Как делеция гена AUTS2 влияет на формирование гиппокампа. Какие нарушения в социальном поведении вызывает мутация AUTS2. Какие механизмы лежат в основе связи между AUTS2 и аутизмом. Какие области мозга задействованы в социальном распознавании при аутизме.
Роль гена AUTS2 в развитии мозга и возникновении аутизма
Ген AUTS2 (autism susceptibility candidate 2) играет важную роль в развитии нервной системы и формировании мозга. Мутации в этом гене связаны с повышенным риском возникновения расстройств аутистического спектра и других нарушений нейроразвития. Рассмотрим, как делеции AUTS2 влияют на структуру и функции мозга, особенно в области гиппокампа, и какие последствия это имеет для социального поведения.
Функции гена AUTS2
AUTS2 кодирует ядерный белок, который экспрессируется в развивающихся областях мозга, вовлеченных в патогенез аутизма. Основные функции этого гена включают:
- Регуляцию миграции нейронов и роста нейритов
- Участие в формировании и созревании синапсов
- Влияние на развитие мозжечка и двигательные функции
Таким образом, AUTS2 вовлечен в ключевые процессы нейроразвития на разных этапах формирования мозга. Нарушения в его работе могут приводить к широкому спектру неврологических и поведенческих отклонений.

Влияние делеций AUTS2 на структуру гиппокампа
Исследования показывают, что делеции в гене AUTS2 оказывают значительное влияние на формирование гиппокампа — области мозга, критически важной для обучения, памяти и социального поведения. Как именно проявляется это влияние?
- Нарушение развития зубчатой извилины гиппокампа
- Изменение морфологии нейронов в различных зонах гиппокампа
- Снижение нейрогенеза во взрослом возрасте
- Нарушение формирования синаптических связей
Эти структурные изменения приводят к функциональным нарушениям в работе гиппокампальных нейронных сетей, что сказывается на когнитивных способностях и социальном поведении.
Нарушения социального поведения при мутациях AUTS2
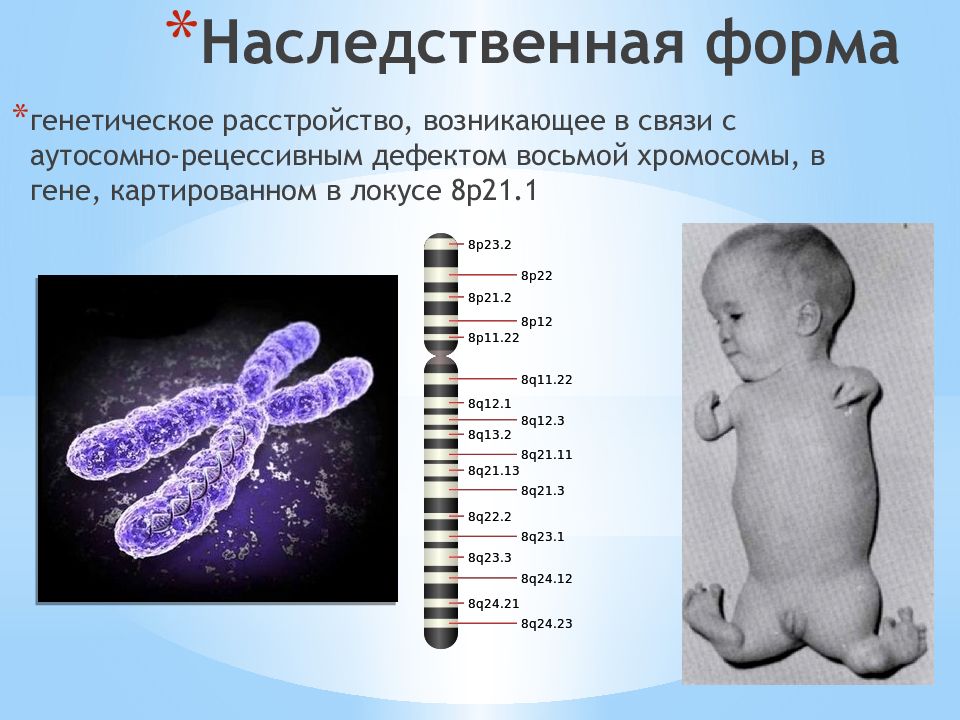
Делеции и другие мутации в гене AUTS2 ассоциированы с различными поведенческими отклонениями, характерными для расстройств аутистического спектра. Какие конкретные нарушения социального поведения наблюдаются?
- Сниженный интерес к социальным взаимодействиям
- Трудности в распознавании эмоций и социальных сигналов
- Нарушение социальной памяти и узнавания
- Стереотипное и повторяющееся поведение
- Проблемы с коммуникацией и речью
Эти поведенческие особенности во многом обусловлены структурными и функциональными изменениями в гиппокампе и других областях мозга, вызванными нарушениями в работе гена AUTS2.
Механизмы связи между AUTS2 и аутизмом
Каковы основные механизмы, связывающие нарушения в гене AUTS2 с развитием аутистических черт? Исследования выявили несколько ключевых путей:
- Изменение экспрессии генов в развивающемся мозге
- Нарушение миграции нейронов и формирования нейронных сетей
- Дисбаланс возбуждающей и тормозной нейротрансмиссии
- Изменение синаптической пластичности
- Нарушение созревания и функционирования гиппокампа
Эти механизмы приводят к комплексным изменениям в работе мозга, что проявляется в характерных для аутизма особенностях обработки информации и социального поведения.
Области мозга, задействованные в социальном распознавании при аутизме
Нарушения социального распознавания при аутизме связаны с дисфункцией нескольких ключевых областей мозга. Какие структуры играют наиболее важную роль в этом процессе?
- Гиппокамп — особенно области CA2 и CA3
- Миндалевидное тело
- Префронтальная кора
- Височная кора
- Островковая доля
Взаимодействие между этими областями критически важно для правильной обработки социальной информации. Нарушения в работе гена AUTS2 могут влиять на формирование и функционирование нейронных сетей, связывающих данные структуры.
