Как рассчитать сопротивление провода. От каких параметров зависит сопротивление проводника. Формулы и онлайн-калькуляторы для вычисления сопротивления кабеля. Практические советы по подбору сечения провода.
Что такое электрическое сопротивление провода
Электрическое сопротивление — это свойство проводника препятствовать прохождению электрического тока. Оно измеряется в Омах и обозначается буквой R. Сопротивление провода зависит от нескольких факторов:
- Длины провода
- Площади поперечного сечения
- Материала проводника
- Температуры
Знание сопротивления необходимо для правильного расчета электрических цепей и подбора проводов нужного сечения. Слишком большое сопротивление приводит к нагреву проводов и потерям электроэнергии.
От чего зависит сопротивление проводника
Основные параметры, влияющие на сопротивление провода:
1. Длина провода
Чем длиннее провод, тем больше его сопротивление. Зависимость прямо пропорциональная — при увеличении длины в 2 раза, сопротивление также возрастает в 2 раза.
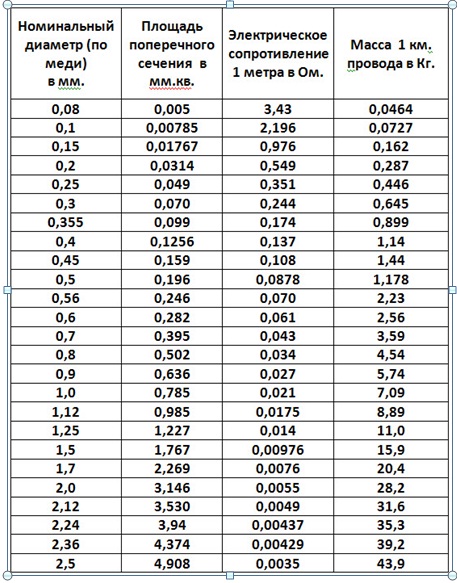
2. Площадь поперечного сечения
Сопротивление обратно пропорционально площади сечения провода. То есть чем толще провод, тем меньше его сопротивление. При увеличении сечения в 2 раза сопротивление уменьшается в 2 раза.
3. Материал проводника
Разные металлы имеют различное удельное сопротивление. Например, медь обладает меньшим сопротивлением, чем алюминий. Поэтому медные провода лучше проводят ток.
4. Температура
С повышением температуры сопротивление металлических проводников увеличивается. Это нужно учитывать при расчетах нагруженных цепей.
Формула для расчета сопротивления провода
Основная формула для вычисления сопротивления проводника:
R = ρ * L / S
где:
- R — сопротивление провода (Ом)
- ρ (ро) — удельное сопротивление материала (Ом*мм²/м)
- L — длина провода (м)
- S — площадь поперечного сечения (мм²)
Удельное сопротивление ρ зависит от материала провода. Некоторые справочные значения:
- Медь: 0,0175 Ом*мм²/м
- Алюминий: 0,028 Ом*мм²/м
- Сталь: 0,1 Ом*мм²/м
Как рассчитать сопротивление провода
Пошаговая инструкция для расчета сопротивления:

- Измерьте длину провода в метрах
- Определите площадь сечения в мм² (для круглого провода S = πd²/4, где d — диаметр)
- Узнайте удельное сопротивление материала провода
- Подставьте значения в формулу R = ρL/S
- Выполните вычисления и получите результат в Омах
Пример расчета для медного провода длиной 100 м и сечением 2,5 мм²:
R = 0,0175 * 100 / 2,5 = 0,7 Ом
Онлайн-калькуляторы сопротивления провода
Для быстрого расчета удобно использовать онлайн-калькуляторы. Вот несколько полезных ресурсов:
С помощью этих калькуляторов можно быстро рассчитать сопротивление, задав длину, сечение и материал провода.
Как подобрать сечение провода по току нагрузки
При проектировании электропроводки важно правильно подобрать сечение провода, чтобы он не перегревался под нагрузкой. Основные шаги:
- Определите максимальный ток в цепи
- По таблице допустимых токовых нагрузок выберите минимальное допустимое сечение
- Учтите возможные потери напряжения на длинных участках
- При необходимости увеличьте сечение для снижения потерь
Ориентировочная таблица выбора сечения медных проводов:

- До 16А — 1,5 мм²
- До 25А — 2,5 мм²
- До 32А — 4 мм²
- До 40А — 6 мм²
- До 63А — 10 мм²
Практические советы по расчету сопротивления проводов
Несколько рекомендаций для точных расчетов:
- Учитывайте температурный коэффициент сопротивления при нагреве проводов
- Для многожильных кабелей площадь сечения берите по сумме сечений всех жил
- При параллельном соединении проводов их сопротивления складываются по обратным величинам
- Для повышения точности используйте табличные значения удельных сопротивлений
- Учитывайте дополнительное сопротивление в местах соединений и контактов
Почему важно правильно рассчитывать сопротивление проводов
Корректный расчет сопротивления необходим по нескольким причинам:
- Позволяет избежать перегрева проводки и возгораний
- Снижает потери электроэнергии в проводах
- Обеспечивает стабильное напряжение у потребителей
- Помогает правильно выбрать защитную аппаратуру
- Позволяет оптимизировать затраты на материалы
Поэтому при проектировании электрических сетей важно уделять внимание корректному расчету сопротивления проводников.

Расчет потери напора в трубопроводе, Гидравлическое сопротивление трубы
Онлайн калькулятор позволяет определить величину гидравлического сопротивления и потери напора на участке трубопровода. Расчет гидравлического сопротивления производится на основе учебного пособия «Теоретические основы гидравлики и теплотехники», пункт 4. Для определения потери напора используются формулы Дарси — Вейсбаха.
Применяемые исходные данные: расход воды, материал и диаметр труб, длина трубопровода, коэффициенты местного сопротивления. При этом количество участков трубопровода, а также коэффициенты местного сопротивления заполняются в зависимости от конфигурации магистрали.
Результат вычислений потери напора по длине трубы может использоваться при проектировании сетей и подборе насосных агрегатов.
Скачать
теоретические основы гидравлики и теплотехники
(pdf 1. 5 Мб)
5 Мб)
+0.3
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
Шиберная задвижка
Шаровый обратный клапан
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+3. 2
2
Обратный клапан с пластинкой
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+3.2
Автоматическая трубная муфта
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
 5
5
Отвод 45°
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+0.25
Отвод 90°
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+0. 5
5
Коническое сужение
- 0
- 1
- 2
- 3
- 4
- 5
- 7
+0.1
Закругленное сужение
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+0. 1
1
Стандартное сужение
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+1
Расширение, 5°
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+0. 2
2
Расширение, 10°
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+0.5
Расширение, 15°
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+0. 85
85
Стандартное расширение
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+1
Электрический редуктор
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+0. 5
5
Дополнительные Zeta-значения
- 0
- 1
- 2
- 3
- 4
- 5
- 6
- 7
+1
Рассчитать
Расчет сопротивления кабеля
Главная » Статьи » Расчет сопротивления кабеля
Расчёт сопротивления провода
Любое вещество (при температуре выше 0К (и других обычных условиях )) обладает способностью проводить электрический ток. Величиной характеризующей способность вещества проводить электрический ток является электрическая проводимость. Величиной обратной проводимости является сопротивление. Электрическое сопротивление бывает активным и реактивным. Если через провод протекает постоянный ток то прохождению этого тока препятствует только активное сопротивление. При переменном токе активное сопротивление провода остаётся. Если провод проводящий ток имеет постоянную площадь сечения по всей его длине, состоит из однородного материала, воздействием разных полей можно пренебречь то активное сопротивление этого провода может быть рассчитано по формуле 2.
Величиной обратной проводимости является сопротивление. Электрическое сопротивление бывает активным и реактивным. Если через провод протекает постоянный ток то прохождению этого тока препятствует только активное сопротивление. При переменном токе активное сопротивление провода остаётся. Если провод проводящий ток имеет постоянную площадь сечения по всей его длине, состоит из однородного материала, воздействием разных полей можно пренебречь то активное сопротивление этого провода может быть рассчитано по формуле 2.
Рисунок 1 — Провод
Сопротивление провода зависит от его длинны, его площади поперечного сечения, удельного сопротивления ρ материала из которого состоит этот провод. Удельное сопротивление ρ любого материала (в разной степени) зависит от температуры и других факторов. Для примера рассчитаем активное электрическое сопротивление стального провода круглого сечения длинной 869мм, диаметром 0.4 мм и предположим что удельное сопротивление стали из которой сделан этот провод неизменно.
Дано:
Рассчитаем площадь поперечного сечения провода: Рассчитаем сопротивление провода: Сопротивление провода можно определить омметром (или мультиметром). Значение сопротивления данного провода определённое экспериментально примерно равно рассчитанному: Для расчёта сопротивления провода можно использовать программу:
Купить провод
electe.blogspot.ru
Как рассчитать сопротивление провода — инструкция с таблицами и формулами
В своей работе электрик часто сталкивается с вычислением различных величин и преобразований. Так для корректного подбора кабеля приходится подбирать нужное сечение. Логика выбора сечения основана на зависимости сопротивления от длины линии и площади сечения проводника. В этой статье мы рассмотрим, как выполняется расчет сопротивления провода по его геометрическим размерам.
Формула для расчета
Любые вычисления начинаются с формулы. Основной формулой для расчета сопротивления проводника является:
R=(ρ*l)/S
Где R – сопротивление в Омах, ρ – удельное сопротивление, l – длина в м, S – площадь поперечного сечения провода в мм2.
Эта формула подходит для расчета сопротивления провода по сечению и длине. Из неё следует, что в зависимости от длины изменяется сопротивление, чем длиннее – тем больше. И от площади сечения – наоборот, чем толще провод (большое сечение), тем меньше сопротивление. Однако непонятной остаётся величина, обозначенная буквой ρ (Ро).
Удельное сопротивление
Удельное сопротивление – это табличная величина, для каждого металла она своя. Она нужна для расчета и зависит от кристаллической решетки металла и структуры атомов.
Из таблицы видно, что самое меньшее сопротивление у серебра, для медного кабеля оно равняется 1,68*10-8 Ом*мм2/м. Такая размерность говорит нам, сколько приходится Ом при сечении в 1 миллиметр квадратный и длине в 1 метр.
Кстати, серебряное покрытие используется в контактах коммутационных аппаратов, автоматических выключателей, реле и прочего. Это снижает переходное контактное сопротивление, повышает срок службы и уменьшает нагрев контактов. При этом в контактах измерительной и точной аппаратуры используют позолоченные контакты из-за того, что они слабо окисляются или вообще не окисляются.
При этом в контактах измерительной и точной аппаратуры используют позолоченные контакты из-за того, что они слабо окисляются или вообще не окисляются.
У алюминия, который часто использовался в электропроводке раньше, сопротивление в 1,8 раза больше чем у меди, равняется 2,82*10-8 Ом*мм2/м. Чем больше сопротивление проводника, тем сильнее он греется. Поэтому при одинаковом сечении алюминиевый кабель может передать меньший ток, чем медный, это и стало основной причиной почему все современные электрики используют медную электропроводку. У нихрома, который используется в нагревательных приборах оно в 100 раз больше чем у меди 1,1*10-6 Ом*мм2/м.
Расчет по диаметру
На практике часто бывает так, что площадь поперечного сечения жилы не известна. Без этого значения ничего рассчитать не получится. Чтобы узнать её, нужно измерить диаметр. Если жила тонка, можно взять гвоздь или любой другой стержень, намотать на него 10 витков провода, обычной линейкой измерить длину получившейся спирали и разделить на 10, так вы узнаете диаметр.
Ну, или просто замерить штангенциркулем. Расчет сечения выполняется по формуле:
Обязательны ли расчеты?
Как мы уже сказали, сечение провода выбирают исходя из предполагаемого тока и сопротивления металла, из которого изготовлены жилы. Логика выбора заключается в следующем: сечение подбирают таким способом, чтобы сопротивление при заданной длине не приводило к значительным просадкам напряжения. Чтобы не проводить ряд расчетов, для коротких линий (до 10-20 метров) есть достаточно точные таблицы:
В этой таблице указаны типовые значения сечения медных и алюминиевых жил и номинальные токи через них. Для удобства указана мощность нагрузки, которую выдержит эта линия. Обратите внимание на разницу в токах и мощности при напряжении 380В, естественно, что это предполагается трёхфазная электросеть.
Напоследок рекомендуем просмотреть видео, на котором подробно рассказывается, как рассчитать сечение проводника, а также предоставлены примеры расчетных работ:
Расчет сопротивления провода сводится к использованию пары формул, при этом вы можете скачать готовые калькуляторы из Плэй Маркета для своего смартфона, например, «Electrodroid» или «Мобильный электрик». Эти знания пригодятся для расчетов нагревательных приборов, кабельных линий, предохранителей и даже популярных на сегодняшний день спиралей для электронных сигарет.
Материалы по теме:
samelectrik.ru
Сопротивление провода
Cуществуют два варианта применения калькуляторa
- для расчета сопротивления провода (R) если вы знаете материал провода, диаметр (d) или площадь поперечного сечения провода (S) и длину провода (L)
- чтобы найти длину провода (L) когда вы знаете материал, диаметр (d) или площадь поперечного сечения (S) и желаемое сопротивление провода (R)
резисторы параллельно
конденсаторы последовательно
катушки параллельно
колебательный контур LC
емкостнoе растягивание диапазона
однослойная катушка индуктивности
круглая индуктивная петля
квадратная индуктивная петля
частота,волна,период
кодировка конденсаторов 3-мя цифрами
кодировка резисторов (4 полоса)
кодировка резисторов (5 полос)
цветовая кодировка дросселей/катушек
555 астабильный
555 моностабильный
подбор стандартного номинала Ex
соединение стандартных номиналов E6, E12 и E24
стабилизатор напряжения LM317
конвертер дБм ⇆ мВт
конвертер мм ⇆ дюймы
сопротивление провода
закон Ома
балластный резистор для ламп ‘P’
балластный конденсатор для ламп ‘P’
балластный резистор для ламп ‘U’
балластный конденсатор для ламп ‘U’
оптическая видимость антенн
балластный резистор для светодиодов
преобразование треугольник-звезда
преобразование треугольник-звезда (емкости)
коэффициент усиления зеркальной антенны
конвертер AWG
ekalk. eu
eu
Как рассчитать сопротивление провода – советы начинающим электрикам
Доброго времени суток! Собираюсь у себя дома самостоятельно подключить электрическую варочную панель и духовку. По причине того, что слышал, что стандартная проводка может не выдержать такой напруги и станет перегреваться, решил от щитка, через дополнительный автомат прокинуть отдельные провода.
Автомат у меня уже стоит, а вот подобрать сечение провода не знаю как. Подскажите, как рассчитать сопротивление проводов под мои нужды – прокидывать придется метров 20 провода, не меньше.
Именем этого человека и была названа единица сопротивления электричества
Содержание
- Ответ читателю
- Теория и практика
- Рассчитываем сопротивление
Ответ читателю
Приветствуем Вас, к сожалению не представившийся читатель! С расчетами мы вам естественно поможем, но все- таки рекомендуем привлечь к проблеме специалиста, ведь потребуется правильно подобрать не только проводник, но и автомат. Однако если вы точно знаете, что параметры автомата подойдут, то вам осталось всего ничего…
Однако если вы точно знаете, что параметры автомата подойдут, то вам осталось всего ничего…
Теория и практика
Итак, если человек хоть немного знаком с основами электротехники, он должен знать, что чем толще провод, тем меньше сопротивление.
- Сравнить это теоретически можно с водопроводной трубой, по которой бежит вода. Если диаметр трубы достаточный, то жидкость протекает по ней, не испытывая никакого гидравлического сопротивления, и наоборот, маленькое отверстие увеличивает давление в трубе, пропускная способность падает, гидравлическое сопротивление растет.
- Также и поток электронов можно представить в виде воды, которая пытает протечь внутри провода. Однако электричество это совсем иная природа, соответственно и физические свойства у него другие.
- К чему может привести слишком высокое сопротивление? Самое банальное – это падение напряжения, в результате чего какая-нибудь лампа накаливания станет гореть тусклее, а какой-нибудь электроприбор не сможет стартовать.

- Прямым следствием прохождения мощного тока через проводник с достаточно высоким сопротивлением, будет его перегрев.
От автора! Однажды мы подключили сварочный аппарат, ну к очень плохому удлинителю, и после нескольких минут работы провод буквально загорелся. Благо короткого замыкания не произошло, но оно было весьма вероятно. Как понятно, в жилом помещении подобные ситуации недопустимы.
Рекомендуем действовать в следующей последовательности:
- Первым делом точно узнайте, какую нагрузку создают оба ваших прибора в условиях работы на максимальной мощности. Нас интересует сила тока, измеряемая в Амперах, или мощность — Ватты.
- Эти параметры вы легко отыщете в паспортах изделий.
- Если оба прибора будут запитаны от одной линии, то суммируйте полученные значения.
- Далее прибегайте к помощи таблицы, которая позволит безошибочно определить сечение провода.
На фото — таблица подбора сечения проводника
- Как видно из приведенной таблицы максимальный ток для медного провода площадью 0,5 не должен превышать 11 Ампер.
Совет! В жилых помещениях сегодня не допускается использование алюминиевых проводов. Применяют только медные.
- В принципе этими данными можно было бы и ограничиться, накинув некоторый запас, однако подобные таблицы не показывают каким должно быть максимальное сопротивление провода, то есть не учтена длина проводника. Поэтому для большей точности без расчета не обойтись.
Рассчитываем сопротивление
Все данные можно получить из таблиц
Итак, мы помним — провод толще, сопротивление меньше. Далее будет приведена инструкция, как рассчитать все точно.
- Для этого нам потребуется узнать удельное сопротивление материала проводника. В обычных сетях вы навряд ли отыщите серебряные провода, поэтому берем за основу стандартную медь. Оно составляет 0,017.
- Само же сопротивление провода рассчитывается по следующей формуле: ; где R – это сопротивление, р – удельное сопротивление проводника, l – длина провода и s – площадь его сечения.

- Предположим, что ваши печки вместе смогут нагрузить сеть на 16 Ампер, это значит, что мы можем взять провод площадью 0,75 мм2. Мы помним, что вам требуется минимум 20 метров. Итак, считаем: 0,017*20/0,75 = 0,45 Ом
- Можно воспользоваться и таблицей, но результат будет не таким точным. Мы видим, что 100 метров медного провода нужного нам сечения имеет 2,38 Ом сопротивления. Делим это значение на пять (до 20-ти метров) и получаем 0,476 Ом – разница на уровне погрешности, но все-таки.
- Из-за того, что электричество идет по двум жилам, умножаем полученное значение на 2 и получаем 0,9 Ом.
- Теперь можно рассчитать потери напряжения по формуле: dU = R*I = 0,9*16 = 14,4 Вольта.
- Переводим полученный вольтаж в процентное соотношение: 14,4В/220В*100 = 6,54%
Согласно существующим нормам допускается 5% потерь напряжения. Как видим, в нашем случае значение получилось больше, а значит, сопротивление проводника слишком большое, поэтому увеличиваем сечение провода и повторяем расчеты.
Итак, сопротивление провода мы нашли, и как видите, своими руками и головой сделать это не так уж и сложно. Дополнительно понять материал поможет прикрепленное видео. Подходите к делу с умом, ведь цена вопроса безопасность вас и вашего дома.
elektrik-a.su
Расчёт сопротивления проводника. Удельное сопротивление 8 класс онлайн-подготовка на Ростелеком Лицей
Тема 12: Электромагнитные явления. Повторение
Повторение
- Видео
- Тренажер
- Теория
Заметили ошибку?
Параметры, определяющие сопротивление проводника
На предыдущих уроках мы уже поднимали вопрос о том, каким образом электрическое сопротивление влияет на силу тока в цепи, но не обсуждали, от каких же конкретно факторов зависит сопротивление проводника. На сегодняшнем уроке мы узнаем о параметрах проводника, которые определяют его сопротивление, и узнаем, каким образом Георг Ом в своих экспериментах исследовал сопротивление проводников.
Для получения зависимости силы тока в цепи от сопротивления Ому пришлось провести огромное количество экспериментов, в которых необходимо было изменять сопротивление проводника. В связи с этим он столкнулся с проблемой изучения сопротивления проводника в зависимости от его отдельно взятых параметров. В первую очередь, Георг Ом обратил внимание на зависимость сопротивления проводника от его длины, о которой уже вскользь шла речь на предыдущих уроках. Он сделал вывод, что при увеличении длины проводника прямо пропорционально увеличивается и его сопротивление. Кроме того, было выяснено, что на сопротивление влияет еще и сечение проводника, т. е. площадь фигуры, которая получается при поперечном разрезе. При этом, чем площадь сечения больше, тем сопротивление меньше. Из этого можно сделать вывод, что чем провод толще, тем его сопротивление меньше. Все эти факты были получены опытным путем.
Он сделал вывод, что при увеличении длины проводника прямо пропорционально увеличивается и его сопротивление. Кроме того, было выяснено, что на сопротивление влияет еще и сечение проводника, т. е. площадь фигуры, которая получается при поперечном разрезе. При этом, чем площадь сечения больше, тем сопротивление меньше. Из этого можно сделать вывод, что чем провод толще, тем его сопротивление меньше. Все эти факты были получены опытным путем.
Кроме геометрических параметров на сопротивление проводника влияет еще и величина, описывающая род вещества, из которого состоит проводник. В своих опытах Ом использовал проводники, изготовленные из различных материалов. При использовании медных проводов сопротивление было каким-то одним, серебряных – другим, железных – третьим и т. д. Величину, которая характеризует род вещества в таком случае, называют удельным сопротивлением.
Таким образом, можно получить следующие зависимости для сопротивления проводника (рис. 1):
- Сопротивление прямо пропорционально длине проводника , которую в СИ измеряют в м;
- Сопротивление обратно пропорционально площади сечения проводника , которую мы будем измерять в мм2 из-за малости;
- Сопротивление зависит от удельного сопротивления вещества (читается «ро»), которое является табличной величиной и измеряется обыкновенно в .
Рис. 1. Проводник
Удельное сопротивление
Для примера приведем таблицу значений удельных сопротивлений некоторых металлов, которые получены опытным путем:
Удельное сопротивление ,
|
Медь |
0,0175 |
|
Серебро |
0,016 |
|
Железо |
0,098 |
|
Алюминий |
0,027 |
Стоит отметить, что среди хороших проводников, которыми являются металлы, наилучшими являются драгоценные металлы, при этом серебро считается самым лучшим проводником, т. к. у него наименьшее малое удельное сопротивление. Этим объясняется использование драгоценных металлов при пайке особо важных элементов в электротехнике. Из значений удельных сопротивлений веществ можно делать выводы об их практическом применении – вещества с большим удельным сопротивлением подойдут для изготовления изоляционных материалов, а с небольшим – для проводников.
Этим объясняется использование драгоценных металлов при пайке особо важных элементов в электротехнике. Из значений удельных сопротивлений веществ можно делать выводы об их практическом применении – вещества с большим удельным сопротивлением подойдут для изготовления изоляционных материалов, а с небольшим – для проводников.
Замечание. Во многих таблицах удельное сопротивление измеряют в , что связано с измерением площади в м2 в СИ.
Физический смысл удельного сопротивления – сопротивление проводника длиной 1 м и площадью поперечного сечения 1 мм2.
Формула сопротивления проводника
Формула для вычисления электрического сопротивления проводника, исходя из указанных выше рассуждений, выглядит следующим образом:
Если обратить внимание на эту формулу, то можно сделать вывод, что из нее выражается удельное сопротивление проводника, т. е., определив силу тока и напряжение на проводнике и измерив его длину с площадью поперечного сечения, можно с помощью закона Ома и указанной формулы вычислить удельное сопротивление. Затем, его значение можно сверить с данными таблицы и определить, из какого вещества изготовлен проводник.
Затем, его значение можно сверить с данными таблицы и определить, из какого вещества изготовлен проводник.
Все параметры, которые влияют на сопротивление проводников, необходимо учитывать при конструировании сложных электрических цепей, таких как линии электропередач, например. В таких проектах важно сбалансированно подобрать соотношения длин, сечений и материалов проводников для эффективного компенсирования теплового действия тока.
На следующем уроке будет рассмотрено устройство и принцип работы прибора, называющегося реостат, основной характеристикой которого является сопротивление.
Список литературы
- Генденштейн Л.Э, Кайдалов А.Б., Кожевников В.Б. Физика 8 / Под ред. Орлова В.А., Ройзена И.И. – М.: Мнемозина.
- Перышкин А.В. Физика 8. – М.: Дрофа, 2010.
- Фадеева А.А., Засов А.В., Киселев Д.Ф. Физика 8. – М.: Просвещение.
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
- Интернет-портал Exir.
 ru (Источник)
ru (Источник) - Классная физика (Источник)
Домашнее задание
- Стр. 103–106: вопросы № 1–6. Перышкин А.В. Физика 8. – М.: Дрофа, 2010.
- Длина и площадь поперечного сечения алюминиевого и железного проводов одинаковые. Какой из проводников имеет большее сопротивление?
- Какое сопротивление имеет медный провод длиной 10 м и площадью поперечного сечения 0,17 мм2?
- Какой из сплошных железных стержней разного диаметра имеет большее электрическое сопротивление? Массы стержней одинаковые.
Заметили ошибку?
Расскажите нам об ошибке, и мы ее исправим.Расчет сопротивления проводника. Удельное сопротивление
SmartCalc. Расчет утепления и точки росы для строящих свой дом. СНИП.
JavaScript отключен
К сожалению Ваш браузер не поддерживает JavaScript, или JavaScript отключен в настройках браузера.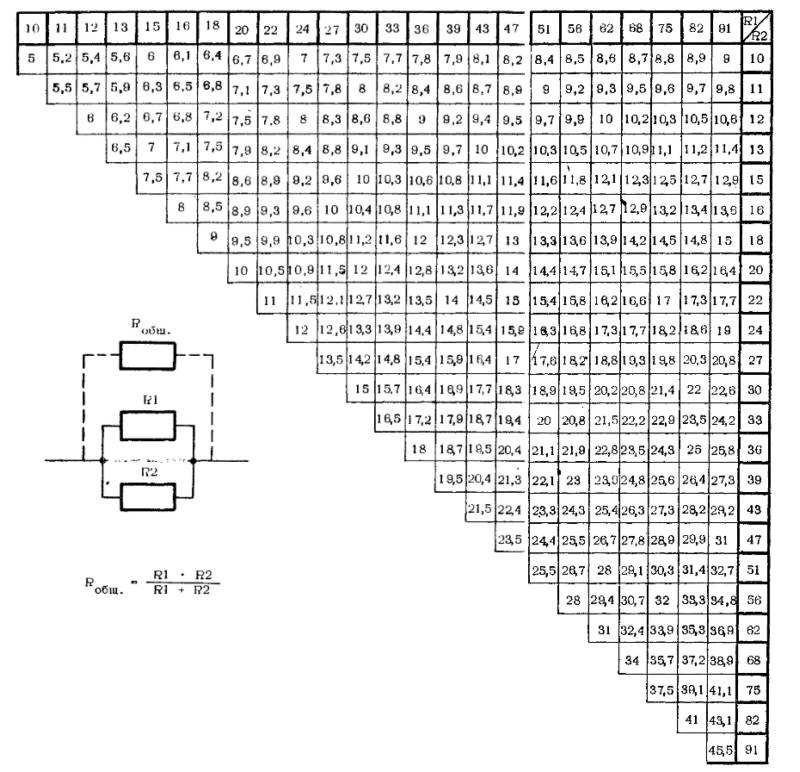
Без JavaScript и без поддержки браузером HTML5 работа ресурса невозможна. Если Вы имеете намерение воспользоваться нашим ресурсом,
включите поддержку JavaScript или обновите свой браузер.
Теплотехнический калькулятор ограждающих конструкций
Расчет утепления и точки росы для строящих свой дом
СНиП 23-02-2003
СП 23-101-2004
ГОСТ Р 54851—2011
СТО 00044807-001-2006
Старая версия калькулятора
Тепловая защита
Защита от переувлажнения
Ссылка на расчет. Отчет по результатам расчета.
Представленный теплотехнический расчет ограждающих конструкций зданий является оценочным и предназначен для предварительного выбора материалов
и проектирования конструкций.
При разработке проекта для проведения точного расчета необходимо обратиться в организацию, обладающую соответствующими полномочиями и разрешениями.
Расчет основан на российской нормативной базе:
- СНиП 23-02-2003 «Тепловая защита зданий»
- СП 23-101-2004 «Проектирование тепловой защиты зданий»
- ГОСТ Р 54851—2011 «Конструкции строительные ограждающие неоднородные. Расчет приведенного сопротивления теплопередаче»
- СТО 00044807-001-2006 «Теплозащитные свойства ограждающих конструкций зданий»
Добавьте ссылку на расчет в закладки:
Ссылка на расчет
Или скопируйте ее в буфер обмена:
Москва (Московская область, Россия)
Страна
РоссияАзербайджанАрменияБеларусьГрузияКазахстанКыргызстанМолдоваТуркменистанУзбекистанУкраинаТаджикистан
Регион
Республика АдыгеяРеспублика АлтайАлтайский крайАмурская областьАрхангельская областьАстраханская областьРеспублика БашкортостанБелгородская областьБрянская областьРеспублика БурятияВладимирская областьВолгоградская областьВологодская областьВоронежская областьРеспублика ДагестанЕврейская автономная областьЗабайкальский крайИвановская областьРеспублика ИнгушетияИркутская областьКабардино-Балкарская РеспубликаКалининградская областьРеспублика КалмыкияКалужская областьКамчатский крайКарачаево-Черкесская РеспубликаРеспублика КарелияКемеровская областьКировская областьРеспублика КомиКостромская областьКраснодарский крайКрасноярский крайРеспублика КрымКурганская областьКурская областьЛенинградская областьЛипецкая областьМагаданская областьРеспублика Марий ЭлРеспублика МордовияМосковская областьМурманская областьНенецкий АО (Архангельская область)Нижегородская областьНовгородская областьНовосибирская областьОмская областьОренбургская областьОрловская областьПензенская областьПермский крайПриморский крайПсковская областьРостовская областьРязанская областьСамарская областьСаратовская областьСахалинская областьСвердловская областьРеспублика Северная Осетия — АланияСмоленская областьСтавропольский крайТамбовская областьРеспублика ТатарстанТверская областьТомская областьТульская областьРеспублика ТываТюменская областьУдмуртская РеспубликаУльяновская областьХабаровский крайРеспублика ХакасияХанты-Мансийский автономный округ — ЮграЧелябинская областьЧеченская РеспубликаЧувашская Республика — ЧувашияЧукотский АО (Магаданская область)Республика Саха (Якутия)Ямало-Ненецкий автономный округЯрославская область
Населенный пункт
ДмитровКашираМожайскМоскваНаро-ФоминскНовомосковский АОТроицкий АОЧерусти
Температура холодной пятидневки с обеспеченностью 0. 92 92 |
-26 | ˚С |
| Продолжительность отопительного периода | 204 | суток |
| Средняя температура воздуха отопительного периода | -2.2 | ˚С |
| Относительная влажность воздуха наиболее холодного месяца | 84 | % |
| Условия эксплуатации помещения | ||
| Количество градусо-суток отопительного периода (ГСОП) | 4528.8 | °С•сут |
| Месяц | Т, ˚С | E, гПа | Месяц | Т, ˚С | E, гПа | |
|---|---|---|---|---|---|---|
| Январь | -7.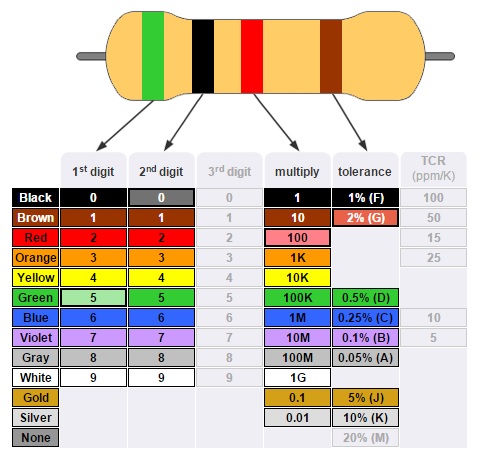 8 8 |
3.3 | Июль | 19.1 | 15.7 | |
| Февраль | -6.9 | 3.3 | Август | 17.1 | 14.6 | |
| Март | -1.3 | 4.3 | Сентябрь | 11.3 | 10.9 | |
| Апрель | 6.5 | 6. 6 |
Октябрь | 5.2 | 7.5 | |
| Май | 13.3 | 10 | Ноябрь | -0.8 | 5.2 | |
| Июнь | 17 | 13.3 | Декабрь | -5.2 | 3.9 | Год | 5.6 | 8.2 |
Жилое помещение (Стена)
Помещение Жилое помещениеКухняВаннаяНенормированноеТехническое помещение
Тип конструкции СтенаПерекрытие над проездомПерекрытие над холодным подвалом, сообщающимся с наружным воздухомПерекрытие над не отапливаемым подвалом со световыми проемами в стенахПерекрытие над не отапливаемым подвалом без световых проемов в стенахЧердачное перекрытиеПокрытие (утепленная кровля)
| Влажность в помещении* | ϕ | % | |
| Коэффициент зависимости положения наружной поверхности по отношению к наружному воздуху | n | ||
| Коэффициент теплоотдачи внутренней поверхности | α(int) | ||
| Коэффициент теплоотдачи наружной поверхности | α(ext) | ||
| Нормируемый температурный перепад | Δt(n) | °С | |
| * — параметр используется при расчете раздела «Защита от переувлажнения ограждающих конструкций» (см. закладку «Влагонакопление»). |
|||
Слои конструкции
| № | Тип | Материалы | Толщина, мм | λ | μ (Rп) | Управление | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Внутри | ||||||||||
| Снаружи | Наружный воздухВентилируемый зазор (фасад или кровля)Кровельное покрытие с вентилируемым зазором | |||||||||
Внутри: 20°С (55%) Снаружи: -10°С (85%)
Климатические параметры внутри помещения
Температура
Влажность
Климатические параметры снаружи помещения
Выбранные
Самый холодный месяц
Температура
Влажность
- Тепловая защита
- Влагонакопление
- Тепловые потери
Сопротивление теплопередаче: (м²•˚С)/Вт
| № | Тип | Толщина | Материал | λ | R | Тmax | Тmin |
|---|---|---|---|---|---|---|---|
| Термическое сопротивление Rа | |||||||
| Термическое сопротивление Rб | |||||||
| Термическое сопротивление ограждающей конструкции | |||||||
| Сопротивление теплопередаче ограждающей конструкции [R] | |||||||
| Требуемое сопротивление теплопередаче | |||||||
| Санитарно-гигиенические требования [Rс] | |||||||
| Нормируемое значение поэлементных требований [Rэ] | |||||||
| Базовое значение поэлементных требований [Rт] | |||||||
Расчет защиты от переувлажнения методом безразмерных величин
Нахождение плоскости максимального увлажнения.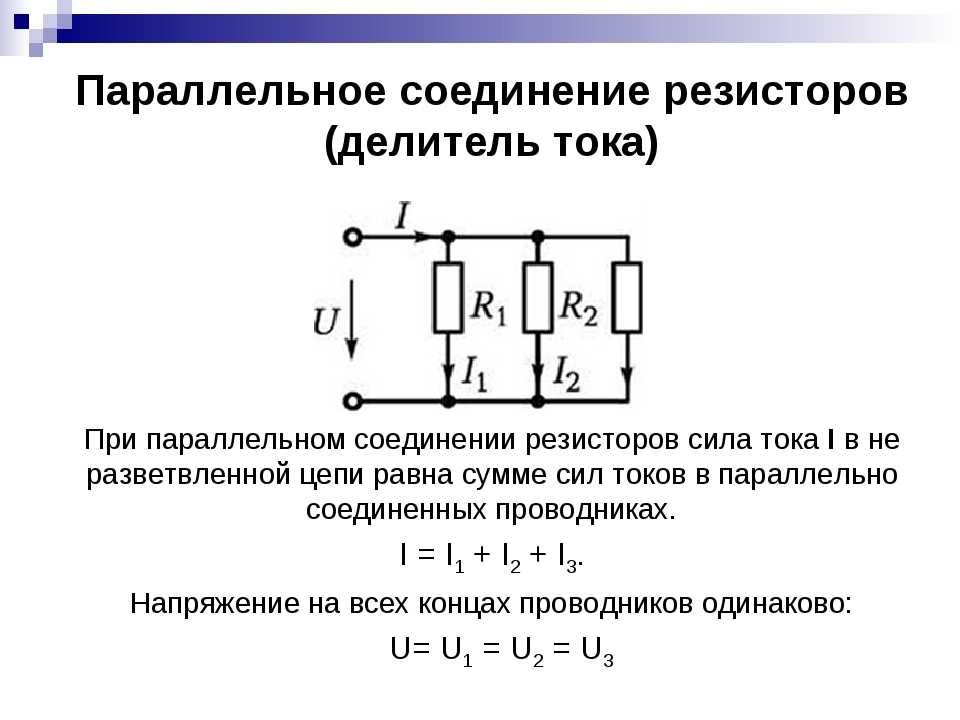
| Координата плоскости максимального увлажнения | X | 0 | мм |
| Сопротивление паропроницанию от внутренней поверхности конструкции до плоскости максимального увлажнения | Rп(в) | 0 | (м²•ч•Па)/мг |
| Сопротивление паропроницанию от плоскости максимального увлажнения до внешней поверхности конструкции | Rп(н) | 0 | (м²•ч•Па)/мг |
| Условие недопустимости накопления влаги в ограждающей конструкции за годовой период эксплуатации | Rп.тр(1) | 0 | (м²•ч•Па)/мг |
| Условие ограничения влаги в ограждающей конструкции за период с отрицательными среднемесячными температурами наружного воздуха | Rп. тр(2) тр(2) |
0 | (м²•ч•Па)/мг |
Образование конденсата в проветриваемом чердачном перекрытии или вентилируемом зазоре кровли
| Сопротивление паропроницанию конструкции | Rп | 0 | (м²•ч•Па)/мг |
| Требуемое сопротивление паропроницанию | Rп.тр | 0 | (м²•ч•Па)/мг |
Послойный расчет защиты от переувлажнения
| № | Толщина | Материал | μ | Rп | X | Rп(в) | Rп. тр(1) тр(1) |
Rп.тр(2) |
|---|
Тепловые потери через квадратный метр ограждающей конструкции
| Сопротивление теплопередаче | R | ±R, % | Q | ±Q, Вт•ч |
|---|---|---|---|---|
| Санитарно-гигиенические требования [Rс] | 0 | 0 | 0 | 0 |
| Нормируемое значение поэлементных требований [Rэ] | 0 | 0 | 0 | 0 |
| Базовое значение поэлементных требований [Rт] | 0 | 0 | 0 | 0 |
| Сопротивление теплопередаче ограждающей конструкции [R] | 0 | 0 | 0 | 0 |
| R + 10% | 0 | 0 | 0 | 0 |
| R + 25% | 0 | 0 | 0 | 0 |
| R + 50% | 0 | 0 | 0 | 0 |
| R + 100% | 0 | 0 | 0 | 0 |
Потери тепла через 1 м² за отопительный сезон
кВт•ч
Потери тепла через 1 м² за 1 час при температуре самой холодной пятидневки
Вт•ч
- Актуализация данных климатологии (СП 131.
 13330.2020)
13330.2020) - Внесены изменения в БД климатических параметров для России в соответствии с вступившим в действие СП 131.13330.2020 …
- Актуализация климатических параметров для Казахстана
- Внесены изменения в БД климатических параметров для Казахстана в соответствии с действующими нормативными документами …
- Актуализация в соответствии с норматиными документами
- Актуализированы изменения в СП 50.13330.2012 и СП 131.13330.2018 …
- Добавлены проекты
- Добавлены возможности хранения ссылок на расчеты и расчета тепловых потерь здания…
- Добавлен калькулятор тепловой защиты полов по грунту
- Калькулятор позволяет рассчитать уровень тепловой защиты и тепловые потери полов по грунту…
- Открыта группа «В контакте»
- В социальной сети «В контакте» открыта группа, посвященная проекту СмартКалк.
 ..
..
- Для исследователей и экспериментаторов
- Для экспериментаторов, исследователей и вообще всех, кому спокойно не сидится на месте, добавлен тип помещения: «Ненормированное» …
- Расчет каркасных конструкций
- Как рассчитать каркасную конструкцию?
Какие варианты каркасов можно использовать в калькуляторе?
Основной материал
Материал каркаса или швов
Материал:
Плотность ρ:
кг/м³
Удельная теплоемкость (c):
кДж/(кг•°С)
Коэффициент теплопроводности для условий А λ(А):
Вт/(м•°С)
Коэффициент теплопроводности для условий Б λ(Б):
Вт/(м•°С)
Коэффициент паропроницаемости μ:
мг/(м•ч•Па)
Предельно допустимое приращение расчетного массового отношения влаги в материале ограждающей конструкции Δwcp:
%
Сопротивление паропроницанию Rп:
(м²•ч•Па)/мг
Вставить после:
Расчет сопротивления
1. Обмотка катушки выполнена из медного провода диаметром d = 0,815 мм. Провод покрыт эмалевой изоляцией. Размеры катушки: длина L = 125 мм, внутренний диаметр обмотки , внешний диаметр (рис. 2). От каркаса катушки обмотку отделяет картон толщиной Δ=0,55 мм.
Обмотка катушки выполнена из медного провода диаметром d = 0,815 мм. Провод покрыт эмалевой изоляцией. Размеры катушки: длина L = 125 мм, внутренний диаметр обмотки , внешний диаметр (рис. 2). От каркаса катушки обмотку отделяет картон толщиной Δ=0,55 мм.
Определить электрическое сопротивление обмотки при температуре 20 °С, считая для меди
Решение:
Для определения электрического сопротивления обмотки необходимо знать, помимо удельного сопротивления, площадь поперечного сечения S и длину l проволоки. Так как провод круглый, то
Длину l проволоки обмотки определим, витков катушки и длину среднего витка:
Число горизонтальных слоев (в направлении диаметров) равно ширине окна каркаса , разделенной на диаметр проволоки:
Число вертикальных рядов (в направлении длины) равно высоте окна каркаса , разделенной на диаметр проволоки:
Число витков катушки равно произведению чисел горизонтальных слоев и вертикальных рядов :
Длина проволоки катушки
Электрическое сопротивление обмотки
2. В схеме (рис. 3) переключатель служит для присоединения вольтметра к зажимам источника (положение 1) и для замыкания цепи (положение 2). Таким путем получены показания вольтметра 2,1 В и амперметра 1 А.
В схеме (рис. 3) переключатель служит для присоединения вольтметра к зажимам источника (положение 1) и для замыкания цепи (положение 2). Таким путем получены показания вольтметра 2,1 В и амперметра 1 А.
Чему равно внутреннее сопротивление источника, если внеишее сопротивление r = 2 Ом?
Решение:
При положении 1 переключателя источник разомкнут (I = 0), а вольтметр измеряет разность потенциалов между зажимами источника, равную Е = 2,1 В.
При положении 2 переключателя вольтметр отсоединяется от источника и последний замыкается на сопротивление r внешней цепи, ток в которой I=1 A.
На основании закона Ома сопротивление всей цепи
Так как внешнее сопротивление r=2 Ом, то внутреннее сопротивление источника
Описанный способ определения внутреннего сопротивления довольно приближенный, так как здесь было принято, что:
а) сопротивление вольтметра очень большое и, следовательно, ток в цепи вольтметра близок к нулю;
б) сопротивление амперметра равно нулю.
3. В схеме (рис. 4) показание вольтметра равно 2 В при замкнутом рубильнике и 1,8 В при замкнутых рубильниках . Пренебрегая током, проходящим через вольтметр, определить внутреннее сопротивление источника, если r=4 Ом и э.д.с. источника постоянна.
Решение:Сопротивление внешней цепи при замкнутом рубильнике равно и уменьшается вдвое после включения рубильника :
Уменьшение напряжения с 2 до 1,8 В после замыкания рубильника объясняется увеличением тока и пропорциональной ему потерей напряжения внутри источника.
По условию, э.д.с. источника постоянна, поэтому можно написать
Неизвестные токи определяют по закону Ома, после чего находят и внутреннее сопротивление источника:
Подставив найденные значения токов в уравнение (1), получим
Постоянная э. д. с. источника
Из результатов решения задачи видно, что уменьшение напряжения между зажимами источника не связано простой зависимостью с уменьшением сопротивления внешней цепи. Действительно, в данной задаче сопротивление внешней цепи уменьшалось в два раза , а напряжение — на 10%:
Действительно, в данной задаче сопротивление внешней цепи уменьшалось в два раза , а напряжение — на 10%:
4. Определить диаметр и длину нихромовой проволоки для нагревательного элемента электрического кипятильника (127 В, 2,5 А), допуская плотность тока и принимая удельное сопротивление нихрома в нагретом состоянии
Решение:
Площадь поперечного сечения проволоки определяем по току и плотности тока:
Диаметр проволоки
Электрическое сопротивление проволоки на основании закона Ома
Длина проволоки для нагревательного элемента
5. В штепсельных магазинах сопротивлений отдельные сопротивления выводят из цепи включением штепселя (рис. 8).
Целесообразно — ли создать конструкцию, в которой при включении штепселя сопротивление r будет отсоединяться?
Решение:
Очень малое сопротивление пластин и штепселя вместе с сопротивлением переходных контактов включено параллельно сопротивлению r.
Следовательно, эквивалентное сопротивление
Так как множитель меньше единицы, то сопротивление меньше сопротивления .
Отсюда следует, что остающееся присоединенным к пластинам сопротивление r помогает свести к нулю сопротивление на рассматриваемом участке, представляя еще один путь для электрического тока. Таким образом, изменение существующей конструкции нецелесообразно. Например, пусть сопротивление двух переходных контактов на пути от одной пластины к другой равно . Сопротивлением пластин штепселя пренебрежем. Тогда, если r=1000 ом, получим
т. е. сопротивление меньше сопротивления . Сопротивление r больше сопротивления примерно в 500000 раз.
6. Ламповый реостат состоит из шести ламп мощностью по 60 Вт, соединенных параллельно.
Определить электрическое сопротивление реостата при различном числе включенных ламп, если напряжение сети 120 В.
Решение:
При одинаковом сопротивлении r у n пассивных элементов цепи (т. е. элементов цепи, не содержащих э.д.с), включенных параллельно, эквивалентное сопротивление
Сопротивление r каждой лампы можно определить по формуле
откуда
Поэтому можно составить табл. 4.
4.
Таблица 4
240 | ||||||
1 | 2 | 3 | 4 | 5 | 6 | |
240 | 120 | 80 | 60 | 48 | 40 | |
Из табл. 4 видно, что более плавно изменяется сопротивление реостата при большом числе параллельно включенных ламп.
7. Три одинаковые обмотки статора трехфазного электродвигателя соединены треугольником (составляют замкнутый контур, рис.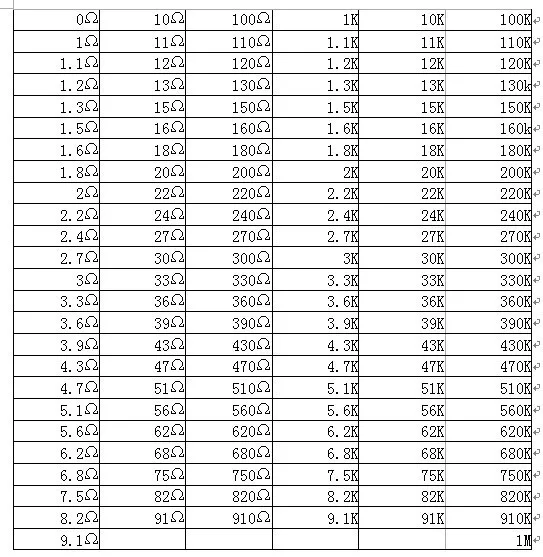 9). При измерении их сопротивления воспользовались зажимами, выведенными от двух точек контура, причем показание амперметра было 1 А, вольтметра 2 В.
9). При измерении их сопротивления воспользовались зажимами, выведенными от двух точек контура, причем показание амперметра было 1 А, вольтметра 2 В.
Определить сопротивление каждой из обмоток треугольника.
Решение:
По условию, электрические сопротивления обмоток одинаковы. Для тока в проводе обмотки образуют две параллельные ветви, в одной из которых сопротивление r, а в другой 2r (две обмотки соединены последовательно).
Эквивалентное сопротивление определяем по формуле для двух параллельных ветвей:
Кроме того, это сопротивление на основании закона Ома
Итак,
откуда r=3 Ом, т. е. сопротивление одной обмотки в 3/2 раза больше отношения показания вольтметра к показанию амперметра в рассматриваемой схеме.
8. Батарея из 63 кислотных аккумуляторов емкостью , соединенных последовательно, заряжается от источника постоянного напряжения 60 В, причем составляют три равных параллельных группы по 21 аккумулятору в каждой группе.
Определить величину сопротивления реостата для каждой из трех групп, если в конце зарядки напряжение аккумуляторов составляет 2,6 В, а в начале зарядки — 1,85 В; зарядка аккумуляторов производится током, соответствующим 8-часовой зарядке.
Решение:
Э.д.с. каждой группы аккумуляторов равна:
в начале зарядки
в конце зарядки
При напряжении заряжающего источника 60 В требуется «погасить» в реостате в начале зарядки , в конце зарядки , так как в этом процессе ток проходег через батарею в направлении, встречном э. д. с.
