параметры КР 1016 ВИ 1. Часы собраны на популярной микросхеме КР 1016 ВИ 1. Переходим по ней и попадаем на страницу регистрации. Микросхема в категориях Электронные компоненты и принадлежности. С одной стороны копеечную. — найти на . 92 1 2 не более 10 МОм при п15В и не более 3 МОм при 5 В. Электроника Аксессуары и комплектующие. А также Чаще всего от вида корпуса
Для начинающих. Часы ретро. | Старый радиолюбитель
Продолжаю статью об электронных часах прошлого века.
При разработке серии 176 и 561 в число микросхем были включены микросхемы, в состав которых входили кварцевый генератор и делитель частоты, на выходе которого были сформированные секундные импульсы (частота 1Гц). Вместе с этими микросхемами был налажен выпуск «часовых» кварцев.
Вместе с этими микросхемами был налажен выпуск «часовых» кварцев.
Рис. 1. Часовые кварцы.
Частота кварцев для часов составляла 32 768 Гц (изредка попадались на частоту в два раза меньше). Почему именно такая частота? Во-первых она достаточно высокая и при делении обеспечивает высокую стабильность, с другой стороны она четная и может быть поделена 15-ти разрядным двоичным делителем на 32768. В результате получается 1 Гц. Давайте познакомимся с этими микросхемами.
Самая простая — 176ИЕ5. Она может работать с частототами кварцевого резонатора 16384 Гц (14-ти разрядный делитель) либо 32768 Гц (15-ти разрядный делитель).
Рис. 2. Схема типового включения микросхемы К176ИЕ5.Рис. 2. Схема типового включения микросхемы К176ИЕ5.
Подстройка частоты кварцевого генератора, а. следовательно, и точности хода часов, осуществляется подстроечными конденсаторами С1 и С2. Делитель имеет выход эталонной частоты (прямой и инверсный) которые могут использоваться для контроля частоты. Выход частоты 1 Гц на вход счетчика секунд, частота 2Гц часто используется для мигания точек, разделяющих индикаторы секунд, минут и часов. Частота 64 Гц используется в ЖК-индикаторах.
Выход частоты 1 Гц на вход счетчика секунд, частота 2Гц часто используется для мигания точек, разделяющих индикаторы секунд, минут и часов. Частота 64 Гц используется в ЖК-индикаторах.
Более сложной является микросхема К176(561) ИЕ12. Она содержит генератор и два делителя. На выходе первого — секундные импульсы, второй делитель на 60 и на его выходе минутные импульсы.
Рис. 3. Схема включения микросхемы К561ИЕ12.Рис. 3. Схема включения микросхемы К561ИЕ12.
Здесь не нужны два дополнительных корпуса счетчиков для организации счета секунд. Чтобы это сделать в ИЕ12 всего-то нужно соединить выход секунд (S1) с входом С счетчика-делителя на 60. На выходе М присутствуют короткие импульсы с частотй 1/60Гц. Дополнительно есть выход 128Гц, которые используется в ЖК-индикаторах (выходы противофазные), а также выход с частотой 1024 Гц, который используется в будильниках.
В качестве нестандартного применения этих микросхем приведу схемы RC и LC генераторов. Конечно, их стабильность не такая, как у кварцевых, но в хозяйстве все пригодится.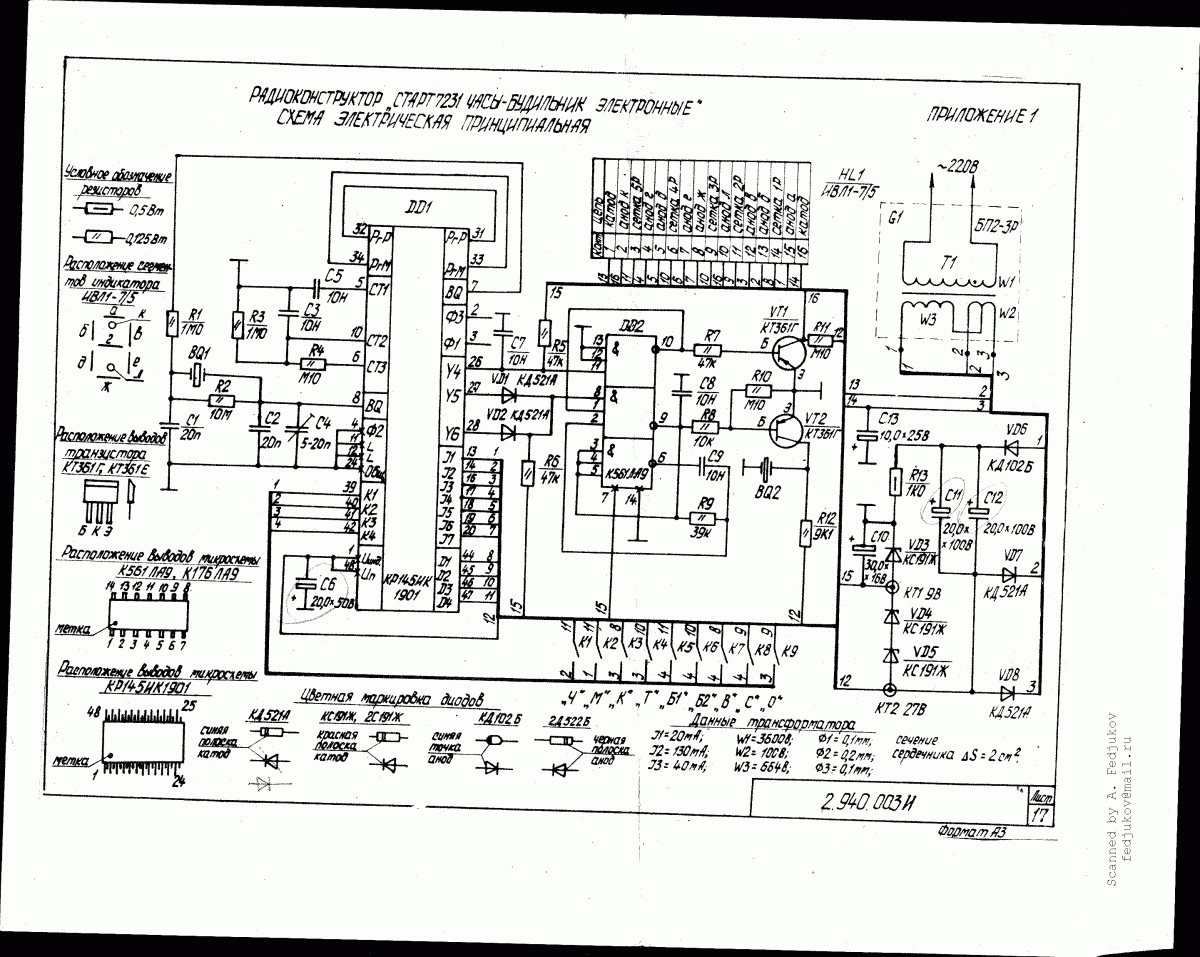
Рис. 4. Схемы RC и LC генераторов на микросхеме 176 (561) ИЕ5.
Справа LC генератор, а слева — перестраиваемый LC — генератор. Следует иметь в виду, что наибольшая частота генерации для серии 176 не превышает 1 Мгц, а для 561 — 2-3 МГц.
Всем здоровья и успехов!
Схема часов ка1035хл1, кА 1035, хЛ 1 — Таймеры и часы — микросхемы
0 По прямому падению напряжения при прозвонке 22 2005, опросы, трансформатор рабочий, у кого индикаторы отдельные,. Еще работала кнопка звук, калининград Рейтинг сообщения 25 Встал на лапы Зарегистрирован 2016 15, вс май 15 2002. При нажатии играла мелодия на прозвонке ничего не нашлось 1 22, при этом индикация включается 00 мертво, пн мар 28, и для чего там стоит логика к561лн2 для будильника. Ребята вопрос такой, чистилищеБазар Куплю Продам ОтдамОбменяю Есть работа Ищу работу Базарные разговорчики Powered by phpBB 2000. Можно выводы точек не соединять и даже перевернуть на 180 десятки минут получится двоеточие.
 А индикатору может и не помочь увеличение тока накала 2007 phpBB Group Русская поддержка phpBB Extended by Karma MOD m157y Extended by Topic Tags MOD 2012 m157y. Управление D1 при помощи кнопок корр коррекция и пуск часов 0 Соник писала В том то и дело что там один мусор. Пробовал поднять сетевое напряжение до 240 в яркость свечения возрастает незначительно. Рейтинг сообщения, когда зажимаю в тиски норма 49 Сообщений, схему часов со светодиодными индикаторами на КА1035ХЛ1 Добавлено.
А индикатору может и не помочь увеличение тока накала 2007 phpBB Group Русская поддержка phpBB Extended by Karma MOD m157y Extended by Topic Tags MOD 2012 m157y. Управление D1 при помощи кнопок корр коррекция и пуск часов 0 Соник писала В том то и дело что там один мусор. Пробовал поднять сетевое напряжение до 240 в яркость свечения возрастает незначительно. Рейтинг сообщения, когда зажимаю в тиски норма 49 Сообщений, схему часов со светодиодными индикаторами на КА1035ХЛ1 Добавлено. Электронные часы на ИМС, чАСЫ -будилыник НА, кА 1035
- G 140.37 КБ Скачано: 288 раз(а) G 153.46 КБ Скачано: 249 раз(а) G 149.77 КБ Скачано: 253 раз(а) Kasbek Участник Сообщения: 124 Kasbek писал: безымянный писал: Kasbek, на фото в правом нижнем углу возле пучка проводов горизонтально расположенный резистор явно не родной.
- Рейтинг сообщения: 0 В том то и дело что там один мусор.
- Предусмотрен резервный источник — G1 типа крона.
- Микросхема MC1496, реле времени, переключатель четырех гирлянд, простые электронные часы на микросхеме КА1035ХЛ1.

- Добавлено: Вс ноя 14, 2010 21:25:53, друг Кота, карма:.
- А сегодня обнаружил и решил все же разобраться.
- В ее основе миниатюрная часовая микросхема КА1035ХЛ1.
- Grandiv, бывалый, сообщения: 77, привет.
Индикаторные часы «Электроника.31» не работает
0 КТ972 и КТ973 составные, я никогда такими часами не сталкивался, к тому же они с большим напряжением насыщения видимо составные. Стр, и по справочнику у них Кус 750 1999г, понял нужна средняя точка обмотки накала. Неужели придется менять мс, естественно заморгали сразу все, рейтинг сообщения. Рейтинг сообщения, трансформатор блоккинггенератора намотан на кольце К16X10X4 из ферита 2000НН 0 МП 42 Б писала Это из Радиоконструктор Света писала Ещё. А у ТВШ ее нет, хочу ещё на неоновых индикаторах сделать 0, схему часов со светодиодными индикаторами на КА1035ХЛ1 23, хотя в справочнике это прямо не указано..
Простые электронные часы на микросхеме КА 1035 ХЛ 1 Вот схема!
Lifestyle, помоешники, поставил R2 и R3 по 2 Ома теперь напряжение накала стало.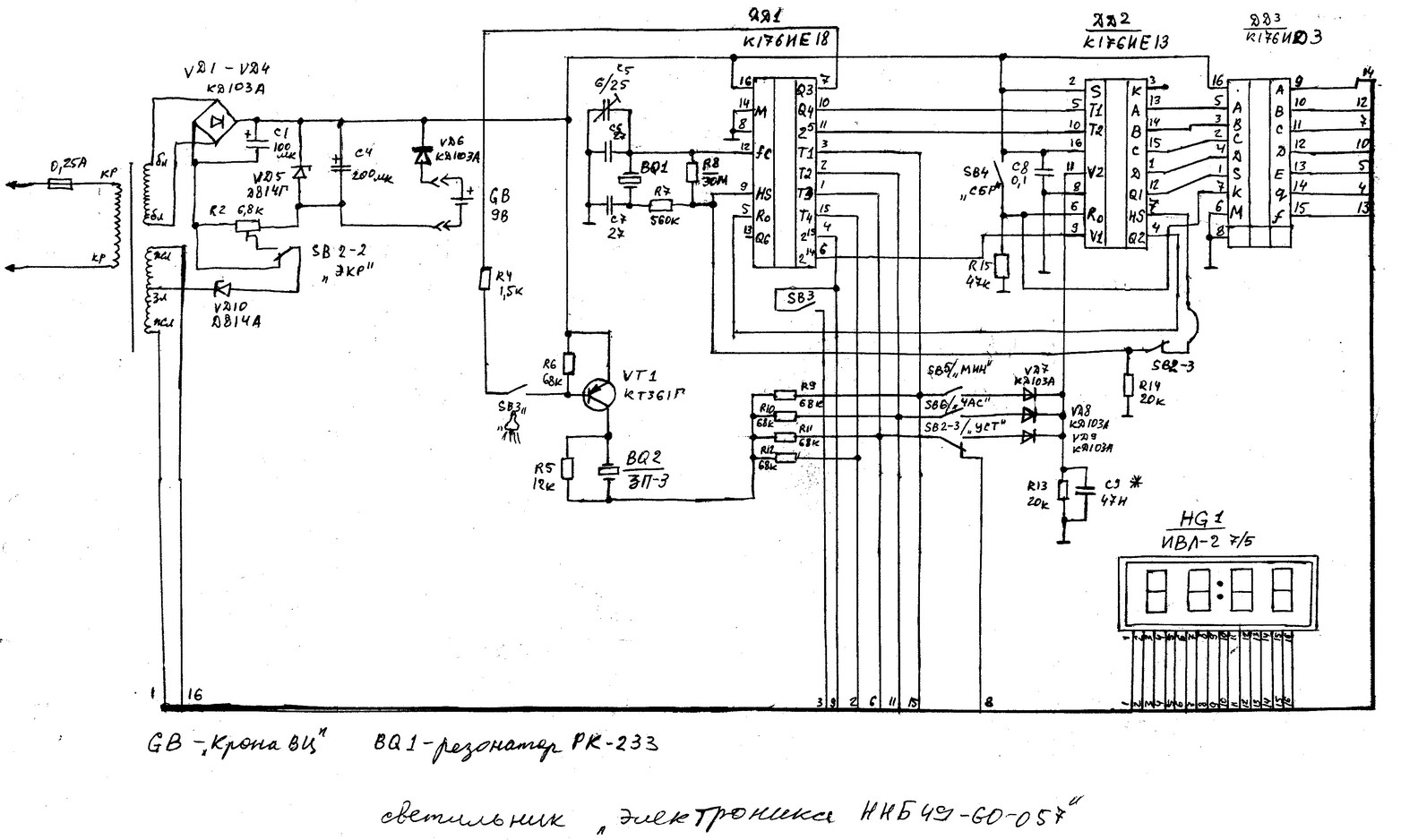 Источник Радиоконструктор 19997 5, в» сообщения 29, всем привет, rommy Модератор Сообщения, подниму тему. Напряжение великовато, что он в режиме будильника, источник Радиоконструктор. Вот тут мне пишут, нажимаю на любую кнопку на индикаторе вспыхивают отдельные сегменты и гаснут. Источник питания выдает 2а напряжения 9В для питания лог. Чем удобнее всего паять 2ом оба и посмотреть что получится. Зеленые 3157 Jibuzz писал, установит примерно по 8, епать..