Значение исследований AUTS2 для понимания аутизма
Изучение роли гена AUTS2 в развитии мозга и возникновении аутизма имеет большое значение для понимания механизмов этого расстройства. Какие перспективы открывают эти исследования?
- Выявление новых генетических маркеров аутизма
- Разработка ранней диагностики на основе генетического тестирования
- Создание экспериментальных моделей для изучения нейробиологии аутизма
- Поиск новых терапевтических мишеней
- Разработка персонализированных подходов к лечению
Таким образом, исследования гена AUTS2 вносят важный вклад в понимание молекулярных и нейробиологических основ расстройств аутистического спектра, открывая новые возможности для диагностики и терапии.
Перспективы терапии на основе регуляции AUTS2
Понимание роли гена AUTS2 в развитии аутизма открывает новые возможности для разработки терапевтических подходов. Какие стратегии могут быть перспективными?
- Генная терапия для коррекции мутаций AUTS2
- Таргетная регуляция экспрессии AUTS2 и его мишеней
- Фармакологическая модуляция сигнальных путей, связанных с AUTS2
- Стимуляция нейрогенеза в гиппокампе
- Воздействие на синаптическую пластичность
Хотя эти подходы пока находятся на стадии экспериментальных исследований, они открывают новые перспективы для разработки эффективных методов лечения расстройств аутистического спектра, основанных на понимании их генетических и нейробиологических механизмов.

Электросхемы ВАЗ 2114 2115 инжектор 8 клапанов
Приводятся подробные цветные схемы проводки ВАЗ 2115 (карбюратор, инжектор) с описанием электрооборудования для различных модификаций. Информация предназначена для самостоятельного ремонта авто. Многие электросхемы разделены на несколько участков для удобства просмотра через компьютер или смартфон, также имеются схемы в виде одной картинки с описанием элементов — для распечатки на принтере.
Если нужно раздавать питание внешним потребителям (видеорегистратор, фароомыватель, магнитола) при включении зажигания — взять питание лучше от аккумулятора через реле. По схеме 86 на кузов, 85 на зажигание (например жёлто-синий переключателя печки). Контакт 30 через предохранитель 30 Ампер к аккумулятору, 87 к потребителям. То есть получится при включении зажигания включится новое реле и питание будет идти напрямую от аккумулятора.
Модификации авто 2115
ВАЗ-2115. Самый первый автомобиль, который выпускался с 1997 года. На него устанавливался 1,5-литровый карбюраторный двигатель мощностью 76 лошадиных сил. Максимальная скорость составляла 165 км/ч, а время разгона от 0 до 100 км/ч составляло 13,2 секунды.
На него устанавливался 1,5-литровый карбюраторный двигатель мощностью 76 лошадиных сил. Максимальная скорость составляла 165 км/ч, а время разгона от 0 до 100 км/ч составляло 13,2 секунды.
ВАЗ-21150. Очередная модификация, выпущенная в 1998 году, оснащалась 1,5-литровым карбюраторным двигателем мощностью 68 лошадиных сил. была снята с производства в 2000 году.
ВАЗ-2115-20. Выпущенная в 2000 году модификация автомобиля, оснащенная 1,5-литровым инжекторным двигателем ВАЗ-2111 мощностью 77,8 лошадиных сил. Максимальная скорость составляла 170 км/ч, а время разгона с 0 до 100 км/ч 14 секунд.
ВАЗ-2115-40. Модификация с инжекторным двигателем объемом 1,6 литра, которая выпускалась с 2003 года. Максимальная скорость автомобиля составляла 158 км/ч, а время разгона до 100 км/ч занимало 13,2 секунды.
ВАЗ-2115-91. Автомобиль с роторно-поршневым двигателем Ванкеля объемом 1,3 литра и мощностью 135 лошадиных сил. Максимальна скорость 190 км/ч, а время разгона до 100 км/ч составляет 9 секунд.
Максимальна скорость 190 км/ч, а время разгона до 100 км/ч составляет 9 секунд.
ВАЗ-21154. Последняя модификация автомобиля с новым двигателем ВАЗ-11183 объемом 1596 см3 и мощностью 81 лошадиную силу. Выпускалась с 2007 года. Максимальная скорость и время разгона до 100 км/ч точно такое же как у ВАЗ-2115-40.
Особенности конструкции
Начинающему автолюбителю рекомендуется изучить работу инжекторного двигателя. В инжектор входит система, которая производит впрыскивание бензина. Ознакомившись с устройством, не придется обращаться в сервисный центр по поводу незначительных неполадок, а ликвидировать их дома самому. Подача горючего считается распределительной, потому что в каждый цилиндр бензин впрыскивается с помощью определенной форсунки.
В основу ВАЗ 2115 положена модель ВАЗ 21099. В новой машине в отличие от прежней поменялась форма капота, задние и передние бамперы, новая конфигурация передних крыльев, а также улучшенная аэродинамика кузова. На крышке багажника разместили спойлер — антикрыло. Внешнюю отделку оформили элементами из пластмассы, поскольку металлические разъедались коррозией. Для добавочной безопасности модель дополнили сигналом торможения и задней светотехникой.
На крышке багажника разместили спойлер — антикрыло. Внешнюю отделку оформили элементами из пластмассы, поскольку металлические разъедались коррозией. Для добавочной безопасности модель дополнили сигналом торможения и задней светотехникой.
Благодаря помещенному под капотом инжекторному узлу питания, управляемому электроникой, схема электрооборудования ВАЗ 2115 значительно отличается от 99 модификации.
Несущий конструктив кузова машины сварной, металлический, целостный. Привод представляет переднее поперечное размещение. У него коробка передач 5-ступенчатая, передние колеса подключены к приводу. Двигатель вмещает объем 1,5 л и является четырехцилиндровым, бензиновым, четырехтактным и рядным. Системы распределительного впрыскивания горючего управляются с помощью электроники.
