Как происходит транскрипция генетической информации. Какие основные этапы включает процесс синтеза РНК на матрице ДНК. Чем отличается транскрипция у прокариот и эукариот.
Что такое транскрипция и зачем она нужна
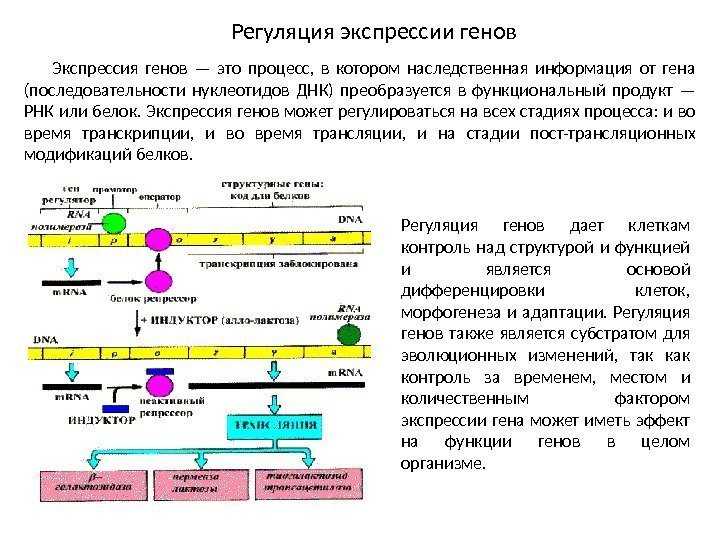
Транскрипция — это процесс синтеза молекулы РНК на матрице ДНК. Она является первым этапом реализации генетической информации и необходима для:
- Синтеза матричных РНК (мРНК), которые служат матрицей для синтеза белков
- Образования транспортных РНК (тРНК), участвующих в трансляции
- Синтеза рибосомных РНК (рРНК), входящих в состав рибосом
- Образования малых ядерных РНК (мяРНК), участвующих в сплайсинге
- Синтеза микроРНК и других некодирующих РНК с регуляторными функциями
Таким образом, транскрипция обеспечивает передачу генетической информации от ДНК к РНК и является ключевым этапом экспрессии генов.
Основные этапы транскрипции
Процесс транскрипции можно разделить на три основных этапа:
1. Инициация
На этом этапе происходит:
- Узнавание промоторной последовательности РНК-полимеразой
- Связывание РНК-полимеразы с промотором
- Локальное расплетение двойной спирали ДНК
- Образование открытого комплекса
2. Элонгация
Основные события элонгации:
- Продвижение РНК-полимеразы вдоль матрицы ДНК
- Синтез комплементарной цепи РНК в направлении 5′ → 3′
- Образование и перемещение «транскрипционного пузырька»
- Временное разделение цепей ДНК
3. Терминация
На этом этапе происходит:
- Узнавание сигнала терминации транскрипции
- Остановка синтеза РНК
- Высвобождение синтезированной молекулы РНК
- Диссоциация РНК-полимеразы от матрицы ДНК
Строение и функции РНК-полимеразы
РНК-полимераза — ключевой фермент транскрипции. Она обладает следующими свойствами:
- Катализирует образование фосфодиэфирных связей между нуклеотидами РНК
- Обладает 5′ → 3′ направленностью синтеза
- Способна к коррекции ошибок
- Имеет ДНК-зависимую активность
- Не требует праймера для начала синтеза
У прокариот существует один тип РНК-полимеразы, а у эукариот — три (РНК-полимеразы I, II и III).
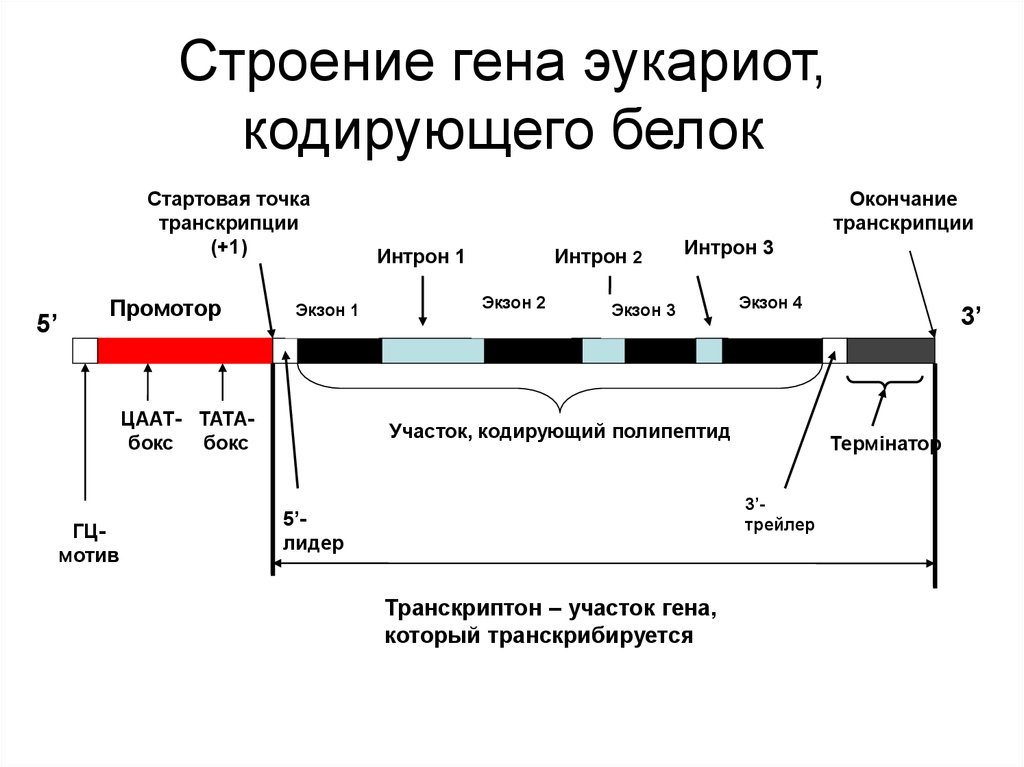
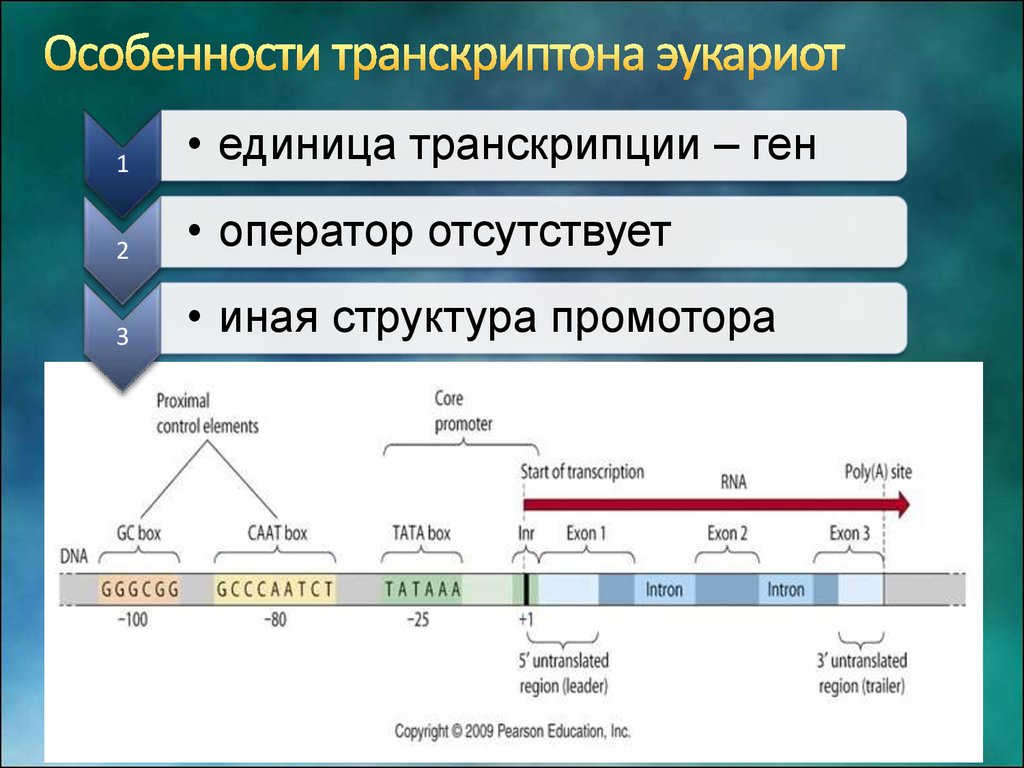
Особенности промоторов прокариот и эукариот
Промоторы — это специфические последовательности ДНК, с которых начинается транскрипция. Они имеют ряд отличий у прокариот и эукариот:
Промоторы прокариот:
- Расположены непосредственно перед геном
- Имеют консенсусные последовательности в положениях -10 и -35
- Узнаются σ-фактором РНК-полимеразы
- Относительно просто устроены
Промоторы эукариот:
- Могут располагаться на значительном расстоянии от гена
- Содержат ТАТА-бокс и другие регуляторные элементы
- Имеют сложное строение
Регуляция транскрипции у прокариот
Регуляция транскрипции у прокариот осуществляется несколькими способами:
- С помощью белков-репрессоров и активаторов
- Путем аттенюации транскрипции
- За счет изменения структуры хроматина
- Через альтернативные σ-факторы
- С участием малых регуляторных РНК
Ключевую роль играют оперонные системы, позволяющие координированно регулировать экспрессию нескольких генов.
Особенности регуляции транскрипции у эукариот
Регуляция транскрипции у эукариот более сложная и включает:
- Эпигенетические механизмы (метилирование ДНК, модификации гистонов)
- Ремоделирование хроматина
- Взаимодействие с энхансерами и сайленсерами
- Регуляцию на уровне инициации, элонгации и терминации
- Посттранскрипционные механизмы (альтернативный сплайсинг, редактирование РНК)
Важную роль играют транскрипционные факторы, связывающиеся с регуляторными элементами генов.
Посттранскрипционные модификации РНК
После синтеза молекулы РНК подвергаются ряду модификаций:
У прокариот:
- Процессинг рРНК и тРНК
- Модификация отдельных нуклеотидов
У эукариот:
- Кэпирование 5′-конца мРНК
- Полиаденилирование 3′-конца мРНК
- Сплайсинг пре-мРНК
- Редактирование РНК
Эти модификации необходимы для созревания и стабилизации молекул РНК.