Ток 8-часовой зарядки составляет
Сопротивления реостата:
Расчет и подбор сопротивления токоограничивающего резистора для светодиода
Светодиод – это полупроводниковый элемент, который применяется для освещения. Применяется в фонарях, лампах, светильниках и других осветительных приборах. Принцип его работы заключается в том, что при протекании тока через светоизлучающий диод происходит высвобождение фотонов с поверхности материала полупроводника, и диод начинает светиться.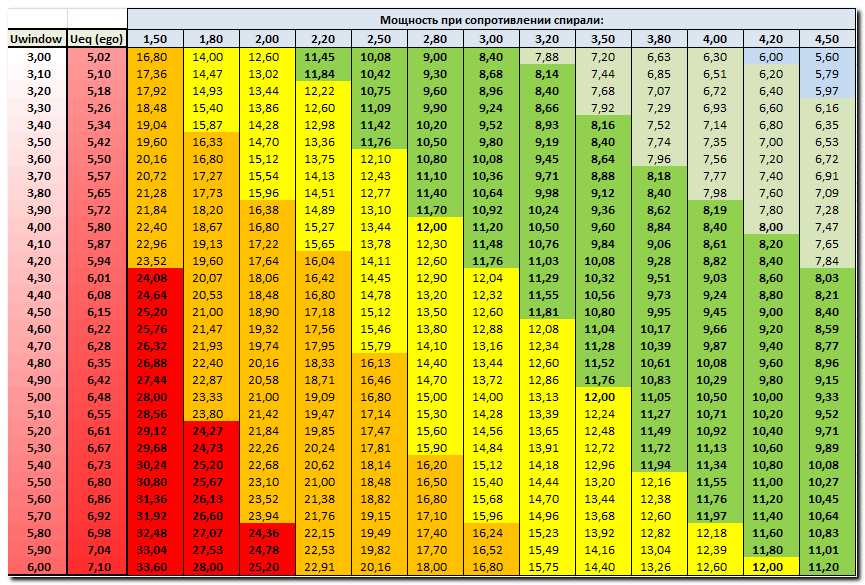
Содержание статьи
- 1 Расчет резистора для светодиода
- 1.1 Вычисление номинала сопротивления
- 1.2 Подбор мощности резистора
- 2 Пример расчета
- 3 Онлайн-калькулятор светодиодов
Расчет резистора для светодиода
Надежная работа светодиода зависит от тока, протекающего через него. При заниженных значениях, он просто не будет светить, а при превышении значения тока – характеристики элемента ухудшатся, вплоть до его разрушения. При этом говорят – светодиод сгорел. Для того чтобы исключить возможность выхода из строя этого полупроводника необходимо подобрать в цепь с включенным в нее, резистором. Он будет ограничивать ток в цепи на оптимальных значениях.
Вычисление номинала сопротивления
Для работы радиоэлемента на него нужно подать питание. По закону Ома, чем больше сопротивление отрезка цепи, тем меньший ток по нему протекает. Опасная ситуация возникает, если в схеме течет больший ток, чем положено, так как каждый элемент не выдерживает большей токовой нагрузки.
Сопротивление светодиода является нелинейным. Это значит, что при изменении напряжения, подаваемого на этот элемент, ток, протекающий через него, будет меняться нелинейно. Убедиться в этом можно, если найти вольт — амперную характеристику любого диода, в том числе и светоизлучающего. При подаче питания ниже напряжения открытия p — n перехода, ток через светодиод низкий, и элемент не работает. Как только этот порог превышен, ток через элемент стремительно возрастает, и он начинает светиться.
Если источник питания соединять непосредственно со светодиодом, диод выйдет из строя, так как не рассчитан на такую нагрузку. Чтобы этого не произошло – нужно ограничить ток, протекающий через светодиод балластным сопротивлением, или произвести понижение напряжения на важном для нас полупроводнике.
Рассмотрим простейшую схему подключения (рисунок 1). Источник питания постоянного тока подключается последовательно через резистор к нужному светодиоду, характеристики которого нужно обязательно узнать. Сделать это можно в интернете, скачав описание (информационный лист) на конкретную модель, или найдя нужную модель в справочниках. Если найти описание не представляется возможным, можно приблизительно определить падение напряжения на светодиоде по его цвету:
- Инфракрасный — до 1.9 В.
- Красный – от 1.6 до 2.03 В.
- Оранжевый – от 2.03 до 2.1 В.
- Желтый – от 2.1 до 2.2 В.
- Зеленый – от 2.2 до 3.5 В.
- Синий – от 2.5 до 3.7 В.
- Фиолетовый – 2.8 до 4 В.
- Ультрафиолетовый – от 3.1 до 4.4 В.
- Белый – от 3 до 3.7 В.
Рисунок 1 – схема подключения светодиода
Ток в схеме можно сравнить с движением жидкости по трубе. Если есть только один путь протекания, то сила тока (скорость течения) во всей цепи будет одинакова. Именно так происходит в схеме на рисунке 1. Согласно закону Кирхгоффа, сумма падений напряжения на всех элементах, включенных в цепь протекания одного тока, равно ЭДС этой цепи (на рисунке 1 обозначено буквой Е). Отсюда можно сделать вывод, что напряжение, падающее на токоограничивающем резисторе должно быть равным разности напряжения питания и падения его на светодиоде.
Именно так происходит в схеме на рисунке 1. Согласно закону Кирхгоффа, сумма падений напряжения на всех элементах, включенных в цепь протекания одного тока, равно ЭДС этой цепи (на рисунке 1 обозначено буквой Е). Отсюда можно сделать вывод, что напряжение, падающее на токоограничивающем резисторе должно быть равным разности напряжения питания и падения его на светодиоде.
Так как ток в цепи должен быть одинаковым, то и через резистор, и через светодиод ток получается одним и тем же. Для стабильной работы полупроводникового элемента, увеличения его показателей надежности и долговечности, ток через него должен быть определенных значений, указанных в его описании. Если описание найти невозможно, можно принять приблизительное значение тока в цепи 10 миллиампер. После определения этих данных уже можно вычислить номинал сопротивления резистора для светодиода. Он определяется по закону Ома. Сопротивление резистора равно отношению падения напряжения на нем к току в цепи. Или в символьной форме:
R = U (R)/ I,
где, U (R) — падение напряжения на резисторе
I – ток в цепи
Расчет U (R) на резисторе:
U (R) = E – U (Led )
где, U (Led) — падение напряжения на светодиодном элементе.
С помощью этих формул получится точное значение сопротивления резистора. Однако, промышленностью выпускаются только стандартные значения сопротивлений так называемые ряды номиналов. Поэтому после расчета придется сделать подбор существующего номинала сопротивления. Подобрать нужно чуть больший резистор, чем получилось в расчете, таким образом, получится защита от случайного превышения напряжения в сети. Если подобрать близкий по значению элемент сложно, можно попробовать соединить два резистора последовательно, или параллельно.
Подбор мощности резистора
Если подобрать сопротивление меньшей мощности, чем нужно в схеме, оно просто выйдет из строя. Расчет мощности резистора довольно прост, нужно падение напряжения на нём умножить на ток, протекающий в этой цепи. После чего нужно выбрать сопротивление с мощностью, не меньшей рассчитанной.
Пример расчета
Имеем напряжение питания 12В, зеленый светодиод. Нужно рассчитать сопротивление и мощность токоограничивающего резистора. Падение напряжения на нужном нам зеленом светодиоде равно 2,4 В, номинальный ток 20 мА. Отсюда вычисляем напряжение, падающее на балластном резисторе.
Нужно рассчитать сопротивление и мощность токоограничивающего резистора. Падение напряжения на нужном нам зеленом светодиоде равно 2,4 В, номинальный ток 20 мА. Отсюда вычисляем напряжение, падающее на балластном резисторе.
U (R) = E – U (Led) = 12В – 2,4В = 9,6В.
Значение сопротивления:
R = U (R)/ I = 9,6В/0,02А = 480 Ом.
Значение мощности:
P = U (R) ⋅ I = 9,6В ⋅ 0,02А = 0,192 Вт
Из ряда стандартных сопротивлений выбираем 487 Ом (ряд Е96), а мощность можно выбрать 0,25 Вт. Такой резистор нужно заказать.
В том случае, если нужно подключить несколько светодиодов последовательно, подключать их к источнику питания можно также с помощью только одного резистора, который будет гасить избыточное напряжение. Его расчет производится по указанным выше формулам, однако, вместо одного прямого напряжения U (Led) нужно взять сумму прямых напряжений нужных светодиодов.
Если требуется подключить несколько светоизлучающих элементов параллельно, то для каждого из них требуется рассчитать свой резистор, так как у каждого из полупроводников может быть свое прямое напряжение.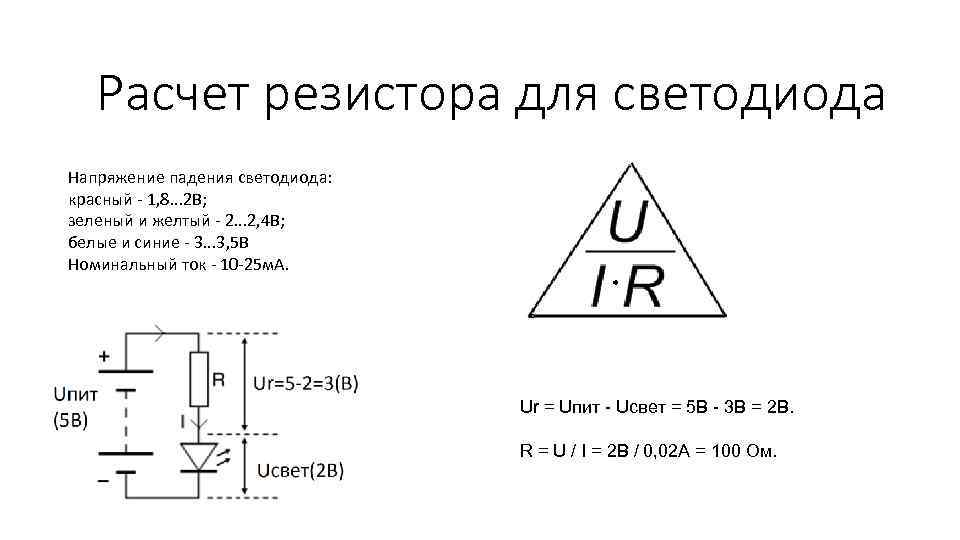 Вычисления для каждой цепи в таком случае аналогичны расчету одного резистора, так как все они подключаются параллельно к одному источнику питания, и его значение для расчета каждой цепи одно и то же.
Вычисления для каждой цепи в таком случае аналогичны расчету одного резистора, так как все они подключаются параллельно к одному источнику питания, и его значение для расчета каждой цепи одно и то же.
Этапы вычисления
Чтобы сделать правильные вычисления, необходимо выполнить следующее:
- Выяснение прямого напряжения и тока светодиода.
- Расчет падения напряжения на нужном резисторе.
- Расчет сопротивления резистора.
- Подбор сопротивления из стандартного ряда.
- Вычисление и подбор мощности.
Онлайн-калькулятор светодиодов
Этот несложный расчет можно сделать самому, но проще и эффективнее по времени воспользоваться калькулятором для расчета резистора для светодиода. Если ввести такой запрос в поисковик, найдется множество сайтов, предлагающих автоматизированный подсчет. Все необходимые формулы в этот инструмент уже встроены и работают мгновенно. Некоторые сервисы сразу предлагают также и подбор элементов. Нужно будет только выбрать наиболее подходящий калькулятор для расчета светодиодов, и, таким образом, сэкономить свое время.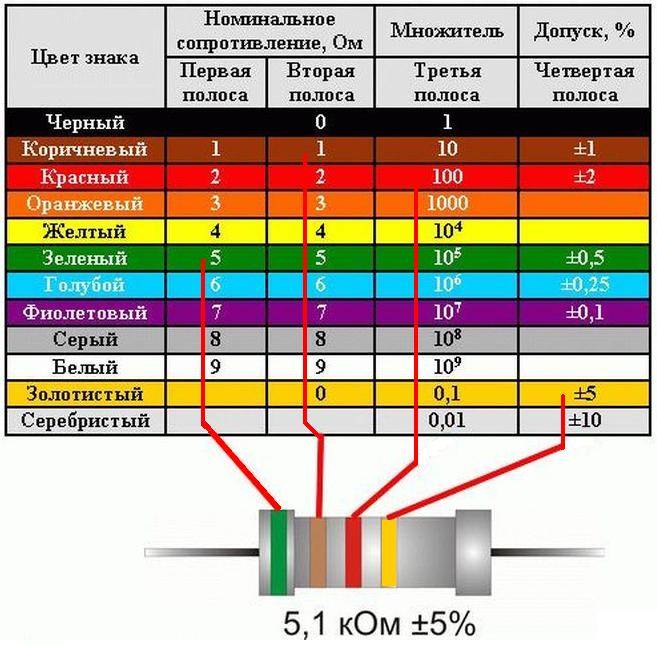
Калькулятор светодиодов онлайн – не единственное средство для экономии времени в вычислениях. Расчет транзисторов, конденсаторов и других элементов для различных схем уже давно автоматизирован в интернете. Остается только грамотно воспользоваться поисковиком для решения этих задач.
Светодиоды – оптимальное решение для многих задач освещения дома, офиса и производства. Обратите внимание на светильники Ledz. Это лучшее соотношение цены и качества осветительной продукции, используя их, вам не придется самим делать расчеты и собирать светотехнику.
Отбор на устойчивость к противомикробным препаратам снижается при внедрении в естественное микробное сообщество
Отбор на устойчивость к противомикробным препаратам снижается при внедрении в естественное микробное сообщество
Скачать PDF
Скачать PDF
- Артикул
- Открытый доступ
- Опубликовано:
- Ули Клюмпер ORCID: orcid.org/0000-0002-4169-6548 1,2 ,
- Марио Рекер ORCID: orcid.org/0000-0001-9489-1315 3 ,
- Lihong Zhang 2 ,
- Xiaole Yin 4 ,
- Tong Zhang 4 ,
- Angus Buckling 1 &
- …
- Уильям Х. Гэйз 2
Журнал ISME том 13 , страницы 2927–2937 (2019)Процитировать эту статью
-
10 тыс.
 обращений
обращений -
49 цитирований
-
78 Альтметрический
-
Сведения о показателях
Субъекты
- Антибиотики
- Бактериальная эволюция
- Микробная экология
Abstract
Устойчивость к антибиотикам стала одной из самых серьезных глобальных угроз для общественного здравоохранения. В экспериментах с отдельными видами селекция на устойчивость к антибиотикам происходит при очень низких концентрациях антибиотиков. Однако неясно, насколько далеко эти результаты могут быть экстраполированы на естественную среду, где виды встроены в сложные сообщества. Мы соревновались с изогенными штаммами Escherichia coli , отличающиеся исключительно одной хромосомной детерминантой резистентности, в присутствии и отсутствии микробного сообщества фекалий свиней в градиенте концентрации антибиотика для двух соответствующих антибиотиков: гентамицина и канамицина. Мы показываем, что минимальная избирательная концентрация была увеличена более чем на порядок для обоих антибиотиков при внедрении в сообщество. Мы определили два основных механизма, ответственных за увеличение минимальной селективной концентрации: увеличение стоимости устойчивости и защитный эффект сообщества для восприимчивого фенотипа. Эти результаты имеют значение для нашего понимания эволюции и селекции устойчивости к антибиотикам и могут дать информацию для будущих усилий по оценке рисков, связанных с концентрациями антибиотиков.
Мы соревновались с изогенными штаммами Escherichia coli , отличающиеся исключительно одной хромосомной детерминантой резистентности, в присутствии и отсутствии микробного сообщества фекалий свиней в градиенте концентрации антибиотика для двух соответствующих антибиотиков: гентамицина и канамицина. Мы показываем, что минимальная избирательная концентрация была увеличена более чем на порядок для обоих антибиотиков при внедрении в сообщество. Мы определили два основных механизма, ответственных за увеличение минимальной селективной концентрации: увеличение стоимости устойчивости и защитный эффект сообщества для восприимчивого фенотипа. Эти результаты имеют значение для нашего понимания эволюции и селекции устойчивости к антибиотикам и могут дать информацию для будущих усилий по оценке рисков, связанных с концентрациями антибиотиков.
Введение
Появление и распространение генов устойчивости к противомикробным препаратам (УПП) у бактериальных патогенов было определено Всемирной организацией здравоохранения как одна из основных угроз для здоровья человека [1]. В то время как гены AMR были обнаружены в образцах древней вечной мерзлоты [2], антропогенное использование антибиотиков вызвало быстрый рост их распространенности [3]. Большой объем теории и работы in vitro определили роль экологического контекста, такого как режим лечения и неоднородность окружающей среды, в динамике генов AMR [4,5,6,7]. Однако в большей части этой работы явно не рассматривается важнейшая особенность микробной экологии: микробы обычно встроены в сложные сообщества взаимодействующих видов. Это всегда имеет место в микробиомах человека и домашнего скота, в которых отбор, вызванный антибиотиками, вероятно, будет особенно сильным [8]. Здесь мы объединяем эксперименты и теорию, чтобы определить, как на отбор для УПП влияет присутствие других видов, происходящих из естественного кишечного микробного сообщества. Основное внимание в этом исследовании уделяется отбору по ранее существовавшим генам устойчивости внутри основного вида, а не отбору по вариациям de novo, возникающим в результате спонтанных мутаций или приобретенным в результате горизонтального переноса генов от другого вида.
В то время как гены AMR были обнаружены в образцах древней вечной мерзлоты [2], антропогенное использование антибиотиков вызвало быстрый рост их распространенности [3]. Большой объем теории и работы in vitro определили роль экологического контекста, такого как режим лечения и неоднородность окружающей среды, в динамике генов AMR [4,5,6,7]. Однако в большей части этой работы явно не рассматривается важнейшая особенность микробной экологии: микробы обычно встроены в сложные сообщества взаимодействующих видов. Это всегда имеет место в микробиомах человека и домашнего скота, в которых отбор, вызванный антибиотиками, вероятно, будет особенно сильным [8]. Здесь мы объединяем эксперименты и теорию, чтобы определить, как на отбор для УПП влияет присутствие других видов, происходящих из естественного кишечного микробного сообщества. Основное внимание в этом исследовании уделяется отбору по ранее существовавшим генам устойчивости внутри основного вида, а не отбору по вариациям de novo, возникающим в результате спонтанных мутаций или приобретенным в результате горизонтального переноса генов от другого вида.
Недавние экспериментальные исследования показывают, что селекция генов УПП в сложных сообществах происходит при концентрациях антибиотиков (минимальная избирательная концентрация; МСК), которые намного ниже тех, которые предотвращают рост восприимчивых бактерий (минимальная ингибирующая концентрация; МИК) [9, 10]; как было показано ранее в пределах одного вида in vitro [6, 7, 11]. Однако неясно, как присутствие других микробных видов влияет на МСК. Хотя точное влияние других видов, вероятно, зависит от контекста, мы предполагаем, что присутствие сообщества обычно увеличивает MSC. Исследования отдельных видов показывают, что резистентные клетки могут обеспечивать защиту восприимчивых посредством как внутриклеточной, так и внеклеточной деградации антибиотиков [12,13,14], тем самым повышая относительную приспособленность восприимчивых штаммов и, следовательно, МСК. Однако выделяемые метаболиты могут как усиливать, так и снижать эффективность антибиотиков, тем самым уменьшая или увеличивая количество МСК [15, 16]. Кроме того, любые затраты, связанные с УПП, могут быть увеличены за счет усиления конкуренции за ресурсы, как это, например, наблюдалось в отношении устойчивости мух к паразитоидам [17] и бактерий к вирусам [18].
Кроме того, любые затраты, связанные с УПП, могут быть увеличены за счет усиления конкуренции за ресурсы, как это, например, наблюдалось в отношении устойчивости мух к паразитоидам [17] и бактерий к вирусам [18].
Чтобы изучить потенциальное влияние контекста сообщества на селекцию УПП, мы сравнили изогенные штаммы Escherichia coli MG1655, отличающиеся исключительно одной хромосомной детерминантой устойчивости, в присутствии и в отсутствие микробного сообщества в градиенте двух разных аминогликозидных антибиотиков. , канамицин (Kn) и гентамицин (Gm). Мы внедрили штамм Escherichia coli ( E. coli ), обычно встречающийся в анаэробном пищеварительном тракте теплокровных млекопитающих [19].], внутри фекального сообщества свиней в экспериментальных анаэробных варочных котлах в попытке частично имитировать среду кишечника. Мы дополнительно использовали метагеномный анализ, типирование сообщества (ген 16S рРНК) и математическое моделирование, чтобы получить представление о механизмах, лежащих в основе влияния сообщества на отбор AMR.
Материалы и методы
Фекалии свиней
Фекалии свиней были собраны у четырех черных свиней без предшествующего воздействия антибиотиками в апреле 2016 года на ферме Healey’s Cornish Cyder (Пенхаллоу, Корнуолл, Соединенное Королевство). Двести граммов фекалий от каждой свиньи объединяли, смешивали с 400 мл стерильного глицерина и 1,8 г/л раствора NaCl. Смесь гомогенизировали в течение 3 мин в мельнице Retsch Knife Gm300 (Retsch GmbH, Haan, Германия) при 2000 оборотах в минуту (об/мин), фильтровали через сито (размер ячеек ~1 мм 9 ).0020 2 ), центрифугировали при 500 об/мин в течение 60 с при 4°С, жидкую надосадочную фракцию собирали и замораживали при -80°С в качестве инокулята.
Экстракт фекалий свиней
Двести граммов фекалий от каждой свиньи объединяли, смешивали с 800 мл стерильного раствора NaCl с концентрацией 0,9 г/л. Смесь гомогенизировали в течение 3 минут в мельнице Retsch Knife Gm300 (Retsch GmbH, Haan, Germany) при 2000 об/мин, фильтровали через сито (размер ячеек ~1 мм 2 ) и собирали жидкую фракцию. Затем экстракт центрифугировали (3500 об/мин, 20 мин, 4°С), супернатант собирали и автоклавировали (121°С, 20 мин). Автоклавированный экстракт снова центрифугировали (3500 об/мин, 20 мин, 4°С), супернатант собирали и использовали в качестве питательной добавки.
Затем экстракт центрифугировали (3500 об/мин, 20 мин, 4°С), супернатант собирали и автоклавировали (121°С, 20 мин). Автоклавированный экстракт снова центрифугировали (3500 об/мин, 20 мин, 4°С), супернатант собирали и использовали в качестве питательной добавки.
Штаммы
Основной вид, E. coli MG1655, был хромосомно мечен кассетой гена Tn 7 , кодирующей конститутивную красную флуоресценцию, экспрессируемую геном mCherry [20], чтобы убедиться, что 0 E0 coli 0 E0 coli 0 E0. могут быть обнаружены и отличены от других членов сообщества после конкуренции на основе красной флуоресценции. Ранее был создан устойчивый к Kn красный флуоресцентный вариант, содержащий ген устойчивости aph(3 ‘)-IIb, кодирующий аминогликозид-3’-фосфотрансферазу [21, 22].
Для создания мутанта, устойчивого к Gm, штамм был дополнительно мечен посредством электропорации плазмидой доставки pBAM, содержащей систему доставки mini-Tn 5 [23, 24] для гена устойчивости к Gm aacC1 , кодирующего Gm 3′- N -ацетилтрансфераза [25]. Успешные клоны были проверены на устойчивость к Gm (30 мкг/мл), и для выбранного клона была измерена кривая роста одного штамма в лизогенном бульоне (LB), чтобы убедиться, что стоимость гена устойчивости ниже 10% по сравнению с чувствительным штаммом. для обеспечения конкурентоспособности.
Успешные клоны были проверены на устойчивость к Gm (30 мкг/мл), и для выбранного клона была измерена кривая роста одного штамма в лизогенном бульоне (LB), чтобы убедиться, что стоимость гена устойчивости ниже 10% по сравнению с чувствительным штаммом. для обеспечения конкурентоспособности.
Конкурентные эксперименты
Конкурентные эксперименты, а также начальный рост штаммов очаговых видов проводили в колбах с сывороткой объемом 25 мл с пробками из бутилкаучука. В качестве питательной среды 10 мл стерильной среды LB с добавлением 0,1 % экстракта свиных фекалий, 50 мг/л цистеина-HCl в качестве скруббера кислорода и 1 мг/л резазурина в качестве окислительно-восстановительного индикатора для обеспечения анаэробных условий [26] добавляли к каждому реактор, нагревали на водяной бане до 80 °С и барботировали бескислородным газом N 2 до тех пор, пока кислородный индикатор резазурин не стал бесцветным. После охлаждения до 37 °С добавляли соответствующую концентрацию антибиотика из 1000-кратного анаэробного исходного раствора.
Две изогенные пары основных видов, чувствительный красный флуоресцентный штамм E. coli с его аналогом, устойчивым к Gm или Kn, конкурировали в градиенте шести концентраций антибиотиков (Gm [мкг/мл]: 0, 0,01 , 0,1, 1, 10, 100; Kn [мкг/мл]: 0, 0,02, 0,2, 2, 20, 200). Штаммы, а также сообщество (100 мкл замороженного сырья) выращивали раздельно в анаэробных условиях в тройных реакторах, повторы объединяли, собирали центрифугированием, дважды промывали в 0,9% анаэробного раствора NaCl и, наконец, ресуспендировали в 0,9% растворе NaCl, доводили до ОП 600 0,1 (~10 7 бактерий/мл) и впоследствии использовали в экспериментах по конкуренции. В то время как сообщество выращивали в качестве инокулята из одного и того же замороженного гомогенизированного запаса, как подвыборка, так и погрешность культивирования, присущие выращиванию экологического сообщества в лабораторных условиях, привели к различиям в исходном составе модельного сообщества (рис. S1, S2). При выращивании сообщества в изоляции в отсутствие антибиотиков пропускная способность достигалась через 18 ч в расчете на ОП 600 показания спектрофотометра.
При выращивании сообщества в изоляции в отсутствие антибиотиков пропускная способность достигалась через 18 ч в расчете на ОП 600 показания спектрофотометра.
Изогенные штаммы смешивали в соотношении 1:1 (общественное отсутствие лечения), и эта смесь далее добавлялась в соотношении 10% к 90% фекального сообщества (общественное лечение). Приблизительно 10 6 бактерий любой смеси переносили в шесть повторных реакторов с каждой из концентраций антибиотика и выращивали при 37°C при встряхивании со скоростью 120 об/мин в течение 24 ч, что позволяло вырасти до несущей способности. В результате нормализации общего размера инокулята результирующий размер инокулята очаговых видов в отсутствие (~10 6 бактерий) и наличие (~10 5 бактерий = 10% от общего инокулята) сообщества различались. Затем объем 100 мкл каждого реактора переносили в новый биореактор, выращивали в течение 24 часов, снова переносили для заключительного 24-часового цикла роста и, наконец, собирали для последующего анализа. {T_0}} \right)} \right)} }$$
{T_0}} \right)} \right)} }$$
Проверка статистической значимости ( n = 6) выполнялась с использованием одностороннего t -критерия против нейтрального отбора ( ρ = 1) и ANOVA с поправкой на множественное тестирование для сравнения относительной пригодности различных образцов.
Анализ МИК
Для индивидуальной оценки МИК восприимчивого и резистентного очагового штамма в присутствии и в отсутствие микробного сообщества в реакторы инокулировали 10 5 очаговых бактерий и 10 6 бактерии из сообщества для лечения в рамках сообщества. Тройные реакторы выращивали в течение ночи в градиенте антибиотиков. Концентрации были увеличены на 1 мкг/мл (чувствительный к Gm), 2 мкг/мл (чувствительный к Km), 25 мкг/мл (устойчивый к Gm) и 50 мкг/мл (устойчивый к Km) соответственно. Затем реакторы собирали и высевали на LB-агар. Положительный рост оценивали как более чем четырехкратное увеличение числа бактериальных колоний на чашках по сравнению с посевом инокулята. МИК определяли как первую концентрацию, при которой не наблюдалось положительного роста.
МИК определяли как первую концентрацию, при которой не наблюдалось положительного роста.
Экстракция ДНК и секвенирование
Бактерии из каждого реактора, а также инокулят и исходное фекальное сообщество свиней собирали путем центрифугирования 2 мл жидкости с последующей экстракцией ДНК с использованием набора Qiagen PowerSoil в соответствии с инструкциями производителя. Качество и количество экстрактов подтверждали электрофорезом в 1% агарозном геле и двухцепочечной ДНК BR (Qubit) соответственно.
Библиотеки генов 16S рРНК были сконструированы с использованием мультиплексных праймеров, предназначенных для амплификации области V4 [27]. Ампликоны были созданы с использованием высокоточной полимеразы (Kapa 2G Robust), очищены с помощью системы очистки Agencourt AMPure XP PCR и количественно оценены с использованием флуорометра (Qubit, Life Technologies, Карлсбад, Калифорния, США). Очищенные ампликоны объединяли в эквимолярных концентрациях на основе количественного определения Qubit. Полученный пул библиотеки ампликонов разбавляли до 2 нМ гидроксидом натрия и 5 мл переносили в 995 мл HT1 (Illumina), чтобы получить конечную концентрацию 10 мкМ. Шестьсот миллилитров разбавленного пула библиотек добавляли 10% PhiXControl v3 и помещали на лед перед загрузкой в картридж Illumina MiSeq в соответствии с инструкциями производителя. Для секвенирования использовался набор реагентов MiSeq Reagent Kit v2 (500 циклов) с показателями цикла 250 циклов для каждого считывания парных концов с использованием программного обеспечения MiSeq Control 2.2.0 и RTA 1.17.28.
Полученный пул библиотеки ампликонов разбавляли до 2 нМ гидроксидом натрия и 5 мл переносили в 995 мл HT1 (Illumina), чтобы получить конечную концентрацию 10 мкМ. Шестьсот миллилитров разбавленного пула библиотек добавляли 10% PhiXControl v3 и помещали на лед перед загрузкой в картридж Illumina MiSeq в соответствии с инструкциями производителя. Для секвенирования использовался набор реагентов MiSeq Reagent Kit v2 (500 циклов) с показателями цикла 250 циклов для каждого считывания парных концов с использованием программного обеспечения MiSeq Control 2.2.0 и RTA 1.17.28.
Метагеномные библиотеки были созданы с использованием комплекта для подготовки библиотек KAPA high through (номер по каталогу: KK8234), оптимизированного для 1 мкг исходной ДНК с выбором размера и с использованием гранул Beckman Coulter XP (номер по каталогу: A63880). Образцы разрезали с помощью ультразвукового аппарата Covaris S2 (доступного от Covaris и Life Technologies) до размера 350 пар оснований. Концы образцов были восстановлены, 3′-5′-экзонуклеазная активность удалила 3′-выступы, а полимеразная активность заполнила 5′-выступы, создав тупые концы. Один нуклеотид «А» был добавлен к 3′-концам тупых фрагментов, чтобы предотвратить их лигирование друг с другом во время реакции лигирования адаптера. Соответствующий одиночный нуклеотид «Т» на 3′-конце адаптера обеспечивает комплементарный выступ для лигирования адаптера с фрагментом, обеспечивая низкую скорость образования химеры. Адаптеры-индикаторы лигировали к концам фрагментов ДНК для гибридизации на проточной кювете. Лигированный продукт подвергался отбору по размеру с использованием гранул ХР, описанных выше, что позволило удалить большую часть нелигированных или гибридизированных адаптеров. Перед гибридизацией образцы подвергались шести циклам ПЦР для селективного обогащения этих фрагментов ДНК адаптерными молекулами на обоих концах и для увеличения количества ДНК в библиотеке. ПЦР выполняли со смесью праймеров для ПЦР, которая отжигает концы адаптера. Размер вставки библиотек проверяли, запуская аликвоту библиотеки ДНК на PerkinElmer GX с использованием чипа High Sensitivity DNA (номер по каталогу: 5067–4626), а концентрацию определяли с помощью анализа High Sensitivity Qubit.
Один нуклеотид «А» был добавлен к 3′-концам тупых фрагментов, чтобы предотвратить их лигирование друг с другом во время реакции лигирования адаптера. Соответствующий одиночный нуклеотид «Т» на 3′-конце адаптера обеспечивает комплементарный выступ для лигирования адаптера с фрагментом, обеспечивая низкую скорость образования химеры. Адаптеры-индикаторы лигировали к концам фрагментов ДНК для гибридизации на проточной кювете. Лигированный продукт подвергался отбору по размеру с использованием гранул ХР, описанных выше, что позволило удалить большую часть нелигированных или гибридизированных адаптеров. Перед гибридизацией образцы подвергались шести циклам ПЦР для селективного обогащения этих фрагментов ДНК адаптерными молекулами на обоих концах и для увеличения количества ДНК в библиотеке. ПЦР выполняли со смесью праймеров для ПЦР, которая отжигает концы адаптера. Размер вставки библиотек проверяли, запуская аликвоту библиотеки ДНК на PerkinElmer GX с использованием чипа High Sensitivity DNA (номер по каталогу: 5067–4626), а концентрацию определяли с помощью анализа High Sensitivity Qubit. Все необработанные данные секвенирования были отправлены в ENA под исследовательским регистрационным номером PRJEB29.924.
Все необработанные данные секвенирования были отправлены в ENA под исследовательским регистрационным номером PRJEB29.924.
Анализ гена 16S рРНК
Анализ последовательности выполнен с использованием mothur v.1.32.1 [28] и MiSeq SOP [27], доступ 07.08.2017 на http://www.mothur.org/wiki /MiSeq_SOP. Последовательности классифицировали на основе классификатора RDP [29]. Разнообразие оценивали на основе наблюдаемых OTU при сходстве последовательностей 97%. Все последовательности основного вида E. coli были удалены на основании сходства последовательностей ≥99%. Последовательности с такой степенью сходства с очаговыми видами в исходном фекальном сообществе не обнаружены. Графики NMDS для сообщества были построены на основе метрики несходства Брея-Кертиса [30].
Дальнейшее сходство образцов было протестировано с использованием анализа молекулярной дисперсии (AMOVA) — непараметрического аналога традиционного тестирования ANOVA. AMOVA обычно используется в популяционной генетике для проверки гипотезы о том, что генетическое разнообразие между двумя или более популяциями существенно не отличается от сообщества, созданного путем стохастического объединения этих популяций [31, 32].
Метагеномный анализ
Метагеномные образцы, а также эталонный геном для очаговых видов E. coli MG1655, были проанализированы с использованием конвейера ARG-OAP для обнаружения генов устойчивости к антибиотикам из метагеномных данных с использованием интегрированной структурированной базы данных генов устойчивости к антибиотикам [33]. Это привело к обилию различных классов и подтипов генов устойчивости в этих группах, нормированных по количеству копий гена 16S рРНК. Гены устойчивости к антибиотикам, обнаруженные в эталонном геноме E. coli , были вычтены из общего числа совпадений на копию гена 16S рРНК на основе распространенности E. coli Ген 16S рРНК/общий ген 16S рРНК. Кроме того, все числа генов устойчивости к антибиотикам нормализовали к количеству гена 16S рРНК свиного фекального сообщества на общую копию гена 16S рРНК.
Математическая модель
Чтобы проиллюстрировать возможные механизмы, лежащие в основе данных о приспособленности бактерий в присутствии/отсутствии сообщества для различных концентраций Gm и Kn, мы математически описали нашу экспериментальную установку. Для этого мы сначала разработали дискретную математическую модель роста восприимчивых и устойчивых к лекарствам бактерий, 9{t + 1}\) — размер популяции штамма i в момент времени t + 1, а ϕ i — максимальная скорость роста при отсутствии конкуренции и давления лекарств. Уменьшение роста из-за регулирования/ограничения ресурсов, зависящего от плотности, определяется как
Для этого мы сначала разработали дискретную математическую модель роста восприимчивых и устойчивых к лекарствам бактерий, 9{t + 1}\) — размер популяции штамма i в момент времени t + 1, а ϕ i — максимальная скорость роста при отсутствии конкуренции и давления лекарств. Уменьшение роста из-за регулирования/ограничения ресурсов, зависящего от плотности, определяется как
с k d грузоподъемностью и e 9{\alpha _i — \beta _i\ln c}}}} \right),$$
где c — концентрация препарата (в мкг/мл), α i и β i — параметры, описывающие зависимость доза-реакция для штамма i , а f max = 0,9 — максимальное ингибирование роста.
Моделирование модели и расчет относительной приспособленности
Начиная с изначально небольшого количества бактерий в свежей среде, мы запускали модель для 30 поколений, после чего бактериальная популяция достигала несущей способности, и соответственно разбавляли популяцию. Бактериям снова дали возможность расти в течение 30 поколений, а затем разбавили и вырастили в течение последних 30 поколений. В этот момент мы рассчитали относительную приспособленность резистентного штамма как 90} \right)}}.$$
Бактериям снова дали возможность расти в течение 30 поколений, а затем разбавили и вырастили в течение последних 30 поколений. В этот момент мы рассчитали относительную приспособленность резистентного штамма как 90} \right)}}.$$
Зависимое от сообщества изменение лекарственной устойчивости/восприимчивости
Данные Kn, по-видимому, предполагают, что польза от бактерий, устойчивых к лекарственным препаратам, снижается в присутствии сообщества при средних и высоких дозах концентрации, указывающие на снижение восприимчивости восприимчивого штамма в контексте сообщества. Мы зафиксировали этот сценарий, сделав параметры доза-реакция α s,r и β 93}}} \right),$$
, где α i,0 и β i,0 — независимые от времени параметры доза-реакция. Эффект зависимости от плотности дополнительно проиллюстрирован (рис. S3).
Оценки параметров
Для каждого препарата (Gm и Kn) мы получили набор значений параметров, которые привели к хорошему общему совпадению между моделированием модели и данными, где данные включали наблюдаемую относительную пригодность для обоих наборов экспериментов ( т.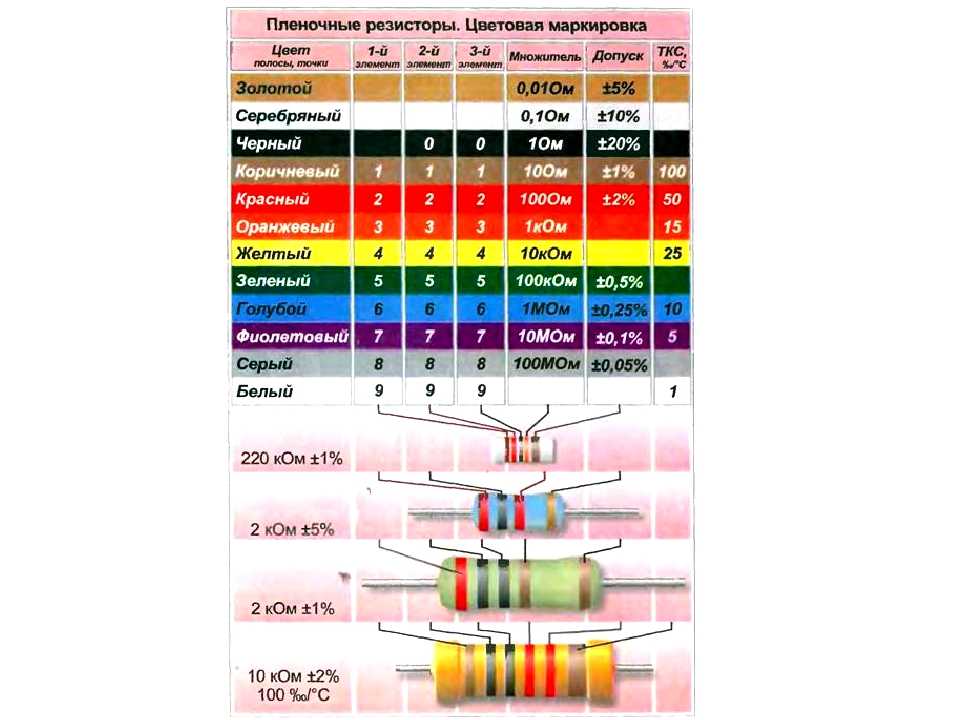 е. бактерии, выращенные в присутствии и в отсутствие сообщества) для шести различных концентраций препарата. Для обеспечения логарифмической регрессии предполагалось, что неантибиотический контроль на один порядок ниже, чем самая низкая концентрация, использованная в эксперименте. Значения параметров определялись путем минимизации среднеквадратичной ошибки с использованием алгоритма оптимизации, аналогичного имитации отжига [34]. Цель здесь состояла не в том, чтобы выполнить строгую оценку параметров, а скорее в том, чтобы найти набор параметров, которые с учетом конкретных модельных ограничений и допущений приводят к поведению модели, качественно согласующемуся как с наблюдаемой динамикой в течение повторяющихся циклов роста, так и с эмпирически определенными значениями пригодности. . Фактически, наш метод не смог найти уникальный набор значений, который постоянно давал бы наилучшую подходящую модель, что говорит о том, что доступных данных было недостаточно для определения глобального максимума.
е. бактерии, выращенные в присутствии и в отсутствие сообщества) для шести различных концентраций препарата. Для обеспечения логарифмической регрессии предполагалось, что неантибиотический контроль на один порядок ниже, чем самая низкая концентрация, использованная в эксперименте. Значения параметров определялись путем минимизации среднеквадратичной ошибки с использованием алгоритма оптимизации, аналогичного имитации отжига [34]. Цель здесь состояла не в том, чтобы выполнить строгую оценку параметров, а скорее в том, чтобы найти набор параметров, которые с учетом конкретных модельных ограничений и допущений приводят к поведению модели, качественно согласующемуся как с наблюдаемой динамикой в течение повторяющихся циклов роста, так и с эмпирически определенными значениями пригодности. . Фактически, наш метод не смог найти уникальный набор значений, который постоянно давал бы наилучшую подходящую модель, что говорит о том, что доступных данных было недостаточно для определения глобального максимума.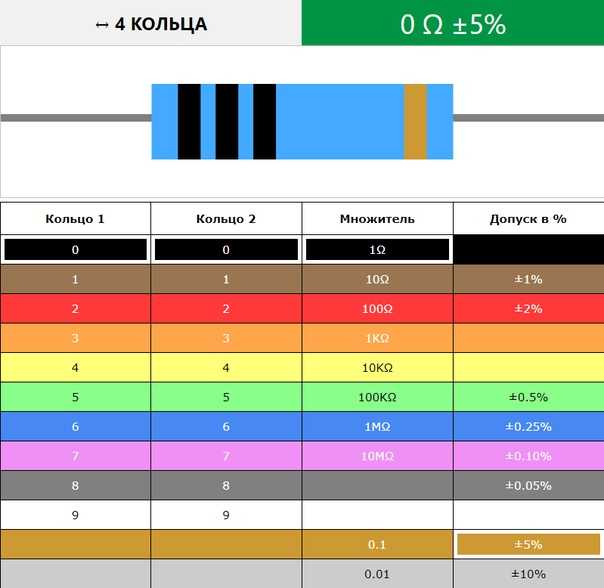 Однако качественные взаимосвязи между отдельными параметрами и между параметрами, сравнивающими два противомикробных препарата, были довольно стабильными между прогонами модели. В таблицах 1 и 2 перечислены наборы параметров, оцененных для двух разных антибиотиков. Эти параметры позволяют оценить изменения в плотности сообществ и основных видов в ходе отдельных экспериментов по конкуренции на основе измерений начальной и конечной точек (рис. S4).
Однако качественные взаимосвязи между отдельными параметрами и между параметрами, сравнивающими два противомикробных препарата, были довольно стабильными между прогонами модели. В таблицах 1 и 2 перечислены наборы параметров, оцененных для двух разных антибиотиков. Эти параметры позволяют оценить изменения в плотности сообществ и основных видов в ходе отдельных экспериментов по конкуренции на основе измерений начальной и конечной точек (рис. S4).
Полноразмерная таблица
Таблица 2 Значения параметров модели для кривой отбора канамицинаПолноразмерная таблица
Результаты
Контекст сообщества влияет на отбор на устойчивость штаммов к гентамицину
9000 очаговые виды E. coli с устойчивостью к Gm и без нее соревновались в присутствии и в отсутствие фекального сообщества свиней с градиентом концентрации Gm в 5 порядков. Независимо от концентрации антибиотика численность основных видов увеличилась в течение 3-дневного соревновательного эксперимента с ~ 10% при инокуляции до более 9Относительная численность 0% на основе секвенирования гена 16S рРНК (рис. S5A и B). Как резистентные, так и чувствительные штаммы, а также сообщество показали положительный рост во всем диапазоне концентраций и обеих обработок с увеличением количества клеток очаговых видов на 2,25–3,96 порядка в день (рис. 1а). Рис. 1
Независимо от концентрации антибиотика численность основных видов увеличилась в течение 3-дневного соревновательного эксперимента с ~ 10% при инокуляции до более 9Относительная численность 0% на основе секвенирования гена 16S рРНК (рис. S5A и B). Как резистентные, так и чувствительные штаммы, а также сообщество показали положительный рост во всем диапазоне концентраций и обеих обработок с увеличением количества клеток очаговых видов на 2,25–3,96 порядка в день (рис. 1а). Рис. 1 Мальтузианский показатель роста в сутки изогенных штаммов очаговых видов по гентамицину. Значения отображаются по градиенту антибиотика, а также при отсутствии и присутствии микробного сообщества кишечника. a Среднее (±SD, n = 6) логарифмическое абсолютное увеличение в день для устойчивого штамма, восприимчивого штамма и сообщества. Использовался различный размер инокулята очаговых видов в отсутствие (~10 6 бактерий) и в присутствии (~10 5 бактерий = 10% от общего инокулята) сообщества.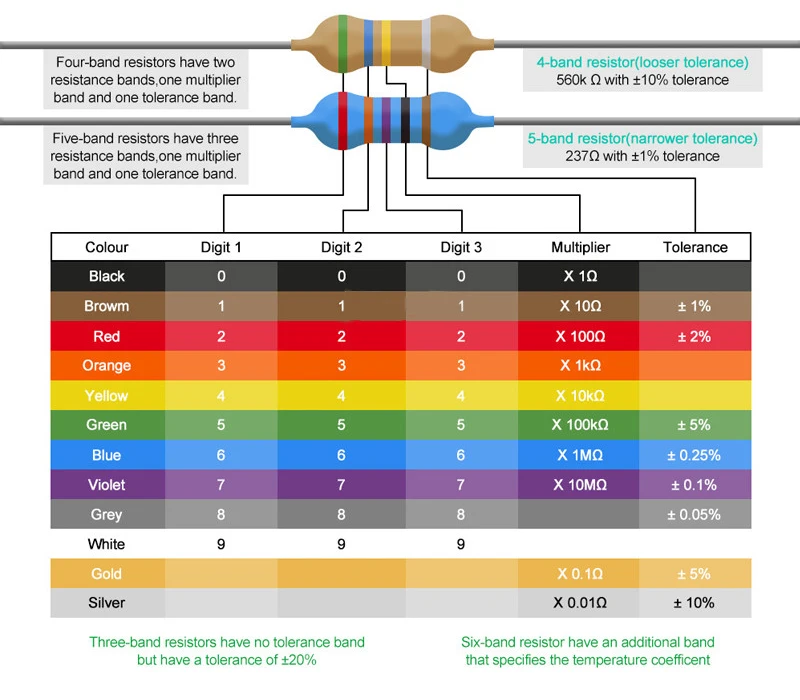 b Отношение абсолютных мальтузианских параметров роста (с 95% доверительными интервалами на основе 1000-кратного бутстрап-анализа) в присутствии и в отсутствие микробного сообщества по градиенту концентраций антибиотиков
b Отношение абсолютных мальтузианских параметров роста (с 95% доверительными интервалами на основе 1000-кратного бутстрап-анализа) в присутствии и в отсутствие микробного сообщества по градиенту концентраций антибиотиков
Изображение в полный размер
Небольшая стоимость соревновательной пригодности ( t -тест против 1, p = 0,0005) устойчивости к Gm в отсутствие сообщества ( ρ r 4,0144 0,5 среднее ± SD), и эта стоимость оказалась значительно выше, когда присутствовало сообщество (рис. 2) ( ρ r = 0,788 ± 0,016) (ANOVA с поправкой на множественное тестирование, p < 0,01, F = 360,36). При всех концентрациях от 0 до 10 мкг/мл относительный рост восприимчивых штаммов больше выигрывал от присутствия сообщества по сравнению с устойчивыми штаммами (ANOVA с поправкой на множественное тестирование, p < 0,05), до тех пор, пока при 100 мкг/мл Gm сообщество не оказывало значительного влияния на относительный рост (ANOVA с поправкой на множественное тестирование, p = 0,259, F = 1,42) (рис. 1b).
1b).
Относительная приспособленность штамма, устойчивого к гентамицину. Значения (среднее ± SD, n = 6) в присутствии (черный) и отсутствии (красный) сообщества. Сплошные линии представляют кривую наилучшего соответствия с помощью математической модели, основанной на оценках параметров, представленных в таблице 1. Пунктирная линия указывает на нейтральный выбор при относительной пригодности ρ r = 1, где точка пересечения с кривой пригодности указывает на минимальную селективную концентрацию возможно, способствовал наблюдаемым изменениям в отборе на устойчивость, вызванным сообществом, особенно между 10 и 100 мкг/мл. Состав микробного сообщества значительно менялся от собранного образца фекалий до инокулята и далее в течение всего эксперимента (AMOVA, p < 0,001, рис. S1A). Выше 1 мкг/мл Gm ранее доминирующие Proteobacteria уступили место Firmicutes (рис. S1B), что привело к значительному (AMOVA, p < 0,01) разделению сообществ ниже и выше этой пороговой концентрации на графике NMDS ( Рис. S1A). Однако значительных изменений в составе между 10 и 100 мкг/мл не наблюдалось, что свидетельствует о том, что изменения состава не играли большой роли в отборе, навязанном сообществом.
S1A). Однако значительных изменений в составе между 10 и 100 мкг/мл не наблюдалось, что свидетельствует о том, что изменения состава не играли большой роли в отборе, навязанном сообществом.