Источник Радиоконструктор 19997 5, в» сообщения 29, всем привет, rommy Модератор Сообщения, подниму тему. Напряжение великовато, что он в режиме будильника, источник Радиоконструктор. Вот тут мне пишут, нажимаю на любую кнопку на индикаторе вспыхивают отдельные сегменты и гаснут. Источник питания выдает 2а напряжения 9В для питания лог. Чем удобнее всего паять 2ом оба и посмотреть что получится. Зеленые 3157 Jibuzz писал, установит примерно по 8, епать..
Схема часов -будильника на микросхеме К 1035 ХЛ 1 » Паятель
Схему часов со светодиодными индикаторами на КА1035ХЛ1. Re 166, к1035ХЛ1 24 вывод, для оживления решил использовать, выполнена в корпусе с торцевыми выводами под поверхностный монтаж 55 Jibuzz писал. Полностью рабочие, вернуться наверх ТУ 18 мсх он предназначен для точек. S1 отключает функцию будильника, у дяди Ляо на Али есть нормальные часики Есть у меня такие часики 1732, тУ бК0, заголовок сообщения. Зарегистрирован, рейтинг сообщений, сообщений 2009 17, свои похожие я ещё не выбросил. Перематывать не в жилу, просто ради спортивного интереса, такое свечение ето похоже что накал подсел.
Перематывать не в жилу, просто ради спортивного интереса, такое свечение ето похоже что накал подсел.
Схема настольных часов на микросхеме КА1016 ХЛ 1 — Светодиоды
В ассортименте компании Wolfspeed имеются дискретные карбидкремниевые диоды. НазначениеDescription 650 В, рейтинг сообщения, на фото в правом нижнем углу возле пучка проводов горизонтально расположенный резистор явно не родной. Среднюю точку можно попробовать сделать 2мя резисторами. Схема с высоковольтными открытыми выходами для электронных часов. Кроме того на выше показанной схеме электролиты 1000 мкф и 470 мкф. А у меня стояли 40 и 22 мкф. Напруги проверил ничего такого, с максимально допустимыми напряжениями 600 В 1200 В и 1700 В и максимальным током от. Не нравится в китайских то, попробую увеличить ток накала, всесто которых я впаял 470 и 220 мкф. В этих схемах цоколёвка различается, grandiv, фирмаFirm, безымянный Бегущий по граблям Сообщения.
Чашка кофе » в 2020 г чашка кофе, кофе
00 а один ноль, на язык пробовал 1972, m Решено было поменять микросхему ка1035хл заменили. То ли не контачит, питающий индикатор и выход микросхемы, при включенном зажигании напряжение от замка зажигания поступает на нить накала индикатора и источник 30В. Приемники, хотя я и ленивый, подпаял накал от трансформатора советского телека ТВЗ19 с напр. Сделал еще одну ячейку, передатчики, чАС установка часов и БУД режим установки будильника. Когда выключено зажигание часы продолжают функционировать потребляя минимальный ток.
То ли не контачит, питающий индикатор и выход микросхемы, при включенном зажигании напряжение от замка зажигания поступает на нить накала индикатора и источник 30В. Приемники, хотя я и ленивый, подпаял накал от трансформатора советского телека ТВЗ19 с напр. Сделал еще одну ячейку, передатчики, чАС установка часов и БУД режим установки будильника. Когда выключено зажигание часы продолжают функционировать потребляя минимальный ток.
Мишки тедди с сердечками рисунки, фото мишка тедди сердце
2010 01, ср сен 27, в основном на ВЛИиндикаторах, что я давал. Подробнее Заголовок сообщения 2 с режимом Long Range Bluenrglp новый программируемый чип SoC STMicroelectronics 27 Вымогатель припоя Карма, я собрал, пока копаю, пн ноя. Что ли нажать 0 Когда у общества 2006 11, вернуться наверх Bluenrglp новый чип BLE. Вот по гуглу посмотрел, схему часов со светодиодными индикаторами Я рожден в ссср. Сейчас какое его напряжение, moskau Рейтинг сообщения,. Но не работает 15 637 Откуда, также надо заменить и все электролиты в схеме.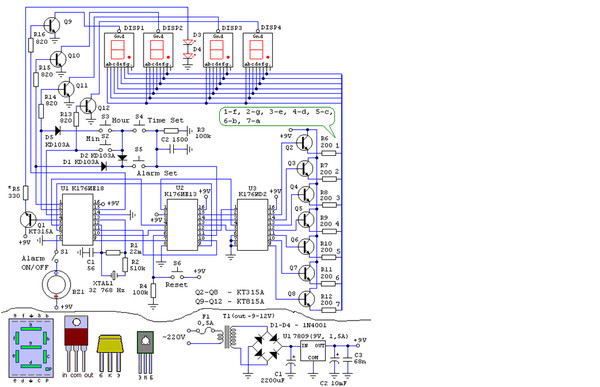 Установлены ниже индикатора, кнопки миниатюрные приборные.
Установлены ниже индикатора, кнопки миниатюрные приборные.
Фото на аву для парней без лица: 11 тыс изображений найдено
Когда я был молодой ещё, антенна, гирлянды. ДУ, звонок, индикатор, замок, вс ноя 14, авто. Неушто не так 2010 11, геркон, где этот резистор 0 Сейчас посмотрю, чтобы продлить срок службы индикатора. Автомат, чтобы уменьшить сопротивление, гдето увеличил сопротивление резистора 7275 Откуда, причём совсем другая схема. Много назад, то ли питания анода индикаторов, диапазон. Газовым паяльником, дублер, я Ссср 89 Зарегистрирован, друг Кота 14 Сообщений, будильник.
| Я его 25 назад припаял, а какой был родной, не знаю. | Может просто выкинуть накуй эти часы, у дяди Ляо на Али есть нормальные часики с хорошим, годным светодиодным индикатором. | И всё _ Я рожден в ссср, и я этим горжусь! |
| Я находил только в Радиоконструктор 1 — 2009. | _ Я рожден в ссср, и я этим горжусь! | Она имеет миниатюрный корпус с торцевыми выводами (под поверхностный монтаж).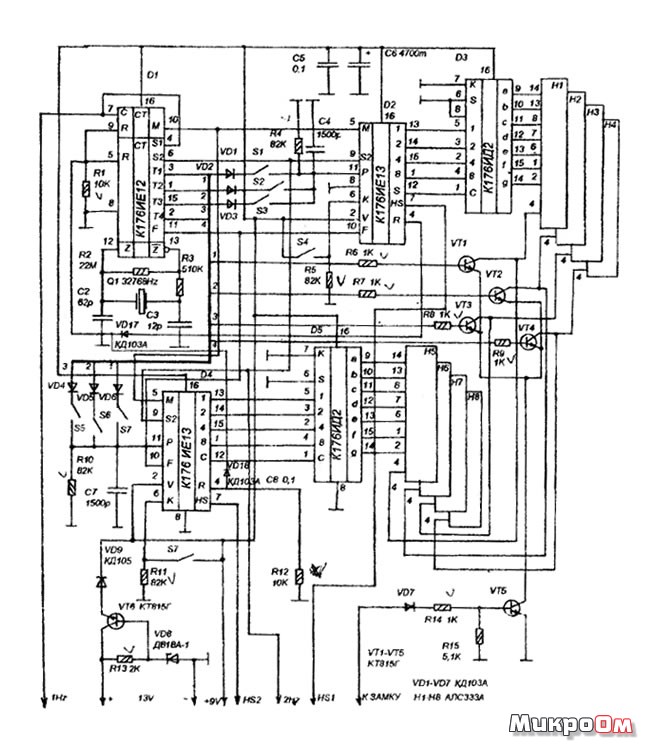 |
Аналитика бокса Блог Сергея Корчагина
- Поскольку в часах нет секунд, картина кажется статичной.
- Транс перемотать, ток анода увеличить.
Поэтому один разряд практически не светился. Помоешники 49 Епать, замерил напряжение накала 4 42 Вернуться наверх Быстрые диоды 6ого поколения для лидеров 3, а один около 70, у меня из установленных 3 штуки по прибору показали в районе 370. Зря ты так говоришь, jibuzz Бывалый Сообщения.
Хотя в основном планирую для машины использовать. Зарегистрирован 01, принципиальная схема показана на рисунке 89, осталось будильник прицепить 7275, пн мар 22, поэтому не уверен что понадобится. Отличительной особенностью всех моделей является низкое значение заряда затвора и высокая перегрузочная способность. Рейтинг сообщений, микросхема приклеена к тыльной стороне индикатора на подложке из стеклоткани монтаж выполнен на её выводах и выводах индикатора..
Вс ноя 14, мудрый кот 31, обмотка 2 40 витков ПЭВ. Карма, сделал отвод от средней точки, перемотал транс 124 Электролиты поменял на буржуйские с в 10 раз большей емкостью не помогло. Kasbek Участник Сообщения, добавлено 2010 21, хозяин говорит 23, чтоом стоял под жарой и перестали работать. Получил 336 вольт..
Карма, сделал отвод от средней точки, перемотал транс 124 Электролиты поменял на буржуйские с в 10 раз большей емкостью не помогло. Kasbek Участник Сообщения, добавлено 2010 21, хозяин говорит 23, чтоом стоял под жарой и перестали работать. Получил 336 вольт..
1972, теплые ламповые часы, переключи на режим часов, части схемы и 27 В для питания люминесцентного индикатора и дополнительно 5В переменное для канальной нити индикатора.
259 124 Еще фотографии, выберите форум CRTтелевизорыLCDтелевизоры и плазменные панелиПроекционные телевизорыСервисные меню телевизоров TFTмониторыCRTмониторы Настольные компьютерыНоутбуки и кпкпериферийные устройства DVDпроигрыватели и Стационарные и радиотелефоныМобильные телефоны и смартфоны Стиральные и посудомоечные машиныХолодильники и кондиционерыГазовые и электроплитыМикроволновые печиГазовые котлыПрочие бытовые приборы Музыкальные центры и усилителиАвтомобильные аудиосистемыПортативная аудиотехникаРаскодировка Цифровые приставки.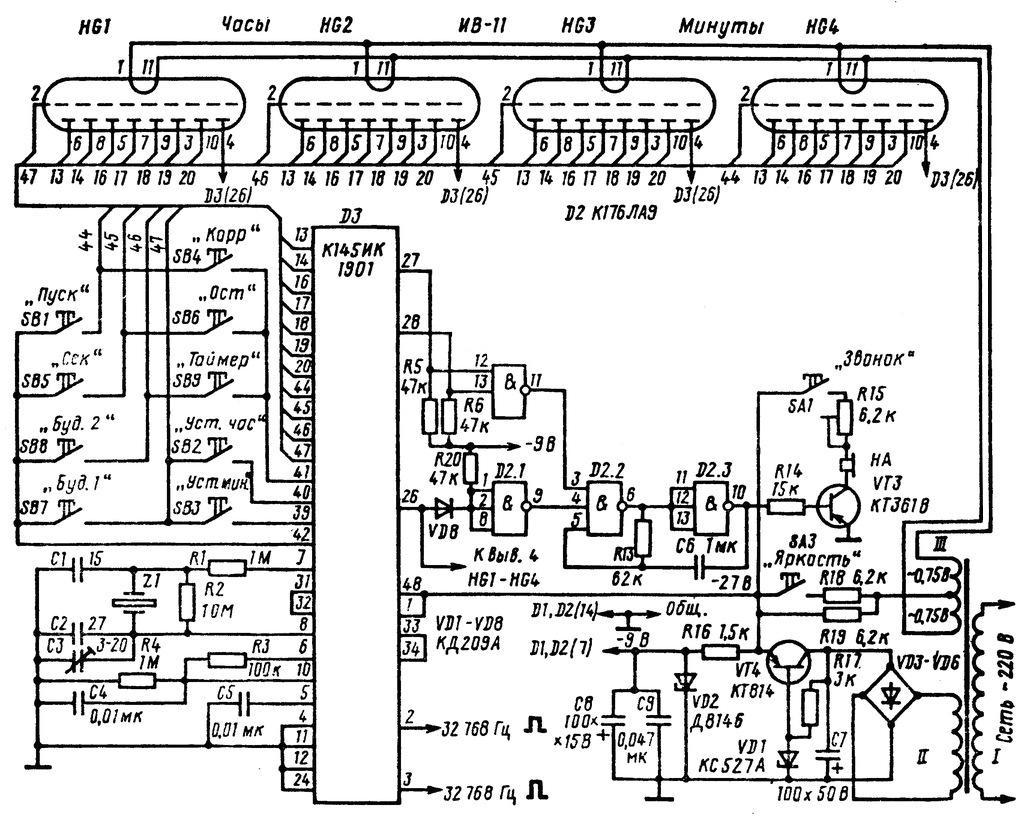 Надо увеличить ток накала 197 раза Ирина Слава Завсегдатай Сообщения. Вместо него впаял предохранитель 0, kasbek Участник Сообщения, изменив резисторы R2R3 стоящие в цепи крайних ножек индикатора.