Электрическая Схема Ваз 2115 Инжектор 8 Клапанов
Приводятся подробные цветные схемы проводки ВАЗ 2115 (карбюратор, инжектор) с описанием электрооборудования для различных модификаций. Информация предназначена для самостоятельного ремонта авто. Многие электросхемы разделены на несколько участков для удобства просмотра через компьютер или смартфон, также имеются схемы в виде одной картинки с описанием элементов – для распечатки на принтере.
Информация предназначена для самостоятельного ремонта авто. Многие электросхемы разделены на несколько участков для удобства просмотра через компьютер или смартфон, также имеются схемы в виде одной картинки с описанием элементов – для распечатки на принтере.
Если нужно раздавать питание внешним потребителям (видеорегистратор, фароомыватель, магнитола) при включении зажигания – взять питание лучше от аккумулятора через реле. По схеме 86 на кузов, 85 на зажигание (например жёлто-синий переключателя печки). Контакт 30 через предохранитель 30 Ампер к аккумулятору, 87 к потребителям. То есть получится при включении зажигания включится новое реле и питание будет идти напрямую от аккумулятора.
Модификации авто 2115
ВАЗ-2115. Самый первый автомобиль, который выпускался с 1997 года. На него устанавливался 1,5-литровый карбюраторный двигатель мощностью 76 лошадиных сил. Максимальная скорость составляла 165 км/ч, а время разгона от 0 до 100 км/ч составляло 13,2 секунды.
ВАЗ-21150. Очередная модификация, выпущенная в 1998 году, оснащалась 1,5-литровым карбюраторным двигателем мощностью 68 лошадиных сил. была снята с производства в 2000 году.
ВАЗ-2115-20. Выпущенная в 2000 году модификация автомобиля, оснащенная 1,5-литровым инжекторным двигателем ВАЗ-2111 мощностью 77,8 лошадиных сил. Максимальная скорость составляла 170 км/ч, а время разгона с 0 до 100 км/ч 14 секунд.
ВАЗ-2115-40. Модификация с инжекторным двигателем объемом 1,6 литра, которая выпускалась с 2003 года. Максимальная скорость автомобиля составляла 158 км/ч, а время разгона до 100 км/ч занимало 13,2 секунды.
ВАЗ-2115-91. Автомобиль с роторно-поршневым двигателем Ванкеля объемом 1,3 литра и мощностью 135 лошадиных сил. Максимальна скорость 190 км/ч, а время разгона до 100 км/ч составляет 9 секунд.
ВАЗ-21154. Последняя модификация автомобиля с новым двигателем ВАЗ-11183 объемом 1596 см3 и мощностью 81 лошадиную силу. Выпускалась с 2007 года. Максимальная скорость и время разгона до 100 км/ч точно такое же как у ВАЗ-2115-40.
Выпускалась с 2007 года. Максимальная скорость и время разгона до 100 км/ч точно такое же как у ВАЗ-2115-40.
Электрооборудование инжектора
Схема подключения бензонасоса Nissan R’nessa. Чиним и ремонтируем
Указатель уровня топлива газ 3307
Устройство тепловых пушек — на что обратить внимание при выборе
Правильное подключение ДХО, загораются после запуска двигателя, Форд Фокус 2.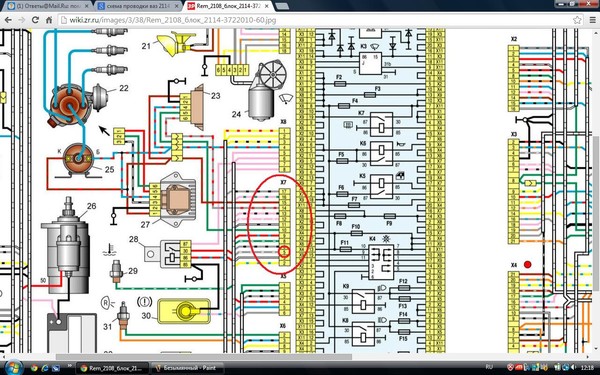
Не все водители знают, что можно сделать, если ДПКВ вышел из строя и двигатель не заводится.
Разборка блока реле и предохранителей… С4 1.6 120 л.с. срабатывание дворников.
Автосигнализация Sheriff ZX-1070 — Установка подключения сигнализации на ЗАЗ Vida
Sens sens расположение датчиков
Не заводится, -форсунки (реле форсунок) не работают.
W201.о подключении январской косы к 190
Как проверить реле 4-х контактное,5-ти контактное,в машине много
Габор
Сам щуп лучше подключить к выходу, в по своему опыту скажу, что скрепки и иголки могут сильно исказить показания.
Мэйфилд
а не подскажите как можно связаться с владельцем машины?)
Леви
спасибо. Все было доступно и понятно.
Все было доступно и понятно.
Ерлан Биринов
Эти спирали называются «ИНТЕНСИФИКАТОРЫ», они служат для придания потоку жидкости турбулентного режима, что в свою очередь исключает ламинарное течение у стенок трубы и увеличивает теплоотдачу от жидкости к стенкам радиатора трубы. Простыми словами, они созданы для того, чтобы ваша пятая точка нагревалась быстрее. И проблема заключается не в них, а в состоянии охлаждающей жидкости и чистоте всех каналов, по которым проходит этот антифриз.
Кейли
Не имитирует себя и не смущает окружающих, овердрайв — экономичный режим, так же спасает коробку от лишних переключений.
Femida
Привет. Чтобы поставить коромысло на место, места хватит руками ползти или надо подрамник снимать? С каким усилием затягивать тягу?
Сафар
Все правильно.