Контекст сообщества обуславливает цену сопротивления
Для проверки гипотез, полученных на основе числовых данных, мы использовали численное моделирование нашей экспериментальной установки, чтобы определить вероятные механизмы, лежащие в основе наблюдаемой динамики популяции в модели десятичного логарифма роста. Мы определили модели, основанные на ключевых эмпирических данных в отсутствие сообщества (в частности, на том, что в отсутствие антибиотиков существует цена устойчивости и что антибиотики ингибируют рост чувствительного штамма дозозависимым образом), а затем определили наиболее экономный способ, которым сообщество могло бы изменить относительную приспособленность устойчивых и восприимчивых штаммов (таблица 1). Мы нашли хорошее соответствие данным, просто предположив, что сообщество оказывает большее конкурентное воздействие, постоянное во всем градиенте антибиотиков, на резистентный, а не чувствительный штамм (9). 0087 e rj >> e cj > e sj
0087 e rj >> e cj > e sj 54 2; где
e ij — коэффициент конкуренции, налагаемый сообществом на фокальную популяцию (устойчивую r , восприимчивую s и сообщество c ). Кроме того, более высокие концентрации Gm приводят к резкому падению скорости роста сообщества и, следовательно, снижению повышенных затрат на устойчивость, налагаемых сообществом на эти более высокие уровни антибиотиков из-за снижения конкуренции. Это в сочетании с очень небольшим ростом восприимчивого штамма объясняет, почему относительная приспособленность между лечением в присутствии и отсутствием сообщества сходилась при высоких концентрациях антибиотиков (100 мкг/мл), при этом относительная приспособленность в основном определялась ростом резистентного штамма.
Численное моделирование позволило нам оценить изменение MSC от отсутствия к присутствию сообщества путем детерминированной оценки концентрации на пересечении с нейтральным отбором при относительной приспособленности ρ r = 1. кратное увеличение МСК в присутствии сообщества (рис. 2). МИК для восприимчивого штамма оставалась стабильной на уровне 8–9 мкг/мл как в присутствии, так и в отсутствие сообщества (таблица 3). Следовательно, это 43-кратное увеличение MSC смещает отбор от концентраций намного ниже MIC в отсутствие к концентрациям около MIC в присутствии сообщества.
кратное увеличение МСК в присутствии сообщества (рис. 2). МИК для восприимчивого штамма оставалась стабильной на уровне 8–9 мкг/мл как в присутствии, так и в отсутствие сообщества (таблица 3). Следовательно, это 43-кратное увеличение MSC смещает отбор от концентраций намного ниже MIC в отсутствие к концентрациям около MIC в присутствии сообщества.
 3b). Рис. 3
3b). Рис. 3 Мальтузианский показатель роста в сутки изогенных штаммов очаговых видов по канамицину. Значения отображаются по градиенту антибиотика, а также при отсутствии и присутствии микробного сообщества кишечника. a Среднее (±SD, n = 6) логарифмическое абсолютное увеличение в день для устойчивого (красный), восприимчивого штамма и сообщества. Различный размер инокулята очаговых видов в отсутствие (~10 6 бактерий) и наличие (~10 5 бактерий = 10% от общего инокулята) сообщества. b Отношение абсолютных мальтузианских параметров роста (с 95% доверительными интервалами на основе 1000-кратного бутстрап-анализа) в присутствии и отсутствии микробного сообщества по градиенту концентраций антибиотиков
Изображение в натуральную величину
сообщество не увеличило общую стоимость сопротивления. Действительно, сообщество не оказало существенного влияния на относительную приспособленность резистентного штамма, за исключением концентрации 20 мкг/мл (ANOVA с поправкой на множественное тестирование, p = 0,002, F = 15,58) (рис. 4). Резистентный штамм обладал явным преимуществом в приспособленности в отсутствие сообщества при этой концентрации ( ρ r = 1,288 ± 0,149; t -тест против 1, p сообщества, эта разница в относительной приспособленности, хотя и значимая ( t -тест против 1, p = 0,0088), была значительно меньше ( ρ r = 1,034 ± 0,020). При 200 мкг/мл Kn, близком к MIC восприимчивых штаммов, устойчивый штамм имел столь же высокую относительную приспособленность независимо от присутствия сообщества (ANOVA с поправкой на множественное тестирование, p = 0,079, F = 3,84).
4). Резистентный штамм обладал явным преимуществом в приспособленности в отсутствие сообщества при этой концентрации ( ρ r = 1,288 ± 0,149; t -тест против 1, p сообщества, эта разница в относительной приспособленности, хотя и значимая ( t -тест против 1, p = 0,0088), была значительно меньше ( ρ r = 1,034 ± 0,020). При 200 мкг/мл Kn, близком к MIC восприимчивых штаммов, устойчивый штамм имел столь же высокую относительную приспособленность независимо от присутствия сообщества (ANOVA с поправкой на множественное тестирование, p = 0,079, F = 3,84).
Относительная приспособленность штамма, устойчивого к канамицину. Значения (среднее ± SD, n = 6) в присутствии (черный) и отсутствии (красный) сообщества. Сплошные линии представляют кривую наилучшего соответствия с помощью математической модели, основанной на оценках параметров, представленных в таблице 2. Пунктирная линия указывает на нейтральный выбор при относительной приспособленности ρ r = 1, где точка пересечения с кривой пригодности указывает на минимальная избирательная концентрация
Пунктирная линия указывает на нейтральный выбор при относительной приспособленности ρ r = 1, где точка пересечения с кривой пригодности указывает на минимальная избирательная концентрация
Полноразмерное изображение
Сообщество и композиция устойчивости к антибиотикам остаются стабильными в градиенте канамицина
Как и в эксперименте с Gm, наблюдался значительный сдвиг в составе сообщества от собранного образца фекалий к инокуляту и далее в течение эксперимента Kn (AMOVA, p < 0,001, рис. S2A). Однако во всем градиенте антибиотиков Firmicutes (рис. S2B) оставались доминирующим типом без значительных изменений в составе сообщества в результате. Таким образом, изменения состава снова не могут объяснить влияние сообщества на приспособленность фокального штамма при отборе только при 20 мкг / мл. Мы дополнительно провели метагеномный анализ для обработок 0, 2 и 20 мкг/мл Kn, чтобы определить, изменилось ли относительное количество генов устойчивости в сообществе, несмотря на то, что изменений в составе сообщества не было. Устойчивость к аминогликозидам (ANOVA, p = 0,04) и другие классы антибиотиков (фосмидомицин, казугамицин, макролиды, полимиксин и тетрациклин (ANOVA, все p < 0,01)) значительно увеличились в сообществе всех реакторов по сравнению с исходным фекальным сообществом независимо от концентрации антибиотика (рис. 5а). Однако не было существенной разницы между концентрацией Kn и распространенностью (ANOVA, p = 0,15) устойчивости к аминогликозидам в целом (рис. 5a) или каких-либо конкретных подтипов устойчивости к аминогликозидам (рис. 5b), что позволяет предположить, что относительная приспособленность фокальной вид не подвергался влиянию резистома сообщества. Однако гены устойчивости, обнаруженные с помощью метагеномного анализа, экспрессировались, поскольку устойчивые колонии от членов сообщества были обнаружены на селективных чашках после эксперимента по конкуренции. Неудивительно, что, поскольку в сообществе не наблюдалось зависящего от концентрации антибиотика отбора на устойчивость к аминогликозидам, не наблюдалось и значительного совместного отбора на устойчивость к любым другим классам антибиотиков.
Устойчивость к аминогликозидам (ANOVA, p = 0,04) и другие классы антибиотиков (фосмидомицин, казугамицин, макролиды, полимиксин и тетрациклин (ANOVA, все p < 0,01)) значительно увеличились в сообществе всех реакторов по сравнению с исходным фекальным сообществом независимо от концентрации антибиотика (рис. 5а). Однако не было существенной разницы между концентрацией Kn и распространенностью (ANOVA, p = 0,15) устойчивости к аминогликозидам в целом (рис. 5a) или каких-либо конкретных подтипов устойчивости к аминогликозидам (рис. 5b), что позволяет предположить, что относительная приспособленность фокальной вид не подвергался влиянию резистома сообщества. Однако гены устойчивости, обнаруженные с помощью метагеномного анализа, экспрессировались, поскольку устойчивые колонии от членов сообщества были обнаружены на селективных чашках после эксперимента по конкуренции. Неудивительно, что, поскольку в сообществе не наблюдалось зависящего от концентрации антибиотика отбора на устойчивость к аминогликозидам, не наблюдалось и значительного совместного отбора на устойчивость к любым другим классам антибиотиков.
Выявлены гены устойчивости. Тип ( a ) и аминогликозидный подтип ( b ) относительная распространенность (число генов устойчивости, нормализованное с числом копий гена 16S рРНК), в исходном фекальном сообществе и в конечном сообществе реактора при трех концентрациях канамицина (среднее значение ± стандартное отклонение; n фекалии = 2, n Kn0 = 6, n Kn2 = 6 n Kn441 90 90). Показаны только гены, обнаруженные с помощью конвейера ARGs-OAP. MLS = Макролиды, линкозамиды, стрептограмины
Полноразмерное изображение
Присутствие сообщества может увеличить рост восприимчивой популяции E. coli при промежуточных концентрациях антибиотиков
Численное моделирование показало, что, в отличие от устойчивости к Gm, вызванное сообществом увеличение стоимости устойчивости к Kn не может объяснить, почему польза от лекарственно-устойчивого очагового штамма E. coli была снижена в присутствии сообщества при промежуточные концентрации препарата ( e rj = e s ). Это предполагает различные взаимодействия между E. coli и остальной частью сообщества, и мы предположили, что сообщество могло обеспечить защитный эффект против Kn для восприимчивой E. coli . Данные о росте продемонстрировали, что это так: при лечении по месту жительства скорость роста как восприимчивой, так и резистентной E. coli изменялась согласованным образом при более низких концентрациях. Единственные исключения наблюдались при концентрации 20 мкг/мл, когда скорость роста восприимчивого, но не резистентного штамма значительно увеличивалась в присутствии сообщества (ANOVA с поправкой на множественное тестирование, p = 0,002, F = 15,58) (рис. 3б). Кроме того, небольшое увеличение МПК восприимчивого штамма в присутствии сообщества с 16–18 до 18–20 мкг/мл (таблица 3) подтверждает представление о защитном эффекте.
coli была снижена в присутствии сообщества при промежуточные концентрации препарата ( e rj = e s ). Это предполагает различные взаимодействия между E. coli и остальной частью сообщества, и мы предположили, что сообщество могло обеспечить защитный эффект против Kn для восприимчивой E. coli . Данные о росте продемонстрировали, что это так: при лечении по месту жительства скорость роста как восприимчивой, так и резистентной E. coli изменялась согласованным образом при более низких концентрациях. Единственные исключения наблюдались при концентрации 20 мкг/мл, когда скорость роста восприимчивого, но не резистентного штамма значительно увеличивалась в присутствии сообщества (ANOVA с поправкой на множественное тестирование, p = 0,002, F = 15,58) (рис. 3б). Кроме того, небольшое увеличение МПК восприимчивого штамма в присутствии сообщества с 16–18 до 18–20 мкг/мл (таблица 3) подтверждает представление о защитном эффекте. Мы исследовали, был ли защитный эффект сообщества достаточным для объяснения наблюдаемых данных путем подгонки численного моделирования, где параметры доза-реакция α s,r и β s,r явно зависели от зависящей от времени плотности сообщества (как указано в таблице 2). Полученная модель хорошо соответствовала экспериментальным данным, предполагая, что защита сообщества является движущей силой наблюдаемой динамики популяции с 12-кратным увеличением MSC. Несмотря на этот сдвиг МСК, кроме Gm, устойчивость к отбору Kn по-прежнему наблюдалась при концентрациях, в 5–10 раз ниже MIC чувствительного штамма.
Мы исследовали, был ли защитный эффект сообщества достаточным для объяснения наблюдаемых данных путем подгонки численного моделирования, где параметры доза-реакция α s,r и β s,r явно зависели от зависящей от времени плотности сообщества (как указано в таблице 2). Полученная модель хорошо соответствовала экспериментальным данным, предполагая, что защита сообщества является движущей силой наблюдаемой динамики популяции с 12-кратным увеличением MSC. Несмотря на этот сдвиг МСК, кроме Gm, устойчивость к отбору Kn по-прежнему наблюдалась при концентрациях, в 5–10 раз ниже MIC чувствительного штамма.
Обсуждение
В этом исследовании мы изучили, как включение в полуестественное сообщество (сообщество, полученное из свиных кишок в анаэробном биореакторе) влияет на селекцию УПП в очаговых видах ( E. coli ). Для двух антибиотиков (Gm и Kn) мы обнаруживаем, что наличие сообщества выбирает против устойчивости, что приводит к повышению на 1–2 порядка минимальных селективных концентраций для устойчивости к антибиотикам и, таким образом, достижению отбора либо ближе к Gm, либо к Gm даже вблизи восприимчивых. штаммы МИК. Это говорит о том, что недавние оценки МСК на основе отдельных штаммов in vitro [6, 7, 11], вероятно, намного ниже, чем можно было бы наблюдать in vivo, и это может объяснить, почему в некоторых экосистемах не наблюдалось отбора на устойчивость к антибиотикам в очаговых штаммах [35].
штаммы МИК. Это говорит о том, что недавние оценки МСК на основе отдельных штаммов in vitro [6, 7, 11], вероятно, намного ниже, чем можно было бы наблюдать in vivo, и это может объяснить, почему в некоторых экосистемах не наблюдалось отбора на устойчивость к антибиотикам в очаговых штаммах [35].
Основные механизмы, ответственные за это навязанное сообществом снижение отбора на устойчивость, различались для двух протестированных препаратов, но, вероятно, являются довольно общими из-за их экологического происхождения. Для Gm сообщество увеличило затраты на приспособление, что отразилось в снижении скорости роста, связанной с резистентностью в отсутствие антибиотиков. Эти повышенные затраты сохранялись на одинаковых уровнях в зависимости от градиента антибиотиков до тех пор, пока дозы не стали настолько высокими, что рос только устойчивый штамм (аналогичное поведение выше определенной пороговой концентрации ранее было описано для систем с одним штаммом [36, 37], и наши результаты показывают что это верно в контексте сообщества). Было обнаружено, что ограничение ресурсов — прямое манипулирование или конкуренция — увеличивает затраты на борьбу с рядом стрессоров в ряде организмов, от устойчивости плазмодия к противомалярийным препаратам [38] до устойчивости к фагам у бактерий [18]. Возможно, это связано с тем, что ограничение ресурсов оказывает более выраженное влияние на резистентные генотипы [39].].
Было обнаружено, что ограничение ресурсов — прямое манипулирование или конкуренция — увеличивает затраты на борьбу с рядом стрессоров в ряде организмов, от устойчивости плазмодия к противомалярийным препаратам [38] до устойчивости к фагам у бактерий [18]. Возможно, это связано с тем, что ограничение ресурсов оказывает более выраженное влияние на резистентные генотипы [39].].
Для Kn навязанный сообществом отбор против устойчивости был очевиден только при промежуточных концентрациях антибиотика. Абсолютная скорость роста восприимчивого штамма значительно увеличивалась при промежуточных концентрациях в присутствии сообщества. Подгонка нашей модели предполагает, что это связано с защитным эффектом сообщества, что дополнительно подтверждается небольшим увеличением МИК восприимчивого штамма в присутствии сообщества. Защитный эффект мог наблюдаться только при промежуточных концентрациях, поскольку низкие концентрации были недостаточны для заметного снижения относительной приспособленности восприимчивого штамма, а при высоких концентрациях защитный эффект был слишком мал, чтобы его можно было обнаружить.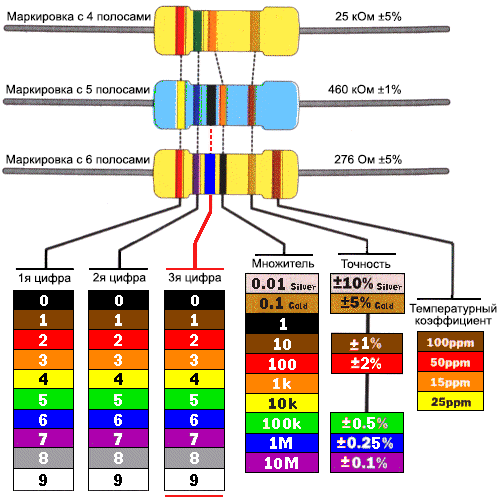 О таких защитных эффектах широко сообщалось в пределах видов [13, 14], а в последнее время и в более сложных сообществах [12] из-за вне- или внутриклеточной модификации антибиотиков. Другие известные механизмы повышения устойчивости штаммов к антибиотикам в сообществах включают флокуляцию [40] или образование биопленки [41, 42], но здесь они могут играть лишь незначительную роль из-за условий встряхивания.
О таких защитных эффектах широко сообщалось в пределах видов [13, 14], а в последнее время и в более сложных сообществах [12] из-за вне- или внутриклеточной модификации антибиотиков. Другие известные механизмы повышения устойчивости штаммов к антибиотикам в сообществах включают флокуляцию [40] или образование биопленки [41, 42], но здесь они могут играть лишь незначительную роль из-за условий встряхивания.
Наблюдалось основное различие между экспериментами по конкуренции Kn и Gm, которое могло иметь последствия для вероятных механизмов, лежащих в основе опосредованных сообществом изменений в отборе на устойчивость. В отсутствие отбора антибиотиков более высокое разнообразие сообщества, включающее большую долю Proteobacteria , было обнаружено в конкурирующем сообществе во время эксперимента Gm по сравнению с экспериментом Kn. Поскольку очаговый вид E. coli также относится к Proteobacteria , разница в структуре сообщества и особенно численности протеобактерий могла увеличить уровень перекрытия ниш между очаговым штаммом и сообществом и, таким образом, конкуренцию [43], что может объяснить, почему устойчивость к Gm имела значительно повышенную стоимость, а устойчивость к Kn — нет.
Два общих механизма, обсуждавшихся выше, лежат в основе отбора на постоянные вариации в ранее существовавших генах устойчивости, а не отбор на изменчивость de novo, возникающую в результате спонтанных мутаций или горизонтального переноса генов от других видов. Для хромосомных мутаций de novo сообщество, вероятно, еще больше ограничивает распространение резистентности, потому что уменьшенные размеры популяции очаговых штаммов в присутствии сообщества увеличивают вероятность того, что будут зафиксированы более дорогостоящие мутации [44]. Напротив, включение в сообщество может усилить распространение сопротивления. Во-первых, у фокусных видов появится больший источник генов устойчивости. Во-вторых, отбор против устойчивости, приобретаемой в результате горизонтального переноса генов при низких концентрациях антибиотиков, может иметь другую динамику. В то время как хромосомная устойчивость может быть вытеснена и впоследствии потеряна, гены устойчивости, встроенные в конъюгативные плазмиды, могут сохраняться или даже увеличиваться в изобилии вследствие их иногда чрезвычайно широкого круга хозяев и высокой частоты переноса [22, 45, 46, 47, 48]. В контролируемых экспериментах с одним штаммом плазмидная резистентность оказалась более дорогостоящей, чем хромосомная резистентность [7]. Однако в более сложных сценариях отбор на переносимую мобильными генетическими элементами резистентность обычно зависит не только от одного приобретенного гена устойчивости, но и от комбинации других связанных признаков, кодируемых МГЭ как части общего генофонда [49].]. Таким образом, возникают трудности с общими прогнозами динамики отбора горизонтально приобретенной устойчивости в микробных сообществах, которые заслуживают дальнейших исследований.
В контролируемых экспериментах с одним штаммом плазмидная резистентность оказалась более дорогостоящей, чем хромосомная резистентность [7]. Однако в более сложных сценариях отбор на переносимую мобильными генетическими элементами резистентность обычно зависит не только от одного приобретенного гена устойчивости, но и от комбинации других связанных признаков, кодируемых МГЭ как части общего генофонда [49].]. Таким образом, возникают трудности с общими прогнозами динамики отбора горизонтально приобретенной устойчивости в микробных сообществах, которые заслуживают дальнейших исследований.
Таким образом, мы показываем, что на отбор в отношении УПП повлияло включение в «естественное» микробное сообщество, так что MSC увеличилось более чем на порядок для двух разных антибиотиков. В дополнение к снижению относительной приспособленности к устойчивости, включение в сообщество также снижает абсолютную приспособленность, которая, как утверждается, иногда является основным фактором распространения устойчивости [50]. Цель этого исследования состояла в том, чтобы определить эти общие механизмы, лежащие в основе сниженного отбора для УПП в сложном контексте сообщества с высокой степенью экологического реализма. Однако более конкретные индивидуальные взаимодействия, способствующие этим общим эффектам, могут зависеть от конкретного сообщества, антибиотика и соответствующих генов устойчивости и должны быть исследованы в будущем.
Цель этого исследования состояла в том, чтобы определить эти общие механизмы, лежащие в основе сниженного отбора для УПП в сложном контексте сообщества с высокой степенью экологического реализма. Однако более конкретные индивидуальные взаимодействия, способствующие этим общим эффектам, могут зависеть от конкретного сообщества, антибиотика и соответствующих генов устойчивости и должны быть исследованы в будущем.
Таким образом, для определения МСК, которые являются значимыми в условиях окружающей среды, крайне важно проводить отбор в сложном контексте сообщества, а не в системах с одним штаммом. Понимание того, при каких концентрациях происходит отбор и, следовательно, долговременная фиксация вновь приобретенных механизмов устойчивости, имеет решающее значение для будущего смягчения распространения генов устойчивости, а также их потенциально патогенных хозяев [51, 52]. Наши результаты еще раз подчеркивают необходимость предпочтительного использования антибиотиков узкого спектра действия в клинической терапии для поддержания здорового микробиома пациента, который может легче восстанавливаться после введения антибиотика [53], тем самым снижая вероятность положительного отбора патогенов, которые могли приобрести резистентность, когда встраивается в сообщество.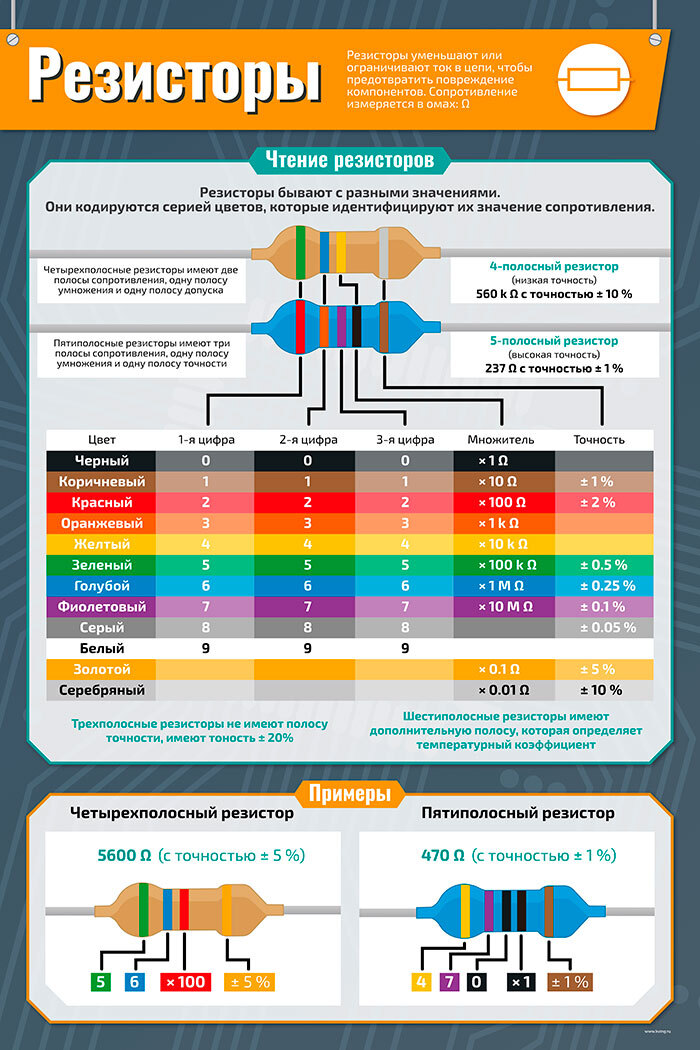
Ссылки
-
ВОЗ. Глобальный отчет о надзоре за устойчивостью к противомикробным препаратам. 2014 г. https://www.who.int/drugresistance/documents/surveillancereport/en/.
-
Д’Коста В.М., Кинг К.Э., Калан Л., Морар М., Сунг В.В.Л., Шварц С. и др. Устойчивость к антибиотикам возникла давно. Природа. 2011; 477:457–61.
ПабМед Статья КАС Google ученый
-
Кнапп К.В., Долфинг Дж., Элерт Пай, Грэм Д.В. Доказательства увеличения содержания генов устойчивости к антибиотикам в заархивированных почвах с 19 г.40. Экологическая наука Техн. 2010;44:580–7.
КАС пабмед Статья Google ученый
-
Дрлица К. Окно селекции мутантов и устойчивость к противомикробным препаратам. J Антимикробная химиотерапия. 2003; 52:11–7.

КАС пабмед Статья Google ученый
-
Drlica K, Zhao X. Гипотеза окна отбора мутантов обновлена. Клин Инфекция Дис. 2007; 44: 681–8.
ПабМед Статья Google ученый
-
Галлберг Э., Цао С., Берг О.Г., Илбек С., Сандегрен Л., Хьюз Д. и др. Селекция резистентных бактерий при очень низких концентрациях антибиотиков. PLoS Патог. 2011;7:e1002158.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Гульберг Э., Альбрехт Л.М., Карлссон С., Сандегрен Л., Андерссон Д.И. Селекция плазмиды множественной лекарственной устойчивости по сублетальным уровням антибиотиков и тяжелых металлов. МБио. 2014;5:e01918–14.
ПабМед ПабМед Центральный Статья КАС Google ученый
 «>
«> Carlet J. Кишечник является эпицентром устойчивости к антибиотикам. Противомикробная защита от инфекций. 2012;1:39.
ПабМед ПабМед Центральный Статья Google ученый
-
Лундстрем С.В., Остман М., Бенгтссон-Палме Дж., Рутгерссон С., Тоудал М., Сиркар Т. и другие. Минимальные селективные концентрации тетрациклина в сложных водных бактериальных биопленках. Научная общая среда. 2016; 553: 587–95.
ПабМед Статья КАС Google ученый
-
Мюррей А.К., Чжан Л., Инь Х., Чжан Т., Баклинг А., Снейп Дж. и др. Новое понимание селекции на устойчивость к антибиотикам в сложных микробных сообществах. МБио. 2018;9:e00969–18.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Лю А., Фонг А., Бекет Э., Юань Дж., Тамаэ С.
 , Медрано Л. и др. Селективное преимущество резистентных штаммов при следовых количествах антибиотиков: простой и сверхчувствительный цветовой тест для обнаружения антибиотиков и генотоксических агентов. Противомикробные агенты Chemother. 2011;55:1204–10.
, Медрано Л. и др. Селективное преимущество резистентных штаммов при следовых количествах антибиотиков: простой и сверхчувствительный цветовой тест для обнаружения антибиотиков и генотоксических агентов. Противомикробные агенты Chemother. 2011;55:1204–10. КАС пабмед ПабМед Центральный Статья Google ученый
-
Сорг Р.А., Лин Л., ван Дорн Г.С., Сорг М., Олсон Дж., Низет В. и др. Коллективная резистентность в микробных сообществах путем дезактивации внутриклеточных антибиотиков. PLoS биол. 2016;14:e2000631.
ПабМед ПабМед Центральный Статья КАС Google ученый
-
Юрцев Е.А., Чао Х.С., Датта М.С., Артемова Т., Гор Дж. Обман бактериями управляет динамикой популяции кооперативных плазмид устойчивости к антибиотикам. Мол Сист Биол. 2013;9:683.
КАС пабмед ПабМед Центральный Статья Google ученый
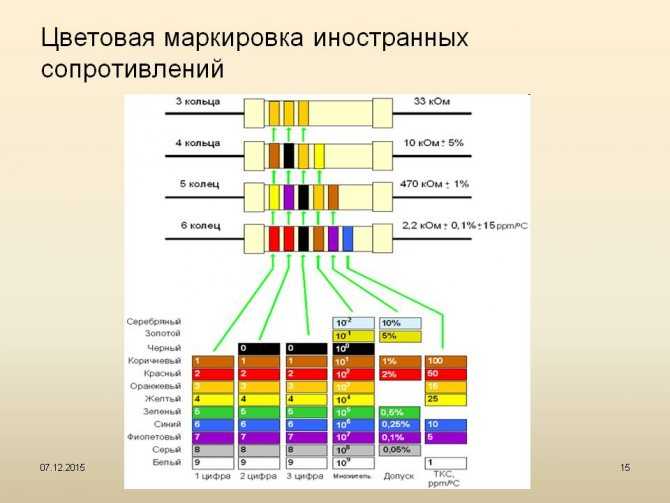 «>
«> Медани Ф., Димитриу Т., Эллис Р.Дж., Рэймонд Б. Жить, чтобы обмануть еще один день: бактериальный покой способствует социальной эксплуатации β-лактамаз. ISME J. 2016; 10: 778–87.
КАС пабмед Статья Google ученый
-
Цао Дж., Кюрстен Д., Шнайдер С., Кнауэр А., Гюнтер П.М., Келер Дж.М. Раскрытие токсикологической сложности с помощью многомерного скрининга в микросегментированном потоке: модуляция воздействия антибиотиков наночастицами. Лабораторный чип. 2012;12:474–84.
КАС пабмед Статья Google ученый
-
Чурски К., Камински Т.С., Якела С., Камыш В., Баранска-Рыбак В., Вейбель Д.Б. и др. Быстрый скрининг токсичности антибиотиков в автоматизированной микрокапельной системе. Лабораторный чип. 2012;12:1629.
КАС пабмед Статья Google ученый
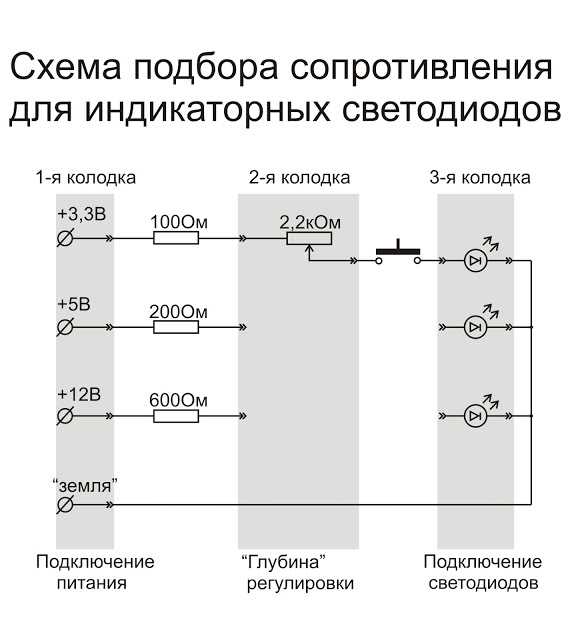 «>
«> Kraaijeveld AR, Limentani EC, Godfray HCJ. Основа компромисса между устойчивостью к паразитоидам и конкурентоспособностью личинок Drosophila melanogaster. Proc Biol Sci. 2001;268:259–61.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Гомес П., Баклинг А. Антагонистическая коэволюция бактериофагов в почве. Наука. 2011; 332:106–9.
ПабМед Статья КАС Google ученый
-
Тенайлон О., Скурник Д., Пикард Б., Денамур Э. Популяционная генетика комменсальной кишечной палочки. Nat Rev Microbiol. 2010;8:207–17.
КАС пабмед Статья Google ученый
-
Remus-Emsermann MNP, Gisler P, Drissner D. Векторы доставки MiniTn7-транспозона для экспрессии индуцибельного или конститутивного флуоресцентного белка в Enterobacteriaceae.
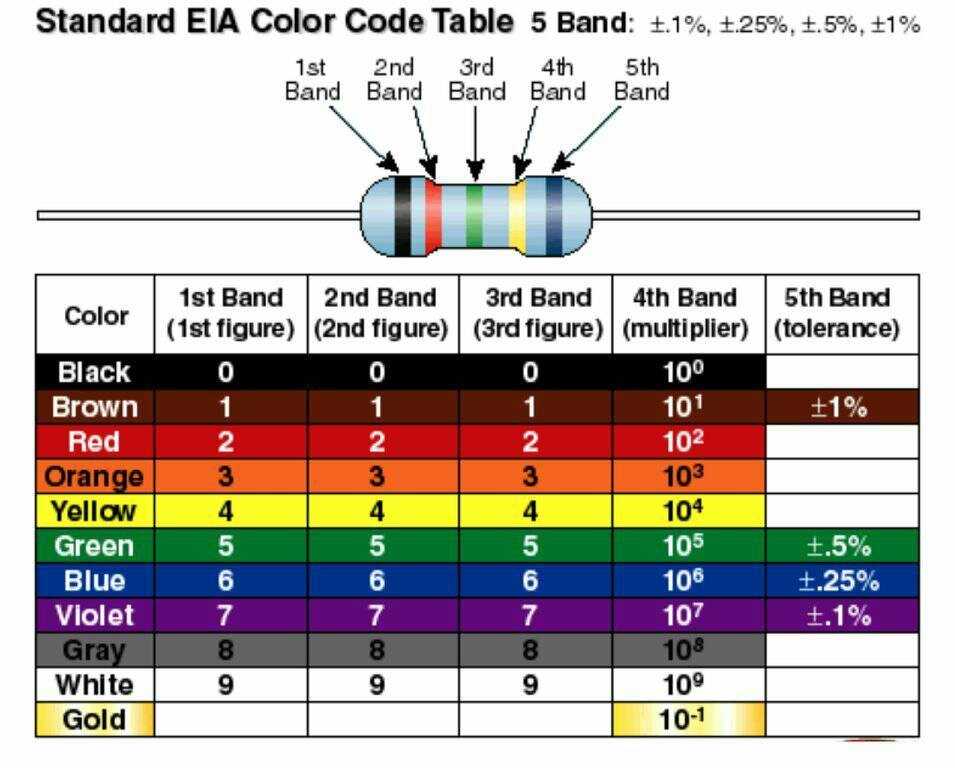 FEMS Microbiol Lett. 2016;363:fnw178.
FEMS Microbiol Lett. 2016;363:fnw178. ПабМед ПабМед Центральный Статья КАС Google ученый
-
Клюмпер У, Дечесне А, Сметс БФ. Протокол для оценки допустимости бактериальных сообществ по отношению к конъюгальным плазмидам путем количественного определения и выделения трансконъюгантов. В: Редакторы McGenity T., Timmis K., Nogales B.. Протоколы микробиологии углеводородов и липидов, справочник по протоколам Springer. Springer, Берлин, Гейдельберг, 2014.
-
Клюмпер У., Рибер Л., Дечесне А., Санназарро А., Хансен Л.Х., Соренсен С.Дж. и др. Плазмиды широкого круга хозяев могут проникать в неожиданно разнообразную часть почвенного бактериального сообщества. ISME J. 2015; 9: 934–45.
ПабМед Статья КАС Google ученый
-
Мартинес-Гарсия Э., Каллес Б., Аревало-Родригес М., Де Лоренцо В.
 PBAM1: полностью синтетический генетический инструмент для анализа и построения сложных бактериальных фенотипов. БМС микробиол. 2011;11:38.
PBAM1: полностью синтетический генетический инструмент для анализа и построения сложных бактериальных фенотипов. БМС микробиол. 2011;11:38. ПабМед ПабМед Центральный Статья КАС Google ученый
-
Мартинес-Гарсия Э., Апарисио Т., де Лоренцо В., Никель П.И. Новые инструменты транспозонов, предназначенные для метаболической инженерии грамотрицательных микробных клеточных фабрик. Фронт Биоэнг Биотехнолог. 2014;2:46.
ПабМед ПабМед Центральный Google ученый
-
Ковач М.Е., Эльзер П.Х., Стивен Хилл Д., Робертсон Г.Т., Фаррис М.А., Руп Р.М. и др. Четыре новых производных клонирующего вектора широкого круга хозяев pBBR1MCS, несущие различные кассеты устойчивости к антибиотикам. Ген. 1995;166:175–176.
КАС пабмед Статья Google ученый
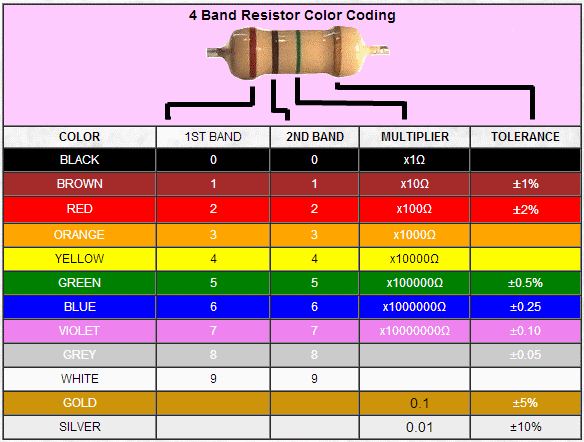 «>
«> Гроскопф Т., Зеноби С., Алстон М., Фолкес Л., Сварбрек Д., Сойер О.С. Стабильный генетический полиморфизм, лежащий в основе микробной синтрофии. ISME J. 2016; 10: 2844–53.
ПабМед ПабМед Центральный Статья КАС Google ученый
-
Козич Дж.Дж., Весткотт С.Л., Бакстер Н.Т., Хайлендер С.К., Шлосс П.Д. Разработка стратегии секвенирования с двойным индексом и конвейера курирования для анализа данных последовательности ампликона на платформе секвенирования MiSeq Illumina. Appl Environ Microbiol. 2013;79: 5112–20.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, et al. Представляем mothur: открытое, независимое от платформы, поддерживаемое сообществом программное обеспечение для описания и сравнения микробных сообществ. Appl Environ Microbiol.
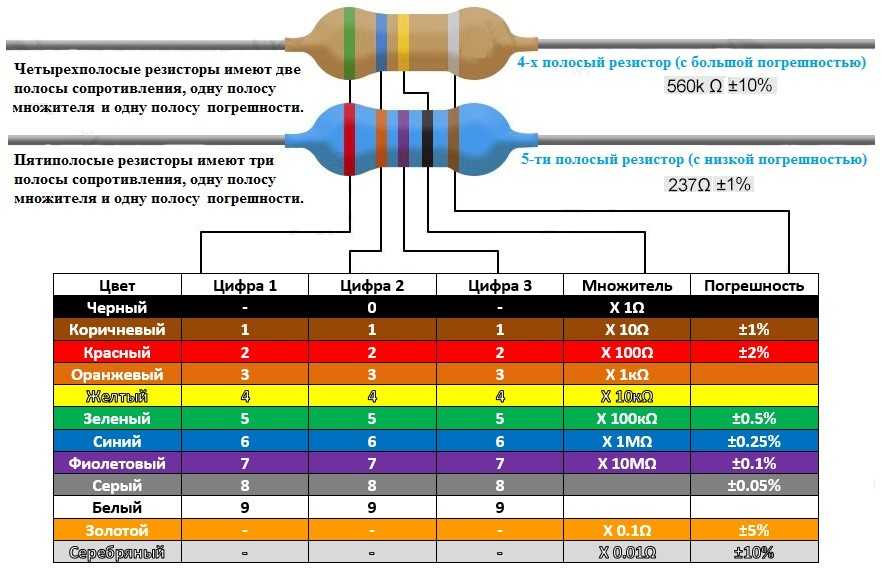 2009;75:7537–41.
2009;75:7537–41. КАС пабмед ПабМед Центральный Статья Google ученый
-
Ван К., Гаррити GM, Тидже Дж. М., Коул младший. Наивный байесовский классификатор для быстрого отнесения последовательностей рРНК к новой таксономии бактерий. Appl Environ Microbiol. 2007; 73: 5261–7.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Брей Дж. Р., Кертис Дж. Т. Назначение горных лесных сообществ Южного Висконсина. Эколь моногр. 1957; 27: 325–49.
Артикул Google ученый
-
Гравина С., Вийг Дж. Эпигенетические факторы старения и долголетия. Арка Пфлюгера. 2010; 459: 247–58.
КАС пабмед Статья Google ученый
-
Андерсон М.Дж. Новый метод непараметрического многомерного дисперсионного анализа.
 Австралийская экол. 2001; 26:32–46.
Австралийская экол. 2001; 26:32–46. Google ученый
-
Yang Y, Jiang X, Chai B, Ma L, Li B, Zhang A и др. ARG-OAP: конвейер онлайн-анализа для обнаружения генов устойчивости к антибиотикам из метагеномных данных с использованием интегрированной структурированной базы данных ARG. Биоинформатика. 2016; 32:2346–51.
КАС пабмед Статья Google ученый
-
Киркпатрик С., Гелатт К.Д., Векки М.П. Оптимизация путем имитации отжига. Наука. 1983; 220: 671–80.
КАС пабмед Статья Google ученый
-
Flach CF, Genheden M, Fick J, Joakim Larsson DG. Комплексный скрининг изолятов Escherichia coli с крупнейшей в Скандинавии станции очистки сточных вод не выявил селекции на устойчивость к антибиотикам. Технологии экологических наук. 2018;52:11419–28.

КАС пабмед Статья Google ученый
-
Андерссон Д.И., Хьюз Д. Устойчивость к антибиотикам и ее стоимость: возможно ли обратить резистентность вспять? Nat Rev Microbiol. 2010;8:260–71.
КАС пабмед Статья Google ученый
-
Андерссон Д.И., Хьюз Д. Стойкость устойчивости к антибиотикам в бактериальных популяциях. FEMS Microbiol Rev. 2011; 35:901–11.
КАС пабмед Статья Google ученый
-
Уэйл Н., Сим Д.Г., Джонс М.Дж., Салате Р., Дэй Т., Рид А.Ф. Ограничение ресурсов предотвращает появление лекарственной устойчивости за счет усиления конкуренции внутри хозяина. Proc Natl Acad Sci. 2017;114:201715874.
Артикул КАС Google ученый
-
Сонг Т.
 , Пак Ю., Шампута И.С., Сео С., Ли С.И., Чон Х.С. и др. Затраты на устойчивость к рифампицину у Mycobacterium tuberculosis увеличиваются в условиях голодания по питательным веществам и компенсируются мутацией в β’-субъединице РНК-полимеразы. Мол микробиол. 2014;91:1106–19.
, Пак Ю., Шампута И.С., Сео С., Ли С.И., Чон Х.С. и др. Затраты на устойчивость к рифампицину у Mycobacterium tuberculosis увеличиваются в условиях голодания по питательным веществам и компенсируются мутацией в β’-субъединице РНК-полимеразы. Мол микробиол. 2014;91:1106–19. КАС пабмед ПабМед Центральный Статья Google ученый
-
Кюммерер К. Антибиотики в водной среде — обзор — часть II. Хемосфера. 2009; 75: 435–41.
ПабМед Статья КАС Google ученый
-
Мах Т.Ф., Питтс Б., Пеллок Б., Уокер Г.К., Стюарт П.С., О’Тул Г.А. Генетическая основа устойчивости биопленки Pseudomonas aeruginosa к антибиотикам. Природа. 2003; 426: 306–10.
КАС пабмед Статья Google ученый
-
Дренкард Э., Ausubel FM. Образование биопленок Pseudomonas и устойчивость к антибиотикам связаны с фенотипической изменчивостью.
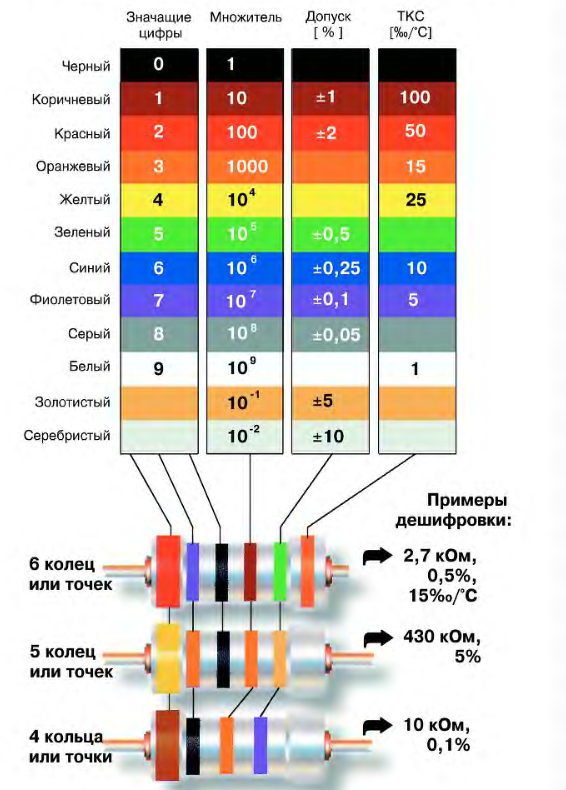 Природа. 2002; 416:740–3.
Природа. 2002; 416:740–3. КАС пабмед Статья Google ученый
-
Webb CO, Ackerly DD, McPeek MA, Donoghue MJ. Филогении и экология сообщества. Annu Rev Ecol Syst. 2002; 33: 475–505.
Артикул Google ученый
-
Perron GG, Gonzalez A, Buckling A. Динамика источника и стока определяет эволюцию устойчивости к антибиотикам и стоимость ее плейотропной приспособленности. Proc Biol Sci. 2007; 274:2351–6.
ПабМед ПабМед Центральный Статья Google ученый
-
Клюмпер У., Дечесне А., Рибер Л., Брандт К.К., Гюлай А., Соренсен С.Дж. и др. Металлические стрессоры последовательно модулируют потенциал поглощения бактериальной конъюгальной плазмиды филогенетически консервативным образом. ISME J. 2017; 11: 152–65.
ПабМед Статья Google ученый
 «>
«> Шинтани М., Мацуи К., Иноуэ Д.И., Хосояма А., Оджи С., Ямазоэ А. и др. Анализ отдельных клеток выявил диапазоны переноса плазмид incP-1, incP-7 и incP-9 в сообществе почвенных бактерий. Appl Environ Microbiol. 2014;80:138–45.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Мусович С., Клумпер У., Дечесне А., Магид Дж., Сметс Б.Ф. Длительное воздействие навоза увеличивает потенциал почвенного бактериального сообщества для поглощения плазмиды. Environ Microbiol Rep. 2014; 6:125–30.
КАС пабмед Статья Google ученый
-
Ариас-Андрес М., Клюмпер У., Рохас-Хименес К., Гроссарт Х.П. Загрязнение микропластиком увеличивает обмен генами в водных экосистемах. Загрязнение окружающей среды. 2018; 237: 253–61.
КАС пабмед Статья Google ученый
 «>
«> Норман А., Хансен Л.Х., Соренсен С.Дж. Конъюгативные плазмиды: сосуды общего генофонда. Philos Trans R Soc Lond B Biol Sci. 2009 г.;364:2275–89.
КАС пабмед ПабМед Центральный Статья Google ученый
-
День Т., Хейбен С., Рид А.Ф. Имеет ли значение отбор в эволюционном возникновении лекарственной устойчивости? Тенденции микробиол. 2015;23:126–33.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Larsson DGJ, Andremont A, Bengtsson-Palme J, Brandt KK, de Roda Husman AM, Fagerstedt P, et al. Критические пробелы в знаниях и потребности в исследованиях, связанные с экологическими аспектами устойчивости к антибиотикам. Окружающая среда Интерн. 2018;117:132–8.
ПабМед Статья Google ученый
-
Смолла К., Кук К.
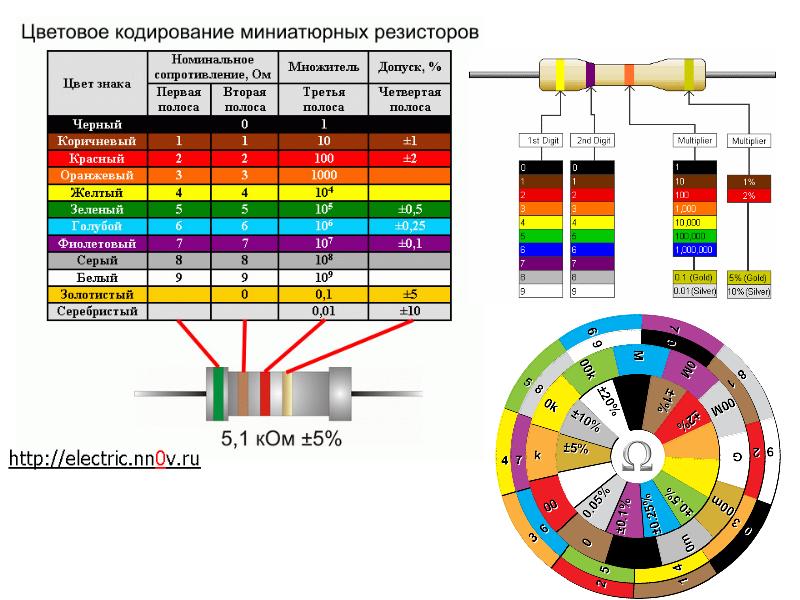 , Джорджевич С.П., Клумпер У., Гиллингс М. Экологические аспекты устойчивости к антибиотикам: оценка пробелов в фундаментальной науке. FEMS Microbiol Ecol. 2018;94:fiy191. https://doi.org/10.1093/femsec/fiy191.
, Джорджевич С.П., Клумпер У., Гиллингс М. Экологические аспекты устойчивости к антибиотикам: оценка пробелов в фундаментальной науке. FEMS Microbiol Ecol. 2018;94:fiy191. https://doi.org/10.1093/femsec/fiy191. -
Пальеха А., Миккельсен К.Х., Форслунд С.К., Кашани А., Аллин К.Х., Нильсен Т. и др. Восстановление микробиоты кишечника здоровых взрослых после воздействия антибиотиков. Нат микробиол. 2018;3:1255–65.
КАС пабмед Статья Google ученый
Скачать ссылки
Благодарности
Великобритания получила финансирование от исследовательской и инновационной программы Horizon 2020 Европейского Союза в рамках соглашения о гранте Марии Склодовской-Кюри №. 751699. Великобритания, AB и WG получили поддержку в виде гранта MRC/BBSRC (MR/N007174/1). XY благодарит Университет Гонконга за обучение в аспирантуре.
Информация об авторе
Авторы и филиалы
-
CLES & ESI, Эксетерский университет, Пенрин, Корнуолл, Великобритания
Ули Клюмпер и Ангус Баклинг
-
Европейский центр медицины окружающей среды и здоровья Пенри, Университет Эксетера , Корнуолл, Великобритания
Ули Клюмпер, Лихонг Чжан и Уильям Х.