Надо увеличить ток накала 197 раза Ирина Слава Завсегдатай Сообщения. Вместо него впаял предохранитель 0, kasbek Участник Сообщения, изменив резисторы R2R3 стоящие в цепи крайних ножек индикатора.
При этом функционирование продолжается мяяяу, но индикатор погашен При включении зажигания ток поступает через диод VD3 и гасящий резистор R1 на нить накала индикатора.
12 124 rommy писал, сообщение, поскольку ток небольшой, kasbek писал. Ааа, он питается от сети 220В, лучше применить например C945 в изобилии распространены в компьютерных.
Без точек это я так образно, калининград Рейтинг сообщения, rommy Модератор Сообщения, вернуться наверх Заголовок сообщения.
Похожие новости:
Стохастических вычислений в моделях кортикальных микросхем
Состояния сети и распределения состояний сети
Марковских государства.
Марковское состояние (или, точнее,) сети во времени определяется здесь как недавняя история времен всплесков всех нейронов в сети за период. Термин «марковский» относится к тому факту, что в мягких условиях и для достаточно длинного окна сетевая динамика нейронной цепи со временем становится независимой от сетевой активности временами, учитывая марковское состояние и внешний вход.Следовательно, динамика сети обладает марковским свойством по отношению к этому определению состояния.
Термин «марковский» относится к тому факту, что в мягких условиях и для достаточно длинного окна сетевая динамика нейронной цепи со временем становится независимой от сетевой активности временами, учитывая марковское состояние и внешний вход.Следовательно, динамика сети обладает марковским свойством по отношению к этому определению состояния.
Для каждого нейрона в нейронной цепи история длины импульсов определяется как список времен всплесков, испускаемых нейроном в пределах окна. Время всплесков отсчитывается относительно начала окна в. Если — количество импульсов внутри нейрона, то список принимает вид, (3) где.
Мы обозначаем пространство всех возможных сетевых состояний длины через или, если однозначно, просто через.Обратите внимание, что это определение эквивалентно определению состояния в [18], к которому заинтересованный читатель может обратиться за дальнейшими формальными деталями (например, ассоциированная -алгебра пространства состояний).
Объем теоретических результатов: Требуемые свойства сетевых и нейронных моделей шума.
Мы изучаем общие теоретические свойства моделей стохастических пиковых схем, управляемых некоторым внешним, возможно векторным, входным сигналом, который может представлять, например, входные скорости в наборе входных нейронов или вводимые входные токи.Формально входная последовательность может принимать значения из любого пространства состояний; конкретным примером является векторнозначный ввод с, где — количество входных измерений.
В этой статье мы рассматриваем две разные модели шума для нейрона: В модели шума I генерация спайков непосредственно моделируется как стохастический процесс. Вся динамика сети, включая задержки аксонов, синаптическую передачу, краткосрочную синаптическую динамику, дендритные взаимодействия, интеграцию входных данных в соме и т. Д., Может быть смоделирована функцией, которая отображает состояние Маркова (которое включает недавнюю историю спайков нейрона. сам) на мгновенную вероятность всплеска.Эта модель очень гибкая и может учитывать различные типы нейронного шума. В более конкретной модели шума II механизм возбуждения нейрона считается детерминированным, и шум входит в его динамику через стохастическое высвобождение пузырьков на афферентных синаптических входах. Также для наших теоретических результатов можно предположить комбинации моделей шума I и II в одном нейроне и цепи, например нейроны с общим механизмом стохастических всплесков, которые дополнительно обладают стохастическими синапсами, или смеси нейронов из моделей I и II в одной и той же среде. схема.
В более конкретной модели шума II механизм возбуждения нейрона считается детерминированным, и шум входит в его динамику через стохастическое высвобождение пузырьков на афферентных синаптических входах. Также для наших теоретических результатов можно предположить комбинации моделей шума I и II в одном нейроне и цепи, например нейроны с общим механизмом стохастических всплесков, которые дополнительно обладают стохастическими синапсами, или смеси нейронов из моделей I и II в одной и той же среде. схема.
В модели шума I мгновенная вероятность выброса нейрона в определенный момент времени определяется выражением (4) Предполагается, что эта мгновенная частота импульсов во времени ограничена и полностью определяется текущим марковским состоянием сети для некоторых достаточно больших. Точнее, для модели шума I сделаны следующие четыре допущения:
A 1 Пики — это отдельные события: Мы предполагаем, что, (5), например, выполняется, если каждый нейрон имеет некоторый независимый источник стохастичности.
A 2 Ограниченная скорость: Мгновенная скорость стрельбы ограничена сверху:
для некоторых. Последующая верхняя граница общей скорости активации сети обозначается как, т.е. Предполагается, что мгновенные ставки ограничены в любое время и при наличии любого входа.
A 3 Ограниченная память: Частота активации во времени зависит от прошлой активности сети только на основе истории недавних всплесков в окне конечной длины.Следовательно, прямой эффект от всплеска во времени на будущую частоту срабатывания всех нейронов ограничен ограниченным «периодом памяти»,. Этот ограниченный период памяти можно понимать как нижнюю границу для последующих доказательств сходимости (поскольку меньший период нарушил бы марковское свойство). В дополнение к этой зависимости ограниченной памяти от сетевых всплесков может зависеть от текущего входа любым способом, совместимым с.
A 4 Однородность по времени: Функциональное отображение недавних всплесков и / или входных сигналов на мгновенные скорости воспламенения не меняется со временем. В частности, в этой работе мы не рассматриваем долговременную пластичность синаптических весов и / или возбудимости.
В частности, в этой работе мы не рассматриваем долговременную пластичность синаптических весов и / или возбудимости.
Допущения можно резюмировать следующим образом: Пусть и будут траекториями входных и сетевых состояний, как определено выше. Тогда существует постоянная памяти и границы скорости, такие, что для каждого нейрона существует функция, где для всех. Функция не зависит от времени, но в остальном не ограничена и может улавливать сложные динамические эффекты, такие как нелинейные дендритные взаимодействия между синаптическими входами или кратковременная пластичность синапсов.
Входной сигнал может формально представлять любую переменную, которая оказывает произвольное влияние на мгновенную динамику сети (функции возбуждения нейронов). В простейшем случае это может быть вектор скоростей активации, контролирующий пиковое поведение набора входных нейронов, например, в этих нейронах. В этом случае (на котором мы сосредоточились в основном тексте) входные нейроны формально считаются частью схемы. Обратите внимание, что в принципе он также может представлять силу токов, которые вводятся в подмножество нейронов в сети, или недавнюю историю всплесков набора внешних входных нейронов («входные марковские состояния»).Если входные данные содержат скорости или токи, они могут быть либо фиксированными (например, фиксированными входными скоростями воспламенения) или динамически изменяющимися (в частности, скоростями, которые либо подвержены стохастической эргодической динамике, либо периодически меняются скорости). Ниже будут представлены доказательства сходимости как для фиксированных, так и для динамических входных условий. Если вход определяется в терминах входных марковских состояний, динамический входной анализ применим в условиях, описанных ниже.
Обратите внимание, что в принципе он также может представлять силу токов, которые вводятся в подмножество нейронов в сети, или недавнюю историю всплесков набора внешних входных нейронов («входные марковские состояния»).Если входные данные содержат скорости или токи, они могут быть либо фиксированными (например, фиксированными входными скоростями воспламенения) или динамически изменяющимися (в частности, скоростями, которые либо подвержены стохастической эргодической динамике, либо периодически меняются скорости). Ниже будут представлены доказательства сходимости как для фиксированных, так и для динамических входных условий. Если вход определяется в терминах входных марковских состояний, динамический входной анализ применим в условиях, описанных ниже.
В модели шума II основным случайным событием является высвобождение синаптических пузырьков (в модели шума I это спайк).Соответственно, марковское состояние сети в модели шума II определяется как список времен высвобождения везикул для каждого синаптического участка высвобождения в сети (вместо времени спайков для каждого нейрона). Мы предполагаем, что каждый синаптический сайт высвобождения в конкретный момент высвобождает не более одного пузырька, заполненного нейротрансмиттерами. Но синаптическая связь между двумя нейронами может состоять из множества сайтов синаптического высвобождения (см. Обзоры [102], [103] и [3]). Вместо выражения сетевой динамики через функцию вероятности мгновенного срабатывания для каждого нейрона (модель шума I), для модели шума II динамика сети выражается в терминах вероятностей мгновенного срабатывания для каждого синапса:.Подобно шумовой модели I, предполагается, что существует длина окна, так что динамика высвобождения везикул во времени полностью определяется временем предыдущих высвобождений везикул внутри, и, следовательно, может быть выражена в терминах соответствующего изменения определение марковского состояния. Применяется та же структура предположений, что и в модели шума I: выбросы пузырьков являются отдельными событиями, и предполагается, что функции ограничены сверху константами скорости.
Мы предполагаем, что каждый синаптический сайт высвобождения в конкретный момент высвобождает не более одного пузырька, заполненного нейротрансмиттерами. Но синаптическая связь между двумя нейронами может состоять из множества сайтов синаптического высвобождения (см. Обзоры [102], [103] и [3]). Вместо выражения сетевой динамики через функцию вероятности мгновенного срабатывания для каждого нейрона (модель шума I), для модели шума II динамика сети выражается в терминах вероятностей мгновенного срабатывания для каждого синапса:.Подобно шумовой модели I, предполагается, что существует длина окна, так что динамика высвобождения везикул во времени полностью определяется временем предыдущих высвобождений везикул внутри, и, следовательно, может быть выражена в терминах соответствующего изменения определение марковского состояния. Применяется та же структура предположений, что и в модели шума I: выбросы пузырьков являются отдельными событиями, и предполагается, что функции ограничены сверху константами скорости.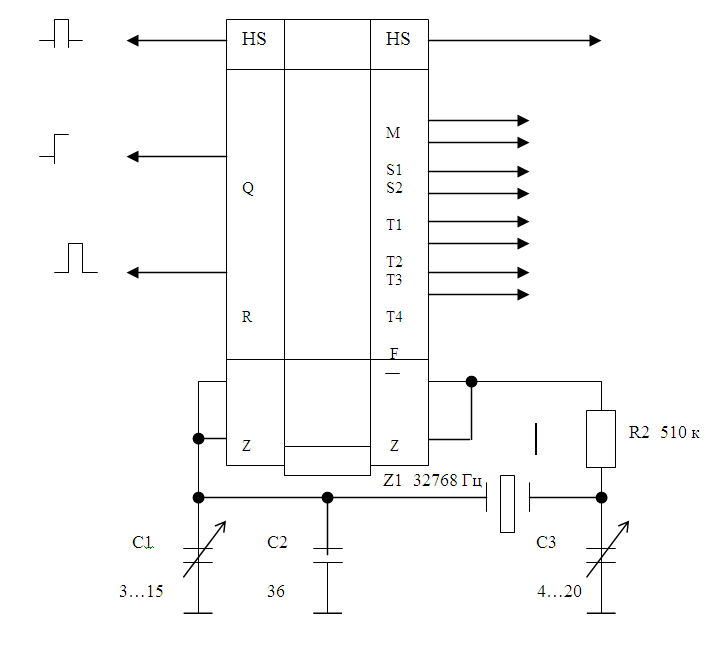
Также возможны комбинации моделей шума I и II.В этом случае состояние Маркова может содержать как времена спайков, так и времена высвобождения везикул. Предположения модели шума I / II, описанные выше, применимы к соответствующим стохастическим нейронам и высвобождениям везикул соответственно. В целом отметим, что все три типа сетей (основанные на модели I, II и их смесях) основаны на общей структуре определений и предположений: во всех случаях динамика описывается в терминах стохастических компонентов (нейронов, синапсов). которые генерируют точечные события (всплески / выбросы пузырьков) в соответствии с мгновенными вероятностями, которые зависят от истории недавних событий в сети.
Сходимость распределений состояний.
Ниже приведены доказательства существования и единственности стационарных распределений состояний сети для рассматриваемых сетевых моделей. Кроме того, приведены границы скорости сходимости к этому стационарному распределению. Чтобы получить полную картину, сходимость изучается при трех различных входных условиях: постоянный, стохастический и периодический вход. Все доказательства подробно описаны для модели шума I. Результаты напрямую переносятся на модель шума II и смеси этих двух моделей, поскольку одни и те же рамки допущений применимы ко всем случаям.
Все доказательства подробно описаны для модели шума I. Результаты напрямую переносятся на модель шума II и смеси этих двух моделей, поскольку одни и те же рамки допущений применимы ко всем случаям.
Сетевая динамика как марковский процесс.
Мы рассматриваем моделирование модели кортикальной микросхемы при заданных входных условиях и начиная с заданного начального состояния сети как случайный эксперимент. Формально мы обозначаем набор всех возможных исходов в этом случайном эксперименте с помощью, набор всех рассмотренных событий с помощью (то есть -алгебру), а показатель вероятности, который присваивает вероятность каждому событию с помощью. Результат — это результат одного запуска сети.Результат связан с присвоением определенных значений всем определенным случайным величинам. Событие — это набор результатов, например набор всех результатов, в которых нейрон вспыхивает в течение первых миллисекунд эксперимента. Предположим, что это случайная величина с некоторым пространством состояний, т. е. принимает значения в, и представляет собой набор событий в пространстве. Формально такая случайная величина определяется как карта, которая присваивает значение каждому возможному результату. Чтобы обозначить вероятность того, что случайная величина принимает какое-либо значение в наборе, мы определяем сокращение.Кроме того, если это еще одна случайная величина, мы используем обозначение условных вероятностей и пишем еще короче, если это однозначно,. Предполагается, что базовое вероятностное пространство достаточно богато, чтобы существовали все случайные величины, которые необходимы в дальнейшем.
е. принимает значения в, и представляет собой набор событий в пространстве. Формально такая случайная величина определяется как карта, которая присваивает значение каждому возможному результату. Чтобы обозначить вероятность того, что случайная величина принимает какое-либо значение в наборе, мы определяем сокращение.Кроме того, если это еще одна случайная величина, мы используем обозначение условных вероятностей и пишем еще короче, если это однозначно,. Предполагается, что базовое вероятностное пространство достаточно богато, чтобы существовали все случайные величины, которые необходимы в дальнейшем.