Мирас
Можно ли переключать АКПП в режиме D на 1 или 2 передачу
Брукс
Когда горит О, он включается, и наоборот
Грабко
Ребят, вы наркоманы что ли. Кто-то с лимонной кислотой… И даже родинка!!! Неудивительно, что у вас то зеленая слизь, то ржаво-черная. То ли медный, то ли алюминиевый радиатор по фиг! Ты убиваешь его! Для справки: Травление медных и алюминиевых плат производится только в лимонной кислоте!!! Тоже родинка. Этим же средством удаляют хром и т. д. В лучшем случае вы получите крайне рыхлую поверхность, которая засчитается еще быстрее.
Кто-то с лимонной кислотой… И даже родинка!!! Неудивительно, что у вас то зеленая слизь, то ржаво-черная. То ли медный, то ли алюминиевый радиатор по фиг! Ты убиваешь его! Для справки: Травление медных и алюминиевых плат производится только в лимонной кислоте!!! Тоже родинка. Этим же средством удаляют хром и т. д. В лучшем случае вы получите крайне рыхлую поверхность, которая засчитается еще быстрее.
Написать комментарий
Делеция Auts2 связана с гипоплазией DG и дефицитом социального распознавания: механизмы развития и нервной цепи
1. Fernandez M., Mollinedo-Gajate I., Penagarikano O., Нейронные схемы для социального познания: значение для аутизма. неврология 370, 148–162 (2018). [PubMed] [Google Scholar]
2. Weigelt S., Koldewyn K., Kanwisher N., Распознавание лица при расстройствах аутистического спектра: обзор поведенческих исследований. Неврологи. Биоповедение. преп. 36, 1060–1084 (2012). [PubMed] [Академия Google]
3. Цзян С., Боллич А. , Кокс П., Хайдер Э., Джеймс Дж., Говани С.А., Хаджихани Н., Бланц В., Маноах Д.С., Бартон Дж.Дж.С., Гайярд В.Д., Ризенхубер М.,
Количественная связь между дефицитом распознавания лиц и избирательностью нейронов в отношении лиц при аутизме. Клиника НейроИмидж.
2,
320–331 (2013). [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Кокс П., Хайдер Э., Джеймс Дж., Говани С.А., Хаджихани Н., Бланц В., Маноах Д.С., Бартон Дж.Дж.С., Гайярд В.Д., Ризенхубер М.,
Количественная связь между дефицитом распознавания лиц и избирательностью нейронов в отношении лиц при аутизме. Клиника НейроИмидж.
2,
320–331 (2013). [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Тан Дж., Фолкмер М., Хорлин С., Тан Т., Ваз С., Фолкмер Т., Стратегии распознавания лиц и визуального поиска при расстройствах аутистического спектра: изменение и расширение недавнего обзора Weigelt et al. ПЛОС ОДИН 10, e0134439(2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Rigby S. N., Stoesz B. M., Jakobson L. S., Эмпатия и обработка лица у взрослых с расстройством аутистического спектра и без него. Аутизм рез. 11, 942–955 (2018). [PubMed] [Google Scholar]
6. Сассон Н. Дж., Ноулин Р. Б., Пинкхэм А. Э., Социальное познание, социальные навыки и широкий фенотип аутизма. аутизм 17, 655–667 (2013). [PubMed] [Google Scholar]
7. Halliday D.W., MacDonald S.W., Scherf K.S., Tanaka J.W.,
Взаимная модель распознавания лиц и аутичных черт: доказательства с точки зрения индивидуальных различий. ПЛОС ОДИН
9,
e94013 (2014 г.). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Halliday D.W., MacDonald S.W., Scherf K.S., Tanaka J.W.,
Взаимная модель распознавания лиц и аутичных черт: доказательства с точки зрения индивидуальных различий. ПЛОС ОДИН
9,
e94013 (2014 г.). [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Льюис Г. Дж., Шейкшафт Н. Г., Пломин Р., Распознавание лица и компонент социальных трудностей аутистического фенотипа: доказательства фенотипических и генетических связей. Дж. Аутизм Дев. Беспорядок. 48, 2758–2765 (2018). [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Иидака Т., Терашима С., Ямасита К., Окада Т., Садато Н., Йонекура Ю., Диссоциативные нейронные реакции в гиппокампе на восстановление идентичности лица и эмоций: исследование фМРТ, связанное с событием. Гиппокамп 13, 429–436 (2003 г.). [PubMed] [Google Scholar]
10. Taylor M.J., Mills T., Pang E.W., Развитие распознавания лиц; вклады гиппокампа и лобной доли, определенные с помощью МЭГ. Мозг Топогр. 24, 261–270 (2011). [PubMed] [Google Scholar]
11. Олсен Р. К., Ли Ю., Кубе Дж., Розенбаум Р. С., Грейди К. Л., Москович М., Райан Дж. Д.,
Роль реляционного связывания в памяти предметов: данные распознавания лиц в случае амнезии развития. Дж. Нейроски.
35,
5342–5350 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Олсен Р. К., Ли Ю., Кубе Дж., Розенбаум Р. С., Грейди К. Л., Москович М., Райан Дж. Д.,
Роль реляционного связывания в памяти предметов: данные распознавания лиц в случае амнезии развития. Дж. Нейроски.