Как транскрипция связана с другими клеточными процессами
Транскрипция тесно связана с другими важными процессами в клетке:
- Репликацией ДНК (координация экспрессии генов с клеточным циклом)
- Репарацией ДНК (транскрипционно-сцепленная репарация)
- Трансляцией (сопряжение транскрипции и трансляции у прокариот)
- Сплайсингом (котранскрипционный сплайсинг у эукариот)
- Транспортом РНК из ядра в цитоплазму

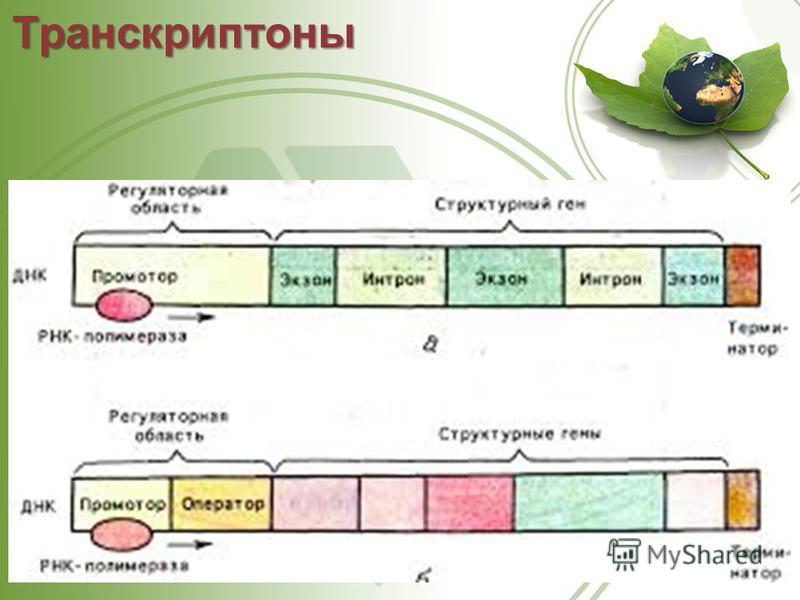
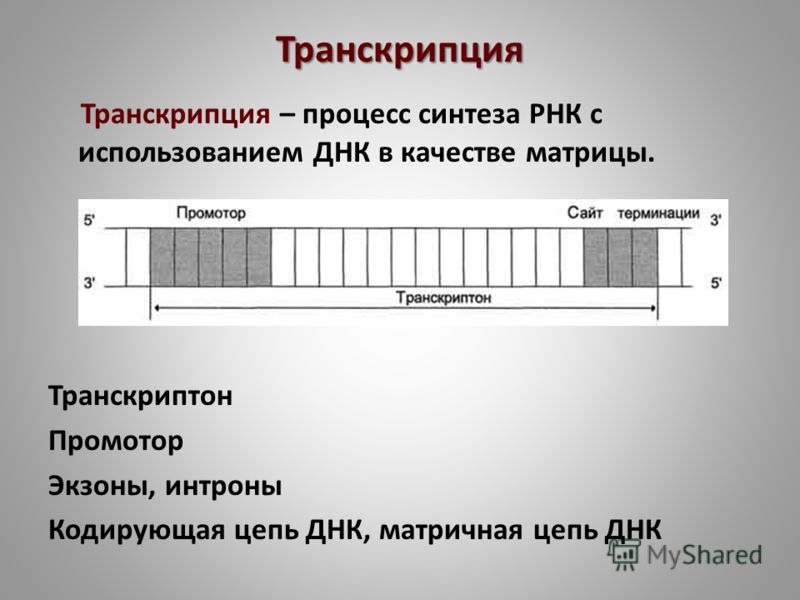
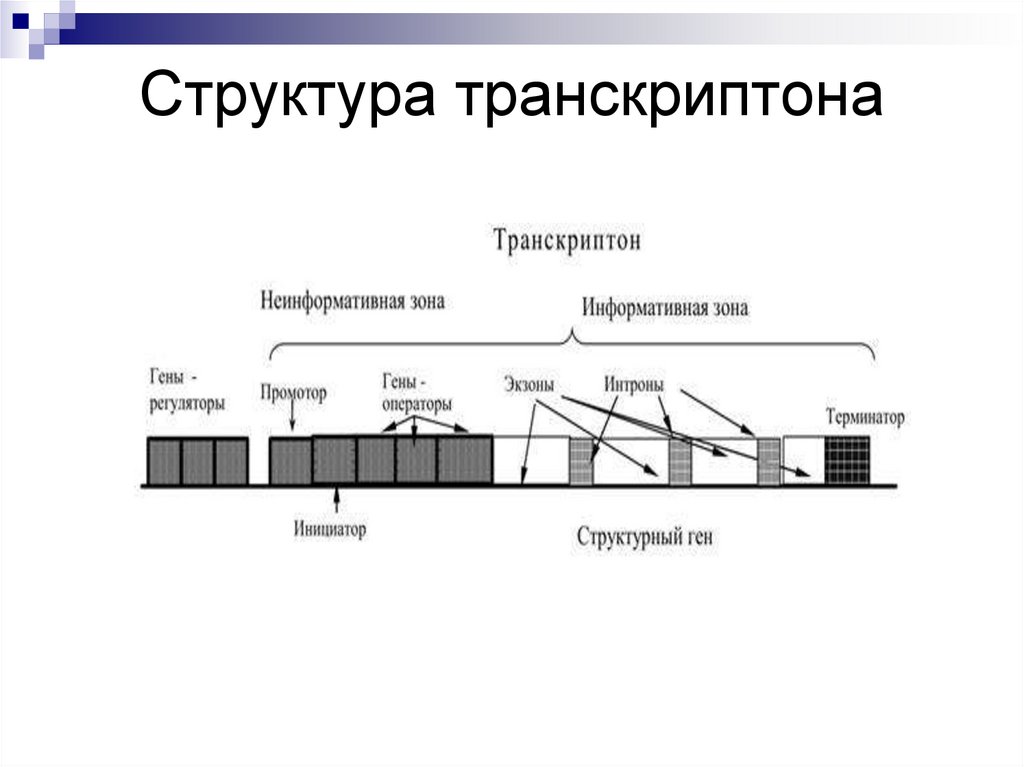
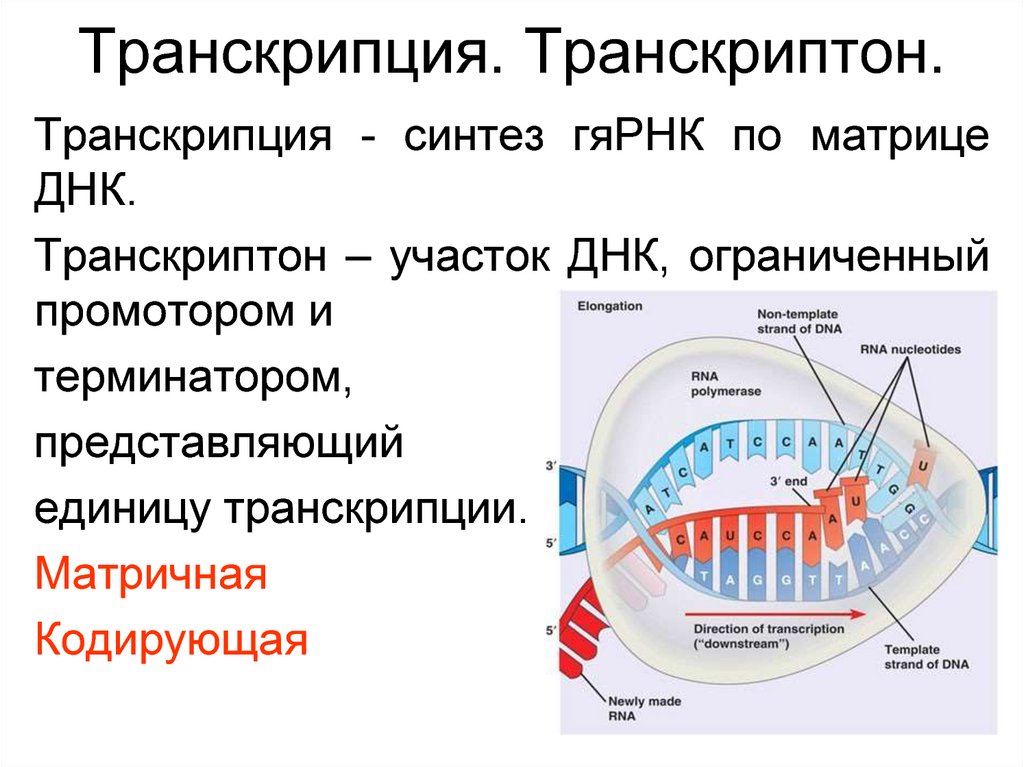
Транскриптон (оперон) — единица транскрипции.
Синтез молекул РНК начинается в определенных местах ДНК, называемых промоторами и завершается в терминаторах. Участок ДНК, ограниченный промотором и терминатором, представляет собой единицу транскрипции — транскриптон (оперон у прокариот). В пределах каждого транскриптона копируется только одна из двух нитей ДНК, которая называется значащей или матричной. Во всех транскриптонах, считываемых в одном направлении, значащей является одна нить ДНК, в транскриптонах, считываемых в противоположном направлении, значащей является другая нить ДНК.
Разбиение ДНК на множество транскриптонов обеспечивает возможность независимого считывания разных генов, их индивидуального включения и выключения.
2
коротких консервативных участка
последовательностей, расположенных на
расстоянии 35 нуклеотидных пар в сторону
5’ конца от начала транскрипции (область
-30 на рис.) и на расстоянии 10 нуклеотидов
в ту же сторону (область -10). АТ богатый
участок области -10 получил название
ТАТА бокса или Прибнов бокса. Считается,
что эта последовательность может быть
легко денатурирована РНК-полимеразой
и является удобным местом начала
транскрипции.
АТ богатый
участок области -10 получил название
ТАТА бокса или Прибнов бокса. Считается,
что эта последовательность может быть
легко денатурирована РНК-полимеразой
и является удобным местом начала
транскрипции.
Первый шаг в транскрипции — связывание РНК-полимеразы с ДНК, с последующим перемещением к инициирующему участку ДНК. Поиск промотора РНК-полимеразой начинается (на рисунке этап 1) с неспецифического связывания фермента с ДНК, и последующего перемещения по ДНК, без диссоциации до достижения последовательности промотора, с которым фермент связывается со значительно более высоким сродством. Этому способствует
Первоначальное взаимодействие РНК-полимеразы и промотора образует закрытый комплекс, цепь ДНК в этом комплексе не раскручена.
РНК-полимераза
раскручивает несколько пар оснований
ДНК, от приблизительно -10 до -1, образуя
открытый промоторный
комплекс, названный
так потому, что в нем цепи ДНК открыты,
или раскручены, фермент готов начать
синтез цепи РНК.
После синтеза первых 10 нуклеотидов, субъединица отделяется от комплекса транскрипции, и дальнейший процесс транскрипции катализируется базовой (коровой) полимеразой (см. рисунок, шаги 5 и 6). Хотя и субъединица покинула фермент, элонгирующий комплекс является довольно устойчивым. Элонгация, таким образом, продолжается до заключительного этапа.
Во время элонгации (см. рисунок выше, шаги 5 и 6), коровый феремент перемещается по двуцепочечной матрице ДНК, одновременно раскручивает ДНК, копируя одноцепочечную матрицу ДНК с образованием транскрипта (вновь синтезируемая РНК). Фермент скручивает матрицу позади 3 ‘ конца растущей цепи РНК.
В областях окончания транскрипции происходит диссоциации РНК-полимеразы и ДНК.
31Процессинг и сплайсинг рнк Кэпирование и полиаденилирование иРнк называется процессингом (посттранскрип-ционной модификацией).
Кэпирование:
К 5 ‘ концу всех эукариотических
иРНК присоединяется во время процессинга
остаток 7-метилгуанозина
с образованием уникальной
5 ‘
5 ‘ фосфодиэфирной связи.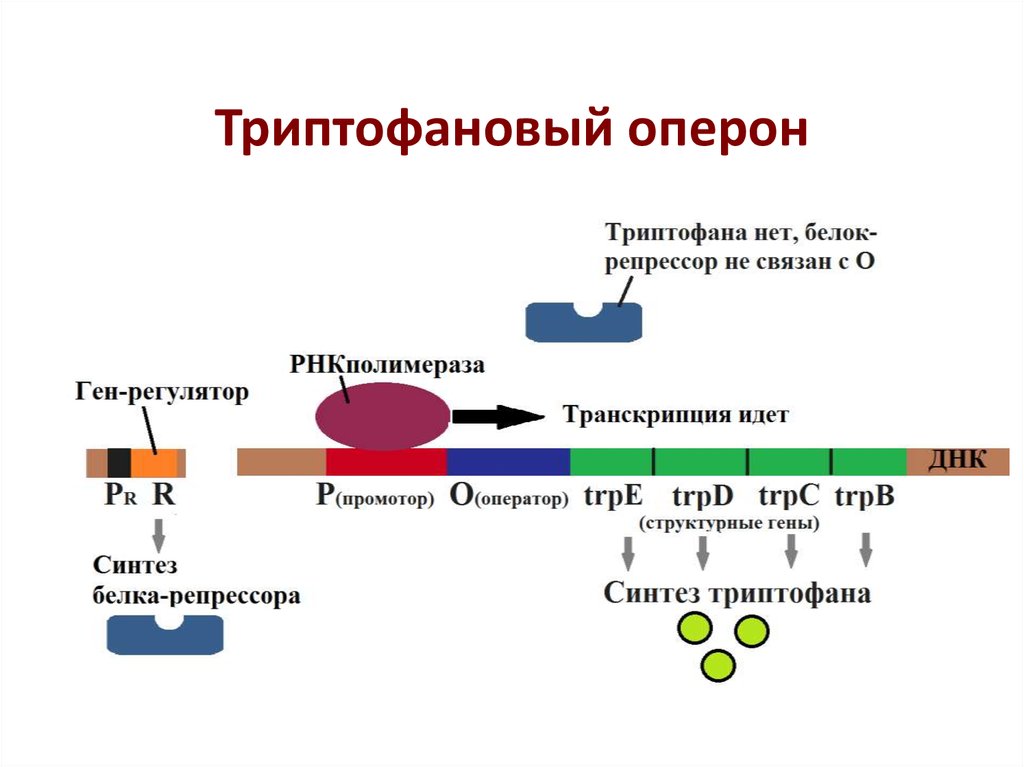 Этот дополнительный нуклеотид получил
название кэп
или колпачек.
Этот дополнительный нуклеотид получил
название кэп
или колпачек.
Функции кэпа:
он защищает РНК от экзонуклеаз
помогает связыванию молекулы мРНК с рибосомой.
Полиаденилирование:
3′-конец также модифицируется сразу после завершения транскрипции. Специальный фермент – полиаденилат-полимераза присоединяет к 3′-концу каждого РНК-транскрипта от 20 до 250 остатков адениловой кислоты (поли(А)). Полиаденилатполимераза узнает специфическую последовательность AAУAAA, отщепляет от первичного транскрипта небольшой фрагмент в 11-30 нуклеотидов и затем присоединяет поли(А) последовательность. Принято считать, что такой «хвост» способствует последующему процессингу РНК и экспорту зрелых молекул мРНК из ядра.