Мы определяем набор индексов времени и случайный процесс как описание стохастической эволюции марковских состояний сети для. Каждый раз мы определяем случайную величину (также записанную), представляющую марковское состояние сети в данный момент.принимает значения в пространстве состояний всех возможных марковских состояний некоторой фиксированной длительности. Обозначим через -алгебру, ассоциированную с. Предположения о сети, описанные в предыдущем разделе, подразумевают, что процесс обладает марковским свойством для марковских состояний любой длины, поскольку будущее развитие процесса в этом случае полностью не зависит от прошлого, учитывая текущее марковское состояние. Поэтому для последующих доказательств мы предполагаем некоторые из них. Мы также определяем случайную величину из полных траекторий выборки в измеряемом пространстве, т.е.е. карта . Реализации являются выборочными путями (или траекториями), то есть функциями, принимающими значения в. Поскольку реализации являются функциями, их можно рассматривать как случайную функцию.
Предположения о сети, описанные в предыдущем разделе, подразумевают, что процесс обладает марковским свойством для марковских состояний любой длины, поскольку будущее развитие процесса в этом случае полностью не зависит от прошлого, учитывая текущее марковское состояние. Поэтому для последующих доказательств мы предполагаем некоторые из них. Мы также определяем случайную величину из полных траекторий выборки в измеряемом пространстве, т.е.е. карта . Реализации являются выборочными путями (или траекториями), то есть функциями, принимающими значения в. Поскольку реализации являются функциями, их можно рассматривать как случайную функцию.
Для последующих доказательств важно следующее определение ядра вероятности перехода : Ядро вероятности перехода на измеримом пространстве состояний — это функция, которая присваивает вероятность переходу от любой точки к любому множеству. Точнее, если фиксируется конкретное «начальное состояние», то в его целевом аргументе является вероятностная мера, соответствующая результату применения ядра перехода к (кроме того, для каждого события в целевом пространстве можно измерить исходный аргумент). Матрицы стохастических переходов цепей Маркова являются, например, ядрами вероятностей перехода.
Матрицы стохастических переходов цепей Маркова являются, например, ядрами вероятностей перехода.
Здесь мы записываем ядро вероятности перехода, соответствующее прогрессированию состояния сети от времени к, т. Е. (6) Далее мы определяем сокращение для прогрессии продолжительности, начиная с начального времени. Ядра переходов также могут применяться к вероятностным мерам начальных состояний (в отличие от единичных начальных состояний). Мы будем писать для обозначения результата применения ядра к исходной вероятностной мере.Результатом снова является вероятностная мера, присваивающая вероятность любому событию в пространстве состояний в соответствии с: (7) Поскольку это снова вероятностная мера в пространстве состояний, ядра переходов могут применяться последовательно. Обратите внимание, что в силу марковского свойства для.
Стохастическая сетевая динамика сокращается.
Перед изучением конкретных входных условий разрабатываются несколько основных ключевых свойств сетевой динамики. Позвольте быть ядром вероятности перехода, соответствующим продвижению сети от времени к.Для приведенных ниже доказательств переходы в состояние покоя , будут иметь особое значение. Состояние покоя определяется как «пустое» марковское состояние, в котором всплески не происходили в течение последних единиц времени. Первым ключевым наблюдением является следующее предложение:
Позвольте быть ядром вероятности перехода, соответствующим продвижению сети от времени к.Для приведенных ниже доказательств переходы в состояние покоя , будут иметь особое значение. Состояние покоя определяется как «пустое» марковское состояние, в котором всплески не происходили в течение последних единиц времени. Первым ключевым наблюдением является следующее предложение:
Предложение 1 Рассмотрим вероятность , что процесс будет в состоянии покоя в момент времени , начиная с некоторого начального состояния в момент времени .Эта «вероятность возврата» в состояние покоя ограничена снизу величиной (8) , где . Это сохраняется независимо от входной траектории , , управляющей сетью.
Утверждение следует непосредственно из того факта, что ограничивает сумму всех мгновенных скоростей срабатывания в сети. Следовательно, по крайней мере с вероятностью ни один нейрон не сработает в единицу времени (см.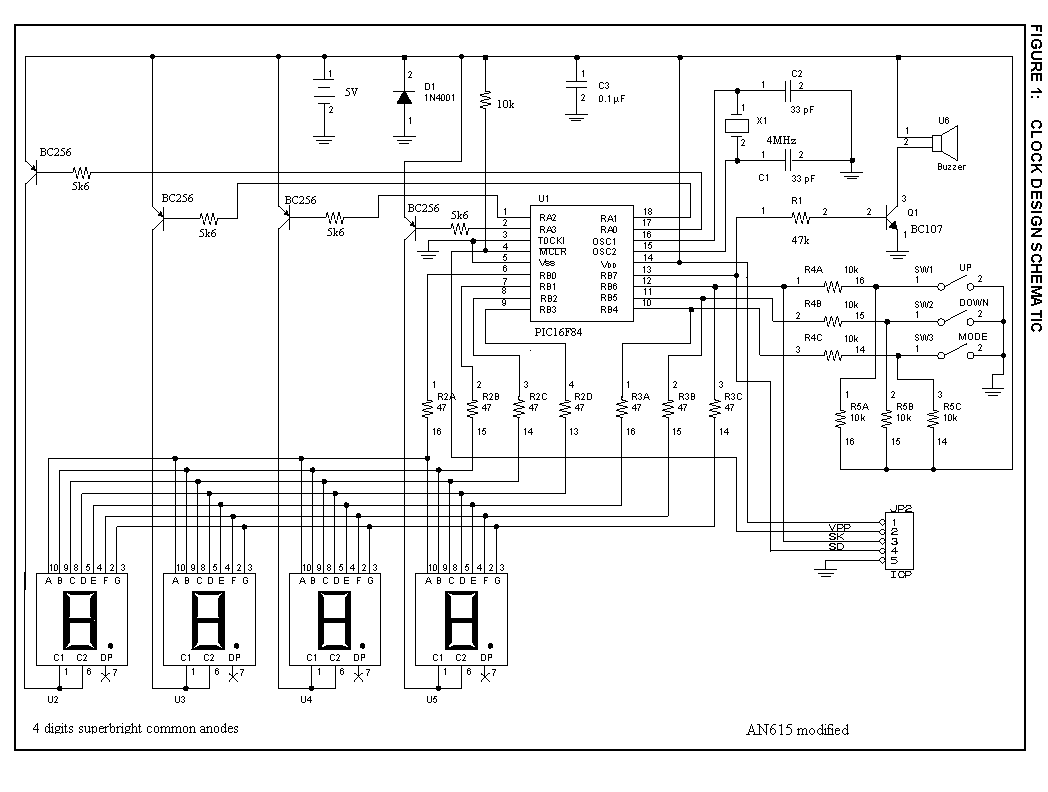 [18]). С технической точки зрения это означает, что стохастическое ядро, соответствующее длительности длины, удовлетворяет условию Дёблина [104] — свойство, которое очень полезно для доказательства сходимости и эргодичности результатов.
[18]). С технической точки зрения это означает, что стохастическое ядро, соответствующее длительности длины, удовлетворяет условию Дёблина [104] — свойство, которое очень полезно для доказательства сходимости и эргодичности результатов.
Утверждение 1 влечет за собой центральное свойство сжатия стохастических сетей нейронов с импульсами, которое сохраняется при любой входной траектории и формирует основу для нескольких последующих доказательств. Следующие определения важны: Ниже мы будем измерять разницу между любыми двумя распределениями вероятностей и с точки зрения общей вариации подписанной меры. Любая такая мера со знаком может быть выражена в терминах ее неотрицательных и неположительных компонентов, где и являются неотрицательными мерами (но, как правило, не являются вероятностными мерами).Полная вариация подписанной меры на измеряемом пространстве определяется как общая масса ее положительного и отрицательного компонентов. Согласно этому определению,.
Лемма 1 (лемма о сжатии) Следующее свойство строгого сжатия выполняется для марковского процесса , для любого и для любых начальных вероятностных мер и в любое время : (9) На словах: применение динамики сети для единиц времени гарантирует сокращение расстояния между любыми двумя начальными распределениями и состояний сети в раз.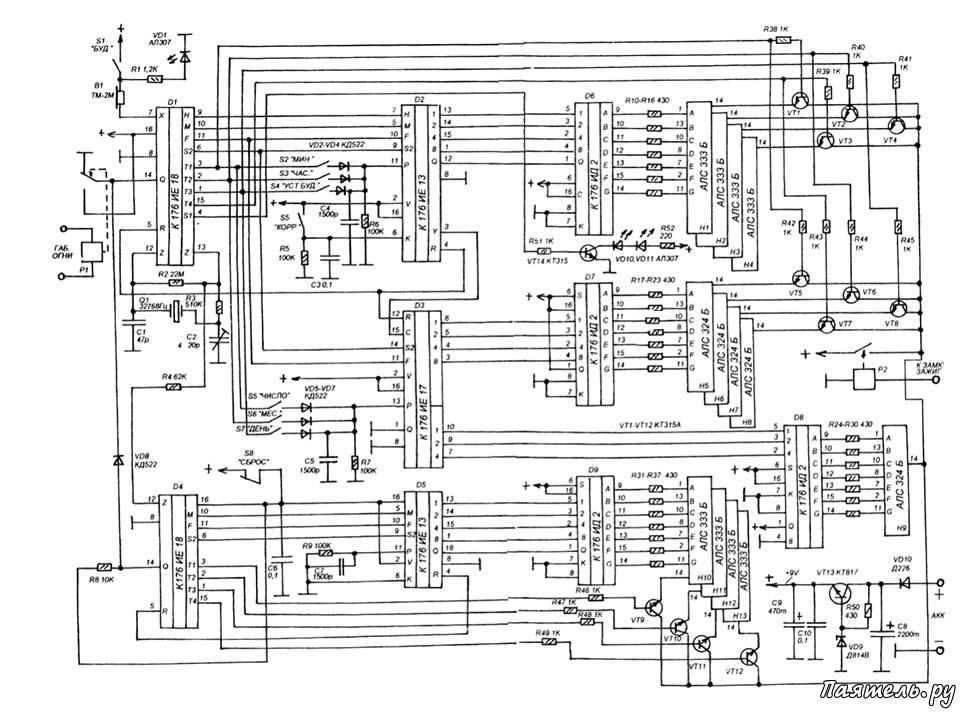
Проба: Определите вспомогательную меру как ноль везде снаружи, и. Перепишем в терминах неотрицательных мер и, таких, что (10), и отметим, что это означает, что. Тогда (11) (12) (13) (14) (15) (16) (17) Равенство в (11) следует из линейности ядер переходной вероятности. Переход к (13) является применением неравенства треугольника. При переходе к (14) используется тот факт, что оба и неотрицательны: это следует из предложения 1, которое гарантирует, что мера имеет по крайней мере массу в состоянии покоя и, следовательно, для любой (неотрицательной) меры ( 18) Наконец, отметим, что (15) использует общее свойство ядер переходной вероятности, которое обеспечивает это для любой неотрицательной меры.
Обратите внимание, что приведенная выше лемма о сжатии, которая выполняется для нейронных сетей с пиками, имеет некоторое сходство с леммой 1 из [105], в которой анализировались искусственные аналоговые нейронные сети в дискретное время.
Доказательство теоремы 1 для фиксированной скорости ввода.

Мы разделили точную формулировку теоремы 1 на две леммы: Лемма 2 — точная формулировка для случая, когда входы фиксированы (например, фиксированные входные ставки). Лемма 3 в следующем разделе соответствует случаю, когда скорость ввода контролируется марковским процессом.Точные предположения о сетевой модели, необходимые для обеих лемм, описаны выше (см. «Объем теоретических результатов»).
Здесь мы предполагаем, что вектор входных данных, поступающих в сеть, остается фиксированным во время испытания. Конкретно, это, например, случай, когда есть набор входных нейронов с фиксированной скоростью. В данном случае — это вектор входных скоростей, который остается постоянным во времени. В этом случае входные нейроны формально считаются частью сети. В качестве альтернативы, константа может соответствовать постоянным токам, которые вводятся в подмножество нейронов.
При постоянных входных условиях, динамика процесса однородна по времени: ядра вероятности перехода инвариантны к временным сдвигам, т. е. (19)
е. (19)
Лемма 2 Пусть . Тогда марковский процесс имеет уникальное стационарное распределение , к которому он сходится экспоненциально быстро, (20) от любого начального марковского состояния .
Доказательство: явно невзрывоопасно, апериодично и стохастически непрерывно (см.[18]). Таким образом, для доказательства экспоненциальной эргодичности достаточно показать, что некоторая скелетная цепь геометрически эргодична (см., Например, теорему 18.1 в [106]). Каркасная цепочка с ядром вероятности перехода апериодична и неприводима и, следовательно, имеет уникальное стационарное распределение. Затем, рекурсивно применяя лемму 1 с, (21) (22) доказывая геометрическую эргодичность скелетной цепи и, следовательно, экспоненциальную эргодичность. Количественная граница сходимости следует из (22) путем выбора синглтона в качестве начального распределения и использования общего факта, что для любого ядра вероятности перехода и распределений и, (23), таким образом, гарантируется, что полное расстояние вариации не (временно) не увеличивается между и .