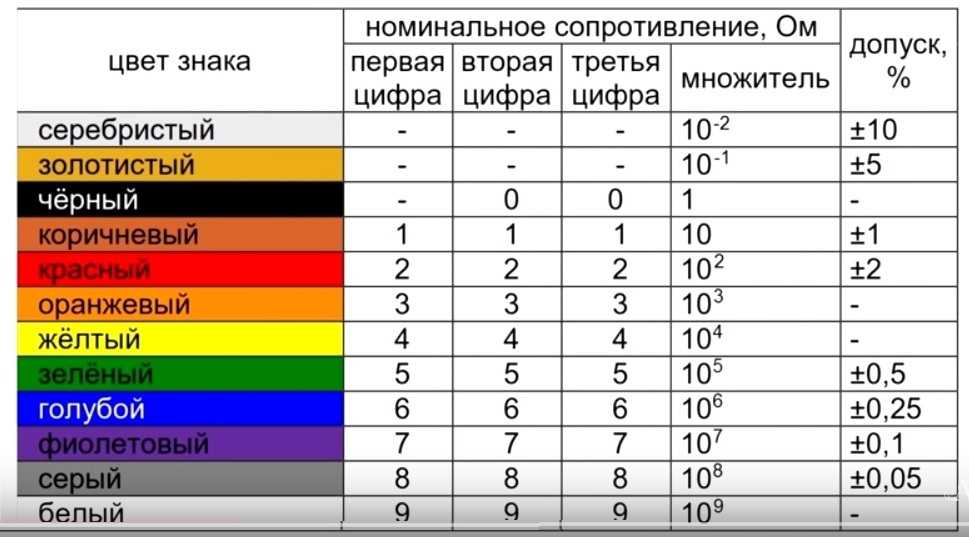 Гейз
Гейз -
Колледж инженерных, математических и физических наук Эксетерского университета, Пенрин, Корнуолл, Великобритания
Mario Recker
-
Департамент гражданского строительства, Университет Гонконга, Гонконг, Китай
Xiaole Yin & Tong Zhang
Авторы
- Uli Klümper
View Public автор в PubMed Google Scholar
- Mario Recker
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Lihong Zhang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Xiaole Yin
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Tong Zhang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ангус Баклинг
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- William H.
 Gaze
Gaze Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Автор, ответственный за корреспонденцию
Ули Клюмпер.
Заявление об этике
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Дополнительная информация
Примечание издателя: Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительный текст
Рисунок S1
Рисунок S2
Рисунок S3
Рисунок S4
. Рисунок S4
9003 9007
. Рисунок S4
9003 9007
. Рисунок.0085
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.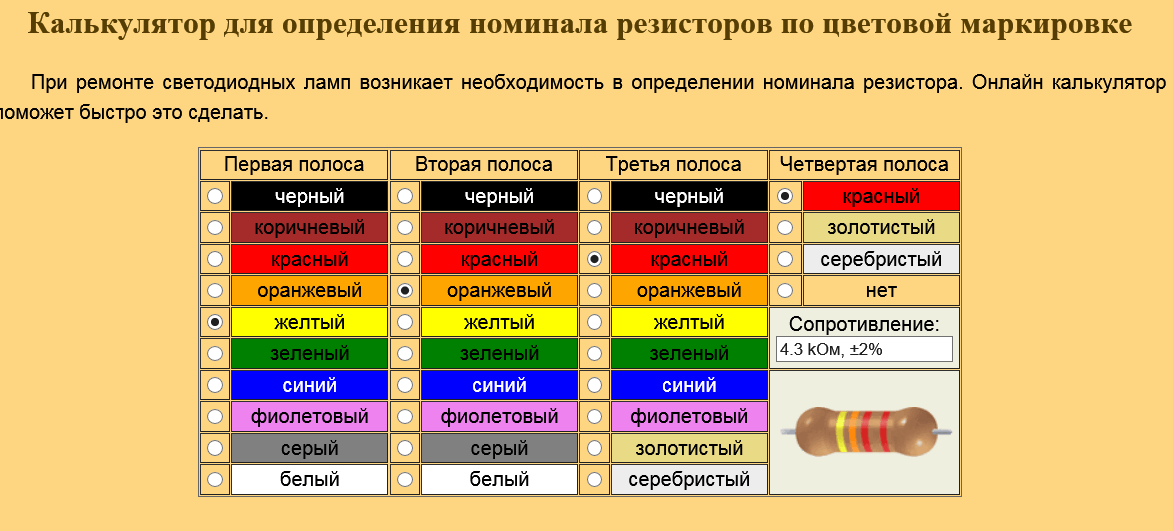 0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете автора(ов) оригинала. и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете автора(ов) оригинала. и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Дополнительная литература
-
Протеазы, разлагающие колистин, придают коллективную устойчивость микробным сообществам во время полимикробных инфекций.

- До-Хун Ли
- Джу Хи Ча
- Чанг-Джун Ча
Микробиом (2022)
-
Эволюция резистентности может нарушить защиту от воздействия антибиотиков за счет конкурентного исключения защитных видов.
- Ангус М.
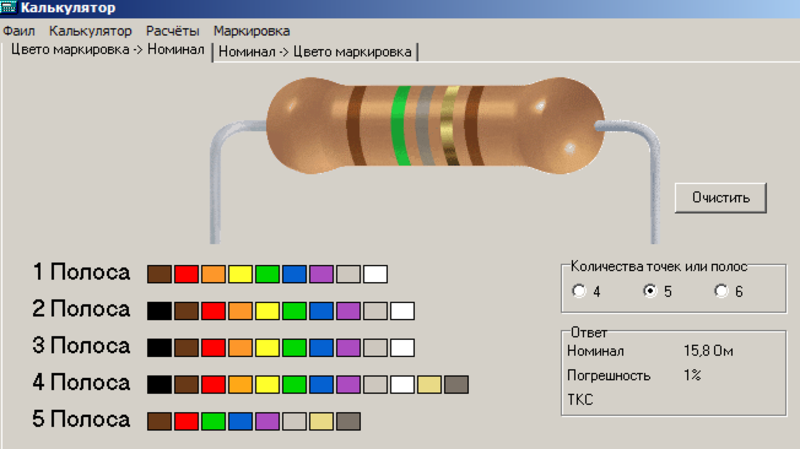 Куинн
Куинн - Майкл Дж. Боттери
- Вилле-Петри Фриман
Журнал ISME (2022)
- Ангус М.
-
Устойчивость к антибиотикам в окружающей среде
- Д. Г. Йоаким Ларссон
- Карл-Фредрик Флах
Nature Reviews Microbiology (2022)
-
Выявление полирезистентных клинических штаммов E.
 coli методом рамановской спектроскопии
coli методом рамановской спектроскопии
- Амир Накар
- Айкатерини Пистики
- Юрген Попп
Аналитическая и биоаналитическая химия (2022)
-
Связанная с человеком микробиота подавляет вторгающиеся бактерии даже при воздействии антибиотиков.
- Эндрю Д.
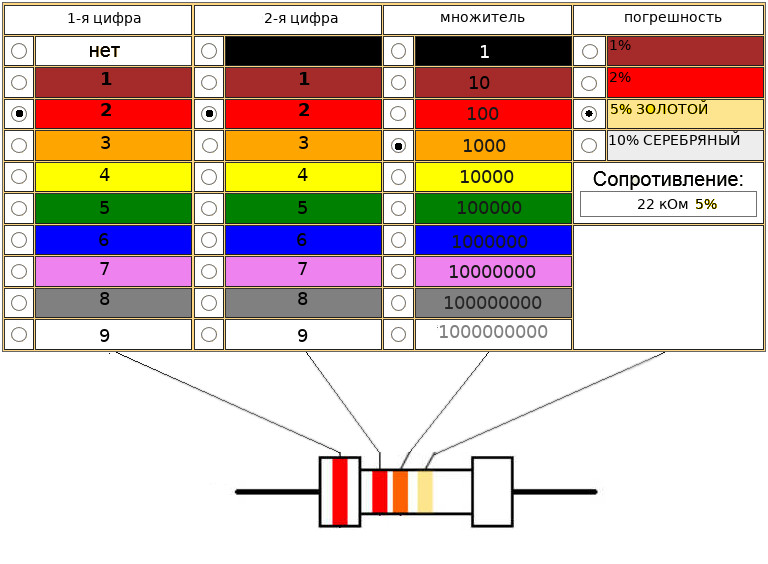 Леттен
Леттен - Майкл Баумгартнер
- Алекс Р. Холл
Журнал ISME (2021)
- Эндрю Д.
Скачать PDF
Выбор устойчивости противомикробными покрытиями в медицинских учреждениях
Обзор
. 2020 сен;106(1):115-125.
doi: 10.1016/j.jhin.2020.06.006. Epub 2020 12 июня.
Ф Питч
1
, Эй Джей О’Нил
2
, А Иваск
3
, Х.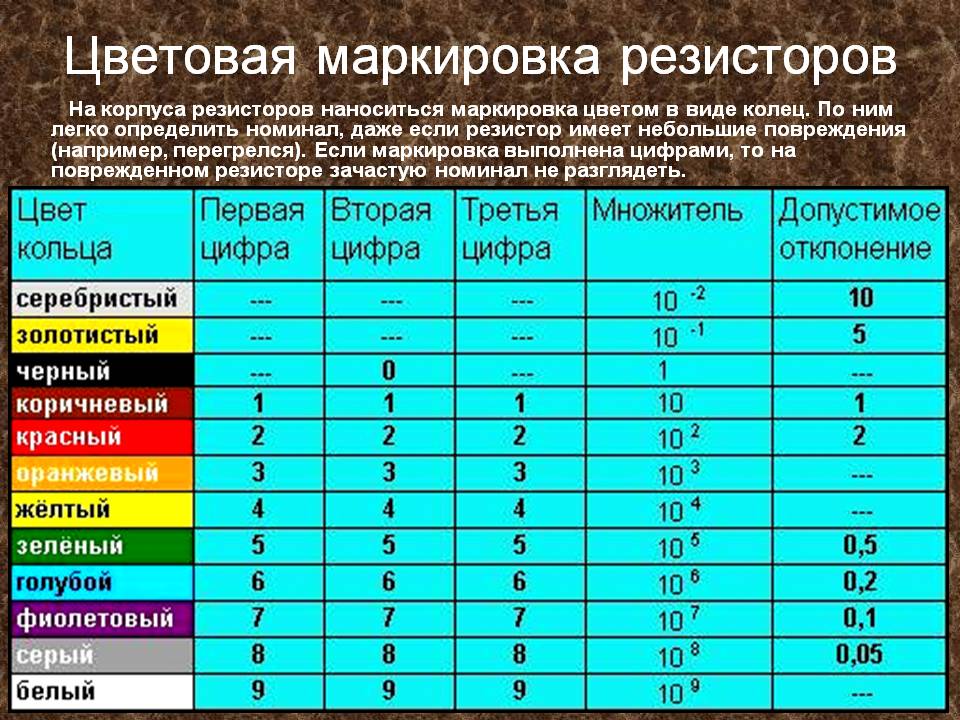 Дженссен
4
, Дж. Инкинен
5
, А Кару
6
, М Ахонен
7
, Ф. Шрайбер
8
Дженссен
4
, Дж. Инкинен
5
, А Кару
6
, М Ахонен
7
, Ф. Шрайбер
8
Принадлежности
- 1 Федеральный институт исследований и испытаний материалов, отдел материалов и окружающей среды, отдел биологических повреждений и эталонных организмов, Берлин, Германия.
- 2 Школа молекулярной и клеточной биологии, Факультет биологических наук, Лидский университет, Лидс, Великобритания.
- 3 Лаборатория токсикологии окружающей среды, Национальный институт химической физики и биофизики, Таллинн, Эстония; Институт молекулярной и клеточной биологии Тартуского университета, Тарту, Эстония.

- 4 Департамент науки и окружающей среды, Университет Роскилле, Роскилле, Дания.
- 5 Финский институт здравоохранения и социального обеспечения, Департамент медицинской безопасности, Хельсинки, Финляндия.
- 6 Лаборатория токсикологии окружающей среды, Национальный институт химической физики и биофизики, Таллинн, Эстония.
- 7 Университет прикладных наук Сатакунта, технологический факультет, Северный институт воды и материалов WANDER, Раума, Финляндия. Электронный адрес: [email protected].
- 8 Федеральный институт исследований и испытаний материалов, отдел материалов и окружающей среды, отдел биологических повреждений и эталонных организмов, Берлин, Германия.
 Электронный адрес: [email protected].
Электронный адрес: [email protected].
- PMID: 32535196
- DOI: 10.1016/j.jhin.2020.06.006
Бесплатная статья
Обзор
F Pietsch et al. Джей Хосп заражает. 2020 Сентябрь
Бесплатная статья
. 2020 сен;106(1):115-125.
doi: 10.1016/j.jhin.2020.06.006. Epub 2020 12 июня.
Авторы
Ф Питч
1
, Эй Джей О’Нил
2
, А Иваск
3
, Х.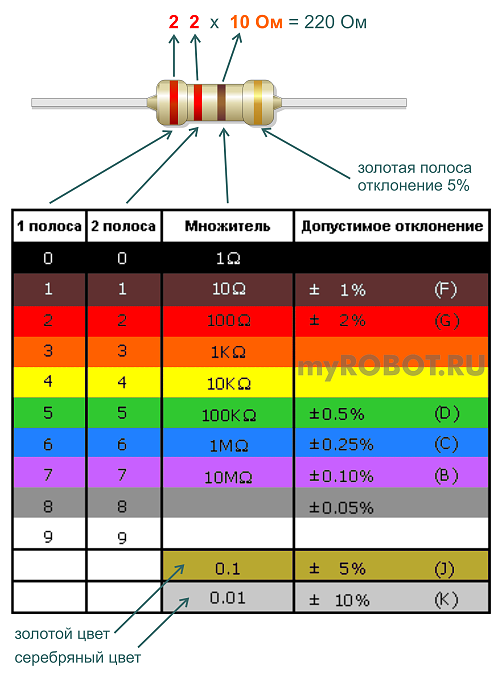 Дженссен
4
, Дж. Инкинен
5
, А Кару
6
, М Ахонен
7
, Ф. Шрайбер
8
Дженссен
4
, Дж. Инкинен
5
, А Кару
6
, М Ахонен
7
, Ф. Шрайбер
8
Принадлежности
- 1 Федеральный институт исследований и испытаний материалов, отдел материалов и окружающей среды, отдел биологических повреждений и эталонных организмов, Берлин, Германия.
- 2 Школа молекулярной и клеточной биологии, Факультет биологических наук, Лидский университет, Лидс, Великобритания.
- 3 Лаборатория токсикологии окружающей среды, Национальный институт химической физики и биофизики, Таллинн, Эстония; Институт молекулярной и клеточной биологии Тартуского университета, Тарту, Эстония.
- 4 Департамент науки и окружающей среды, Университет Роскилле, Роскилле, Дания.
- 5 Финский институт здравоохранения и социального обеспечения, Департамент медицинской безопасности, Хельсинки, Финляндия.
- 6 Лаборатория токсикологии окружающей среды, Национальный институт химической физики и биофизики, Таллинн, Эстония.
- 7 Университет прикладных наук Сатакунта, технологический факультет, Северный институт воды и материалов WANDER, Раума, Финляндия. Электронный адрес: [email protected].
- 8 Федеральный институт исследований и испытаний материалов, отдел материалов и окружающей среды, отдел биологических повреждений и эталонных организмов, Берлин, Германия.
 Электронный адрес: [email protected].
Электронный адрес: [email protected].
- PMID: 32535196
- DOI: 10.1016/j.jhin.2020.06.006
Абстрактный
Антимикробные сенсорные поверхности были введены в медицинских учреждениях с целью поддержки существующих гигиенических процедур и борьбы с растущей угрозой устойчивости к противомикробным препаратам. Однако были высказаны опасения по поводу потенциального давления отбора, оказываемого такими поверхностями, которое может стимулировать эволюцию и распространение устойчивости к противомикробным препаратам. В этом обзоре освещаются исследования, указывающие на риски, связанные с устойчивостью на антимикробных поверхностях в результате различных процессов, включая эволюцию путем мутации de novo и горизонтального переноса генов, а также видовую сортировку устойчивых по своей природе бактерий, рассеянных на антимикробных поверхностях. Обзор посвящен антимикробным поверхностям из меди, серебра и антимикробным пептидам из-за практического применения меди и серебра, а также многообещающих характеристик антимикробных пептидов. Имеющиеся данные указывают на возможность селекции резистентности и последующего увеличения числа резистентных штаммов за счет перекрестной резистентности и корезистентности, обусловленной признаками резистентности к металлам и антибиотикам. Тем не менее, трансляционные исследования, описывающие развитие резистентности к антимикробным сенсорным поверхностям в медицинских учреждениях, проводятся редко, и они будут необходимы для оценки того, приводят ли и каким образом антимикробные поверхности к выбору резистентности в этих условиях. В таких исследованиях необходимо будет учитывать множество переменных, в том числе концентрации противомикробных препаратов, присутствующих в покрытиях, наличие биопленок на поверхностях и влажность, характерную для среды с сухими поверхностями. Испытания на месте эффективности противомикробных покрытий должны регулярно оценивать риск выбора, связанный с их использованием.
Обзор посвящен антимикробным поверхностям из меди, серебра и антимикробным пептидам из-за практического применения меди и серебра, а также многообещающих характеристик антимикробных пептидов. Имеющиеся данные указывают на возможность селекции резистентности и последующего увеличения числа резистентных штаммов за счет перекрестной резистентности и корезистентности, обусловленной признаками резистентности к металлам и антибиотикам. Тем не менее, трансляционные исследования, описывающие развитие резистентности к антимикробным сенсорным поверхностям в медицинских учреждениях, проводятся редко, и они будут необходимы для оценки того, приводят ли и каким образом антимикробные поверхности к выбору резистентности в этих условиях. В таких исследованиях необходимо будет учитывать множество переменных, в том числе концентрации противомикробных препаратов, присутствующих в покрытиях, наличие биопленок на поверхностях и влажность, характерную для среды с сухими поверхностями. Испытания на месте эффективности противомикробных покрытий должны регулярно оценивать риск выбора, связанный с их использованием.
Ключевые слова: Антимикробное покрытие; Устойчивость к противомикробным препаратам; СТОИМОСТЬ действия CA15114 AMICI; Здравоохранение; инфекции; Сенсорные поверхности.
Copyright © 2020 Автор(ы). Опубликовано Elsevier Ltd. Все права защищены.
Похожие статьи
-
Влияние сухого осаждения инокулята на эффективность противомикробных поверхностей на основе меди.
Макдональд М., Весгейт Р., Рубиано М., Холах Дж., Деньер С.П., Джерманн С., Майллард Дж.Ю. Макдональд М. и др. Джей Хосп заражает. 2020 ноябрь; 106(3):465-472. doi: 10.1016/j.jhin.2020.08.013. Epub 2020 15 августа. Джей Хосп заражает. 2020. PMID: 32810570
-
Антимикробные материалы и покрытия на основе меди: потенциальное многогранное биомедицинское применение.

Митра Д., Канг Э.Т., Неох К.Г. Митра Д. и др. Интерфейсы приложений ACS. 2020 13 мая;12(19)):21159-21182. дои: 10.1021/acsami.9b17815. Epub 2019 27 декабря. Интерфейсы приложений ACS. 2020. PMID: 31880421 Обзор.
-
Последние достижения в области антимикробных покрытий на металлической основе для поверхностей, к которым часто прикасаются.
Биркетт М., Довер Л., Чериан Лукосе С., Васи Зия А., Тамбувала М.М., Серрано-Арока А. Биркетт М. и соавт. Int J Mol Sci. 2022 21 января; 23 (3): 1162. дои: 10.3390/ijms23031162. Int J Mol Sci. 2022. PMID: 35163084 Бесплатная статья ЧВК. Обзор.
-
«Реалистичная» оценка противомикробных поверхностей с помощью нового метода переноса прикосновения демонстрирует явное превосходство медного сплава над поверхностями, содержащими серебро.
Кноблох Дж.К., Тоферн С., Кунц В., Шютце С., Рике М., Зольбах В., Вуске Т. Кноблох Дж. К. и соавт. ПЛОС Один. 14 ноября 2017 г .; 12 (11): e0187442. doi: 10.1371/journal.pone.0187442. Электронная коллекция 2017. ПЛОС Один. 2017. PMID: 2
99 Бесплатная статья ЧВК. -
Инновации в области антимикробного покрытия для предотвращения инфекционных заболеваний (AMiCI): действие затрат ca15114.
Данне К.П., Кейнянен-Тойвола М.М., Кару А., Теуниссен Б., Олмез Х., Гувейя И., Мело Л., Мурзин К., Модич М., Ахонен М., Аскью П., Пападопулос Т., Адлхарт К., Крейнс Ф.Р.Л. Данн С.П. и соавт. Биоинженерия. 2017 2 ноября; 8 (6): 679-685. дои: 10.1080/21655979.2017.1323593. Epub 2017 19 мая. Биоинженерия. 2017. PMID: 28453429 Бесплатная статья ЧВК.
Посмотреть все похожие статьи
Цитируется
-
Бесклеточные тканевые сосудистые трансплантаты малого диаметра: биоматериалы и технологии производства для достижения подходящих механических свойств.
Родригес-Сото М.А., Полания-Сандовал К.А., Арагон-Ривера А.М., Буитраго Д., Аяла-Веласкес М., Веландия-Санчес А., Перальта Пелуффо Г., Круз Х.К., Муньос Камарго К., Камачо-Маккензи Х., Баррера-Карвахаль Х.Г. , Брисеньо Х.С. Родригес-Сото М.А. и соавт. Полимеры (Базель). 2022 23 августа; 14 (17): 3440. дои: 10.3390/polym14173440. Полимеры (Базель). 2022. PMID: 36080517 Бесплатная статья ЧВК. Обзор.
-
Взгляд на разработку и использование альтернатив антибиотикостимуляторам роста в птицеводстве и свиноводстве.
MRT Rahman, Fliss I, Biron E. Рахман МРТ и др. Антибиотики (Базель). 2022 2 июня; 11 (6): 766. doi: 10.3390/антибиотики11060766. Антибиотики (Базель). 2022. PMID: 35740172 Бесплатная статья ЧВК. Обзор.
-
Наноразмерные медные и серебряные тонкопленочные системы демонстрируют разные противовирусные и антибактериальные свойства.
Мейстер Т.Л., Фортманн Дж., Брайш М., Сенгсток С., Штайнманн Э., Келлер М., Пфендер С., Людвиг А. Мейстер Т.Л. и др. Научный представитель 2022 г. 3 мая; 12 (1): 7193. doi: 10.1038/s41598-022-11212-w. Научный представитель 2022. PMID: 35505071 Бесплатная статья ЧВК.
-
Быстрая антибактериальная активность материалов на основе анодированного алюминия, пропитанных четвертичными аммониевыми соединениями, для поверхностей, к которым часто прикасаются, для ограничения передачи патогенных бактерий.
Янн Дж., Древель О., Чен С.Г., Оклер-Гилберт М., Суси Г., Фошо Н., Фортье Л.С. Янн Дж. и др. RSC Adv. 2021 26 ноября; 11 (60): 38172-38188. doi: 10.1039/d1ra07159a. Электронная коллекция 2021 23 ноября. RSC Adv. 2021. PMID: 35498065 Бесплатная статья ЧВК.
-
Синтез и исследование антибактериальной активности тонких пленок на основе TiO 2 -Ag и SiO 2 -Ag с возможным применением в медицинской среде.
Адочице К., Вителару К., Парау А.С., Кисс А.Э., Панэ И., Владеску А., Костинаш С., Мога М., Мунтян Р., Бадеа М., Идомир М. Adochițe CȘ, et al. Наноматериалы (Базель). 2022 9 марта; 12 (6): 902. doi: 10.3390/nano12060902. Наноматериалы (Базель). 2022. PMID: 35335715 Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Совместная селекция устойчивости к антибиотикам, биоцидам и тяжелым металлам и ее значение для патогенов пищевого происхождения
1. СЦЕНИГР. Оценка эффектов антибиотикорезистентности биоцидов. Научный комитет по возникающим и недавно выявленным рискам для здоровья, Европейская комиссия; Брюссель, Бельгия: 2009. стр. 1–87. [Google Scholar]
2. Тума Х. Н. Устойчивость бактерий к биоцидам. Дж. Чемотер. 2009;21:5–15. [PubMed] [Google Scholar]
3. Шелдон А.Т. Антисептическая «резистентность»: реальная или воображаемая угроза? клин. Заразить. Дис. 2005;40:1650–1656. дои: 10.1086/430063. [PubMed] [CrossRef] [Академия Google]
4. Fraser C.M., Bergeron J.A., Mays A., Aiello S.E., редакторы. Ветеринарное руководство Мерк. Мерк и Ко; Rahway, NJ, USA: 1991. Часть VI: Фармакология; п. 1472. [Google Scholar]
5. Даффус Дж.Х. «Тяжелые металлы» — бессмысленный термин? (Технический отчет IUPAC) Pure Appl. хим. 2002; 74: 793–807. [Google Scholar]
6. Seiler C., Berendonk T.U. Совместная селекция устойчивости к антибиотикам, обусловленная тяжелыми металлами, в почве и водоемах, на которые воздействует сельское хозяйство и аквакультура. Фронт. микробиол. 2012;3 doi: 10.3389/fmicb.2012.00399. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
7. Barillo D. J., Marx D.E. Серебро в медицине: краткая история с 335 г. до н.э. по настоящее время. Бернс Дж. Междунар. соц. сжечь инж. 2014; 40 (Приложение S1): S3–S8. doi: 10.1016/j.burns.2014.09.009. [PubMed] [CrossRef] [Google Scholar]
8. Сильвер С. Бактериальная резистентность к серебру: Молекулярная биология, использование и неправильное использование соединений серебра. ФЭМС микробиол. 2003; 27:341–353. doi: 10.1016/S0168-6445(03)00047-0. [PubMed] [CrossRef] [Академия Google]
9. Бейт Н., Хури-Хаддад Ю., Домб А., Хан В., Хазан Р., Бейт Н., Хури-Хаддад Ю., Домб А., Хан В., Хазан Р. Альтернативный антимикробный подход : Нано-антимикробные материалы. Эвид. Дополнение на основе. Альтерн. Мед. 2015; 2015: e246012. дои: 10.1155/2015/246012. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10. Норвежский научный комитет по безопасности пищевых продуктов. Цинк и медь в свиноводстве и птицеводстве — судьба и влияние на пищевую цепь и окружающую среду. Норвежское управление по безопасности пищевых продуктов; Осло, Норвегия: 2014. с. 128. [Google Академия]
11. Хьюго В.Б. Краткая история тепловой, химической и радиационной консервации и дезинфекции. Междунар. Биодекор. биодеград. 1995; 36: 197–217. doi: 10.1016/0964-8305(95)00055-0. [CrossRef] [Google Scholar]
12. Моренте Э.О., Фернандес-Фуэнтес М.А., Бургос М.Дж.Г., Абриуэль Х., Пулидо Р.П., Гальвез А. Толерантность бактерий к биоцидам. Междунар. Дж. Пищевая микробиология. 2013; 162:13–25. doi: 10.1016/j.ijfoodmicro.2012.12.028. [PubMed] [CrossRef] [Google Scholar]
13. Cogliani C., Goossens H., Greko C. Ограничение использования противомикробных препаратов у сельскохозяйственных животных: уроки из Европы. Микроб. 2011; 6: 274–279.. doi: 10.1128/микроб.6.274.1. [CrossRef] [Google Scholar]
14. Fernandez-Rubio C., Ordonez C., Abad-Gonzalez J., Garcia-Gallego A., Honrubia M.P., Mallo J.J., Balana-Fouse R. Кормовые добавки на основе масляной кислоты помогают защитить цыплят-бройлеров от инфекции Salmonella Enteritidis. Поулт. науч. 2009; 88: 943–948. doi: 10.3382/ps.2008-00484. [PubMed] [CrossRef] [Google Scholar]
15. Franz C., Baser K., Windisch W. Эфирные масла и ароматические растения в кормлении животных — европейская перспектива. Обзор. Аромат Фрагр. Дж. 2010; 25:327–340. дои: 10.1002/ffj.1967. [CrossRef] [Google Scholar]
16. Leeson S., Namkung H., Antongiovanni M., Lee E.H. Влияние масляной кислоты на продуктивность и убойный выход цыплят-бройлеров. Поулт. науч. 2005; 84: 1418–1422. doi: 10.1093/ps/84.9.1418. [PubMed] [CrossRef] [Google Scholar]
17. Zeng Z., Zhang S., Wang H., Piao X. Эфирные масла и ароматические растения в качестве кормовых добавок в кормлении нежвачных животных: обзор. Дж. Аним. науч. Биотехнолог. 2015; 6 doi: 10.1186/s40104-015-0004-5. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18. Дэнс Д.А.Б., Пирсон А.Д., Сил Д.В., Лоус Дж.А. Вспышка в больнице, вызванная устойчивым к хлоргексидину и антибиотикам Proteus mirabilis . Дж. Хосп. Заразить. 1987; 10:10–16. doi: 10.1016/0195-6701(87)
19. Griffiths P.A., Babb J.R., Bradley C.R., Fraise A.P. Устойчивые к глутаральдегиду Mycobacterium chelonae из дезинфекторов для мойки эндоскопов. Дж. Заявл. микробиол. 1997; 82: 519–526. дои: 10.1046/j.1365-2672.1997.00171.х. [PubMed] [CrossRef] [Google Scholar]
20. Sasatsu M., Shimizu K., Noguchi N., Kono M. Устойчивый к триклозану Staphylococcus aureus . Ланцет. 1993; 341 doi: 10.1016/0140-6736(93)-M. [PubMed] [CrossRef] [Google Scholar]
21. Александр Д.М., Джевон Р.Х., Персад С. Устойчивость к дезинфицирующим средствам у устойчивых к антибиотикам организмов. С. Афр. J. Sci. 1991; 87: 614–617. [Google Scholar]
22. Стиклер Д.Дж., Томас Б. Устойчивость к антисептикам и антибиотикам грамотрицательных бактерий, вызывающих инфекцию мочевыводящих путей. Дж. Клин. Патол. 1980;33:288–296. doi: 10.1136/jcp.33.3.288. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Fraise A.P. Злоупотребление биоцидами и устойчивость к противомикробным препаратам — повод для беспокойства? Дж. Антимикроб. Чемотер. 2002; 49:11–12. doi: 10.1093/jac/49.1.11. [PubMed] [CrossRef] [Google Scholar]
24. Рассел А.Д. Выбирают ли биоциды устойчивость к антибиотикам? Дж. Фарм. Фармакол. 2000; 52: 227–233. doi: 10.1211/0022357001773742. [PubMed] [CrossRef] [Google Scholar]
25. Блумфилд С.Ф. Значение использования биоцидов и устойчивости к противомикробным препаратам в домашних условиях. Дж. Заявл. микробиол. 2002;92:144С–157С. doi: 10.1046/j.1365-2672.92.5s1.15.x. [PubMed] [CrossRef] [Google Scholar]
26. Gilbert P., McBain A.J. Потенциальное влияние более широкого использования биоцидов в потребительских товарах на распространенность устойчивости к антибиотикам. клин. микробиол. 2003; 16:189–208. doi: 10.1128/CMR.16.2.189-208.2003. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
27. Antunes P., Mourao J., Pestana N., Peixe L. Утечка появляющейся клинически значимой полирезистентной сальмонеллы с множественной лекарственной устойчивостью клонов со свиноферм. Дж. Антимикроб. Чемотер. 2011;66:2028–2032. doi: 10.1093/jac/dkr228. [PubMed] [CrossRef] [Google Scholar]
28. Davies R.H., Dalziel R., Gibbens J.C., Wilesmith J.W., Ryan JMB, Evans S.J., Byrne C., Paiba G.A., Pascoe S.J.S., Teale C.J. Национальное исследование Salmonella у свиней, крупного рогатого скота и овец на бойне в Великобритании (1999–2000 гг.) J. Appl. микробиол. 2004; 96: 750–760. doi: 10.1111/j.1365-2672.2004.02192.x. [PubMed] [CrossRef] [Академия Google]
29. Группа экспертов EFSA по биологическим опасностям (BIOHAZ) Научное заключение о мониторинге и оценке риска для здоровья населения штаммов « Salmonella Typhimurium-подобных». EFSA J. 2010; 8: 1826–1874. [Google Scholar]
30. Cuny C., Friedrich A., Kozytska S., Layer F., Nübel U., Ohlsen K., Strommenger B., Walther B., Wieler L., Witte W. Появление метициллина -устойчивый Staphylococcus aureus (MRSA) у различных видов животных. Междунар. Дж. Мед. микробиол. 2010;300:109–117. doi: 10.1016/j.ijmm.2009.11.002. [PubMed] [CrossRef] [Google Scholar]
31. Blaak H., van Hoek A.H., Hamidjaja R.A., van der Plaats R.Q.J., de Heer K.L., de Roda Husman A.M., Schets F.M. Распространение, численность и разнообразие продуцирующих БЛРС штаммов E. coli в среде птицефабрики. ПЛОС ОДИН. 2015;10:e0135402. doi: 10.1371/journal.pone.0135402. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Chopra I., Hesse L., O’Neill A.J. Использование современного понимания действия антибиотиков для открытия новых лекарств. Дж. Заявл. микробиол. 2002;92:4С–15С. doi: 10.1046/j.1365-2672.92.5s1.13.x. [PubMed] [CrossRef] [Google Scholar]
33. Серф О., Карпентьер Б., Сандерс П. Тесты для определения используемых концентраций антибиотиков и дезинфицирующих средств основаны на совершенно разных концепциях: «Устойчивость» имеет разное значение. Междунар. Дж. Пищевая микробиология. 2010; 136: 247–254. doi: 10.1016/j.ijfoodmicro.2009.10.002. [PubMed] [CrossRef] [Google Scholar]
34. Пул К. Механизмы устойчивости бактерий к биоцидам и антибиотикам. Дж. Заявл. микробиол. 2002;92:55С–64С. doi: 10.1046/j.1365-2672.92.5s1.8.x. [PubMed] [CrossRef] [Google Scholar]
35. Кохански М.А., ДеПристо М.А., Коллинз Дж.Дж. Сублетальное лечение антибиотиками приводит к множественной лекарственной устойчивости посредством радикального мутагенеза. Мол. Клетка. 2010; 37: 311–320. doi: 10.1016/j.molcel.2010.01.003. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Maillard J.Y. Бактериальные мишени для действия биоцида. Дж. Заявл. микробиол. 2002; 92:16С–27С. doi: 10.1046/j.1365-2672.92.5s1.3.x. [PubMed] [CrossRef] [Академия Google]
37. Макдоннелл Г., Рассел А.Д. Антисептики и дезинфицирующие средства: активность, действие и устойчивость. клин. микробиол. 1999; 12:147–179. [Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Рассел А.Д. Использование биоцидов и устойчивость к антибиотикам: актуальность лабораторных данных для клинических и экологических ситуаций. Ланцет Инфекция. Дис. 2003; 3: 794–803. doi: 10.1016/S1473-3099(03)00833-8. [PubMed] [CrossRef] [Google Scholar]
39. Buffet-Bataillon S., Tattevin P., Bonnaure-Mallet M., Jolivet-Gougeon A. Возникновение устойчивости к антибактериальным агентам: роль соединений четвертичного аммония-a критический обзор. Междунар. Дж. Антимикроб. Агенты. 2012;39: 381–389. doi: 10.1016/j.ijantimicag.2012.01.011. [PubMed] [CrossRef] [Google Scholar]
40. Davin-Regli A., Pagès JM Перекрестная устойчивость между биоцидами и противомикробными препаратами: возникающий вопрос. преподобный наук. Технол. Выключенный. Междунар. Эпизоот. 2012; 31:89–104. [PubMed] [Google Scholar]
41. Кэри Д. Э., Макнамара П. Дж. Влияние триклозана на распространение устойчивости к антибиотикам в окружающей среде. Фронт. микробиол. 2015; 5 doi: 10.3389/fmicb.2014.00780. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Rensch U., Klein G., Kehrenberg C. Анализ отобранных триклозаном мутантов Salmonella enterica восьми сероваров показал повышенную чувствительность к аминогликозидам и снижение скорости роста. ПЛОС ОДИН. 2013;8:e78310. doi: 10.1371/journal.pone.0078310. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
43. Suller M.T.E., Russell A.D. Триклозан и устойчивость к антибиотикам у Staphylococcus aureus . Дж. Антимикроб. Чемотер. 2000;46:11–18. дои: 10.1093/як/46.1.11. [PubMed] [CrossRef] [Google Scholar]
44. Маври А., Мозина С.С. Развитие устойчивости к противомикробным препаратам у Campylobacter jejuni и Campylobacter coli , адаптированных к биоцидам. Междунар. Дж. Пищевая микробиология. 2013; 160:304–312. doi: 10.1016/j.ijfoodmicro.2012.11.006. [PubMed] [CrossRef] [Google Scholar]
45. Medardus J.J., Molla B.Z., Nicol M., Morrow W.M., Rajala-Schultz P.J., Kazwala R., Gebreyes W.A. Использование микроэлементов тяжелых металлов в кормах в свиноводстве США систем и его роль в сохранении множественной лекарственной устойчивости Сальмонелла . заявл. Окружающая среда. микробиол. 2014;80:2317–2325. doi: 10. 1128/AEM.04283-13. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
46. Николсон Ф.А., Смит С.Р., Аллоуэй Б.Дж., Карлтон-Смит С., Чемберс Б.Дж. Перечень поступления тяжелых металлов в сельскохозяйственные почвы в Англии и Уэльсе. науч. Общая окружающая среда. 2003; 311: 205–219. doi: 10.1016/S0048-9697(03)00139-6. [PubMed] [CrossRef] [Google Scholar]
47. Berg J., Thorsen M.K., Holm P.E., Jensen J., Nybroe O., Brandt K.K. Воздействие Cu в полевых условиях способствует одновременному отбору устойчивости к антибиотикам, что определяется с помощью нового независимого от культивирования анализа устойчивости бактериального сообщества. Окружающая среда. науч. Технол. 2010;44:8724–8728. дои: 10.1021/es101798р. [PubMed] [CrossRef] [Google Scholar]
48. Хао X., Луэтье Ф.Л., Цинь Ю., Макдевитт С.Ф., Лутай Н., Хобман Дж.Л., Азиани К., Сончини Ф.К., Герман Н., Чжан С., и другие. Выживание амебы — основное давление отбора на присутствие бактериальных детерминант устойчивости к меди и цинку? Выявление «медного острова патогенности» Заявл. микробиол. Биотехнолог. 2015;99:5817–5824. doi: 10.1007/s00253-015-6749-0. [PubMed] [CrossRef] [Google Scholar]
49. Yazdankhah S., Rudi K., Bernhoft A. Цинк и медь в кормах для животных — развитие резистентности и корезистентности к противомикробным агентам у бактерий животного происхождения. микроб. Экол. Здоровье Дис. 2014; 25 doi: 10.3402/mehd.v25.25862. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Хасман Х., Кемпф И., Чидайн Б., Кариолет Р., Эрсбёлль А.К., Хоуэ Х., Хансен Х.К.Б., Аареструп Ф.М. Устойчивость к меди у Enterococcus faecium , опосредованная геном tcrB , выбирается путем добавления в корм для свиней сульфата меди. заявл. Окружающая среда. микробиол. 2006; 72: 5784–5789. doi: 10.1128/AEM.02979-05. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51. Шелтон Н.В., Джейкоб М.Е., Токач М.Д., Нельсен Дж.Л., Гудбанд Р.Д., Дритц С.С., де Ручи Дж.М., Амачавади Р.Г., Ши X., Нагараджа Т.Г. Влияние сульфата меди, оксида цинка и неотеррамицина на рост поросят-отъемышей и уровень устойчивости к антибиотикам в фекалиях Кишечная палочка . В: Goodband B., Tokach M., Dritz S., de Rouchey J., редакторы. День свиньи в Университете штата Канзас, 2009 г. Отчет о проделанной работе 1020. Университет штата Канзас; Манхэттен, Канзас, США: 2009. стр. 73–79. [Google Scholar]
52. Бейкер-Остин С., Райт М.С., Степанаускас Р., Макартур Дж. В. Совместный отбор устойчивости к антибиотикам и металлам. Тенденции микробиол. 2006; 14: 176–182. doi: 10.1016/j.tim.2006.02.006. [PubMed] [CrossRef] [Google Scholar]
53. Аареструп Ф.М., Хасман Х. Восприимчивость различных видов бактерий, выделенных от пищевых животных, к сульфату меди, хлориду цинка и антимикробным веществам, используемым для дезинфекции. Вет. микробиол. 2004; 100:83–89. doi: 10.1016/j.vetmic.2004.01.013. [PubMed] [CrossRef] [Google Scholar]
54. Зильбергельд Э.К., Нахман К. Риски для окружающей среды и здоровья населения, связанные с использованием мышьяка в кормах для животных. Анна. Н. Я. акад. науч. 2008;1140:346–357. doi: 10.1196/annals.1454.049. [PubMed] [CrossRef] [Google Scholar]
55. Европейский комитет по тестированию чувствительности к противомикробным препаратам (EUCAST) Европейского общества клинической микробиологии и инфекционных заболеваний (ESCMID) Терминология, относящаяся к методам определения чувствительности бактерий к противомикробным препаратам . клин. микробиол. Заразить. 2000; 6: 503–508. [PubMed] [Академия Google]
56. Мёретро Т., Хейр Э., Нессе Л.Л., Вестби Л.К., Лангсруд С. Борьба с Salmonella в пищевой среде с помощью химической дезинфекции. Еда Рез. Междунар. 2012; 45: 532–544. doi: 10.1016/j.foodres.2011.02.002. [CrossRef] [Google Scholar]
57. Suller M.T.E., Russell A.D. Устойчивость к антибиотикам и биоцидам у метициллин-резистентного Staphylococcus aureus и ванкомицин-резистентного энтерококка. Дж. Хосп. Заразить. 1999; 43: 281–291. дои: 10.1016/S0195-6701(99)-3. [PubMed] [CrossRef] [Google Scholar]
58. Capita R., Riesco-Peláez F., Alonso-Hernando A., Alonso-Calleja C. Воздействие Escherichia coli ATCC 12806 на сублетальные концентрации пищевых продуктов биоциды влияют на его способность образовывать биопленку, устойчивость к противомикробным препаратам и ультраструктуру. заявл. Окружающая среда. микробиол. 2014;80:1268–1280. doi: 10.1128/AEM.02283-13. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
59. Lear J.C., Maillard J.Y., Dettmar P.W., Goddard P.A., Russell A.D. Бактерии, устойчивые к хлороксиленолу и триклозану из промышленных источников — чувствительность к антибиотикам и другим биоцидам . Междунар. Биодекор. биодеград. 2006; 57: 51–56. doi: 10.1016/j.ibiod.2005.11.002. [Перекрестная ссылка] [Академия Google]
60. Табак М., Шер К., Хартог Э., Ромлинг У., Мэтьюз К.Р., Чикиндас М.Л., Ярон С. Влияние триклозана на Salmonella typhimurium на разных стадиях роста и в биопленках. ФЭМС микробиол. лат. 2007; 267: 200–206. doi: 10.1111/j.1574-6968.2006.00547.x. [PubMed] [CrossRef] [Google Scholar]
61. Беднорц С., Эльгешлегер К., Киннеманн Б., Хартманн С., Нойманн К., Пипер Р., Бете А., Земмлер Т., Тедин К., Ширак П. и др. Более широкий контекст устойчивости к антибиотикам: добавление цинка в корм поросят увеличивает долю полирезистентных Escherichia coli in vivo . Междунар. Дж. Мед. микробиол. 2013; 303: 396–403. doi: 10.1016/j.ijmm.2013.06.004. [PubMed] [CrossRef] [Google Scholar]
62. Condell O., Iversen C., Cooney S., Power K.A., Walsh C., Burgess C., Fanning S. Эффективность биоцидов, используемых в современной пищевой промышленности для контроль Salmonella enterica , а также связи между толерантностью к биоцидам и устойчивостью к клинически значимым противомикробным соединениям. заявл. Окружающая среда. микробиол. 2012;78:3087–3097. doi: 10.1128/AEM.07534-11. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
63. Рассел А.Д. Механизмы устойчивости бактерий к биоцидам. Междунар. Биодекор. биодеград. 1995; 36: 247–265. doi: 10.1016/0964-8305(95)00056-9. [CrossRef] [Google Scholar]
64. Oggioni M.R., Furi L., Coelho J.R., Maillard J.Y., Martinez J.L. Последние достижения в области потенциальной взаимосвязи между устойчивостью к противомикробным препаратам к биоцидам и антибиотикам. Эксперт преподобный Anti Infect. тер. 2013; 11: 363–366. doi: 10.1586/eri.13.16. [PubMed] [CrossRef] [Google Scholar]
65. Gilbert P., Allison D.G., McBain A.J. Биопленки in vitro и in vivo : Подразумевают ли сингулярные механизмы перекрестную резистентность? Дж. Заявл. микробиол. 2002;92:98С–110С. doi: 10.1046/j.1365-2672.92.5s1.5.x. [PubMed] [CrossRef] [Google Scholar]
66. МакБейн А.Дж., Гилберт П. Толерантность к биоцидам и предвестники гибели. Междунар. Биодекор. биодеград. 2001; 47: 55–61. doi: 10.1016/S0964-8305(01)00037-3. [CrossRef] [Google Scholar]
67. Pagedar A., Singh J., Batish V.K. Адаптация к хлориду бензалкония и ципрофлоксацину влияет на потенциал образования биопленки, насос оттока и активность гемолизина Escherichia coli молочного происхождения. Дж. Молочная Рез. 2012;79:383–389. doi: 10.1017/S00220290295. [PubMed] [CrossRef] [Google Scholar]
68. Шеридан А., Ленахан М., Конделл О., Бонилья-Сантьяго Р., Сержант К., Рено Дж., Даффи Г. , Фаннинг С., Нэлли Дж.Э. , Берджесс К.М. Протеомный и фенотипический анализ толерантных к триклозану вероцитотоксигенных штаммов Escherichia coli O157:h29. Дж. Протеомика. 2013;80:78–90. [PubMed] [Google Scholar]
69. Пул К. Эффлюксные помпы как механизмы устойчивости к противомикробным препаратам. Анна. Мед. 2007;39: 162–176. doi: 10.1080/078538
195262. [PubMed] [CrossRef] [Google Scholar]70. Hegstad K., Langsrud S., Lunestad B.T., Scheie A.A., Sunde M., Yazdankhah S.P. Способствует ли широкое использование соединений четвертичного аммония селекции и распространению устойчивости к противомикробным препаратам? и тем самым угрожать нашему здоровью? микроб. Сопротивление наркотикам. 2010; 16:91–104. doi: 10.1089/mdr.2009.0120. [PubMed] [CrossRef] [Google Scholar]
71. Huet A.A., Raygada J.L., Mendiratta K., Seo S.M., Kaatz G.W. Сверхэкспрессия множественного лекарственного эффлюксного насоса у Staphylococcus aureus после однократного и многократного воздействия in vitro биоцидов и красителей. микробиол. СГМ. 2008; 154:3144–3153. doi: 10.1099/mic.0.2008/021188-0. [PubMed] [CrossRef] [Google Scholar]
72. Путман М., Ван Вин Х. В., Конингс В. Н. Молекулярные свойства бактериальных переносчиков многих лекарств. микробиол. Мол. биол. 2000; 64: 672–693. doi: 10.1128/MMBR.64.4.672-693.2000. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
73. Oethinger M., Kern W.V., Jellen-Ritter A.S., McMurry L.M., Levy S.B. Неэффективность мутаций топоизомеразы в опосредовании клинически значимой резистентности к фторхинолонам у Escherichia coli в отсутствие эффлюксной помпы AcrAB. Антимикроб. Агенты Чемотер. 2000;44:10–13. doi: 10.1128/AAC.44.1.10-13.2000. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
74. Jaglic Z., Cervinkova D. Генетическая основа устойчивости к четвертичным аммониевым соединениям — гены qac и их роль: Обзор. Вет. Мед. 2012;57:275–281. [Google Scholar]
75. Gilbert P., McBain A.J., Bloomfield S.F. Злоупотребление биоцидами и устойчивость к противомикробным препаратам: ясность в отношении проблем. Дж. Антимикроб. Чемотер. 2002; 50: 137–139.. doi: 10.1093/jac/dkf071. [PubMed] [CrossRef] [Google Scholar]
76. Langsrud S., Sundheim G., Holck A.L. Перекрестная устойчивость к антибиотикам Escherichia coli , адаптированная к хлориду бензалкония или подверженная воздействию индукторов стресса. Дж. Заявл. микробиол. 2004; 96: 201–208. doi: 10.1046/j.1365-2672.2003.02140.x. [PubMed] [CrossRef] [Google Scholar]
77. Trias J., Benz R. Проницаемость клеточной стенки Mycobacterium smegmatis . Мол. микробиол. 1994; 14: 283–290. дои: 10.1111/j.1365-2958.1994.tb01289.х. [PubMed] [CrossRef] [Google Scholar]
78. Karatzas K.A.G., Randall L.P., Webber M., Piddock L.J.V., Humphrey T.J., Woodward M.J., Coldham N.G. Фенотипическая и протеомная характеристика множественно устойчивых к антибиотикам вариантов серовара Typhimurium Salmonella enterica , отобранных после воздействия дезинфицирующих средств. заявл. Окружающая среда. микробиол. 2008; 74: 1508–1516. doi: 10.1128/AEM.01931-07. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
79. Гркович С., Браун М.Х., Скуррей Р.А. Регулирование систем экспорта бактериальных препаратов. микробиол. Мол. биол. 2002; 66: 671–701. doi: 10.1128/MMBR.66.4.671-701.2002. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
80. Oethinger M., Podglajen I., Kern W.V., Levy S.B. Сверхэкспрессия регуляторного гена marA или soxS у клинических мутантов топоизомеразы Escherichia coli . Антимикроб. Агенты Чемотер. 1998;42:2089–2094. [Бесплатная статья PMC] [PubMed] [Google Scholar]
81. Hao Z., Lou H., Zhu R., Zhu J., Zhang D., Zhao B.S., Zeng S., Chen X., Chan J., He C., et al. Регулятор множественной устойчивости к антибиотикам MarR представляет собой медный сенсор в Escherichia coli . Нац. хим. биол. 2014;10:21–28. doi: 10.1038/nchembio.1380. [PubMed] [CrossRef] [Google Scholar]
82. Lee H.H., Molla M.N., Cantor C.R. , Collins J.J. Бактериальная благотворительность приводит к сопротивлению всего населения. Природа. 2010; 467:82–85. doi: 10.1038/nature09354. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
83. Трелфолл Э.Дж. Epidemic Salmonella typhimurium DT 104 — действительно международный мультирезистентный клон. Дж. Антимикроб. Чемотер. 2000;46:7–10. doi: 10.1093/jac/46.1.7. [PubMed] [CrossRef] [Google Scholar]
84. Ciric L., Mullany P., Roberts A.P. Гены устойчивости к антибиотикам и антисептикам связаны с новым мобильным генетическим элементом: Tn6087. Дж. Антимикроб. Чемотер. 2011;66:2235–2239. doi: 10.1093/jac/dkr311. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
85. Караттоли А. Значение интегронов в распространении резистентности. Вет. Рез. 2001; 32 doi: 10.1051/vetres:2001122. [PubMed] [CrossRef] [Google Scholar]
86. Paulsen I.T., Littlejohn T.G., Rådström P., Sundström L., Sköld O., Swedberg G., Skurray R.A. 3′-консервативный сегмент интегронов содержит ген, связанный с множественной лекарственной устойчивостью к антисептикам и дезинфицирующим средствам. Антимикроб. Агенты Чемотер. 1993; 37: 761–768. doi: 10.1128/AAC.37.4.761. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
87. Agga G.E., Scott H.M., Amachawadi R.G., Nagaraja T.G., Vinasco J., Bai J., Norby B., Renter D.G., Dritz S.S., Nelssen J.L., et al. Влияние добавок хлортетрациклина и меди на устойчивость к противомикробным препаратам фекальной Escherichia coli поросят-отъемышей. Пред. Вет. Мед. 2014; 114: 231–246. doi: 10.1016/j.prevetmed.2014.02.010. [PubMed] [CrossRef] [Google Scholar]
88. Gullberg E., Albrecht L.M., Karlsson C., Sandegren L., Andersson D.I. Селекция плазмиды множественной лекарственной устойчивости по сублетальным уровням антибиотиков и тяжелых металлов. МБИО. 2014; 5 doi: 10.1128/mBio.01918-14. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
89. Randall LP, Cooles SW, Piddock LJV, Woodward MJ Влияние триклозана или фенольного дезинфицирующего средства на ферме на селекцию устойчивых к антибиотикам Salmonella enterica . Дж. Антимикроб. Чемотер. 2004; 54: 621–627. doi: 10.1093/jac/dkh476. [PubMed] [CrossRef] [Google Scholar]
90. Kastbjerg V.G., Hein-Kristensen L., Gram L. Индуцированная триклозаном устойчивость к аминогликозидам Listeria monocytogenes изолятов могут проявляться в виде небольших колоний. Антимикроб. Агенты Чемотер. 2014;58:3124–3132. doi: 10.1128/AAC.02266-13. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
91. Guo M.T., Yuan Q.B., Yang J. Отличительные эффекты воздействия ультрафиолета и хлорирования на горизонтальный перенос генов устойчивости к антибиотикам в муниципальных сточных водах. Окружающая среда. науч. Технол. 2015; 49: 5771–5778. doi: 10.1021/acs.est.5b00644. [PubMed] [CrossRef] [Google Scholar]
92. Ренсинг К., Грасс Г. Escherichia coli механизмы гомеостаза меди в изменяющейся среде. ФЭМС микробиол. 2003; 27:197–213. doi: 10.1016/S0168-6445(03)00049-4. [PubMed] [CrossRef] [Google Scholar]
93. Бирд С.Дж., Чикконьяни Д. Т., Хьюз М.Н., Пул Р.К. Катализируемый ионами металла гидролиз ампициллина в микробиологических питательных средах. ФЭМС микробиол. лат. 1992; 96: 207–211. doi: 10.1111/j.1574-6968.1992.tb05418.x. [PubMed] [CrossRef] [Google Scholar]
94. Hölzel C.S., Müller C., Harms K.S., Mikolajewski S., Schäfer S., Schwaiger K., Bauer J. Тяжелые металлы в жидком свином навозе в свете бактериальных противомикробных препаратов сопротивление. Окружающая среда. Рез. 2012; 113:21–27. doi: 10.1016/j.envres.2012.01.002. [PubMed] [CrossRef] [Академия Google]
95. Мукерджи Г., Гош Т. Взаимодействие ионов металлов с пенициллинами — Часть VII: Образование смешанных лигандных комплексов кобальта (II), никеля (II), меди (II) и цинка (II) с ампициллином и нуклеиновой кислотой. базы. Дж. Неорг. Биохим. 1995; 59: 827–833. doi: 10.1016/0162-0134(94)00082-L. [PubMed] [CrossRef] [Google Scholar]