По мере участия иРНК в
процессах трансляции, длина полиА
фрагмента уменьшается.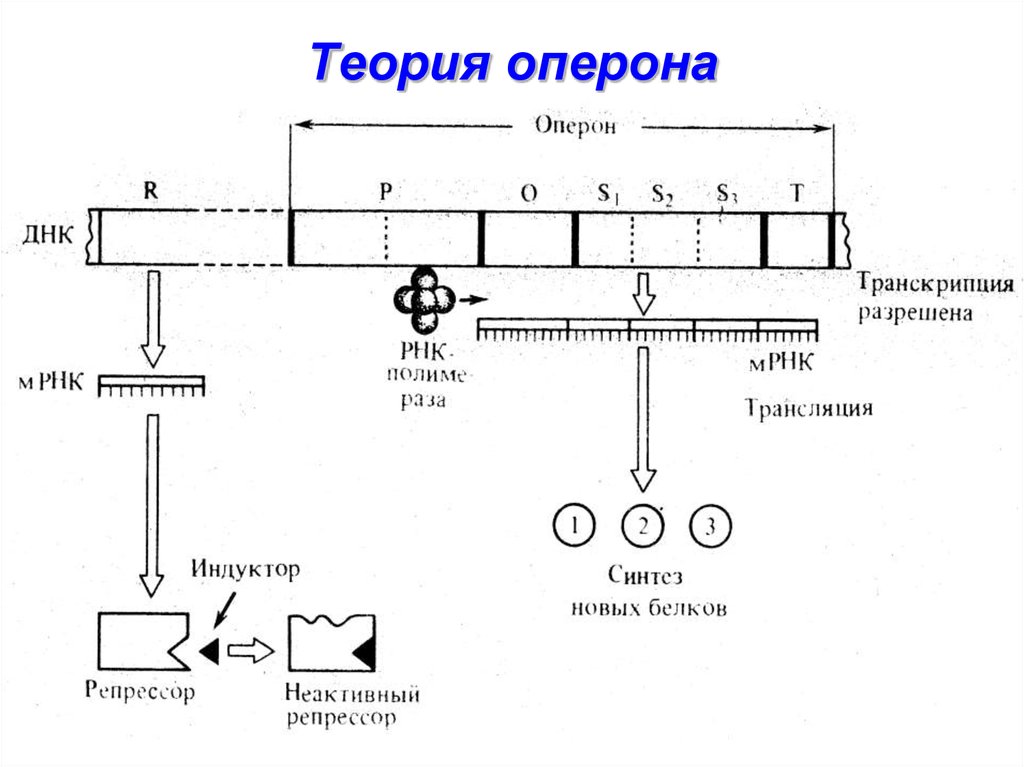 Критическим для
стабильности считается 30 адениловых
нуклеотидов.
Критическим для
стабильности считается 30 адениловых
нуклеотидов.
Вся совокупность ядерных транскриптов РНК-полимеразы II известна как гетерогенная ядерная РНК (гяРНК).
Все 3 класса РНК транскрибируются с генов, которые содержат интроны (неинформативные участки) и экзоны (участки ДНК, несущие информацию). Последовательности, кодируемые интронами ДНК, должны быть удалены из первичного транскрипта до того, как РНК станет биологически активной. Процесс удаления копий интронных последовательностей получил название сплайсинга РНК.
Сплайсинг РНК катализируется комплексами белков с РНК, известными как «малые ядерные рибонуклеопротеидные частицы» (мяРНП, англ. small nuclear ribonucleic particles, snRNP).Такие каталитические РНК носят название рибозимов.
Функции интронов:
защищают функционально активную часть генома клетки от повреждающего действия химических или физических (лучевых) факторов
позволяет при помощи так называемого альтернативного сплайсинга увеличить генетическое разнообразие генома без увеличения числа генов.

Альтернативный сплайсинг:
В результате изменения распределение экзонов одного транскрипта во время сплайсинга возникают различные РНК и следовательно различные белки.
Известны уже более 40 генов, транскрипты которых подвергаются альтернативному сплайсингу. Например, транскрипт гена кальцитонина, в результате альтернативного сплайсинга дает РНК, которая служит матрицей для синтеза кальцитонина (в щитовидной железе) или специфического белка, отвечающего за вкусовое восприятие (в мозге). Еще более сложному альтернативному сплайсингу подвергается транскрипт гена -тропомиозина. Были идентифицированы по крайней мере 8 различных тропомиозиновых иРНК, полученных из одного транскрипта (см рис)
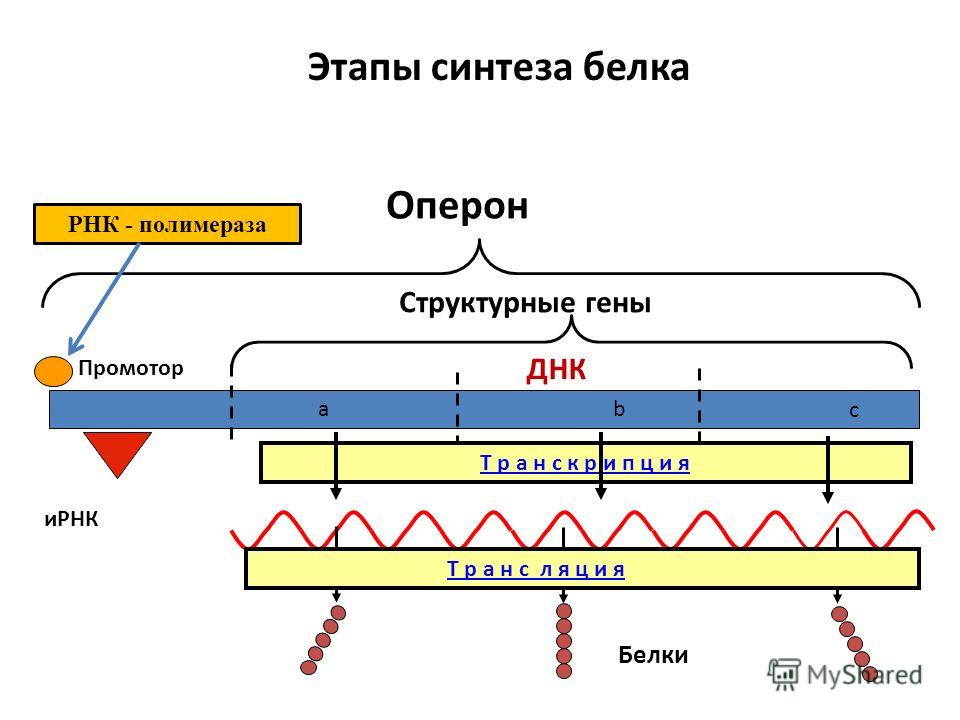
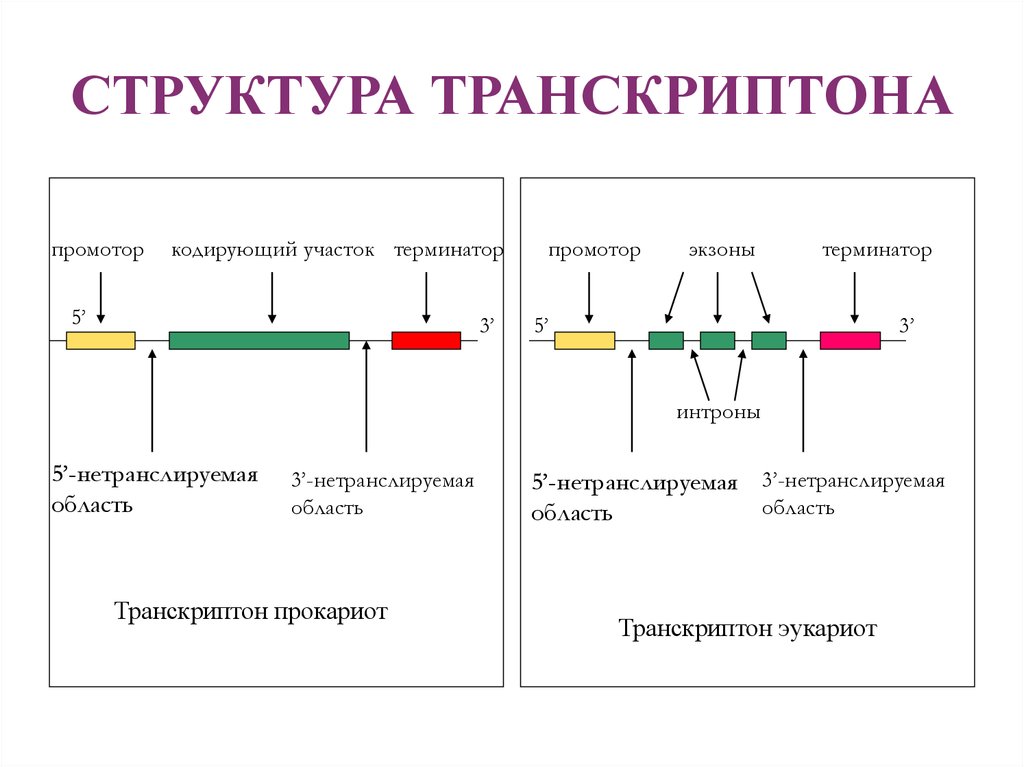
Презентация на тему: Строение транскриптона
Прокариоты
Промотор — участок оперона, служащий для узнавания ферментом РНК-полимеразой. Последовательность оснований по ходу цепи ДНК ниже сайта промотора с направлением 3’5’ используется в качестве матрицы для синтеза РНК.
После промотора в опероне находится акцепторная зона (у эукариот) или оператор (у прокариот), которая служит для связывания с регуляторами транскрипции (например, усилителями – энхансерами или репрессорами). Оператор разрешает или запрещает транскрипцию.
Цистрон — последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев — белок) или одну тРНК, или одну рРНК.
В результате транскрипции образуется полицистронная мРНК.
Лактозный оперон
lacZ кодирует β-галактозидазу |
lacA кодирует β-галактозид |
lacY кодирует β-галактозид пермеазу |
трансацетилазу |
Лактозный оперон
4-О-β-D-галактопиранозил-D-глюкоза
Эукариоты
Процессинг включает кэпирование (присоединение к 5’- концевому метилированному звену предшественника мРНК 7- метилгуанозина), полиаденилирование (присоединение к 3’-концу сегмента поли(А)), сплайсинг РНК (вырезание интронов и соединение экзонов – кодирующих последовательностей – в непрерывную последовательность)
Схема процессинга пре-мРНК в ядре
Процессинг – совокупность ферментативных процессов, в результате которых синтезируемая РНК превращается в функционально полноценную молекулу
мРНК эукариот
Функции кэпа и полиА хвоста
Функции кэпа:
1. Необходим для узнавания мРНК при инициации трансляции. 2.Для защиты 5′ конца РНК от рибонуклеаз, специфически разрезающих фосфодиэфирные связи в направлении 5’→3‘ и атакующих незащищённый 5′ конец.
Необходим для узнавания мРНК при инициации трансляции. 2.Для защиты 5′ конца РНК от рибонуклеаз, специфически разрезающих фосфодиэфирные связи в направлении 5’→3‘ и атакующих незащищённый 5′ конец.
3.Участвует в созревании 3′ конца мРНК.
4.Осуществляет экспорт мРНК из ядра в цитоплазму.
5.Участвует в сплайсинге.
6.Участвует в регуляции транскрипции.
Функции полиА хвоста
1.Способствуют экспорту зрелых мРНК из ядра.
2.Защищают мРНК от действия нуклеаз в цитоплазме, тем самым увеличивают время их жизни. Таким
образом, время жизни мРНК коррелирует с длиной поли(А) хвоста.
3.Служат в качестве сигнала узнавания для рибосомы.
Транскрипция РНК
Генетический код — это система записи информации о
последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК.
Свойства генетического кода
•- триплетность (43=64 кодона)
•- специфичность (один кодон — одна аминокислота) Исключение составляет кодон AUG.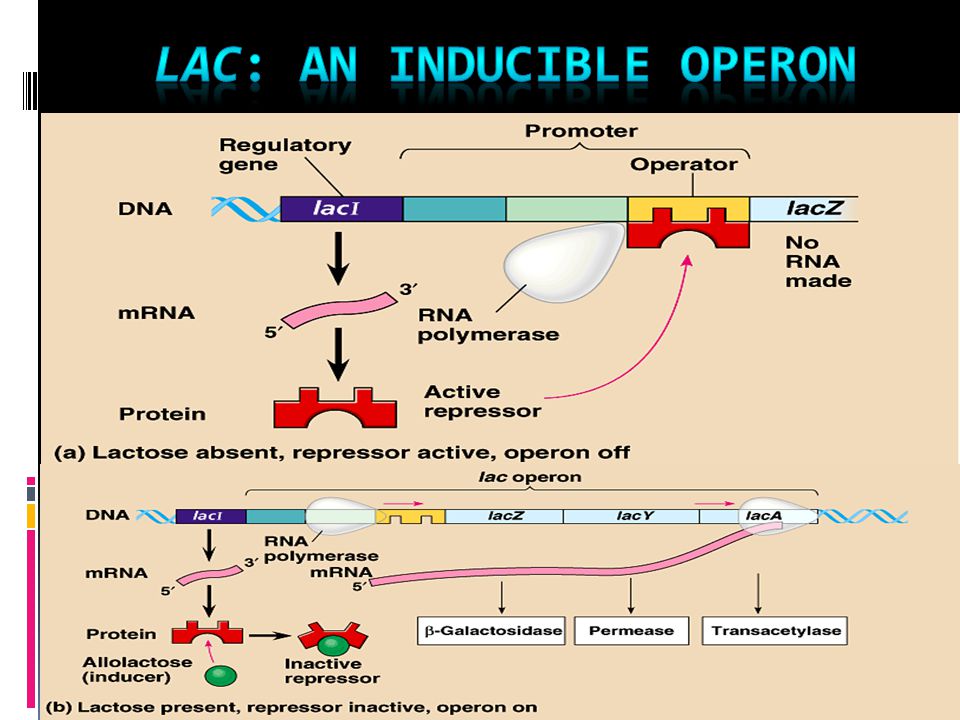 У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой — метионин. )
У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой — метионин. )
•- вырожденность (все аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом. Всего 61 триплет кодирует 20 аминокислот)
•- универсальность
•- однонаправленность (5’3’)
•- непрерывность
•- неперекрываемость
•- линейность
2.6: Транскрипция — Биология LibreTexts
-
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 8423
Поток генетической информации
Основная роль ДНК заключается в хранении наследуемой информации, которая кодирует инструкции по созданию организма.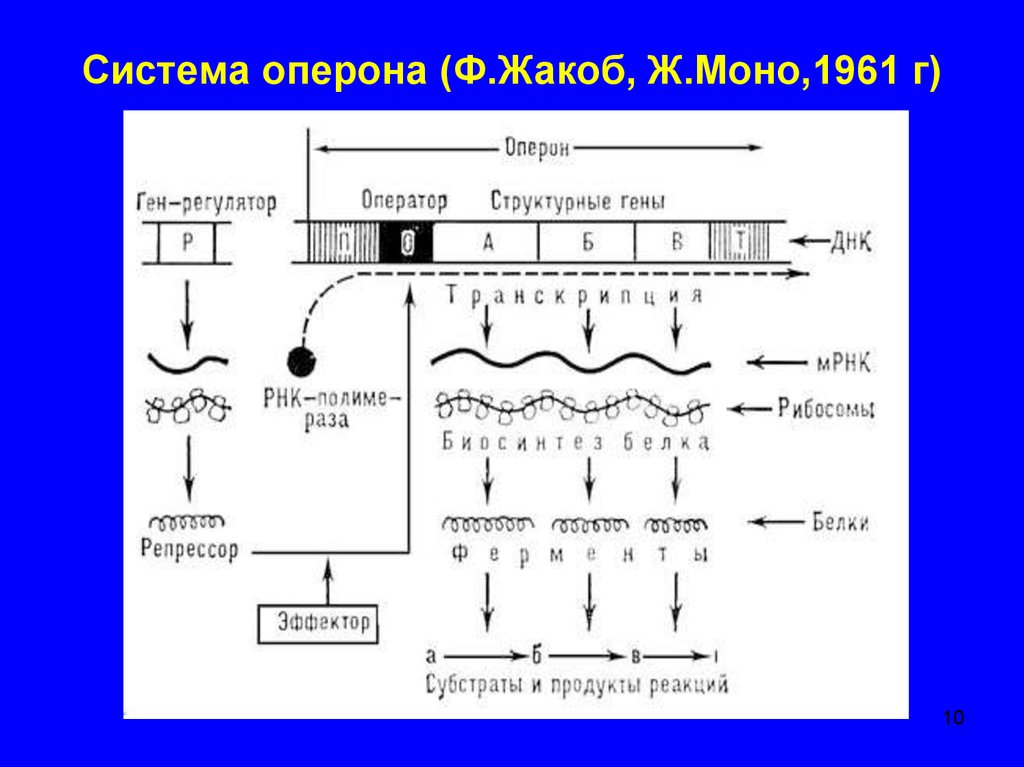 За последнее десятилетие мы очень хорошо научились секвенировать ДНК, но мы до сих пор не знаем, как надежно расшифровать ВСЮ информацию, и мы до сих пор не понимаем ВСЕ механизмы, с помощью которых она выражается.