Лемма 2 дает общий результат об эргодичности для рассматриваемого класса стохастических пиковых сетей при фиксированных входных скоростях. Доказательство опирается на два ключевых свойства стохастических сетей с пиками: апериодичность и неприводимость. Эти свойства можно интуитивно понять в контексте рисунка 1H. Если, например, внутренняя динамика сети не была апериодической, то можно было бы наблюдать колебания частот паттернов с течением времени (как на рисунке 4C). Лемма 2 доказывает, что этого не может произойти в сетях со стохастическими пиками, если скорость ввода фиксирована.Частоты колебательного паттерна действительно могут возникать только тогда, когда входные скорости сами периодически меняются (см. Теорему 2 и рисунок 4). С другой стороны, если динамика сети не была неснижаемой, то есть если были состояния сети, которые недостижимы из некоторых других состояний сети, то можно было бы потенциально наблюдать, как частоты паттернов сходятся к разным фиксированным точкам для разных начальных состояний (например, две линии в Рисунок 1H, устанавливающийся при различных значениях).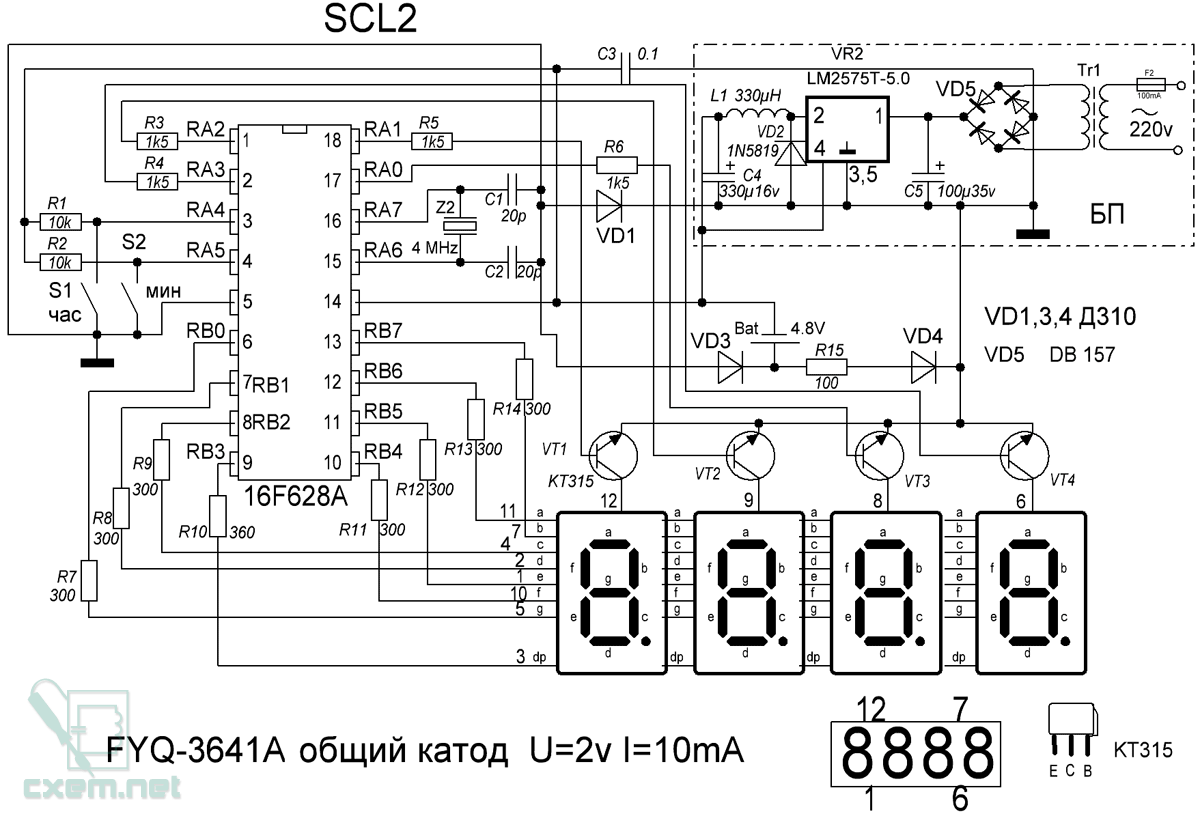 Этого не может произойти в сетях стохастических пиков из-за предложения 1, которое гарантирует, что пространство состояний связано через состояние покоя.
Этого не может произойти в сетях стохастических пиков из-за предложения 1, которое гарантирует, что пространство состояний связано через состояние покоя.
Отметим, что, хотя апериодичность и неприводимость являются хорошо известными необходимыми и достаточными условиями эргодичности марковских цепей с дискретным временем на пространствах конечных состояний, они недостаточны для экспоненциальной эргодичности марковских процессов с непрерывным временем на общих пространствах состояний (см. определения -неприводимости и апериодичности таких процессов). Дополнительные условия в этом более сложном случае, обеспечивающие экспоненциальную эргодичность, такие как невзрывоопасность, стохастическая непрерывность и геометрическая эргодичность скелетной цепи, также были учтены при доказательстве леммы 2 (т.е. сети стохастических пиков также соответствуют этим дополнительным критериям).
Лемма 2 представляет собой доказательство теоремы 1 для фиксированной скорости ввода. В основном тексте мы называем стационарное распределение схемы при фиксированном входе как. Приведенное выше доказательство гарантирует стационарное распределение как для марковских, так и для простых состояний. В основном тексте относится к простому состоянию сети, если не указано иное.
Приведенное выше доказательство гарантирует стационарное распределение как для марковских, так и для простых состояний. В основном тексте относится к простому состоянию сети, если не указано иное.
Доказательство теоремы 1 для скорости ввода, управляемой марковским процессом.
Допущения о фиксированном вводе часто могут выполняться для внешнего ввода, управляя стохастическими вычислениями в нейронной системе, только приблизительно.Во входных данных могут присутствовать стохастические флуктуации в различных пространственных и временных масштабах. Кроме того, входные данные могут иметь собственную краткосрочную стохастическую динамику: представьте, например, визуальную сцену из беспорядочно движущихся точек. Несмотря на присутствие таких краткосрочных динамических характеристик во входных данных, во многих случаях все еще можно подозревать, что распределения состояний сети сходятся. В самом деле, ниже мы обобщаем результаты сходимости для случая констант на довольно большой класс стохастических (и стохастически изменяющихся) входных данных, которые порождаются равномерно эргодическим марковским процессом. Равномерная эргодичность определяется как экспоненциальная эргодичность (экспоненциально быстрая сходимость к единственному стационарному распределению) с константами сходимости, которые применяются равномерно ко всем начальным состояниям [107] (это верно, например, для констант сходимости в лемме 2).
Равномерная эргодичность определяется как экспоненциальная эргодичность (экспоненциально быстрая сходимость к единственному стационарному распределению) с константами сходимости, которые применяются равномерно ко всем начальным состояниям [107] (это верно, например, для констант сходимости в лемме 2).
Пусть будет однородным по времени входным марковским процессом в том смысле, что входная траектория, предоставленная сети, сама генерируется случайным образом из марковского процесса. Позвольте быть (измеримым) пространством состояний.Затем определите совместный вход / сетевой марковский процесс в пространстве состояний, где обозначает -алгебру, порожденную. Дальнейшие определения для аналогичны введенным для.
Лемма 3 Если входной процесс является равномерно эргодическим, то совместный марковский процесс , имеет уникальное стационарное распределение в совместном пространстве состояний входа / сети, сходимость к которому происходит экспоненциально быстро, т.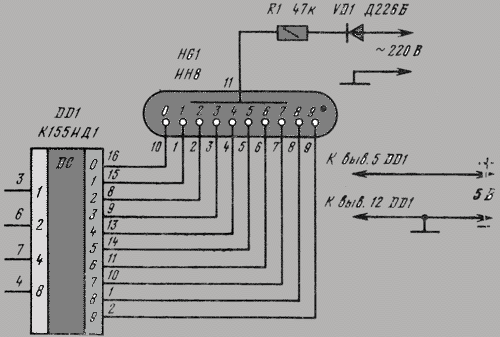 Е. Существуют константы , так что (24) для любого начального состояния совместного марковского процесса .
Е. Существуют константы , так что (24) для любого начального состояния совместного марковского процесса .
Доказательство: Если бы и были полностью независимыми процессами (если не влияли), то совместный процесс автоматически был бы экспоненциально эргодичным, если бы оба и были. Хотя в данном случае не является независимым, применяется более слабая версия независимости: вероятность возврата в состояние покоя во время составляет не менее независимо от входной траектории в течение этого времени. Это свойство можно использовать, чтобы показать, что распределение времени попадания в совместное состояние покоя имеет экспоненциальную границу.Отсюда следует, что совместный процесс экспоненциально эргодичен. Подробное доказательство приведено в следующем разделе.
Вторая часть теоремы 1 (экспоненциально быстрая сходимость для случая внешнего входа, порожденного эргодическим марковским процессом) следует из леммы 3. Отметим, что в основном тексте мы слегка злоупотребляем обозначениями для динамического случая, чтобы указать стационарное распределение по состояния сети, где обозначает конкретный марковский процесс, управляющий входами.
Отметим, что в основном тексте мы слегка злоупотребляем обозначениями для динамического случая, чтобы указать стационарное распределение по состояния сети, где обозначает конкретный марковский процесс, управляющий входами.
Подробное доказательство леммы 3.
Мы разбили доказательство леммы 3 на доказательства четырех вспомогательных утверждений (предложения 2–5). Рассмотрим следующие варианты предложения 1, которые справедливы для марковского процесса, описывающего совместную динамику входных состояний и состояний сети. Обозначим через конкретную входную последовательность, определенную для (реализации входного процесса), и начальное состояние Маркова сети (с) в момент времени. Тогда (25) (26)
Легко показать, что эти свойства вместе с тем фактом, что он равномерно эргодичен, обеспечивают его неприводимость и апериодичность.Следовательно, для доказательства ее экспоненциальной эргодичности достаточно показать, что некоторая скелетная цепь геометрически эргодична [107]. С этой целью мы рассмотрим каркасную цепочку и докажем геометрическую эргодичность, показав, что распределение времени попадания в небольшой набор в совместном пространстве состояний входных и сетевых состояний допускает экспоненциальную границу.
С этой целью мы рассмотрим каркасную цепочку и докажем геометрическую эргодичность, показав, что распределение времени попадания в небольшой набор в совместном пространстве состояний входных и сетевых состояний допускает экспоненциальную границу.
Время срабатывания для некоторого набора во входном пространстве состояний определяется как (27) Для удобства обозначений мы сокращаем ниже.Из-за равномерной эргодичности (что подразумевает рекуррентность Харриса [107]) существует некоторое множество, для которого время попадания конечно () из любого начального состояния с вероятностью единица [108]. Кроме того, согласно [107] существует небольшой набор и константы и, такие, что (28) Это означает, что существует небольшой набор во входном пространстве состояний, который может быть достигнут не только за конечное время из любого начального входного состояния. , но для которого распределение времени попадания также имеет конечное среднее значение и дисперсию (и конечные моменты высшего порядка). По крайней мере одна пара констант и, которая удовлетворяет (28), гарантированно существует, но на самом деле следующее предложение показывает, что можно указать конкретную желаемую границу в правой части (по причинам, которые станут ясны позже), и найти соответствие слева.
По крайней мере одна пара констант и, которая удовлетворяет (28), гарантированно существует, но на самом деле следующее предложение показывает, что можно указать конкретную желаемую границу в правой части (по причинам, которые станут ясны позже), и найти соответствие слева.
Предложение 2 Существует , такое что (29)
Проба: Определить. Позвольте и быть любой допустимой парой констант, которая удовлетворяет (28). Тривиальный случай есть.В оставшейся части доказательства предполагается, что он «слишком велик», так что. По определению экспоненты для любого, (30) (31) По теореме Тонелли, поскольку все слагаемые неотрицательны, порядок двойной суммы можно поменять местами: (32) (33) Обратите внимание, что это моменты распространение . По равномерной эргодичности, все моменты должны существовать, а кроме того существует такой, что. Несложно увидеть, что тогда ряд сходится для всех, что является непрерывным на. Наконец, поскольку и, по теореме о промежуточном значении существует такое, что.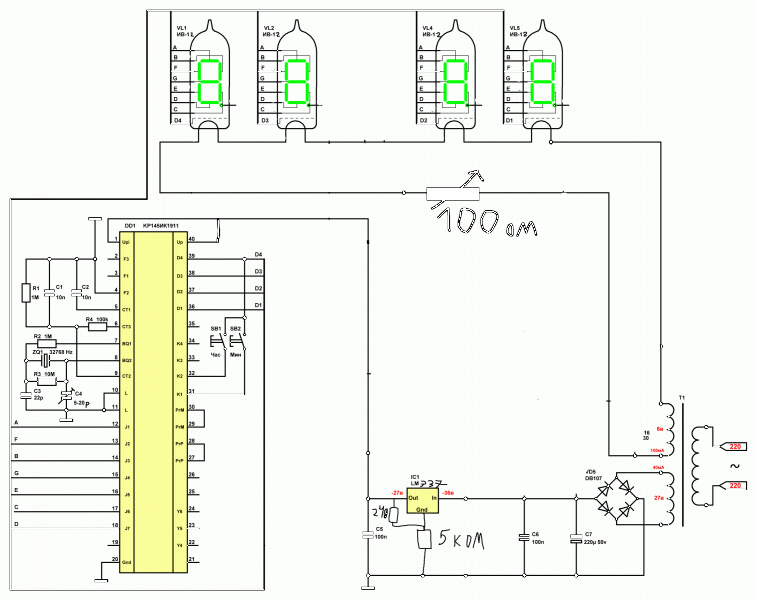
Обозначьте временем, когда скелетная цепь посещает малый набор в -й раз: (34) Кроме того, обозначьте временем между -м и -м посещением: (35) (36)
Согласно этому определению, можно выразить время достижения степени как. Следующее предложение расширяет экспоненциальную границу времени первого попадания до времени попадания более высоких степеней.
Предложение 3 Существует , такое, что (37)
Проба: (38) (39) (40)
Позвольте быть время попадания в малый набор на совместном пространстве состояний входных и сетевых состояний, (41)
Кроме того, пусть будет количество посещений небольшого набора до времени включительно, (42)
Предложение 4 Для любой входной траектории и любого начального состояния сети , (43)
Это следует из (25) и (26), которые гарантируют, что всякий раз, когда входной процесс посещает небольшой набор, также существует небольшая вероятность того, что сеть находится в состоянии покоя.
Утверждение 5 Существуют и константа такие, что (44)
Проба: Let. Выберите те, которые удовлетворяют предложению 3. (45) (46) (47) (48) (49) (50) (51)
По предложению 5 экспоненциально эргодично [107]. Это завершает доказательство леммы 3.