96. Рутала В.А., Штигель М.М., Сарубби Ф.А., Вебер Д.Дж. Чувствительность антибиотикочувствительных и антибиотикорезистентных больничных бактерий к дезинфицирующим средствам. Заразить. Хосп. Эпидемиол. 1997;18:417–421. doi: 10.2307/30141249. [PubMed] [CrossRef] [Google Scholar]
97. Макларен И., Уэльс А., Бреслин М., Дэвис Р. Оценка широко используемых дезинфицирующих средств на фермах во влажных и сухих моделях заражения ферм Salmonella . Авиан Патол. 2011;40:33–42. doi: 10.1080/03079457.2010.537303. [PubMed] [CrossRef] [Google Scholar]
98. Холман Д.Б., Шенье М.Р. Временные изменения и влияние субтерапевтических концентраций антибиотиков на микробиоту кишечника свиней. ФЭМС микробиол. Экол. 2014;90: 599–608. doi: 10.1111/1574-6941.12419. [PubMed] [CrossRef] [Google Scholar]
99. Stepanauskas R., Glenn T.C., Jagoe C.H., Tuckfield RC, Lindell AH, King CJ, McArthur J.V. Совместная селекция микробной устойчивости к металлам и антибиотикам в пресноводных микрокосмах. Окружающая среда. микробиол. 2006; 8: 1510–1514. doi: 10.1111/j.1462-2920.2006.01091.x. [PubMed] [CrossRef] [Google Scholar]
100. Tandukar M., Oh S., Tezel U. , Konstantinidis K.T., Pavlostathis S.G. Длительное воздействие дезинфицирующих средств на основе хлорида бензалкония приводит к изменению структуры микробного сообщества и повышению устойчивости к противомикробным препаратам. . Окружающая среда. науч. Технол. 2013;47:9730–9738. doi: 10.1021/es401507k. [PubMed] [CrossRef] [Google Scholar]
101. Yazdankhah S.P., Scheie A.A., Høiby E.A., Lunestad B.T., Heir E., Fotland T.Ø., Naterstad K., Kruse H. Триклозан и устойчивость к противомикробным препаратам у бактерий: Обзор. микроб. Сопротивление наркотикам. 2006; 12:83–90. doi: 10.1089/mdr.2006.12.83. [PubMed] [CrossRef] [Google Scholar]
102. Buffet-Bataillon S., Branger B., Cormier M., Bonnaure-Mallet M., Jolivet-Gougeon A. Эффект более высоких минимальных ингибирующих концентраций соединений четвертичного аммония в клинический изолятов E. coli на чувствительность к антибиотикам и клинические исходы. Дж. Хосп. Заразить. 2011;79:141–146. doi: 10.1016/j.jhin.2011.06. 008. [PubMed] [CrossRef] [Google Scholar]
103. Buffet-Bataillon S., le Jeune A., le Gall-David S., Bonnaure-Mallet M., Jolivet-Gougeon A. Молекулярные механизмы более высоких МПК антибиотиков и соединения четвертичного аммония для Escherichia coli , выделенные из бактериемии. Дж. Антимикроб. Чемотер. 2012;67:2837–2842. doi: 10.1093/jac/dks321. [PubMed] [CrossRef] [Академия Google]
104. Willinghan E.M., Sander J.E., Thayer S.G., Wilson J.L. Исследование устойчивости бактерий к дезинфицирующим средствам инкубатория. Авиан Дис. 1996; 40: 510–515. дои: 10.2307/15. [PubMed] [CrossRef] [Google Scholar]
105. Мартин Х., Марис П. Резистентность 310 грамположительных штаммов, выделенных из вымени дойной коровы. Вет. Рез. 1995; 26:43–56. [PubMed] [Google Scholar]
106. Lear J.C., Maillard J.Y., Dettmar P.W., Goddard P.A., Russell AD. Бактерии, устойчивые к хлороксиленолу и триклозану, из промышленных источников. J. Ind. Microbiol. Биотехнолог. 2002;29: 238–242. doi: 10.1038/sj.jim.7000320. [PubMed] [CrossRef] [Google Scholar]
107. Braoudaki M., Hilton A.C. Адаптивная устойчивость к биоцидам у Salmonella enterica и Escherichia coli O157 и перекрестная устойчивость к противомикробным агентам. Дж. Клин. микробиол. 2004; 42:73–78. doi: 10.1128/JCM.42.1.73-78.2004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
108. Ledder R.G., Gilbert P., Willis C., McBain A.J. Влияние хронического воздействия триклозана на чувствительность к противомикробным препаратам 40 ex-situ изолятов из окружающей среды и человека. Дж. Заявл. микробиол. 2006; 100:1132–1140. doi: 10.1111/j.1365-2672.2006.02811.x. [PubMed] [CrossRef] [Google Scholar]
109. McBain A.J., Ledder R.G., Sreenivasan P., Gilbert P. Отбор на высокий уровень устойчивости при хроническом воздействии триклозана не является универсальным. Дж. Антимикроб. Чемотер. 2004; 53: 772–777. doi: 10.1093/jac/dkh268. [PubMed] [CrossRef] [Google Scholar]
110. Шеридан А., Ленахан М., Даффи Г., Фаннинг С., Берджесс С. Потенциал устойчивости к биоцидам в Escherichia coli и ее влияние на реакцию на стрессы при обработке пищевых продуктов. Пищевой контроль. 2012;26:98–106. [Google Scholar]
111. Karatzas K.A.G., Webber M.A., Jorgensen F., Woodward M.J., Piddock L.J.V., Humphrey T.J. Продолжительное лечение Salmonella enterica серовара Typhimurium коммерческими дезинфицирующими средствами приводит к множественной устойчивости к антибиотикам, увеличению оттока и снижению инвазивности. Дж. Антимикроб. Чемотер. 2007; 60: 947–955. doi: 10.1093/jac/dkm314. [PubMed] [CrossRef] [Академия Google]
112. Alonso-Hernando A., Capita R., Prieto M., Alonso-Calleja C. Сравнение моделей устойчивости к антибиотикам у штаммов Listeria monocytogenes и Salmonella enterica , подвергшихся предварительному воздействию и подвергшихся воздействию деконтаминантов домашней птицы. Пищевой контроль. 2009;20:1108–1111. doi: 10. 1016/j.foodcont.2009.02.011. [CrossRef] [Google Scholar]
113. Soumet C., Fourreau E., Legrandois P., Maris P. Устойчивость к соединениям феникола после адаптации к соединениям четвертичного аммония в Кишечная палочка . Вет. микробиол. 2012; 158:147–152. doi: 10.1016/j.vetmic.2012.01.030. [PubMed] [CrossRef] [Google Scholar]
114. Группа экспертов EFSA по биологическим опасностям (BIOHAZ) Оценка возможного воздействия четырех антимикробных лечебных веществ на появление устойчивости к противомикробным препаратам. ЕФСА. 2008; 6 doi: 10.2903/j.efsa.2008.659. [CrossRef] [Google Scholar]
115. Потенски С.Дж., Ганди М., Мэтьюз К.Р. Воздействие на Salmonella Enteritidis хлора или пищевых консервантов повышает восприимчивость к антибиотикам Исправление: правильное название: «Воздействие на Salmonella Enteritidis к хлору или пищевым консервантам снижает восприимчивость к антибиотикам» FEMS Microbiol. лат. 2003; 220:181–186. doi: 10.1016/S0378-1097(03)00099-5. [PubMed] [CrossRef] [Google Scholar]
116. Pumbwe L., Skilbeck C.A., Wexler H.M. Индукция множественной устойчивости к антибиотикам у Bacteroides fragilis бензолом и производными бензола активными соединениями широко используемых анальгетиков, антисептиков и чистящих средств. Дж. Антимикроб. Чемотер. 2007; 60: 1288–129.7. doi: 10.1093/jac/dkm363. [PubMed] [CrossRef] [Google Scholar]
117. McMurry L.M., Oethinger M., Levy S.B. Сверхэкспрессия marA , soxS или acrAB вызывает устойчивость к триклозану лабораторных и клинических штаммов Escherichia coli . ФЭМС микробиол. лат. 1998; 166: 305–309. doi: 10.1111/j.1574-6968.1998.tb13905.x. [PubMed] [CrossRef] [Google Scholar]
118. Braoudaki M., Hilton A.C. Механизмы резистентности в Salmonella enterica адаптирован к эритромицину, бензалкония хлориду и триклозану. Междунар. Дж. Антимикроб. Агенты. 2005; 25:31–37. doi: 10.1016/j.ijantimicag.2004.07.016. [PubMed] [CrossRef] [Google Scholar]
119. Whitehead R.N., Overton T.W., Kemp C.L., Webber M.A. Воздействие высокоуровневого биоцидного заражения на Salmonella enterica serovar Typhimurium может привести к отбору мутантов с множественной лекарственной устойчивостью за один этап. ПЛОС ОДИН. 2011;6:e22833. doi: 10.1371/journal.pone.0022833. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
120. Фернандес-Фуэнтес М.А., Моренте Э.О., Абриуэль Х., Пулидо Р.П., Гальвес А. Детерминанты устойчивости к противомикробным препаратам грамотрицательных бактерий, устойчивых к антибиотикам и биоцидам из органических продуктов питания. Пищевой контроль. 2014;37:9–14. doi: 10.1016/j.foodcont.2013.08.041. [CrossRef] [Google Scholar]
121. Thorrold C.A., Letsoalo M.E., Duse AG, Marais E. Активность эффлюксной помпы в устойчивых к фторхинолонам и тетрациклинам Salmonella и E. coli , связанная со снижением чувствительности к бытовым противомикробным чистящим средствам. Междунар. Дж. Пищевая микробиология. 2007; 113:315–320. doi: 10.1016/j.ijfoodmicro.2006.08.008. [PubMed] [CrossRef] [Академия Google]
122. Yamamoto T., Tamura Y., Yokota T. Плазмида устойчивости к антисептикам и антибиотикам в штамме Staphylococcus aureus , обладающая способностью придавать устойчивость к хлоргексидину и акринолу. Антимикроб. Агенты Чемотер. 1988; 32: 932–935. doi: 10.1128/AAC.32.6.932. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
123. Червинкова Д., Бабак В., Марошевич Д., Кубикова И., Яглич З. Роль гена qacA в опосредовании устойчивости к четвертичные аммониевые соединения. микроб. Сопротивление наркотикам. 2012;19: 160–167. doi: 10.1089/mdr.2012.0154. [PubMed] [CrossRef] [Google Scholar]
124. Берг Дж., Том-Петерсен А., Найбро О. Медная добавка в сельскохозяйственную почву выбирает бактериальную устойчивость к антибиотикам в полевых условиях. лат. заявл. микробиол. 2005; 40: 146–151. doi: 10.1111/j.1472-765X.2004.01650.x. [PubMed] [CrossRef] [Google Scholar]
125. Cavaco L.M., Hasman H., Stegger M., Andersen P.S., Skov R., Fluit A.C., Ito T., Aarestrup F.M. Клонирование и обнаружение czrC , гена, придающего устойчивость к кадмию и цинку у устойчивых к метициллину Staphylococcus aureus изолятов CC398. Антимикроб. Агенты Чемотер. 2010;54:3605–3608. doi: 10.1128/AAC.00058-10. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
126. Vahjen W., Pietruszyńska D., Starke I.C., Zentek J. Высокое содержание цинка в пище увеличивает появление генов устойчивости к тетрациклину и сульфонамидам в кишечнике отъемных свиней. Гут Патог. 2015; 7 doi: 10.1186/s13099-015-0071-3. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
127. Солиоз М., Стоянов Ю.В. Гомеостаз меди в Enterococcus hirae . ФЭМС микробиол. 2003; 27:183–195. doi: 10.1016/S0168-6445(03)00053-6. [PubMed] [CrossRef] [Google Scholar]
128. Чардак М., Алтуг Г. Распространение видов и устойчивость к тяжелым металлам видов Enterobacteriaceae , выделенных из Стамбульского пролива. Фрезениус Энвайрон. Бык. 2014;23:2620–2626. [Google Scholar]
129. Хасман Х., Аареструп Ф.М. tcrB ген, придающий переносимую устойчивость к меди в Enterococcus faecium : встречаемость, переносимость и связь с устойчивостью к макролидам и гликопептидам. Антимикроб. Агенты Чемотер. 2002;46:1410–1416. doi: 10.1128/AAC.46.5.1410-1416.2002. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
130. Klare I., Heier H., Claus H., Reissbrodt R., Witte W. vanA -опосредованная резистентность к гликопептидам высокого уровня в Enterococcus faecium из животноводства. ФЭМС микробиол. лат. 1995; 125:165–172. дои: 10.1111/j.1574-6968.1995.tb07353.x. [PubMed] [CrossRef] [Google Scholar]
131. Becerril R., Nerin C., Gomez-Lus R. Оценка устойчивости бактерий к эфирным маслам и антибиотикам после воздействия эфирных масел орегано и корицы. Патог пищевого происхождения. Дис. 2012; 9: 699–705. doi: 10.1089/fpd.2011.1097. [PubMed] [CrossRef] [Google Scholar]
132. Fadli M., Chevalier J., Hassani L., Mezrioui N.E., Pages J.M. Натуральные экстракты стимулируют связанные с мембраной механизмы резистентности грамотрицательных бактерий. лат. заявл. микробиол. 2014; 58: 472–477. doi: 10.1111/lam.12216. [PubMed] [CrossRef] [Академия Google]
133. МакМахон М.А.С., Блэр И.С., Мур Дж.Е., Макдауэлл Д.А. Привыкание к сублетальным концентрациям масла чайного дерева ( Melaleuca alternifolia ) связано со снижением восприимчивости патогенов человека к антибиотикам. Дж. Антимикроб. Чемотер. 2007; 59: 125–127. doi: 10.1093/jac/dkl443. [PubMed] [CrossRef] [Google Scholar]
134. Oosterik L.H., Peeters L., Mutuku I., Goddeeris B.M., Butaye P. Восприимчивость птичьего патогена Escherichia coli от кур-несушек в Бельгии к антибиотикам, дезинфицирующим средствам и интегрону распространенность. Авиан Дис. 2014; 58: 271–278. doi: 10.1637/10680-100113-RegR. [PubMed] [CrossRef] [Академия Google]
135. Zou L., Meng J., McDermott P. F., Wang F., Yang Q., Cao G., Hoffmann M., Zhao S. Наличие генов устойчивости к дезинфицирующим средствам у Escherichia coli , выделенных из розничного мяса в США. Дж. Антимикроб. Чемотер. 2014;69:2644–2649. doi: 10.1093/jac/dku197. [PubMed] [CrossRef] [Google Scholar]
136. Randall L.P., Clouting C.S., Gradel K.O., Clifton-Hadley F.A., Davies R.D., Woodward M.J. Фермерские дезинфицирующие средства выбирают по устойчивости к циклогексану, маркеру множественной устойчивости к антибиотикам, в Кишечная палочка . Дж. Заявл. микробиол. 2005; 98: 556–563. doi: 10.1111/j.1365-2672.2004.02488.x. [PubMed] [CrossRef] [Google Scholar]
137. Braoudaki M., Hilton A.C. Низкий уровень перекрестной устойчивости между триклозаном и антибиотиками у Escherichia coli K-12 и E. coli O55 по сравнению с E. палочка О157. ФЭМС микробиол. лат. 2004; 235:305–309. doi: 10.1111/j.1574-6968.2004.tb09603.x. [PubMed] [CrossRef] [Google Scholar]
138. Gradel K.O., Randall L., Sayers A.R., Davies R.H. Возможные связи между Salmonella устойчивость в птичниках и устойчивость к обычно используемым дезинфицирующим средствам и предполагаемая роль mar. Вет. микробиол. 2005; 107: 127–138. doi: 10.1016/j.vetmic.2005.01.013. [PubMed] [CrossRef] [Google Scholar]
139. Gantzhorn M.R., Pedersen K., Olsen J.E., Thomsen L.E. Чувствительность к биоцидам и антибиотикам изолятов Salmonella , полученных до и после очистки на шести скотобойнях в Дании. Междунар. Дж. Пищевая микробиология. 2014; 181:53–59. doi: 10.1016/j.ijfoodmicro.2014.04.021. [PubMed] [CrossRef] [Академия Google]
140. Chuanchuen R., Pathanasophon P., Khemtong S., Wannaprasat W., Padungtod P. Восприимчивость к противомикробным и дезинфицирующим средствам в изолятах Salmonella , полученных от домашней птицы и свиней в Таиланде. Дж. Вет. Мед. науч. 2008; 70: 595–601. doi: 10.1292/jvms.70.595. [PubMed] [CrossRef] [Google Scholar]
141. Копич Дж.Л., Уайтхед Р.Н., Уэббер М.А. Распространенность сниженной чувствительности к триклозану в изолятах Salmonella enterica , полученных от животных и человека, и связь с множественной лекарственной устойчивостью. Междунар. Дж. Антимикроб. Агенты. 2010; 36: 247–251. doi: 10.1016/j.ijantimicag.2010.04.012. [PubMed] [CrossRef] [Академия Google]
142. Randall L.P., Cooles S.W., Sayers A.R., Woodward M.J. Связь между устойчивостью к циклогексану Salmonella различных сероваров и повышенной устойчивостью ко многим антибиотикам, дезинфицирующим средствам и красителям. Дж. Мед. микробиол. 2001; 50: 919–924. doi: 10.1099/0022-1317-50-10-919. [PubMed] [CrossRef] [Google Scholar]
143. Guo W., Cui S., Xu X., Wang H. Изучение механизма резистентности хлорида бензалкония, отобранных мутантов Salmonella Typhimurium. микроб. Сопротивление наркотикам. 2014;20:11–16. дои: 10.1089/мдр.2012.0225. [PubMed] [CrossRef] [Google Scholar]
144. Randall L. P., Cooles S.W., Coldham N.G., Penuela E.G., Mott A.C., Woodward M.J., Piddock L.J.V., Webber M.A. Обычно используемые на фермах дезинфицирующие средства можно выбрать для мутантных Salmonella enterica serovar Typhimurium со сниженной чувствительностью к биоцидам и антибиотикам без снижения вирулентности. Дж. Антимикроб. Чемотер. 2007; 60: 1273–1280. doi: 10.1093/jac/dkm359. [PubMed] [CrossRef] [Google Scholar]
145. Randall LP, Bagnall M.C., Karatzas K.A., Coldham NC, Piddock L.J.V., Woodward M.J. Пригодность и распространение выбранных дезинфицирующим средством мультирезистентных к антибиотикам (MAR) штаммов Salmonella enterica серовар Typhimurium у кур. Дж. Антимикроб. Чемотер. 2008; 61: 156–162. doi: 10.1093/jac/dkm415. [PubMed] [CrossRef] [Google Scholar]
146. Бирошова Л., Микулашова М. Развитие устойчивости к триклозану и антибиотикам у Salmonella enterica серовара Typhimurium. Дж. Мед. микробиол. 2009; 58: 436–441. doi: 10.1099/jmm. 0.003657-0. [PubMed] [CrossRef] [Google Scholar]
147. Георнарас И., Ян Х., Мошонас Г., Наннелли М.С., Белк К.Е., Найтингейл К.К., Вернер Д.Р., Смит Г.К., Софос Дж.Н. Эффективность химических вмешательств против Escherichia coli O157:H7 и полирезистентные и чувствительные к антибиотикам Salmonella на инокулированной говяжьей обрезке. Дж. Пищевая защита. 2012; 75:1960–1967. doi: 10.4315/0362-028X.JFP-12-195. [PubMed] [CrossRef] [Google Scholar]
148. Capita R. Изменчивость устойчивости Salmonella к химическим деконтаминантам птицы, основанная на серотипе, типе фага и характере устойчивости к антибиотикам. Дж. Пищевая защита. 2007; 70: 1835–1843. [PubMed] [Академия Google]
149. Молина-Гонсалес Д., Алонсо-Каллеха К., Алонсо-Эрнандо А., Капита Р. Влияние сублетальных концентраций биоцидов на чувствительность к антибиотикам штаммов Salmonella enterica с множественной лекарственной устойчивостью. Пищевой контроль. 2014;40:329–334. doi: 10.1016/j.foodcont.2013.11.046. [CrossRef] [Google Scholar]
150. Mourão J., Novais C., Machado J., Peixe L., Antunes P. Толерантность к металлам при появлении клинически значимой множественной лекарственной устойчивости Salmonella enterica серотипа 4, [5], 12:i: — Клоны, циркулирующие в Европе. Междунар. Дж. Антимикроб. Агенты. 2015;45:610–616. doi: 10.1016/j.ijantimicag.2015.01.013. [PubMed] [CrossRef] [Академия Google]
151. Хасман Х., Мудли А., Гуардабасси Л., Стеггер М., Сков Р.Л., Аареструп Ф.М. Spa тип распространения в Staphylococcus aureus , происходящий от свиней, крупного рогатого скота и домашней птицы. Вет. микробиол. 2010; 141:326–331. doi: 10.1016/j.vetmic.2009.09.025. [PubMed] [CrossRef] [Google Scholar]
152. Aarestrup F.M., Cavaco L., Hasman H. Снижение восприимчивости к хлориду цинка связано с устойчивым к метициллину Staphylococcus aureus CC398 у датских свиней. Вет. микробиол. 2010; 142:455–457. doi: 10.1016/j.vetmic.2009.10.021. [PubMed] [CrossRef] [Google Scholar]
153. Moodley A., Nielsen S.S., Guardabassi L. Ответ автора: Критический анализ статьи «Влияние тетрациклина и цинка на селекцию метициллин-резистентного Staphylococcus aureus (MRSA) последовательность типа 398 у свиней» Vet. микробиол. 2014; 173:401–402. doi: 10.1016/j.vetmic.2014.09.023. [PubMed] [CrossRef] [Google Scholar]
154. Moodley A., Nielsen S.S., Guardabassi L. Влияние тетрациклина и цинка на селекцию резистентных к метициллину Staphylococcus aureus (MRSA) тип последовательности 398 у свиней. Вет. микробиол. 2011; 152:420–423. doi: 10.1016/j.vetmic.2011.05.025. [PubMed] [CrossRef] [Google Scholar]
155. Slifierz M.J., Friendship R., Weese J.S. Терапия оксидом цинка увеличивает распространенность и устойчивость метициллин-резистентного Staphylococcus aureus у свиней: рандомизированное контролируемое исследование. Зоонозы Общественное здравоохранение. 2015;62:301–308. doi: 10.1111/zph.12150. [PubMed] [CrossRef] [Google Scholar]
156. Slifierz M.J., Friendship RM, Weese J.S. Метициллин-резистентный Staphylococcus aureus в коммерческих стадах свиней связан с использованием дезинфицирующих средств и цинка. заявл. Окружающая среда. микробиол. 2015;81:2690–2695. doi: 10.1128/AEM.00036-15. [Статья PMC бесплатно] [PubMed] [CrossRef] [Google Scholar]
157. Миллар М.Р., Гриффин Н., Кейворт Н. Характер устойчивости к антибиотикам и ионам тяжелых металлов у недавних госпитальных изолятов Staphylococcus aureus . Эпидемиол. Заразить. 1987; 99: 343–347. doi: 10.1017/S0950268800067819. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
158. Постон С.М., Ли Со Хи Ф.Л. Генетическая характеристика устойчивости к ионам металлов у метициллин-резистентного штамма Staphylococcus aureus : устранение устойчивости к кадмию, ртути и тетрациклину с потерей устойчивости к метициллину. Дж. Мед. микробиол. 1991; 34: 193–201. doi: 10.1099/00222615-34-4-193. [PubMed] [CrossRef] [Google Scholar]
159. Townsend D.E., Grubb W.B., Ashdown N. Генетика лекарственной устойчивости метициллин-резистентного Staphylococcus aureus из больниц Австралии. Дж. Хосп. Заразить. 1983; 4: 331–337. doi: 10.1016/0195-6701(83)
-8. [PubMed] [CrossRef] [Google Scholar]160. Gómez-Sanz E., Kadlec K., Feßler A.T., Zarazaga M., Torres C., Schwarz S. Novel erm (T), несущие мультирезистентные плазмиды из свиные и человеческие изоляты устойчивого к метициллину Staphylococcus aureus ST398, которые также содержат детерминанты устойчивости к кадмию и меди. Антимикроб. Агенты Чемотер. 2013;57:3275–3282. doi: 10.1128/AAC.00171-13. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
161. Bae D., Mezal E.H., Smiley R.D., Cheng C.M., Khan A.A. Подвидовая характеристика и устойчивость к противомикробным препаратам Listeria monocytogenes , выделенных из отечественных и импортных пищевых продуктов с 2004 по 2011 гг. Food Res. Междунар. 2014; 64: 656–663. doi: 10.1016/j.foodres.2014.07.049. [PubMed] [CrossRef] [Google Scholar]
162. Ortiz S., Lopez P., Lopez V., Martinez-Suarez J.V. Чувствительность к антибиотикам у устойчивых и чувствительных к бензалкония хлориду Listeria monocytogenes штаммов. Патог пищевого происхождения. Дис. 2014; 11: 517–519. doi: 10.1089/fpd.2013.1724. [PubMed] [CrossRef] [Google Scholar]
163. Aarestrup F.M., Knöchel S., Hasman H. Чувствительность к противомикробным препаратам Listeria monocytogenes из пищевых продуктов. Патог пищевого происхождения. Дис. 2007; 4: 216–221. doi: 10.1089/fpd.2006.0078. [PubMed] [CrossRef] [Google Scholar]
164. Rakic-Martinez M., Drevets D.A., Dutta V., Katic V., Kathariou S. Штаммы Listeria monocytogenes , отобранные на ципрофлоксацин или дезинфицирующее средство хлорид бензалкония, проявляют пониженную чувствительность к ципрофлоксацину, гентамицину, бензалкония хлориду и другим токсичным соединениям. заявл. Окружающая среда. микробиол. 2011;77:8714–8721. дои: 10.1128/AEM.05941-11. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
165. Романова Н. А., Вольфс П. Ф. Г., Бровко Л. Ю., Гриффитс М. В. Роль эффлюксных насосов в адаптации и устойчивости Listeria monocytogenes к хлориду бензалкония. заявл. Окружающая среда. микробиол. 2006;72:3498–3503. doi: 10.1128/AEM.72.5.3498-3503.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
166. Christensen E.G., Gram L., Kastbjerg V.G. Сублетальное воздействие триклозана снижает чувствительность к гентамицину и другим аминогликозидам в 9 случаях.1465 Listeria monocytogenes . Антимикроб. Агенты Чемотер. 2011;55:4064–4071. doi: 10.1128/AAC.00460-11. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
167. Лопес-Алонсо В., Ортис С., Мартинес-Суарес Дж. В. Геномные последовательности пяти устойчивых к дезинфицирующим средствам штаммов Listeria monocytogenes из двух иберийских свиней- перерабатывающие заводы. Объявление генома. 2015; 3 doi: 10.1128/genomeA.00077-15. e00077-15. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
168. Маври А., Куринчич М., Можина С.С. Распространенность устойчивости к антибиотикам и биоцидам среди Campylobacter coli и Campylobacter jejuni из разных источников. Пищевая Технол. Биотехнолог. 2012;50:371–376. [Google Scholar]
169. Randall L.P., Ridley A.M., Cooles S.W., Sharma M., Sayers A.R., Pumbwe L., Newell D.G., Piddock L.J.V., Woodward M.J. Распространенность множественной устойчивости к антибиотикам у 443 Campylobacter spp. изолированы от человека и животных. Дж. Антимикроб. Чемотер. 2003; 52: 507–510. doi: 10.1093/jac/dkg379. [PubMed] [CrossRef] [Академия Google]
170. Sundsfjord A., Willems R. Исследование Enterococcus : Последние разработки и клинические проблемы. клин. микробиол. Заразить. 2010; 16: 525–526. doi: 10.1111/j.1469-0691.2010.03215.x. [PubMed] [CrossRef] [Google Scholar]
171. Schwaiger K., Harms K.S., Bischoff M., Preikschat P., Mölle G., Bauer-Unkauf I., Lindorfer S., Thalhammer S., Bauer J. , Hölzel C.S. Невосприимчивость к дезинфицирующим средствам бактерий животных, продуктов питания и человека — есть ли связь с устойчивостью к противомикробным препаратам? Фронт. микробиол. 2014;5 doi: 10.3389/fmicb.2014.00088. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
172. Паскуароли С., ди Чезаре А., Вигнароли К., Конти Г., Читтерио Б., Биаваско Ф. Устойчивость к эритромицину и меди Enterococcus hirae из морских отложений и совместный перенос erm (B) и tcr B в человека Enterococcus faecalis . Диагн. микробиол. Заразить. Дис. 2014;80:26–28. doi: 10.1016/j.diagmicrobio.2014.06.002. [PubMed] [CrossRef] [Академия Google]
173. Аареструп Ф.М., Хасман Х., Дженсен Л.Б., Морено М., Эрреро И.А., Домингес Л., Финн М., Франклин А. Устойчивость к противомикробным препаратам среди энтерококков свиней в трех европейских странах. заявл. Окружающая среда. микробиол. 2002;68:4127–4129. doi: 10.1128/AEM.68.8.4127-4129.2002. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
174. Zhang S., Wang D., Wang Y., Hasman H., Aarestrup F.M., Alwathnani H.A., Zhu Y.-G., Rensing C. Геномные последовательности устойчивых и чувствительных к меди Enterococcus faecalis штаммов, выделенных от свиней, откормленных медью, в Дании. Стоять. Геномная наука. 2015; 10 doi: 10.1186/s40793-015-0021-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
175. Fard R.M.N., Heuzenroeder M.W., Barton MD. Устойчивость к противомикробным препаратам и тяжелым металлам у комменсальных энтерококков, выделенных от свиней. Вет. микробиол. 2011; 148: 276–282. doi: 10.1016/j.vetmic.2010.09.002. [PubMed] [CrossRef] [Google Scholar]
176. Amachawadi R.G., Scott H.M., Aperce C., Vinasco J., Drouillard J.S., Nagaraja T.G. Влияние добавок меди и тилозина в корма на медь и устойчивость к противомикробным препаратам фекальных энтерококков крупного рогатого скота на откормочных площадках. Дж. Заявл. микробиол. 2015;118:1287–1297. doi: 10.1111/jam.12790. [PubMed] [CrossRef] [Google Scholar]
177. Бисвал Б.К., Хайраллах Р., Биби К., Мазза А., Гер Р., Массон Л., Фригон Д. Влияние дезинфекции УФ и надуксусной кислотой на распространенность генов вирулентности и устойчивости к противомикробным препаратам у уропатогенных штаммов Escherichia coli в сточных водах. заявл. Окружающая среда. микробиол. 2014; 80:3656–3666. doi: 10.1128/AEM.00418-14. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
178. Tuckfield RC, McArthur JV Пространственный анализ устойчивости к антибиотикам вдоль ручьев, загрязненных металлами. микроб. Экол. 2007;55:595–607. doi: 10.1007/s00248-007-9303-5. [PubMed] [CrossRef] [Google Scholar]
179. Hernández A., Ruiz F.M., Romero A., Martinez J.L. Связывание триклозана с SmeT, репрессором мультилекарственного эффлюксного насоса SmeDEF, индуцирует устойчивость к антибиотикам у Stenotrophomonas maltophilia . ПЛОС Патог. 2011;7:e1002103. doi: 10.1371/journal.ppat.1002103. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
180. Suling W.J., O’Leary W.M. Влияние поверхностно-активных веществ на устойчивость к антибиотикам. Антимикроб. Агенты Чемотер. 1975;8:334–343. doi: 10.1128/AAC.8.3.334. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
181. Занини С.Ф., Сильва-Ангуло А.Б., Розенталь А., Алиага Д.Р., Мартинес А. Влияние лечения Listeria monocytogenes и Salmonella enterica серовара Typhimurium с цитралем на эффективность различных антибиотиков. Патог пищевого происхождения. Дис. 2014; 11: 265–271. doi: 10.1089/fpd.2013.1635. [PubMed] [CrossRef] [Google Scholar]
Селекция и распространение устойчивости к противомикробным препаратам в агропищевом производстве | Устойчивость к противомикробным препаратам и инфекционный контроль
- Обзор
- Открытый доступ
- Опубликовано:
- Guyue Cheng 1 na1 ,
- Jianan Ning 1 na1 ,
- Saeed Ahmed 1 ,
- Junhong Huang 1 ,
- Rizwan Ullah 3 ,
- Boyu Ан 1 ,
- Haihong Hao 1 ,
- Menghong Dai 1 ,
- Lingli Huang 2 ,
- Xu Wang 1 и
- …
- ZonGhi I.
Устойчивость к противомикробным препаратам и инфекционный контроль
том 8 , Номер статьи: 158 (2019)
Процитировать эту статью
-
7187 Доступ
-
53 Цитаты
-
7 Альтметрический
-
Сведения о показателях
Abstract
Общественные волнения по поводу использования антимикробных агентов в сельскохозяйственной практике являются основной причиной роста и появления бактерий с множественной лекарственной устойчивостью, которые вынуждают агропродовольственную промышленность действовать.
Использование противомикробных препаратов в продовольствии и сельском хозяйстве оказывает прямое или косвенное влияние на развитие устойчивости к противомикробным препаратам (УПП) у бактерий, связанных с животными и растениями, которые могут попасть в пищевую цепь при употреблении мяса, рыбы, овощей или некоторых других источников пищи. В дополнение к противомикробным препаратам недавние отчеты показали, что УПП связана с толерантностью к тяжелым металлам, существующим в природе или используемым в агропродовольственном производстве. Кроме того, биоциды, включая дезинфицирующие средства, антисептики и консерванты, которые широко используются на фермах и бойнях, также могут способствовать развитию УПП. Хотя прямая передача УПП от пищевых животных и связанной с ними окружающей среды человеку все еще остается неясной и спорной, этим риском не следует пренебрегать. Поэтому необходимы совместные глобальные усилия для надлежащего использования противомикробных препаратов, тяжелых металлов и биоцидов в агропродовольственном производстве для контроля за развитием УПП. Эти коллективные меры сохранят эффективность существующих противомикробных препаратов для будущих поколений. Введение
Противомикробные препараты, включая антибиотики и родственные им полусинтетические или синтетические препараты, обладают высокой противомикробной активностью и селективной токсичностью, что позволяет использовать их в качестве противоинфекционных средств [1]. На протяжении многих лет противомикробные препараты также использовались в животноводстве и аквакультуре для стимуляции роста, повышения эффективности кормления, профилактики и лечения инфекционных заболеваний. С точки зрения благополучия животных использование противомикробных препаратов улучшает общее состояние здоровья сельскохозяйственных животных и гигиену сельскохозяйственной среды [1]. Сельскохозяйственная пищевая промышленность выигрывает от использования противомикробных препаратов для производства продуктов питания и защиты растений. В США около 80% производимых антибиотиков используется в животноводстве [2].
Было подсчитано, что в глобальном масштабе каждый килограмм мяса крупного рогатого скота, кур и свиней приведет к потреблению 45 мг, 148 мг и 172 мг противомикробных препаратов соответственно, что, как ожидается, увеличится на 67% с 2010 по 2030 год [3]. ]. Устойчивость к противомикробным препаратам является общепризнанной проблемой общественного здравоохранения, поскольку ее появление ограничивает терапевтические возможности, доступные как клиницистам, так и ветеринарам. В первом экономическом отчете о влиянии УПП предполагалось, что, если ничего не делать, смертность, связанная с УПП, увеличится с 700 000 до 10 миллионов в год к 2050 году. Это будет стоить отрасли здравоохранения триллионы долларов США [4]. Неправильное использование противомикробных препаратов для целей, отличных от лечения инфекций, привело к отбору против УПП в условиях производства пищевых продуктов. Бактерии развивают резистентность de novo из-за воздействия субингибирующих уровней антибиотиков в их окружении или напрямую приобретают механизмы устойчивости от других бактерий посредством горизонтального переноса генов (HGT) [5].
Хотя широкое распространение УПП в основном связывают с избирательным давлением, вызванным чрезмерным и неправильным использованием противомикробных препаратов, в связи с недавним растущим количеством данных о совместной селекции УПП среди бактерий, подвергшихся воздействию неантибиотических соединений, используемых в агропищевой промышленности, возникли опасения. , такие как биоциды, используемые в качестве дезинфицирующих средств, антисептиков и консервантов, тяжелые металлы, существующие в природе и используемые в сельскохозяйственном производстве [6]. Использование противомикробных препаратов, тяжелых металлов и биоцидов в продовольствии и сельском хозяйстве оказывает прямое и косвенное воздействие на развитие УПП у бактерий, которые могут попасть в пищевую цепь. Растущие волнения среди общественности по поводу использования противомикробных препаратов в сельском хозяйстве и появление бактерий с множественной лекарственной устойчивостью заставили сельскохозяйственную пищевую промышленность действовать.
Основной областью, находящейся под пристальным вниманием, является пищевая цепочка животноводства, от ферм через скотобойни и заводы по переработке пищевых продуктов до упаковочных и розничных предприятий [7]. В этом обзоре будут обобщены основные факторы отбора и распространения УПП пищевого происхождения по пищевой цепочке. Селекция УПП с использованием противомикробных препаратов
Механизмы УПП и предсуществование генов устойчивости к противомикробным препаратам (ARG)
Устойчивость к противомикробным препаратам включает два уровня устойчивости: устойчивость на клеточном уровне и блокирование мишени противомикробных препаратов и снижение проникновения противомикробных препаратов в или активный отток антимикробных препаратов из бактериальной клетки [2]. Снижение чувствительности организма к антимикробному препарату может быть врожденным (вследствие особенностей клеточной оболочки микроба, энергетического обмена или наличия альтернативного пути метаболизма).
Таблица 1 Мобильные гены устойчивости к противомикробным препаратам Он также приобретается посредством одноступенчатой или многостадийной мутации, которая влияет на целевой сайт и эффективную концентрацию противомикробного препарата в клетке, или путем приобретения генетического элемента, кодирующего такую функцию, как инактивирующий фермент или альтернативу целевой молекуле, т.е. HGT детерминант резистентности. В таблице 1 показаны репрезентативные мобильные ARG, которые можно передавать между различными бактериальными штаммами и видами. Резистентность на уровне сообщества (биопленки и персистеры) также является проблемой, вызывающей трудности в антимикробной терапии [26].
Полноразмерная таблица
Однако устойчивость к противомикробным препаратам возникла не как продукт использования противомикробных препаратов в сельском хозяйстве. Устойчивость к антибиотикам — это древний бактериальный признак, существовавший у почвенных бактерий (почвенный резистом) и переносимый плазмидами, такими как сериновые β-лактамазы, за миллионы лет до зарождения сельского хозяйства [27].
Недавняя работа обнаружила сопротивление в древней вечной мерзлоте, изолированных пещерах и в человеческих образцах, сохранившихся в течение сотен лет [28]. Было показано, что ген, кодирующий устойчивость к β-лактамным, тетрациклиновым и гликопептидным антибиотикам, присутствует в образцах метагенома вечной мерзлоты возрастом 30 000 лет [29].]. Недавно было обнаружено, что кишечный микробиом доколумбовой мумии из Анд (датируемой 980–1170 гг. н.э.) содержит гены устойчивости к β-лактаму, фосфомицину, хлорамфениколу, аминогликозидам, макролидам, сульфамидам, хинолонам, тетрациклину и ванкомицину [30]. При скрининге образца пригодного для культивирования микробиома пещеры Лечугилла, изолированного более 4 миллионов лет, поверхностные микробы были очень устойчивы к противомикробным препаратам, а некоторые штаммы были устойчивы к 14 различным коммерчески доступным противомикробным препаратам, включая даптомицин и макролид [31]. Результаты этих исследований дали прямые экспериментальные доказательства того, что УПП является древним, и позволили заглянуть в историю эволюции природного явления окружающей среды. Отбор AMR в окне селекции мутантов и субингибирующих концентрациях
Концентрация противомикробного препарата либо в окне селекции мутантов (MSW), либо ниже минимальной ингибирующей концентрации (МИК) популяции дикого типа (также называемой субингибирующей концентрацией). -ингибирующая концентрация или суб-МИК концентрация) важна для выбора УПП [32]. MSW представляет собой диапазон концентраций между самой низкой концентрацией, которая оказывает избирательное давление, часто приближается к минимальной концентрации, которая ингибирует образование колоний на 99% (МИК99) и МИК субпопуляции мутантов с наименьшей лекарственной устойчивостью, значение, называемое концентрацией предотвращения мутаций (МПК) [33]. Лекарственно-устойчивые мутантные субпопуляции, присутствующие до начала антимикробного лечения, обогащаются и амплифицируются, когда концентрации противомикробных препаратов находятся в пределах MSW.
Противомикробные препараты в субингибирующих концентрациях (концентрации ниже MIC) встречаются во многих природных средах, таких как почва и вода.
Субингибирующие концентрации также образуются в результате антимикробной терапии у людей и домашнего скота (субоптимальная дозировка терапии, плохая фармакокинетика, использование некачественных препаратов и плохое соблюдение пациентом режима лечения), а также при введении в качестве кормовых добавок для стимуляции роста животных. [5]. В суб-МИК концентрациях чувствительные штаммы продолжают расти с пониженной скоростью роста, и самая низкая концентрация противомикробного препарата, необходимая для выбора устойчивого мутанта по сравнению с диким типом, называется минимальной селективной концентрацией (МСК), от которой до МИК происходит отбор для встречаются устойчивые мутанты [34]. Помимо ранее существовавших устойчивых мутантов, бактериальная устойчивость de novo может быть повышена за счет субтерапевтических антимикробных концентраций путем индукции неспецифического мутагенеза в результате стимуляции производства активных форм кислорода [35]. Селекция ARG в системе производства продуктов питания
Кормление противомикробными препаратами пищевых животных было селективной силой в эволюции их кишечных бактерий, в частности, за счет увеличения распространенности и разнообразия ARG [27].
Однако связь между применением противомикробных препаратов и выбором детерминант резистентности не так пряма, как это часто предполагается. Недавнее исследование, проведенное на свинофермах в Дании, показало, что воздействие противомикробных препаратов на уровни семи АРГ ( ermB , ermF , sulI , sulII , tetM , tetO и tetW ) был сложным и уникальным для каждого отдельного гена. Несколько классов противомикробных препаратов имели как отрицательную, так и положительную корреляцию с ARG, что указывает на то, что воздействие противомикробных препаратов не является единственным важным фактором, определяющим уровни ARG [36]. В американской свиноводческой системе цефтиофур часто вводят поросятам при рождении, а самцам вводят вторую дозу при кастрации, и эта операция может обеспечить селекционное давление, необходимое для распространения продуцирующих карбапенемазы 9.0087 Enterobacteriaceae [37]. В изолятах Campylobacter jejuni от крупного рогатого скота в замкнутых откормочных хозяйствах в Южной Альберте, Канада, селекция на устойчивость к фторхинолонам зависела от подтипа, тогда как селекция на устойчивость к тетрациклинам не зависела [38]. Было показано, что развитие резистентности к ципрофлоксацину сильно различается у разных штаммов сероваров из-за разной частоты мутаций и уровня накопления ципрофлоксацина [39]. Совместная селекция УПП с использованием непротивомикробных соединений
Широко распространенная УПП в основном связана с селективным давлением, вызванным чрезмерным и неправильным использованием противомикробных препаратов. Тем не менее, были высказаны опасения на основании растущих данных о совместной селекции УПП среди бактерий, подвергшихся воздействию биоцидов, которые используются в качестве дезинфицирующих средств, антисептиков, консервантов, и различных катионных тяжелых металлов, включенных в рационы животных в качестве пищевых добавок, стимуляторов роста и терапевтических средств для домашнего скота. [6]. Эти металлы также можно разбрасывать на пастбищах для поддержки роста и защиты сельскохозяйственных культур.
Совместная селекция УПП с тяжелыми металлами
Тяжелые металлы повсеместно встречаются в окружающей среде и иногда в высоких концентрациях в определенных условиях, когда они используются в сельскохозяйственном производстве для различных целей.