За последнее десятилетие мы очень хорошо научились секвенировать ДНК, но мы до сих пор не знаем, как надежно расшифровать ВСЮ информацию, и мы до сих пор не понимаем ВСЕ механизмы, с помощью которых она выражается.
Однако существуют некоторые основные принципы и механизмы, связанные с чтением и выражением генетического кода, основные шаги которых понятны и которые должны стать частью концептуального инструментария для всех биологов. Два из этих процессов — транскрипция и трансляция ; копирование частей генетического кода, записанного в ДНК, в молекулы родственной полимерной РНК с последующим чтением и декодированием последовательности РНК (нуклеотидов) в последовательность белков (аминокислот).
В BIS2A мы в основном сосредоточены на развитии понимания процесса транскрипции (напомним, что Energy Story — это просто способ описания процесса) и его роли в выражении генетической информации. Мы мотивируем наше обсуждение транскрипции, сосредотачиваясь на функциональных проблемах (привнося часть нашей задачи по решению проблем/проектированию), которые должны быть решены, чтобы процесс состоялся.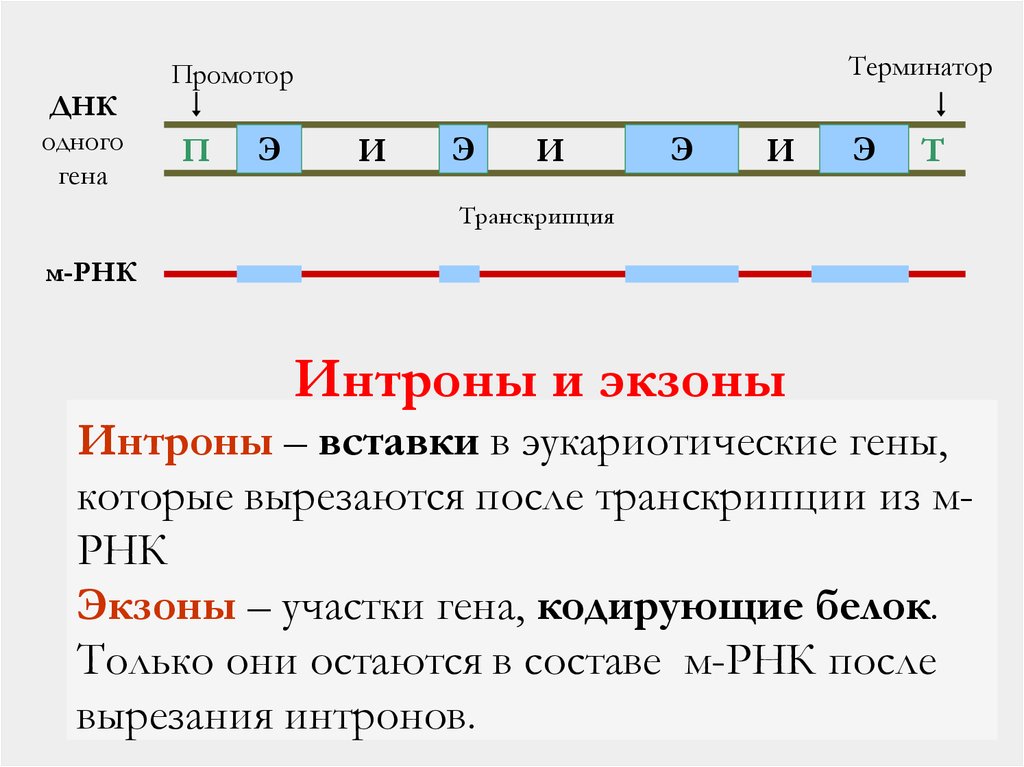 Затем мы переходим к описанию того, как природа использует этот процесс для создания множества функциональных молекул РНК (которые могут выполнять различные структурные, каталитические или регуляторные роли), включая молекулы информационной РНК (мРНК), которые несут информацию, необходимую для синтеза белков. Точно так же мы сосредоточимся на проблемах и вопросах, связанных с процессом трансляции, процессом, посредством которого рибосомы синтезируют белки.
Затем мы переходим к описанию того, как природа использует этот процесс для создания множества функциональных молекул РНК (которые могут выполнять различные структурные, каталитические или регуляторные роли), включая молекулы информационной РНК (мРНК), которые несут информацию, необходимую для синтеза белков. Точно так же мы сосредоточимся на проблемах и вопросах, связанных с процессом трансляции, процессом, посредством которого рибосомы синтезируют белки.
Основной поток генетической информации в биологических системах часто изображают в виде схемы, известной как «центральная догма» (см. рисунок ниже). Эта схема утверждает, что информация, закодированная в ДНК, передается в РНК посредством транскрипции и, в конечном счете, в белки посредством трансляции. Такие процессы, как обратная транскрипция (создание ДНК и матрицы РНК) и репликация, также представляют собой механизмы распространения информации в различных формах. Эта схема, однако, ничего не говорит о том, как кодируется информация или о механизмах, с помощью которых регуляторные сигналы перемещаются между различными слоями типов молекул, изображенных в модели.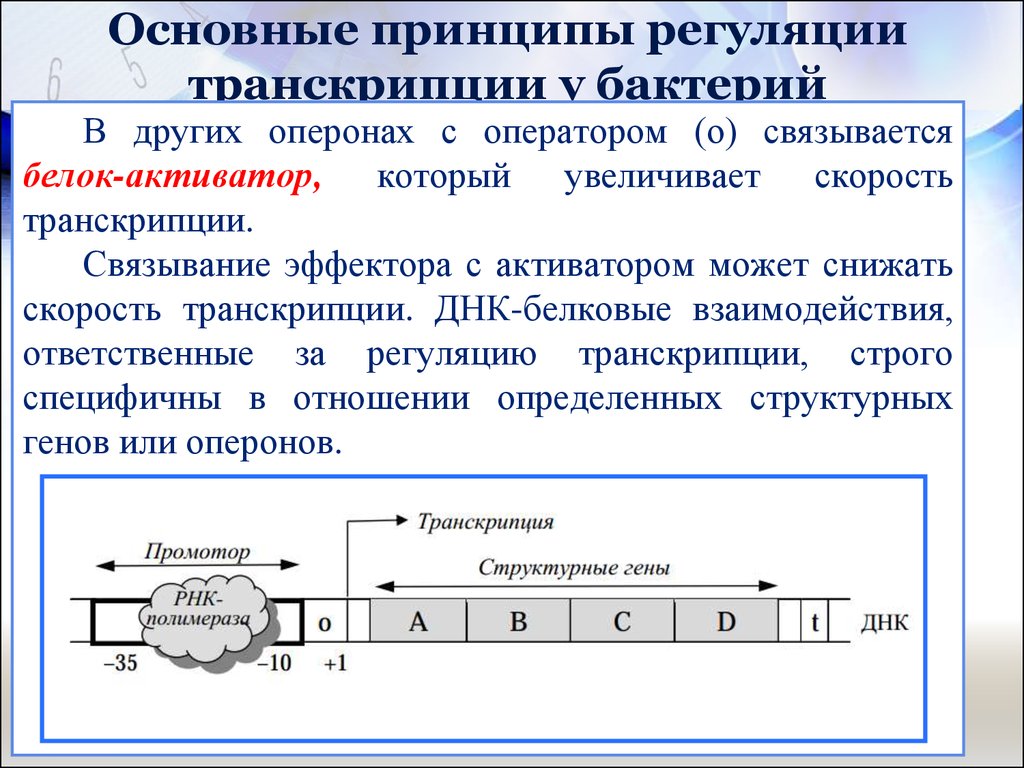 Поэтому, хотя приведенная ниже схема является почти обязательной частью лексикона любого биолога, возможно, оставшейся от старой традиции, учащиеся также должны знать, что механизмы потока информации более сложны (о некоторых мы узнаем по ходу дела, а что «центральная догма» представляет лишь некоторые основные пути9.0028
Поэтому, хотя приведенная ниже схема является почти обязательной частью лексикона любого биолога, возможно, оставшейся от старой традиции, учащиеся также должны знать, что механизмы потока информации более сложны (о некоторых мы узнаем по ходу дела, а что «центральная догма» представляет лишь некоторые основные пути9.0028
Генотип к фенотипу
Важным понятием в следующих разделах является взаимосвязь между генетической информацией, генотипом , и результатом ее выражения, фенотипом . Эти два термина и механизмы, которые их связывают, будут неоднократно обсуждаться в течение следующих нескольких недель — начните осваивать этот словарь.
Информация, хранящаяся в ДНК, находится в последовательности отдельных нуклеотидов при чтении в направлении от 5′ к 3′.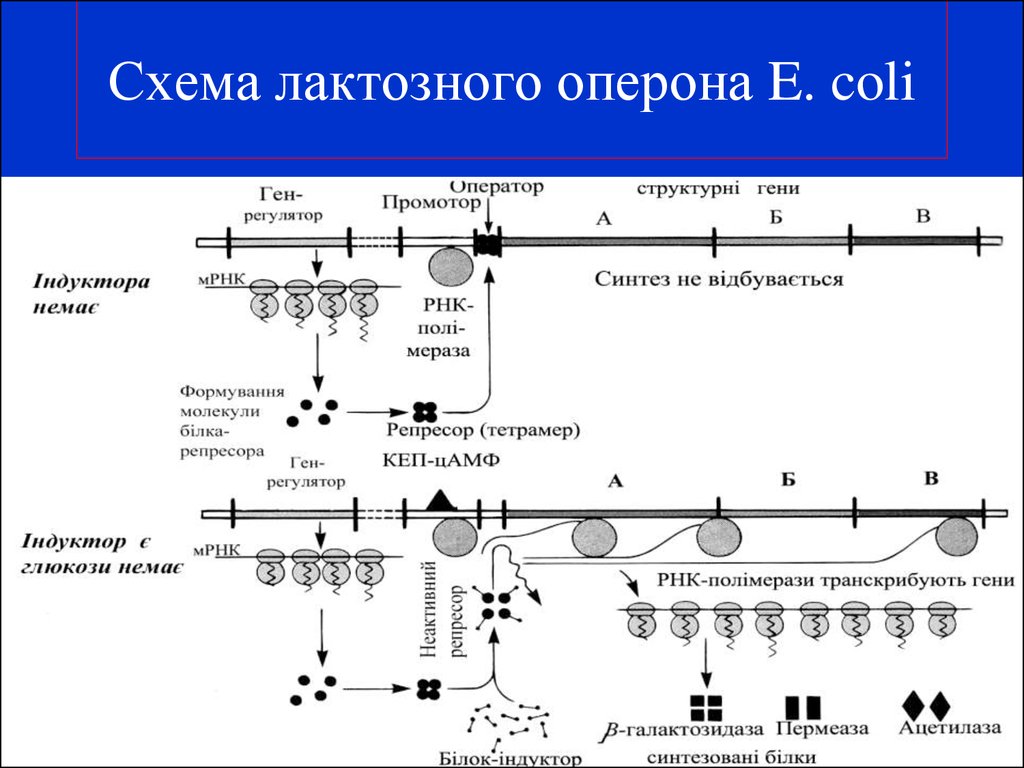 Преобразование информации из ДНК в РНК (процесс, называемый транскрипцией) создает вторую форму, которую информация принимает в клетке. мРНК используется в качестве матрицы для создания аминокислотной последовательности белков (в трансляции). Здесь показаны два разных набора информации. Последовательность ДНК немного отличается, в результате чего образуются две разные мРНК, за которыми следуют два разных белка и, в конечном итоге, два разных цвета шерсти у мышей.