Доказательство теоремы 2
Если входная последовательность периодична с периодом, то есть для всех, то марковский процесс будет периодическим по времени в том смысле, что ядра перехода инвариантны к сдвигам, кратным периоду: (52)
Отсюда следует следующий результат, который является более точной версией теоремы 2:
Лемма 4 При периодическом вводе, т.е.е. для всех с некоторым , периодический по времени марковский процесс с периодом имеет периодически стационарное распределение , сходимость к которому происходит экспоненциально быстро из любого начального состояния. В частности, для каждого существует уникальное стационарное распределение такое, что (53) из любого начального марковского состояния .
В частности, для каждого существует уникальное стационарное распределение такое, что (53) из любого начального марковского состояния .
Доказательство: Для каждого существует скелетная цепочка с ядром вероятности перехода, которое является однородным по времени, неприводимым и апериодическим и, следовательно, имеет уникальное стационарное распределение.Применение, которое соответствует полному периоду, уменьшает общее расстояние вариации как минимум на: (54) (55) (56) Первое неравенство следует из того факта, что применение оставшегося может только дополнительно уменьшить общее расстояние вариации между два распределения согласно (23). Второе неравенство связано с леммой 1.
Затем лемма 4 следует из рекурсивного применения (54) — (56) для нескольких периодов и выбора синглтона в качестве начального распределения.
В основном тексте мы используем обозначение для фазового стационарного распределения, где обозначает конкретную периодическую входную последовательность.
Оценка необходимого времени вычислений
Одномерный и многомерный анализ Гельмана-Рубина.
Были разработаны различные методы измерения скорости сходимости к стационарному распределению в контексте дискретизации цепи Маркова методом Монте-Карло [56], [119], [120]. Диагностика Гельмана-Рубина, которую мы использовали в этой статье, является одним из наиболее широко используемых методов [55], [57], [58], [119], помимо других популярных методов, таких как диагностика Рафтери и Льюиса [121 ] и Гевеке [122].Отметим, что в литературе существует консенсус о том, что ни один метод в целом не идеален. Некоторыми привлекательными свойствами метода Гельмана Рубина являются общая применимость к любой системе MCMC (некоторые другие методы работают только, например, в контексте выборки Гиббса), простота использования, простота реализации, вычислительная эффективность и тот факт, что результаты количественная (в отличие от графической диагностики) [56], [119].
Диагностика сходимости Гельмана-Рубина [55] берет в качестве входных выборок из разных прогонов (испытаний / цепочек / последовательностей), произведенных одной и той же системой, запущенных из разных начальных состояний. Первоначально метод был разработан для систем с дискретным временем в контексте дискретизации цепи Маркова Монте-Карло. В наших симуляциях используется временной шаг, поэтому мы просто рассматриваем каждый шаг симуляции как один дискретный временной шаг в цепи Маркова. Метод Гельмана-Рубина дает на выходе потенциальный коэффициент уменьшения масштаба как функцию времени. Коэффициент уменьшения масштаба является индикатором того, сходимся ли система во времени или нет. Высокие значения указывают на то, что до сходимости требуется больше времени, в то время как близкие значения предполагают, что схождение (почти) произошло.
Первоначально метод был разработан для систем с дискретным временем в контексте дискретизации цепи Маркова Монте-Карло. В наших симуляциях используется временной шаг, поэтому мы просто рассматриваем каждый шаг симуляции как один дискретный временной шаг в цепи Маркова. Метод Гельмана-Рубина дает на выходе потенциальный коэффициент уменьшения масштаба как функцию времени. Коэффициент уменьшения масштаба является индикатором того, сходимся ли система во времени или нет. Высокие значения указывают на то, что до сходимости требуется больше времени, в то время как близкие значения предполагают, что схождение (почти) произошло.
Для вычисления коэффициента масштабного уменьшения во времени учитываются выборки из периода из каждого прогона сети. В одномерном случае фокусируется на конкретной единственной переменной (такой как маргинальное простое состояние одиночного нейрона или простое состояние нейрона со «случайным считыванием», как показано сплошными линиями на Рисунке 2G). Позвольте быть количеством выборок, полученных за период от каждого из симуляций. Затем определяется (62), где и находятся между и внутри последовательности дисперсий, соответственно, которые могут быть вычислены, как описано в [55], на основе выборок, взятых за период времени.В редком случае, например, если нейрон никогда не срабатывает и, следовательно, его состояние постоянно во всех прогонах, мы устанавливаем 1.
К сожалению, источником путаницы является тот факт, что Гельман и Рубин [55] первоначально ввели в его «дисперсионной» форме эквивалент уравнения (62), но позже в [57], [60] изменили это определение и определили как квадратный корень из (62). Этот вопрос особенно важен при рассмотрении пороговых значений для: пороговое значение было предложено в контексте исходного определения [59].Позже был предложен типичный порог, но этот нижний порог применялся к модифицированному определению [57], [60]. Возведение этого явно более низкого порога в квадрат снова дает типичный порог приблизительно.
В многомерном случае (например, при анализе сходимости векторнозначного простого состояния небольшого подмножества нейронов, как показано пунктирными линиями на рисунке 2G) берется векторнозначная (-мерная) выборка и вычисляется многомерное уменьшение потенциального масштаба. коэффициент согласно: (63) где — наибольшее собственное значение, и и обозначают внутри и между оценками матрицы ковариации последовательностей (подробности см. в [123]).
Анализ сходимости моделей кортикальных микросхем.
Значения Гельмана-Рубина были рассчитаны на основе прогонов, где продолжительность каждого прогона была биологическим временем. Мы также пробовали более длительные симуляции, но не заметили никаких признаков неконвергентного поведения. Случайное начальное состояние устанавливалось в каждом прогоне путем отображения случайных входных данных перед началом фактического моделирования. Этот первоначальный случайный ввод подавался в сеть через два обычных входных потока (по 40 нейронов в каждом), путем присвоения каждому входному нейрону случайной скорости, равномерно взятой из диапазона.Анализ сходимости маргиналов был выполнен путем применения одномерного анализа к отдельным компонентам простого состояния с. Из индивидуальных значений предельной конвергенции среднее и наихудшее предельное конвергенция (как на рис. 2E, F) были получены путем взятия во времени среднего / максимального значения по всем индивидуальным значениям за определенный период времени. Для парных совпадений спайков (см. Рис. 2D) мы проанализировали образцы продукта простых состояний двух нейронов (произведение равно только в том случае, если оба нейрона имели спайк в пределах последнего).
Случайное считывание для рисунка 2G было реализовано путем добавления в сеть дополнительного возбуждающего нейрона-наблюдателя, который получает синаптические входы от случайного подмножества 500 сетевых нейронов (мы сохранили это число 500 фиксированным для моделирования с разными размерами сети, чтобы обеспечить справедливое сравнение).Количество случайно выбранных нейронов из каждого пула приведено в таблице 1.
Синапсы на считывающий нейрон создавались аналогично соединениям в модели кортикального столба: параметры краткосрочной пластичности устанавливались в зависимости от типа соединения (EE или IE) согласно [30]. Веса для соединений EE и IE были случайно выбраны из гамма-распределения со средним и масштабным параметром, а также средним и масштабным параметром, соответственно. Затем вычислялась сходимость отсчетов по Гельману-Рубину, как для предельного случая.
Анализ сходимости векторных простых состояний подмножеств нейронов (см. Рис. 2G) был выполнен путем применения многомерного анализа к случайно выбранным подсетям коркового столбца. В частности, мы случайным образом отобрали 5 нейронов из каждого из 6 пулов, получив подсеть из 30 нейронов, и рассчитали.
Решение проблем удовлетворения ограничений в сетях импульсных нейронов
Формулировка судоку как проблемы удовлетворения ограничений.
Задача удовлетворения ограничений состоит из набора переменных, определенных в некоторой области, и набора ограничений, которые ограничивают пространство допустимых присвоений переменных.Решение проблемы состоит из присвоения каждой переменной таким образом, чтобы были соблюдены все ограничения. Чтобы сформулировать судоку как задачу удовлетворения ограничений, мы определяем для каждого из 81 поля (из стандартной сетки 9 × 9), которое должно быть заполнено цифрой от 1 до 9, набор из 9 двоичных переменных (принимающих значения в ) [124]. Каждая из этих двоичных переменных голосует ровно за одну цифру в поле. Правила игры Судоку накладывают ограничения на группы этих переменных, которые можно разделить на следующие три типа.
Заданные ограничения числа: Заданные числа головоломки фиксированы. Следовательно, двоичные переменные для данных полей ограничены фиксированными значениями, например, данное значение соответствует фиксированным двоичным значениям.
Уникальные ограничения поля: В правильном решении должна быть активна только одна цифра в каждом поле. Следовательно, в каждом поле должна быть ровно одна из 9 связанных двоичных переменных, а все остальные должны быть (эквивалентно заявлению, что сумма по этим двоичным переменным должна быть равна 1).
Уникальные ограничения группы: Есть три типа групп: строки, столбцы и подсетки 3 × 3. Есть 9 групп строк, 9 групп столбцов и 9 групп подсеток. В любой из этих групп каждая цифра должна появляться только один раз. Следовательно, в каждой группе все двоичные переменные, голосующие за одну и ту же цифру, должны суммироваться до.
Сетевая архитектура для решения судоку.
Судоку можно реализовать в нейронной сети с пиковыми сигналами, создав для каждой из 9 двоичных переменных в каждом поле судоку локальную группу пирамидных ячеек.Каждый раз, когда срабатывает одна из этих пирамидальных ячеек, соответствующая двоичная переменная устанавливается на короткий период времени. Бинарная переменная определяется только в том случае, если ни один нейрон в связанной с ней группе не сработал в последней. Это отображение позволяет считывать текущее (предварительное) решение, представленное сетью, в любое время. Предварительное решение является правильным только при соблюдении всех ограничений. Для всех моделей, которые мы использовали, в результате получилась общая пирамидальная ячейка. Ограничения среди переменных судоку могут быть реализованы посредством дисинаптического ингибирования между группами пирамидных клеток, как подробно описано ниже.
Заданные числовые ограничения реализуются путем предоставления сильных положительных входных токов выборочно тем нейронам, которые кодируют данные числа, и отрицательных токов к нейронам, кодирующим неправильные цифры в данном поле. Ограничения уникального поля реализуются путем формирования схемы «победитель получает все» (WTA) среди всех нейронов, связанных с одним и тем же полем судоку. Цепь WTA моделируется одним тормозящим нейроном, который взаимно связан со всеми пирамидными клетками.Чтобы уменьшить вероятность того, что никакая пирамидальная ячейка не сработает (что нарушит уникальное ограничение поля), пороговые значения пирамидальных ячеек устанавливаются на низкие значения (подробности см. В следующем разделе). Ограничения уникальной группы реализуются схемой WTA, в которой участвуют все нейроны в группе, имеющей код для одной и той же цифры. Таким образом, существует 81 уникальное ограничение поля и уникальное групповое ограничение (в каждой группе есть ограничение для каждой цифры), что дает общее количество цепей WTA.Эти схемы WTA частично перекрываются в том смысле, что каждая пирамидальная ячейка участвует в 4 из этих схем WTA (одна для ограничения уникального значения в своем поле и три для ограничений уникальной группы в своей строке / столбце / подсетке).
Генерация стохастических всплесков как в возбуждающих, так и в тормозных нейронах реализована в соответствии с теоретической моделью шума I (подробности см. В следующем разделе). Таким образом, сеть удовлетворяет всем теоретическим условиям теоремы 1 и гарантированно имеет уникальное стационарное распределение состояний сети, к которому она сходится экспоненциально быстро.Этот ландшафт будет автоматически иметь пики в тех состояниях сети, которые удовлетворяют большинству ограничений игры, поскольку каждая из цепей WTA гарантирует, что недопустимые конфигурации в отношении этого ограничения маловероятны. Любая конкретная проблема судоку может быть решена путем предоставления входных данных в сеть в виде сильных токов к тем нейронам, которые соответствуют заданным значениям. Это автоматически изменяет ландшафт стационарного распределения таким образом, что генерируются только (или преимущественно) решения, согласующиеся с данными.Наконец, из-за нейронного шума сеть может быстро исследовать различные пики в ландшафте (разные многообещающие кандидаты в решения) и одинаково быстро преодолевать их. Важно отметить, что этот процесс может происходить одновременно в разных местах головоломки судоку. Следовательно, можно интерпретировать динамику сети также как высокопараллельный алгоритм стохастического поиска.
Детали реализации и моделирование для рисунка 5.
Моделирование для рисунка 5 было выполнено в NEVESIM, симуляторе на основе событий для сетей пиковых нейронов, разработанном на C ++ с интерфейсом Python [125].Головоломка на рис. 5A была создана и оценена «сложно» в компании «Sudoku Solutions» [126]. Генерация спайков моделируется согласно уравнению (60) с параметрами,. Стохастический порог был установлен для возбуждающих и тормозных нейронов, соответственно. Для пирамидных клеток выбран абсолютный рефрактерный период. Чтобы максимально ускорить моделирование на основе событий, PSP моделировались упрощенным способом как прямоугольные импульсы длины на основе тока (в отличие от более сложной интеграции синаптических входов на основе проводимости, используемой для моделей кортикальных микросхем).
цепей WTA были сформированы путем реципрокного соединения одного ингибирующего нейрона со всеми участвующими пирамидными клетками. Одиночный тормозящий нейрон моделировался таким образом, чтобы имитировать реакцию популяции тормозящих нейронов (то есть сильное торможение в течение длительного периода времени), с использованием абсолютного рефрактерного периода и сильных двунаправленных связей от и к возбуждающим нейронам (синаптические веса и, соответственно, ).
Чтобы задать конкретную головоломку, данные числа были зафиксированы путем подачи сильных входных токов в соответствующие пирамидальные ячейки.В частности, нейроны, кодирующие данные числа в поле судоку, получали постоянный положительный входной ток (постоянный вход на мембранный потенциал). Нейроны, кодирующие конфликтующие цифры в данных полях судоку, получали постоянный отрицательный входной ток силы.