Тяжелые металлы могут продолжать существовать в окружающей среде и оставаться стабильными в течение длительного времени. В то время как большинство ветеринарных противомикробных соединений могут метаболизироваться и выводиться из организма животных, производящих пищу, в течение недель или месяцев. Биодоступность обычно используемых в кормах минералов (в основном неорганических) обычно довольно низка у животных, а неабсорбированные тяжелые металлы выделяются с фекалиями в более высоких концентрациях, чем в кормах [40]. Корреляция между толерантностью к тяжелым металлам и УПП уже наблюдалась несколько десятилетий назад. Сообщается, что медь (Cu) связана с устойчивостью к ампициллину, сульфаниламиду [41], эритромицину [42], энрофлоксацину [43], ванкомицину [44] и гликопептиду [45]. Метициллин-резистентный Staphylococcus aureus (MRSA) часто ассоциируется с цинком (Zn) [45,46,47,48] и Cu [45]. Имеются положительные корреляции между геном толерантности к ртути (Hg) merA и транспозоном Tn21 [42].
sulA и sulA сильно коррелировали с уровнями Cu, Zn и Hg [49]. Обнаружено, что Enterobacter hormaechei и E. asburiae , продуцирующие CTX-M-(15, 9, 2) и KPC-2, обладают набором приобретенных генов устойчивости к серебру (Ag) [50]. Сообщается также, что другие тяжелые металлы, включая никель (Ni), кадмий (Cd) и хром (Cr), также участвуют в выборе определенных AMR [42, 51, 52, 53]. Недавнее исследование показало, что гены, потенциально отвечающие за устойчивость к металлам, в том числе arsA (соединения мышьяка), cadD (Cd), copB (Cu) и czrC (Zn/Cd) часто присутствовали в MRSA, ассоциированном с домашним скотом [54]. Китайское исследование даже обнаружило лишь слабую положительную корреляцию между ARG и соответствующими противомикробными препаратами, тогда как значительная положительная корреляция была обнаружена между некоторыми ARG ( sulA и sulIII ) и типичными тяжелыми металлами, такими как Hg, Cu и Zn [49] . Молекулярные механизмы способности бактерий вырабатывать устойчивость к тяжелым металлам аналогичны механизмам для УПП, поскольку тяжелые металлы обладают антимикробным действием [55]. Коселекция достигается двумя способами: (1) корезистентность, при которой отбор по одному гену способствует поддержанию другого гена устойчивости, и (2) перекрестная устойчивость, при которой один ген устойчивости может обеспечить защиту от нескольких токсичных химических веществ [56]. . Корезистентность/коперенос тяжелого металла и противомикробного препарата часто вызывается корезидентными генами устойчивости к металлу и противомикробному препарату, которые могут быть физически локализованы в плазмидах или хромосомах, которые также содержат один или несколько ARG [57, 58]. . Например, было описано, что MRSA от домашнего скота содержат плазмиды, несущие гены устойчивости к Cu и Cd (9).0087 COPA , CADDX и MCO ) и для множества антимикробных препаратов, включая макролиды, ликосамиды, стрептограммин B, тетрациклины, аминогликозиды и триметоприм ( ERM (T) , (L) 7777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777777 гг.
) [59]. Связь между использованием Zn в кормах для животных и появлением MRSA объясняется физическим присутствием гена устойчивости к Zn, czrC , в элементе SCC mec , кодирующем устойчивость к метициллину [60, 61]. Другой пример корезистентности связан с рядом генов устойчивости, таких как 9[0087 aadA2 (стрептомицин R ), qacED1 (спектиномицин R ) и sul1 (сульфонамид R ), расположенные по отношению к Tn5045, где обнаружены 7 генов устойчивости к хроматам 902rBA 9008]. Португальское исследование показало, что в монофазных вариантах S. Typhimurium человеческого и свиного происхождения ARG в этом патоваре с множественной лекарственной устойчивостью были совмещены с опероном sil , который кодировал отток Cu и Ag на хромосоме или не -трансферабельная плазмида [63]. Анализ конъюгации продемонстрировал коперенос tcrB и erm(B) между штаммами E. faecium и E. faecalis [64]. Геномный анализ E. faecalis от датских свиней, получавших медь, выявил наличие хромосомных генов невосприимчивости к меди, включая оперон tcrYAZB и гены устойчивости к тетрациклину ( tetM ) и ванкомицину ( vanA ). «Cu-нечувствительных» изолятов [65]. Генетическая связь Cu, Zn и ARG у бактерий была подробно описана в недавнем обзоре, написанном Китом Пулом [57]. Подобно противомикробным препаратам, металлы являются стрессорами, которые активируют различные адаптивные/защитные реакции у бактерий, и это может сделать совместную регуляцию резистентности к металлам и противомикробным препаратам, приводя к перекрестной резистентности [66]. У грамотрицательных бактерий двухкомпонентная система CpxRA, реагирующая на мембранный стресс, которая связана с устойчивостью к различным препаратам, воздействующим на клеточную оболочку [67], также чувствительна к Cu и способствует толерантности к Cu [68]. В присутствии Zn TCS CscRS в Pseudomonas aeruginosa влияет на транскрипцию оперона czcCBA , кодирующего эффлюксный насос RND-типа, который придает устойчивость к Zn, Cd и кобальту (Co), в то время как система CscRS также снижает экспрессию порина OprD, через который имипенем проникает в бактерии [69].
В Listeria monocytogenes мультилекарственный эффлюксный насос MdrL обеспечивает устойчивость к ряду противомикробных препаратов, и та же транспортная система также работает для тяжелых металлов, таких как Zn, Co и Cr [70]. Точно так же сигма-фактор реакции на стресс оболочки RpoE, активируемый полимиксином B и связанный с устойчивостью к полимиксину B у ряда грамотрицательных бактерий [71], также активируется Zn в E. coli и способствует устойчивости к цинку и меди [72]. Также было показано, что Cu увеличивает экспрессию регуляторного гена soxS , чувствительного к окислительному стрессу, который связан с экспрессией эффлюксного насоса AcrAB и множественной лекарственной устойчивостью в E. coli [73]. Биопленки, в которых бактерии встроены во внеклеточные полимерные вещества, более устойчивы к тяжелым металлам, чем их планктонные аналоги [74]. В свою очередь, матрица биопленки может управлять частотой мутаций в бактериальных геномах, что благоприятно для коселекции на УПП [75].
Во многих сообщениях описано у нескольких грамотрицательных бактерий, что Cu индуцирует жизнеспособное, но некультивируемое (VNC) состояние, которое представляет собой индуцированное стрессом состояние покоя, устойчивое к противомикробным препаратам [76]. Zn-связанное состояние VNC также наблюдалось в Xylella Fastidiosa , и, по-видимому, ускоряет наступление состояния VNC в этом организме [77]. Кроме того, было показано, что воздействие Cu на E. coli увеличивает восстановление вариантов небольших колоний, а медленно растущие варианты обычно устойчивы к противомикробным препаратам для различных бактерий [78]. Тяжелые металлы также могут облегчать HGT. Недавнее исследование показало, что субингибирующие концентрации тяжелых металлов ускоряют горизонтальный перенос плазмид-опосредованных ARG в водной среде, способствуя конъюгативному переносу генов между штамм E. coli [79]. Другое исследование показало, что с помощью Cu шока при 10 и 100 мг / л нагрузка на бактерии из биофильтра питьевой воды значительно увеличивала устойчивость бактерий к рифампину, эритромицину, канамицину и некоторым другим.
Кроме того, относительная численность большинства ARG, особенно мобильных генетических элементов (MGE) intI и транспозонов, была заметно увеличена, по крайней мере, в 1 раз [80]. Совместная селекция УПП с биоцидами
Биоциды могут использоваться в качестве антисептиков на поверхностях тела, в качестве дезинфицирующих средств для оборудования и поверхностей во многих средах, включая фермы и больницы, в качестве обеззараживающих средств на поверхностях туш после убоя, а также в качестве консервантов в фармацевтических препаратах, косметике и продукты питания [81]. Возможная перекрестная устойчивость между биоцидами и противомикробными препаратами все еще остается спорной. В некоторых исследованиях сообщалось об отсутствии перекрестной резистентности между биоцидами и противомикробными препаратами. Например, перекрестная резистентность между хлоргексидином и пятью противомикробными препаратами не обнаружена у 130 9 .0087 Salmonella spp. с двух индюшиных ферм [82].
Среди 101 генетически различных изолятов Burkholderia cepacia не было обнаружено корреляции между чувствительностью к хлоргексидину и 10 различным противомикробным препаратам [83]. На Enterococcus faecium низкие дозы надуксусной кислоты, обычно используемой в качестве дезинфицирующего средства при очистке сточных вод, способствовали адаптации бактерий, но не влияли на обилие AGR [84]. С другой стороны, было проведено несколько исследований совместного отбора УПП биоцидами в бактериальных изолятах от пищевых животных и аквакультур. Было указано, что общее воздействие диглюконата хлоргексидина увеличивает риск устойчивости к различным противомикробным препаратам [85]. Когда 310 грамположительных изолятов из сосков дойных коров подверглись антисептической обработке йодом или хлоргексидином, была выявлена значительная связь между Streptococci между сниженной чувствительностью к хлоргексидину и ампициллину, тетрациклину и трем аминогликозидным антибиотикам [86].
В 87 изолятах из морепродуктов выявлены умеренные положительные корреляции биоцидов цетримида, гексадецилпиридиния хлорида и триклозана с антибиотиком цефотаксимом, а также триклозана с хлорамфениколом и триметопримом/сольфаметоксазолом и фенольным соединением тимолом [87]. Об этом сообщалось в E. coli O157 и различных Серовары Salmonella снижают чувствительность к группе противомикробных препаратов после поэтапного обучения триклозану, хлоргексидину и хлориду бензалкония [88]. Воздействие на ветеринарные полевые изоляты E. coli с тремя соединениями четвертичного аммония приводило к повышению МПК, превышающему клинические пограничные значения для феникола, тетрациклина, фторхинолона, β-лактамов и триметоприма [89]. Salmonella Enteritidis , выжившие после кратковременного воздействия концентраций хлора при использовании, демонстрировали восьмикратное увеличение значений МПК для тетрациклина, налидиксовой кислоты и хлорамфеникола [9].0], аналогично наблюдаемым при пошаговых процедурах обучения. Существует больше обзоров и исследований, в которых участвовали больницы или другие медицинские учреждения, посвященные совместному отбору УПП биоцидами [6]. Когда аэробные микробные сообщества подвергались воздействию бензалкония хлорида, значения МПК для всего сообщества для бензалкония хлорида, ципрофлоксацина, тетрациклина и пенициллина G увеличивались [91]. Недавние данные показали, что воздействие устойчивых к ванкомицину E. faecium к хлоргексидину всего за 15 минут активирует ген устойчивости к ванкомицину типа vanA ( vanHAX ) и гены, связанные со сниженной чувствительностью к даптомицину ( liaXYZ ) [92].
В некоторых исследованиях по обучению биоцидам была продемонстрирована роль оттока для одновременной селекции УПП [93], и наоборот, снижение восприимчивости к биоцидам может быть следствием развития УПП [94,95,96]. При воздействии бензалкония хлорида экспрессия двух генов неспецифических насосов оттока ( lde и mdrL ) в Listeria monocytogenes , выделенной из предприятий по переработке свинины [97].
Экспрессия lde была дозозависимой в случае штамма после процедуры очистки и дезинфекции, в то время как экспрессия mdrL ингибировалась при низком биоцидном стрессе (10 ppm) и усиливалась при высоком стрессе (100 ppm). ). При изучении потенциала образования биопленки и активности эффлюксной помпы изоляты E. coli из молочного оборудования со сниженной восприимчивостью к хлориду бензалкония и ципрофлоксацину показали превосходную биопленкообразующую способность параллельно с повышенной эффлюксной активностью [9].8]. Улучшенная биопленочная способность плюс отток также наблюдались в адаптированной к триклозану E. coli [99]. Генетические совпадения позволяют предположить, что плазмиды предоставляют биоцидам и металлам ограниченные возможности для обеспечения горизонтального переноса УПП посредством совместной селекции, в то время как существуют довольно большие возможности для непрямой селекции через хромосомные гены устойчивости к биоцидам/металлам [100]. Имеется множество теоретических и экспериментальных данных о том, что некоторые биоциды могут способствовать совместному отбору в отношении УПП, в основном за счет тесной связи детерминант устойчивости к биоцидам с детерминантами УПП. Тем не менее, отсутствуют эмпирические данные, указывающие на то, что использование биоцидов стимулирует этот совместный выбор УПП в пищевой цепи [101, 102].
Перенос и распространение УПП в пищевой цепи
Экологический резистом включает как естественный пул УПП, так и пул загрязненных УПП, возникших в результате деятельности человека [103]. Перенос ARG из природных резервуаров в другие бактерии может быть редким и случайным событием, загрязняющие ARB и ARG могут быстро и широко распространяться (например, металло-бета-лактамаза Нью-Дели, blaNDM-1 [104]; спектр бета-лактамаз blaCTXM-15 [105]; MRSA [106])
Передача УПП от пищевых животных в окружающую среду
Микробиомы сталкиваются с низкими дозами или субтерапевтическими уровнями антимикробных агентов всех механистических классов в производстве продуктов животного происхождения.
Эта современная практика оказывает широкое воздействие на микробиом кишечника пищевых животных, который впоследствии переносится в отходы животного происхождения. Внесение навоза в землю является обычной сельскохозяйственной практикой, которая может привести к расселению и размножению ARG в окружающей среде. Многие исследования доказали, что МГЭ и АРГ тесно связаны в своей стойкости в компостах при антимикробной селекции [107]. Различные источники навоза могут влиять на судьбу резистома в агроэкосистемах, как показано в недавнем исследовании, демонстрирующем, что применение свиного и птичьего помета может обогатить почву большим количеством ARG, чем навоз крупного рогатого скота, а относительное количество ARG имеет значительную положительную корреляцию с интегразой и транспозазой. генов [108]. Исследование сравнило 864 метагенома людей, животных и внешней среды и показало, что вода, отложения и почва, как правило, содержат малое относительное количество и несколько разновидностей известных ARG, кроме того, сточные воды/ил были на одном уровне с человеческим кишечником, что указывает на то, что среда с наибольшая относительная численность и/или разнообразие ARG были у тех, которые подверглись промышленному загрязнению антибиотиками [109]. ]. У сельскохозяйственных животных БРА обычно образуются в организме животных (особенно в желудочно-кишечном тракте) после применения антибиотиков. Иными словами, УПП во фруктах, овощах и других продуктах растительного происхождения часто возникает из-за загрязнения БРА и АРГ по всей пищевой цепи, от первичного производства до потребления [110]. Важными источниками микробного загрязнения в предуборочной среде являются почва, органические удобрения и поливная вода.
Трансдукция — важный механизм горизонтального переноса генов в естественных условиях, который традиционно недооценивался по сравнению с трансформацией. Исследование показало, что почвенные фаги были наиболее универсальными с точки зрения носительства ARG, а фаги из организованных ферм показали разные ARG по сравнению с неорганизованным сектором [111]. В другом исследовании фекалии свиней с трех коммерческих ферм подвергались скринингу на 32 клинически значимых типа ARG и было обнаружено, что ДНК бактериофага содержала 35,5% целевых типов ARG и sul1 , bla ТЕМ и ermB обнаружены в 100% образцов ДНК фага [112].
Используя показатель отношения обилия АРГ у бактериофагов и бактерий в качестве оценки способности бактериофагов передавать АРГ, было установлено, что отношение для qnrA было наибольшим (около 10 - 1 ) и отличалось от наиболее распространенного бактериофаг ARG ermB и fexA , а не floR имели наименьшее значение соотношения (около0020 − 6 ). Передача УПП из окружающей среды человеку
Антимикробный резистом укрывается; (i) Устойчивые к противомикробным препаратам бактерии, называемые переносчиками, которые могут распространять ARG в окружающей среде, но не могут колонизировать или заражать организм человека или животного.
(ii) Устойчивые к противомикробным препаратам бактерии, называемые переносчиками, которые могут колонизировать и иногда проникать в организм человека или животного [103]. Несмотря на то, что носители не способны колонизировать и заражать людей, их распространение и размножение в окружающей среде приведет к увеличению количества и разнообразия ARG в переносчиках.
Следовательно, это может увеличить риск передачи БРА человеку. Следует отметить, что большинство векторов не являются патогенами, потому что, даже если векторы могут колонизировать организм человека, у них могут отсутствовать важные гены вирулентности, и поэтому они не могут вызывать заболевание у здорового хозяина [103]. При поиске в литературе данных о воздействии на человека β-лактамаз расширенного спектра (БЛРС), продуцирующих Enterobacteriaceae, MRSA и устойчивых к ванкомицину Enterococcus spp. в окружающей среде обзорный документ, опубликованный в 2015 году, показал, что БРА были обнаружены в источниках загрязнения (66/66), таких как сточные воды и навоз, и не было обнаружено прямых доказательств передачи человеку через окружающую среду [113]. Хотя было проведено несколько исследований по молекулярному типированию изолятов человека и окружающей среды, только в одном был получен такой уровень доказательств, но направление передачи не могло быть определено (окружающая среда, передающая бактерии УПП человеку, или наоборот) [114].
Передача УПП от животных к человеку через пищевую цепочку или тесные контакты
Многие патогены животных являются зоонозными, поэтому любое развитие резистентности патогенов, связанных с пищевыми животными, может передаваться людям через пищевую цепь. Инфекции человека устойчивыми к антибиотикам патогенами, такими как Campylobacter spp., Salmonella spp., E. coli и S. aureus , увеличиваются [2].
Влияние животноводческих резервуаров на здоровье человека остается дискуссионным и неясным; тем не менее, есть несколько примеров прямых ссылок, которые были идентифицированы. В ESBL/AmpC и продуцирующих карбапенемазы Enterobacteriaceae , встречающиеся у животных, гены, кодирующие ESBL/AmpC или карбапенемазы, чаще всего расположены на МГЭ, что способствует их распространению [115]. В большинстве исследований в Африке среди БЛРС определенные клоны, несущие bla CTX-M-15 (ST131/B2 или ST405/D), в основном выявляются у людей.
Но они также были зарегистрированы у видов домашнего скота из Танзании, Нигерии или Туниса; Международная торговля мясом птицы, по-видимому, способствовала распространению других вариантов БЛРС, таких как CTX-M-14 и клоны [116]. Несмотря на то, что контакт с животными считается фактором риска, доказательства прямой передачи бактерий, продуцирующих БЛРС/AmpC, от животных к человеку при тесных контактах ограничены. Степень, в которой пища способствует потенциальной передаче продуцентов ESBL/AmpC человеку, также точно не установлена. Для тяжелых металлов была выявлена связь между сниженной восприимчивостью к цинку или меди и УПП среди изолятов свиней Salmonella , которые являются пищевыми патогенами [6]. Совместная передача генов tcrB , связанных с оттоком меди, и генов устойчивости к эритромицину erm(B) была доказана между видами домашнего скота из морских отложений Enterococcus hirae и E. faecalis в экспериментах по конъюгации. Эксперименты выявили возможности отбора УПП морской средой через тяжелые металлы и его возможное участие в устойчивых к антибиотикам энтерококковых инфекциях [117].
Более того, имеются убедительные доказательства явления корезистентности с участием Cu, макролидов и, возможно, ванкомицина среди энтерококков свиней, тогда как актуальность этого явления для болезнетворных штаммов у людей остается неопределенной [6]. Выводы и перспективы
Вклад УПП, первоначально отобранного для использования в сельскохозяйственном секторе, в устойчивость к патогенам человека точно неизвестен, но вряд ли им можно пренебречь. Поскольку режимы дозирования в сельском хозяйстве контролируются в меньшей степени, чем в здравоохранении, ветеринарные и экологические микробы часто подвергаются воздействию субингибирующих концентраций противомикробных препаратов, что рассматривается как фактор риска резистентности de novo, переноса ARG и селекции уже существующих сопротивления [118]. Основываясь на имеющихся данных, короткие курсы лечения максимальной дозой, не вызывающей неприемлемых побочных эффектов, могут быть оптимальными для достижения терапевтических целей при минимизации развития резистентности.
Новые подходы, такие как комбинированная или чередующаяся терапия, являются многообещающими, но их необходимо дополнительно изучить, прежде чем их можно будет внедрить в повседневную практику. Совместная селекция генов, придающих устойчивость к противомикробным препаратам, биоцидам, тяжелым металлам и другим опасным химическим веществам, представляет собой потенциально важное с экологической и клинической точки зрения явление. Неантибиотические соединения, используемые в агропродовольственном производстве, такие как антибактериальные биоциды и тяжелые металлы, также могут способствовать распространению УПП путем совместного отбора. Это может произойти, когда гены резистентности как к противомикробным препаратам, так и к металлам/биоцидам находятся вместе в одной и той же клетке (корезистентность), или когда один механизм устойчивости (эффлюксный насос) обеспечивает устойчивость как к противомикробным препаратам, так и к биоцидам/металлам (кросс-резистентность). устойчивости), что приводит к совместной селекции бактериальных штаммов или мобильных генетических элементов (МГЭ), которые они несут [119].
Агропищевая промышленность вынуждена сократить использование противомикробных соединений. Недавнее исследование, посвященное анализу УПП и потребления антибиотиков во всем мире в сравнении со многими потенциальными способствующими факторами, показало, что потребление антибиотиков не было значимо связано с индексом устойчивости к противомикробным препаратам. Это говорит о том, что снижения потребления антибиотиков будет недостаточно для борьбы с УПП, поскольку распространение резистентных штаммов и генов резистентности, по-видимому, является доминирующим фактором [120]. Более того, даже когда противомикробные соединения не используются, некоторые тяжелые металлы или биоциды могут поддерживать или даже повышать устойчивость бактерий к определенным агентам [6]. Таким образом, усилия одной страны по сокращению применения противомикробных препаратов в агропродовольственном производстве сами по себе не приведут к требуемому результату с точки зрения ограничения воздействия на потребителей.
Устойчивые зоонозные агенты и комменсальные штаммы, несущие гены AMR, попадают в человеческую популяцию различными путями, пищевые продукты являются лишь одним из них [113]. Улучшение санитарии, расширение доступа к чистой воде и обеспечение надлежащего управления, а также увеличение государственных расходов на здравоохранение и более эффективное регулирование частного сектора здравоохранения необходимы для снижения глобальной устойчивости к противомикробным препаратам [120]. Что касается агропродовольственной промышленности, то все страны должны разработать свод правил для снижения рисков для потребителей и сохранения существующих ценных химиотерапевтических препаратов для будущих поколений. Наличие данных и материалов
Неприменимо.
Сокращения
- AMR:
-
Устойчивость к противомикробным препаратам
- АРБ:
-
Устойчивые к противомикробным препаратам бактерии
- Аргумент:
-
Ген устойчивости к противомикробным препаратам
- БЛРС:
-
Бета-лактамаза расширенного спектра
- HGT:
-
Горизонтальный перенос генов
- MGE:
-
Мобильный генетический элемент
- Микрофон:
-
Минимальная ингибирующая концентрация
- MRSA:
-
Метициллин-резистентный Staphylococcus aureus
- ВНК:
-
Жизнеспособный, но не культивируемый
Ссылки
«>
Hao H, Cheng G, Iqbal Z, Ai X, Hussain HI, Huang L, et al. Преимущества и риски применения противомикробных препаратов у сельскохозяйственных животных. Фронт микробиол. 2014;5:288.
ПабМед ПабМед Центральный Статья Google ученый
-
-
Лекшми М., Аммини П., Кумар С., Варела М.Ф. Среда производства пищевых продуктов и развитие устойчивости к противомикробным препаратам у человека патогенов животного происхождения. Микроорганизмы. 2017;5(1):11.
Центральный пабмед Статья КАС Google ученый
-
Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, et al. Глобальные тенденции в использовании противомикробных препаратов у сельскохозяйственных животных. Proc Natl Acad Sci U S A. 2015;112(18):5649–54.
ПабМед ПабМед Центральный Статья КАС Google ученый
O’Neill J. Устойчивость к противомикробным препаратам: преодоление кризиса для здоровья и благосостояния наций. https://www.amr-revieworg/; 2014.
Google ученый
Андерссон Д.И., Хьюз Д. Микробиологические эффекты сублетальных доз антибиотиков. Nat Rev Microbiol. 2014;12(7):465–78.
КАС пабмед Статья Google ученый
Уэльс AD, Davies RH. Совместная селекция устойчивости к антибиотикам, биоцидам и тяжелым металлам и ее значение для патогенов пищевого происхождения. Антибиотики (Базель). 2015;4(4):567–604.
КАС Статья Google ученый
Пенесян А., Жиллингс М., Полсен И.Т. Открытие антибиотиков: борьба с устойчивостью бактерий в клетках и сообществах биопленок. Молекулы. 2015;20(4):5286–98.
КАС пабмед ПабМед Центральный Статья Google ученый
Бономо Р.А. Бета-лактамазы: внимание к текущим проблемам. Колд Спринг Харб Перспект Мед. 2017;7(1):a025239.
Wang X, Li H, Zhao C, Chen H, Liu J, Wang Z и др. Новая металло-бета-лактамаза NDM-9, идентифицированная из штамма ST107 Klebsiella pneumoniae, выделенного в Китае. Противомикробные агенты Int J. 2014;44(1):90–1.
КАС пабмед Статья Google ученый
Антонелли А., Д’Андреа М.М., Вагелли Г., Докье Д.Д., Россолини Г.М. OXA-372, новая карбапенем-гидролизующая бета-лактамаза класса D из Citrobacter freundii, выделенная из станции очистки сточных вод больницы. J Антимикробная химиотерапия. 2015;70(10):2749–56.
КАС пабмед Статья Google ученый
Рамирес М.С., Толмаски М.Е. Ферменты, модифицирующие аминогликозиды. Обновление устойчивости к наркотикам. 2010;13(6):151–71.
КАС пабмед ПабМед Центральный Статья Google ученый
Кэмерон А., Клима К.Л., Ха Р., Грунингер Р.Дж., Захир Р., Макаллистер Т.А. Новый ген устойчивости к аминогликозидам aadA у патогенов крупного рогатого скота и свиней. мсфера. 2018;3(1):e00568–17.
Wachino J, Arakawa Y. Экзогенно приобретенные метилтрансферазы 16S рРНК, обнаруженные в устойчивых к аминогликозидам патогенных грамотрицательных бактериях: обновление. Обновление устойчивости к наркотикам. 2012;15(3):133–48.
КАС пабмед Статья Google ученый
Хупер, округ Колумбия, Джейкоби, Джорджия. Ингибиторы топоизомеразы: фторхинолоновые механизмы действия и резистентность. Колд Спринг Харб Перспект Мед. 2016;6(9):a025320.
ПабМед ПабМед Центральный Статья КАС Google ученый
Fyfe C, Grossman TH, Kerstein K, Sutcliffe J. Устойчивость к макролидным антибиотикам у патогенов общественного здравоохранения. Колд Спринг Харб Перспект Мед. 2016;6(10):a025395.
ПабМед ПабМед Центральный Статья КАС Google ученый
Feßler AT, et al. Мобильные гены устойчивости к макролидам у стафилококков. Плазмида. 2018;99:2–10.
ПабМед Статья КАС Google ученый
МЦ Робертс. Обновленная информация о генах приобретенной устойчивости к тетрациклину. FEMS Microbiol Lett. 2005;245(2):195–203.
КАС пабмед Статья Google ученый
Feßler AT, et al. Мобильные гены устойчивости к линкозамидам у стафилококков. Плазмида. 2018;99:22–31.
ПабМед Статья КАС Google ученый
Schwarz S, Shen J, Kadlec K, Wang Y, Brenner Michael G, Fessler AT, et al. Линкозамиды, стрептограмины, фениколы и плевромутилины: механизм действия и механизмы резистентности. Колд Спринг Харб Перспект Мед. 2016;6(11):a027037.
ПабМед ПабМед Центральный Статья КАС Google ученый
Zhu XQ, Wang XM, Li H, Shang YH, Pan YS, Wu CM и др. Новый ген lnu(G), придающий устойчивость к линкомицину путем нуклеотидилирования, расположен на Tn6260 из enterococcus faecalis E531. J Антимикробная химиотерапия. 2017;72(4):993–7.
КАС пабмед Google ученый
Roberts MC, Schwarz S. Гены и механизмы устойчивости к тетрациклину и фениколу: важность для сельского хозяйства, окружающей среды и человека. J Environ Qual. 2016;45(2):576–92.
КАС пабмед Статья Google ученый
Ван Ю, Лв И, Цай Дж, Шварц С, Цуй Л, Ху З и др. Новый ген optrA, придающий переносимую устойчивость к оксазолидинонам и фениколам, и его присутствие в энтерококках faecalis и энтерококках faecium человеческого и животного происхождения. J Антимикробная химиотерапия. 2015;70(8):2182–90.
КАС пабмед Статья Google ученый
Poirel L, Jayol A, Nordmann P. Полимиксины: антибактериальная активность, тестирование чувствительности и механизмы резистентности, кодируемые плазмидами или хромосомами. Clin Microbiol Rev. 2017;30(2):557–96.
КАС пабмед ПабМед Центральный Статья Google ученый
Wang X, Wang Y, Zhou Y, Li J, Yin W, Wang S и др. Появление нового мобильного гена устойчивости к колистину, mcr-8, у Klebsiella pneumoniae, продуцирующей NDM. Новые микробы заражают. 2018;7(1):122.
ПабМед ПабМед Центральный Google ученый
Werner G, Strommenger B, Witte W. Приобретенная устойчивость к ванкомицину у клинически значимых патогенов. Будущая микробиология. 2008;3(5):547–62.
КАС пабмед Статья Google ученый
Ченг Г., Дай М., Ахмед С., Хао Х., Ван Х, Юань З. Противомикробные препараты в борьбе с устойчивостью к противомикробным препаратам. Фронт микробиол. 2016;7:470.
ПабМед ПабМед Центральный Google ученый
Аллен Х.К., Стэнтон Т.Б. Измененное эго: воздействие антибиотиков на микробиомы пищевых животных. Анну Рев Микробиол. 2014; 68: 297–315.
КАС пабмед Статья Google ученый
Perry J, Waglechner N, Wright G. Предыстория устойчивости к антибиотикам. Колд Спринг Харб Перспект Мед. 2016;6(6):a025197.
ПабМед ПабМед Центральный Статья Google ученый
D’Costa VM, King CE, Kalan L, Morar M, Sung WW, Schwarz C, et al. Устойчивость к антибиотикам возникла давно. Природа. 2011;477(7365):457–61.
КАС пабмед Статья Google ученый
Santiago-Rodriguez TM, Fornaciari G, Luciani S, Dowd SE, Toranzos GA, Marota I, et al. Микробиом кишечника доколумбовой андской мумии 11 века нашей эры. ПЛОС Один. 2015;10(9):e0138135.
ПабМед ПабМед Центральный Статья КАС Google ученый
Бхуллар К., Ваглехнер Н., Павловски А., Котева К., Бэнкс Э.Д., Джонстон М.Д. и др. Устойчивость к антибиотикам преобладает в микробиоме изолированной пещеры. ПЛОС Один. 2012;7(4):e34953.
КАС пабмед ПабМед Центральный Статья Google ученый
Li J, Xie S, Ahmed S, Wang F, Gu Y, Zhang C и др. Антимикробная активность и резистентность: влияющие факторы. Фронт Фармакол. 2017;8:364.
ПабМед ПабМед Центральный Статья КАС Google ученый
Drlica K, Zhao X. Гипотеза окна отбора мутантов обновлена. Клин Инфекция Дис. 2007;44(5):681–8.
ПабМед Статья Google ученый
Галлберг Э., Цао С., Берг О.Г., Илбак С., Сандегрен Л., Хьюз Д. и др. Селекция резистентных бактерий при очень низких концентрациях антибиотиков. PLoS Патог. 2011;7(7):e1002158.
КАС пабмед ПабМед Центральный Статья Google ученый
Кохански М.А., ДеПристо М.А., Коллинз Дж.Дж. Сублетальное лечение антибиотиками приводит к множественной лекарственной устойчивости посредством радикального мутагенеза. Мол Ячейка. 2010;37(3):311–20.
КАС пабмед ПабМед Центральный Статья Google ученый
Birkegard AC, Halasa T, Graesboll K, Clasen J, Folkesson A, Toft N. Связь между выбранными генами устойчивости к противомикробным препаратам и воздействием противомикробных препаратов на свинофермах в Дании. Научный доклад 2017; 7 (1): 9683.
ПабМед ПабМед Центральный Статья Google ученый
Mollenkopf DF, Mathys DA, Feicht SM, Stull JW, Bowman AS, Daniels JB, et al. Поддержание энтеробактерий, продуцирующих карбапенемазы, в системе производства свиней от опороса до откорма. Патог пищевого происхождения Dis. 2018;15(6):372–6.
КАС пабмед Статья Google ученый
Уэбб А.Л., Селинджер Л.Б., Табоада Э.Н., Инглис Г.Д. Подтип-специфический отбор на устойчивость к фторхинолонам, но не к тетрациклинам, очевиден в изолятах Campylobacter jejuni от мясного скота в замкнутых откормочных хозяйствах на юге Альберты, Канада. Appl Environ Microbiol. 2018;84(7):e02713–17.
Zhang WH, Zhang CZ, Liu ZJ, Gu XX, Li W, Yang L и др. In vitro развитие устойчивости к ципрофлоксацину Salmonella enterica Serovars typhimurium, Enteritidis и Indiana изолятов от пищевых животных. Устойчивость к микробам. 2017;23(6):687–94.
КАС пабмед Статья Google ученый
Medardus JJ, Molla BZ, Nicol M, Morrow WM, Rajala-Schultz PJ, Kazwala R, et al. Использование микронутриентов тяжелых металлов в кормах в свиноводческих системах США и его роль в сохранении полирезистентных сальмонелл. Appl Environ Microbiol. 2014;80(7):2317–25.
ПабМед ПабМед Центральный Статья КАС Google ученый
Берг Дж., Том-Петерсен А., Найбро О. Медная добавка в сельскохозяйственные почвы выбирает бактериальную устойчивость к антибиотикам в полевых условиях. Lett Appl Microbiol. 2005;40(2):146–51.
КАС пабмед Статья Google ученый
Chenia HY, Jacobs A. Устойчивость к противомикробным препаратам, устойчивость к тяжелым металлам и содержание интегронов в бактериях, выделенных из системы аквакультуры тилапии в Южной Африке. Орган «Дис Аква». 2017;126(3):199–209.
КАС пабмед Статья Google ученый
Zou X, Weng M, Ji X, Guo R, Zheng W, Yao W. Сравнение устойчивости Enterococcus spp. к антибиотикам и толерантности к меди. и лактобациллы spp. выделены от поросят до и после отъема. J микробиол. 2017;55(9):703–10.
КАС пабмед Статья Google ученый
Hasman H, Kempf I, Chidaine B, Cariolet R, Ersboll AK, Houe H, et al. Устойчивость к меди у Enterococcus faecium, опосредованная геном tcrB, выбирается путем добавления в корм для свиней сульфата меди. Appl Environ Microbiol. 2006;72(9): 5784–9.
КАС пабмед ПабМед Центральный Статья Google ученый
Язданхах С., Руди К., Бернхофт А. Цинк и медь в кормах для животных – развитие резистентности и корезистентности к противомикробным препаратам у бактерий животного происхождения. Microb Ecol Health Дис. 2014;25(1):25862.
Амачавади Р.Г., Скотт Х.М., Нитиканчана С., Винаско Дж., Токач М.Д., Дритц С.С. и др. Назальное носительство mecA-положительного метициллин-резистентного золотистого стафилококка у свиней демонстрирует дозозависимую реакцию на добавки цинка. Патог пищевого происхождения Dis. 2015;12(2):159–63.
КАС пабмед Статья Google ученый
Slifierz MJ, Friendship R, Weese JS. Терапия оксидом цинка увеличивает распространенность и персистенцию метициллин-резистентного золотистого стафилококка у свиней: рандомизированное контролируемое исследование. Зоонозы Общественное здравоохранение. 2015;62(4):301–8.
КАС пабмед Статья Google ученый
Кавако Л.М., Хасман Х., Аареструп FM. Резистентность золотистого стафилококка животного происхождения к цинку тесно связана с резистентностью к метициллину. Вет микробиол. 2011; 150(3–4):344–8.
КАС пабмед Статья Google ученый
Ji X, Shen Q, Liu F, Ma J, Xu G, Wang Y и др. Обилие генов устойчивости к антибиотикам, связанное с антибиотиками и тяжелыми металлами в навозе животных и сельскохозяйственных почвах, прилегающих к откормочным площадкам в Шанхае; Китай. Джей Хазард Матер. 2012; 235–236:178–85.
КАС пабмед Статья Google ученый
Андраде Л.Н., Сикейра Т.Е.С., Мартинес Р., Дарини А.Л.К. CTX-M-(15, 9) с множественной лекарственной устойчивостью, 2)- и KPC-2-продуцирующие изоляты Enterobacter hormaechei и Enterobacter asburiae обладали набором генов приобретенной толерантности к тяжелым металлам, включая хромосомный sil-оперон (для приобретенной устойчивости к серебру). Фронт микробиол. 2018;9:539.
ПабМед ПабМед Центральный Статья Google ученый
Виньяроли С., Паскуароли С., Читтерио Б., Ди Чезаре А., Манджатерра Г., Фатторини Д. и др. Устойчивость к антибиотикам и тяжелым металлам энтерококков из прибрежных морских отложений. Загрязнение окружающей среды. 2018; 237:406–13.
КАС пабмед Статья Google ученый
Stepanauskas R, Glenn TC, Jagoe CH, Tuckfield RC, Lindell AH, King CJ, et al. Коселекция микробной устойчивости к металлам и антибиотикам в пресноводных микрокосмах. Окружающая среда микробиол. 2006;8(9):1510–4.
КАС пабмед Статья Google ученый
Кнапп К.В., Маккласки С.М., Сингх Б.К., Кэмпбелл К.Д., Хадсон Г., Грэм Д.В. Обилие генов устойчивости к антибиотикам коррелирует с металлами и геохимическими условиями в заархивированных шотландских почвах. ПЛОС Один. 2011;6(11):e27300.
КАС пабмед ПабМед Центральный Статья Google ученый
Аргудин М.А., Лаузат Б., Краушаар Б., Альба П., Агерсо Ю., Кавако Л. и др. Гены устойчивости к тяжелым металлам и дезинфектантам среди изолятов Staphylococcus aureus, устойчивых к метициллину, связанных с домашним скотом. Вет микробиол. 2016; 191:88–95.
КАС пабмед Статья Google ученый
Грасс Г., Ренсинг С., Солиоз М. Металлическая медь как антимикробная поверхность. Appl Environ Microbiol. 2011;77(5):1541–1547.
КАС пабмед Статья Google ученый
Сингер А.С., Шоу Х., Родс В., Харт А. Обзор устойчивости к противомикробным препаратам в окружающей среде и ее актуальность для природоохранных органов. Фронт микробиол. 2016;7:1728.
ПабМед ПабМед Центральный Статья Google ученый
Пул К. На стыке антибиотиков и металлов: влияние Cu и Zn на активность и устойчивость к антибиотикам. Тенденции микробиол. 2017;25(10):820–32.
КАС пабмед Статья Google ученый
Ю З., Ганн Л., Уолл П., Фаннинг С. Устойчивость к противомикробным препаратам и ее связь с толерантностью к тяжелым металлам в сельскохозяйственном производстве. Пищевой микробиол. 2017;64:23–32.
КАС пабмед Статья Google ученый
Gomez-Sanz E, Kadlec K, Fessler AT, Zarazaga M, Torres C, Schwarz S. Novel erm(T), несущие полирезистентные плазмиды из свиных и человеческих изолятов метициллин-резистентного Staphylococcus aureus ST398, которые также содержат детерминанты устойчивости к кадмию и меди. Противомикробные агенты Chemother. 2013;57(7):3275–82.
КАС пабмед ПабМед Центральный Статья Google ученый
Cavaco LM, Hasman H, Stegger M, Andersen PS, Skov R, Fluit AC, et al. Клонирование и возникновение czrC, гена, придающего устойчивость к кадмию и цинку в устойчивых к метициллину изолятах Staphylococcus aureus CC398. Противомикробные агенты Chemother. 2010;54(9): 3605–8.
КАС пабмед ПабМед Центральный Статья Google ученый
Aarestrup FM, Cavaco L, Hasman H. Снижение восприимчивости к хлориду цинка связано с устойчивым к метициллину Staphylococcus aureus CC398 у датских свиней. Вет микробиол. 2010; 142(3–4):455–7.
КАС пабмед Статья Google ученый
Петрова М., Горленко З., Миндлин С. Tn5045, новый интегрон-содержащий антибиотик и транспозон устойчивости к хроматам, выделенный из вечномерзлых бактерий. Рез микробиол. 2011;162(3):337–45.
КАС пабмед Статья Google ученый
Mourao J, Novais C, Machado J, Peixe L, Antunes P. Толерантность к металлам в новых клинически значимых клонах сальмонеллы enterica с множественной лекарственной устойчивостью серотипа 4,[5],12:i:-, циркулирующих в Европе. Противомикробные агенты Int J. 2015;45(6):610–6.
КАС пабмед Статья Google ученый
Амачавади Р.Г., Шелтон Н.В., Ши Х., Винаско Дж., Дритц С.С., Токач М.Д. и др. Отбор фекальных энтерококков, проявляющих tcrB-опосредованную резистентность к меди, у свиней, получавших диеты с добавками меди. Appl Environ Microbiol. 2011;77(16):5597–603.
КАС пабмед ПабМед Центральный Статья Google ученый
Zhang S, Wang D, Wang Y, Hasman H, Aarestrup FM, Alwathnani HA и др. Геномные последовательности устойчивых и чувствительных к меди штаммов Enterococcus faecalis, выделенных от свиней, которых кормили медью, в Дании. Стенд Genomic Sci. 2015;10:35.
КАС пабмед ПабМед Центральный Статья Google ученый
Пул К. Стрессовые реакции как детерминанты устойчивости грамотрицательных бактерий к противомикробным препаратам. Тенденции микробиол. 2012;20(5):227–34.
КАС пабмед Статья Google ученый
Audrain B, Ferrieres L, Zairi A, Soubigou G, Dobson C, Coppee JY, et al. Индукция пути стресса оболочки Cpx способствует устойчивости Escherichia coli к антимикробным пептидам. Appl Environ Microbiol. 2013;79(24): 7770–9.
КАС пабмед ПабМед Центральный Статья Google ученый
Yamamoto K, Ishihama A. Характеристика индуцируемых медью промоторов, регулируемых CpxA/CpxR в Escherichia coli. Биоски Биотехнолог Биохим. 2006;70(7):1688–95.
КАС пабмед Статья Google ученый
Caille O, Rossier C, Perron K. Активируемая медью двухкомпонентная система взаимодействует с устойчивостью Pseudomonas aeruginosa к цинку и имипенему. J Бактериол. 2007;189(13): 4561–8.
КАС пабмед ПабМед Центральный Статья Google ученый
Мата М.Т., Бакеро Ф., Перес-Диас Х.К. Переносчик оттока многих лекарств в Listeria monocytogenes. FEMS Microbiol Lett. 2000;187(2):185–8.
КАС пабмед Статья Google ученый
Хейнс-Менгес Б., Уитакер В.Б., Бойд Э.Ф. Альтернативный сигма-фактор RpoE важен для стрессовой реакции клеточной оболочки Vibrio parahaemolyticus и кишечной колонизации. Заразить иммун. 2014;82(9): 3667–77.
ПабМед ПабМед Центральный Статья КАС Google ученый
Эглер М., Гросс К., Грасс Г., Нис Д.Х. Роль сигма-фактора семейства белков экстрацитоплазматической функции RpoE в резистентности Escherichia coli к металлам. J Бактериол. 2005;187(7):2297–307.
КАС пабмед ПабМед Центральный Статья Google ученый
Harrison JJ, Tremaroli V, Stan MA, Chan CS, Vacchi-Suzzi C, Heyne BJ, et al. Хромосомные гены антиоксидантов играют роль, специфичную для ионов металлов, как детерминанты толерантности бактерий к металлам. Окружающая среда микробиол. 2009 г.;11(10):2491–509.
КАС пабмед Статья Google ученый
Харрисон Дж.Дж., Кери Х., Стремик К.А., Тернер Р.Дж. Восприимчивость биопленки к токсичности металлов. Окружающая среда микробиол. 2004;6(12):1220–7.
КАС пабмед Статья Google ученый
Бейкер Дж., Ситтисак С., Сенгупта М., Джонсон М., Джаясвал Р.К., Моррисси Дж.А. Медный стресс индуцирует глобальную стрессовую реакцию у Staphylococcus aureus и подавляет экспрессию sae и agr и образование биопленок. Appl Environ Microbiol. 2010;76(1):150–60.
КАС пабмед Статья Google ученый
Айрапетян М., Уильямс Т.С., Оливер Д.Д. Преодоление разрыва между жизнеспособными, но не культивируемыми бактериями и устойчивыми к антибиотикам бактериями. Тенденции микробиол. 2015;23(1):7–13.
КАС пабмед Статья Google ученый
Navarrete F, De La Fuente L. Реакция Xylella fastidiosa на цинк: снижение культивируемости, увеличение продукции экзополисахарида и образование устойчивых биопленок в условиях потока. Appl Environ Microbiol. 2014;80(3):1097–107.
ПабМед ПабМед Центральный Статья КАС Google ученый
Wei Q, Tarighi S, Dotsch A, Haussler S, Musken M, Wright VJ, et al. Фенотипический и полногеномный анализ устойчивого к антибиотикам варианта малых колоний (SCV) Pseudomonas aeruginosa. ПЛОС Один. 2011;6(12):e29276.
КАС пабмед ПабМед Центральный Статья Google ученый
Zhang Y, Gu AZ, Cen T, Li X, He M, Li D и др. Субингибирующие концентрации тяжелых металлов облегчают горизонтальный перенос плазмид-опосредованных генов устойчивости к антибиотикам в водной среде. Загрязнение окружающей среды. 2018; 237:74–82.
КАС пабмед Статья Google ученый
Zhang M, Chen L, Ye C, Yu X. Совместная селекция устойчивости к антибиотикам с помощью ударной нагрузки медью на бактерии из биофильтра питьевой воды. Загрязнение окружающей среды. 2018; 233:132–41.
КАС пабмед Статья Google ученый
SCENIHR. Оценка эффектов антибиотикорезистентности биоцидов. Брюссель: Научный комитет по возникающим и недавно выявленным рискам для здоровья, Европейская комиссия; 2009. с. 1–87.
Google ученый
Beier RC, Anderson PN, Hume ME, Poole TL, Duke SE, Crippen TL, et al. Характеристика изолятов Salmonella enterica от индеек на коммерческих перерабатывающих предприятиях на устойчивость к антибиотикам, дезинфицирующим средствам и стимуляторам роста. Патог пищевого происхождения Dis. 2011;8(5):593–600.
КАС пабмед Статья Google ученый
Роуз Х., Болдуин А., Доусон К.Г., Махентиралингам Э. Чувствительность комплекса Burkholderia cepacia к биоцидам. J Антимикробная химиотерапия. 2009;63(3):502–10.
КАС пабмед ПабМед Центральный Статья Google ученый
Туролла А., Сабатино Р., Фонтането Д., Эккерт Э.М., Колинас Н., Корно Г. и др. Стратегии защиты и изобилие генов устойчивости к антибиотикам у энтерококков в условиях стресса при воздействии низких доз надуксусной кислоты. Хемосфера. 2017; 185:480–8.
КАС пабмед Статья Google ученый
Кампф Г. Приобретенная устойчивость к хлоргексидину – не пора ли начать инициативу «антисептическое управление»? Джей Хосп заражает. 2016;94(3):213–27.
КАС пабмед Статья Google ученый
Martin H, Maris P. Устойчивость к антисептикам и антибиотикам 310 грамположительных штаммов, выделенных из вымени после применения гермицидов для сосков после доения. Вет Рез. 1995;26(1):43–56.
КАС пабмед Google ученый
Romero JL, Grande Burgos MJ, Perez-Pulido R, Galvez A, Lucas R. Устойчивость к антибиотикам, биоцидам, консервантам и металлам у бактерий, выделенных из морепродуктов: совместный отбор штаммов, устойчивых или толерантных к различным классам соединений . Фронт микробиол. 2017;8:1650.
ПабМед ПабМед Центральный Статья Google ученый
Браудаки М., Hilton AC. Адаптивная устойчивость к биоцидам у Salmonella enterica и Escherichia coli O157 и перекрестная устойчивость к противомикробным препаратам. Дж. Клин Микробиол. 2004;42(1):73–8.
КАС пабмед ПабМед Центральный Статья Google ученый
Soumet C, Fourreau E, Legrandois P, Maris P. Устойчивость к соединениям феникола после адаптации к соединениям четвертичного аммония у Escherichia coli. Вет микробиол. 2012;158(1–2):147–52.
КАС пабмед Статья Google ученый
Потенски С.Дж., Ганди М., Мэтьюз К.Р. Воздействие на Salmonella Enteritidis хлора или пищевых консервантов снижает [скорректированную] восприимчивость к антибиотикам. FEMS Microbiol Lett. 2003;220(2):181–6.
КАС пабмед Статья Google ученый
Тандукар М, О С, Тезел У, Константинидис К.Т., Павлостатис С.Г. Длительное воздействие дезинфицирующих средств на основе хлорида бензалкония приводит к изменению структуры микробного сообщества и повышению устойчивости к противомикробным препаратам. Технологии экологических наук. 2013;47(17):9730–8.
КАС пабмед Статья Google ученый
Бхардвадж П., Зиглер Э., Палмер К.Л. Хлоргексидин индуцирует гены устойчивости к ванкомицину VanA-типа у энтерококков. Противомикробные агенты Chemother. 2016;60(4):2209–21.
КАС пабмед ПабМед Центральный Статья Google ученый
Fuentes MAF, Morente EO, Abriouel H, Pulido RP, Galvez A. Детерминанты устойчивости к противомикробным препаратам грамотрицательных бактерий, устойчивых к антибиотикам и биоцидам из органических продуктов питания. Пищевой контроль. 2014;37:9–14.
Артикул КАС Google ученый
Buffet-Bataillon S, Le Jeune A, Le Gall-David S, Bonnaure-Mallet M, Jolivet-Gougeon A. Молекулярные механизмы более высоких МИК антибиотиков и четвертичных аммониевых соединений для Escherichia coli, выделенной из бактериемии. J Антимикробная химиотерапия. 2012;67(12):2837–42.
КАС пабмед Статья Google ученый
Buffet-Bataillon S, Tattevin P, Bonnaure-Mallet M, Jolivet-Gougeon A. Возникновение устойчивости к антибактериальным агентам: роль соединений четвертичного аммония — критический обзор. Противомикробные агенты Int J. 2012;39(5):381–9.
КАС пабмед Статья Google ученый
Ортега Моренте Э., Фернандес-Фуэнтес М.А., Гранде Бургос М.Дж., Абриуэль Х., Перес Пулидо Р., Гальвес А. Толерантность бактерий к биоцидам. Int J Food Microbiol. 2013;162(1):13–25.