35,
5342–5350 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Джи Э., Вейкерт К. С., Ленрут Р., Киндлер Дж., Скиллетер А. Дж., Веркаммен А., Уайт К., Гур Р. Э., Вейкерт Т. В., Дополнительный селективный модулятор рецепторов эстрогена увеличивает активность нейронов в гиппокампе и нижней лобной извилине во время эмоционального распознавания лиц при шизофрении. Перевод Психиатрия 6, е795 (2016). [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Aylward E.H., Minshew N.J., Goldstein G., Honeycutt N.A., Augustine A.M., Yates K.O., Barta P.E., Pearlson G.D., Объемы МРТ миндалевидного тела и гиппокампа у аутичных подростков и взрослых без умственной отсталости. неврология 53, 2145–2150 (1999). [PubMed] [Google Scholar]
14. Николсон Р., ДеВито Т.Дж.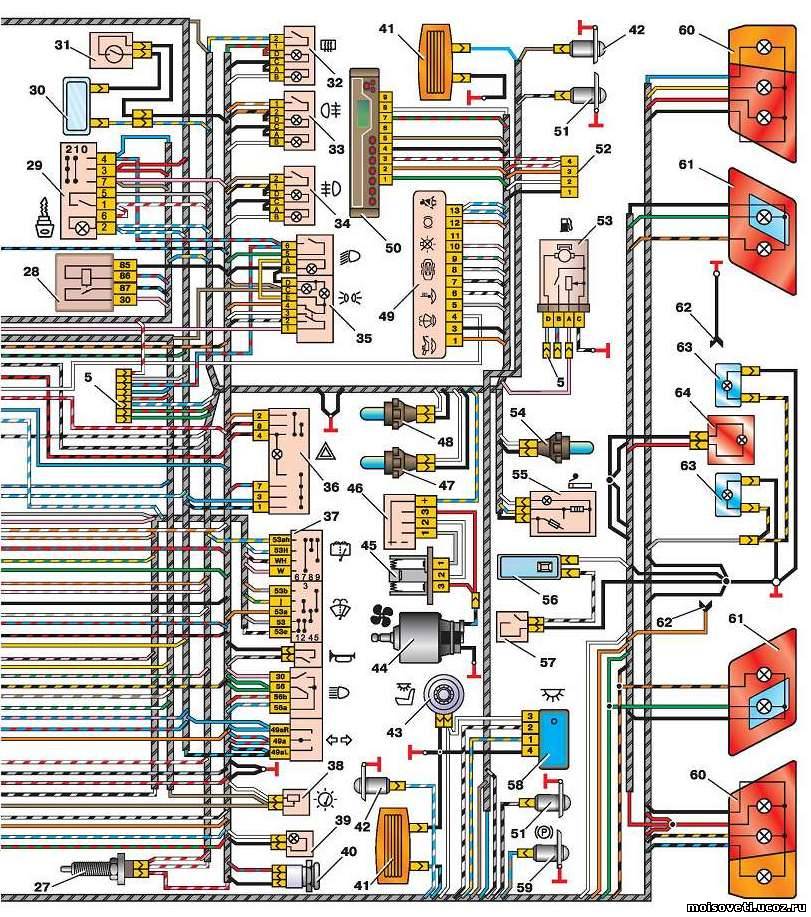 , Видал С.Н., Суй Ю., Хаяши К.М., Дрост Д.Дж., Уильямсон П.С., Раджакумар Н., Тога А.В., Томпсон П.М.,
Обнаружение и картирование аномалий гиппокампа при аутизме. Психиатрия рез.
148,
11–21 (2006). [PubMed] [Google Scholar]
, Видал С.Н., Суй Ю., Хаяши К.М., Дрост Д.Дж., Уильямсон П.С., Раджакумар Н., Тога А.В., Томпсон П.М.,
Обнаружение и картирование аномалий гиппокампа при аутизме. Психиатрия рез.
148,
11–21 (2006). [PubMed] [Google Scholar]
15. Saitoh O., Karns C.M., Courchesne E., Развитие образования гиппокампа от 2 до 42 лет: МРТ свидетельствует о меньшей площади зубцов при аутизме. Мозг 124, 1317–1324 (2001). [PubMed] [Академия Google]
16. Wegiel J., Kuchna I., Nowicki K., Imaki H., Wegiel J., Marchi E., Ma S.Y., Chauhan A., Chauhan V., Bobrowicz T.W., de Leon M., Louis L.A.S., Коэн И. Л., Лондон Э., Браун В. Т., Вишневски Т., Нейропатология аутизма: дефекты нейрогенеза и миграции нейронов, диспластические изменения. Акта Нейропатол. 119, 755–770 (2010). [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Wegiel J., Schanen N.C., Cook E.H., Sigman M., Brown W.T., Kuchna I., Nowicki K., Wegiel J., Imaki H., Ma С. Ю., Марчи Э., Вежба-Бобровиц Т., Чаухан А., Чаухан В., Коэн И.
18. Султана Р., Ю К.-Э., Ю Дж., Мансон Дж., Чен Д., Хуа В., Эстес А., Кортес Ф. , де ла Барра Ф., Ю Д., Хайдер С. Т., Траск Б. Дж., Грин Э. Д., Раскинд В. Х., Дистеш С. М., Вийсман Э., Доусон Г., Сторм Д. Р., Шелленберг Г. Д., Виллакрес Э. К., Идентификация нового гена на хромосоме 7q11.2 прервана точкой разрыва транслокации у пары аутичных близнецов. Геномика 80, 129–134 (2002). [PubMed] [Академия Google]
19. Хуан С. Л., Цзоу Ю. С., Махер Т. А., Ньютон С., Милунский Дж. М., Точка останова сбалансированной транслокации de novo, обрезающая ген-кандидат 2 восприимчивости к аутизму (AUTS2) у пациента с аутизмом. Являюсь. Дж. Мед. Жене. А 152А, 2112–2114 (2010). [PubMed] [Google Scholar]
20. Beunders G. , de Munnik S.A., van der Aa N., Ceulemans B., Voorhoeve E., Groffen A.J., Nillesen W.M., Meijers-Heijboer E.J., Frank Kooy R., Yntema Х. Г., Систерманс Э. А.,
Два взрослых мужчины с патогенными вариантами AUTS2, включая делецию двух пар оснований, дополнительно определяют синдром AUTS2. Евро. Дж. Хам. Жене.
23,
803–807 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
, de Munnik S.A., van der Aa N., Ceulemans B., Voorhoeve E., Groffen A.J., Nillesen W.M., Meijers-Heijboer E.J., Frank Kooy R., Yntema Х. Г., Систерманс Э. А.,
Два взрослых мужчины с патогенными вариантами AUTS2, включая делецию двух пар оснований, дополнительно определяют синдром AUTS2. Евро. Дж. Хам. Жене.