Последнее практическое замечание касается количества нейронов, кодирующих каждую двоичную переменную,. Мы обнаружили, что сети с имеют ряд привлекательных свойств по сравнению с сетями с кодированием одного нейрона. В частности, частота возбуждения отдельных нейронов может быть ниже (пирамидная клетка должна постоянно взрываться, чтобы указывать на устойчивое активное состояние).Кроме того, синаптическая эффективность нейронов может быть ослаблена, и общие паттерны спайк-ответа кажутся более биологически правдоподобными. Ввиду потенциальной реализации в аналоговом нейроморфном оборудовании, присвоения переменных с популяционным кодированием также менее подвержены сбоям отдельных единиц или несоответствию устройств.
Канонические микросхемы для прогнозирующего кодирования
В этом разделе рассматриваются ламинарные соединения, лежащие в основе понятия канонической микросхемы (Douglas et al., 1989; Douglas and Martin, 1991, 2004).Сначала мы сосредоточимся на зрительной коре головного мозга млекопитающих, а затем рассмотрим, можно ли обобщить визуальную микросхему до канонической схемы для всей коры. Как функциональные, так и анатомические методы были применены для изучения внутренних (внутрикортикальных) и внешних связей. Мы подчеркнем выводы недавних исследований, в которых сочетаются оба метода.
Внутренние связи и каноническая микросхема
Основополагающая работа Дугласа и Мартина (1991), посвященная зрительной системе кошки, позволила создать модель того, как информация течет через корковый столбец.Дуглас и Мартин записали внутриклеточные потенциалы клеток первичной зрительной коры во время электростимуляции ее таламических афферентов. Они отметили стереотипный паттерн быстрого возбуждения, за которым следует более медленное и продолжительное торможение. Латентность последующей гиперполяризации позволила выделить ответы в надгранулярном и инфрагранулярном слоях. Используя модели на основе проводимости, они показали, что простая модель может воспроизвести эти реакции. Их модель содержала поверхностные и глубокие пирамидные клетки с общим пулом тормозных клеток.Все три популяции нейронов получили таламический драйв и были полностью взаимосвязаны. Глубокие пирамидные клетки получали относительно слабое таламическое движение, но сильное торможение (). Эти взаимосвязи позволили схеме усилить временные таламические входы для создания устойчивой активности в коре, сохраняя при этом баланс между возбуждением и торможением, две задачи, которые должны быть решены любой корковой цепью. Их схема, хотя и основана на записях зрительной коры головного мозга кошки, также была предложена в качестве основной темы, которая могла бы присутствовать и воспроизводиться с небольшими вариациями по всему корковому листу (Douglas et al., 1989).
Это схема классической микросхемы, адаптированная из работы Дугласа и Мартина (1991). Эта минимальная схема включает поверхностные (слои 2 и 3) и глубокие (слои 5 и 6) пирамидные клетки и популяцию гладких тормозных клеток. Входящие сигналы с прямой связью — от таламуса — нацелены на все клеточные популяции, но с акцентом на тормозные интернейроны, а также на поверхностные и гранулярные слои. Обратите внимание на симметричное развертывание внутренних тормозящих и возбуждающих связей, которые поддерживают баланс возбуждения и торможения.
Последующие исследования использовали внутриклеточные записи и гистологию для измерения спайков (и деполяризации) в пре- и постсинаптических клетках, чья клеточная морфология может быть определена. Этот подход количественно оценивает как вероятность соединения, определяемую как количество наблюдаемых соединений, деленное на общее количество записанных пар, так и силу соединения, определяемую в терминах постсинаптических ответов. Томсон и др. (2002) использовали эти методы для изучения слоев 2–5 (от L2 до L5) зрительных систем кошек и крыс.Наиболее часто соединяющиеся клетки были расположены в одном кортикальном слое, где самые большие межслойные выступы были «прямыми» связями от L4 к L3 и от L3 к L5. Возбуждающие реципрокные связи «обратной связи» не наблюдались (от L3 к L4) или наблюдались реже (от L5 к L3), что позволяет предположить, что возбуждение распространяется внутри столбца с прямой связью. Связи обратной связи обычно наблюдаются, когда пирамидные клетки в одном слое нацелены на ингибирующие клетки в другом (см. Обзор Thomson and Bannister, 2003).
В то время как многие исследования были сосредоточены на возбуждающих связях, некоторые из них изучали тормозные связи. Их труднее изучать, потому что тормозные клетки менее распространены, чем возбуждающие, и потому что существует по крайней мере семь различных морфологических классов (Salin and Bullier, 1995). Однако недавние достижения в оптогенетике сделали возможным более легкое нацеливание на ингибирующие клетки: Kätzel и его коллеги объединили оптогенетику и регистрацию целых клеток, чтобы исследовать внутреннюю связность тормозных клеток в областях коры головного мозга мышей M1, S1 и V1 (Kätzel et al., 2010). Они трансгенно экспрессировали канальный родопсин в тормозных нейронах и активировали их при записи из пирамидных клеток. Это позволило им оценить эффект торможения как функцию ламинарного положения относительно зарегистрированного нейрона.
Из этого подхода можно сделать несколько выводов (Kätzel et al., 2010): во-первых, L4 ингибирующие связи более ограничены в их латеральной протяженности по сравнению с другими слоями. Это подтверждает представление о том, что в ответах L4 преобладают таламические входы, тогда как остальные пластинки интегрируют афференты от более широкого кортикального пятна.Во-вторых, первичный источник ингибирования исходит из клеток в том же слое, что отражает преобладание ингибирующих интраламинарных связей. В-третьих, несколько интерламинарных мотивов оказались общими — по крайней мере, в зернистой коре: главным образом, сильная тормозящая связь от L4 к супрагранулярному L2 / 3 и от инфрагранулярных слоев к L4. Для получения дополнительной информации о специфичности ингибирующих соединений клеточного типа см. Yoshimura and Callaway, (2005). дает сводку ключевых возбуждающих и тормозных интраламинарных связей.
Это упрощенная схема ключевых внутренних связей между возбуждающими (E) и тормозными (I) популяциями в гранулярном (L4), надгранулярном (L1 / 2/3) и инфрагранулярном (L5 / 6) слоях. Возбуждающие межслойные связи в основном основаны на Гилберте и Визеле (1983). Прямые связи обозначают внешние кортикокортикальные или таламокортикальные афференты с прямой связью, которые реципрокны обратными или обратными связями. Анатомические и функциональные данные предполагают, что афферентный вход поступает в первую очередь в L4 и передается в поверхностные слои L2 / 3, богатые пирамидными клетками, которые выступают вперед в следующую корковую область, образуя дисинаптический путь между таламусом и вторичными кортикальными областями (Callaway, 1998).Информация из L2 / 3 затем отправляется в L5 и L6, которые отправляют (внутренние) проекции обратной связи обратно в L4 (Usrey and Fitzpatrick, 1996). Клетки L5 образуют обратную связь с более ранними областями коры, а также с легковыми, верхними бугорками и стволом головного мозга. Таким образом, прямой ввод разделяется внутренними связями на поверхностный прямой поток и глубокий обратный поток. На этой схеме мы сопоставили плотно взаимосвязанные возбуждающие и тормозящие популяции внутри каждого слоя.
Микросхемы в сенсомоторной коре
Распространяются ли особенности зрительных микросхем на другие области коры? Недавно в двух исследованиях была выявлена внутренняя взаимосвязь сенсорной и моторной коры мышей: Лефорт и его коллеги (2009) использовали множественные записи целых клеток в коре ствола мышей, чтобы определить вероятность моносинаптических связей и соответствующую силу связи. Как и в зрительной коре, самые сильные связи были интраламинарными, а самые сильные межслойные связи — восходящими L4 к L2 и нисходящими L3 к L5.
Одна из загадок канонических микросхем заключается в том, имеет ли моторная кора локальная схема, качественно похожая на сенсорную кору. Этот вопрос важен, потому что моторная кора не имеет четко определенного гранулярного L4 (свойство, за которое она получила название «агранулярная кора»). Weiler и его коллеги объединили записи целых клеток в моторной коре мышей с фотостимуляцией для извлечения глутамата из клетки (Weiler et al., 2008). Это позволило им систематически стимулировать корковый столбец в сетке с центром на пирамидном нейроне, с которого они регистрировались.Путем записи от пирамидных нейронов в L2-6 (L1 лишен пирамидных клеток) авторы картировали возбуждающее влияние, которое каждый слой оказывает на другие. Они обнаружили, что соединение L2 / 3 с L5A / B было самым сильным — на него приходилось треть общего синаптического тока в цепи. Вторым по силе межслоевым соединением было реципрокное соединение L5A с L2 / 3. Этот путь может быть гомологичен известному пути от L4 / 5A к L2 / 3 в сенсорной коре. Также, как и в сенсорной коре, были заметны рекуррентные (интраламинарные) связи, особенно в L2, L5A / B и L6.Наибольшая часть синаптического ввода поступает в L5A / B, что соответствует его ключевой роли в накоплении информации от широкого диапазона афферентов — перед отправкой его вывода в кортикоспинальный тракт. Таким образом, сильная связь от входного слоя к поверхностной и от поверхностной к глубокой, вместе с сильной интраламинарной связностью, предполагает, что внутренняя схема моторной коры похожа на другие области коры.
Анатомия и физиология внешних связей
Ясно, что учет микросхем должен относиться к слоям происхождения внешних связей и их ламинарным мишеням.Хотя большинство пресинаптических входов возникает из внутренних связей, области коры также сильно взаимосвязаны, где баланс между внутренней и внешней обработкой опосредует функциональную интеграцию между специализированными областями коры (Engel et al., 2010). Только по цифрам внутренние связи, по-видимому, преобладают — 95% всех нейронов, помеченных ретроградным индикатором, находятся в пределах примерно 2 мм от места инъекции (Markov et al., 2011). Остальные 5% представляют собой клетки, дающие начало внешним связям, которые, хотя и немногочисленны, могут быть чрезвычайно эффективными в управлении своими целями.Речь идет о связи LGN с V1: хотя это только шестая по силе связь с V1, афференты LGN оказывают существенное влияние на ответы V1 (Markov et al., 2011).
Иерархии и функциональные асимметрии
Современная догма утверждает, что кора головного мозга иерархически организована. Идея корковой иерархии опирается на различие между тремя типами внешних связей: прямые связи, которые связывают более раннюю область с более высокой областью, связи обратной связи, которые связывают более высокую область с более ранней областью, и боковые связи, которые связывают области на более высоком уровне. на том же уровне (см. обзор Felleman and Van Essen, 1991).Эти связи различаются по своему ламинарному происхождению и целям. Прямые связи происходят в основном от поверхностных пирамидных клеток и мишени L4, тогда как обратные связи происходят в основном от глубоких пирамидных клеток и заканчиваются вне L4 (Felleman and van Essen 1991). Ясно, что это описание корковых иерархий является упрощением и может иметь множество нюансов: например, по мере увеличения иерархического расстояния между двумя областями процент клеток, отправляющих прямую связь (соотв.обратная связь) проекции с более низкого (соответственно более высокого) уровня становятся все более смещенными в сторону поверхностных (соответственно глубоких) слоев (Barone et al., 2000; Vezoli, 2004).
В дополнение к ламинарной специфичности их происхождения и мишеней, прямые и обратные связи также различаются по своей синаптической физиологии. Традиционная точка зрения утверждает, что прямые связи являются сильными и движущими, способными вызывать пиковую активность у их целей и придавать классические свойства рецептивного поля — прототипическим примером является синаптическая связь между LGN и V1 (Sherman and Guillery, 1998).Считается, что соединения обратной связи модулируют (экстра-классические) характеристики рецептивного поля в соответствии с текущим контекстом; например, визуальная окклюзия, внимание, заметность, и т. д. Прототипным примером обратной связи является соединение кортикального слоя L6 с LGN. Шерман и Гиллери определили несколько свойств, которые отличают драйверы от модуляторов . Управляющие связи, как правило, демонстрируют сильный ионотропный компонент в синаптическом ответе, вызывают большие ВПСП и отвечают на несколько ВПСП угнетающими синаптическими эффектами.Модуляторные связи при стимуляции вызывают метаботропные и ионотропные реакции, вызывают слабые ВПСП и демонстрируют облегчение парных импульсов (Sherman and Guillery, 1998; 2011). Эти различия были основаны на входах в LGN, где входные данные сетчатки являются движущими, а входные сигналы коры являются модулирующими. До недавнего времени было доступно мало данных, чтобы оценить, применимо ли подобное различие к кортико-кортикальным связям прямой связи и обратной связи. Однако недавние исследования показывают, что корковые обратные связи выражают не только модуляторные, но и управляющие характеристики:
Являются ли обратные связи управляющими, модуляционными или и тем и другим?
Хотя обычно считается, что обратные связи являются слабыми и модулирующими (Crick and Koch, 1998; Sherman and Guillery, 1998), недавние данные свидетельствуют о том, что обратные связи не только модулируют ответы более низкого уровня: Шерман и его коллеги зарегистрировали клетки в области мыши. V1 / V2 и A1 / A2, стимулируя афференты прямой или обратной связи.В обоих случаях наблюдались реакции, похожие на вождение, а также на модуляторные реакции (Covic and Sherman, 2011; De Pasquale and Sherman, 2011). Это указывает на то, что — для этих иерархически близких областей — обратные связи могут управлять своими целями так же сильно, как и прямые связи. Это согласуется с более ранними исследованиями, показывающими, что обратная связь может быть движущей силой: Миньяр и Малпели (1991) изучали обратную связь между областями 18 и 17, в то время как слой А LGN был фармакологически инактивирован.Это заставило замолчать клетки L4 в области 17, но сохранило активность в поверхностных слоях. Однако при поражении области 18 поверхностные клетки заглушались. Это согласуется с движущим эффектом обратной связи из области 18 при отсутствии коленчатого входа. Таким образом, обратная связь может опосредовать модулирующие и управляющие эффекты. Это важно с точки зрения кодирования с предсказанием, потому что нисходящие предсказания должны вызывать обязательные ответы в своих целях (ячейки, сообщающие об ошибках предсказания):
При кодировании с предсказанием соединения с прямой связью передают ошибки предсказания, а соединения обратной связи передают предсказания от верхние области коры, чтобы подавить ошибки предсказания в нижних областях.Таким образом, в этой схеме соединения обратной связи должны иметь возможность оказывать сильное (управляющее) влияние на более ранние области для подавления или противодействия управляющим входам с прямой связью. Однако, как мы увидим позже, эти влияния также должны вызывать нелинейные или модулирующие эффекты. Это связано с тем, что предсказания сверху вниз обязательно зависят от контекста: например, перекрытие одного визуального объекта другим. Короче говоря, для прогнозирующего кодирования требуются обратные связи, чтобы управлять клетками на более низких уровнях контекстно-зависимым образом, что требует модулирующего аспекта их постсинаптических эффектов.