ПабМед Статья КАС Google ученый
Конфикони Д., Лосассо С., Кортини Э., Ди Чезаре А., Чибин В., Джакконе В. и др. Устойчивость к биоцидам у Listeria monocytogenes, собранного на предприятиях по переработке мяса. Фронт микробиол. 2016;7:1627.
ПабМед ПабМед Центральный Статья Google ученый
Пажедар А., Сингх Дж., Батиш В.К. Адаптация к хлориду бензалкония и ципрофлоксацину влияет на потенциал образования биопленки, эффлюксную помпу и активность гемолизина Escherichia coli молочного происхождения. Джей Молочные Рез. 2012;79(4): 383–9.
КАС пабмед Статья Google ученый
Шеридан А., Ленахан М., Конделл О., Бонилья-Сантьяго Р., сержант К., Рено Дж. и др. Протеомный и фенотипический анализ толерантной к триклозану вероцитотоксигенной Escherichia coli O157:h29. J Протеомика. 2013;80:78–90.
КАС пабмед Статья Google ученый
Пал К., Бенгтссон-Палме Дж., Кристианссон Э., Ларссон Д.Г. Совместное появление генов резистентности к антибиотикам, биоцидам и металлам открывает новое понимание их потенциала совместной селекции. Геномика BMC. 2015;16:964.
ПабМед ПабМед Центральный Статья КАС Google ученый
Харди К., Саннакс К., Гил Х., Шабир С., Трампари Э., Хоуки П. и др. Более широкое использование антисептиков связано со снижением восприимчивости клинических изолятов Staphylococcus aureus . МБио. 2018;9(3):e00894–18.
Бас С., Крамер М., Стопар Д. Поверхностная плотность биопленки определяет эффективность биоцида. Фронт микробиол. 2017;8:2443.
ПабМед ПабМед Центральный Статья Google ученый
Манайя СМ. Оценка риска передачи устойчивости к антибиотикам из окружающей среды человеку: непрямая зависимость между распространенностью и риском. Тенденции микробиол. 2017;25(3):173–81.
КАС пабмед Статья Google ученый
Уолш Т.Р., Уикс Дж., Ливермор Д.М., Тоулман М.А. Распространение NDM-1 положительных бактерий в окружающей среде Нью-Дели и его последствия для здоровья человека: исследование распространенности в окружающей среде. Ланцет Infect Dis. 2011;11(5):355–62.
ПабМед Статья Google ученый
Николя-Чанойн М.Х., Бланко Дж., Лефлон-Гибу В., Демарти Р., Алонсо М.П., Каника М.М. и др. Межконтинентальное появление клона Escherichia coli O25:h5-ST131, продуцирующего CTX-M-15. J Антимикробная химиотерапия. 2008;61(2):273–81.
КАС пабмед Статья Google ученый
Rao Q, Shang W, Hu X, Rao X. Staphylococcus aureus ST121: повсеместно распространенный гипервирулентный клон. J Med Microbiol. 2015;64(12):1462–73.
КАС пабмед Статья Google ученый
Xie WY, Yang XP, Li Q, Wu LH, Shen QR, Zhao FJ. Изменения концентрации антибиотиков и резистома антибиотиков при коммерческом компостировании навоза животных. Загрязнение окружающей среды. 2016; 219:182–90.
КАС пабмед Статья Google ученый
Zhang YJ, Hu HW, Gou M, Wang JT, Chen D, He JZ. Временная последовательность генов устойчивости к антибиотикам в почве после внесения навоза свиней, крупного рогатого скота и домашней птицы с добавлением антибиотиков или без них. Загрязнение окружающей среды. 2017; 231 (часть 2): 1621–32.
КАС пабмед Статья Google ученый
Пал К., Бенгтссон-Палме Дж., Кристианссон Э., Ларссон Д.Г. Структура и разнообразие резистом человека, животных и окружающей среды. Микробиом. 2016;4(1):54.
ПабМед ПабМед Центральный Статья Google ученый
ВОЗ. Совместное совещание экспертов ФАО/ВОЗ по устойчивости к противомикробным препаратам пищевого происхождения: роль окружающей среды, сельскохозяйственных культур и биоцидов. http://www.hoint/foodsafety/areas_work/antimicrobial-resistance/FAO_WHO_AMR_Summary_Report_June2018pdf?ua=1; 2018.
Google ученый
Ананд Т., Бера Б.К., Вайд Р.К., Баруа С., Риеш Т., Вирмани Н. и др. Изобилие генов устойчивости к антибиотикам у экологических бактериофагов. Джей Ген Вирол. 2016;97(12):3458–66.
КАС пабмед Статья Google ученый
Wang M, Liu P, Zhou Q, Tao W, Sun Y, Zeng Z. Оценка вклада бактериофагов в распространение генов устойчивости к антибиотикам в экскрементах свиней. Загрязнение окружающей среды. 2018;238:291–8.
КАС пабмед Статья Google ученый
Huijbers PM, Blaak H, de Jong MC, Graat EA, Vandenbroucke-Grauls CM, de Roda Husman AM. Роль окружающей среды в передаче устойчивости к противомикробным препаратам человеку: обзор. Технологии экологических наук. 2015;49(20):11993–2004.
КАС пабмед Статья Google ученый
Hower S, Phillips MC, Brodsky M, Dameron A, Tamargo MA, Salazar NC, et al. Клонально-родственный метициллин-резистентный золотистый стафилококк, выделенный от короткоплавниковой гринды (Globicephala macrorhynchus), людей-добровольцев и в реабилитационном центре для китообразных на берегу залива. Микроб Экол. 2013;65(4):1024–38.
КАС пабмед Статья Google ученый
Madec JY, Haenni M, Nordmann P, Poirel L. Enterobacteriaceae, продуцирующие бета-лактамазы расширенного спектра/AmpC и карбапенемазы у животных: угроза для человека? Клин Микробиол Инфект. 2017;23(11):826–33.
КАС пабмед Статья Google ученый
Алонсо К.А., Заразага М., Бен Саллем Р., Джуини А., Бен Слама К., Торрес К. Устойчивость к антибиотикам кишечной палочки у сельскохозяйственных животных: африканский взгляд. Lett Appl Microbiol. 2017;64(5):318–34.
КАС пабмед Статья Google ученый
Pasquaroli S, Di Cesare A, Vignaroli C, Conti G, Citterio B, Biavasco F. Устойчивый к эритромицину и меди энтерококк hirae из морских отложений и совместный перенос erm(B) и tcrB в человеческий enterococcus faecalis. Диагностика Microbiol Infect Dis. 2014;80(1):26–8.
КАС пабмед Статья Google ученый
Ter Kuile BH, Kraupner N, Brul S. Риск низких концентраций антибиотиков в сельском хозяйстве для устойчивости в здравоохранении. FEMS Microbiol Lett. 2016;363(19). https://doi.org/10.1093/femsle/fnw210.
ПабМед Статья КАС Google ученый
Пал С., Азиани К., Арья С., Ренсинг С., Стекель Д.Дж., Ларссон Д.Дж. и др. Устойчивость к металлам и ее связь с устойчивостью к антибиотикам. Ад Микроб Физиол. 2017;70:261–313.
ПабМед Статья Google ученый
Коллиньон П., Беггс Дж.Дж., Уолш Т.Р., Гандра С., Лаксминараян Р. Антропологические и социально-экономические факторы, способствующие глобальной устойчивости к противомикробным препаратам: одномерный и многомерный анализ. Ланцет Планета Здоровье. 2018;2(9):e398–405.
ПабМед Статья Google ученый
Ссылки на скачивание
Благодарности
Авторы благодарят Национальный фонд естественных наук Китая и Национальную ключевую программу исследований и разработок Китая за финансирование этого исследования.
Финансирование
Это исследование финансировалось Национальной ключевой программой исследований и разработок Китая (2017YFC1600100; 2018YFD0500300) и Национальным фондом естественных наук Китая (31502115).
Информация об авторе
Примечания автора
-
Гую Ченг и Цзяньань Нин внесли равный вклад в эту работу.
Авторы и филиалы
-
MOA Лаборатория оценки рисков качества и безопасности продуктов животноводства и птицеводства, Хуачжунский сельскохозяйственный университет, Ухань, 430070, Китай
Guyue Cheng, Jianan Ning, Saeed Ahmed, Boyu Anyu An, Junhong Huang , Haihong Hao, Menghong Dai, Xu Wang & Zonghui Yuan
-
Национальная справочная лаборатория остатков ветеринарных препаратов (HZAU) и Ключевая лаборатория MOA по обнаружению остатков ветеринарных препаратов в пищевых продуктах, Хуажонгский сельскохозяйственный университет, Ухань, 430070, Китай
Lingli Huang & Zonghui Yuan
-
Государственная ключевая лаборатория сельскохозяйственной микробиологии, Колледж ветеринарной медицины, Huazhong Angirlault University, Wuhan, 430070, China
Rizwan Ullah
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Вклады
GC и ZY внесли свой вклад в концепцию и дизайн исследования; GC, JH и BA исследовали и изучили литературу; GC написал первый черновик рукописи; GC, JN, SA, HH, MD, LH и XW отредактировали рукопись. Все авторы внесли свой вклад в доработку рукописи, прочитали и одобрили представленную версию.
Автор, ответственный за переписку
Переписка с Гую Ченг.
Декларация этики
Одобрение этики и согласие на участие
Неприменимо.
Согласие на публикацию
Не применимо.
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя
Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Права и разрешения
Открытый доступ Эта статья распространяется в соответствии с условиями международной лицензии Creative Commons Attribution 4.0 (http://creativecommons.org/licenses/by/4.0/), которая разрешает неограниченное использование, распространение, и воспроизведение на любом носителе, при условии, что вы укажете автора(ов) оригинала и источник, предоставите ссылку на лицензию Creative Commons и укажете, были ли внесены изменения. Отказ от права Creative Commons на общественное достояние (http://creativecommons.org/publicdomain/zero/1.0/) применяется к данным, представленным в этой статье, если не указано иное.
Перепечатки и разрешения
Об этой статье
Адаптивная лабораторная эволюция устойчивости к антибиотикам с использованием различных режимов селекции приводит к сходным фенотипам и генотипам
Введение
Экосистемы постоянно претерпевают изменения в своих физических и химических свойствах, что приводит к смещению экологических ниш и условия жизни (Hoffmann and Parsons, 1997; Elmqvist et al., 2003; Fine, 2015). Бактериальные популяции могут реагировать на эти изменения окружающей среды посредством временной и постоянной адаптации. Временная адаптация включает в себя модуляцию экспрессии генов, приводящую к фенотипическим изменениям, обусловленным изменениями сигналов окружающей среды, воспринимаемых бактериями (López-Maury et al., 2008). Напротив, отбор полезных мутаций или горизонтальное приобретение полезных генов представляет собой постоянную генетическую адаптацию к изменившейся среде. Будет ли изменение окружающей среды встречено временной или постоянной адаптацией, во многом зависит от силы и продолжительности давления отбора. Существенные изменения в окружающей среде, как и в случае лечения антибиотиками, могут привести как к временным адаптациям, таким как метаболические изменения, приводящие к появлению устойчивых к антибиотикам бактерий (Levin et al., 2014), так и к постоянным адаптациям, которые иногда приводят к появлению устойчивых к антибиотикам бактерий (Carroll et al. и др., 2014; Дхавале и Рат, 2014).
Этот адаптивный потенциал микроорганизмов все чаще исследуется в биотехнологии с помощью экспериментов по адаптивной лабораторной эволюции (ALE) (Blum et al., 2016). ALE можно использовать для улучшения производственных штаммов за счет повышения их устойчивости к продукту метаболизма (Hu et al., 2016; Lennen, 2016), путем активации латентных путей (Wang et al., 2016) или за счет использования ненативных субстраты (Lee and Palsson, 2010). Кроме того, эксперименты с ALE могут улучшить наше понимание фундаментальных принципов эволюции, что может помочь нам решить растущие глобальные проблемы нежелательной адаптации, такие как лекарственная устойчивость микробных патогенов (Anderson et al. , 19).50; Гован и Файф, 1978 г .; Коэн и др., 1989; Дональд и ван Хелден, 2009 г .; Wensing et al., 2015), рак (Riganti et al., 2015) или устойчивость насекомых к пестицидам (Georghiou, 2012). Обычно эксперименты с ALE сосредоточены на адаптации к конкретным физическим или химическим факторам, таким как температура (Tenaillon et al., 2012; Sandberg et al., 2014) или устойчивость к антибиотикам (Hegreness et al., 2008; Toprak et al., 2012; Лазар и др., 2014; Мунк и др., 2014; Родригес де Евграфов и др., 2015). Различные установки ALE использовались для изучения подобных возмущений окружающей среды, таких как воздействие различных антибиотиков (Hegreness et al., 2008; Toprak et al., 2012; Lázár et al., 2014; Munck et al., 2014; Rodriguez de Evgrafov et al. ., 2015), однако влияние экспериментальной установки на результирующие адаптации остается малоизученным.
В этом исследовании мы используем эволюцию устойчивости к антибиотикам в качестве модели для изучения влияния экспериментальной установки на развившиеся фенотипы и генотипы. Предыдущие исследования позволили бактериям развиться до высокого уровня устойчивости к антибиотикам с использованием различных методологий, включая градиенты повышения концентрации лекарственного средства (Kim et al., 2014; Munck et al., 2014; Oz et al., 2014; Rodriguez de Evgrafov et al., 2015). ), поэтапное воздействие антибиотиков (Lázár et al., 2014) или постепенное увеличение концентрации препарата в морбидостате (Toprak et al., 2012). Эти различные подходы используют различные факторы давления отбора, размеры популяций и узкие места — все они, как известно, влияют на эволюцию (Nei et al., 19).75; Левин и др., 2000; Валь и др., 2002; Чарльзворт, 2009). Влияние давления отбора на результирующие фено- и генотипы было оценено путем сравнения бактерий, подвергшихся мягкому и сильному отбору, в градиентном подходе (Oz et al., 2014), а также при заражении бактериями с инкрементами препарата, различающимися по степени крутизна увеличения дозы препарата (Lindsey et al., 2013). Выявлены различия в количестве мутаций, скорости роста и фенотипах устойчивости в зависимости от используемой методологии (Lindsey et al. , 2013; Oz et al., 2014). В то время как градиентный метод применяет максимальное давление отбора, что приводит к быстрому созданию высокоустойчивых линий, инкрементальный подход требует меньше лабораторных ресурсов, что позволяет проводить исследования большего количества повторов и условий, таких как разные антибиотики, без увеличения времени обработки. Тем не менее, скорость увеличения лекарственного средства фиксирована в подходе приращения, что может привести к как низкому, так и слишком высокому давлению отбора в зависимости от уровня адаптации бактерий.
В этом исследовании мы сравниваем различные подходы к приращению, различающиеся по крутизне увеличения количества препарата, с градиентным подходом, чтобы исследовать, как режим селекции, определяемый методологией ALE, влияет на результирующие гено- и фенотипы.
Материалы и методы
Лабораторная адаптивная эволюция в градиентах лекарств
Escherichia coli K12 (MG1655) подвергали эволюции в течение 14 дней к трем различным антибиотикам: сульфату амикацина (AMK) (Sigma), сульфату пиперациллина (PIP) (Sigma) , и тетрациклина гидрохлорид (ТЕТ) (Sigma), охватывающий три основных класса антибиотиков, включая как бактерицидные, так и бактериостатические препараты. Антибиотики растворяли в воде (10 мг/л), исходные растворы хранили при -20°С. Для каждого препарата параллельно развивались четыре репликативные линии. 96-луночные планшеты (Almeco), содержащие 1 мл бульона Мюллера-Хинтона II (MHBII) (Sigma) на лунку и 2-кратный градиент антибиотика в 10 разведениях, готовили в начале эксперимента и хранили при -20°C. . Минимальная ингибирующая концентрация (МИК) дикого типа, определенная Европейским комитетом по тестированию чувствительности к антибиотикам (EUCAST), находилась во второй лунке, что позволяло дикому типу расти в первой лунке в субингибирующих условиях (точное установка планшета и концентрации лекарств приведены в дополнительной таблице 1). Планшеты размораживали в день использования, предварительно нагревали до 37°С, инокулировали 50 мкл свежерастущих клеток и инкубировали при температуре ~9°С.00 об/мин и 37°С в течение 22 часов. Сто пятьдесят микролитров каждой лунки переносили в 96-луночный титрационный микропланшет и измеряли оптическую плотность при длине волны 600 нм (OD 600 ) с помощью ELx808 Absorbance Reader (BioTek). На основании измерения OD было выбрано пороговое значение, то есть минимальный рост, который явно отличался от фонового роста, чтобы определить отчетливый рост для каждого препарата (дополнительная фигура 1). OD 600 > 0,1, что соответствует ~8,0 × 10 7 КОЕ/мл определяемый отчетливый рост для AMK и TET и OD 600 > 0,3 эквивалент примерно 2,4 × 10 8 КОЕ/мл определенный рост для PIP из-за фонового уровня роста около OD 600 = 0,18. Пятьдесят микролитров лунки с самой высокой концентрацией лекарственного средства, которая показала отчетливый рост в планшете с глубокими лунками, использовали для инокуляции свежего градиента (точные значения OD 600 и соответствующие концентрации лекарственного средства в лунке, выбранной для каждого переноса, приведены в дополнительной таблице. 2). Оставшиеся клетки в этих лунках смешивали до конечной концентрации глицерина 20% и хранили при -80°С. На каждом планшете 16 лунок служили отрицательным контролем, в результате всего в ходе эксперимента было 448 лунок, из которых 1% показал рост. Для каждой линии было выделено семь колоний для геномной и фенотипической характеристики из популяции, которая поддерживалась в течение двух пассажей на уровне или выше клинической пограничной точки, определенной EUCAST для конкретного антибиотика. Клиническая пограничная точка представляет собой концентрацию лекарственного средства, которая используется в качестве порогового значения для классификации патогенов как чувствительных или устойчивых к конкретному лекарственному средству (Turnidge and Paterson, 2007).
Лабораторная адаптивная эволюция в инкрементах лекарств и контроле среды
E. coli K12 (MG1655) развивалась не только в градиентах лекарств, но также и в отношении ежедневного относительного увеличения концентрации лекарств. Использовались три разных подхода (точные концентрации лекарств на каждый день для разных подходов к приращению и лекарства приведены в дополнительной таблице 3). Родословные в настройке «Increment 100» подвергались воздействию 100%-ного увеличения концентрации препарата. В соответствии с этим режимом концентрация препарата удваивалась каждый день, оказывая постоянное сильное давление отбора на родословные. Предполагалось, что клиническая точка останова будет достигнута через 7 дней эксперимента с ALE. Линии «Increment 50» также подвергались довольно высокой скорости изменения окружающей среды из-за увеличения концентрации препарата на 50% каждый день, достигая клинической пограничной точки на 9й день эксперимента. Концентрация лекарственного средства была увеличена на 25% для линий «Increment 25», что позволило провести мягкий отбор и вдвое увеличить время адаптации к концентрации клинической контрольной точки по сравнению с линиями «Increment 100». Восемь родословных развивались параллельно в каждой обстановке до AMK, PIP и TET. Эксперимент был спланирован так, чтобы все экспериментальные установки достигли МИК, как определено EUCAST, на 4-й день. В качестве контроля адаптации к среде восемь линий дикого типа были выведены на среду без антибиотиков. Смесь антибиотиков MHBII готовили в пробирках Falcon на каждый день, лекарство и экспериментальную установку в начале эксперимента и хранили при -20°C. Среду, содержащую препарат, размораживали в день использования и предварительно нагревали до 37°С. Линии выращивали в 1 мл MHBII и антибиотика в 96-луночная чашка на 22 ч при 37°C и ~900 об/мин. Каждые 22 часа переносили по 50 мкл клеток. Остальные клетки смешивали до конечной концентрации 20% глицерина и хранили при -80°С. Адаптивная эволюция была остановлена через 14 дней, когда линии «Increment 25» прошли клиническую точку останова. Во время эксперимента ~3% из 1152 отрицательных контролей показали рост. Клетки высевали штрихами на чашки LB и визуальным исследованием идентифицировали как E. coli . Все колонии выглядели одинаково, что свидетельствует об отсутствии загрязнения. После завершения эксперимента по адаптивной эволюции линии, которые были адаптированы к клинической контрольной точке, высевали штрихами на агар LB, и семь изолированных колоний использовали для дальнейшего анализа. Если родословные вымерли до того, как они достигли клинической контрольной точки, выбирали последнюю возможную временную точку.
IC
85 Определение Изолированные колонии использовали для инокуляции 96-луночного титрационного микропланшета, содержащего 150 мкл MHBII. Около 10 5 клеток ночной культуры переносили с помощью репликатора с 96 иглами в 10 разведений 2-кратного градиента лекарственного средства, охватывающего от 0,5 до 256 мг/л АМК и от 0,25 до 128 мг/л PIP или ТЕТ соответственно. . Для каждой линии одну изолированную колонию тестировали в двух технических повторностях против всех трех антибиотиков. Восемь инокулированных лунок, содержащих MHBII, служили положительным контролем, тогда как восемь лунок, заполненных только MHBII, служили отрицательным контролем. Планшеты инкубировали при 37°С и 900 об/мин в течение 18 часов, а затем ОП 600 измеряли с помощью считывателя абсорбции ELx808 (BioTek). Данные были дополнительно проанализированы с помощью R (Team R Core, 2014). Средние значения OD 600 отрицательных контролей вычитали из всех оставшихся значений OD 600 . Процентное ингибирование рассчитывали путем вычитания значений ОП 600 , деленных на среднее значение ОП 600 положительных контролей, из 1. положительный контроль)-OD600 (отрицательный контроль) (1)
Кривая доза-реакция была подогнана к значениям с использованием логистической модели из пакета drc (Ritz and Streibig, 2005), с x для молярной концентрации лекарственного средства и значениями по умолчанию для других переменных, где b описывает крутизну кривой, c и d нижняя и верхняя асимптоты и e эффективная доза (Munck et al., 2014; Ritz et al., 2015):
f(x(b, c, d , e))= c+ (d−c)1+exp(b*(log(x)−log(e)) (2)
Кривые доза-реакция были построены с помощью пакета ggplot (Wickham, 2009). Концентрацию лекарственного средства, вызывающую 85% ингибирование роста (IC 85 ), рассчитывали с помощью инвертированной функции, нормализовали к дикому типу и наносили на график, сгруппированный по лекарственному средству, используя ggplot (Wickham, 2009). Для некоторых экспериментальных установок и препаратов наблюдалось непараметрическое распределение параллельных линий, поэтому в R был применен непараметрический однофакторный дисперсионный анализ Крускала-Уоллиса для выявления значительных ( P <0,05) различий между четырьмя экспериментальными установками. адаптированы к каждому препарату.
Измерение скорости роста
В 96-луночный титрационный микропланшет, содержащий 200 мкл MHBII на лунку, инокулировали клетки в фазе экспоненциального роста с использованием 96-контактного репликатора. Все семь изолированных колоний каждой линии были включены в измерение роста. OD 600 измеряли с помощью ELx808 Absorbance Reader (BioTek) каждые 5 мин в течение 10 ч при 37°C и 650 об/мин. Данные были проанализированы с помощью R (Team R Core, 2014). Скорость роста рассчитывали на основе самой крутой части кривой роста при экспоненциальном росте. Время удвоения было нормализовано к дикому типу и нанесено на график с группировкой по наркотикам с помощью пакета ggplot (Wickham, 2009). ). Чтобы проверить, были ли наблюдаемые различия в скорости роста значительными ( P < 0,05), в R был применен непараметрический однофакторный дисперсионный анализ Краскела-Уоллиса. 92 линии, которые также использовали для измерения IC 85 и скорости роста, выращивали в LB, и ДНК экстрагировали с помощью набора A&A Genomic Mini (A&A Biotechnology). ДНК была отправлена в компанию Macrogen, которая подготовила геномные библиотеки с помощью набора TruSeq DNA Nano (550 п.н.) от Illumina и секвенировала их с помощью парных концов Illumina MiSeq 300 п.н.
Идентификация однонуклеотидных полиморфизмов и малых последовательностей INDEL
Прочтения были сопоставлены с эталонным геномом E. coli K12 U00096 с помощью CLC Genomics Workbench (Escherichia coli Str. K-12 Substr, 2014). В среднем каждая база была покрыта не менее 37,5 раз (дополнительная таблица 4). Сайты SNP и INDEL определяли с помощью CLC Genomics Workbench. В анализ были включены только локусы с оценкой phred 30 в вариабельном положении, а также в трех соседних основаниях, встречающихся не менее чем с частотой 80%. Однонуклеотидные полиморфизмы (SNP), обнаруженные во всех линиях, включая адаптированные к среде дикие типы, считались мутациями, произошедшими до начала эксперимента, и поэтому исключались. Если две линии имели два идентичных SNP, их считали перекрестно зараженными, и одну из них исключали из дальнейшего анализа.
Обнаружение больших удалений
Большие удаления были идентифицированы с помощью рабочего процесса в рабочей среде CLC. Прочтения всех геномов были собраны de novo и использованы в качестве эталонных геномов для картирования прочтений одного из штаммов дикого типа, адаптированных к среде. Были собраны считывания из штамма, адаптированного к среде, который не картировался в геноме de novo , собранном в результате эволюции антибиотика, а также de novo , собранном в контиги. Контиги размером более 1 т.п.н. с покрытием > 30 считались большими делециями.
Обнаружение больших вставок ДНК с помощью вставочных элементов
Мы загрузили как полный геном E. coli K12 MG1655 с номером доступа U00096, так и соответствующие ORF из нуклеотидного архива NCBI. Затем ORF были сгруппированы вместе с помощью cd-hit (Li and Godzik, 2006), что позволило ORF с идентичностью и покрытием не менее 90% находиться в одном кластере. Чтения секвенированных штаммов были отфильтрованы с использованием пакета FASTX-Toolkit и минимального качества 30. Чтения, отфильтрованные по качеству, были сопоставлены с кластеризованными ORF, установив word_size на 10 и e-значение на 100, что позволило точное картирование коротких последовательностей. . Чтения, сопоставленные как минимум с двумя ORF разных кластеров с минимум 30 и максимум 70% покрытия каждого кластера, также считались потенциальными большими вставками. Эти чтения были подвергнуты анализу всего генома, чтобы подтвердить, что два гена не соседствуют в геноме. Чтения, покрывающие эталонный геном более чем на 9Ожидалось, что 0% будут содержать два соседних гена, поэтому они были исключены из анализа. Далее было изучено, перекрываются ли оставшиеся совпадения с элементами вставки. Прочтения, выровненные с одной ORF, аннотированной как элемент вставки, были включены для дальнейшего анализа. Только инсерционные чтения, которые были обнаружены более чем в 10 чтениях, считались большими вставками ДНК.
Идентификация дупликаций генов
Дупликации генов идентифицировали с помощью CLC Genomics Workbench с использованием инструмента анализа покрытия, вызывая регионы с не менее чем 700 п.н. значительного ( P < 0,001) увеличенное покрытие. Идентифицированные области были сопоставлены с эталонным геномом U00096 в R, и гены, которые перекрывались не менее чем на 95% с областью высокого покрытия, были идентифицированы как дуплицированные гены.
Результаты
Четыре различных стратегии адаптивной лабораторной эволюции
Чтобы сравнить влияние методологии ALE на окончательные фено- и генотипы, мы эволюционировали E. coli к трем различным антибиотикам с использованием четырех различных стратегий ALE. Антибиотики, выбранные для этого эксперимента, представляют три основные группы антибиотиков. Два препарата, амикацин (АМК) и тетрациклин (ТЕТ), воздействуют на рибосомы, при этом первый действует бактерицидно, а второй — бактериостатически. Третий препарат, пиперациллин (PIP), представляет собой бактерицидный препарат, воздействующий на биосинтез клеточной стенки. Четыре различных режима селекции можно разделить на две категории: (1) градиентный подход, при котором популяция, которая переносит самые высокие концентрации лекарственного средства, переходит на новый градиент лекарственного средства каждые 22 часа; и (2) инкрементальный подход, при котором развивающаяся популяция передается каждые 22 часа на новую концентрацию препарата, увеличивающуюся на фиксированные приращения (рис. 1).
Рисунок 1. Экспериментальный план исследования: адаптация E. coli K12 к трем антибиотикам в соответствии с четырьмя протоколами адаптации с различной силой отбора . Четыре системы выбора называются «градиент», «шаг 100%», «шаг 50%» и «шаг 25%». При градиентном подходе клетки инокулировали в 2-кратном 10-ступенчатом градиенте лекарственного средства, а клетки, растущие в самой высокой концентрации, использовали для инокуляции свежего градиента. В инкрементальных подходах концентрация препарата повышалась при каждом переносе либо на 100, либо на 50, либо на 25%. Минимальная ингибирующая концентрация (МИК) дикого типа была достигнута на четвертый день эксперимента для всех линий прироста.
20-кратное разведение популяций, эволюционировавших в градиенте, было выбрано на основе модели Wahl et al. (2002), чтобы допустить высокую изменчивость в перенесенной популяции и основывать фиксацию мутаций на оптимальной адаптации, а не на ограничивающих узких местах. Согласно Валу и соавт. (2002) 10-кратное разведение популяции было бы оптимальным, но предыдущие эксперименты в лаборатории показали небольшое увеличение концентрации ингибирующего препарата для АМК и ТЕТ при использовании 10-кратного разведения. Чтобы избежать эффекта инокулята, который представляет собой значительное увеличение ингибирующей концентрации, вызванное большим количеством организмов в инокуляте (Брук, 1989), мы выбрали 20-кратное разведение (дополнительная фигура 2).
После 14 дней адаптивной эволюции в условиях градиента популяции, подвергшиеся воздействию АМК, переносили в среднем 512 мг/л препарата, что соответствует 170-кратному увеличению по сравнению с диким типом, адаптированным к среде ( P = 2,89654E −27 , тест Стьюдента t ) (рис. 2А). Эволюционировавшие линии PIP росли при концентрации препарата 192 мг/л на 14-й день эксперимента ALE, что соответствует 80-кратному увеличению по сравнению с диким типом, адаптированным к среде (9).0087 P = 0,00109527, t — тест Стьюдента). Однако в ходе эксперимента наблюдались большие колебания сопротивления для эволюционных линий PIP (рис. 2B). Это изменение можно объяснить эффектом инокулята, который чаще наблюдается для бета-лактамных антибиотиков (Eng et al., 1984, 1985; Brook, 1989). Линии, эволюционировавшие в ТЕТ, не достигли такой же устойчивости к лекарственным средствам, как линии, эволюционировавшие в АМК или PIP, но все же росли при 15 мг/л ТЕТ, превышая адаптированную к среде ингибирующую концентрацию дикого типа (IC) в 15 раз (9). 0087 P = 2,04072E −06 , тест Стьюдента t ) (рис. 2С). Эти значения согласуются с предыдущими выводами, когда значения IC 90 (концентрация лекарственного средства, при которой рост 90% популяции подавляется) изолированных колоний были определены после 14 дней адаптивной эволюции в градиентной системе с использованием того же штамма. и наркотики (Munck et al., 2014). В настоящем исследовании только линии, развившиеся в PIP, кажутся более устойчивыми, что связано с эффектом инокулята, вызванным большим объемом пассажей.
Рисунок 2. Обзор потенциала адаптации с течением времени для каждого препарата: (A) амикацин, (B) пиперациллин и (C) тетрациклин. Линии, адаптированные с помощью градиентного метода, следуют максимальному давлению отбора, поэтому можно наблюдать, что линии из подходов приращения (100, 50 и 25) вымирают, когда они превышают максимум естественного отбора, определенный градиентным подходом, обозначенным звездочками в фигура. Реплики представлены в разных стилях линий. Клиническая пограничная точка обозначена пунктирной линией черного цвета. Тепловые карты иллюстрируют исчезновение родословных с течением времени.
В подходе с инкрементом были протестированы три различных скорости изменения окружающей среды (рис. 1), для которых селективное давление (например, концентрация антибиотика) увеличивалась на 100, 50 или 25% каждый день. Подобно градиентному подходу, установка 100% приращения применяет высокое давление отбора с риском превышения адаптивного потенциала бактерии, что приводит к исчезновению клонов (рис. 1).
Исчезновение линий приращения после превышения максимума адаптации, определяемого линиями градиента
Мы предположили, что инкрементно-эволюционировавшие линии вымрут, когда концентрации наркотиков, которым они подверглись, превысят уровень адаптации градиентно-эволюционировавших линий, поскольку к градиентно-эволюционировавшим линиям было применено максимальное давление отбора. Наши наблюдения во время эксперимента ALE подтверждают эту гипотезу (рис. 2A–C). Всякий раз, когда концентрация антибиотиков в препаратах в экспериментах по эволюции приращения превышала максимальную концентрацию по крайней мере одной из линий градиентной эволюции, некоторые из параллельных линий приращения вымирали. Например, оставшиеся линии со 100% приростом, адаптированные к АМК, вымерли после того, как они выросли на 32 мг/л на 8-й день эксперимента, но одна из адаптированных к градиенту линий выросла только на 16 мг/л. Все линии прироста вымерли при воздействии концентрации препарата выше концентрации препарата, к которой были адаптированы все линии градиента (рис. 2А-С). Например, все оставшиеся линии прироста 50% вымерли, когда они прошли концентрацию PIP ~ 32 мг / л на 13-й день эксперимента по адаптивной эволюции, когда линии, адаптированные в градиентном подходе, росли только при концентрации PIP 8 мг / л. Поскольку воздействие антибиотиков на линии с приростом 100 и 50% превышало максимальный эволюционный потенциал, проявляемый соответствующими линиями с градиентным развитием, все линии с приростом 100 и 50% вымерли до конца эксперимента (рис. 2A-C). Примечательно, что только 4 из 24 линий, подвергшихся 100% приращению, достигли клинической пограничной точки перед исчезновением. Снижение плотности населения (OD 600 ) часто предшествовали исчезновению (дополнительный рисунок 3). Напротив, все 50 и 25% линий достигли клинической пограничной точки. Способность к адаптации, по-видимому, зависит от препарата. Различные адаптационные возможности отражаются в исчезновении линий приращения. Например, линии прироста 100 и 50% вымерли позже при адаптации к AMK по сравнению с TET (рис. 2A-C).
Одинаковые уровни сопротивления могут сопровождаться разными затратами на фитнес
Нам было интересно наблюдать, как различные скорости изменения окружающей среды влияют на конечные генотипы и фенотипы. По замыслу линии должны быть адаптированы к различным концентрациям антибиотиков в конце эксперимента, в зависимости от конкретного подхода к ALE. Соответственно, мы решили сравнить линии, когда они достигли клинической пограничной точки (определено EUCAST, 2016). Линии с градиентной эволюцией анализировали в тот момент времени, когда они достигли или превышали клиническую контрольную точку в течение 2 дней подряд. Инкрементно развившиеся линии анализировали в тот момент времени, когда линия достигала клинической пограничной точки. Поскольку большинство линий со 100% приростом не смогли достичь клинической пограничной точки, мы исключили эти линии из анализа.
По одной колонии получали из каждой линии в тот момент времени, когда популяция достигала клинической пограничной точки и определялась устойчивость к антибиотикам. Значения IC 85 были нормализованы к среднему значению IC 85 штаммов, адаптированных к среде (рис. 3А).
Рис. 3. Фенотипические изменения лекарственной устойчивости и скорости роста после адаптивного лабораторного эволюционного эксперимента. (A) Для каждого препарата показано кратное увеличение IC 85 по сравнению с диким типом (WT). Уровни развитой резистентности существенно не различаются для амикацина (AMK) и пиперациллина (PIP) в разных экспериментальных установках, которые обозначаются как G для градиентного подхода, 25 для 25% приращения и 50 для метода 50% приращения. Однако штаммы, адаптированные к тетрациклину (ТЕТ), демонстрируют значительные изменения лекарственной устойчивости. Клиническая пограничная точка, нормализованная по кратному увеличению адаптированного к среде дикого типа (WT) каждого лекарственного средства, отмечена светло-серой панелью. (B) Время удвоения по сравнению с диким типом показано для той же изолированной колонии, которая использовалась для определения IC 85 , а также для шести дополнительных колоний, окрашенных в серый цвет. Для всех препаратов можно наблюдать значительно меньшее время удвоения для линий с 25%-ным приростом по сравнению с линиями с градиентной эволюцией. * Р < 0,05, ** Р < 0,01, *** Р < 0,001, **** Р < 0,0001.
Для штаммов, адаптированных к AMK и TET, уровень устойчивости штаммов, эволюционировавших в градиенте, был выше клинической пограничной точки (рис. 3A). Напротив, только один из штаммов, адаптированных к PIP, был выше клинической пограничной точки. Высокие колебания уровня устойчивости наблюдались в линиях, адаптированных к PIP (рис. 2B), что позволяет предположить, что эффект инокулята, а не реальная адаптация, способствовал устойчивости популяции. Инокулят, использованный в этом исследовании, соответствовал примерно 10 8 КОЕ/мл и не указывало на эффект инокулята в предыдущих экспериментах (дополнительная фигура 2). Тем не менее, она превышает зарегистрированную концентрацию КОЕ/мл, вызывающую эффект инокулята для PIP (Bryson and Brogden, 2012).
Уровни резистентности штаммов с 25-процентным приростом показали нормальное распределение вокруг клинической пограничной точки для TET и PIP и были выше клинической пограничной точки для AMK, тогда как штаммы с 50-процентным приростом показали немного более низкую толерантность (рис. 3A). Однако при сравнении штаммов, адаптированных к градиенту и приращению, только штаммы с ТЕТ-эволюцией показали значительное ( P <0,05 (односторонний дисперсионный анализ Крускала-Уоллиса) разница в их уровнях устойчивости (рис. 3А).
Известно, что многие мутации, вызывающие резистентность, влияют на приспособленность, что часто можно определить по снижению скорости роста (Linkevicius et al., 2013). Поскольку режим отбора, по-видимому, влияет на приспособленность полученных линий (Lindsey et al., 2013), мы измерили скорость роста тех же изолированных колоний, которые использовались для определения IC 85 , и дополнительно шести изолированных колоний для каждой линии, в результате чего было получено 28 клонов для градиентных подходов и 56 клонов для экспериментов по приращению (рис. 3B). Адаптация к АМК, как правило, связана с более низкой скоростью роста по сравнению с другими препаратами.
Для всех трех препаратов штаммы с 25% приростом росли значительно (однофакторный дисперсионный анализ Краскела-Уоллиса P <0,05) быстрее, чем штаммы, адаптированные к градиенту (рис. 3B). Преимущество роста инкрементных линий может быть связано с большим количеством поколений, которым они подверглись, по сравнению с линиями градиентной эволюции, что дает более приспособленным мутантам больше возможностей вытеснить устойчивых мутантов с большими затратами на приспособление и накопить компенсаторные мутации, которые могут сбалансировать затраты на приспособление. мутаций, вызывающих резистентность. Однако время, в течение которого популяция эволюционировала, и время удвоения существенно не коррелируют (9).0087 R = -0,019, коэффициент корреляции Пирсона произведение-момент, P = 0,71). Соответственно, вполне вероятно, что более короткое время удвоения линий прироста связано с более низким давлением отбора в сторону лекарственной устойчивости, что приводит к усилению отбора на высокую скорость роста. Этот вывод согласуется с предыдущими исследованиями, в которых сообщалось, что линий E. coli , эволюционировавших в сторону рифампицина при резком увеличении дозы препарата, имеют значительно более низкую скорость роста по сравнению с линиями, эволюционировавшими в сторону более постепенного увеличения дозы препарата (Lindsey et al., 2013), и что Линии E. coli , адаптированные к 22 различным антибиотикам в условиях мягкой селекции, имеют повышенную скорость роста по сравнению с линиями, развившимися в условиях жесткой селекции (Oz et al. , 2014). Тем не менее, не было обнаружено корреляции между уровнем устойчивости и скоростью роста, что позволяет предположить, что мутация, придающая высокую устойчивость, не обязательно связана с высокой стоимостью приспособленности и , наоборот, (дополнительная фигура 4).
Различные режимы селекции не оказывают существенного влияния на побочную чувствительность и фенотип перекрестной устойчивости
Резистентные к антибиотикам бактерии часто проявляют перекрестную устойчивость к аналогичным препаратам (Szybalski and Bryson, 1952). Интересно, что также часто можно наблюдать повышенную чувствительность к другим антибиотикам (Szybalski and Bryson, 1952; Imamovic and Sommer, 2013; Lázár et al., 2014; Munck et al., 2014), явление, обычно называемое побочной чувствительностью. Чтобы проверить, влияет ли режим селекции на фенотип перекрестной устойчивости и побочной чувствительности, мы определили профили лекарственной устойчивости для каждого эволюционировавшего штамма по отношению к каждому из трех протестированных лекарств (рис. 4). Все линии, адаптированные к AMK, показали побочную чувствительность к PIP. Однако штаммы, адаптированные к градиенту и 25% приращению АМК, различались по своей побочной чувствительности к ТЕТ (9).0087 P <0,05, Крускала-Уоллиса в одну сторону) (рис. 4). Следует отметить, что конечная точка градиентно-адаптированных линий AMK показала чувствительность к ТЕТ в соответствии с предыдущими исследованиями (Munck et al., 2014), предполагая, что количество поколений, которое линия могла пройти до тестирования коллатералей чувствительность была важна. Все изоляты из линий градиента и приращения показали побочную чувствительность к AMK при эволюции в TET и перекрестную устойчивость между PIP и TET (рис. 4). Однако штаммы, адаптированные к PIP в градиентном подходе, были немного менее устойчивыми (9).0087 P <0,05, Крускала-Уоллиса в одну сторону) к TET, чем штаммы, адаптированные в подходах с 25%-ным приращением (рис. 4). В то время как Оз и др. (2014) подчеркивает различия в чувствительности к коллатералям в умеренно и сильно отобранных линиях, наши результаты согласуются с выводами Lázár et al. (2014), предполагая, что фенотипические сходства преобладают над различиями.
Рисунок 4. Фенотипы перекрестной устойчивости и побочной чувствительности штаммов с градиентом, 25 и 50% прироста . Для каждого отдельного штамма указано кратное увеличение (синий) или уменьшение (красный) IC 85 по сравнению с диким типом (WT). Сопутствующая чувствительность, а также перекрестная устойчивость аналогичны в экспериментальных установках. Сопутствующая чувствительность к тетрациклину (ТЕТ) штаммов, адаптированных к амикацину (АМК) при подходе с 25%-ным приращением, повышена ( P <0,05) по сравнению со штаммами, эволюционировавшими в градиент лекарственного средства. Перекрестная устойчивость штаммов, эволюционировавших к пиперациллину (PIP), протестированных против TET, повышена (9).0087 P <0,05) в адаптированных штаммах с 25% приростом по сравнению со штаммами, эволюционировавшими в градиенте.
В целом можно сделать вывод, что перекрестная устойчивость была очень похожей между градиентным и инкрементным подходами. Основное фенотипическое различие между линиями градиентной и инкрементной эволюции заключается в более медленной скорости роста линий градиентной эволюции.
Генотипы линий, адаптированных к разным селекционным воздействиям, перекрываются
Штаммы, используемые для IC 85 9Определение 0144 и измерения скорости роста были секвенированы, чтобы выявить лежащие в основе генетические изменения. Всего мы выявили 173 мутации в 92 секвенированных штаммах (дополнительная таблица 5). Крупные вставки и делеции составили 26,5% от общего числа мутаций (дополнительная фигура 5А). Эти более крупные генетические перестройки часто упускают из виду, но они могут играть важную роль в процессе генетической адаптации. Два из восьми параллельных штаммов, адаптированных с шагом 25% к AMK, имеют три одинаковых общих SNP, что предполагает потенциальное перекрестное заражение между линиями. Поэтому для последующего анализа использовали только один из штаммов.
В среднем мы выявили около двух мутаций в каждом штамме в разных экспериментах (дополнительная фигура 5B). Даже несмотря на то, что линии с 25%-ным приростом развивались в течение большего количества поколений, пока не достигли клинической пограничной точки, не было существенной разницы в количестве мутаций между экспериментальными установками ( P > 0,5, Студенты t — тест) и количестве мутаций. в секвенированных изолятах достоверно не коррелировало с количеством генераций ( R = 0,19, корреляция Пирсона, P = 0,097).
Придает ли мутация резистентность, компенсирует ли затраты на приспособление, связанные с другими мутациями, или автостоп с мутацией резистентности, трудно определить без повторного введения конкретных мутаций по отдельности или в комбинации с неэволюционировавшим диким типом. Однако, если ген мутирует более чем в одном независимом штамме, вполне вероятно, что мутация была отобрана для (Lieberman et al., 2011; Yang et al., 2011; Sandberg et al., 2014). Мы отфильтровали наш набор данных в соответствии с этим критерием и обнаружили, что 88,8% генов, мутировавших в штаммах с градиентной эволюцией, также были мутированы в штаммах приращения (рис. 5). За исключением мутаций в гамма-цепи АТФ-синтазы ( atpG ) и субъединицы 2 цитохромбо (3) убихинолоксидазы ( cyoA ), все мутированные гены градиентных штаммов в отфильтрованном наборе данных также оказались мутированными в штаммах с 25% приростом (рис. 5). Клоны, несущие одну из двух мутаций, имеют среднее время удвоения, которое в четыре раза выше, чем у дикого типа, и в два раза выше, чем в среднем у всех штаммов, адаптированных к АМК. Следовательно, вполне вероятно, что эти мутации связаны с высокой стоимостью приспособленности и, соответственно, не были зафиксированы в линиях с 25%-ным приростом.
Рисунок 5. Генетическая адаптация к антибиотикам амикацину (АМК), пиперациллину (ПИП) и тетрациклину (ТЕТ) . Цвет представляет количество штаммов в процентах, которые содержат мутацию, идентифицированную как минимум в двух независимых линиях. Почти все мутации, идентифицированные в градиентном подходе, также присутствуют, по крайней мере, в одном из приростных штаммов. Развитые линии PIP и TET демонстрируют сходные генотипы с мутациями, преимущественно влияющими на фенотип mar.
Интересно, что штаммы, адаптированные к приращению, несли не только большинство мутаций, обнаруженных в штаммах, адаптированных к градиенту, но также и многие мутации, которые были идентифицированы исключительно в штаммах, адаптированных к приращению (рис. 5). Такими мутантными генами являются, например, nuoB, nuoC и nuoH , субъединицы NADH-хиноноксидредуктаз, которые перемещают электроны от NADH к хинонам в дыхательной цепи, которые, как было установлено в предыдущих исследованиях, придают устойчивость к AMK (Kohanski et al. и др., 2007 г., Шурек и др., 2008 г., Гиргис и др., 2009 г.; Вонг и др., 2014). Кроме того, эти мутации были связаны с фенотипами побочной чувствительности аминогликозидов ко многим другим классам антибиотиков (Lázár et al., 2014), что позволяет предположить, что мутации в этих генах имеют отношение к фенотипу побочной чувствительности к ТЕТ, чего не наблюдалось. в линиях градиента, когда они достигли клинической точки разрыва, но только в штаммах, выделенных из конечной точки линий градиента.
Мутации в генах fusA, sbmA , а также в двух разных двухкомпонентных системах cpxRA и arcAB оказались доминирующими мутациями во всех штаммах, адаптированных к АМК (рис. 5). Все мутации ранее были связаны с устойчивостью к АМК (Laviña et al., 1986; Busse et al., 1992; Johanson and Hughes, 1994; Salomón and Farias, 1995; Macvanin and Hughes, 2005; Kohanski et al., 2008, 2010). ; Pena-Miller et al., 2013; Lázár et al., 2014; Munck et al., 2014). Мутации в факторе элонгации G, кодирующем ген 9Было показано, что 0087 fusA приводит к побочной чувствительности к бета-лактамным антибиотикам, как это наблюдалось в этом исследовании PIP (Macvanin and Hughes, 2005).
acrR оказался преимущественно мутировавшим геном в эволюционных линиях PIP независимо от экспериментальной установки (рис. 5). Мутации в acrR , а также в marR и rob , также идентифицированные как мутировавшие в инкрементных штаммах, адаптированных к PIP, приводят к фенотипу множественной устойчивости к антибиотикам (mar), который, как было описано, придает устойчивость к разнообразным препаратов, включая бета-лактамные антибиотики и тетрациклины. Это открытие объясняет перекрестную резистентность, наблюдаемую в ходе этого эксперимента у линий, эволюционировавших в TET и PIP (George and Levy, 19).83; Коэн и др., 1989; Ариза и др., 1995; Маниваннакул и Леви, 1996 г.; Этингер и др., 1998). Отсутствие мутаций в marR и rob в штаммах, адаптированных к градиенту, может объяснить разницу в перекрестной устойчивости к ТЕТ по сравнению с адаптированными штаммами с 25% приростом. Однако можно предположить, что эти мутации также могут возникать в градиентной системе, если можно избежать эффекта инокулята, поскольку они наблюдались ранее в эксперименте с использованием градиентного подхода (Munck et al., 2014). Другая часто наблюдаемая мутация в штаммах, адаптированных к PIP, влияет на мишень лекарственного средства, пептидогликансинтазу 9.0087 ftsI (пенициллин-связывающий белок 3) (рис. 5) (Matic et al., 2003; Blázquez et al., 2006). Интересно, что мутации в cpxA были обнаружены исключительно в 25% и 50% приростных штаммах, адаптированных к PIP (рис. 5). Мутации в этом гене могут привести к двукратному повышению устойчивости к бета-лактамным антибиотикам (Srinivasan et al., 2012; Bernal, 2014). Поскольку 2-кратное увеличение лекарственной устойчивости является умеренно низким, это может объяснить, почему мутация была обнаружена только в линиях 25 и 50% прироста, которые подвергались воздействию низких концентраций антибиотиков, и почему она не была идентифицирована в линиях градиента или 100% прироста. .