Преобразование информации из ДНК в РНК (процесс, называемый транскрипцией) создает вторую форму, которую информация принимает в клетке. мРНК используется в качестве матрицы для создания аминокислотной последовательности белков (в трансляции). Здесь показаны два разных набора информации. Последовательность ДНК немного отличается, в результате чего образуются две разные мРНК, за которыми следуют два разных белка и, в конечном итоге, два разных цвета шерсти у мышей.
Генотип относится к информации, хранящейся в ДНК организма, последовательности нуклеотидов, компиляции его генов. Фенотип относится к любой физической характеристике, которую вы можете измерить, такой как рост, вес, количество продуцируемого АТФ, способность метаболизировать лактозу, реакция на стимулы окружающей среды и т. д. Различия в генотипе, даже незначительные, могут привести к различным фенотипам, которые подвергается естественному отбору. Рисунок выше иллюстрирует эту идею.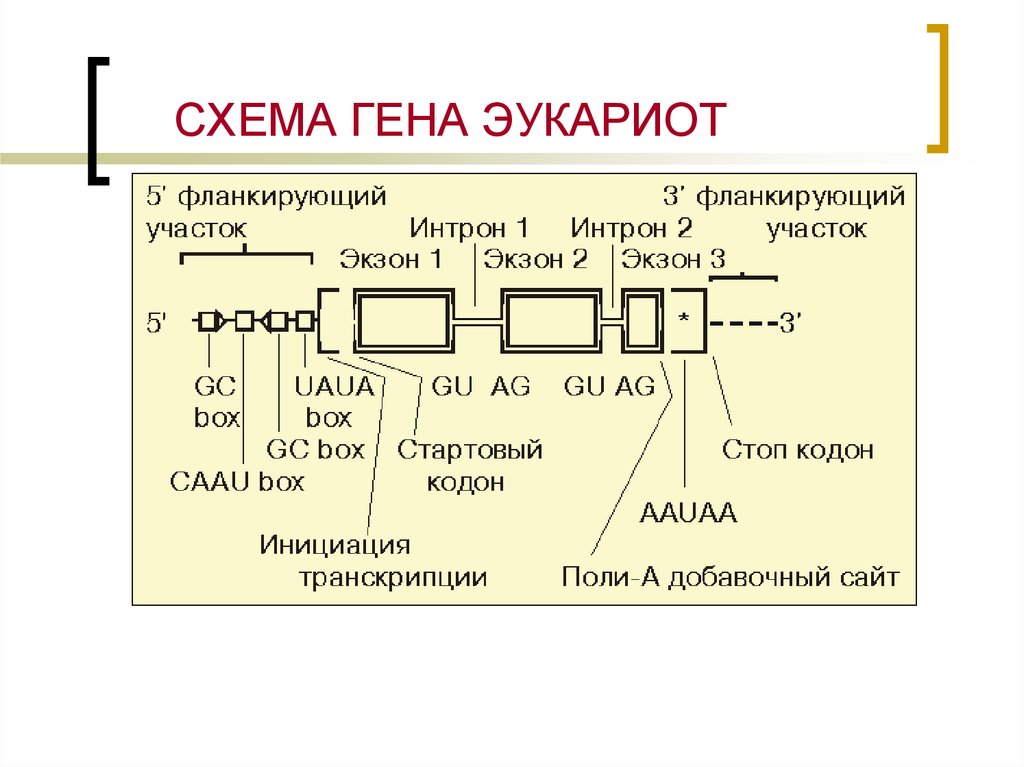 Заметим также, что, хотя классические обсуждения взаимосвязей генотипа и фенотипа ведутся в контексте многоклеточных организмов, эта номенклатура и лежащие в ее основе концепции применимы ко всем организмам, даже к одноклеточным организмам, таким как бактерии и археи.
Заметим также, что, хотя классические обсуждения взаимосвязей генотипа и фенотипа ведутся в контексте многоклеточных организмов, эта номенклатура и лежащие в ее основе концепции применимы ко всем организмам, даже к одноклеточным организмам, таким как бактерии и археи.
Предлагаемое обсуждение
Можно ли считать фенотипом то, что не видно «на глаз»?
Предлагаемое обсуждение
Могут ли одноклеточные организмы иметь одновременно несколько фенотипов? Если да, можете ли вы предложить пример? Если нет, то почему?
Гены
Что такое ген? Ген представляет собой сегмент ДНК в геноме организма, который кодирует функциональную РНК (такую как рРНК или тРНК и т. д.) или белковый продукт (ферменты, тубулин и т. д.). Общий ген содержит элементы, кодирующие регуляторные области (которые часто не транскрибируются), и область, кодирующую транскрибируемую единицу.
Гены могут приобретать мутации , определяемые как изменения в составе и/или последовательности нуклеотидов, либо в кодирующих, либо в регуляторных областях.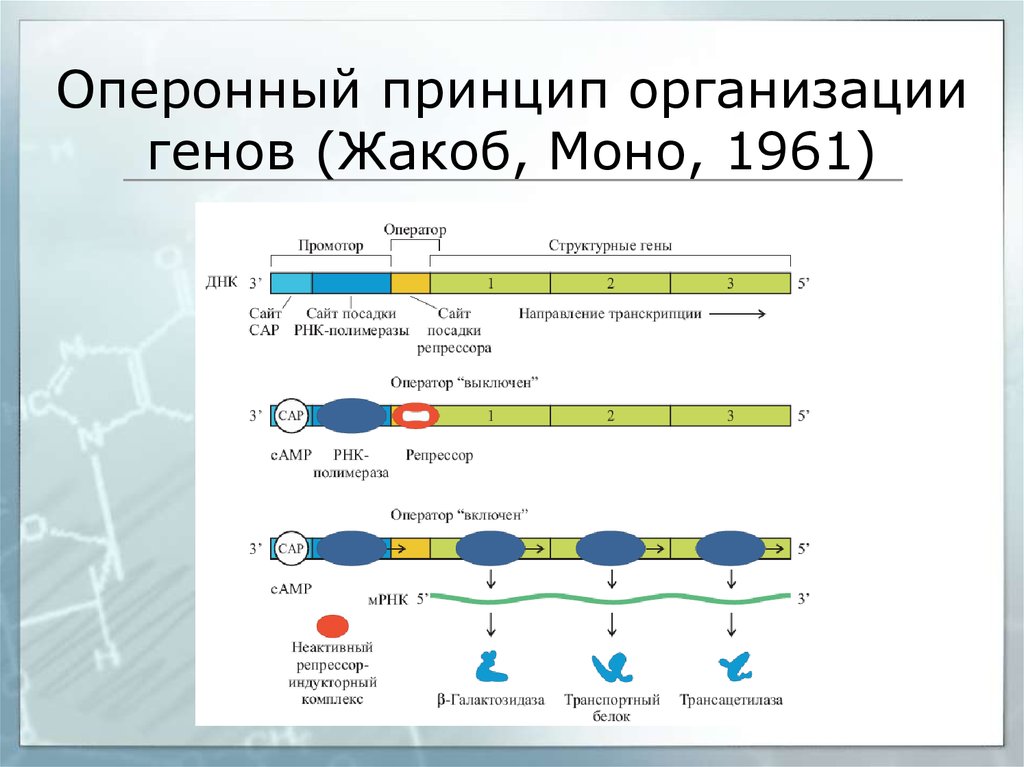 Эти мутации могут привести к нескольким возможным результатам: (1) в результате не происходит ничего измеримого; (2) ген больше не экспрессируется; или (3) экспрессия или поведение продукта(ов) гена отличаются. В популяции организмов, имеющих один и тот же ген, разные варианты гена известны как аллеля 9.0025 . Различные аллели могут приводить к различиям в фенотипах индивидуумов и способствовать разнообразию биологии, находящейся под давлением отбора.
Эти мутации могут привести к нескольким возможным результатам: (1) в результате не происходит ничего измеримого; (2) ген больше не экспрессируется; или (3) экспрессия или поведение продукта(ов) гена отличаются. В популяции организмов, имеющих один и тот же ген, разные варианты гена известны как аллеля 9.0025 . Различные аллели могут приводить к различиям в фенотипах индивидуумов и способствовать разнообразию биологии, находящейся под давлением отбора.
Начните изучать эти словарные термины и связанные с ними понятия. Вы будете в некоторой степени знакомы с ними, когда мы начнем углубляться в них более подробно в следующих лекциях.

Резюме раздела
Все живые существа должны транскрибировать гены из своего генома. Хотя клеточное расположение может быть различным (эукариоты осуществляют транскрипцию в ядре, бактерии и археи, не имеющие ядра, осуществляют транскрипцию в цитоплазме), механизмы, с помощью которых организмы из каждой из этих ветвей осуществляют этот процесс, принципиально одинаковы и могут выделяют три стадии: инициацию, элонгацию и терминацию.
Транскрипция: от ДНК к РНК
A краткий обзор транскрипции
Транскрипция – это процесс создания РНК-копии сегмента ДНК. Поскольку это процесс , мы хотим применить Энергетическую Историю, чтобы развить функциональное понимание транскрипции. Как выглядит система молекул до начала транскрипции? Как это выглядит в конце? Какие превращения материи и переносы энергии происходят при транскрипции и что катализирует этот процесс? Мы также хотим рассмотреть этот процесс с точки зрения Design Challenge.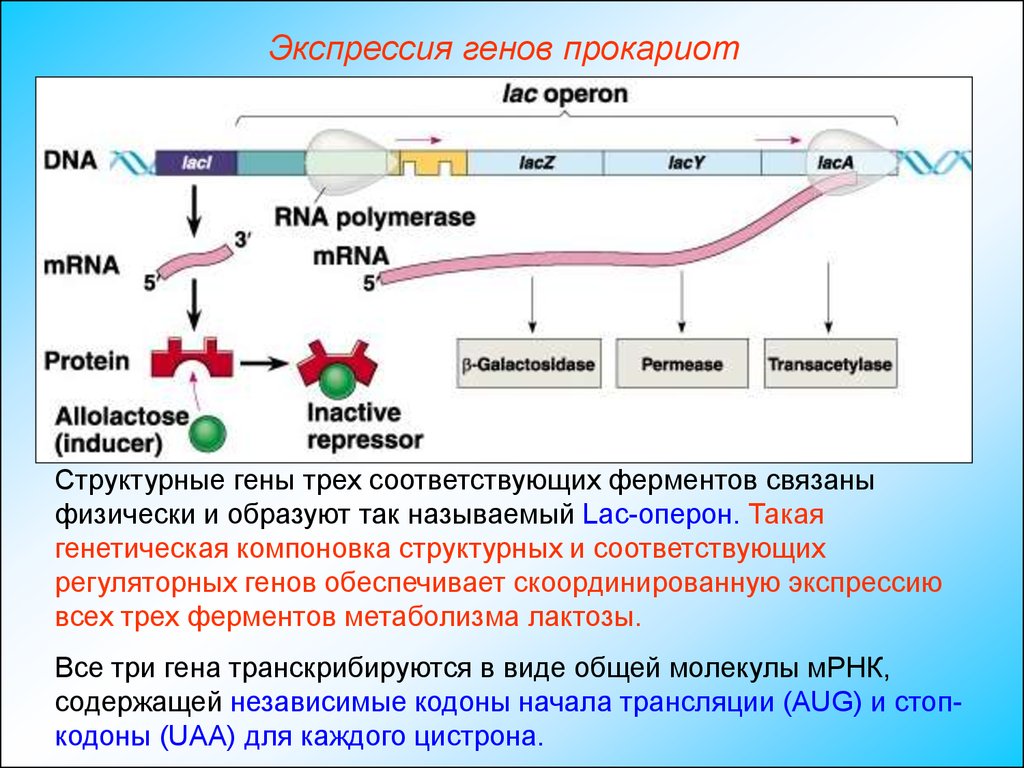 Если биологическая задача состоит в том, чтобы создать копию ДНК на химическом языке РНК, какие проблемы мы можем разумно предположить или предвидеть, учитывая наши знания о других процессах нуклеотидных полимеров, которые необходимо преодолеть? Есть ли свидетельства того, что Природа решала эти проблемы по-разному? Каковы, по-видимому, критерии успеха транскрипции? Вы поняли идею.
Если биологическая задача состоит в том, чтобы создать копию ДНК на химическом языке РНК, какие проблемы мы можем разумно предположить или предвидеть, учитывая наши знания о других процессах нуклеотидных полимеров, которые необходимо преодолеть? Есть ли свидетельства того, что Природа решала эти проблемы по-разному? Каковы, по-видимому, критерии успеха транскрипции? Вы поняли идею.
Перечислим некоторые основные требования к транскрипции
Давайте сначала рассмотрим поставленные задачи, используя некоторые из наших фундаментальных знаний и представив, что может произойти в процессе транскрипции, если целью является создание копии РНК. фрагмент одной нити двухцепочечной молекулы ДНК. Мы увидим, что использование некоторой базовой логики позволяет нам сделать вывод о многих важных вопросах и вещах, о которых нам нужно знать, чтобы правильно описать процесс.