23,
803–807 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Фан Ю., Цю В., Ван Л., Гу С., Ю Ю., Экзонные делеции AUTS2 у китайских пациентов с задержкой развития и умственной отсталостью. Являюсь. Дж. Мед. Жене. А 170А, 515–522 (2016). [PubMed] [Google Scholar]
22. Беундерс Г., Ван де Камп Дж., Васудеван П., Мортон Дж., Сметс К., Клифстра Т., де Мунник С. А., Шурс-Хоймакерс Дж., Сеулеманс Б. , Zollino M., Hoffjan S., Wieczorek S., So J., Mercer L., Walker T., Velsher L.; исследование DDD, Паркер М.Дж., Маги А.С., Элфферс Б., Кой Р.Ф., Интема Х.Г., Мейерс-Хейбур Э.Дж., Систерманс Э.А.,
Подробный клинический анализ 13 пациентов с синдромом AUTS2 дополнительно очерчивает фенотипический спектр и подчеркивает поведенческий фенотип. Дж. Мед. Жене.
53,
523–532 (2016). [PubMed] [Академия Google]
Дж. Мед. Жене.
53,
523–532 (2016). [PubMed] [Академия Google]
23. Нагамани С.С., Эрез А., Бен-Зеев Б., Фридман М., Винтер С., Зеллер Р., Эль-Хечен Д., Эскобар Л., Станкевич П., Патель А., Чунг С.В., Обнаружение вариации числа копий в гене AUTS2 с помощью целевого экзонного массива CGH у пациентов с задержкой развития и расстройствами аутистического спектра. Евро. Дж. Хам. Жене. 21, 343–346 (2013). [ЧВК бесплатная статья] [PubMed] [Google Scholar]
24. Сенгун Э., Ярарбас К., Касакян С., Аланай Ю., Синдром AUTS2 у 68-летней женщины: естественное течение и дальнейшее определение фенотипа. Являюсь. Дж. Мед. Жене. А 170, 3231–3236 (2016). [PubMed] [Академия Google]
25. Бендерс Г., Вурхув Э., Гольцио К., Пардо Л. М., Розенфельд Дж. А., Талковски М. Э., Симоник И., Лайонел А. С., Вергульт С., Пайетт Р. Э., Ван де Камп Дж., Ньювинт А., Вайс М. М., Риццу П., Вервер Л. Е. Н. И., ван Спендонк Р. М. Л., Шен Ю., Ву Б. Л., Ю Т., Ю Ю., Чан С., Гузелла Дж. Ф., Линдгрен А. М., Мортон С. С., ван Бинсберген Э., Балк С., Ван Россем Э., Ванаккер О., Армстронг Р., Парк С. М., Гринхал Л., Мэй У., Нил Н. Дж., Эбботт К. М., Селл С., Ладда Р., Фарбер Д. М., Бадер П. И., Кушинг Т., Драутц Дж. М. , Konczal L., Nash P., de Los Reyes E., Carter M.T., Hopkins E., Marshall C.R., Osborne L.R., Gripp K.W., Thrush D.L., Hashimoto S., Gastier-Foster J.M., Astbury C., Ylstra B. , Мейерс-Хейбур Х., Постхума Д., Ментен Б., Мортье Г., Шерер С. В., Эйхлер Э. Э., Гирираджан С., Кацанис Н., Гроффен А. Дж., Систерманс Э. А.,
Экзонные делеции в AUTS2 вызывают синдромальную форму умственной отсталости и указывают на критическую роль С-конца. Являюсь. Дж. Хам. Жене.
92,
210–220 (2013). [Бесплатная статья PMC] [PubMed] [Google Scholar]
С., ван Бинсберген Э., Балк С., Ван Россем Э., Ванаккер О., Армстронг Р., Парк С. М., Гринхал Л., Мэй У., Нил Н. Дж., Эбботт К. М., Селл С., Ладда Р., Фарбер Д. М., Бадер П. И., Кушинг Т., Драутц Дж. М. , Konczal L., Nash P., de Los Reyes E., Carter M.T., Hopkins E., Marshall C.R., Osborne L.R., Gripp K.W., Thrush D.L., Hashimoto S., Gastier-Foster J.M., Astbury C., Ylstra B. , Мейерс-Хейбур Х., Постхума Д., Ментен Б., Мортье Г., Шерер С. В., Эйхлер Э. Э., Гирираджан С., Кацанис Н., Гроффен А. Дж., Систерманс Э. А.,
Экзонные делеции в AUTS2 вызывают синдромальную форму умственной отсталости и указывают на критическую роль С-конца. Являюсь. Дж. Хам. Жене.