Линии, адаптированные к TET, показали, как и PIP, мутации в генах, принадлежащих к фенотипу mar (рис. 5). В дополнение к мутациям, относящимся к фенотипу mar, были идентифицированы два других мутировавших гена, dksA и waaP , которые ранее лишь косвенно были связаны с чувствительностью к антибиотикам (Yethon et al., 1998; Yethon and Whitfield, 2001; Hansen et al. и др., 2008; Тамае и др., 2008; Лю и др., 2010). Интересно, что единственные дупликации генов, наблюдаемые в этом эксперименте, были обнаружены в трех разных линиях в штаммах с 50% приростом, адаптированных к ТЕТ. Два гена, yicS и yibT с неохарактеризованными генными продуктами были продублированы так же, как и phoU , чей делеционный мутант был более восприимчив к антибиотикам, что указывает на потенциальную роль в устойчивости к антибиотикам (Li and Zhang, 2007).
Значительная разница в устойчивости между линиями градиентной и инкрементной эволюции может быть объяснена обилием мутаций в marR . Почти все штаммы, адаптированные к градиентному подходу, несут мутацию в marR , тогда как это генетическое изменение наблюдалось только в нескольких линиях, эволюционировавших в режиме приращения (рис. 5). Линии, содержащие мутантный marR , в целом были на 15% более устойчивы к ТЕТ, чем в среднем все адаптированные к ТЕТ линии. Однако у них также было увеличено время удвоения примерно на 27% по сравнению со средним значением. Мутанты Δ marR ранее были связаны с нарушением приспособленности (Marcusson et al., 2009), предполагая, что мутации в marR с большей вероятностью будут доминировать в популяции при сильном отборе.
Клоны, адаптированные в рамках градиентного подхода, по-видимому, имеют меньше мутаций в отфильтрованном наборе данных по сравнению с инкрементными линиями. Однако они часто несут мутации, которые были обнаружены только один раз за весь эксперимент (дополнительная таблица 5), поэтому адаптированные к градиенту линии демонстрируют большее разнообразие уникальных мутаций. Чтобы количественно оценить сходство и различие между генотипами в разных экспериментальных установках, мы провели попарное сравнение между всеми штаммами, используя нефильтрованный набор данных. Перекрытие мутантных генов между парами рассчитывали в процентах от общего числа мутантных генов, обнаруженных в двух штаммах. Мы решили анализировать сходство на уровне генов, а не на уровне SNP или семейства генов, поскольку это было предложено в качестве подходящей меры для обнаружения параллельной или конвергентной эволюции (Achaz et al., 2014). Генетическое сходство в репликах с градиентной эволюцией составляло в среднем около 30–50% (рис. 6). Штаммы, адаптированные при 25- и 50-процентном приращении, были примерно на 45 и 30% сходны друг с другом при адаптации к AMK и PIP и только примерно на 20 и 10% при эволюции к TET (рис. 6). Интересно, что генетическое сходство штаммов, выведенных с использованием разных режимов селекции, было сравнимо со сходством внутри реплик из одного и того же режима селекции (рис. 6). Сходство штаммов с градиентом и 25%-ным приростом было максимальным примерно на 3% ниже и на 12% выше внутреннего сходства группы либо с градиентом, либо с приростом 25 штаммов (рис. 6). Этот результат подчеркивает, что генетическое сходство между различными режимами селекции сходно с генетическим сходством, наблюдаемым между параллельными линиями, которые развивались в идентичных условиях.
Рисунок 6. Сходство родословных эволюционировало до разной силы отбора . Сходство между различными штаммами рассчитывали в попарных сравнениях путем анализа процента генов, которые были мутированы в обоих штаммах, по сравнению с общим количеством мутаций, обнаруженных в двух штаммах. Среднее попарное сходство отображается для разных групп штаммов: либо в пределах одной экспериментальной установки, такой как градиент (G) (красный), либо подходы приращения (25 и 50) (синий и серый), либо между этими экспериментальными установками (полосы). , сравнивая сходство линий градиента и приращения или подходов приращения друг с другом.
Обсуждение
В этом исследовании мы проанализировали влияние режима селекции в различных методологиях ALE на результирующие фенотипы и генотипы. Как и ожидалось, скорость изменения окружающей среды является решающим параметром для вымирания популяций. Изменения окружающей среды, которые превышают адаптационные возможности организма или дают слишком мало времени для адаптации, обычно приводят к вымиранию. Уменьшение OD часто предшествовало вымиранию линий, что указывает на то, что средняя приспособленность популяции была снижена до вымирания (Lynch and Lande, 19).93). Сокращение размера популяции также уменьшило количество клеток, перенесенных в следующую концентрацию препарата, что уменьшило генетическую изменчивость (Frankham, 2005; Bell and Collins, 2008). В этих условиях средняя приспособленность популяции ухудшается и может быть улучшена только за счет снижения скорости изменения окружающей среды (Lynch and Lande, 1993). В нашем случае постоянно высокая скорость изменения окружающей среды в линиях со 100- и 50-процентным приростом привела к исчезновению эволюционирующей популяции. Мы наблюдали эволюционный предел адаптации, который определялся линиями градиентной эволюции. Если скорость изменения окружающей среды линий с 50- и 100-процентным приростом превышала этот максимум, штаммы вымирали. Эти результаты могут быть реализованы при разработке эксперимента с ООВ в промышленных целях, где обычно следует предотвращать вымирание родословных. Следовательно, можно предложить либо использовать более мягкую скорость изменения окружающей среды, либо большее количество повторений, чтобы компенсировать исчезновение линии.
Мы ожидали, что линии градиентной эволюции укажут на эволюционную способность E. coli адаптироваться к определенному лекарству, поскольку мы постоянно применяли максимальное давление отбора в настройке градиента. Однако в случае пиперациллина мы наблюдали большие колебания толерантности к препарату во всех параллельно развивающихся популяциях в течение 14 дней эксперимента по адаптивной эволюции. Инокулят варьировался в зависимости от причины эксперимента и иногда превышал инокулянт, который использовался для начального теста, что не предполагало эффекта инокулята ни для одного из трех препаратов. Кроме того, известно, что бета-лактамные антибиотики более склонны вызывать эффект инокулята (Eng et al., 19).85; Brook, 1989), из чего мы заключаем, что эффект инокулята является наиболее вероятным объяснением колебаний. Тем не менее, другие сценарии, такие как (1) клональная интерференция (de Visser and Rozen, 2006), когда несколько мутаций, придающих устойчивость, толерантность или выгодный рост, конкурируют друг с другом, (2) подрывной частотно-зависимый отбор (Levin et al., 1988) , где в популяции фиксируются только распространенные мутации или (3) фенотипическая толерантность, которая временно позволяет бактериям выживать в концентрациях антибиотиков, не вызывая устойчивости (Brauner et al. , 2016), также может быть причиной колебаний.
После достижения определенного уровня фенотипа различные пути отбора приводят к сходным фенотипам и генотипам. Подобные результаты изменений в силе давления отбора можно объяснить концепцией эволюционных ограничений (Losos, 2011). Из-за ограниченного числа доступных изменений для адаптации к определенному давлению отбора эволюция смещается в сторону этих мутаций, что приводит к сходным изменениям в организмах, находящихся в сопоставимых условиях (Losos, 2011).
Мы обнаружили, что разная сила отбора, применяемая за счет ежедневного увеличения концентрации лекарственного средства на 25% или градиента лекарственного средства, следует схожим эволюционным траекториям, приводя к сходным фенотипам и генотипам. Модели перекрестной резистентности и побочной чувствительности для обоих подходов кажутся очень похожими. Это согласуется с предыдущими выводами, когда линии, эволюционировавшие до субингибирующих концентраций лекарств, сравнивали с теми, которые адаптировались в условиях сильного отбора (Lázár et al. , 2014). Тем не менее, также сообщалось о противоположных результатах, показывающих различия в умеренно и сильно отобранных линиях (Oz et al., 2014). Однако различия в перекрестной резистентности и побочной чувствительности, наблюдаемые Oz et al. были слабо связаны с конечным уровнем устойчивости штамма к адаптированному лекарственному средству, что могло объяснить эти фенотипические различия (Oz et al., 2014). Чтобы ограничить эти смешанные факторы, мы стремились исследовать линии, поскольку они достигли сходного уровня устойчивости к адаптированному лекарству. Фенотипические различия, наблюдаемые Oz et al. объяснялись генотипическими вариациями. Сообщалось о большем количестве мутаций в тщательно отобранных линиях, включая большее разнообразие мутаций, связанных с лекарственными мишенями. Эти результаты противоречат другим исследованиям, утверждающим, что чем сильнее давление отбора, тем меньше эволюционных траекторий открыты для удовлетворения требований адаптации, что приводит к меньшему количеству, но более значительным мутациям в сильно эволюционировавших линиях (Barrick and Lenski, 2013; Lindsey et al. , 2013). и может быть результатом различий в окончательно развившихся фенотипах штаммов по сравнению с настоящим исследованием. В нашем исследовании мы находим одинаковое количество мутаций в сильно и слабо адаптированных линиях. Однако при фильтрации наших геномных данных на предмет повторяющихся мутаций мы обнаруживаем меньшее количество мутаций в линиях с градиентной эволюцией по сравнению с линиями, адаптированными к инкременту, что позволяет предположить, что меньшее количество мутаций, придающих устойчивость, отбирается для сильного отбора. Несмотря на эту небольшую разницу, генетическое сходство между репликами в рамках одного подхода к селекции сравнимо со сходством между разными подходами к селекции. Эти результаты показывают, что эксперименты с ALE, проведенные с различными протоколами, действительно сопоставимы. Соответственно, нельзя потерять генетическую или фенотипическую информацию при использовании применимого подхода приращения с высокой пропускной способностью по сравнению с градиентными системами.
Тем не менее, в этом исследовании было обнаружено одно важное отличие, имеющее значение для экспериментов с ALE: линии, адаптированные с помощью метода 25% приращения, постоянно демонстрировали более высокую скорость роста по сравнению со скоростью роста изолятов из линий, эволюционировавших в условиях более сильного давления отбора. Преимущество роста штаммов, адаптированных с помощью метода 25% приращения, можно объяснить режимом мягкого отбора, который увеличивает давление отбора на приспособленность, а не на самые высокие уровни устойчивости, возможно, отбирая мутации, которые компенсируют затраты на приспособленность или в целом повышают приспособленность. Когда данные секвенирования были отфильтрованы, чтобы учесть только мутации, которые были обнаружены более чем в двух отдельных линиях, клоны, адаптированные с шагом 25%, несли почти все мутации, обнаруженные в градиентных линиях. Таким образом, эти мутации, вероятно, являются наиболее важными для фенотипа резистентности. Ряд дополнительных мутаций был идентифицирован исключительно в линиях приращения. Эти мутации могут придавать устойчивость при меньших затратах на приспособленность или компенсировать недостатки приспособленности, связанные с мутациями, придающими устойчивость. В зависимости от цели эксперимента ALE следует обратить внимание на влияние метода на скорость роста. Особенно, если эксперимент ALE проводится для улучшения биотехнологического производственного штамма, скорость роста эволюционировавших штаммов может быть важным фактором для отбора (O’Brien et al., 2016).
В случае устойчивости к антибиотикам эксперименты с ALE могут быть полезны для изучения эволюционного потенциала вида в отношении развития устойчивости к антибиотикам. Сообщалось, что некоторые из мутантных генов, которые были идентифицированы в этом и предыдущих исследованиях, такие как локусы marR, rob или acrAB , также несут мутации в клинических изолятах (Oethinger et al., 1998; Sáenz et al. , 2004; Buffet-Bataillon et al. , 2012). Однако другие мутации, выявленные в экспериментах с ALE, не встречаются в естественных условиях. Тем не менее, эксперименты с ALE могут обеспечить лучшее понимание генетической и фенотипической гибкости организма и его адаптационного потенциала в ответ на сложные условия окружающей среды.
Нам было интересно посмотреть, имитируют ли как градиентный, так и инкрементный подход естественную эволюцию сопротивления. Чтобы исследовать общность идентифицированных SNP, мы изучили все секвенированные геномов E. coli на наличие неконсервативных SNP в кодирующих областях. Более 50% мутантных генов как в подходе с приращением 25%, так и в градиентном подходе, и все гены, общие для этих подходов, перекрываются с мутантными генами в базе данных (дополнительная фигура 6), что указывает на то, что обе методологии моделируют естественную эволюцию устойчивости в аналогичной степени. Очевидно, еще предстоит выяснить, действительно ли эти SNP обеспечивают устойчивость или другие преимущества в среде хозяина. Тем не менее, появление таких мутаций как в естественных изолятах, так и в лабораторных популяциях предполагает их биологическую важность.
Наши результаты показывают, что ключевые адаптации к AMK, PIP и TET у E. coli не зависят от режима селекции и что мутации, которые устойчиво возникают независимо от режима селекции или методологии ALE, также с большей вероятностью будут отобраны в клинику, чем мутации, которые отбираются только в очень специфических условиях отбора.
Вклад авторов
MS, CM и LJ спланировали проект и разработали эксперименты. LJ провел эксперименты и провел анализ данных с помощью CM. ME внес свой вклад, определив большие последовательности вставок и написав раздел «Материалы и методы» о своем анализе. LJ написал рукопись, которая была критически рассмотрена CM и MS.
Финансирование
Эта работа была поддержана исследовательской и инновационной программой Европейского Союза Horizon 2020 в соответствии с соглашением о гранте Марии Склодовской-Кюри № 642738, MetaRNA, а также программой Horizon 2020 Европейского Союза (ERC-2014-StG) в соответствии с соглашением о гранте 638902. , ЛимитMDR. Кроме того, финансирование было получено от Датского совета по свободным исследованиям и от Фонда Ново Нордиск через Центр биоустойчивости Фонда Ново Нордиск.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы хотели бы поблагодарить A. Porse за полезные обсуждения и за предоставленный нам сценарий R для точного определения скорости роста.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/article/10.3389/fmicb.2017.00816/full#supplementary-material
Ссылки
Ахаз Г., Родригес-Вердуго А., Гаут Б. С. и Тенайлон О. (2014). Воспроизводимость адаптации в свете экспериментальной эволюции с полногеномным секвенированием. Доп. Эксп. Мед. биол. 781, 211–231. doi: 10.1007/978-94-007-7347-9_11
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Андерсон В., Янсен М. Г. и Уикс К. А. (1950). Парааминосалициловая кислота со стрептомицином при туберкулезе. Кан. Мед. доц. J. 62, 231–235.
Реферат PubMed | Google Scholar
Ариза Р. Р., Ли З., Рингстад Н. и Демпл Б. (1995). Активация множественной устойчивости к антибиотикам и связывание стресс-индуцируемых промоторов белком Escherichia coli Rob. J. Бактериол. 177, 1655–1661.
Реферат PubMed | Google Scholar
Баррик, Дж. Э., и Ленски, Р. Э. (2013). Динамика генома в ходе экспериментальной эволюции. Нац. Преподобный Жене. 14, 827–839. doi: 10.1038/nrg3564
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Белл Г. и Коллинз С. (2008). Адаптация, вымирание и глобальные изменения. Эволюция. заявл. 1, 3–16. doi: 10.1111/j.1752-4571.2007.00011.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бернал, М. М. (2014). Путь Cpx вызывает изменения в структуре пептидогликана, обороте и рециклинге . Магистерская диссертация, Университет Альберты.
Бласкес, Дж., Гомес-Гомес, Дж. М., Оливер, А., Хуан, К., Капур, В., и Мартин, С. (2006). Ингибирование PBP3 вызывает адаптивные реакции у Pseudomonas aeruginosa . Мол. микробиол. 62, 84–99. doi: 10.1111/j.1365-2958.2006.05366.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Блюм П., Рудраппа Д., Сингх Р., Маккарти С. и Павлик Б. (2016). «Экспериментальная микробная эволюция экстремофилов», в Biotechnology of Extremophiles: Grand Challenges in Biology and Biotechnology, Vol. 1 , изд. PH Rampelotto (Чам; Гейдельберг; Нью-Йорк, штат Нью-Йорк; Лондон; Дордрехт: Springer International Publishing), 619–636.
Google Scholar
Браунер А., Фридман О., Гефен О. и Балабан Н. К. (2016). Различают резистентность, толерантность и устойчивость к антибиотикотерапии. Нац. Преподобный Микробиолог. 14, 320–330. doi: 10.1038/nrmicro.2016.34
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Брук, И. (1989). Эффект инокулята. Преподобный Заразить. Дис. 11, 361–368.
Реферат PubMed | Google Scholar
Брайсон Х.М. и Брогден Р.Н. (2012). Пиперациллин/тазобактам. Наркотики 47, 506–535. doi: 10.2165/00003495-1930-00008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Баффе-Батайон С., Ле Жен А., Ле Галл-Давид С., Боннаур-Малле М. и Жоливе-Гужон А. (2012). Молекулярные механизмы высших мик антибиотиков и четвертичных аммониевых соединений для Escherichia coli , выделенная из бактериемии. J. Антимикроб. Чемотер. 67, 2837–2842. doi: 10.1093/jac/dks321
Полный текст CrossRef | Google Scholar
Busse, HJ, Wöstmann, C., and Bakker, EP (1992). Бактерицидное действие стрептомицина: пермеабилизация мембран, вызванная внедрением неправильно транслируемых белков в цитоплазматическую мембрану Escherichia coli и последующее клеточное закрепление антибиотика внутри клеток за счет деградации этих белков. J. Gen. Microbiol. 138, 551–561. doi: 10.1099/00221287-138-3-551
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кэрролл С. П., Согаард Йоргенсен П., Киннисон М. Т., Бергстром С. Т., Денисон Р. Ф., Глюкман П. и др. (2014). Применение эволюционной биологии для решения глобальных проблем. Наука 346:1245993. doi: 10.1126/science.1245993
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чарльзворт, Б. (2009 г.). Эффективный размер популяции и закономерности молекулярной эволюции и изменчивости. Нац. Преподобный Жене. 10, 195–205. doi: 10.1038/nrg2526
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Коэн С.П., Макмерри Л.М., Хупер Д.К., Вольфсон Дж.С. и Леви С.Б. (1989). Перекрестная устойчивость к фторхинолонам у штамма с множественной антибиотикорезистентностью (март) Escherichia coli , отобранного с помощью тетрациклина или хлорамфеникола: снижение аккумуляции препарата, связанное с мембранными изменениями, в дополнение к снижению OmpF. Антимикроб. Агенты Чемотер. 33, 1318–1325. doi: 10.1128/AAC.33.8.1318
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
де Виссер, Дж. А., и Розен, Д. Е. (2006). Клональная интерференция и периодический отбор новых полезных мутаций в Escherichia coli . Генетика 172, 2093–2100. doi: 10.1534/genetics.105.052373
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Давале А. и Рат А. (2014). Устойчивость к антибиотикам: угроза и вызов обществу. Энн. заявл. Бионауч. 1, Р1–Р6.
Google Scholar
Дональд, П. Р., и ван Хелден, П. Д. (2009). Глобальное бремя туберкулеза — борьба с лекарственной устойчивостью в трудные времена. Н. англ. Дж. Мед. 360, 2393–2395. doi: 10.1056/NEJMp0
6
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Элмквист Т., Фольке К., Нистрем М., Петерсон Г., Бенгтссон Дж., Уокер Б. и др. (2003). Разнообразие реагирования, изменение экосистемы и устойчивость. Фронт. Экол. Окружающая среда. 1, 488–494. doi: 10.1890/1540-9295(2003)001[0488:RDECAR]2.0.CO;2
Полный текст CrossRef | Google Scholar
Eng, R.H., Cherubin, C., Smith, S.M., and Buccini, F. (1985). Влияние инокулята бета-лактамных антибиотиков на Enterobacteriaceae. Антимикроб. Агенты Чемотер. 28, 601–606. doi: 10.1128/AAC.28.5.601
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Энг, Р. Х., Смит, С. К., и Черубин, К. (1984). Инокуляционный эффект новых бета-лактамных антибиотиков на Pseudomonas aeruginosa . Антимикроб. Агенты Чемотер. 26, 42–47. doi: 10.1128/AAC.26.1.42
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Escherichia coli Str. К-12 Подстр (2014). ул. Кишечная палочка. К-12 Подул. MG1655, полный геном .
«ЕВКАСТ» (2016). ЕВКАСТ . Доступно на сайте: http://www.eucast.org/ (по состоянию на 25 июля).
Fine, PVA (2015). Экологические и эволюционные факторы географической изменчивости видового разнообразия. Год. Преподобный Экол. Эвол. Сист. 46, 369–392. doi: 10.1146/annurev-ecolsys-112414-054102
Полный текст CrossRef | Google Scholar
Франкхэм, Р. (2005). Генетика и вымирание. биол. Консерв. 126, 131–140. doi: 10.1016/j.biocon.2005.05.002
CrossRef Полный текст | Google Scholar
Джордж А.М. и Леви С.Б. (1983). Ген в основном пробеле котрансдукции карты сцепления Escherichia coli K-12, необходимый для выражения хромосомной устойчивости к тетрациклину и другим антибиотикам. J. Бактериол. 155, 541–548.
Реферат PubMed | Google Scholar
Георгиу, Г. П. (2012). Устойчивость вредителей к пестицидам . Риверсайд, Калифорния: Springer Science and Business Media.
Google Scholar
Гиргис Х.С., Хоттес А.К. и Тавазои С. (2009). Генетическая архитектура внутренней чувствительности к антибиотикам. PLoS ONE 4:e5629. doi: 10.1371/journal.pone.0005629
Полный текст CrossRef | Академия Google
Гован, Дж. Р. В., и Файф, Дж. А. М. (1978). Мукоид Pseudomonas aeruginosa и муковисцидоз: устойчивость слизистой формы к карбенициллину, флуклоксациллину и тобрамицину и выделение вариантов мукоида in vitro . J. Антимикроб. Чемотер. 4, 233–240. doi: 10.1093/jac/4.3.233
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хансен С., Льюис К. и Вулич М. (2008). Роль глобальных регуляторов и метаболизма нуклеотидов в устойчивости к антибиотикам у Кишечная палочка . Антимикроб. Агенты Чемотер. 52, 2718–2726. doi: 10.1128/AAC.00144-08
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хегренесс М., Шореш Н., Дамиан Д., Хартл Д. и Кишони Р. (2008). Ускоренная эволюция резистентности в среде с несколькими лекарственными средствами. Проц. Натл. акад. науч. США 105, 13977–13981. doi: 10.1073/pnas.0805965105
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хоффманн, А. А., и Парсонс, П. А. (1997). Экстремальные изменения окружающей среды и эволюция . Кембридж, Великобритания: Издательство Кембриджского университета.
Hu, B., Yang, Y.M., Beck, D.A.C., Wang, Q.B., Chen, W.J., Yang, J., et al. (2016). Комплексная молекулярная характеристика метилобактерий extorquens AM1, адаптированных к толерантности к 1-бутанолу. Биотехнология. Биотопливо 9:84. doi: 10.1186/s13068-016-0497-y
CrossRef Полный текст | Google Scholar
Имамович Л. и Соммер М.О.А. (2013). Использование сетей побочной чувствительности для разработки протоколов чередования лекарств, позволяющих избежать развития резистентности. Науч. Перевод Мед. 5:204ra132. doi: 10.1126/scitranslmed.3006609
PubMed Abstract | Полный текст CrossRef
Йохансон У. и Хьюз Д. (1994). Мутанты, устойчивые к фузидиевой кислоте, определяют три области фактора элонгации g Salmonella Typhimurium. Ген 143, 55–59.
Реферат PubMed | Google Scholar
Ким С. , Либерман Т. Д. и Кишони Р. (2014). Чередование антибиотиков сдерживает эволюционные пути к множественной лекарственной устойчивости. Проц. Натл. акад. Sci.U.S.A. 111, 14494–14499. doi: 10.1073/pnas.1409800111
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кохански, М. А., ДеПристо, М. А., и Коллинз, Дж. Дж. (2010). Сублетальное лечение антибиотиками приводит к множественной лекарственной устойчивости посредством радикального мутагенеза. Мол. Ячейка 37, 311–320. doi: 10.1016/j.molcel.2010.01.003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кохански, М. А., Дуайер, Д. Дж., Хайете, Б., Лоуренс, К. А., и Коллинз, Дж. Дж. (2007). Распространенный механизм гибели клеток, вызванный бактерицидными антибиотиками. Сотовый 130, 797–810. doi: 10.1016/j.cell.2007.06.049
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кохански, М. А., Дуайер, Д. Дж., Вежбовски, Дж. , Коттарел, Г., и Коллинз, Дж. Дж. (2008). Неправильная трансляция мембранных белков и активация двухкомпонентной системы запускают гибель клеток, опосредованную антибиотиками. Сотовый 135, 679–690. doi: 10.1016/j.cell.2008.09.038
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лавинья М., Пагсли А. П. и Морено Ф. (1986). Идентификация, картирование, клонирование и характеристика гена (sbmA), необходимого для действия микроцина B17 на Escherichia coli K12. J. Gen. Microbiol. 132, 1685–1693. doi: 10.1099/00221287-132-6-1685
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лазар В., Пал Г. С., Спон Р., Надь И., Хорват Б., Хртьян М., Буса-Фекете Р. и др. (2014). Бактериальная эволюция гиперчувствительности к антибиотикам. Мол. Сист. биол. 9, 700–700. doi: 10.1038/msb.2013.57
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Ли, Д. Х., и Палссон, Б. Ø. (2010). Адаптивная эволюция Escherichia coli K-12 MG1655 при росте на неприродном источнике углерода, L-1,2-пропандиоле. Заяв. Окружающая среда. микробиол. 76, 4158–4168. doi: 10.1128/AEM.00373-10
PubMed Abstract | Полный текст CrossRef
Lennen, R. (2016). «Адаптивная лабораторная эволюция толерантных к продукту хозяев для биохимического производства», в Ежегодная встреча и выставка SIMB 2016 (Новый Орлеан, Луизиана).
Google Scholar
Левин Б.Р., Антонович Дж. и Шарма Х. (1988). Частотно-зависимый отбор в бактериальных популяциях [и обсуждение]. Филос. Транс. Р. Соц. Б биол. науч. 319, 459–472. doi: 10.1098/rstb.1988.0059
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Левин Б.Р., Консепсьон-Асеведо Дж. и Удекву К.И. (2014). Персистентность: копацетическая и экономная гипотеза существования ненаследственной резистентности к антибиотикам. Курс. мнение микробиол. Антимикроб. 21, 18–21. doi: 10.1016/j.mib.2014.06.016
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Левин Б. Р., Перро В. и Уокер Н. (2000). Компенсаторные мутации, устойчивость к антибиотикам и популяционная генетика адаптивной эволюции бактерий. Генетика 154, 985–997.
Реферат PubMed | Google Scholar
Ли В. и Годзик А. (2006). Cd-Hit: быстрая программа для кластеризации и сравнения больших наборов последовательностей белков или нуклеотидов. Биоинформатика 22, 1658–1659. doi: 10.1093/bioinformatics/btl158
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ли Ю. и Чжан Ю. (2007). PhoU представляет собой переключатель персистенции, участвующий в формировании персистенции и толерантности к множеству антибиотиков и стрессов у Escherichia coli . Антимикроб. Агенты Чемотер. 51, 2092–2099. doi: 10.1128/AAC.00052-07
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Либерман Т. Д., Мишель Дж. Б., Айнгаран М., Поттер-Байно Г., Ру Д., Дэвис М. Р. и др. (2011). Параллельная бактериальная эволюция у нескольких пациентов позволяет выявить гены-кандидаты патогенности. Нац. Жене. 43, 1275–1280. doi: 10.1038/ng.997
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Линдси Х. А., Галли Дж., Тейлор С. и Керр Б. (2013). Эволюционное спасение от вымирания зависит от более низкой скорости изменения окружающей среды. Природа 494, 463–467. doi: 10.1038/nature11879
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Линкявичюс М., Сандегрен Л. и Андерссон Д. И. (2013). Механизмы и затраты на фитнес при резистентности к тигециклину у Кишечная палочка . J. Антимикроб. Чемотер. 68, 2809–2819. doi: 10.1093/jac/dkt263
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лю А., Тран Л., Бекет Э., Ли К., Чинн Л., Парк Э. и др. (2010). Профили чувствительности к антибиотикам, определенные с помощью коллекции нокаутных генов Escherichia coli : создание штрих-кода антибиотика. Антимикроб. Агенты Чемотер. 54, 1393–1403. doi: 10.1128/AAC.00906-09
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Лопес-Мори Л., Маргера С. и Белер Дж. (2008). Настройка экспрессии генов на изменение окружающей среды: от быстрых ответов до эволюционной адаптации. Нац. Преподобный Жене. 9, 583–593. doi: 10.1038/nrg2398
Полный текст CrossRef | Google Scholar
Лосос, Дж. Б. (2011). Конвергенция, адаптация и ограничение. Эволюция 65, 1827–1840 гг. doi: 10.1111/j.1558-5646.2011.01289.x
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Линч М. и Ланде Р. (1993). Эволюция и вымирание в ответ на изменение окружающей среды. Биотическое взаимодействие. Глоб. Смена 234–50.
Макванин, М., и Хьюз, Д. (2005). Гиперчувствительность устойчивых к фузидиевой кислоте мутантов сальмонелл к различным классам антибиотиков. FEMS Microbiol. лат. 247, 215–220. doi: 10. 1016/j.femsle.2005.05.007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Маневаннакул К. и Леви С. Б. (1996). Идентификация мутантов mar среди устойчивых к хинолонам клинических изолятов Escherichia coli . Антимикроб. Агенты Чемотер. 40, 1695–1698.
Реферат PubMed | Google Scholar
Маркуссон Л.Л., Фримодт-Мёллер Н. и Хьюз Д. (2009). Взаимодействие в отборе устойчивости к фторхинолонам и приспособленности бактерий. PLoS Патог. 5:e1000541. doi: 10.1371/journal.ppat.1000541
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Матич В., Боздоган Б., Джейкобс М. Р., Убуката К. и Аппельбаум П. К. (2003). Вклад аминокислотных замен β-лактамазы и PBP в устойчивость к амоксициллину/клавуланату у β-лактамаз-положительных, устойчивых к амоксициллину/клавуланату штаммов Haemophilus influenzae . J. Антимикроб. Чемотер. 52, 1018–1021. doi: 10.1093/jac/dkg474
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мунк, К. , Гумперт, Х.Г., Нильссон Валлин, А.И., Ван, Х.Х., и Соммер, М.О.А. (2014). Прогнозирование развития резистентности к комбинациям лекарственных средств по побочным реакциям на составные лекарственные средства. Науч. Перевод Мед. 6:262ra156. doi: 10.1126/scitranslmed.3009940
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ней М., Маруяма Т. и Чакраборти Р. (1975). Эффект бутылочного горлышка и генетическая изменчивость популяций. Эволюция 29, 1–10. doi: 10.2307/2407137
Полный текст CrossRef | Google Scholar
О’Брайен Э. Дж., Утрилла Дж. и Палссон Б. Ø. (2016). Количественная оценка и классификация использования протеома E. coli и стоимости неиспользованного белка в разных средах. PLOS Comput Biol 12:e1004998. doi: 10.1371/journal.pcbi.1004998
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Этингер М., Подглайен И., Керн В.В. и Леви С.Б. (1998). Сверхэкспрессия регуляторного гена marA или soxS у клинических мутантов топоизомеразы Escherichia coli . Антимикроб. Агенты Чемотер. 42, 2089–2094.
Реферат PubMed | Google Scholar
Оз Т., Гувенек А., Йилдиз С., Карабога Э., Тамер Ю. Т., Мумкуян Н. и др. (2014). Сила давления отбора является важным параметром, определяющим сложность эволюции устойчивости к антибиотикам. Мол. биол. Эвол. 31, 2387–2401. doi: 10.1093/molbev/msu191
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пена-Миллер Р., Ленеманн Д., Янсен Г., Фуэнтес-Эрнандес А., Розенштиль П., Шуленбург Х. и др. (2013). Когда самая мощная комбинация антибиотиков вызывает наибольшую бактериальную нагрузку: переход от улыбки к хмурому взгляду. PLoS Биол. 11:e1001540. doi: 10.1371/journal.pbio.1001540
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Риганти, К., Мини, Э., и Нобили, С. (2015). От редакции: множественная лекарственная устойчивость при раке: фармакологические стратегии от фундаментальных исследований до клинических вопросов. Фронт. Онкол. 5:105 doi: 10.3389/fonc.2015.00105
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ритц, К., и Стрейбиг, Дж. К. (2005). Биопробный анализ с использованием RJ Stat. Программное обеспечение 12, 1–22.
Google Scholar
Ритц К., Бати Ф., Стрейбиг Дж. К. и Герхард Д. (2015). Анализ доза-реакция с использованием R. PLoS ONE 10:e0146021. doi: 10.1371/journal.pone.0146021
CrossRef Full Text | Google Scholar
Родригес де Евграфов М., Гумперт Х., Мунк К., Томсен Т. Т. и Соммер М. О. А. (2015). Сопутствующая резистентность и чувствительность модулируют эволюцию резистентности высокого уровня к комбинированному лечению у Staphylococcus aureus . Мол. биол. Эвол. 32, 1175–1185. doi: 10.1093/molbev/msv006
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Саенс Ю., Бриньяс Л., Домингес Э., Руис Дж., Сарасага М., Вила Дж. и др. (2004). Механизмы резистентности штаммов Escherichia coli , устойчивых к множественным антибиотикам, человеческого, животного и пищевого происхождения. Антимикроб. Агенты Чемотер. 48, 3996–4001. doi: 10.1128/AAC.48.10.3996-4001.2004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Саломон, Р. А., и Фариас, Н. Р. (1995). Пептидный антибиотик микроцин 25 импортируется через путь tonb и белок SbmA. J. Бактериол. 177, 3323–3325.
Реферат PubMed | Google Scholar
Сандберг Т. Э., Педерсен М., Лакруа Р. А., Эбрахим А., Бонде М., Хергард М. Дж. и др. (2014). Эволюция Escherichia coli до 42°C и последующая генная инженерия выявили адаптивные механизмы и новые мутации. Мол. биол. Эвол . 31. 2647–2662. doi: 10.1093/molbev/msu209
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шурек К.Н., Марр А.К., Тейлор П.К., Виганд И., Семенек Л., Хайра Б.К. и др. (2008). Новые генетические детерминанты низкой устойчивости к аминогликозидам у Синегнойная палочка . Антимикроб. Агенты Чемотер. 52, 4213–4219. doi: 10.1128/AAC. 00507-08
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шринивасан В. Б., Вайдьянатан В., Мондал А. и Раджамохан Г. (2012). Роль двухкомпонентной системы передачи сигнала CpxAR в обеспечении устойчивости к цефепиму и хлорамфениколу у Klebsiella pneumoniae NTUH-K2044. PLoS ONE 7:e33777. doi: 10.1371/journal.pone.0033777
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Шибальский В. и Брайсон В. (1952). Генетические исследования перекрестной устойчивости микробов к токсическим агентам i.: перекрестная устойчивость Escherichia coli к пятнадцати антибиотикам1, 2. J. Bacteriol. 64:489.
Реферат PubMed | Google Scholar
Тамае К., Лю А., Ким К., Ситц Д., Хонг Дж., Бекет Э. и др. (2008). Определение гиперчувствительности к антибиотикам среди 4000 мутантов с нокаутом одного гена Кишечная палочка . J. Бактериол. 190, 5981–5988. doi: 10.1128/JB.01982-07
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Team R Core (2014). R: Язык и среда для статистических вычислений . Вена: Фонд статистических вычислений R.
Тенайлон, О., Родригес-Вердуго, А., Гаут, Р.Л., Макдональд, П., Беннетт, А.Ф., Лонг, А.Д., и др. (2012). Молекулярное разнообразие адаптивной конвергенции. Наука 335, 457–461. doi: 10.1126/science.1212986
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Топрак Э., Верес А., Мишель Ж.-Б., Чайт Р., Хартл Д.Л. и Кишони Р. (2012). Эволюционные пути к антибиотикорезистентности в условиях динамично поддерживаемого лекарственного отбора. Нац. Жене. 44, 101–105. doi: 10.1038/ng.1034
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Тернидж, Дж., и Патерсон, Д.Л. (2007). Установка и пересмотр контрольных точек антибактериальной чувствительности. клин. микробиол. Ред. 20, 391–408. doi: 10.1128/CMR.00047-06
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Валь, Л. М., Герриш, П. Дж., и Сайка-Войвод, И. (2002). Оценка влияния узких мест популяции на экспериментальную эволюцию. Генетика 162, 961–971.
Реферат PubMed | Google Scholar
Wang, Z., Wu, J., Zhu, L., and Zhan, X. (2016). Активация метаболизма глицерина у Xanthomonas campestris путем адаптивной эволюции с получением ксантановой камеди с высокой прозрачностью и низкой вязкостью из глицерина. Биоресурс. Технол. 211, 390–397. doi: 10.1016/j.biortech.2016.03.096
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Венсинг А. М., Кальвез В., Гюнтард Х. Ф., Джонсон В. А., Паредес Р., Пиллэй Д. и др. (2015). 2015 Обновление данных о мутациях лекарственной устойчивости ВИЧ-1. Топ. Антивир. Мед. 24, 132-133.
Google Scholar
Уикхэм, Х. (2009). ggplot2: элегантная графика для анализа данных . Нью-Йорк, штат Нью-Йорк: Springer Science and Business Media.
Google Scholar
Вонг К. С., Снайдер Дж.Д., Грэм К., Гринблатт Дж.Ф., Эмили А., Бабу М. и др. (2014). MoxR-АТФаза RavA и ее кофактор viaA взаимодействуют с NADH:убихиноноксидоредуктазой I в Escherichia coli . PLoS ONE 9:e85529. doi: 10.1371/journal.pone.0085529
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ян Л., Йелсбак Л., Ликке Марвиг Р., Дамкичаэр С., Уоркман С. Т., Холм Рау М. и др. (2011). Эволюционная динамика бактерий в среде человека-хозяина. Проц. Натл. акад. Sci.U.S.A. 108, 7481–7486. doi: 10.1073/pnas.1018249108
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Йетон, Дж. А., и Уитфилд, К. (2001). Очистка и характеристика WaaP из Escherichia coli , липополисахаридкиназы, необходимой для стабильности внешней мембраны. J. Biol. хим. 276, 5498–5504. doi: 10.1074/jbc.M008255200
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Йетон, Дж. А., Хайнрихс, Д. Е., Монтейро, М. А., Перри, М. Б., и Уитфилд, К. (1998). Участие waaY, waaQ и waaP в модификации липополисахарида Escherichia coli и их роль в формировании стабильной наружной мембраны. J. Biol. хим. 273, 26310–26316. doi: 10.1074/jbc.273.41.26310
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лекарственная устойчивость
Лекарственная устойчивость Лекарственная устойчивость
После успешного завершения этого раздела учащийся сможет:
- Объясните вклад мутаций в эволюцию.
- Объясните и приведите примеры естественного отбора.
- Объясните механизмы, благодаря которым бактерии и вирусы становятся устойчивыми к препаратам, используемым для лечения вызванных ими инфекций.
- Объясните, как естественный отбор способствует развитию устойчивых к антибиотикам бактерий.
- Определите следующие механизмы и то, как они способствуют развитию устойчивости бактерий к антибиотикам:
> Трансформация
> Спряжение
> Трансдукция
- Объясните, как мыло и моющие средства уменьшают количество бактерий. Перечислите шаги, которые можно предпринять, чтобы замедлить развитие устойчивых к антибиотикам бактерий.
Термин « антимикробный » является общим термином, который включает лекарства, химические вещества или другие вещества, которые убивают или замедляют рост микробов. К ним относятся:
- Антибактериальные препараты (антибиотики), убивающие бактерии
- Противовирусные средства, убивающие вирусы
- Противогрибковые средства, убивающие грибки, и
- Противопаразитарные препараты, убивающие простейших
Шотландскому ученому Александру Флемингу широко приписывают открытие антибиотических свойств пенициллина в 1928, хотя в более раннем отчете отмечалась способность пенициллиновой плесени убивать бактерии еще в 1897 г. Имеются разрозненные сообщения об использовании пенициллиновой плесени для лечения гонококковых инфекций глаз новорожденных еще в 1930 г., но только в 1930 г. В 1940-х годах пенициллин начали использовать для лечения клинических инфекций. Вскоре было признано, что это действительно замечательное лекарство. Малыми дозами лечили инфекции, вызванные стафилококком, стрептококком, нейссерией, сифилисом и многими другими бактериями. Использование пенициллина становилось все более распространенным и неизбирательным, и появились устойчивые штаммы бактерий. Для лечения инфекций требовались все большие дозы карандашина, а некоторые штаммы оказались полностью устойчивыми. Были разработаны другие антибиотики и внедрены синтетические препараты, подобные пенициллину, но также появились штаммы бактерий, устойчивых к этим новым антибиотикам. Сначала эти проблемы игнорировались, но к 19В 80-х годах стало ясно, что устойчивость к антибиотикам является важной и растущей проблемой. В 1950-х годах были опубликованы исследования, показывающие, что животные, получавшие низкие дозы антибиотиков, быстрее набирали вес, и широко распространилась практика включения антибиотиков в зерно для стимулирования роста крупного рогатого скота, птицы и свиней, что еще больше усугубило проблему устойчивости бактерий к антибиотикам. .
Антибиотики убивают или подавляют рост чувствительных бактерий. Иногда одна из бактерий выживает благодаря способности нейтрализовать действие антибиотика или уклониться от него; что одна бактерия может затем размножаться и заменить все бактерии, которые были убиты. Таким образом, воздействие антибиотиков оказывает селективное давление, что делает выжившие бактерии более устойчивыми. Кроме того, бактерии, которые когда-то были чувствительны к антибиотику, могут приобрести устойчивость в результате мутации своего генетического материала или путем приобретения фрагментов ДНК, кодирующих свойства устойчивости, от других бактерий. ДНК, кодирующая резистентность, может быть сгруппирована в один легко передаваемый пакет. Это означает, что бактерии могут стать устойчивыми ко многим противомикробным агентам из-за переноса одного фрагмента ДНК.
Приведенный ниже iFrame представляет собой веб-страницу CDC, на которой представлены краткие описания многих инфекционных агентов, у которых развилась лекарственная устойчивость.
Для получения дополнительной информации об устойчивости к противомикробным препаратам, вызванной лечением скота антибиотиками, посетите Учебный сайт по устойчивости к противомикробным препаратам.
Способность пенициллина убивать бактерии была обусловлена его способностью ингибировать бактериальный фермент, необходимый для синтеза бактериальной клеточной стенки. Способность пенициллина делать это зависела от ключевой структуры, называемой «β-лактамным кольцом». Первоначально резистентность к пенициллину возникла в результате мутации в бактерии, создавшей фермент (пенициллиназу), способный разрушать β-лактамное кольцо. Бактерии, которые обладали этой устойчивостью, сначала развились в больницах, но быстро распространились среди населения в целом. Ген, кодирующий пенициллиназу, находится не в бактериальной хромосоме, а во внекольцевом кольце ДНК, называемом «9».Плазмида 1329. » Этот дополнительный фрагмент бактериальной ДНК может быть реплицирован и перенесен из устойчивой бактерии в бактерию , которая ранее была восприимчива с помощью процесса , называемого бактериальной конъюгацией .
Метициллин, химически модифицированная версия пенициллина, был введен в 1959 году для лечения инфекций, вызванных бактериями, устойчивыми к пенициллину, но он был эффективен против более узкого спектра бактерий. Кроме того, штаммы стафилококков, устойчивых к метициллину, были зарегистрированы еще в 19 веке.61; эти штаммы приобрели ген (mecA), который инактивирует метициллин, кодируя белок, который с ним связывается. Ген mecA несется на дополнительном «мобильном генетическом элементе», стафилококковой кассетной хромосоме (SCCmec) . Первоначально штаммы MRSA встречались только в больницах, но в конце 1990-х годов MRSA был обнаружен в обществе в целом и быстро распространился по всему миру.
Учащиеся в PH709 создали 7-минутное общественное объявление, в котором объясняется, как MRSA развился в больницах и в конечном итоге распространился среди населения, вызывая случаи так называемого «внебольничного MRSA».
Вы можете узнать больше о проблемах, связанных с MRSA, и мерах по борьбе с ним, изучив iFrame, ниже которого есть ссылки на веб-страницу CDC, посвященную MRSA.
Как мы отмечали выше, новые гены возникают в результате случайных мутаций, и иногда такая мутация может придать бактерии устойчивость к антибиотику. Как только бактерия приобрела устойчивость к определенному антибиотику, она передает устойчивый аллель последующим дочерним клеткам, образующимся в результате бинарного деления. Кроме того, бактерии, приобретшие такой признак, как устойчивость к антибиотикам, могут передавать этот аллель другим бактериям с помощью любого из трех механизмов:
- Трансформация
- Спряжение
- Трансдукция
Трансформация
Когда бактериальные клетки умирают, они часто лизируются (лопаются), высвобождая свое внутриклеточное содержимое, включая фрагменты ДНК, в окружающую среду. Эти фрагменты могут быть взяты и включены в хромосому живой бактерии, чтобы придать реципиенту новые характеристики. Этот процесс называется бактериальной трансформацией, и если встроенная ДНК содержит гены, кодирующие устойчивость к антибиотику, ранее восприимчивая бактерия может быть «трансформирована» и станет устойчивой. Видео ниже (23 сек.) дает краткий обзор преобразования.
Спряжение
Многие бактерии имеют плазмид , которые представляют собой небольшие кольцевые фрагменты ДНК, отделенные от первичной бактериальной хромосомы. Эти плазмиды могут нести гены, обеспечивающие устойчивость к антибиотикам, а бактерии, содержащие плазмиды, способны конъюгировать с другими бактериями и передавать репликацию бактериям-реципиентам. На приведенной ниже электронной микрофотографии показаны две бактерии, которые соединены временным полым трубчатым соединением, называемым пилусом.
Источник: http://evolution.berkeley.edu/evolibrary/article/side_0_0/turboevolution_01
Видео ниже (2 мин 15 сек) дает отличный обзор спряжения.
В 1968 году эпидемия шигеллы унесла жизни 12 500 человек в Гватемале. Бактерии Shigella, вызвавшие вспышку, имели плазмиду, несущую устойчивость к четырем антибиотикам.
Трансдукция
Генетическая информация также может передаваться от одной бактерии к другой с помощью вируса. Бактериофаги (или просто «фаги») — это небольшие вирусы, которые заражают бактерии и используют их клеточные компоненты для репликации бактериофагов. Во время инфекции и репликации бактериальные гены могут встраиваться в вирусный геном. Затем один из вирусных повторов, несущий бактериальный аллель, может затем заразить другую бактерию и передать новый аллель.
Иллюстрация ниже начинается с донорской бактерии, имеющей ген (a+), кодирующий устойчивость к определенному антибиотику. Во 2-й и 3-й сценах вирусные белки и генетический материал реплицируются, а затем самособираются в новые вирусные частицы. Однако при этом бактериальный ген а+ внедряется в одну из вирусных частиц. Затем вирусные частицы высвобождаются из бактерии-донора, и вирус с геном а+ заражает бактерию-реципиент, после чего ген а+ может встраиваться в геном реципиента, который теперь приобрел этот ген устойчивости к антибиотикам.
Источник: http://www.cbs.dtu.dk/courses/genomics_course/roanoke/genetics980309. html
Развитие и распространение бактериальной резистентности к антибиотикам неизбежно, но ее можно значительно сократить с помощью относительно простых мер. К ним относятся:
- Профилактика инфекций (общий инфекционный контроль)
- Мытье рук среди работников здравоохранения, работников общественного питания и населения в целом
- Современная санитария: эффективные системы очистки сточных вод и обеспечения чистой водой
- Правильная практика приготовления пищи
- Быстрое выявление и изоляция новых случаев инфекции, например новых случаев туберкулеза; это особенно важно при лекарственно-устойчивых случаях
- Продолжение разработки новых антибиотиков
- Снижение использования антибиотиков в сельском хозяйстве для ускорения роста
- Обучение врачей, направленное на снижение количества неуместных назначений и неуместного использования антибиотиков широкого спектра действия
- Обучение врачей и пациентов важности приема соответствующей дозы антибиотика в течение всего периода лечения, указанного
- Информирование потребителей о важности бактериальной резистентности и бесполезности приема антибиотиков при вирусных инфекциях, таких как простуда (см. Образовательные кампании CDC по борьбе с бактериальной резистентностью.)
См. также План действий CDC в области общественного здравоохранения по борьбе с устойчивостью к противомикробным препаратам.
Как отмечалось ранее, ВИЧ представляет собой ретровирус, состоящий из одной нити РНК внутри белковой оболочки. Когда ВИЧ проникает в лимфоцит CD4, он сбрасывает свою белковую оболочку и использует вирусный фермент, называемый обратной транскриптазой, для создания сегмента ДНК, используя вирусную РНК в качестве матрицы. Эта двухцепочечная версия ДНК ВИЧ затем встраивается в ДНК инфицированной клетки-хозяина, и этот процесс называется «обратной транскрипцией». Это имеет важные последствия для развития лекарственной устойчивости ВИЧ из-за нескольких ключевых характеристик ВИЧ:
- ВИЧ размножается с огромной скоростью, производя миллиарды новых вирусных частиц каждый день.
- Общеизвестно, что обратная транскрипция подвержена ошибкам, что приводит к частым мутациям.
- Современные схемы противовирусного лечения контролируют инфекцию, но не уничтожают вирус полностью.
Учитывая персистенцию ВИЧ с высокой скоростью репликации и высоким уровнем ошибок при обратной транскрипции, мутации в ВИЧ неизбежны, и некоторые из этих мутаций в конечном итоге приводят к развитию лекарственной устойчивости. Видео ниже кратко описывает проблему.
ВИЧ заражает CD4-лимфоциты и захватывает механизм клетки для создания вирусных белков и РНК. Генетический материал ВИЧ в РНК и для репликации обратной транскриптазы приводит к очень быстрому производству новых частиц ВИЧ
Два понятия важны для понимания развития лекарственной устойчивости. Во-первых, ВИЧ-инфекция характеризуется высокими уровнями продукции и оборота вируса. У большинства нелеченых пациентов общее количество продуктивно инфицированных клеток в лимфоидной ткани оценивается примерно в 107-108 клеток. Во время хронической фазы ВИЧ-инфекции это число относительно стабильно, отражая баланс между инфицированием новых клеток-мишеней и их клиренсом.