Давайте представим, что мы хотим спроектировать наномашину/нанобота, который будет выполнять транскрипцию. Мы можем использовать некоторые подходы к проектированию, чтобы определить проблемы и подзадачи, которые должен решить наш маленький робот.
Мы можем использовать некоторые подходы к проектированию, чтобы определить проблемы и подзадачи, которые должен решить наш маленький робот.
• Первое, что нам нужно, чтобы наша машина знала, это с чего начать. Куда должна быть направлена машина от миллионов до миллиардов пар оснований?
• Точно так же нам нужно знать, где остановиться.
• Если у нас есть начальный и конечный сайты, нам потребуются способы кодирования этой информации, чтобы наши машины могли прочитать эту информацию — как это будет достигнуто?
• Сколько РНК-копий ДНК нам нужно будет сделать?
• Как быстро нужно делать копии РНК?
• Насколько точно должны быть сделаны копии?
• Сколько энергии потребуется для процесса и откуда она будет браться?
Конечно, это лишь некоторые из основных вопросов. При желании можно копнуть глубже. Тем не менее, они уже достаточно хороши, чтобы мы начали хорошо чувствовать этот процесс. Заметьте также, что многие из этих вопросов удивительно похожи на те, которые, как мы предполагали, могут быть необходимы для понимания репликации ДНК.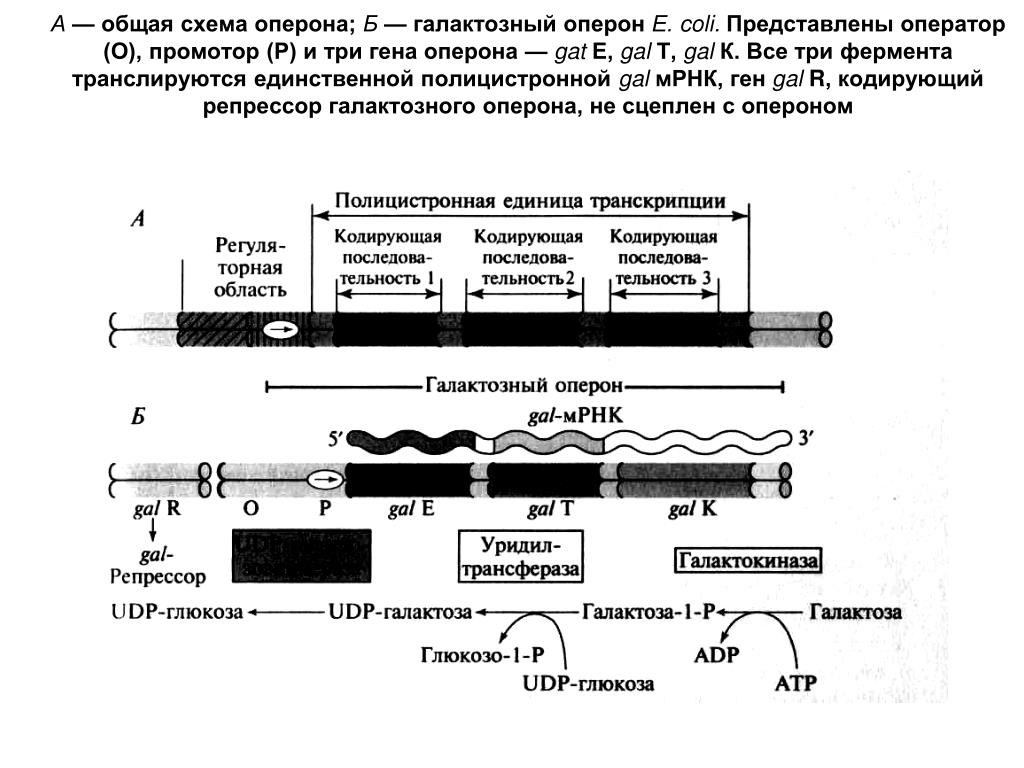
Строительные блоки транскрипции
Строительные блоки РНК
Вспомним из нашего обсуждения структуры нуклеотидов, что строительные блоки РНК очень похожи на блоки ДНК. В РНК строительные блоки состоят из нуклеотидтрифосфатов, которые состоят из сахара рибозы, азотистого основания и трех фосфатных групп. Ключевое различие между строительными блоками ДНК и РНК заключается в том, что молекулы РНК состоят из нуклеотидов с рибозными сахарами (в отличие от дезоксирибозных сахаров) и что вместо тимидина (тиминсодержащего нуклеотида) РНК использует уридин (урацилсодержащий нуклеотид). нуклеотид). Обратите внимание, что урацил и тимин структурно очень похожи — в урациле просто отсутствует метил (CH 3 ) функциональная группа по сравнению с тимином.
Основные химические компоненты нуклеотидов.
Атрибуция: Marc T. Facciotti (оригинальная работа)
Инициация транскрипции
Промоторы
Белки, ответственные за создание РНК-копии определенного фрагмента ДНК (начало транскрипции), должны сначала быть в состоянии распознать начало копируемый элемент. Промотор представляет собой последовательность ДНК, с которой различные белки, известные под общим названием «машина транскрипции», связываются и инициируют транскрипцию. В большинстве случаев промоторы существуют выше (5′ от кодирующей области) генов, которые они регулируют. Конкретная последовательность промотора очень важна, потому что она определяет, будет ли соответствующая кодирующая часть гена транскрибироваться все время, время от времени или нечасто.
Промотор представляет собой последовательность ДНК, с которой различные белки, известные под общим названием «машина транскрипции», связываются и инициируют транскрипцию. В большинстве случаев промоторы существуют выше (5′ от кодирующей области) генов, которые они регулируют. Конкретная последовательность промотора очень важна, потому что она определяет, будет ли соответствующая кодирующая часть гена транскрибироваться все время, время от времени или нечасто.
В бактерии E. coli в областях -10 и -35 выше сайта инициации (сайта первого нуклеотида транскрипта) имеются две промоторные консенсусные последовательности или области, которые сходны через многие промоторы и через различные родственные виды. Некоторые промоторы будут иметь последовательность, очень похожую на консенсусную последовательность (последовательность, содержащую наиболее распространенные элементы последовательности), а другие будут выглядеть совсем иначе. Эти вариации последовательности влияют на силу, с которой механизм транскрипции может связываться с промотором, чтобы инициировать транскрипцию. Это помогает контролировать количество сделанных расшифровок и частоту их выполнения.
Это помогает контролировать количество сделанных расшифровок и частоту их выполнения.
(а) Общая схема гена. Ген включает промоторную последовательность, нетрансляционную область (UTR) и кодирующую последовательность. (b) Список нескольких сильных промоторных последовательностей E. coli. Блок -35 и блок -10 представляют собой высококонсервативные последовательности во всем списке сильных промоторов. Более слабые промоторы будут иметь больше различий в парах оснований по сравнению с этими последовательностями. Источник: http://www.discoveryandinnovation.co…lecture12.html
Предлагаемое обсуждение
Какие типы взаимодействий между механизмом транскрипции и ДНК изменяются при изменении нуклеотидной последовательности промотора? Почему некоторые последовательности создают «сильный» промотор, а другие — «слабый»?
Бактериальные и эукариотические промоторы
В бактериальных клетках область -10 богата АТ, часто ТАТААТ.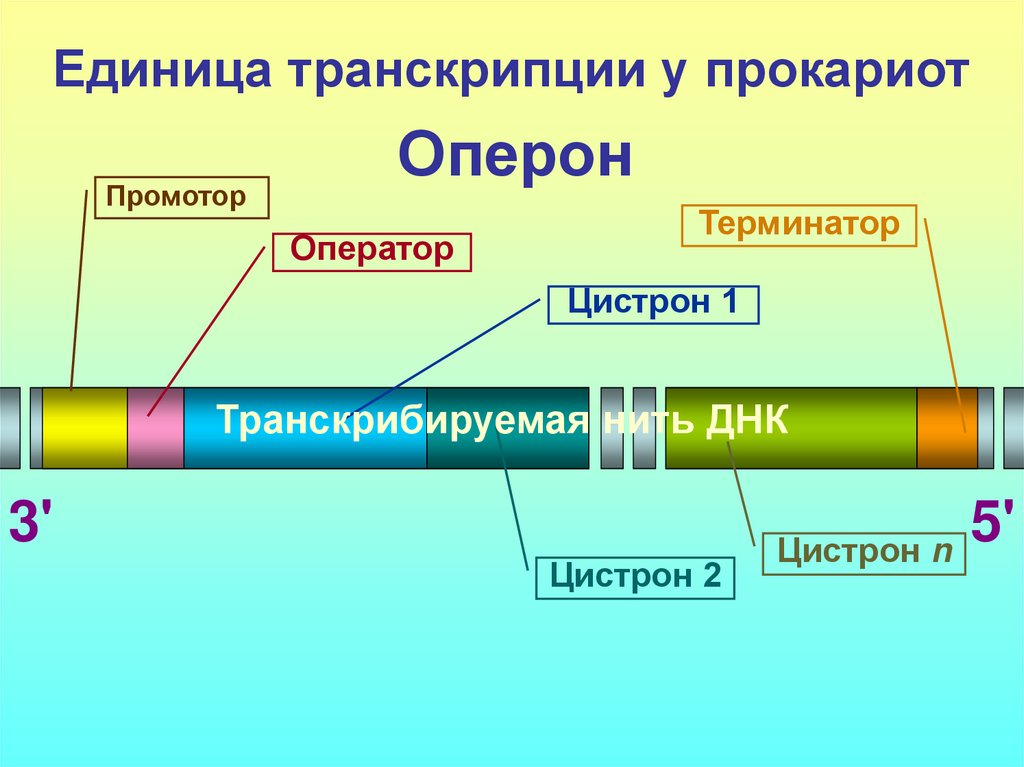 Последовательности непосредственно перед областью -10, а также областью -35 (TTGACA) распознаются и связываются белком σ («сигма-фактор»), который является одним из компонентов холофермента РНК-полимеразы. Как только это взаимодействие белок-ДНК установлено, сигма-фактор облегчает раскручивание области -10 и загружает полимеразу на цепь матрицы. Таким образом, сигма-фактор помогает полимеразе распознавать промоторные последовательности и загружать полимеразу в нужное место, указывая в правильном направлении. Интересно, что E. coli производит несколько различных сигма-факторов. В разных ситуациях, например при определенном стрессе, активация другого сигма-фактора будет изменять типы генов, наиболее часто транскрибируемых РНК-полимеразой. Некоторые бактериофаги воспользовались этим аспектом бактериальной транскрипции, производя свои сигма-факторы, специфичные для фаговых генов, которые перехватывают РНК-полимеразу хозяина и перенаправляют ее в геном фага.
Последовательности непосредственно перед областью -10, а также областью -35 (TTGACA) распознаются и связываются белком σ («сигма-фактор»), который является одним из компонентов холофермента РНК-полимеразы. Как только это взаимодействие белок-ДНК установлено, сигма-фактор облегчает раскручивание области -10 и загружает полимеразу на цепь матрицы. Таким образом, сигма-фактор помогает полимеразе распознавать промоторные последовательности и загружать полимеразу в нужное место, указывая в правильном направлении. Интересно, что E. coli производит несколько различных сигма-факторов. В разных ситуациях, например при определенном стрессе, активация другого сигма-фактора будет изменять типы генов, наиболее часто транскрибируемых РНК-полимеразой. Некоторые бактериофаги воспользовались этим аспектом бактериальной транскрипции, производя свои сигма-факторы, специфичные для фаговых генов, которые перехватывают РНК-полимеразу хозяина и перенаправляют ее в геном фага.
Эукариотические промоторы намного крупнее и сложнее, чем прокариотические промоторы, но оба имеют AT-богатую область — у эукариот ее обычно называют «боксом ТАТА». Например, в гене тимидинкиназы мыши бокс ТАТА расположен примерно на -30. Для этого гена точной последовательностью ТАТА-бокса является ТАТАААА, прочитанная в направлении от 5′ к 3′ на нематричной цепи. Эта последовательность не идентична области E. coli -10, но обе они имеют общее качество элементов, богатых А-Т.
Например, в гене тимидинкиназы мыши бокс ТАТА расположен примерно на -30. Для этого гена точной последовательностью ТАТА-бокса является ТАТАААА, прочитанная в направлении от 5′ к 3′ на нематричной цепи. Эта последовательность не идентична области E. coli -10, но обе они имеют общее качество элементов, богатых А-Т.
Вместо одной бактериальной полимеразы геномы большинства эукариот кодируют три разные РНК-полимеразы, каждая из которых состоит из 10 или более белковых субъединиц. Каждой эукариотической полимеразе также требуется отдельный набор белков, известных как факторов транскрипции , чтобы привлечь ее к промотору. К сожалению, терминология факторов транскрипции весьма непоследовательна. Достаточно сказать, что существует много белков, которые всегда должны действовать одновременно, чтобы загрузить, например, любую РНК-полимеразу II (полимеразу, транскрибирующую мРНК). Их называют «базальными» (или «общими») факторами транскрипции. Кроме того, армия белков может влиять на частоту притяжения этих базальных факторов к промоторам, частоту загрузки РНК pol II и даже «ускользание» комплекса pol II от эукариотических промоторов (чтобы он мог фактически выполнить транскрипцию). Усилители и глушители — оба Последовательности ДНК , а не белки, распознаются этими регуляторными факторами транскрипции. Базальные факторы транскрипции имеют решающее значение в формировании преинициаторного комплекса на ДНК-матрице, который впоследствии привлекает РНК-полимеразу для инициации транскрипции.