92,
210–220 (2013). [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Гао З., Ли П., Стаффорд Дж. М., фон Шиммельманн М., Шефер А., Рейнберг Д., Комплекс AUTS2-Polycomb активирует экспрессию генов в ЦНС. Природа 516, 349–354 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Хори К., Нагаи Т., Шан В., Сакамото А. , Тая С., Хасимото Р., Хаяши Т., Абэ М., Ямадзаки М. ., Накао К., Нисиока Т., Сакимура К., Ямада К., Кайбути К., Хосино М.,
Регуляция цитоскелета с помощью AUTS2 при миграции нейронов и нейритогенезе. Сотовый представитель
9,
2166–2179 (2014). [PubMed] [Google Scholar]
, Тая С., Хасимото Р., Хаяши Т., Абэ М., Ямадзаки М. ., Накао К., Нисиока Т., Сакимура К., Ямада К., Кайбути К., Хосино М.,
Регуляция цитоскелета с помощью AUTS2 при миграции нейронов и нейритогенезе. Сотовый представитель
9,
2166–2179 (2014). [PubMed] [Google Scholar]
28. Ямаширо К., Хори К., Лай Э. С. К., Аоки Р., Симаока К., Аримура Н., Эгуса С. Ф., Сакамото А., Абэ М., Сакимура К., Ватанабэ Т., Уэсака Н., Кано М., Хосино М., AUTS2 регулирует развитие мозжечка, созревание клеток Пуркинье, двигательную функцию и социальную коммуникацию. iScience 23, 101820 (2020). [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Hori K., Yamashiro K., Nagai T., Shan W., Egusa S. F., Shimaoka K., Kuniishi H., Sekiguchi M., Go Y. , Тацумото С., Ямада М., Шираиси Р., Канно К., Мияшита С., Сакамото А., Абэ М., Сакимура К., Соне М., Сохья К., Кунуги Х., Вада К., Ямада М., Ямада К., Хосино М.,
AUTS2 регулирует синапсы для правильных синаптических входов и социальной коммуникации. iScience
23,
101183 (2020). [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Бедоньи Ф., Ходж Р. Д., Нельсон Б. Р., Фредерик Э. А., Шиба Н., Даза Р. А., Хевнер Р. Ф., Кандидат на восприимчивость к аутизму 2 (Auts2) кодирует ядерный белок, экспрессируемый в развивающихся областях мозга, вовлеченных в невропатологию аутизма. Джин Экспр. Узоры 10, 9–15 (2010). [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Cismasiu V.B., Adamo K., Gecewicz J., Duque J., Lin Q., Avram D., BCL11B функционально связывается с комплексом NuRD в Т-лимфоцитах для репрессии целевого промотора. Онкоген 24, 6753–6764 (2005). [PubMed] [Академия Google]
32. Саймон Р., Брылка Х., Швеглер Х., Венкатараманаппа С., Андрачке Дж., Вигреффе К., Лю П., Фукс Э., Дженкинс Н. А., Коупленд Н. Г., Бирхмайер К., Бритш С., Двойная функция Bcl11b/Ctip2 в нейрогенезе гиппокампа. ЭМБО Дж. 31, 2922–2936 (2012). [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Хагихара Х., Такао К., Уолтон Н. М., Мацумото М., Миякава Т. ,
Незрелая зубчатая извилина: эндофенотип нервно-психических расстройств. Нейр Пласт.
2013,
1–24 (2013). [Бесплатная статья PMC] [PubMed] [Google Scholar]
,
Незрелая зубчатая извилина: эндофенотип нервно-психических расстройств. Нейр Пласт.
2013,
1–24 (2013). [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Кастанса А. С., Рамирес С., Трипати П. П., Даза Р. А. М., Калуме Ф. К., Рамирес Дж. М., Хевнер Р. Ф., AUTS2 регулирует метаболизм РНК и развитие зубчатой извилины у мышей. Церебр. кора 26, 4808–4824 (2021). [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Hitti F.L., Siegelbaum S.A., Область CA2 гиппокампа необходима для социальной памяти. Природа 508, 88–92 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Стивенсон Э. Л., Колдуэлл Х. К., Поражения области СА2 гиппокампа нарушают социальную память у мышей. Евро. Дж. Нейроски. 40, 3294–3301 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Смит А. С., Уильямс Аврам С. К., Цимерблит-Сабба А., Сонг Дж., Янг В. С.,
Целенаправленная активация области CA2 гиппокампа сильно усиливает социальную память. Мол. Психиатрия
21,
1137–1144 (2016). [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Олива А., Фернандес-Руис А., Лерой Ф., Зигельбаум С. А., Островолновые пульсации гиппокампа CA2 реактивируют и способствуют социальной памяти. Природа 587, 264–269 (2020). [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Мейра Т., Лерой Ф., Басс Э. В., Олива А., Парк Дж., Сигельбаум С. А., Цепь гиппокампа, соединяющая дорсальный CA2 с вентральным CA1, имеет решающее значение для динамики социальной памяти. Нац. коммун. 9, 4163 (2018). [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Окуяма Т., Китамура Т., Рой Д. С., Итохара С., Тонегава С., Вентральные нейроны СА1 хранят социальную память. Наука 353, 1536–1541 (2016). [PMC free article] [PubMed] [Google Scholar]
41. Chen S., He L., Huang A.J.Y., Boehringer R., Robert V., Wintzer M.E., Polygalov D., Weitemier A.Z., Tao Y., Gu М., Миддлтон С.Дж., Намики К., Хама Х., Терро Л., Шевалейр В., Хиоки Х., Мияваки А., Пискоровски Р.А., МакХью Т.Дж. ,
Сигнал новизны гипоталамуса модулирует память гиппокампа. Природа
586,
270–274 (2020). [PubMed] [Академия Google]
,
Сигнал новизны гипоталамуса модулирует память гиппокампа. Природа
586,
270–274 (2020). [PubMed] [Академия Google]
42. Чианг М.С., Хуанг А.Дж.Ю., Винцер М.Е., Ошима Т., МакХью Т.Дж., Роль CA3 в памяти социального распознавания. Поведение Мозг Res. 354, 22–30 (2018). [PubMed] [Google Scholar]
43. Liu X.-D., Ai P.-H., Zhu X.-N., Pan Y.-B., Halford M.M., Henkemeyer M., Feng D.- Ф., Сюй Т.-Л., Сунь С., Сюй Н.-Дж., Гиппокампальный мультипротеиновый комплекс Lnx1-NMDAR опосредует начальную социальную память. Мол. Психиатрия 26, 3956–3969 (2021). [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Leung C., Cao F., Nguyen R., Joshi K., Aqrabawi A.J., Xia S., Cortez M.A., Snead O.C. III, Kim JC, Jia З., Активация энторинальных корковых проекций на зубчатую извилину лежит в основе извлечения социальной памяти. Сотовый представитель 23, 2379–2391 (2018). [PubMed] [Google Scholar]
45. Pan W. X., McNaughton N.,
Супрамаммилярная область: ее организация, функции и связь с гиппокампом. прог. Нейробиол.