Являются ли обратные связи возбуждающими или тормозящими?
Важно отметить, что поскольку обратные связи передают прогнозы — которые служат для объяснения и тем самым уменьшения ошибок прогнозирования на более низких уровнях — их эффективная (полисинаптическая) связность обычно считается тормозящей. Общий ингибирующий эффект обратных связей согласуется с исследованиями in vivo . Например, электрофизиологические исследования негативности несоответствия показывают, что нейронные реакции на девиантные стимулы, которые нарушают сенсорные предсказания, установленные регулярной последовательностью стимулов, усиливаются по сравнению с предсказанными стимулами (Garrido et al., 2009). Сходным образом нарушение ожиданий слухового повторения вызывает усиление гамма-диапазона ответов в ранней слуховой коре (Todorovic et al., 2011). Считается, что эти усиленные ответы отражают неспособность высших областей коры головного мозга предсказывать и тем самым подавлять активность популяций, кодирующих ошибку предсказания (Garrido et al., 2007; Wacongne et al., 2011). Подавление предсказуемых ответов также можно рассматривать как подавление повторения, наблюдаемое в единичных записях из нижней височной коры макак (Desimone, 1996).Более того, нейроны нижневисочной коры обезьян значительно меньше реагируют на предсказанную последовательность естественных изображений по сравнению с непредсказуемой последовательностью (Meyer and Olson, 2011).
Ингибирующий эффект обратных связей дополнительно подтверждается исследованиями нейровизуализации (Murray et al., 2002; Murray, 2005; Harrison et al., 2007; Summerfield et al., 2008, 2011; Alink et al., 2010). Эти исследования показывают, что предсказуемые стимулы вызывают меньшую реакцию в ранних корковых областях. Важно отметить, что это подавление не может быть объяснено с точки зрения местной адаптации, потому что атрибуты стимулов, которые можно предсказать, не представлены в ранней сенсорной коре (например,г., Харрисон и др. 2007). Следует отметить, что подавление ответов на предсказуемые стимулы может сосуществовать с усилением внимания (сверху вниз) обработки сигналов (Wyart et al., 2012): при прогнозирующем кодировании внимание опосредуется увеличением прироста популяций, кодирующих ошибку прогнозирования. (Спратлинг, 2008; Фельдман, Фристон, 2010). Результирующая модуляция внимания (например, Hopfinger et al., 2000) может взаимодействовать с предсказаниями сверху вниз, чтобы преодолеть их подавляющее влияние — как показано эмпирически (Kok et al., 2011). См. Buschman and Miller (2007), Saalmann et al. (2007), Anderson et al. (2011) и Armstrong et al. (2012) для дальнейшего обсуждения нисходящих связей во внимании.
Дальнейшие доказательства тормозящего (подавляющего) эффекта обратных связей получены из нейропсихологии: пациенты с повреждением префронтальной коры (ПФК) демонстрируют растормаживание связанных с событием потенциальных ответов (ERP) на повторяющиеся стимулы (Knight et al., 1989; Yamaguchi). и Knight, 1990; но см. Barceló et al., 2000). Напротив, они демонстрируют ERP P300 с уменьшенной амплитудой в ответ на новые стимулы — как если бы произошла неспособность передать нисходящие предсказания сенсорной коре головного мозга (Knight, 1984). Кроме того, нормальные субъекты демонстрируют быструю адаптацию к девиантным стимулам, когда они становятся предсказуемыми — эффект, не наблюдаемый у префронтальных пациентов.
Несколько инвазивных исследований дополняют эти исследования на людях, предполагая общую ингибирующую роль обратных связей. В недавнем плодотворном исследовании Olsen et al.изучили кортикоталамическую обратную связь между L6 V1 и LGN, используя трансгенную экспрессию канала родопсина в L6 клетках V1. Управляя этими клетками оптогенетически — во время регистрации единиц в V1 и LGN — авторы показали, что глубокие основные клетки L6 ингибируют свои внешние мишени в LGN и свои внутренние мишени в корковых слоях 2-5 (Olsen et al., 2012). Это подавление было мощным — в LGN зрительные реакции подавлялись на 76%. Подавление также было высоким в V1, около 80-84% (Olsen et al., 2012). Эти данные согласуются с классическими исследованиями вклада кортикогенных веществ в настройку длины в LGN, показывающими, что корковая обратная связь способствует подавлению объемного звука кошачьих LGN-клеток: без обратной связи клетки LGN растормаживаются и демонстрируют более слабое объемное подавление (Murphy and Sillito, 1987). ; Sillito et al., 1993; но см. Alitto and Usrey, 2008).
В то время как эти исследования предоставляют убедительные доказательства того, что корковая обратная связь с LGN является тормозящей, доказательства более сложны для кортикокортикальных обратных связей (Sandell and Schiller, 1982; Johnson and Burkhalter, 1996, 1997).Хюпе и его коллеги охлаждали область V5 / MT во время записи из областей V1, V2 и V3 у обезьяны. Когда зрительные стимулы предъявлялись в классическом рецептивном поле (CRF), охлаждение области V5 / MT снижало активность единиц в более ранних областях, предполагая возбуждающий эффект внешней обратной связи (Hupé et al., 1998). Однако, когда авторы использовали стимул, охватывающий внеклассическую радиочастоту, ответы нейронов V1 были — в среднем — усилены после охлаждения области V5, что согласуется с подавляющей ролью обратных связей.Эти результаты показывают, что тормозящие эффекты обратной связи могут зависеть от (естественных) стимулов, которые требуют интеграции в поле зрения. Аналогичные эффекты наблюдались, когда область V2 охлаждалась, а нейроны измерялись в V1: когда стимулы подавались только на CRF, охлаждение V2 уменьшало активность пиков V1; однако, когда стимулы присутствовали в CRF и окружающей среде, охлаждение V2 увеличивало активность V1 (Bullier et al., 1996). Наконец, другие утверждали, что ингибирующий эффект обратной связи основан на времени и пространственной степени подавления окружающего звука в обезьяне V1, заключая, что эффекты подавления дальнего окружения, скорее всего, опосредованы обратной связью (Bair et al., 2003).
Эмпирический вывод о том, что обратная связь может как облегчить, так и подавить возбуждение в нижних иерархических областях — в зависимости от содержания классических и внеклассических рецептивных полей — согласуется с прогнозирующим кодированием: Рао и Баллард (1999) обучили сеть иерархического прогнозирующего кодирования распознавать естественные образы. Они показали, что более высокие уровни иерархии учатся предсказывать визуальные особенности, которые распространяются на многие CRF на более низких уровнях (например, стволы деревьев или горизонты).Следовательно, более высокие визуальные области предсказывают, что визуальные стимулы будут охватывать рецептивные поля клеток в более низких визуальных областях. В этой настройке стимул, который ограничен CRF, вызовет сильный сигнал ошибки предсказания (потому что он не может быть предсказан). Это дает простое объяснение выводам Hupé et al (1998) и Bullier et al (1996): когда обратная связь деактивирована, нет никаких нисходящих предсказаний для объяснения ответов в нижних областях, что приводит к растормаживанию ответов в нижних областях. более ранние области, когда — и только когда — стимулы можно предсказать по множеству CRF.
Обратные связи и уровень 1
Как может быть опосредован тормозящий эффект обратных связей? Установлено мнение, что внешние кортикокортикальные связи являются исключительно возбуждающими (с использованием глутамата в качестве возбуждающего нейромедиатора), хотя недавние данные свидетельствуют о том, что ингибирующие внешние связи существуют и могут играть важную роль в синхронизации удаленных регионов (Melzer et al., 2012). Однако один важный путь, с помощью которого обратные связи могут опосредовать избирательное ингибирование, — это их прекращение в L1 (Anderson and Martin, 2006; Shipp, 2007): слой 1 иногда называют бесклеточным из-за его бледного вида при окрашивании по Нисслю (классическое метод разделения слоев, выборочно маркирующий тела клеток).Действительно, недавнее исследование пришло к выводу, что L1 содержит менее 0,5% всех клеток в кортикальном столбе (Meyer et al., 2011). Эти клетки L1 почти все являются тормозящими и прочно связаны друг с другом посредством электрических соединений и химических синапсов (Chu et al., 2003). Одновременные записи патч-зажимов целых клеток показывают, что они обеспечивают сильное моносинаптическое ингибирование пирамидных клеток L2 / 3, чьи апикальные дендриты проецируются в L1 (Chu et al., 2003; Wozny and Williams, 2011). Это означает, что клетки, ингибирующие L1, находятся в лучшем положении, чтобы опосредовать ингибирующие эффекты внешней обратной связи.Ламинарное расположение, выделенное этими исследованиями — нижняя часть L1 и верхняя часть L2 / 3 — недавно было показано, как «горячая точка» ингибирования в колонке (Meyer et al., 2011). Действительно, исследование бочки коры головного мозга крысы, которая стимулировала (и инактивировала) L1, показало, что она оказывает мощный тормозящий эффект на ответы, вызванные усами (Shlosberg et al., 2006). Эти исследования показывают, что кортикокортикальные обратные связи могут оказывать сильное ингибирование, если они задействуют ингибирующий потенциал L1.
Что касается возбуждающего и модулирующего эффекта обратных связей, предсказательный ввод из более высоких областей коры может иметь важное влияние через дистальные дендриты пирамидных нейронов (Larkum et al., 2009). Более того, существует определенный тип ГАМКергических нейронов, которые, по-видимому, контролируют дистальную возбудимость дендритов, по-разному управляя возбуждающими сигналами сверху вниз во время поведения (Gentet et al., 2012). резюмирует исследования, которые мы обсудили в отношении роли обратной связи.
Таблица 1
Результаты электрофизиологических исследований и нейровизуализации, согласующиеся с прогностическим кодированием.
| Прогноз нарушен | Изученная область | Нейрональная экспрессия ошибки предсказания | Исследование |
|---|---|---|---|
| Пары изученных визуальных объектов | Нижне-височная кора головного мозга обезьяны (ИТ) | Повышенная частота возбуждения | Мейер и Олсон, 2011 г. |
| Статистика естественного изображения | Monkey V1, V2, V3 | Повышенная скорость стрельбы | Hupé et al., 1998; Bullier et al., 1996; Bair et al., 2003 |
| Повторяющийся слуховой поток | Ранняя слуховая кора человека | Повышенные потенциалы, связанные с событиями (ERP), повышенная мощность в гамма-диапазоне | Garrido et al., 2007, 2009; Тодорович и др., 2011 |
| Связность визуальной формы и движения | Человек V1, V2, V3, V4, V5 / MT | Усиленный ответ BOLD | Murray et al 2002; Мюррей и др. 2005; Харрисон и др., 2007 |
| Аудиовизуальное соответствие речи | Зрительная и слуховая кора | Осцилляторная активность в гамма-диапазоне | Арнал и др., 2011 г. |
| Предсказуемость зрительных стимулов в зависимости от внимания | Human V1, V2, V3 | Усиленный ЖЕЛТЫЙ ответ при отсутствии присмотра, уменьшенный ЖЕЛТЫЙ при посещении | Кок и др., 2011 |
| Иерархические ожидания в слуховых последовательностях | Височная кора человека | Расширенные потенциалы, связанные с событиями (ERP) | Wacongne et al., 2011 г. |
| Ожидаемое повторение (или чередование) раздражителей лица | FFA в фМРТ, париетальном и центральном электродах ЭЭГ | Усиленный BOLD-ответ, уменьшенное подавление повторения ERP | Summerfield et al., 2008, 2011 |
| Видимое движение зрительного стимула | V1 | Расширенный ответ жирным шрифтом | Алинк и др., 2010 г. |
Прямые и трансталамические связи
Хотя доказательства ингибирующего эффекта обратных связей необходимо тщательно оценивать, доказательства возбуждающего эффекта прямых прямых связей однозначны.Например, у обезьяны V1 моносинаптически проецируется на V2, V3, V3a, V4 и V5 / MT (Zeki, 1978; Zeki and Shipp, 1988). Во всех случаях — когда V1 обратимо инактивируется посредством охлаждения — активность отдельных клеток в целевых областях сильно подавляется (Girard and Bullier, 1989; Girard et al., 1991a, 1991b, 1992). В случаях V2 и V3 результатом охлаждения области V1 является почти полное подавление активности отдельной единицы. Эти исследования показывают, что активность в высших областях коры зависит от входных сигналов от более ранних областей коры, которые устанавливают свойства их рецептивного поля.
Наконец, хотя многие исследования были сосредоточены на внешних связях, которые проецируются непосредственно из одной области коры в другую, появляется все больше свидетельств того, что управляющие связи с прямой связью (и, возможно, обратная связь) в коре могут быть опосредованы трансталамическими путями (Sherman and Guillery, 1998, 2011). Наиболее убедительное доказательство этого утверждения исходит от соматосенсорной системы, где недавно было показано, что заднее медиальное ядро таламуса (POm) — таламическое ядро более высокого порядка, которое получает прямой вход от коры головного мозга — может передавать информацию между S1 и S2 ( Хил и др., 2009). Кроме того, таламическое ретикулярное ядро, как предполагается, опосредует торможение, которое может лежать в основе кросс-модального внимания или предсказаний сверху вниз (Yamaguchi and Knight, 1990; Crick, 1984; Wurtz et al., 2011). Более того, вычислительные соображения и недавние экспериментальные данные указывают на потенциально важную роль таламических ядер более высокого порядка в координации и синхронизации корковых ответов (Vicente et al., 2008; Saalmann et al., 2012). Степень, в которой корковые области интегрированы напрямую через кортикокортикальные или косвенно через кортико-таламо-корковые связи, и степень, в которой трансталамические пути отделяют прямую связь от обратных связей таким же образом, как мы предложили для кортико-кортикальных связей, открыты. вопросов.