Усилители и глушители — оба Последовательности ДНК , а не белки, распознаются этими регуляторными факторами транскрипции. Базальные факторы транскрипции имеют решающее значение в формировании преинициаторного комплекса на ДНК-матрице, который впоследствии привлекает РНК-полимеразу для инициации транскрипции.
изображение из Kelvinsong
Независимо от деталей локализации и ориентации бактериальной и эукариотической полимеразы, инициация транскрипции начинается со связывания РНК-полимеразы с промоутер . Для транскрипции требуется, чтобы двойная спираль ДНК частично раскручивалась, чтобы одну из цепей можно было использовать в качестве матрицы для синтеза РНК. Обратите внимание, что раскручивание происходит внутри полимеразы; РНК-полимераза, в отличие от ДНК-полимеразы, обладает внутренней геликазной активностью. Двухцепочечная ДНК всасывается в фермент (как и НТФ), и появляется двухцепочечная ДНК плюс РНК.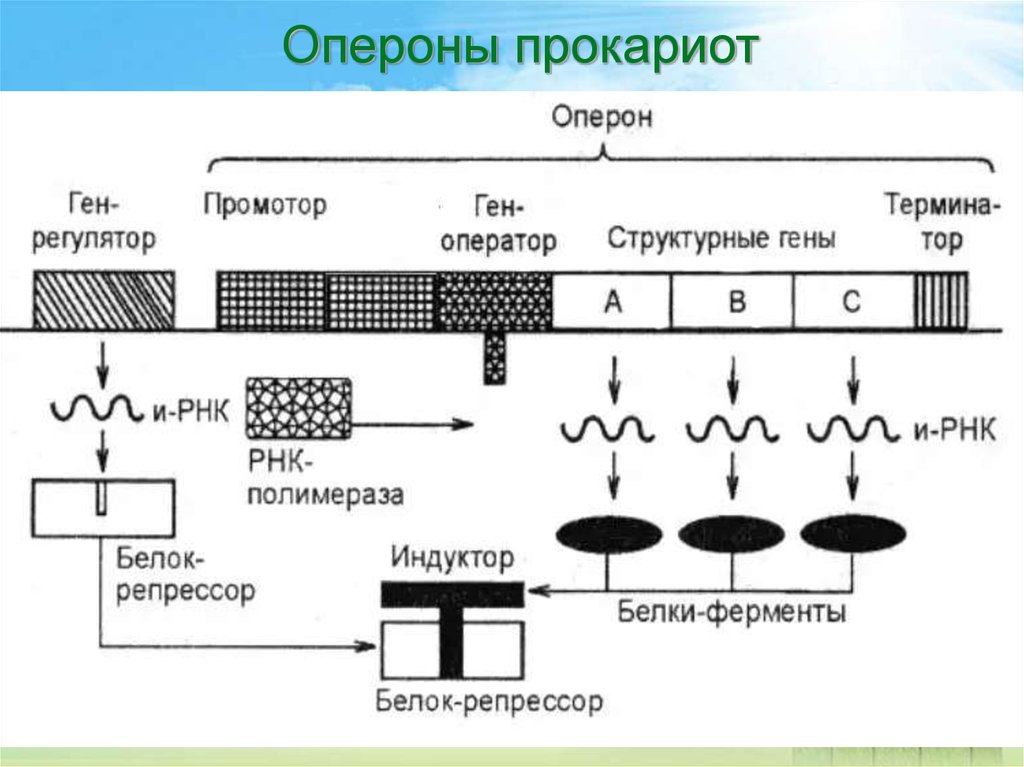 Область раскручивания называется транскрипционным пузырем.
Область раскручивания называется транскрипционным пузырем.
Элонгация
Транскрипция всегда происходит с одной из двух цепей ДНК, которая называется матричной нитью . Продукт РНК комплементарен матричной цепи и почти идентичен нематричной цепи, называемой кодирующей цепью , за исключением того, что РНК содержит урацил (U) вместо тимина (T), обнаруженного в ДНК.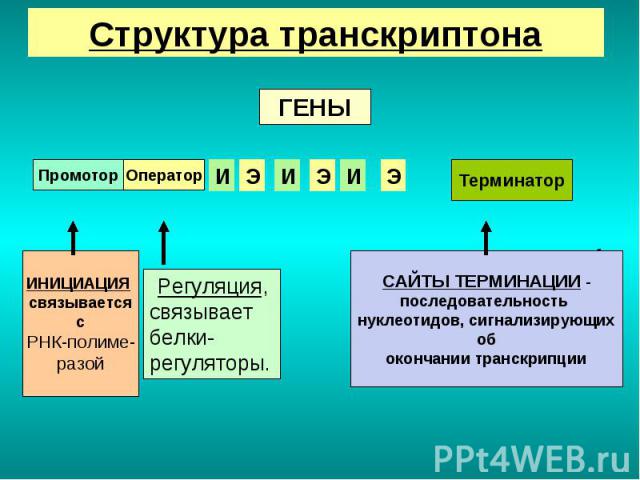 Во время элонгации фермент, называемый РНК-полимеразой , движется вдоль матрицы ДНК, добавляя нуклеотиды путем спаривания оснований с матрицей ДНК, аналогично репликации ДНК, с той разницей, что синтезируется цепь РНК, которая не остается связанной с ДНК. шаблон. По мере удлинения ДНК непрерывно раскручивается перед коровым ферментом и закручивается позади него. Обратите внимание, что направление синтеза идентично направлению синтеза в ДНК — от 5′ к 3′.
Во время элонгации фермент, называемый РНК-полимеразой , движется вдоль матрицы ДНК, добавляя нуклеотиды путем спаривания оснований с матрицей ДНК, аналогично репликации ДНК, с той разницей, что синтезируется цепь РНК, которая не остается связанной с ДНК. шаблон. По мере удлинения ДНК непрерывно раскручивается перед коровым ферментом и закручивается позади него. Обратите внимание, что направление синтеза идентично направлению синтеза в ДНК — от 5′ к 3′.
Во время элонгации РНК-полимераза отслеживает ДНК-матрицу, синтезирует мРНК в направлении от 5′ к 3′ и раскручивает, а затем перематывает ДНК по мере ее считывания. Обратите внимание, что РНК-полимераза «всасывает» двухцепочечную ДНК и рибонуклеотиды и удаляет двухцепочечную ДНК плюс РНК. Как это соотносится с ДНК-полимеразой?
 РНК полимеризуется от 5′ к 3′, двигаясь антипараллельно прямо относительно матрицы. С каждым добавлением нуклеотида фосфоангидридная связь гидролизуется ферментом, что приводит к более длинному полимеру и высвобождению пирофосфата, который позже расщепляется до фосфата.
РНК полимеризуется от 5′ к 3′, двигаясь антипараллельно прямо относительно матрицы. С каждым добавлением нуклеотида фосфоангидридная связь гидролизуется ферментом, что приводит к более длинному полимеру и высвобождению пирофосфата, который позже расщепляется до фосфата. Предлагаемое обсуждение
Сравните и сопоставьте энергетическую историю добавления нуклеотида в репликации ДНК с добавлением нуклеотида в транскрипцию.
Удлинение бактерий и эукариот
У бактерий удлинение начинается с высвобождения субъединицы σ холофермента РНК-полимеразы. Диссоциация σ позволяет коровому ферменту двигаться вдоль матрицы ДНК, синтезируя мРНК в направлении от 5′ к 3′ со скоростью примерно 40 нуклеотидов в секунду. По мере удлинения ДНК непрерывно раскручивается перед коровым ферментом и закручивается позади него. Спаривание оснований между ДНК и РНК недостаточно стабильно, чтобы поддерживать стабильность компонентов синтеза мРНК.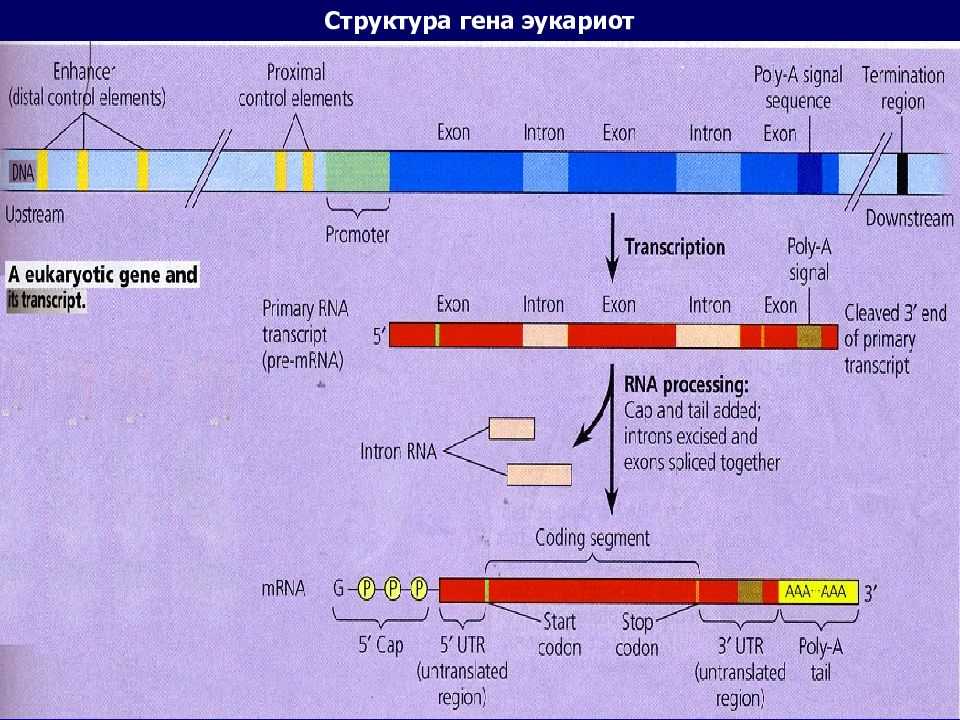 Вместо этого РНК-полимераза действует как стабильный линкер между матрицей ДНК и зарождающимися цепями РНК, чтобы гарантировать, что удлинение не прервется преждевременно.
Вместо этого РНК-полимераза действует как стабильный линкер между матрицей ДНК и зарождающимися цепями РНК, чтобы гарантировать, что удлинение не прервется преждевременно.
У эукариот после образования преинициаторного комплекса полимераза высвобождается из других факторов транскрипции, и элонгация может происходить, как это происходит у прокариот, когда полимераза синтезирует пре-мРНК в направлении от 5′ к 3′. Как обсуждалось ранее, РНК-полимераза II транскрибирует большую часть эукариотических генов, поэтому в этом разделе основное внимание будет уделено тому, как эта полимераза осуществляет элонгацию и терминацию.
Завершение
В бактериях
После того, как ген транскрибирован, бактериальной полимеразе необходимо дать указание диссоциировать от матрицы ДНК и высвободить вновь созданную мРНК. В зависимости от транскрибируемого гена существует два вида сигналов терминации. Один основан на белке, а другой на основе РНК. Rho-зависимая терминация контролируется белком rho, который следует за полимеразой на растущей цепи мРНК. Ближе к концу гена полимераза встречает серию нуклеотидов G на матрице ДНК и останавливается. В результате белок rho сталкивается с полимеразой. Взаимодействие с rho высвобождает мРНК из транскрипционного пузыря.
Rho-зависимая терминация контролируется белком rho, который следует за полимеразой на растущей цепи мРНК. Ближе к концу гена полимераза встречает серию нуклеотидов G на матрице ДНК и останавливается. В результате белок rho сталкивается с полимеразой. Взаимодействие с rho высвобождает мРНК из транскрипционного пузыря.
Rho-независимая терминация контролируется специфическими последовательностями в матричной цепи ДНК. Когда полимераза приближается к концу транскрибируемого гена, она сталкивается с областью, богатой нуклеотидами C-G. мРНК сворачивается сама по себе, и комплементарные нуклеотиды C-G связываются вместе. В результате получается стабильная шпилька , которая заставляет полимеразу останавливаться, как только она начинает транскрибировать область, богатую нуклеотидами AT. Комплементарная U-A-область транскрипта мРНК образует лишь слабое взаимодействие с матричной ДНК. Это, в сочетании с остановленной полимеразой, вызывает достаточную нестабильность для того, чтобы коровый фермент оторвался и высвободил новый транскрипт мРНК.
У эукариот
Терминация транскрипции различна для разных полимераз. В отличие от прокариот, элонгация РНК-полимеразой II у эукариот происходит на 1000–2000 нуклеотидов за конец транскрибируемого гена. Этот хвост пре-мРНК впоследствии удаляется расщеплением во время процессинга мРНК. С другой стороны, РНК-полимеразы I и III нуждаются в сигналах терминации. Гены, транскрибируемые РНК-полимеразой I, содержат специфическую последовательность из 18 нуклеотидов, которая распознается терминирующим белком. Процесс терминации у РНК-полимеразы III включает шпильку мРНК, сходную с ро-независимой терминацией транскрипции у прокариот.
У архей
Терминация транскрипции у архей изучена гораздо меньше, чем у двух других доменов жизни, и до сих пор недостаточно изучена. Хотя функциональные детали, вероятно, напоминают механизмы, которые наблюдались в других сферах жизни, детали выходят за рамки этого курса.