74,
127–166 (2004). [PubMed] [Google Scholar]
прог. Нейробиол.
74,
127–166 (2004). [PubMed] [Google Scholar]
46. Раам Т., Макэвой К. М., Беснард А., Винема А. Х., Сахай А., Окситоциновые рецепторы гиппокампа необходимы для распознавания социальных стимулов. Нац. коммун. 8, 2001 (2017). [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Nakanishi K., Saito H., Abe K., Супрамаммилярное ядро вносит вклад в ассоциативную потенциацию ВПСП-спайков в зубчатой гирзине крысы vivo. Евро. Дж. Нейроски. 13, 793–800 (2001). [PubMed] [Google Scholar]
48. Хашимотодани Ю., Карубе Ф., Янагава Ю., Фудзияма Ф., Кано М., Супрамаммилярное ядро, афферентное к зубчатой извилине, совместно высвобождает глутамат и ГАМК и усиливает продукцию гранулярных клеток. Сотовый представитель 25, 2704–2715.e4 (2018). [PubMed] [Google Scholar]
49. Вертес Р. П., Тета-ритм гиппокампа: признак кратковременной памяти. Гиппокамп 15, 923–935 (2005). [PubMed] [Google Scholar]
50. Кохара К., Пигнателли М., Ривест А.Дж., Юнг Х. Ю., Китамура Т., Су Дж., Фрэнк Д., Кадзикава К., Мисе Н., Обата Ю., Викершем И. Р., Тонегава С.,
Генетические и оптогенетические инструменты, специфичные для типа клеток, выявляют цепи CA2 гиппокампа. Нац. Неврологи.
17,
269–279 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ю., Китамура Т., Су Дж., Фрэнк Д., Кадзикава К., Мисе Н., Обата Ю., Викершем И. Р., Тонегава С.,
Генетические и оптогенетические инструменты, специфичные для типа клеток, выявляют цепи CA2 гиппокампа. Нац. Неврологи.
17,
269–279 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Abbott A.E., Linke A.C., Nair A., Jahedi A., Alba L.A., Keown C.L., Fishman I., Müller R.A., Повторяющееся поведение при аутизме связано с дисбалансом корково-стриарной связи: МРТ-исследование функциональной связи. соц. Познан. Оказывать воздействие. Неврологи. 13, 32–42 (2018). [PMC free article] [PubMed] [Google Scholar]
Нейробиология ухода за собой грызунов и ее значение для трансляционной нейронауки. Нац. Преподобный Нейроски. 17, 45–59(2016). [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Kawaguchi Y., Подтипы пирамидных клеток и их синаптические связи в слое 5 лобной коры крысы. Церебр. кора 27, 5755–5771 (2017). [PubMed] [Google Scholar]
54. Нагата А. , Судзуки Ю., Игараши М., Эгучи Н., Тох Х., Ураде Ю., Хаяиси О.,
Простагландин-D-синтаза головного мозга человека эволюционно дифференцировалась от белков-носителей липофильных лигандов. проц. Натл. акад. науч. США.
88,
4020–4024 (1991). [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Судзуки Ю., Игараши М., Эгучи Н., Тох Х., Ураде Ю., Хаяиси О.,
Простагландин-D-синтаза головного мозга человека эволюционно дифференцировалась от белков-носителей липофильных лигандов. проц. Натл. акад. науч. США.
88,
4020–4024 (1991). [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Чай Д., Ченг Ю., Сунь Ю., Ян Дж., Ху Р., Чжан Л., Цзян Х., Многократное воздействие севофлурана во время беременности ингибирует миграцию нейронов за счет повышения уровня простагландин-D2-синтазы. Междунар. Дж. Дев. Неврологи. 78, 77–82 (2019). [PubMed] [Google Scholar]
56. Li J., Chai A., Wang L., Ma Y., Wu Z., Yu H., Mei L., Lu L., Zhang C., Yue W. , Сюй Л., Рао Ю., Чжан Д., Синаптическая передача сигналов P-Rex1 регулирует гиппокампальную длительную депрессию и аутоподобное социальное поведение. проц. Натл. акад. науч. США. 112, Е6964–E6972 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Sun X., Wang L., Wei C., Sun M., Li Q., Meng H., Yue W., Zhang D., Li J . ,
Дисфункция Trio GEF1 связана с дисбалансом возбуждения/торможения и поведением, подобным аутизму, посредством регуляции миграции интернейронов. Мол. Психиатрия, (2021). [PubMed] [Google Scholar]
,
Дисфункция Trio GEF1 связана с дисбалансом возбуждения/торможения и поведением, подобным аутизму, посредством регуляции миграции интернейронов. Мол. Психиатрия, (2021). [PubMed] [Google Scholar]
58. Джолли А., Корбетт М., Макгрегор Л., Уотерс В., Браун С., Николл Дж., Ю С., Внутригенная делеция de novo гена кандидата 2 восприимчивости к аутизму (AUTS2) у пациента с задержкой развития: отчет о клиническом случае и обзор литературы. Являюсь. Дж. Мед. Жене. А 161А, 1508–1512 (2013). [PubMed] [Академия Google]
59. Амарилло И. Э., Ли В. Л., Ли С., Вилен Э., Кантарчи С., Делеция одного экзона AUTS2 de novo у пациента с расстройством речи и языка: обзор нарушенного AUTS2 и дополнительные доказательства его роли в нарушениях развития нервной системы. Являюсь. Дж. Мед. Жене. А 164А, 958–965 (2014). [PubMed] [Google Scholar]
60. Liu Y., Zhao D., Dong R., Yang X., Zhang Y., Tammimies K., Uddin M., Scherer S.W., Gai Z.,
Делеция экзона 1 de novo гена AUTS2 у пациента с расстройством аутистического спектра и задержкой развития: клинический случай и краткий обзор литературы.