Расположение в клетке
У бактерий и архей
У бактерий и архей транскрипция происходит в цитоплазме, где находится ДНК.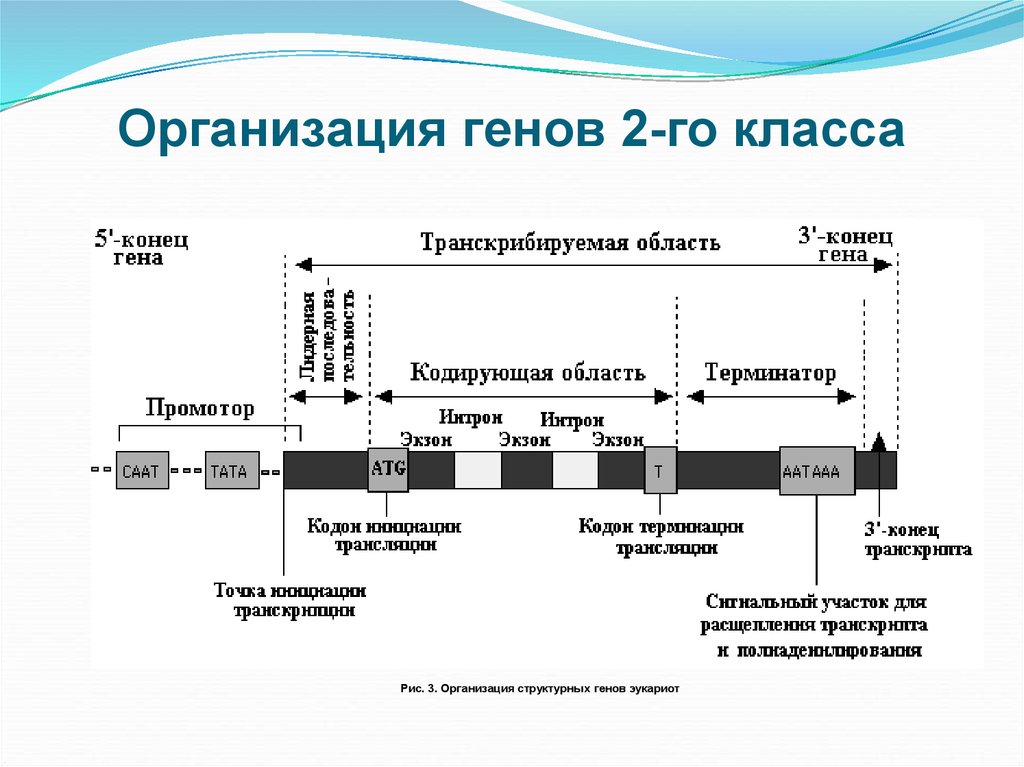 Поскольку расположение ДНК и, следовательно, процесс транскрипции физически не отделены от остальной части клетки, трансляция часто начинается до завершения транскрипции. Это означает, что мРНК у бактерий и архей используется в качестве матрицы для белка до того, как будет произведена вся мРНК. Отсутствие пространственной сегрегации также означает, что для этих процессов очень мало временной сегрегации. На изображении ниже показаны процессы транскрипции и трансляции, происходящие одновременно.
Поскольку расположение ДНК и, следовательно, процесс транскрипции физически не отделены от остальной части клетки, трансляция часто начинается до завершения транскрипции. Это означает, что мРНК у бактерий и архей используется в качестве матрицы для белка до того, как будет произведена вся мРНК. Отсутствие пространственной сегрегации также означает, что для этих процессов очень мало временной сегрегации. На изображении ниже показаны процессы транскрипции и трансляции, происходящие одновременно.
У эукариот….
У эукариот процесс транскрипции физически отделен от остальной части клетки, изолирован внутри ядра. Это приводит к двум вещам: мРНК завершается до того, как может начаться трансляция, и есть время «настроить» или «отредактировать» мРНК до начала трансляции. Физическое разделение этих процессов дает эукариотам возможность изменить мРНК таким образом, чтобы: увеличить продолжительность жизни мРНК или даже изменить белковый продукт, который будет произведен из мРНК.
Процессинг мРНК
5′ G-Cap и 3′ Poly-A хвост
При транскрипции эукариотического гена первичный транскрипт обрабатывается в ядре несколькими способами. Эукариотические мРНК модифицируются на 3′-конце путем добавления поли-А-хвоста. Эта серия остатков А добавляется ферментом, который не использует геномную ДНК в качестве матрицы. Кроме того, мРНК имеют химическую модификацию 5′-конца, называемую 5′-кэпом. Данные свидетельствуют о том, что эти модификации помогают увеличить продолжительность жизни мРНК (предотвращают ее преждевременную деградацию в цитоплазме), а также помогают мРНК инициировать трансляцию.
Данные свидетельствуют о том, что эти модификации помогают увеличить продолжительность жизни мРНК (предотвращают ее преждевременную деградацию в цитоплазме), а также помогают мРНК инициировать трансляцию.
Рисунок: Пример почти полностью эукариотического транскрипта и сигналов, необходимых для добавления полиА-хвоста (еще не добавленного!). Эти сигналы будут отщеплены от транскрипта при добавлении хвоста.
Рисунок: 5′-кэп эукариотических транскриптов. Часто также модифицируются первые 2 нуклеотида (здесь фиолетовые) мРНК. Обратите внимание на нечетную связь 5 ‘на 5’.
Сплайсинг
«Сплайсинг» эукариотических мРНК относится к удалению некодирующих последовательностей (называемых интронами), встроенных в кодирующую область транскрипта (экзоны) (см. ниже). В разделе доктора Бритта сплайсинг подробно не обсуждается, за исключением того, что он рассматривается как процесс, который должен произойти до того, как мРНК полностью созреет и сможет быть доставлена в цитоплазму для трансляции.
2.6: Транскрипция распространяется по незаявленной лицензии и была создана, изменена и/или курирована LibreTexts.
- Наверх
-
- Была ли эта статья полезной?
-
- Тип изделия
- Раздел или Страница
- Показать оглавление
- нет
-
- Теги
-
- транскрипция
|
: Связаться с нами : Часто задаваемые вопросы |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Каждая четверть имеет десять недель обучения. В среднем набор студентов на курс составляет четыре курса (16 единиц).
Минимальный набор на очную форму обучения составляет 12 единиц. Оценка A+ была введена осенью 19 квартала.89; все остальные плюс и
минусовые оценки были введены осенью 1973 года.
Каждая четверть имеет десять недель обучения. В среднем набор студентов на курс составляет четыре курса (16 единиц).
Минимальный набор на очную форму обучения составляет 12 единиц. Оценка A+ была введена осенью 19 квартала.89; все остальные плюс и
минусовые оценки были введены осенью 1973 года.
 ..отлично
..отлично  Не рассчитывается в среднем балле.
Не рассчитывается в среднем балле. 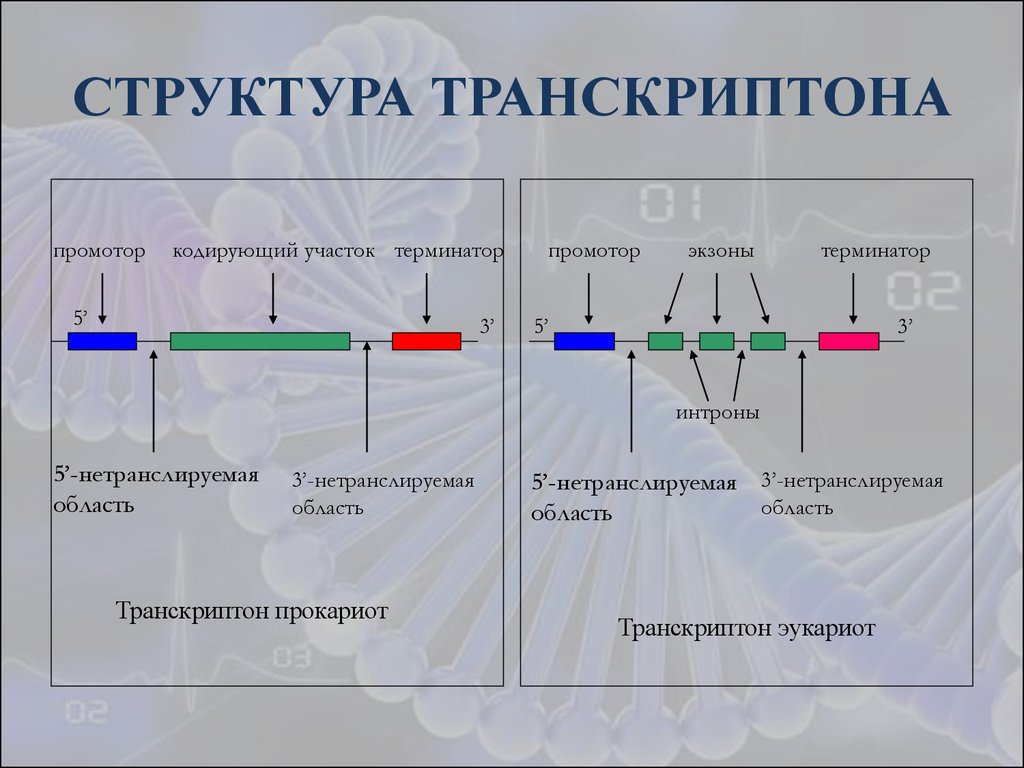
 (Студенты)
(Студенты) 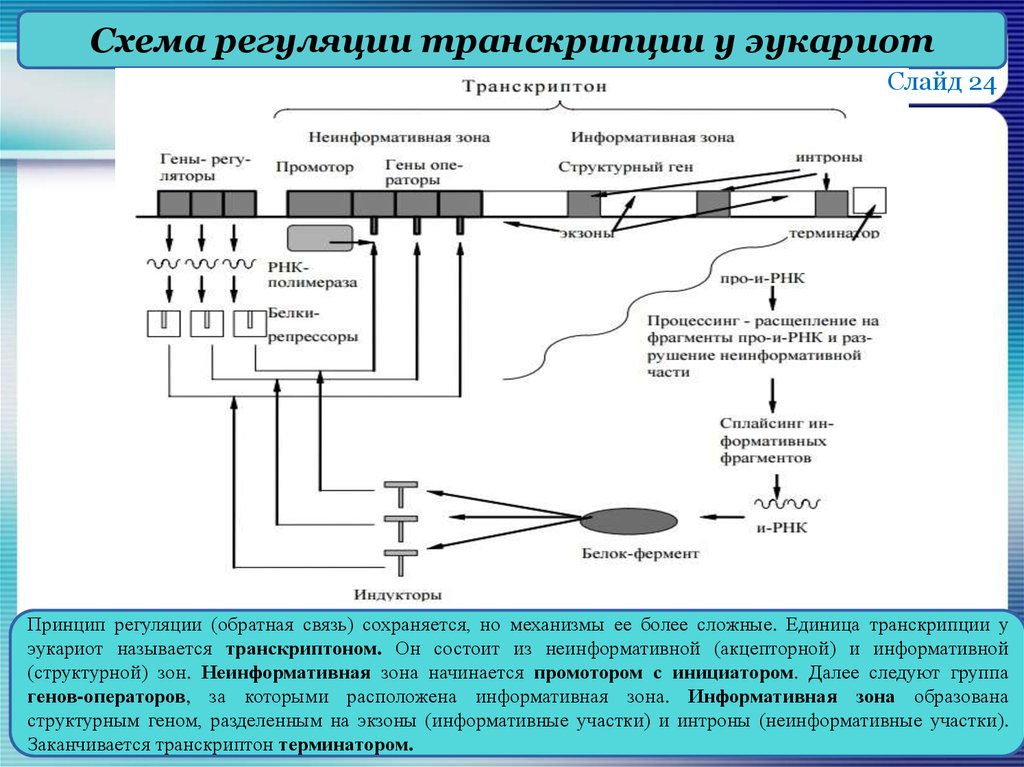


 , исходный класс C-, D+, D или D-;
до осени 1984 г. исходная оценка D+, D или D-.
, исходный класс C-, D+, D или D-;
до осени 1984 г. исходная оценка D+, D или D-. 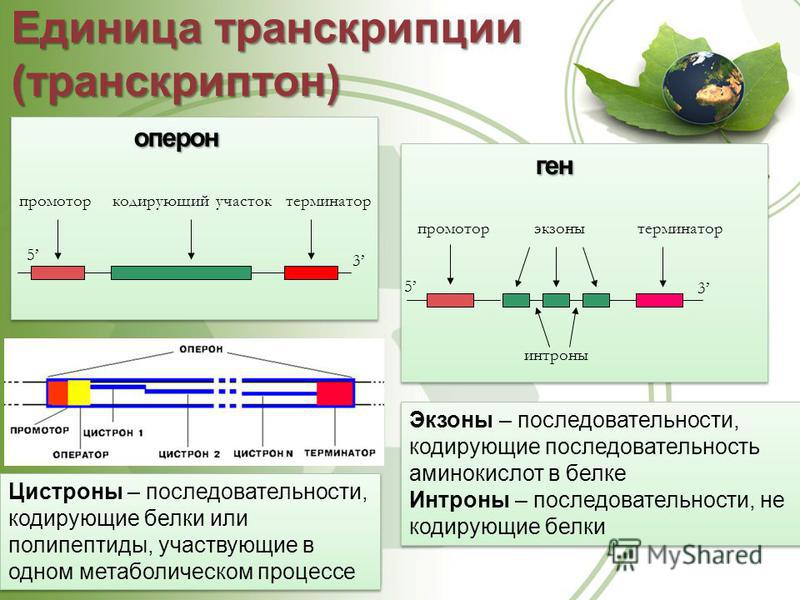
 раздел меморандумов стенограммы.
раздел меморандумов стенограммы.